Embed Size (px)
Citation preview

Developmental Science. 2017;e12523. wileyonlinelibrary.com/journal/desc | 1 of 14https://doi.org/10.1111/desc.12523
© 2017 John Wiley & Sons Ltd
Received:26April2016 | Accepted:23September2016DOI:10.1111/desc.12523
P A P E R
Reach tracking reveals dissociable processes underlying inhibitory control in 5- to 10- year- olds and adults
Christopher D. Erb1 | Jeff Moher2,3 | Joo-Hyun Song3,4 | David M. Sobel3
1DepartmentofPsychology,UniversityofNorthCarolinaatGreensboro,Greensboro,NorthCarolina,USA2PsychologyDepartment,WilliamsCollege,Williamstown,Massachusetts,USA3DepartmentofCognitive,Linguistic,andPsychologicalSciences,BrownUniversity,Providence,RhodeIsland,USA4BrownInstituteforBrainScience,BrownUniversity,RhodeIsland,USA
CorrespondenceChristopherD.Erb,DepartmentofPsychology,UniversityofNorthCarolinaatGreensboro,296EberhartBuilding,Greensboro,NC27412,USA.Email:[email protected]
Funding informationNationalScienceFoundation,Grant/AwardNumber:1223777;NationalInstituteofGeneralMedicalSciences,Grant/AwardNumber:P20GM103645
AbstractResearchershaveproposedthattwoprocessesfeaturingdistincttypesofinhibitionsupportinhibitorycontrol:aresponse threshold adjustment processinvolvingtheglobalinhibitionofmotoroutputandaconflict resolution processinvolvingcompetitiveinhibi-tionamongco-activeresponsealternatives.Totargetthedevelopmentofthesepro-cesses,wemeasuredthereachingbehaviorof5-to10-year-olds(Experiment1)andadults(Experiment2)astheyperformedanEriksenflankertask.Thismethodprovidedtwokeymeasures:initiation time(thetimeelapsedbetweenstimulusonsetandmove-mentonset)and reach curvature (thedegreetowhichamovementdeviates fromadirect path to the selected target).We suggest that initiationtime reflects the re-sponsethresholdadjustmentprocessbyindexingthedegreeofmotoricstoppingex-periencedbeforeamovement isstarted,whilereachcurvaturereflectstheconflictresolutionprocessbyindexingthedegreeofco-activationbetweenresponsealterna-tivesoverthecourseofamovement.Ourresultssupportthisclaim,revealingdifferentpatternseffectsininitiationtimeandcurvature,anddivergentdevelopmentaltrajec-toriesbetweenchildhoodandadulthood.Thesefindingsprovidebehavioralevidenceforthedissociationbetweenglobalandcompetitiveinhibition,andoffernewinsightintothedevelopmentofinhibitorycontrol.
RESEARCH HIGHLIGHTS
• Twoofthemeasuresaffordedbyreachtracking,initiationtimeandcurvature,revealeddistinctpatternsoftrialsequenceeffectsintheEriksenflankertaskinchildren5to10yearsofageandadults.Weproposethatthepatternofeffectsobservedin initiationtimere-flectsaresponsethresholdadjustmentprocessinvolvingtheglobalinhibitionofmotoroutput,whilethepatternobservedinreachcur-vaturereflectsaconflict resolutionprocess involvingcompetitiveinhibitionamongco-activeresponsealternatives.
• Initiationtimeandreachcurvaturerevealedsimilargainsininhibi-torycontrolbetween5and10yearsofage,butonlycurvaturere-vealedperformancegainsbetween8-to10-year-oldsandadults.
• Thegainsobservedbetweenchildhoodandadulthoodweredrivenbya specific subsetof trials, suggesting that age-relatedgains ininhibitorycontrolhavebeenmischaracterizedinthepast.
• Theseresultspresenta frameworkfor futureresearchtoexplorehowthe response thresholdadjustmentprocessandconflict res-olution process contribute to individual and group differences ininhibitorycontrol.
1 | INTRODUCTION
Inordertobehaveinaflexible,adaptivemanner,childrenandadultsmustbeabletosuppresshabitualorprepotentresponsesselectively.This capacity, known as inhibitory control, undergoes a protracteddevelopment, improvingrapidlyduringearlychildhoodandreachingitspeakduring lateadolescenceoradulthood (e.g.,Carver,Livesey,& Charles, 2001; Davidson, Amso, Anderson, & Diamond, 2006;Diamond,2002;Luna,2009;Luna,Garver,Urban,Lazar,&Sweeney,2004;Waszak, Li,&Hommel, 2010; Zelazo etal., 2013). Inhibitory

2 of 14 | ERB Et al .
controlhasbeenlinkedtoarangeofsocialandcognitivecapacitiesduringchildhood,includingtheoryofmind(Carlson,Moses,&Breton,2002), emotion regulation (Carlson&Wang,2007), andearlymathandreadingability(Blair&Razza,2007).Individualdifferencesinin-hibitorycontrolhavealsobeenlinkedtoimportantoutcomesinadult-hood,includingsuccessinschoolandatworkandlevelsofmentalandphysicalhealth(seeDiamond,2013,forareview).
While theterm inhibition isoftenusedtorefer toaunitarypro-cessorcapacity,agrowingbodyofresearchindicatesthatinhibitorycontrolissupportedbyanumberofdissociableprocessesthatfeaturedistincttypesof inhibition (forareview,seeMunakataetal.,2011).Given the important role that inhibitory control plays in supportingadaptivethoughtandbehavioracrossthelifespan,akeychallengefac-ingresearchersistoidentifyhowthesedissociableprocessesfunctionat different points in development.However, it is difficult to targettheseprocesseswithtraditionalbehavioralmethods,asaccuracyandresponsetimeprovidelimitedinsightintohowdifferentprocessesun-fold leadinguptoaresponse (Song&Nakayama,2009).Toaddressthis limitation, we use a technique known as reach tracking (e.g.,Diedrich,Thelen,Smith,&Corbetta,2000;Song&Nakayama,2007)totargethowtwokeyprocessesunderlyinginhibitorycontrol–are-sponse threshold adjustment process and a conflict resolution process–arereflectedinparticipants’handmovementsastheyreachtotoucharesponsetarget.
Tooutlineourargument,wedescribeaprominentmodelofin-hibitory control (Botvinick, Braver, Barch, Carter, & Cohen, 2001;Cohen, Dunbar, & McClelland, 1990; Cohen & Huston, 1994;Shenhav, Botvinick,&Cohen, 2013) to contextualize how the re-sponse threshold adjustment process and conflict resolution pro-cess function during performance of the Eriksen flanker task(Eriksen&Eriksen,1974).Wethenreviewevidenceindicatingthattheseprocesses(a)featuredifferenttypesofinhibitionand(b)gen-eratedifferentpatternsof trial sequence effects – effects inwhichqualitiesofaprevioustrail(e.g.,trialn-1)influenceperformanceonthe current trial (trial n) (see e.g., Egner, 2007). Finally, followingrecentresearchusingreachtrackingwithadults(Erb,Moher,Sobel&Song,2016),wepropose that twoof themeasuresaffordedbyreachtracking–initiation time and curvature–canbeusedtotargethoweachoftheseprocessescontributestoinhibitorycontrolacrossthelifespan.
1.1 | Inhibitory control and the flanker task
TheEriksenflankertask(Eriksen&Eriksen,1974)isoneofthemostwidelyusedmeasuresofinhibitorycontrol,andwasrecentlystandard-izedforuseintheNIHToolbox(seeZelazoetal.,2013).Inthetask,participantsidentifyacentrallypresentedtargetstimulusinthepres-ence of distracting stimuli known as ‘flankers’. On congruent trials,thetargetanddistractorscuethesameresponse (e.g.,)andtheneedforinhibitorycontrolisminimal.Onincongruenttrials,thestimulicuecompetingresponses(e.g.,)andinhibitorycontrol isrequiredtooverridetheprepotentresponsegeneratedbythedistractors.Acongruency effect is typicallyobserved in the task,
withhighererrorratesandresponsetimesonincongruentrelativetocongruenttrials.
Contemporarymodelsofinhibitorycontrolproposethatstimuliintheflankertaskareprocessedalongtwodifferentpathways:anau-tomatic pathway that is sensitive to the overall stimulus array, anda control-demanding pathway that can be directed to focus on thecentermost element in the array (e.g., Botvinick etal., 2001;Cohenetal., 1990;DeJong, Liang,& Lauber, 1994; Ridderinkhof, van derMolen,&Bashore,1995;Shenhavetal., 2013).On incongruent tri-als, these pathways generate competing response activations, andtheresultingconflictengagesthreekeyprocesses.First,amonitoringprocess linked to the dorsal anterior cingulate cortex (dACC) regis-tersconflictbetweenthecompetingresponseactivationsgeneratedby the automaticand control-demandingpathways (Botvinick etal.,2001;Yeung,Botvinick,&Cohen,2004).Second,aresponsethresholdadjustmentprocesstemporarilyinhibitsmotoroutputinresponsetosignalsofconflictfromthedACC(Cavanaghetal.,2011;Frank,2006;Munakataetal.,2011;Shenhavetal.,2013;Wiecki&Frank,2013).Thisprocessisproposedtohelpbalancespeed–accuracytrade-offsbytemporarilyhaltingresponding.Thisinturnallowsadditionaltimeforathirdconflictresolutionprocessassociatedwiththelateralprefrontalcortex(LPFC)toswayactivationinfavoroftheappropriateresponseby providing top-down support to the control-demanding pathway(Shenhavetal.,2013).
Twodistincttypesofinhibitioncanbeidentifiedwithinthismodelof inhibitorycontrol (Munakataetal.,2011).Global inhibition occurs in the response threshold adjustment processwhen signals of con-flict fromthedACC leadtothedirectsuppressionofmotoroutput.Competitive inhibition, on the other hand, occurs during the con-flict resolution process, when increased activity along the control-demandingpathwaysuppressesactivity in thecompetingautomaticpathwaythroughlateralinhibitoryconnections.
Theresponsethresholdadjustmentprocessandconflictresolutionprocesshavealsobeenlinkedtodifferentpatternsoftrialsequenceeffects.Forexample,Shethandcolleagues(2012)usedsingle-unitre-cordingstomeasureactivityinthedACCwhileadultparticipantsper-formedaStroop-likeinterferencetask.Theyobservedmaineffectsofbothcurrentandprevioustrialcongruency,resultinginthefollowingpatternofeffectsinthemagnitudeofdACCactivation:cC<iC<cI<iI(where lowercase letters denote previous trial congruency and up-percase lettersdenotecurrenttrialcongruency).Thispatternoftrialsequence effects has since been suggested to reflect the responsethresholdadjustmentprocess(Erbetal.,2016;Shenhavetal.,2013).Onthisview,responsethresholdsareadjustedoneachtrial,within-congruenttrialsincreasingone’sresponsethresholdfromitspreviouspositionandcongruenttrialsdecreasingit(C<I).Trialsprecededbyanincongruenttrialwillthereforetendtofeaturehigherresponsethresh-olds,while thoseprecededbya congruent trialwill tend to featurelowerthresholds(c<i).
