Embed Size (px)
Citation preview

RESEARCH PAPER New Biotechnology �Volume 28, Number 3 �April 2011
Research
Pap
er
Rational improvement of cell-free proteinsynthesis
Anders Pedersen1,2, Kristofer Hellberg1, Johan Enberg1 and B. Goran Karlsson1
1 Swedish NMR Centre, University of Gothenburg, PO Box 465, SE-40530 Gothenburg, Sweden2Department of Chemistry, University of Gothenburg, PO Box 462, SE-40530 Gothenburg, Sweden
Experimental design principles were applied on cell-free protein synthesis to optimize performance with
regard to the expression yield and the incorporation efficiency of amino acid precursors. A versatile
screening platform based on batch-mode cell-free expression and central composite design was used. The
performance of different extracts (S12 and S30), the concentration dependence of key components and
the effect of different additives were investigated. We find that the initial expression yield can be
enhanced twofold to threefold in this manner. The improved conditions comprise a modified S12
extract, optimized concentrations of creatine phosphate and key amino acids, as well as introduction of
ketoacid additives. Our results show that current cell-free expression technology is far from optimal and
that higher yields and increased utilization of the provided precursors are attainable with further
optimization.
Introduction‘Cell-free’ protein synthesis (CFPS) involves the use of a cellular
extract of prokaryotic (e.g. Escherichia coli) or eukaryotic (e.g. rabbit
reticulocyte, wheat germ) origin to provide the translational
machinery, which is required for production of protein using an
added DNA template. With the advent of preparative level expres-
sion protocols, CFPS has become an attractive alternative to reg-
ular in vivo protein overexpression. Structural proteomics
initiatives (e.g. CESG, RIKEN) have used cell-free expression as
the main tool to produce isotopically labeled proteins for NMR
spectroscopy structure determination. One characteristic of cell-
free expression is that it is fast and malleable in terms of changing
the optimal reaction conditions as the translation machinery is
not enclosed by a membrane. A given added reagent has direct
contact with the nascent polypeptide chain and can be used as
long as it does not interfere with the system, including chaper-
onins, cofactors, lipids, detergents and redox buffers, effectively
making cell-free synthesis the most versatile expression system
currently available. The ability to use PCR products as template has
led to the adoption of the technique for efficient construct screen-
Corresponding author: Karlsson, B.G. ([email protected])
218 www.elsevier.com/locate/nbt 1871-6784/$
ing [1] and proteomics approaches [2,3], and will probably be used
extensively in systems biology applications.
For NMR spectroscopy applications, there are additional advan-
tages that make the method the preferred choice to produce
isotopically labeled proteins. Isotopic scrambling is not a serious
problem as expression is made in the absence of an active meta-
bolism, and incorporation of any isotope, including deuterium, is
simply dictated by the choice of the included amino acids and does
not have an effect on expression yield. In many cases, particularly
for large systems, deuteration is necessary to improve NMR signal
linewidths. For in vivo expression, this necessitates growth in
heavy water, typically with lower yields, and the concomitant
back exchange of protons at peptide amides can be problematic,
for example, for membrane proteins. These problems are not
relevant for cell-free synthesis as it is performed in regular water.
Because cell-free synthesis has been widely used for more than
30 years, it is not surprising that a variety of expression protocols
have been described. It is not evident, however, how the minutiae
of these protocols have been derived; most probably they result
from empirically varying a single or a few parameters in parallel at
a time. For some included components, such an optimization is
sufficient and is not altered much by changing other parts of the
- see front matter � 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.nbt.2010.06.015

New Biotechnology �Volume 28, Number 3 �April 2011 RESEARCH PAPER
ResearchPap
er
reaction mixture. For others, however, it is likely that component
interactions affect the optimal setting. This is a shortcoming we
sought out to investigate in this work. Recent years have seen a
marked increase in the understanding of batch-based CFPS, parti-
cularly by the Swartz group, to the extent that we now know that
central metabolism can be activated and low-cost protein produc-
tion can be accomplished [4]. The ‘Cytomim’ system developed by
Jewett et al. [4] is not suitable for NMR applications, however, as it
uses large amounts of L-glutamate in the buffer as an energy source,
which for labeling purposes would mean a substantial cost and
unwanted possibilities for isotopic scrambling.
We wanted to set up an in-house cell-free protein expression
capability to enable routine use of this technique for NMR appli-
cations in a way that also was economically sound. Here, we
present the use of experimental design on cell-free protein synth-
esis with emphasis on improving the incorporation of amino acids
to facilitate efficient isotope labeling for NMR spectroscopy appli-
cations. We find that by applying this methodology it is possible to
improve expression yield several-fold by adjusting the energy
substrate and amino acid composition of the original protocol,
particularly the concentrations of serine and glutamine, as well as
by the addition of malic acid, 2-oxoglutaric acid and succinic acid.
The generality of our results and future optimization approaches
are discussed.