While single-unit recordings of the dACC suggest that the re-sponsethresholdadjustmentprocess issensitiveto thecongruencyof both the current and previous trial (cC<iC<cI<iI), responsetimesintheflankertaskindicatethattheconflictresolutionprocess

| 3 of 14ERB Et al .
isinfluencedbyadifferentpatternoftrialsequenceeffects.Earlyre-search investigating trial sequenceeffects in theflanker task foundthatresponsetimesonincongruenttrialswerefasterwhenthepre-cedingtrialwasincongruent(iI trials)asopposedtocongruent(cI trials)(e.g.,Gratton,Coles,&Donchin,1992).Accordingtotheconflictadap-tationaccountofthisfinding,responsetimesarefasteroniIrelativetocItrialsbecausetop-downresourcesaremorelikelytohavebeenrecentlyrecruitedontrialsprecededbyanincongruenttrial(forare-view,seeEgner,2007).
The conflict adaptation account of flanker task performance hasbeencalledintoquestion,however,byresearchindicatingthatthere-sponsetimedifferencebetweeniIandcItrials isdrivenbyaspecificsubsetofresponses;namely,thosefeaturingarepeatoftheprevioustrial’sresponse(Mayr,Awh,&Laurey,2003;Nieuwenhuisetal.,2006).ThesestudiesfoundthatresponsetimesweresignificantlylongeroncI-rthaniI-rtrials(where‘-r’indicatesaresponserepeat)butnotcI-crelativetoiI-ctrials(where‘-c’indicatesaresponsechange).Accordingtothefeatureintegrationaccountofthesefindings,elevatedresponsetimesoncI-r trials reflectastimulus–response (S–R)bindingconflictthatoccurswhentheS–Rpairformedontheprevioustrial(e.g.,stim-ulus = , response=right) interfereswiththeformationofa newS–Rpair on the current trial (e.g., stimulus=, re-sponse=right)(Egner,2007;Hommel,2004;Nieuwenhuisetal.,2006).
Onthisview,activatingonememberof theS–Rpair formedontheprevioustrialleadstotheautomaticactivationoftheothermem-ber.ThisgeneratesS–RbindingconflictoncI-rtrialsbecausepartici-pantsmustpairtheresponseprovidedontheprevioustrial(e.g.,right)withadifferentstimulus(e.g.,)fromthatoftheprevioustrial(e.g.,).GiventhattheappropriateS–Rpairmustbeformedbeforetop-downsupportcanswayactivationinfavorofthecontrol-demandingpathway, thisS–Rconflictcanbeunderstoodtodelay the conflict resolution process.Although iC-r trials also allowforS–Rbindingconflict,theconflictresolutionprocessisnotrequiredtoselecttheappropriateresponseoncongruenttrials.NosuchS–RbindingconflictoccursoncC-ror iI-r trials,as these trials featurearepeatofbothstimulusandresponse.Similarly,S–Rbindingdoesnotoccuronresponsechangetrialsbecausethesetrialsnecessarilyfea-tureastimuluschange.
1.2 | Measuring different processes of inhibition
Theresultsreviewedabovesuggestthattwoprocessesfeaturingdis-tincttypesofinhibitionanddifferentpatternsoftrialsequenceeffectssupport inhibitory control. To examine this hypothesis directly, Erbandcolleagues(2016)presentedadultparticipantswithreachtrack-ingversionsoftheStroopandflankertasks.Theyproposedthattwoofthemeasuresaffordedbyreachtracking–initiation time(thetimeelapsedbetweenstimulusonsetandmovementonset)andreach cur-vature(thedegreetowhichamovementdeviatesfromadirectpathtotheselectedtarget)–couldbeusedtotargettheresponsethresholdadjustmentprocessandconflictresolutionprocess,respectively.
Previous researchusing reach tracking suggests that thedegreeofcurvatureinaparticipant’sreachmovementreflectshowcoactive
differentresponsesareoverthecourseofatrial,andthatparticipantsroutinely initiateamovementbefore theconflict resolutionprocesshasswayedactivation infavorofaspecificresponse (e.g.,Freeman,Nakayama, & Ambady, 2013; Song & Nakayama, 2007). Erb etal.(2016)predicted that initiationtimecouldbeused to index the re-sponsethresholdadjustmentprocess,ashigherresponsethresholdsshouldleadtolongerperiodsofmotoricstoppingand,consequently,longerinitiationtimes.Theyalsopredictedthatreachcurvaturecouldbe used to target the conflict resolution process,with larger reachcurvatures indicatingthatparticipantsweremorepulled toward theprepotentresponsebeforetheconflictresolutionprocesscouldswayactivationinfavorofthecorrectresponse.
TheresultsofErbetal.(2016)supportedthesepredictions,withinitiationtimes revealing the samepatternof trial sequenceeffectsthatwasobservedinsingle-unitrecordingsofthedACC(Shethetal.,2012)andwaslaterproposedtoreflecttheresponsethresholdadjust-mentprocess(Shenhavetal.,2013):cC<iC<cI<iI.Reachcurvaturesrevealedamaineffectofcurrenttrialcongruency,withlargercurva-tureson incongruent relative tocongruent trials.An interactionbe-tweencurrentandprevioustrialcongruencywasalsoobserved,withsignificantlylargerreachcurvaturesonthesubsetofincongruenttrialsthatallowedforS–Rbindingconflict(cItrials)relativetothosethatdidnot(iItrials).1Thispatternofresultsisconsistentwiththeclaimthatreachcurvaturecanbeusedtoindexhowtheconflictresolutionpro-cessunfoldsoverthecourseofaresponse.Thus,initiationtimeandcurvatureappear tocapturedistinctprocessesunderlying inhibitorycontrolinadults.
1.3 | The current study
ThecurrentstudybuildsonErbetal.(2016)toaddresstwokeydevel-opmentalquestions.First,do the response threshold adjustment process and conflict resolution process make dissociable contributions to flanker task performance during childhood?Iftheseprocessesfunctionsimilarlyinchildrenasinadults,thenchildren’sinitiationtimesintheflankertaskshouldrevealmaineffectsofbothcurrentandprevioustrialcon-gruency,resultinginthefollowingpatternofeffects:cC<iC<cI<iI.Children’sreachcurvaturesshouldbeuniformlylowoncongruenttri-als,elevatedonincongruenttrialsnotfeaturingS–Rbindingconflict(cI-c,iI-c,andiI-rtrials),andlargestonincongruenttrialsfeaturingS–Rbindingconflict(cI-rtrials).
Second,how do these processes contribute to the age-related changes in flanker task performance?Whileanumberofstudieshavefoundthatflanker task performance continues to improve into adulthood (Li,Hämmerer, Müller, Hommel, & Lindenberger, 2009; Waszak etal.,2010), it is unclear the extent towhich the response threshold ad-justmentprocessandconflictresolutionprocesscontributetothesedevelopmentalgains.Theavailableneurophysiologydataindicatethatkeybrainregionsimplicatedinsupportingtheseprocesses(thedACCandLPFC)undergo relativelyprolongeddevelopment (Gogtayetal.,2004;Sowelletal.,2003;Velanova,Wheeler,&Luna,2008),suggest-ingthatbothinitiationtimeandreachcurvaturewillreveallargercon-gruencyeffectsinchildrenthanadults.

4 of 14 | ERB Et al .
Numerouspaststudieshaveinvestigatedinhibitorycontrolusingcontinuous behavioral and psychophysiologicalmeasureswith bothchildren(e.g.,Checa,Castellanos,Abundis-Gutiérrez,&Rueda,2014;Ridderinkhof&vanderMolen,1995;vandeLaar,vandenWildenberg,vanBoxtel,&vanderMolen,2014)andadults (e.g.,Eriksen,Coles,Morris, & O’Hara, 1985; Gratton etal., 1992; van Boxtel, van derMolen, Jennings, & Brunia, 2001). For example, Ridderinkhof andvan der Molen (1995) had 5- to 12-year-olds and adults performtheflankertaskbysqueezingdynamometerswiththeirleftandrighthands,whichallowedtheresearcherstoseparatesqueezeonsetandsqueeze closure. These measures revealed nearly identical gains inperformance,withsignificantreductionsinthesizeofthecongruencyeffect occurring across childhood and no significant improvementsoccurringbetween10-to12-year-oldsandadults.However,trialse-quenceeffectswerenotanalyzedinthestudyand,consequently,itisunclearwhethersqueezeonsetandsqueezeclosurecaptureddiffer-entunderlyingprocesses.
While trial sequence effects in the flanker task have been in-vestigatedindetail inadults (Grattonetal.,1992;Mayretal.,2003;Nieuwenhuis etal., 2006; Schmidt & deHouwer, 2011; Ullsperger,Bylsma, & Botvinick, 2005; Verbruggen, Notebaert, Liefooghe, &Vandierendonck, 2006;Weissman, Jiang,&Egner, 2014), few stud-ies have evaluated these effects in children, and data are particu-larlysparseforchildrenundertheageof10years(e.g.,Cragg,2016;Nieuwenhuis etal., 2006; Stins, Polderman, Boomsma, & de Geus,2007;Takarae,Schmidt,Tassone,&Simon,2009).2Thus,akeygoalofthepresentstudyistoevaluatetheextenttowhichspecifictrialse-quenceeffectscontributetodevelopmentalgainsinflankertaskper-formance.GiventhatincongruenttrialsfeaturingS–Rbindingconflictplacegreater–andpossiblydifferent–demandsoninhibitorycontrolrelativetoothertrials,weexpectthatage-relatedgainsininhibitorycontrolwillbedriveninlargepartbythesetrials.
2 | EXPERIMENT 1
Five-to10-year-oldscompletedachild-friendlyversionoftheflankertaskby reaching to touch target locationsonadigitaldisplaywhilewearinga smallmotion-tracking sensoron their indexfinger. If theresponse threshold adjustment process and conflict resolutionpro-cessfunctionsimilarlyinchildrenasintheadultsfeaturedinErbetal.(2016), thenchildren’s initiationtimesshouldrevealmaineffectsofcurrentandprevious trial congruency,whilechildren’s reachcurva-turesshouldbeuniformlylowoncongruenttrials,elevatedonincon-gruenttrials,andgreatestonincongruenttrialsfeaturingS–Rbindingconflict (cI-r trials). Further, if these processes undergo significantdevelopment duringmiddle childhood, then the congruency effectsobservedininitiationtimeandcurvatureshoulddecreasewithage.