Materials and methodsChemicals, E. coli strains and vectorsCreatine phosphate (CP), creatine kinase (CK), complete EDTA-
free protease inhibitor cocktail and protector RNAse inhibitor were
purchased from Roche Applied Science, plasmid purification kits
from QIAGEN and TALON affinity resin from Clontech. All other
chemicals were purchased from Sigma–Aldrich. E. coli DH5a(Invi-
trogen) was used for plasmid amplification, BL21(DE3) Rosetta
(Novagen) for extract preparation and BL21(DE3) for T7 RNA
polymerase (T7RNAP) expression. The cycle 3 mutant of GFP
(GFPcyc3) [5] cloned into the vector pIVEX2.4d (Roche Applied
[(Figure_1)TD$FIG]
FIGURE 1
Flowchart of the steps taken to change the original to the final protocol.
Science), the Discosoma sp. DsRed mutant tdTomato [6] cloned
into pRSETB (Invitrogen) and the E. coli elongation factor Ts (EF-
Ts) cloned into pIX3.0 (QIAGEN) were used as reporter proteins of
expression yield. Additionally, constructs of E. coli transhydrogen-
ase domain III (TH dIII) and wild-type GFP (wtGFP) were used to
monitor the overall effect of the protocol optimization. T7RNAP
was prepared according to [7,8].
Extract cultivation and preparationE. coli BL21(DE3) Rosetta was grown at 378C in 2xYTPG medium
[9] in the presence of 30 mg/ml chloramphenicol in a Braun Biostat
C fermentor. The fermentor run was halted when the OD600
reached 3.5. The cell suspension was cooled through a stainless-
steel coil immersed in ice water before being harvested by cen-
trifugation. The temperature of the cell suspension changed from
378C to 10–158C after passage through the coil. The cell pellet was
resuspended in a total of 3 l extract buffer (10 mM Tris–acetate pH
8.2, 14 mM magnesium acetate, 60 mM potassium acetate, 1 mM
DTT) before repeating centrifugation. Finally, the cell pellet was
resuspended in 10 ml S30 buffer/8 g wet cells and stored at �808Cuntil usage. S30 and S12 extracts were prepared from the same
lysate essentially according to [10,11], respectively, but the S12
extract was subjected to an additional dialysis step against extract
buffer before flash-freezing in liquid nitrogen.
Cell-free protein synthesisCFPS reactions were performed in volumes of 100 ml or 200 ml
when using microtiter plates and 500 ml when using 1.5 ml tubes.
The initial cell-free reaction mix was essentially described in [12]
but the reactions were performed in the batch format, PEG8000
was omitted and D-glutamic acid (D-Glu) was used as a counter-ion
to ammonium and potassium instead of acetate, as in [13]. The
initial and final protocol contents of the reaction mixture are listed
in Fig. 1. Expression was performed in an Eppendorf Thermomixer
at 800 rpm for two hours at the relevant temperature. A liquid
handling robot (Xiril X75) was employed to facilitate condition
www.elsevier.com/locate/nbt 219
RESEARCH PAPER New Biotechnology �Volume 28, Number 3 �April 2011
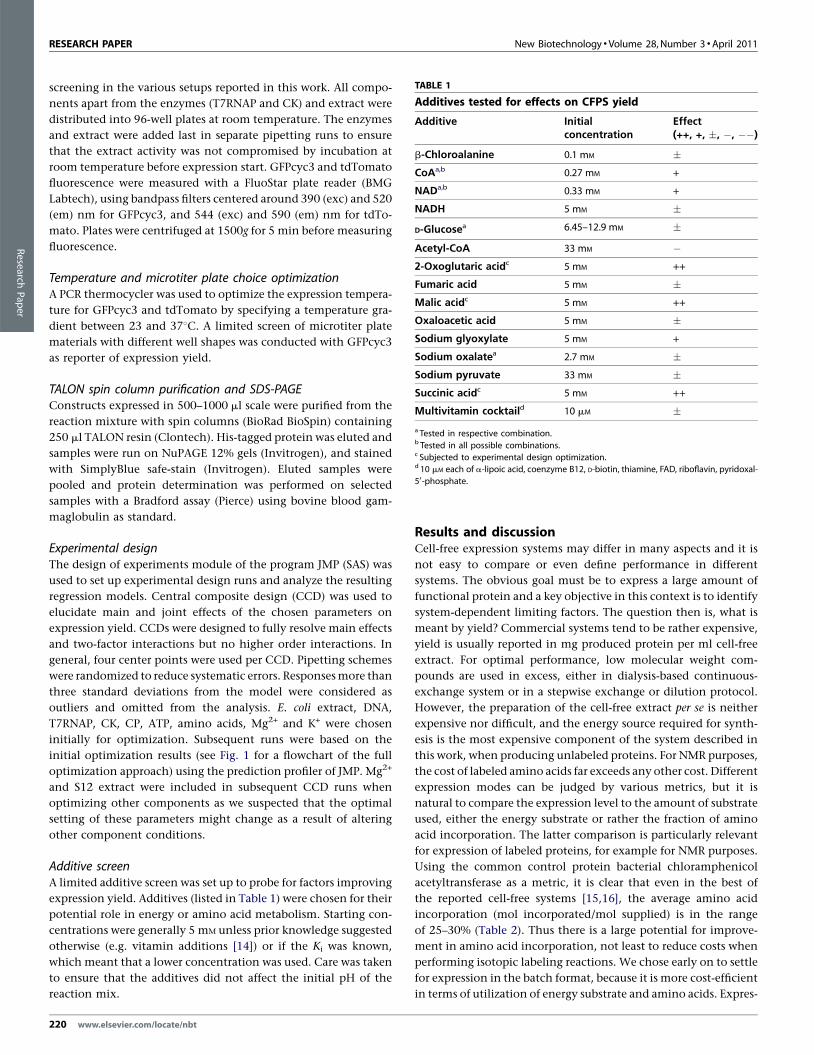
TABLE 1
Additives tested for effects on CFPS yield
Additive Initialconcentration
Effect(++, +, �, �, ��)
b-Chloroalanine 0.1 mM �CoAa,b 0.27 mM +
NADa,b 0.33 mM +
NADH 5 mM �
D-Glucosea 6.45–12.9 mM �
Acetyl-CoA 33 mM �2-Oxoglutaric acidc 5 mM ++
Fumaric acid 5 mM �Malic acidc 5 mM ++
Oxaloacetic acid 5 mM �Sodium glyoxylate 5 mM +
Sodium oxalatea 2.7 mM �Sodium pyruvate 33 mM �Succinic acidc 5 mM ++
Multivitamin cocktaild 10 mM �a Tested in respective combination.b Tested in all possible combinations.c Subjected to experimental design optimization.d 10 mM each of a-lipoic acid, coenzyme B12, D-biotin, thiamine, FAD, riboflavin, pyridoxal-
50-phosphate.
Research
Pap
er
screening in the various setups reported in this work. All compo-
nents apart from the enzymes (T7RNAP and CK) and extract were
distributed into 96-well plates at room temperature. The enzymes
and extract were added last in separate pipetting runs to ensure
that the extract activity was not compromised by incubation at
room temperature before expression start. GFPcyc3 and tdTomato
fluorescence were measured with a FluoStar plate reader (BMG
Labtech), using bandpass filters centered around 390 (exc) and 520
(em) nm for GFPcyc3, and 544 (exc) and 590 (em) nm for tdTo-
mato. Plates were centrifuged at 1500g for 5 min before measuring
fluorescence.
Temperature and microtiter plate choice optimizationA PCR thermocycler was used to optimize the expression tempera-
ture for GFPcyc3 and tdTomato by specifying a temperature gra-
dient between 23 and 378C. A limited screen of microtiter plate
materials with different well shapes was conducted with GFPcyc3
as reporter of expression yield.
TALON spin column purification and SDS-PAGEConstructs expressed in 500–1000 ml scale were purified from the
reaction mixture with spin columns (BioRad BioSpin) containing
250 ml TALON resin (Clontech). His-tagged protein was eluted and
samples were run on NuPAGE 12% gels (Invitrogen), and stained
with SimplyBlue safe-stain (Invitrogen). Eluted samples were
pooled and protein determination was performed on selected
samples with a Bradford assay (Pierce) using bovine blood gam-
maglobulin as standard.
Experimental designThe design of experiments module of the program JMP (SAS) was
used to set up experimental design runs and analyze the resulting
regression models. Central composite design (CCD) was used to
elucidate main and joint effects of the chosen parameters on
expression yield. CCDs were designed to fully resolve main effects
and two-factor interactions but no higher order interactions. In
general, four center points were used per CCD. Pipetting schemes
were randomized to reduce systematic errors. Responses more than
three standard deviations from the model were considered as
outliers and omitted from the analysis. E. coli extract, DNA,
T7RNAP, CK, CP, ATP, amino acids, Mg2+ and K+ were chosen
initially for optimization. Subsequent runs were based on the
initial optimization results (see Fig. 1 for a flowchart of the full
optimization approach) using the prediction profiler of JMP. Mg2+
and S12 extract were included in subsequent CCD runs when
optimizing other components as we suspected that the optimal
setting of these parameters might change as a result of altering
other component conditions.
Additive screenA limited additive screen was set up to probe for factors improving
expression yield. Additives (listed in Table 1) were chosen for their
potential role in energy or amino acid metabolism. Starting con-
centrations were generally 5 mM unless prior knowledge suggested
otherwise (e.g. vitamin additions [14]) or if the Ki was known,
which meant that a lower concentration was used. Care was taken
to ensure that the additives did not affect the initial pH of the
reaction mix.
220 www.elsevier.com/locate/nbt
Results and discussionCell-free expression systems may differ in many aspects and it is
not easy to compare or even define performance in different
systems. The obvious goal must be to express a large amount of
functional protein and a key objective in this context is to identify
system-dependent limiting factors. The question then is, what is
meant by yield? Commercial systems tend to be rather expensive,
yield is usually reported in mg produced protein per ml cell-free
extract. For optimal performance, low molecular weight com-
pounds are used in excess, either in dialysis-based continuous-
exchange system or in a stepwise exchange or dilution protocol.
However, the preparation of the cell-free extract per se is neither
expensive nor difficult, and the energy source required for synth-
esis is the most expensive component of the system described in
this work, when producing unlabeled proteins. For NMR purposes,
the cost of labeled amino acids far exceeds any other cost. Different
expression modes can be judged by various metrics, but it is
natural to compare the expression level to the amount of substrate
used, either the energy substrate or rather the fraction of amino
acid incorporation. The latter comparison is particularly relevant
for expression of labeled proteins, for example for NMR purposes.