2.1 | Method
2.1.1 | Participants
Sixty right-handedchildren (M = 91.4months,SD=19.6months;33females)withnormalreachingbehaviorandnormalorcorrected-to-normalvisionparticipatedinthestudy,with12childrenineachoffiveagegroups(5-,6-,7-,8-and9-to10-year-olds).Theaverageageinmonthsofeachgroupwasasfollows:65.0(SD=3.7),79.0(SD=3.0),90.1 (SD=3.8), 101.6 (SD=2.7), and 120.5 (SD=6.6). Participantswererecruited froma listofhospitalbirthsor throughcontactatalocal children’smuseum.All participantswere tested in the labora-toryonBrownUniversity’scampusandreceivedasmallprizefortheirparticipation.TheInstitutionalReviewBoardatBrownUniversityap-provedtheprotocol.
2.1.2 | Materials
The experiment was conducted using a rear-mounted projector todisplay the taskonaPlexiglas screen thatwasarrangeduprightonatableapproximately48cminfrontoftheparticipant.ParticipantsinitiatedallmovementsfromaStyrofoamstartingblock(2×2×2cm)located27cminfrontofthecenterofthescreen.Reachmovementsand response selectionsweremeasured at a rate of approximately160Hzwith an electromagnetic position and orientation recordingsystem(Liberty,Polhemus).Handpositionwasmeasuredwithasmallmotion-trackingmarker(2.26cmlong,1.27cmwide,and1.14high)weighing0.13ouncesthatwassecuredtotheparticipant’srightindexfinger.ThetaskwasprogrammedinMATLAB(Mathworks).
2.1.3 | Procedure
Participantswerepresentedwithachild-friendlyversionoftheflankertaskfeaturingyellowcartoonfish(adaptedfromRuedaetal.,2004;seeFigure1).Childrenidentifiedwhichdirectionthefishinthecenterofthestimulusarraywasfacingbytouchingoneoftwopiecesoffishfood(orangecircles,2cmindiameter)locatedtowardthetopleftorrightofthescreen.Eachfishwas1.5cmatitstallestand3cmlong.
Duringeachtrial,acrosshairappeared1secondbeforethestimu-lusarray.Thecuewaslocatedinthesamelocationinwhichthecentraltargetappeared,minimizingthedemandsplacedonvisualsearch(seeFigure2).Atrialwouldnotinitiateuntilthechild’sfingerwasrestingontheStyrofoamstartingblockfor1second.Ifthechild’shandmovedfromthislocationbeforestimulusonset,thetaskwaspausedanddidnot resumeuntil thechild returned theirhand to the startingblockfor 1second. Children had up to 10seconds to respond followingstimulusonset.Ahightonesoundedforcorrectresponsesprovidedintheallottedtime(600Hzfor200milliseconds).Alowtonesounded
F IGURE 1 Exampleofanincongruentstimulusarrayintheflankertask

| 5 of 14ERB Et al .
forincorrectresponsesorresponsesthatexceededtheallottedtime(300Hzfor200milliseconds).
Childrenfirstcompleteda9-pointcalibrationsequencefollowedby16baselinetrialsthatrequiredreachingtoanimageatthetopleftorrightofthescreen.Participantsthenreceivedapracticeblockof10flankertrialsbeforebeginningtheexperiment.Theexperimentcon-sistedofthreeblocksoftrials.Eachblockfeaturedfourneutraltrialsinwhichthecentraltargetappearedwithoutflankers,16congruenttrialsinwhichthecentraltargetandflankerscuedthesameresponse,and 16 incongruent trials in which the central target and flankerscuedopposingresponses (36trialstotal).Halfofthecongruentandincongruenttrialsfeaturedtwoflankerswhiletheotherhalffeaturedfour(seeSection1oftheSupportingInformationforresultsrelatingto thismanipulation).Participantswere remindedbeforeeachblocktoperformthetaskquickly.Threechildren(two5-year-oldsandone7-year-old)declinedtocompletethethirdblockoftrials.
2.1.4 | Data processing
The processing procedures used in the current study were largelyadapted fromMoher and Song (2013). Three-dimensional resultantspeedscalarswerecreatedforeachtrialusingadifferentiationproce-dureinMATLAB.Thesescalarswerethensubmittedtoasecond-order,low-passButterworthfilterwithacut-offof10Hz.Movementonsetwascalculatedasthefirstpointoneachtrialafterstimulusonsetatwhichhandmovementspeedexceeded25.4cm/s.Eachindividualtrialwasvisuallyinspectedasinpreviouswork(Song&Nakayama,2006,2007, 2008); for trials inwhich thedefault threshold clearlymissedpartofthemovementorincludedsubstantialmovementbacktothestarting point, thresholds were adjusted manually. Manual adjust-mentsweremosttypicallyrequiredwhenparticipants(a)rapidlypulledtheirfingerawayfromthescreenafterhavingtouchedatargetor(b)stoppedentirelyduringtheirmovement(e.g.,afterrealizingthattheyhadbeenmovingtowardtheincorrecttarget).Anaverageof11.54%(SD =11.13%)oftrialswereadjustedmanuallyforeachparticipant.
Trajectories for calculating curvature were measured in two-dimensional xy spacebycalculatinga line fromthestart to theendpoint of themovement, andmeasuring theorthogonal deviationofthe actualmovement from that line at each sample.Curvaturewas
definedasthemaximumpointofdeviationincentimetersdividedbythelengthofthelinefromthestarttotheendpointsofthemovementincentimeters (followingDesmurget,Jordan,Prablanc,&Jeannerod,1997;Moher&Song,2013).
2.2 | Results
The first trial of each block was excluded from analysis. Averageerror rate was analyzed with a 2 (Current Trial Congruency: C orI)×2(PreviousTrialCongruency:cori)×2(ResponseType:changeorrepeat)×5(AgeGroup:5,6,7,8,or9–10yearsofage)ANOVA.Averageinitiationtimeandcurvaturewerecalculatedforallaccuratetrials that were not preceded by an error andwere then analyzedvia2×2×2×5ANOVAsof the formdescribedabove.Preliminaryanalysesrevealednodifferencebetweenneutralandcongruenttrialsand,consequently,neutraltrialswereincludedascongruenttrialsinthe following analyses. In addition to initiationtime and curvature,movementtime(thetimeelapsedbetweenstimulusonsetandmove-mentonset)andtotaltime(thetimeelapsedbetweenstimulusonsetandresponsecompletion)werealsomeasured.TheresultsofthesemeasuresarepresentedinSection1oftheSupportingInformation.
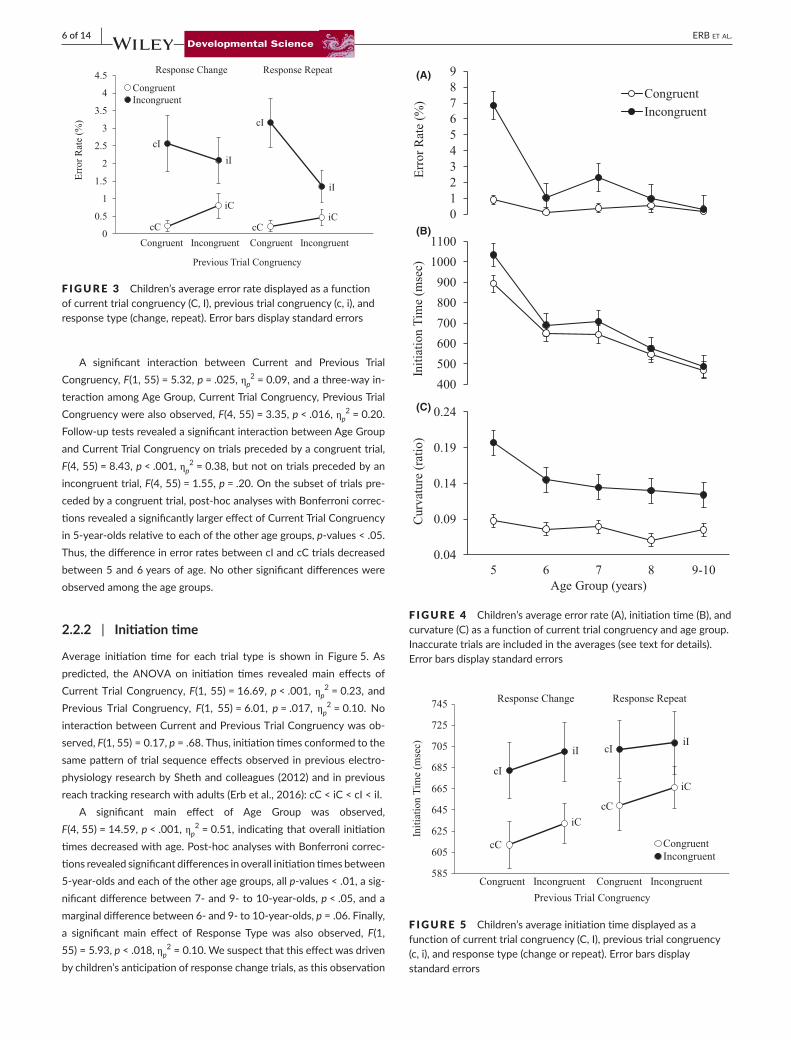
2.2.1 | Error rate
TheaverageerrorrateforeachtrialtypeisshowninFigure3.Giventhe clear floor effects on congruent trials and unequal variancesamong the trial types, we note that the following analyses shouldbeinterpretedwithcaution.SignificantmaineffectsofCurrentTrialCongruency, F(1,55)=20.96, p < .001, ηp
2=0.28, and Age Groupwereobserved,F(4,55)=9.57,p < .001,ηp
2=0.41.Post-hocanaly-ses with Bonferroni corrections revealed significant differences inoverall error rates between 5-year-olds and each of the other agegroups, all p-values<.01 (see Figure4A). A significant interactionbetweenCurrentTrialCongruencyandAgeGroupwasalsoobserved,F(4,55)=6.80,p < .001,ηp
2=0.33.Post-hocanalyseswithBonferronicorrectionsrevealedthattheeffectofCurrentTrialCongruencywassignificantlylargerin5-year-oldsthanineachoftheotheragegroups,p-values<.05. None of the other age groups differed significantlyfromoneanother.
F IGURE 2 IllustrationofacongruenttrialintheflankertaskpresentedinExperiment1.Participantswereinstructedtorespondbytouchingthetargetlocationcuedbythecentermostfishinthestimulusarray
+
6 of 14 | ERB Et al .