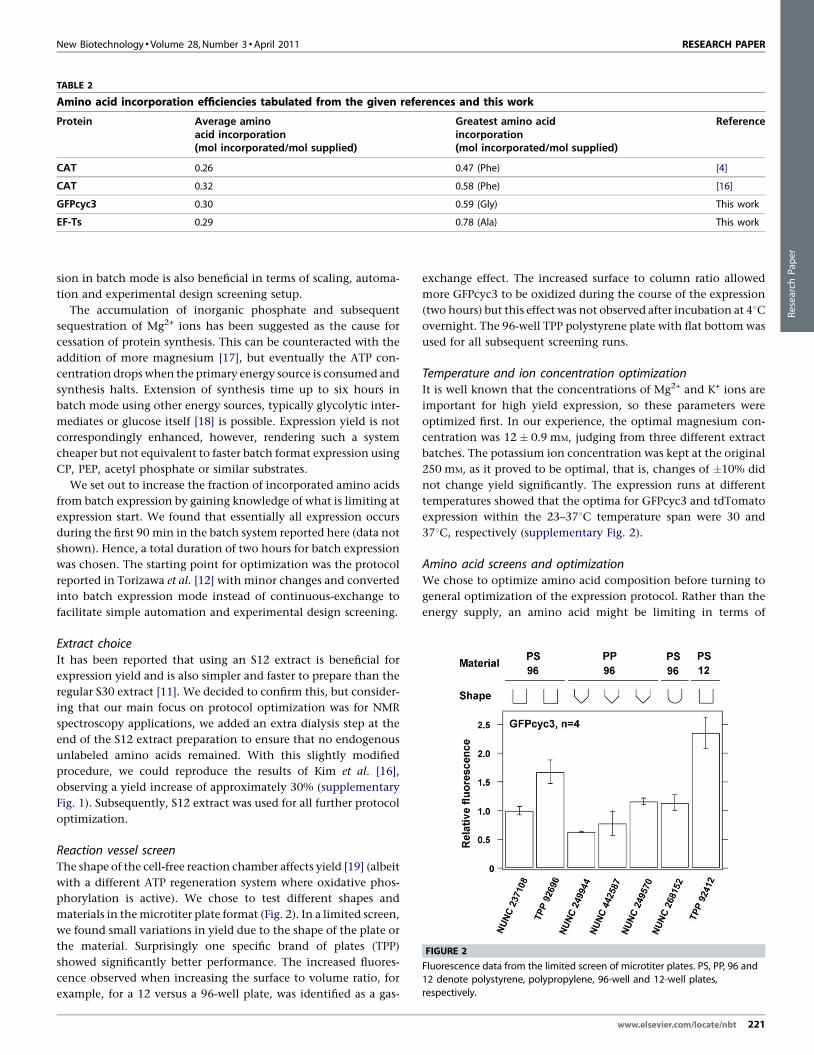
Using the common control protein bacterial chloramphenicol
acetyltransferase as a metric, it is clear that even in the best of
the reported cell-free systems [15,16], the average amino acid
incorporation (mol incorporated/mol supplied) is in the range
of 25–30% (Table 2). Thus there is a large potential for improve-
ment in amino acid incorporation, not least to reduce costs when
performing isotopic labeling reactions. We chose early on to settle
for expression in the batch format, because it is more cost-efficient
in terms of utilization of energy substrate and amino acids. Expres-
New Biotechnology �Volume 28, Number 3 �April 2011 RESEARCH PAPER
TABLE 2
Amino acid incorporation efficiencies tabulated from the given references and this work
Protein Average aminoacid incorporation(mol incorporated/mol supplied)
Greatest amino acidincorporation(mol incorporated/mol supplied)
Reference
CAT 0.26 0.47 (Phe) [4]
CAT 0.32 0.58 (Phe) [16]
GFPcyc3 0.30 0.59 (Gly) This work
EF-Ts 0.29 0.78 (Ala) This work
[(Figure_2)TD$FIG]
FIGURE 2
Fluorescence data from the limited screen of microtiter plates. PS, PP, 96 and
12 denote polystyrene, polypropylene, 96-well and 12-well plates,respectively.
ResearchPap
er
sion in batch mode is also beneficial in terms of scaling, automa-
tion and experimental design screening setup.
The accumulation of inorganic phosphate and subsequent
sequestration of Mg2+ ions has been suggested as the cause for
cessation of protein synthesis. This can be counteracted with the
addition of more magnesium [17], but eventually the ATP con-
centration drops when the primary energy source is consumed and
synthesis halts. Extension of synthesis time up to six hours in
batch mode using other energy sources, typically glycolytic inter-
mediates or glucose itself [18] is possible. Expression yield is not
correspondingly enhanced, however, rendering such a system
cheaper but not equivalent to faster batch format expression using
CP, PEP, acetyl phosphate or similar substrates.
We set out to increase the fraction of incorporated amino acids
from batch expression by gaining knowledge of what is limiting at
expression start. We found that essentially all expression occurs
during the first 90 min in the batch system reported here (data not
shown). Hence, a total duration of two hours for batch expression
was chosen. The starting point for optimization was the protocol
reported in Torizawa et al. [12] with minor changes and converted
into batch expression mode instead of continuous-exchange to
facilitate simple automation and experimental design screening.
Extract choiceIt has been reported that using an S12 extract is beneficial for
expression yield and is also simpler and faster to prepare than the
regular S30 extract [11]. We decided to confirm this, but consider-
ing that our main focus on protocol optimization was for NMR
spectroscopy applications, we added an extra dialysis step at the
end of the S12 extract preparation to ensure that no endogenous
unlabeled amino acids remained. With this slightly modified
procedure, we could reproduce the results of Kim et al. [16],
observing a yield increase of approximately 30% (supplementary
Fig. 1). Subsequently, S12 extract was used for all further protocol
optimization.
Reaction vessel screenThe shape of the cell-free reaction chamber affects yield [19] (albeit
with a different ATP regeneration system where oxidative phos-
phorylation is active). We chose to test different shapes and
materials in the microtiter plate format (Fig. 2). In a limited screen,
we found small variations in yield due to the shape of the plate or
the material. Surprisingly one specific brand of plates (TPP)
showed significantly better performance. The increased fluores-
cence observed when increasing the surface to volume ratio, for
example, for a 12 versus a 96-well plate, was identified as a gas-
exchange effect. The increased surface to column ratio allowed
more GFPcyc3 to be oxidized during the course of the expression
(two hours) but this effect was not observed after incubation at 48Covernight. The 96-well TPP polystyrene plate with flat bottom was
used for all subsequent screening runs.
Temperature and ion concentration optimizationIt is well known that the concentrations of Mg2+ and K+ ions are
important for high yield expression, so these parameters were
optimized first. In our experience, the optimal magnesium con-
centration was 12 � 0.9 mM, judging from three different extract
batches. The potassium ion concentration was kept at the original
250 mM, as it proved to be optimal, that is, changes of �10% did
not change yield significantly. The expression runs at different
temperatures showed that the optima for GFPcyc3 and tdTomato
expression within the 23–378C temperature span were 30 and
378C, respectively (supplementary Fig. 2).
Amino acid screens and optimizationWe chose to optimize amino acid composition before turning to
general optimization of the expression protocol. Rather than the
energy supply, an amino acid might be limiting in terms of
www.elsevier.com/locate/nbt 221
RESEARCH PAPER New Biotechnology �Volume 28, Number 3 �April 2011
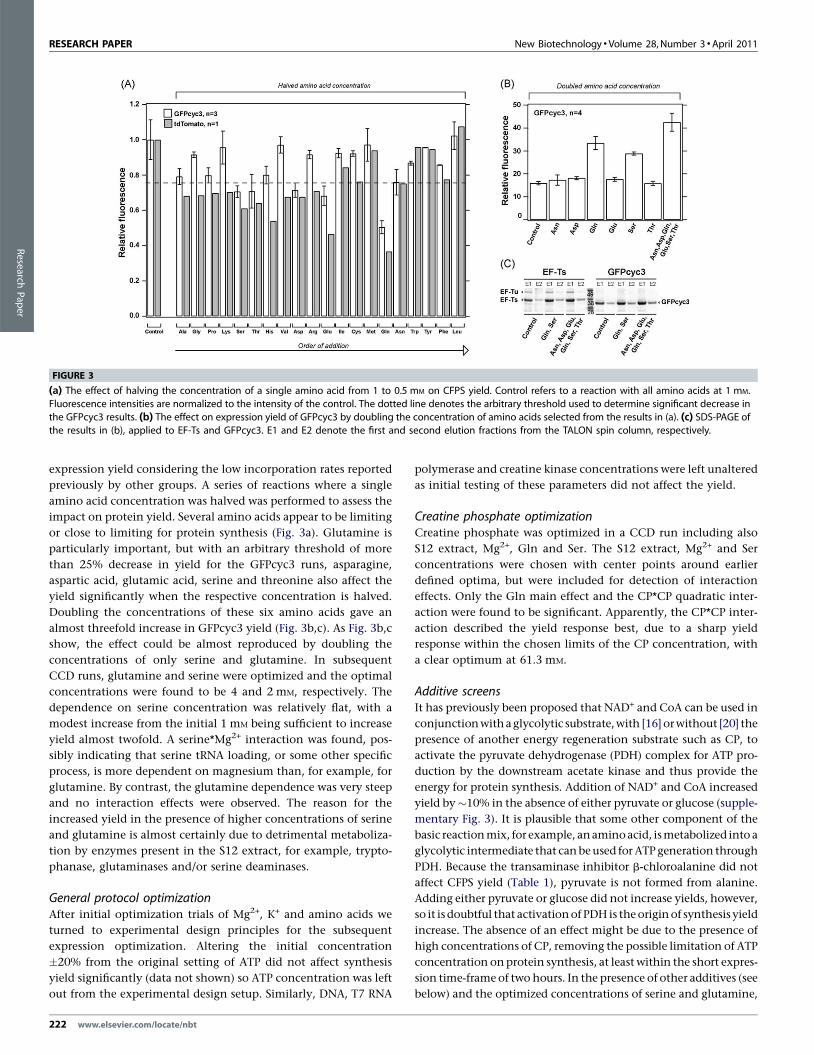
[(Figure_3)TD$FIG]
FIGURE 3
(a) The effect of halving the concentration of a single amino acid from 1 to 0.5 mM on CFPS yield. Control refers to a reaction with all amino acids at 1 mM.Fluorescence intensities are normalized to the intensity of the control. The dotted line denotes the arbitrary threshold used to determine significant decrease in
the GFPcyc3 results. (b) The effect on expression yield of GFPcyc3 by doubling the concentration of amino acids selected from the results in (a). (c) SDS-PAGE ofthe results in (b), applied to EF-Ts and GFPcyc3. E1 and E2 denote the first and second elution fractions from the TALON spin column, respectively.