A significant interaction between Current and Previous TrialCongruency,F(1,55)=5.32,p = .025,ηp
2=0.09,andathree-wayin-teractionamongAgeGroup,CurrentTrialCongruency,PreviousTrialCongruencywerealsoobserved,F(4,55)=3.35,p < .016,ηp
2 = 0.20. Follow-uptestsrevealedasignificantinteractionbetweenAgeGroupandCurrentTrialCongruencyontrialsprecededbyacongruenttrial,F(4,55)=8.43,p < .001,ηp
2=0.38,butnotontrialsprecededbyanincongruenttrial,F(4,55)=1.55,p = .20.Onthesubsetoftrialspre-cededbyacongruenttrial,post-hocanalyseswithBonferronicorrec-tionsrevealedasignificantlylargereffectofCurrentTrialCongruencyin5-year-oldsrelativetoeachoftheotheragegroups,p-values<.05.Thus,thedifferenceinerrorratesbetweencIandcCtrialsdecreasedbetween5and6yearsofage.Noothersignificantdifferenceswereobservedamongtheagegroups.
2.2.2 | Initiation time
Average initiation time for each trial type is shown in Figure5. Aspredicted, the ANOVA on initiation times revealedmain effects ofCurrentTrialCongruency,F(1,55)=16.69,p < .001,ηp
2=0.23,andPrevious Trial Congruency, F(1, 55)=6.01, p = .017, ηp
2 = 0.10. No interactionbetweenCurrentandPreviousTrialCongruencywasob-served,F(1,55)=0.17,p = .68.Thus,initiationtimesconformedtothesamepatternoftrialsequenceeffectsobservedinpreviouselectro-physiologyresearchbyShethandcolleagues(2012)andinpreviousreachtrackingresearchwithadults(Erbetal.,2016):cC<iC<cI<iI.
A significant main effect of Age Group was observed,F(4,55)=14.59,p < .001,ηp
2=0.51,indicatingthatoverall initiationtimesdecreasedwithage.Post-hocanalyseswithBonferronicorrec-tionsrevealedsignificantdifferencesinoverallinitiationtimesbetween5-year-oldsandeachoftheotheragegroups,allp-values<.01,asig-nificantdifferencebetween7-and9-to10-year-olds,p < .05,andamarginaldifferencebetween6-and9-to10-year-olds,p = .06.Finally,a significant main effect of Response Typewas also observed, F(1,55)=5.93,p < .018,ηp
2=0.10.Wesuspectthatthiseffectwasdrivenbychildren’santicipationofresponsechangetrials,asthisobservation
F IGURE 4 Children’saverageerrorrate(A),initiationtime(B),andcurvature(C)asafunctionofcurrenttrialcongruencyandagegroup.Inaccuratetrialsareincludedintheaverages(seetextfordetails).Errorbarsdisplaystandarderrors
0123456789
Erro
r Rat
e (%
) CongruentIncongruent
400500600700800900
10001100
Initi
atio
n Ti
me
(mse
c)
0.04
0.09
0.14
0.19
0.24
5 6 7 8 9-10
Cur
vatu
re (r
atio
)
Age Group (years)
(B)
(C)
(A)
F IGURE 5 Children’saverageinitiationtimedisplayedasafunctionofcurrenttrialcongruency(C,I),previoustrialcongruency(c,i),andresponsetype(changeorrepeat).Errorbarsdisplaystandarderrors
585
605
625
645
665
685
705
725
745
Initi
atio
n Ti
me
(mse
c)
Previous Trial Congruency
CongruentIncongruent
iIcI
cC
iC
iC
cC
cI
iI
Response Change Response Repeat
Congruent Incongruent Congruent Incongruent
F IGURE 3 Children’saverageerrorratedisplayedasafunctionofcurrenttrialcongruency(C,I),previoustrialcongruency(c,i),andresponsetype(change,repeat).Errorbarsdisplaystandarderrors
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5Er
ror R
ate
(%)
Previous Trial Congruency
CongruentIncongruent
Congruent Incongruent Congruent Incongruent
iI
cI
cCiC
iC
cC
cIiI
Response Change Response Repeat

| 7 of 14ERB Et al .
isconsistentwithpreviousresearchinvestigatingalternationbehaviorinchildrenwhichindicatesthatolderchildren(above4.5yearsofage)arebiasedtoalternatebetweenresponseoptionswhileyoungerchil-dren(around3yearsofage)arebiasedtoperseverateononeoption(Jeffrey&Cohen,1965).This interpretationisalsosupportedbyre-searchindicatingthatlongresponse-to-stimulusintervals(RSIs)tendto encourage faster responding on response change than responserepeattrials(e.g.,Soetens,Boer,&Hueting,1985).Whileanumberofstudieshavenotobservedfasterrespondingonresponsechangethanresponse repeat trials in children (for a review, see Smulders etal.,2005),thesestudiesfeaturedshorterRSIsthanthecurrentstudy.
TheinteractionbetweenAgeGroupandCurrentTrialCongruencyapproached significance, F(4, 55)=2.37, p = .064, ηp
2=0.15, as didthe interaction between Age Group and Previous Trial Congruency,F(4,55)=2.16,p = .085,ηp
2=0.14.Given that5-year-oldshadhighererrorratesthantheotheragegroups,itisplausiblethattheseinterac-tioneffectswereweakenedbythedisproportionateexclusionoftrialsfromtheyoungestagegroup.Whentrialsfeaturingerrorswereincludedin the analysis, the interaction betweenAgeGroup andCurrentTrialCongruencyreachedsignificance,F(4,55)=3.13,p = .022,ηp
2=0.18(seeFigure4B),whiletheinteractionbetweenAgeGroupandPreviousTrialCongruencystillremainedatamarginallevel,F(4,55)=2.27,p = .073,ηp
2=0.14.Post-hoc analyseswithBonferroni corrections revealednosignificant differences in the effect of Current Trial Congruency be-tweenanyoftheindividualagegroups,althoughthedifferencebetween5-year-oldsand9–10-year-oldsapproachedsignificance,p = .051.
2.2.3 | Curvature
AveragereachcurvatureforeachtrialtypeisshowninFigure6.TheANOVAon reach curvatures revealedmaineffectsofCurrentTrialCongruency, Previous Trial Congruency, and Response Type, all p-values<.001,allηp
2-values>.19.Further,alltwo-wayinteractionsandthethree-wayinteractionamongthesefactorsweresignificant,all p-values<.001,allηp
2- values > .20.
Toaccount for these interactions,wefirst evaluated the effectsofPreviousTrialCongruency andResponseTypeon congruent andincongruenttrialsseparately.NoeffectofPreviousTrialCongruency,F(1, 59)=1.21, p = .28, or Response Type, F(1, 59)=0.83, p = .37,wereobservedoncongruenttrials.Thesefindingsareconsistentwiththepredictionthatreachcurvaturesoncongruenttrialswouldbeuni-formlylow.
IncongruenttrialsrevealedsignificantmaineffectsofPreviousTrialCongruency,F(1,59)=18.35,p < .001,ηp
2=0.25,andResponseType,F(1,59)=35.45,p < .001,ηp
2=0.39. Inaddition, therewasasignif-icant interaction between Previous Trial Congruency and ResponseType,F(1,59)=24.94,p < .001,ηp
2=0.31.Follow-uptests revealedasignificanteffectofPreviousTrialCongruencyontrialsfeaturingaresponse repeat, F(1,59)=38.07, p < .001, ηp
2=0.39, with largerreachcurvaturesoncI-rrelativetoiI-rtrials.NoeffectofPreviousTrialCongruencywasobservedontrialsfeaturingaresponsechange,F(1,59)=0.005,p = .94.Post-hocanalysesrevealednosignificantdiffer-encebetweeniI-rtrialsandeitherthecI-coriI-ctrials,p- values = .14. TheseresultssupporttheclaimthatS–RbindingconflictoncI-rtri-als delayed the conflict resolution process involving competitiveinhibition.
TheinteractionbetweenAgeGroupandCurrentTrialCongruencyapproached significance, F(4, 55)=2.14, p = .089, ηp
2=0.13.Whentrials featuring errors were included in the analysis, this interac-tion reached significance, F(4, 55)=2.85, p = .032, ηp
2=0.17 (seeFigure4C).Post-hocanalyseswithBonferronicorrectionsrevealedasignificantly larger effectofCurrentTrialCongruency in5-year-oldsrelativeto7-and9–10-year-olds,p-values<.05.Noothersignificantdifferenceswereobservedamongtheagegroups.
2.3 | Discussion
The results of Experiment 1 indicate that two of themeasures af-fordedbyreachtracking, initiationtimeandcurvature,canbeusedtotargetdissociableprocessesunderlyinginhibitorycontrol in5-to10-year-olds, consistentwith Erb etal.’s (2016) resultswith adults.Children’sinitiationtimesrevealedmaineffectsofcurrentandprevi-ous trialcongruency,andmatched thepatternof trial sequenceef-fectsproposedtoreflectthedACC’sroleinsupportingtheresponsethresholdadjustmentprocess:cC<iC<cI<iI (Shenhavetal.,2013;Shethetal., 2012).These results suggest that conflict registeredattheoutsetofatrialresultedinhigherresponsethresholdswithlongerperiodsofmotoricstopping.
Reachcurvaturesalsorevealedamaineffectofcurrenttrialcon-gruency,withlargercurvaturesonincongruentrelativetocongruenttrials.Whilenoeffectofresponsetypeorprevioustrialcongruencywasobservedoncongruenttrials,thesubsetofincongruenttrialsthatfeaturedS–Rbindingconflict(cI-rtrials)generatedsignificantlylargerreachcurvaturesthantheotherincongruenttrialtypes.Theseresultsareconsistentwith theclaim thatS–Rbindingconflict impedes theconflictresolutionprocessonincongruenttrials,whichleadspartici-pantstobepulledtowardtheprepotentresponseforalongerperiodoftimebeforetop-downsupportcanintervene.
F IGURE 6 Children’saveragereachcurvaturedisplayedasafunctionofcurrenttrialcongruency(C,I),previoustrialcongruency(c,i),andresponsetype(change,repeat).Errorbarsdisplaystandarderrors
0.040.060.08
0.10.120.140.160.18
0.20.220.24
Cur
vatu
re (r
atio
)
Previous Trial Congruency
CongruentIncongruent
Congruent Incongruent Congruent Incongruent
iI
cI
cC iCiCcC
cI iI
Response Change Response Repeat

8 of 14 | ERB Et al .
Itisimportanttonotethatbecauseflankernumberwasmanipulatedinthetask,asubsetofiI-rtrialsfeaturedastimuluschange(e.g., followedby).Thus,itispossiblethatS–RbindingconflictcouldhaveoccurredoncertainiI-rtrials,assumingthatparticipantsperceivedthestimulitobesubstantiallydifferentfromoneanother.Totestthispos-sibility,wecomparedreachcurvaturesoniI-rtrialsfeaturingastimuluschangetothosefeaturingastimulusrepeat.Weobservednoevidenceofadifferencebetweenthetwotrialtypes,F(1,59)=0.13,p = .72.Thisfinding,coupledwiththeobservationthatreachcurvaturesdidnotdifferbetweeniI-rtrialsandcI-coriI-ctrials,suggeststhatS–RbindingconflictdidnotsignificantlyimpactperformanceoniI-rtrials.