Research
Pap
er
expression yield considering the low incorporation rates reported
previously by other groups. A series of reactions where a single
amino acid concentration was halved was performed to assess the
impact on protein yield. Several amino acids appear to be limiting
or close to limiting for protein synthesis (Fig. 3a). Glutamine is
particularly important, but with an arbitrary threshold of more
than 25% decrease in yield for the GFPcyc3 runs, asparagine,
aspartic acid, glutamic acid, serine and threonine also affect the
yield significantly when the respective concentration is halved.
Doubling the concentrations of these six amino acids gave an
almost threefold increase in GFPcyc3 yield (Fig. 3b,c). As Fig. 3b,c
show, the effect could be almost reproduced by doubling the
concentrations of only serine and glutamine. In subsequent
CCD runs, glutamine and serine were optimized and the optimal
concentrations were found to be 4 and 2 mM, respectively. The
dependence on serine concentration was relatively flat, with a
modest increase from the initial 1 mM being sufficient to increase
yield almost twofold. A serine*Mg2+ interaction was found, pos-
sibly indicating that serine tRNA loading, or some other specific
process, is more dependent on magnesium than, for example, for
glutamine. By contrast, the glutamine dependence was very steep
and no interaction effects were observed. The reason for the
increased yield in the presence of higher concentrations of serine
and glutamine is almost certainly due to detrimental metaboliza-
tion by enzymes present in the S12 extract, for example, trypto-
phanase, glutaminases and/or serine deaminases.
General protocol optimizationAfter initial optimization trials of Mg2+, K+ and amino acids we
turned to experimental design principles for the subsequent
expression optimization. Altering the initial concentration
�20% from the original setting of ATP did not affect synthesis
yield significantly (data not shown) so ATP concentration was left
out from the experimental design setup. Similarly, DNA, T7 RNA
222 www.elsevier.com/locate/nbt
polymerase and creatine kinase concentrations were left unaltered
as initial testing of these parameters did not affect the yield.
Creatine phosphate optimizationCreatine phosphate was optimized in a CCD run including also
S12 extract, Mg2+, Gln and Ser. The S12 extract, Mg2+ and Ser
concentrations were chosen with center points around earlier
defined optima, but were included for detection of interaction
effects. Only the Gln main effect and the CP*CP quadratic inter-
action were found to be significant. Apparently, the CP*CP inter-
action described the yield response best, due to a sharp yield
response within the chosen limits of the CP concentration, with
a clear optimum at 61.3 mM.
Additive screensIt has previously been proposed that NAD+ and CoA can be used in
conjunction witha glycolytic substrate,with [16]orwithout [20] the
presence of another energy regeneration substrate such as CP, to
activate the pyruvate dehydrogenase (PDH) complex for ATP pro-
duction by the downstream acetate kinase and thus provide the
energy for protein synthesis. Addition of NAD+ and CoA increased
yield by�10% in the absence of either pyruvate or glucose (supple-
mentary Fig. 3). It is plausible that some other component of the
basic reaction mix, for example, an amino acid, is metabolized into a
glycolytic intermediate that can be used for ATP generation through
PDH. Because the transaminase inhibitor b-chloroalanine did not
affect CFPS yield (Table 1), pyruvate is not formed from alanine.
Adding either pyruvate or glucose did not increase yields, however,
so it is doubtful that activation of PDH is the origin of synthesis yield
increase. The absence of an effect might be due to the presence of
high concentrations of CP, removing the possible limitation of ATP
concentration on protein synthesis, at least within the short expres-
sion time-frame of two hours. In the presence of other additives (see
below) and the optimized concentrations of serine and glutamine,