Theresultsofthecurrentexperimentrevealedanumberofage-relateddifferences.Children’soverallerrorratesand initiationtimesdecreasedsignificantlywithage.Whenincorrectresponseswerein-cluded in the analyses, initiationtime and curvature revealedmod-estgainsininhibitorycontrol(asindexedbytheinteractionbetweencurrent trialcongruencyandagegroup),particularlybetween5and6years of age. Movement time and total time also revealed age-related gains in inhibitory control, again driven by differences be-tween5-year-oldsandolderagegroups(seeSection1ofSupportingInformation).Thesefindingsareconsistentwiththeresultsofprevi-ousstudiesusingsimilarchild-friendlystimuli,whichhavefoundlim-iteddevelopmentalgainsinchildren’sresponsetimesbetween6and10yearsofage(e.g.,Checaetal.,2014;Ruedaetal.,2004).
Given that initiation time and reach curvature revealed similarinteractions between age group and current trial congruency, ourresults indicate that thegains in inhibitorycontrolmadebetween5and 10years of age reflect changes in the functioning of both theresponse threshold adjustment process and the conflict resolutionprocess.However,itisuncleartheextenttowhicheachofthesepro-cesses contributes to the developmental gains in inhibitory controlobservedbetweenchildhoodandadulthood(Lietal.,2009;Waszaketal.,2010).WeaddressthisquestioninExperiment2.
3 | EXPERIMENT 2
InExperiment2,wepresentedadultparticipantswithareachtrackingversionoftheflankertaskanalogoustothatofExperiment1.Inlightofpreviousreachtrackingresearchwithadults(Erbetal.,2016),wepre-dictedthatinitiationtimeandreachcurvaturewouldrevealthesameoverallpatternsofeffectsasthoseobserved inExperiment1.Aftertestingthisprediction,we investigatedhowtheresponsethresholdadjustmentprocessandconflictresolutionprocesschangebetweenchildhood and adulthood by directly comparing 8- to 10-year-olds’performancetothatofadults.
3.1 | Method
3.1.1 | Participants
Twenty-four right-handed adults (M = 20.1years,SD = 1.3 years; 14 females) with normal reaching behavior and normal or
corrected-to-normal vision participated in the experiment.Participants received course credit for their participation. TheInstitutional Review Board at Brown University approved theprotocol.
3.1.2 | Materials
ThesamematerialswereusedasinExperiment1.
3.1.3 | Procedure
Participantswerepresentedwithanarrayofdarkgreyarrows thatpointedtotheleftorrightandwereinstructedtoindicatewhichdi-rectionthearrowinthecenterofthearraypointed.Participantsre-spondedbyreachingtotouchoneoftwotargetslocatedtowardthetop leftand rightof adigitaldisplaywhilewearinga small trackingdeviceontheirindexfinger.Thebackgroundofthedisplaywaswhiteandtheresponsetargetswere identicalorangecircleswithadiam-eterof2cm.Halfofthetrialsfeaturedanarrayofthreearrows(e.g.,),while theotherhalf featuredan arrayoffive arrows (e.g.,)(seeSection2oftheSupportingInformationforresultsrelatingtothismanipulation).Thethreearrowarrayswere4.8cmby1.5cm.Thefivearrowarrayswere8.2cmby1.5cm.
The structure of the procedurewas the same as Experiment 1,except that participants had 3seconds to respond following stimu-lusonset insteadof10seconds.Participantscompleted16baselinetrials inwhichasolotargetappearedateachofthetargetlocationsfrom themain task.These trialsprovidedabaselineofparticipants’reaching behavior and familiarized participantswith the procedure.Theexperimentalportionofthetaskwaspresentedinfiveblocksof48trials.Eachblockoftrialsconsistedof24congruentand24incon-gruenttrials.Thecorrectresponsewasdividedequallybetweentheleftandrighttargets.Beforeeachblock,participantswereremindedtorespondquicklywhilemaintainingahighdegreeofaccuracy.Thefirst10trialsofthefirstblockwerepresentedaspracticetrialsandwereexcludedfromfurtheranalysis.
3.1.4 | Data processing
DatawereprocessedinthesamemannerasinExperiment1.Anaver-ageof1.15%(SD =3.62%)oftrialswereadjustedmanuallyforeachparticipant.
3.2 | Results
Thefirst trialofeachblockwasexcluded fromanalysis.Error rateswereatfloor(lessthan1%)andwerenotanalyzedfurther.Averageinitiation time and curvature were calculated for all accurate trialsthatwerenotprecededbyanerrorandwerethenanalyzedviaa2(CurrentTrialCongruency:Cor I)×2 (PreviousTrialCongruency:cori)×2(ResponseType:changeorrepeat)ANOVA.SeeSection2ofSupportingInformationforresultsfromthemovementtimeandtotaltimemeasures.

| 9 of 14ERB Et al .
3.2.1 | Initiation time
Average initiation time for each trial type is shown in Figure7. Aspredicted, the ANOVA on initiation times revealedmain effects ofCurrentTrialCongruency,F(1,23)=60.36,p < .001,ηp
2=0.72,andPreviousTrialCongruency,F(1,23)=18.72,p < .001,ηp
2 = 0.45. No interactionbetweenCurrentandPreviousTrialCongruencywasob-served,F(1,23)=1.63,p = .21.However,asignificantinteractionbe-tweenCurrentTrialCongruencyandResponseTypewasobserved,F(1,23)=7.52,p = .012,ηp
2=0.25.Follow-uptestsrevealedsignifi-cantlyhigherinitiationtimesoncongruenttrialsfeaturingaresponserepeatrelativetothosefeaturingaresponsechange,F(1,23)=9.48,p = .005, ηp
2=0.29. No effect of Response Typewas observed onincongruenttrials,F(1,23)=0.03,p = .86.AsinExperiment1,theseresults support the claim that initiation time reflects the responsethresholdadjustmentprocessinvolvingtheglobalinhibitionofmotoroutput.
3.2.2 | Curvature
AveragereachcurvatureforeachtrialtypeisshowninFigure8.TheANOVAon reach curvatures revealedmaineffectsofCurrentTrialCongruency, Previous Trial Congruency, and Response Type, all p-values<.01, allηp
2-values>.27.Further, all two-way interactionsandthethree-wayinteractionamongthesefactorsweresignificant,all p-values<.01,allηp
2- values > .32.To account for these interaction effects, we first evaluated the
effectsofPreviousTrialCongruencyandResponseTypeoncongru-ent and incongruent trials separately. No effects of Previous TrialCongruency,F(1,23)=0.05,p = .83,orResponseType,F(1,23)=0.31,p = .83,were observed on congruent trials. These findings are con-sistent with the prediction that reach curvatures on congruent tri-alswouldbeuniformly low.SignificantmaineffectsofPreviousTrialCongruency,F(1,23)=14.98,p < .001,ηp
2=0.39,andResponseType,F(1,23)=10.61,p = .003,ηp
2=0.32,wereobservedon incongruent
trials. In addition, a significant interaction between Previous TrialCongruency and Response Type was observed, F(1, 23)=21.02,p < .001, ηp
2=0.48. Follow-up tests revealed a significant effect ofPreviousTrialCongruencyon trials featuringa response repeat,F(1,23)=25.26,p < .001,ηp
2=0.52,with largerreachcurvaturesoncI-rrelativeto iI-r trials.NoeffectofPreviousTrialCongruencywasob-servedontrialsfeaturingaresponsechange,F(1,23)=0.18,p = .67.TheseresultssupporttheclaimthatS–RbindingconflictoncI-rtrialsdelayedtheconflictresolutionprocessinvolvingcompetitiveinhibition.
3.3 | Comparison of child and adult data
Overall, adultperformancepresented the samepatternsof trial se-quenceeffectsobservedinchildreninExperiment1andinadultsinErbetal.(2016).Initiationtimesweresignificantlylongeronincongru-enttrialsandtrialsprecededbyanincongruenttrial.Adults’reachcur-vatureswereuniformlylowoncongruenttrials,highestoncI-rtrials,and intermediateontheremaining incongruenttrial types (cI-c, iI-c,andiI-r).Again,wesawnodifferencebetweeniI-rtrialsfeaturingare-sponsechangeandthosefeaturingaresponserepeat,F(1,23)=0.01,p = .93,indicatingthatS–RbindingconflictdidnotsignificantlyimpactperformanceoniI-rtrialsinthecurrentexperiment.
Next,weevaluatedhowinhibitorycontrolchangesbetweenchild-hoodandadulthoodbydirectlycomparingadultperformancetothatof the twooldest agegroups fromExperiment1with a seriesof2(CurrentTrialCongruency:CorI)×2(PreviousTrialCongruency:cori)×2(ResponseType:changeorrepeat)×2(AgeGroup:8-to10-year-oldsoradults)ANOVAs.Asinourpreviousanalyses,thefirsttrialofeach roundwasexcluded fromanalysis andonly accurate trialsnotprecededbyanerrorwereanalyzed.Inordertoequateforthenumberoftrialspresentedineachagegroup,onlythefirst108experimentaltrials collectedwith adultswere included in the following analyses.Thisstepwastakentoguardagainstthepossibilitythatage-relatedchangesinperformancecouldreflectpracticeorfatigueeffectsintheadultsample.Toseeafullcomparisonofchildandadultperformance,
F IGURE 7 Adults’averageinitiationtimedisplayedasafunctionofcurrenttrialcongruency(C,I),previoustrialcongruency(c,i),andresponsetype(changeorrepeat)foradultparticipants.Errorbarsdisplaystandarderrors
350
360
370
380
390
400
410
420
430
440In
itiat
ion
Tim
e (m
sec)
Previous Trial Congruency
CongruentIncongruent
Congruent Incongruent Congruent Incongruent
iIcI
cC
iC
iCcC
cIiI
Response Change Response Repeat
F IGURE 8 Adults’averagereachcurvaturedisplayedasafunctionofcurrenttrialcongruency(C,I),previoustrialcongruency(c,i),andresponsetype(changeorrepeat)foradultparticipants.Errorbarsdisplaystandarderrors
0.040.050.060.070.080.09
0.10.110.120.130.14
Cur
vatu
re (r
atio
)
Previous Trial Congruency
CongruentIncongruent
Congruent Incongruent Congruent Incongruent
iI
cI
cC iCiCcC
cI iI
Response Change Response Repeat

10 of 14 | ERB Et al .
including all ages tested in Experiment 1 and all trials collected inExperiment2,seeSection3oftheSupportingInformation.Givenourfocusondevelopmentaldifferences,werestrictourdiscussiontoin-teractionswithAgeGroupinthefollowinganalyses.
3.3.1 | Initiation time
AsignificantinteractionbetweenAgeGroupandResponseTypewasobserved, F(1, 46)=4.37, p = .042, ηp
2=0.09, with a larger differ-encebetweenresponserepeatandresponsechangetrialsinchildren(M = 25ms,SD=43ms) thanadults (M = 3ms,SD=21ms).Nosig-nificantinteractionswereobservedwithAgeGroupandCurrentTrialCongruencyorPreviousTrialCongruency,allp- values > .17.
3.3.2 | Curvature
Reachcurvaturerevealedasignificanttwo-wayinteractionbetweenAge Group and Response Type, significant three-way interactionsamongAgeGroup,CurrentTrialCongruency,andResponseType,andAgeGroup,PreviousTrialCongruency,andResponseType,andasig-nificantfour-wayinteractionamongallthefactors,allp-values<.05.
Toaccountfortheseinteractioneffects,wesplitouranalysesbyPreviousTrialCongruency. Follow-up tests revealed no interactionsofAgeGroupwithCurrentTrialCongruencyorResponseTypeontri-alsprecededbyan incongruent trial,p-values> .20.Trialsprecededby a congruent trial revealed a significant interaction betweenAgeGroupandResponseType,F(1,46)=12.53,p < .001,ηp
2=0.21,anda significant three-way interaction amongAge Group, Current TrialCongruency,andResponseType,F(1,46)=9.31,p = .004,ηp
2 = 0.17.To further evaluate these interactions, we analyzed response
changeandresponserepeattrialsseparately.TheinteractionbetweenAgeGroupandCurrentTrialCongruencywasnot significanton re-sponsechangetrials,F(1,46)=0.36,p = .55.However,itwassignifi-cantonresponserepeattrials,F(1,46)=4.62,p = .037,ηp
2=0.09,withalargerdifferencebetweencI-randcC-rtrialsinchildren(M = 0.115,SD=0.090)thanadults(M = 0.067,SD=0.065).
3.4 | Discussion
Thecomparisonofchildandadultperformance revealednosignifi-cantage-relatedgainsininhibitorycontrolininitiationtime.Althoughtheeffectofresponsetypeoninitiationtimediddecreasebetween8-to10-year-oldsandadults,thisimprovementoccurredacrossbothcongruentandincongruenttrials.Incontrasttoinitiationtime,reachcurvaturedidrevealsignificantage-relatedgainsininhibitorycontrol.However,theobservedgainswerespecifictoaparticularsubsetoftrials;namely,thosefeaturingS–Rbindingconflict.Reachcurvatureson cI-r trials decreased significantly between childhood and adult-hood,while theother trial types revealednosignificantage-relatedimprovements.Theseresultsindicatethattheage-relatedgainsinin-hibitorycontrolobservedbetween8to10yearsofageandadulthoodaredrivenbyimprovementsintheconflictresolutionprocess’capac-itytoresolveS–Rbindingconflict.
In a separate experiment, we presented adult participantswiththesamechild-friendlyversionof the taskpresented inExperiment1.When we compared child and adult performance, we observednearly identical results to those presented above (see Section 4 ofSupporting Information). However, it was unclear whether the ob-servedage-relatedchangesinperformanceweredrivenbyimprovedinhibitorycontrolinadultsorbydifferencesintaskdifficulty(e.g.,thechild-friendlyversionmayhavebeentooeasyforadults). In lightofpreviousresearchindicatingthatthearrowversionofthetaskismoredifficult thanthechild-friendlyversionused inExperiment1 (Ruedaetal.,2004),weselectedthearrowversionofthetaskforExperiment2inanefforttoequatetaskdifficultybetweentheagegroups.Takentogether, the results presented above and the results presented inSections3and4oftheSupportingInformationconvergeonthesameconclusion:theage-relatedgainsobservedbetweenmiddlechildhoodandadulthoodintheflankertaskaredrivenbychangesinthefunc-tioningoftheconflictresolutionprocessontrialsfeaturingS–Rbind-ingconflict.
4 | GENERAL DISCUSSION
The results of the current study indicate that twoof themeasuresaffordedby reach tracking, initiationtimeand reach curvature, canbeused to target the functioningof the response thresholdadjust-mentprocess and conflict resolutionprocess in children andadultsalike. Initiationtimes inExperiments1 and2 revealedmaineffectsof current and previous trial congruency, consistentwith the claimthatconflictdetectedattheoutsetofatrialleadstohigherresponsethresholds and, consequently, increased global inhibition of motoroutput. Reach curvatures across both experiments were uniformlylowoncongruent trials,elevatedon incongruent trialswithoutS–Rbindingconflict (cI-c, iI-c,and iI-r trials), and largeston incongruenttrialswithS–Rbindingconflict (cI-r trials).Thispatternofeffects isconsistentwiththeclaimthatreachcurvaturereflectstherelativeco-activationof responsesover thecourseofamovementand, there-fore, canbeused to index the conflict resolutionprocess involvingcompetitiveinhibition(Erbetal.,2016).
Theoverallpatternofeffectsobservedininitiationtimeandreachcurvatureweresimilar inchildrenandadults, indicating that the re-sponsethresholdadjustmentprocessandconflictresolutionprocessfunctioninmuchthesamemannerinbothagegroups.Themeasuresdid, however, present a number of notable developmental differ-ences. Both initiation time and reach curvature revealed relativelymodestgainsininhibitorycontrolduringchildhood(asevidencedbyinteractions between current trial congruency and age group),withpronounced improvementsoccurringbetween5and6yearsofage.These results suggest that both processes of interest follow similardevelopmental trajectories during childhood. However, only reachcurvaturerevealedevidenceofage-relatedgainsininhibitorycontrolbetweenchildhoodandadulthood,indicatingthattheprotractedde-velopmentof inhibitorycontrol intoadulthoodisdrivenprimarilybychanges in the functioningof theconflict resolutionprocess.These

| 11 of 14ERB Et al .
findingssuggestthattheresponsethresholdadjustmentprocessandconflict resolutionprocess followdivergentdevelopmental trajecto-ries, although further research isneeded toexaminewhether theseresultsgeneralizetoothertasksandtestingconditions.
It is important tonote that the inhibitorycontroladvantageob-served in adult reach curvatureswas driven by a specific subset oftrials; namely, incongruent trials featuring S–R binding conflict (cI-rtrials).Thisfindingraisesanumberofimportantquestionsconcerningthenatureofthegainsininhibitorycontrolobservedbetweenchild-hoodandadulthood.Forexample,werethedevelopmentalgainsob-served in thecurrent studydrivenbycI-r trialsbecause these trialswerethemostdifficultand,therefore,themost likelytorevealage-relatedimprovements?Or,werethedevelopmentalgainsspecifictocI-rtrialsbecausethesetrialsplacedunique–notjustgreater–de-mandsonparticipants?
Research by Hommel, Kray, and Lindenberger (2011) providessomeinsightonthisissue.Theypresented9-to10-year-olds,youngeradults(20–31yearsofage),andolderadults(64–76yearsofage)withataskadoptedfromHommel(1998)thatsystematicallymanipulatedstimulus and response repetitions. Hommel and colleagues foundthat theeffectofS–Rbindingconflictwasmorepronounced in theerror rates of 9- to 10-year-olds than adults. In their discussion ofthisage-relatedchange,theresearchersreferencedworkbyColzato,vanWouwe,Lavender,andHommel(2006)indicatingthattheabilityto unbind and rebind event features (e.g., a particular stimulus andresponse)islinkedtofluidintelligence,whichimprovesduringchild-hoodandadolescence(Fry&Hale,2000).Further,Hommeletal.notethatthereconfiguringofeventfeatureshasbeenproposedtoinvolveneurophysiologicalprocesseslinkedtodopaminergicmodulation,thefunctioningofwhichhasalsobeen found to relate toageandfluidintelligence(citingBäckman,Nyberg,Lindenberger,Li,&Farde,2006).Thus,theage-relatedgainsinflankertaskperformanceobservedbe-tween8-to10-year-oldsandadultsinthecurrentstudymayreflectchangesspecifictohowefficientlyeventfeaturescanbereconfiguredat different points in development, rather than improved inhibitorycontrol(orconflictresolution)perse.
This account is consistentwith recentwork byCragg (2016) inwhich 7-year-olds, 10-year-olds, and adults completed a modifiedversion of the flanker task that did not allow stimulus or responserepeats to occur. In contrast to previous developmentalwork (e.g.,Lietal.,2009;Waszaketal.,2010),Craggobservednoage-relatedgains in inhibitorycontrolon responseconflict trials (akin to incon-gruent trials in the current study).The results of the current studysuggestthatthelackofage-relatedchangesinCragg(2016)mayhaveresultedfromtheremovalofassociativeprimingeffectssuchasS–Rbindingconflict.Whilefurtherresearchisneededtoevaluatetheex-tent towhich age-related gains in flanker task performance reflectimproved inhibitory control or an improved capacity to reconfigureeventfeatures,ourresultspresentfurtherevidencethatage-relatedgainsinthesetasksaredriveninpartbyaselectsubsetoftrialsthatfeature greater – and possibly different – demands. Consequently,age-relatedgainsinperformanceonthesetasksmayhavebeenmis-characterizedinthepast.
4.1 | Conflict adaptation
Anongoingdebateintheinhibitorycontrolliteratureconcernstheex-tenttowhichperformanceoncongruencytasksisinfluencedbyconflict-drivenmodulationsoftop-downcontrol.AsnotedintheIntroduction,faster responsetimeson iI relativetocI trialshavebeen interpretedtoreflectaconflictadaptationeffectinwhichtherecentrecruitmentoftop-downsupportononeincongruenttrialservestofacilitateper-formanceonthenext(e.g.,Botvinicketal.,2001).WhileanumberofstudieshavefoundthatthedifferencebetweeniIandcItrialsisbetterexplainedbyS–RbindingconflictoncI-rtrialsthanconflictadaptationoniItrials(Mayretal.,2003;Nieuwenhuisetal.,2006),otherstudiesfeaturingmorethantworesponsesandalargerstimulussetsizehaveobserved enhancedperformance on iI relative to cI trials even aftertrialsfeaturingS–Rbindingconflictwereexcludedfromanalysis(e.g.,Kerns etal., 2004; Ullsperger etal., 2005; Verbruggen etal., 2006).However,recentresearchindicatesthatsuchattemptstocontrolforS–Rbindingconflictcanhavetheunintendedconsequenceofintroduc-ingcontingencylearningeffectsinwhichparticipantslearntoassociateparticularstimulusfeatureswithaspecificresponse(foradiscussion,seeSchmidt,2013).Forexample,whenSchmidtandDeHouwer(2011)controlledforbothS–Rbindingconflictandcontingencylearningef-fects in the flanker task, they observed no performance differencesbetween iI andcI trials, suggesting that conflict adaptationdoesnotsignificantlycontributetoflankertaskperformance.