New Biotechnology �Volume 28, Number 3 �April 2011 RESEARCH PAPER
TABLE 3
Results from the Bradford assay of the initial and final protocolyields
Protein Initial protocolyield (mg/ml)
Final protocolyield (mg/ml)
EF-Ts 280 � 36 630 � 21
GFPcyc3 270 � 14 670 � 33
wtGFP 190 � 16 400 � 14
TH dIII 230 � 18 410 � 15
tdTomato 180 � 16 245 � 8
[(Figure_4)TD$FIG]
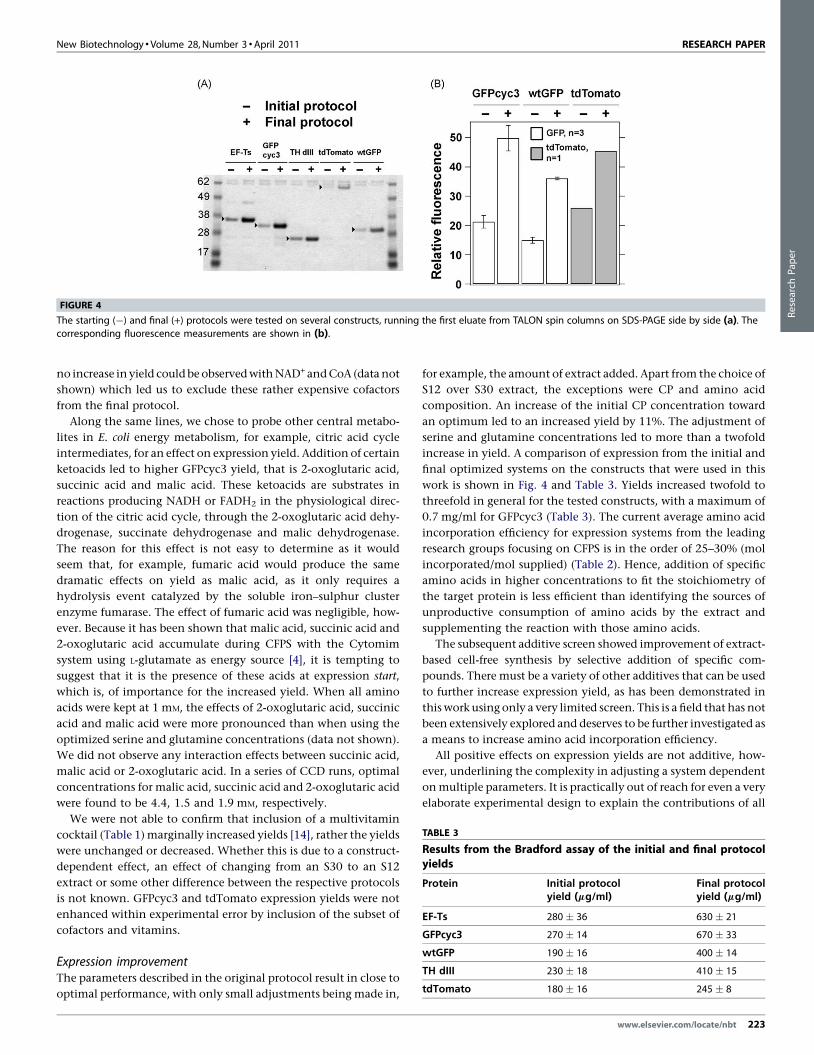
FIGURE 4
The starting (�) and final (+) protocols were tested on several constructs, running the first eluate from TALON spin columns on SDS-PAGE side by side (a). Thecorresponding fluorescence measurements are shown in (b).
ResearchPap
er
no increase in yield could be observed with NAD+ and CoA (data not
shown) which led us to exclude these rather expensive cofactors
from the final protocol.
Along the same lines, we chose to probe other central metabo-
lites in E. coli energy metabolism, for example, citric acid cycle
intermediates, for an effect on expression yield. Addition of certain
ketoacids led to higher GFPcyc3 yield, that is 2-oxoglutaric acid,
succinic acid and malic acid. These ketoacids are substrates in
reactions producing NADH or FADH2 in the physiological direc-
tion of the citric acid cycle, through the 2-oxoglutaric acid dehy-
drogenase, succinate dehydrogenase and malic dehydrogenase.
The reason for this effect is not easy to determine as it would
seem that, for example, fumaric acid would produce the same
dramatic effects on yield as malic acid, as it only requires a
hydrolysis event catalyzed by the soluble iron–sulphur cluster
enzyme fumarase. The effect of fumaric acid was negligible, how-
ever. Because it has been shown that malic acid, succinic acid and
2-oxoglutaric acid accumulate during CFPS with the Cytomim
system using L-glutamate as energy source [4], it is tempting to
suggest that it is the presence of these acids at expression start,
which is, of importance for the increased yield. When all amino
acids were kept at 1 mM, the effects of 2-oxoglutaric acid, succinic
acid and malic acid were more pronounced than when using the
optimized serine and glutamine concentrations (data not shown).
We did not observe any interaction effects between succinic acid,
malic acid or 2-oxoglutaric acid. In a series of CCD runs, optimal
concentrations for malic acid, succinic acid and 2-oxoglutaric acid
were found to be 4.4, 1.5 and 1.9 mM, respectively.
We were not able to confirm that inclusion of a multivitamin
cocktail (Table 1) marginally increased yields [14], rather the yields
were unchanged or decreased. Whether this is due to a construct-
dependent effect, an effect of changing from an S30 to an S12
extract or some other difference between the respective protocols
is not known. GFPcyc3 and tdTomato expression yields were not
enhanced within experimental error by inclusion of the subset of
cofactors and vitamins.
Expression improvementThe parameters described in the original protocol result in close to
optimal performance, with only small adjustments being made in,
for example, the amount of extract added. Apart from the choice of
S12 over S30 extract, the exceptions were CP and amino acid
composition. An increase of the initial CP concentration toward
an optimum led to an increased yield by 11%. The adjustment of
serine and glutamine concentrations led to more than a twofold
increase in yield. A comparison of expression from the initial and
final optimized systems on the constructs that were used in this
work is shown in Fig. 4 and Table 3. Yields increased twofold to
threefold in general for the tested constructs, with a maximum of
0.7 mg/ml for GFPcyc3 (Table 3). The current average amino acid
incorporation efficiency for expression systems from the leading
research groups focusing on CFPS is in the order of 25–30% (mol
incorporated/mol supplied) (Table 2). Hence, addition of specific
amino acids in higher concentrations to fit the stoichiometry of
the target protein is less efficient than identifying the sources of
unproductive consumption of amino acids by the extract and
supplementing the reaction with those amino acids.
The subsequent additive screen showed improvement of extract-
based cell-free synthesis by selective addition of specific com-
pounds. There must be a variety of other additives that can be used
to further increase expression yield, as has been demonstrated in
this work using only a very limited screen. This is a field that has not
been extensively explored and deserves to be further investigated as
a means to increase amino acid incorporation efficiency.
All positive effects on expression yields are not additive, how-
ever, underlining the complexity in adjusting a system dependent
on multiple parameters. It is practically out of reach for even a very
elaborate experimental design to explain the contributions of all
www.elsevier.com/locate/nbt 223

RESEARCH PAPER New Biotechnology �Volume 28, Number 3 �April 2011
Research
Pap
er
involved components at a given time, so a reductionist approach
has to be taken. The parameter choices made in this work were
chosen with the specific aim of reducing cost and increasing
amino acid incorporation, both of which are synonymous as
isotopically labeled amino acids by far constitute the most expen-
sive component in NMR sample production.
ConclusionsWe have applied the experimental design rationale to the task of
improving cell-free protein synthesis. The overall goal of the
optimization was to improve expression yield to make incorpora-
tion of isotopic label more cost-efficient, facilitating the routine
production of NMR samples at costs equivalent to regular in vivo
224 www.elsevier.com/locate/nbt
expression. It is clear from this work that there is a large potential
for increasing expression yield simply by manipulating the amino
acid composition as well as searching for permissible additives.
AcknowledgementsWe would like to acknowledge Jean-Michel Betton and Cordula
Jany for providing the GFP cycle 3 vector and vector sequence,
respectively, and Roger Y. Tsien for providing the tdTomato
vector. This project was funded by the Swedish Research Council.
Appendix A. Supplementary dataSupplementary data associated with this article can be found, in
the online version, at doi:10.1016/j.nbt.2010.06.015.
References
1 Wu, P.S. et al. (2007) Cell-free transcription/translation from PCR-amplified DNA
for high-throughput NMR studies. Angew. Chem. Int. Ed. Engl. 46, 3356–3358
2 Endo, Y. and Sawasaki, T. (2005) Advances in genome-wide protein expression
using the wheat germ cell-free system. Methods Mol. Biol. 310, 145–167
3 Ryabova, L. et al. (2008) A quick in vitro pathway from prokaryotic genomic
libraries to enzyme discovery. Biotechniques 45, 63–68
4 Jewett, M.C. et al. (2008) An integrated cell-free metabolic platform for protein
production and synthetic biology. Mol. Syst. Biol. 4, 220
5 Crameri, A. et al. (1996) Improved green fluorescent protein by molecular
evolution using DNA shuffling. Nat. Biotechnol. 14, 315–319
6 Shaner, N.C. et al. (2004) Improved monomeric red, orange and yellow fluorescent
proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 22,
1567–1572
7 Davanloo, P. et al. (1984) Cloning and expression of the gene for bacteriophage T7
RNA polymerase. Proc. Natl. Acad. Sci. U. S. A. 81, 2035–2039
8 Grodberg, J. and Dunn, J.J. (1988) ompT encodes the Escherichia coli outer
membrane protease that cleaves T7 RNA polymerase during purification. J.
Bacteriol. 170, 1245–1253
9 Kim, R.G. and Choi, C.Y. (2000) Expression-independent consumption of substrates
in cell-free expression system from Escherichia coli. J. Biotechnol. 84, 27–32
10 Liu, D.V. et al. (2005) Streamlining Escherichia coli S30 extract preparation for
economical cell-free protein synthesis. Biotechnol. Prog. 21, 460–465
11 Kim, T.W. et al. (2006) Simple procedures for the construction of a robust and cost-
effective cell-free protein synthesis system. J. Biotechnol. 126, 554–561
12 Torizawa, T. et al. (2005) Efficient production of isotopically labeled proteins by
cell-free synthesis: a practical protocol. J. Biomol. NMR 30, 311–325
13 Matsuda, T. et al. (2007) Improving cell-free protein synthesis for stable-isotope
labeling. J. Biomol. NMR 37, 225–229
14 Woodrow, K.A. et al. (2006) Rapid expression of functional genomic libraries. J.
Proteome Res. 5, 3288–3300
15 Calhoun, K.A. and Swartz, J.R. (2006) Total amino acid stabilization during cell-
free protein synthesis reactions. J. Biotechnol. 123, 193–203
16 Kim, T.W. et al. (2007) Prolonged cell-free protein synthesis using dual energy
sources: combined use of creatine phosphate and glucose for the efficient supply of
ATP and retarded accumulation of phosphate. Biotechnol. Bioeng. 97, 1510–1515
17 Kim, T.W. et al. (2006) Rapid production of milligram quantities of proteins in a
batch cell-free protein synthesis system. J. Biotechnol. 124, 373–380
18 Calhoun, K.A. and Swartz, J.R. (2005) Energizing cell-free protein synthesis with
glucose metabolism. Biotechnol. Bioeng. 90, 606–613
19 Voloshin, A.M. and Swartz, J.R. (2005) Efficient and scalable method for scaling up
cell free protein synthesis in batch mode. Biotechnol. Bioeng. 91, 516–521
20 Calhoun, K.A. and Swartz, J.R. (2005) An economical method for cell-free protein
synthesis using glucose and nucleoside monophosphates. Biotechnol. Prog. 21,
1146–1153