Inthecurrentstudy,weobservednoevidenceofconflictadapta-tion in initiation time or reach curvature.Although reach curvatureswerelargeroncIrelativetoiItrials,thisdifferencewasspecifictotrialsfeaturingaresponserepeat, indicatingthatthedifferencewasdrivenbyS–RbindingconflictoncI-rtrialsandnotconflictadaptationon iItrials. Children’s movement times in Experiment 1were faster on iIrelativetocI trials regardlessof response type, suggestingapossibleroleforconflictadaptation(seeSection1ofSupportingInformation).However,thedifferencebetweencCandiCtrialswasalsomarginallysignificant (p = .066), indicating that children’smovementtimesweregenerallyfasterontrialsfeaturingacongruencyrepeat(cCandiItrials)thanthosefeaturingacongruencychange(iCandcItrials).Interestingly,adults’movementtimesdidnotshowevidenceofthisgeneralperfor-mancedifferenceoncongruencyrepeatandcongruencychangetrialsinExperiment2ortheadditionalexperimentreportedinSection4oftheSupporting Information.Thesefindingssuggestthatchildrenmayex-perienceaswitchcostintheflankertasknotencounteredbyadults,al-thoughfurtherresearchisnecessarytodirectlyevaluatethispossibility.
4.2 | Methodological considerations
Traditionally,inhibitorycontrolintheEriksenflankertaskisassessedbyevaluatingtheeffectofcurrenttrialcongruencyonresponsetimes,accuracy, or a composite score of the two (e.g.,Nieuwenhuis etal.,2006; Rueda etal., 2004;Waszak etal., 2010; Zelazo etal., 2013).Whilethisapproachhascontributedgreatlytoourcurrentunderstand-ingof inhibitorycontroland itsdevelopment, theresultsof thepre-sentstudyhighlightthepromiseof(a)collectingcontinuousbehavioral

12 of 14 | ERB Et al .
measures and (b) evaluating the role of trial sequence effectswheninvestigatinginhibitorycontrolinchildren.Reachtrackingandrelatedtechniques suchasmouse trackinghavebeenused to studyawiderangeoftopicsinadults,includingattention,categorization,numericalcognition, language, anddecisionmaking (for reviews, seeFreeman,Dale,&Farmer,2011,andSong&Nakayama,2009).However,rela-tivelylittleresearchhasbeenconductedusingthesetechniqueswithchildren(e.g.,Anderson,Farmer,Goldstein,Schwade,&Spivey,2011;Diedrichetal.,2000;Rommelseetal.,2007).Ourresultsindicatethatreachtrackingisappropriateforusewithchildrenasyoungas5yearsofage,andpresentsapromisingalternativetothebehavioralmethodstraditionallyusedtoassesschildren’sperception,cognition,andaction.
Theabilitytoisolatedistinctpatternsofeffectsunderlyingperfor-mance isparticularly relevant fordevelopmental research,asoverallmeasures of performance such as response timemay conceal age-relatedchangesofinterest.Forexample,totaltimerevealednoage-relatedgains in inhibitorycontrolbetweenchildren8to10yearsofageandadults(seeSection2ofSupportingInformation),despitecur-vatureandmovementtimerevealingcleargainsinperformance.Thisisbecausetotaltimeistheproductofbothinitiationtimeandmove-menttime,which generateddifferent patternsof trial sequence ef-fectsandfolloweddifferentdevelopmentaltrajectoriesinthecurrentstudy.Thus,ourfindingsindicatethatreachtrackingcanbeusedtotargetdevelopmentalchangesinperformancethatmaybeobscuredbymeasuresthatreflecttheoutcomeofadecisionprocessbutnotitsunfolding.
4.3 | Conclusion
Thecurrentstudypresentsevidencethatreachtrackingcanbeusedtotargethowtwokeyprocessesunderlyinginhibitorycontrolfunc-tionacrossthelifespan.Ourfindingsindicatethataresponsethresh-oldadjustmentprocessinvolvingtheglobalinhibitionofmotoroutputand a conflict resolution process involving competitive inhibitionamongco-activeresponsesfollowdifferentdevelopmental trajecto-riesbetweenchildhoodandadulthood.Thecurrent studyalsopre-sentsaframeworkforfutureresearchtoexplorehoweachoftheseprocessescontributestoindividualandgroupdifferencesininhibitorycontrol.More broadly, our results contribute to a growing body ofdevelopmental research highlighting the importance of evaluatingtrialsequenceeffectsinchildren’scognitiveperformance(e.g.,Cragg,2016;Hommeletal.,2011;Kray,Karbach,&Blaye,2012).
ACKNOWLEDGEMENTS
ThisprojectissupportedbyNSF1223777toDMSandNIGMS-NIHIDeAP20GM103645toJHS.
NOTES1Erbetal.(2016)usedathree-responseversionoftheflankertaskthatre-sultedinalargernumberofresponsechangethanresponserepeattrials.Consequently, the researchersdidnot investigate theeffectof responsetype(repeatvs.change).
2Recently,Cragg(2016)hasreportedresponsetimedataindicatingthatchil-drenasyoungas7yearsof agepresent similar trial sequenceeffects tothoseobservedinolderchildrenandadults(e.g.,Nieuwenhuisetal.,2006),althoughthetaskdidnotassesstheinfluenceofresponsetype(repeatvs.change)and,consequently,theroleofS–Rbindingconflictwasnoteval-uated.Similarly,workbyTakaraeandcolleagues (2009)featuredchildrenyoungerthan10(7–14years),buttheaverageageoftheirtypicallydevel-oping participantswas greater than10years and the effect of responsetypewasnotreported.
REFERENCES
Anderson, S.E., Farmer, T.A., Goldstein, M., Schwade, J., & Spivey, M.(2011). Individual differences in measures of linguistic experienceaccount for variability in the sentence processing skill of five-year-olds. Experience, Variation, and Generalization: Learning a First Language (Trends in Language Acquisition Research),7,203–221.
Bäckman,L.,Nyberg,L.,Lindenberger,U.,Li,S.C.,&Farde,L.(2006).Thecorrelativetriadamongaging,dopamine,andcognition:Currentsta-tusandfutureprospects.Neuroscience and Biobehavioral Reviews,30,791–807.
Blair,C.,&Razza,R.P.(2007).Relatingeffortfulcontrol,executivefunction,andfalsebeliefunderstandingtoemergingmathandliteracyabilityinkindergarten.Child Development,78,647–663.
Botvinick, M.M., Braver, T.S., Barch, D.M., Carter, C.S., & Cohen, J.D.(2001).Conflictmonitoringandcognitivecontrol.Psychological Review,108,624–652.
Carlson,S.M.,Moses, L.J.,&Breton,C. (2002).Howspecific is the rela-tionbetweenexecutivefunctionandtheoryofmind?Contributionsofinhibitorycontrolandworkingmemory.Infant and Child Development,11,73–92.
Carlson,S.M.,&Wang,T.S.(2007).Inhibitorycontrolandemotionregula-tioninpreschoolchildren.Cognitive Development,22,489–510.
Carver,A.C., Livesey,D.J., &Charles,M. (2001).Age related changes ininhibitory control as measured by stop signal task performance.International Journal of Neuroscience,107,43–61.
Cavanagh, J.F., Wiecki, T.V., Cohen, M.X., Figueroa, C.M., Samanta, J.,Sherman, S.J., & Frank, M.J. (2011). Subthalamic nucleus stimula-tion reversesmediofrontal influence over decision threshold.Nature Neuroscience,14,1462–1467.
Checa,P.,Castellanos,M.C.,Abundis-Gutiérrez,A.,&Rueda,M.R.(2014).Development of neural mechanisms of conflict and error process-ing during childhood: Implications for self-regulation. Frontiers in Psychology,5,326.
Cohen,J.D.,Dunbar,K.,&McClelland,J.L. (1990).Onthecontrolofau-tomatic processes: A parallel distributed processing account of theStroopeffect.Psychological Review,97,332–361.
Cohen, J.D., & Huston, T.A. (1994). Progress in the use of interactivemodels forunderstandingattentionandperformance. InC.Umiltà&M. Moscovitch (Eds.), Attention and performance XV (pp. 453–476).Cambridge,MA:MITPress.
Colzato, L.S., Van Wouwe, N.C., Lavender, T.J., & Hommel, B. (2006).Intelligenceandcognitiveflexibility:Fluid intelligencecorrelateswithfeature‘unbinding’acrossperceptionandaction.Psychonomic Bulletin and Review,13,1043–1048.
Cragg,L.(2016).Thedevelopmentofstimulusandresponseinterferencecontrolinmidchildhood.Developmental Psychology,52,242–252.
Davidson, M.C., Amso, D., Anderson, L.C., & Diamond, A. (2006).Developmentofcognitivecontrolandexecutivefunctionsfrom4to13years:Evidencefrommanipulationsofmemory,inhibition,andtaskswitching.Neuropsychologia,44,2037–2078.
DeJong,R.,Liang,C.C.,&Lauber,E.(1994).Conditionalandunconditionalautomaticity:Adual-processmodelofeffectsof spatial stimulus–re-sponse correspondence. Journal of Experimental Psychology: Human Perception and Performance,20,731–750.

| 13 of 14ERB Et al .
Desmurget, M., Jordan, M., Prablanc, C., & Jeannerod, M. (1997).Constrained and unconstrainedmovements involve different controlstrategies.Journal of Neurophysiology,77,1644–1650.
Diamond,A. (2002).Normaldevelopmentofprefrontalcortexfrombirthtoyoungadulthood:Cognitivefunctions,anatomy,andbiochemistry.InD.T.Stuss&R.T.Knight(Eds.),Principles of frontal lobe function(pp.466–503).NewYork:OxfordUniversityPress.
Diamond,A.(2013).Executivefunctions.Annual Review of Psychology,64,135–168.
Diedrich,F.J.,Thelen,E.,Smith,L.B.,&Corbetta,D. (2000).Motormem-oryisafactorininfantperseverativeerrors.Developmental Science,3,479–494.
Egner, T. (2007). Congruency sequence effects and cognitive control.Cognitive, Affective, and Behavioral Neuroscience,7,380–390.
Erb,C.D.,Moher,J.,Sobel,D.M.,&Song,J.H. (2016).Reachtrackingre-veals dissociable processes underlying cognitive control. Cognition,152,114–126.
Eriksen, B.A., & Eriksen, C.W. (1974). Effects of noise letters upon theidentification of a target letter in a nonsearch task. Perception and Psychophysics,16,143–149.
Eriksen,C.W.,Coles,M.G.,Morris, L.R.,&O’Hara,W.P. (1985).An elec-tromyographic examination of response competition. Bulletin of the Psychonomic Society,23,165–168.
Frank,M.J. (2006). Hold your horses:A dynamic computational role forthe subthalamic nucleus in decision making. Neural Networks, 19,1120–1136.
Freeman,J.,Dale,R.,&Farmer,T.(2011).Handinmotionrevealsmindinmotion.Frontiers in Psychology,2,59.
Freeman, J.B., Nakayama, K., & Ambady, N. (2013). Finger in flight re-veals parallel categorizationacrossmultiple social dimensions.Social Cognition,31,792–805.
Fry,A.F.,&Hale,S.(2000).Relationshipsamongprocessingspeed,work-ingmemory,andfluidintelligenceinchildren.Biological Psychology,54,1–34.
Gogtay,N.,Giedd,J.N.,Lusk,L.,Hayashi,K.M.,Greenstein,D.,&Vaituzis,A.C.,…&Rapoport,J.L. (2004).Dynamicmappingofhumancorticaldevelopmentduringchildhoodthroughearlyadulthood.Proceedings of the National Academy of Sciences of the United States of America,101,8174–8179.
Gratton,G.,Coles,M.G.,&Donchin,E. (1992).Optimizingtheuseof in-formation: Strategic control of activation of responses. Journal of Experimental Psychology: General,121,480–506.
Hommel,B.(1998).Eventfiles:Evidenceforautomaticintegrationofstim-ulus–responseepisodes.Visual Cognition,5,183–216.
Hommel,B. (2004).Eventfiles:Featurebinding inandacrossperceptionandaction.Trends in Cognitive Sciences,8,494–500.
Hommel,B.,Kray,J.,&Lindenberger,U.(2011).Featureintegrationacrossthelifespan:Stickierstimulus–responsebindingsinchildrenandolderadults.Frontiers in Psychology,2,268.
Jeffrey,W.E., & Cohen, L.B. (1965). Response tendencies of children ina two-choice situation. Journal of Experimental Child Psychology, 2,248–254.
Kerns,J.G.,Cohen,J.D.,MacDonald,A.W.,Cho,R.Y.,Stenger,V.A.,&Carter,C.S.(2004).Anteriorcingulateconflictmonitoringandadjustmentsincontrol.Science,303,1023–1026.
Kray,J.,Karbach,J.,&Blaye,A.(2012).Theinfluenceofstimulus-setsizeondevelopmentalchangesincognitivecontrolandconflictadaptation.Acta Psychologica,140,119–128.
Li,S.C.,Hämmerer,D.,Müller,V.,Hommel,B.,&Lindenberger,U.(2009).Lifespandevelopmentofstimulus–responseconflictcost:Similaritiesand differences between maturation and senescence. Psychological Research,73,777–785.
Luna, B. (2009). Developmental changes in cognitive control throughadolescence. Advances in Child Development and Behavior, 37,233–278.
Luna, B., Garver, K.E., Urban, T.A., Lazar, N.A., & Sweeney, J.A. (2004).Maturationofcognitiveprocesses from latechildhoodtoadulthood.Child Development,75,1357–1372.
Mayr,U.,Awh,E.,&Laurey,P. (2003).Conflictadaptationeffects in theabsenceofexecutivecontrol.Nature Neuroscience,6,450–452.
Moher, J., & Song, J.H. (2013). Context-dependent sequential effects oftargetselectionforaction.Journal of Vision,13,10.
Munakata, Y., Herd, S.A., Chatham, C.H., Depue, B.E., Banich, M.T., &O’Reilly,R.C.(2011).Aunifiedframeworkforinhibitorycontrol.Trends in Cognitive Sciences,15,453–459.
Nieuwenhuis,S.,Stins,J.F.,Posthuma,D.,Polderman,T.J.,Boomsma,D.I.,&deGeus,E.J. (2006).Accounting for sequential trial effects in theflanker task: Conflict adaptation or associative priming?Memory & Cognition,34,1260–1272.
Ridderinkhof,K.R.,&vanderMolen,M.W.(1995).Apsychophysiologicalanalysisofdevelopmentaldifferences in theability to resist interfer-ence. Child Development,66,1040–1056.
Ridderinkhof,K.R.,vanderMolen,M.W.,&Bashore,T.R.(1995).Limitsontheapplicationofadditivefactorslogic:Violationsofstagerobustnesssuggestadual-processarchitecturetoexplainflankereffectsontargetprocessing.Acta Psychologica,90,29–48.
Rommelse,N.N.,Altink,M.E.,Oosterlaan,J.,Buschgens,C.J.,Buitelaar,J.,DeSonneville,L.M.,&Sergeant,J.A.(2007).MotorcontrolinchildrenwithADHDandnon-affectedsiblings:Deficitsmostpronouncedusingthelefthand.Journal of Child Psychology and Psychiatry,48,1071–1079.
Rueda,M.R.,Fan,J.,McCandliss,B.D.,Halparin,J.D.,Gruber,D.B.,Lercari,L.P., & Posner,M.I. (2004). Development of attentional networks inchildhood. Neuropsychologia,42,1029–1040.
Schmidt, J.R. (2013). The Parallel Episodic Processing (PEP) model:Dissociatingcontingencyandconflictadaptation in the item-specificproportioncongruentparadigm.Acta Psychologica,142,119–126.
Schmidt, J.R., &DeHouwer, J. (2011). Nowyou see it, nowyou don’t:Controlling for contingenciesand stimulus repetitionseliminates theGrattoneffect.Acta Psychologica,138,176–186.
Shenhav,A., Botvinick,M.M.,&Cohen, J.D. (2013).The expectedvalueofcontrol:Anintegrativetheoryofanteriorcingulatecortexfunction.Neuron,79,217–240.
Sheth, S.A., Mian, M.K., Patel, S.R., Asaad, W.F., Williams, Z.M., &Dougherty,D.D.,…&Eskandar,E.N.(2012).Humandorsalanteriorcin-gulatecortexneuronsmediateongoingbehaviouraladaptation.Nature,488,218–221.
Smulders, S.F., Notebaert, W., Meijer, M., Crone, E.A., van der Molen,M.W.,& Soetens, E. (2005). Sequential effects on speeded informa-tionprocessing:Adevelopmental study.Journal of Experimental Child Psychology,90,208–234.
Soetens,E.,Boer,L.C.,&Hueting,J.E.(1985).Expectancyorautomaticfa-cilitation? Separating sequential effects in two-choice reactiontime.Journal of Experimental Psychology: Human Perception and Performance,11,598–616.
Song, J.-H.,&Nakayama,K. (2006). Role of focal attentionon latenciesand trajectories ofvisually guidedmanual pointing. Journal of Vision,6,982–995.
Song,J.-H.,&Nakayama,K.(2007).Automaticadjustmentofvisuomotorreadiness. Journal of Vision,7,2.
Song,J.-H.,&Nakayama,K.(2008).Targetselectioninvisualsearchasre-vealedbymovementtrajectories.Vision Research,48,853–861.
Song, J.-H.,&Nakayama,K. (2009).Hidden cognitive states revealed inchoicereachingtasks.Trends in Cognitive Sciences,13,360–366.
Sowell, E.R., Peterson, B.S., Thompson, P.M.,Welcome, S.E., Henkenius,A.L.,&Toga,A.W.(2003).Mappingcorticalchangeacrossthehumanlifespan.Nature Neuroscience,6,309–315.
Stins, J.F., Polderman, J.T., Boomsma, D.I., & de Geus, E.J. (2007).Conditionalaccuracyinresponseinterferencetasks:EvidencefromtheEriksenflankertaskandthespatialconflicttask.Advances in Cognitive Psychology,3,409–417.

14 of 14 | ERB Et al .
Takarae, Y., Schmidt, L., Tassone, F., & Simon, T.J. (2009). Catechol-O-methyltransferase polymorphism modulates cognitive control inchildren with chromosome 22q11. 2 deletion syndrome. Cognitive, Affective, and Behavioral Neuroscience,9,83–90.
Ullsperger,M.,Bylsma,L.M.,&Botvinick,M.M.(2005).Theconflictadap-tation effect: It’s not just priming.Cognitive, Affective, and Behavioral Neuroscience,5,467–472.
vanBoxtel,G.J.,vanderMolen,M.W.,Jennings,J.R.,&Brunia,C.H.(2001).Apsychophysiologicalanalysisofinhibitorymotorcontrolinthestop-signalparadigm.Biological Psychology,58,229–262.
vandeLaar,M.C.,vandenWildenberg,W.P.,vanBoxtel,G.J.,&vanderMolen,M.W.(2014).Developmentofresponseactivationandinhibi-tioninaselectivestop-signaltask.Biological Psychology,102,54–67.
Velanova,K.,Wheeler,M.E.,& Luna,B. (2008).Maturational changes inanterior cingulate and frontoparietal recruitment support the devel-opmentoferrorprocessingandinhibitorycontrol.Cerebral Cortex,18,2505–2522.
Verbruggen,F.,Notebaert,W.,Liefooghe,B.,&Vandierendonck,A.(2006).Stimulus-andresponse-conflict-inducedcognitivecontrolintheflankertask.Psychonomic Bulletin and Review,13,328–333.
Waszak, F., Li, S.C., & Hommel, B. (2010). The development of atten-tional networks: Cross-sectional findings from a life span sample.Developmental Psychology,46,337–349.
Weissman,D.H.,Jiang,J.,&Egner,T.(2014).Determinantsofcongruencysequence effects without learning and memory confounds. Journal
of Experimental Psychology: Human Perception and Performance, 40,2022–2037.
Wiecki,T.V.,&Frank,M.J.(2013).Acomputationalmodelofinhibitorycontrolinfrontalcortexandbasalganglia.Psychological Review,120,329–355.
Yeung, N., Botvinick, M.M., & Cohen, J.D. (2004). The neural basis oferrordetection:Conflictmonitoringand theerror-relatednegativity.Psychological Review,111,931–959.
Zelazo,P.D.,Anderson,J.E.,Richler,J.,Wallner-Allen,K.,Beaumont,J.L.,&Weintraub,S. (2013).NIHtoolboxcognitionbattery (CB):Measuringexecutivefunctionandattention.Monographs of the Society for Research in Child Development,78,16–33.
SUPPORTING INFORMATION
AdditionalSupportingInformationmaybefoundonlineinthesupport-inginformationtabforthisarticle.
How to cite this article:ErbCD,MoherJ,SongJ-H,andSobelDM.Reachtrackingrevealsdissociableprocessesunderlyinginhibitorycontrolin5-to10-year-oldsandadults.Dev Sci. 2017;00:e12523.https://doi.org/10.1111/desc.12523





























