Embed Size (px)
Citation preview

640
http://journals.tubitak.gov.tr/biology/
Turkish Journal of Biology Turk J Biol(2017) 41: 640-648© TÜBİTAKdoi:10.3906/biy-1702-48
Recovery and amplification of ancient DNA from Herculaneum victims killed by the 79 AD Vesuvius hot surges
Fabio Maria GUARINO1,*, Claudio BUCCELLI2, Vincenzo GRAZIANO2, Pietro LA PORTA1, Marcello MEZZASALMA1, Gaetano ODIERNA1, Mariano PATERNOSTER2, Pierpaolo PETRONE3
1Department of Biology, University of Naples Federico II, Naples, Italy2Department of Advanced Medical Sciences, Section of Legal Medicine, University of Naples Federico II, Naples, Italy
3Department of Advanced Biomedical Sciences, Section of Legal Medicine, Laboratory of Human Osteobiology andForensic Anthropology, University of Naples Federico II, Naples, Italy
* Correspondence: [email protected]
1. IntroductionThe study of ancient DNA (aDNA) presents many challenges. Indeed, DNA extraction and amplification are hampered by degradation processes following the death of an individual, limiting the amount of extractable and amplifiable DNA, and by enzymatic inhibitors that are coextracted with aDNA (Pääbo, 1989; Lindahl, 1993; Hoss et al., 1996; Kaestle and Horsburgh, 2002). In addition, taphonomic conditions, abrupt changes in macroenvironmental conditions such as those occurring during the excavation of remains and their transfer to collections, and the risk of contamination by foreign DNA can also limit DNA extraction and amplification of aDNA (O’Rourke et al., 2000; Gilbert et al., 2005a; Pruvost et al., 2007, Elsner et al., 2015). Preservation and amplification of aDNA are also hindered in thermally damaged archaeological remains (Brown and Brown, 2011), including partially charred skeletons such as those of victims of volcanic eruption.
The possibility to successfully analyze aDNA from thermally damaged archaeological bones (for example as
a consequence of cooking) was highlighted in the study by Cottoni et al. (2009). However, this study concluded that heavily burnt or cremated bones exposed to temperatures above 170 °C are unlikely to produce authentic DNA sequences.
The first analysis of aDNA yielded from archaeological remains of human victims of the Vesuvius eruption in AD 79 was performed on thirteen skeletons from the House of Julius Polybius in Pompeii (Cipollaro et al., 1998). These studies proved that the success of aDNA extraction and amplification was closely related to the degree of bone preservation of the samples, evaluated under a light microscope. Indeed, it was possible to extract and amplify DNA only from well-preserved, nondiagenized bones. In particular, the amelogenin gene and Y-specific alphoid repeat sequences were successfully amplified and individuals were unambiguously sexed, thus implementing anthropometric observations. In a further study, after preliminary histological analysis, the short tandem repeats and von Willebrand factor loci for individual identification were also analyzed (Cipollaro et al., 1999). Favorable
Abstract: In thermally damaged archaeological bones the quantity, quality, and amplifiability of DNA are very much reliant on both the extent of heating and the environmental conditions of the burial context. In this study we tested the possibility of extracting and amplifying ancient DNA from human bone remains of Herculaneum victims of the 79 AD eruption of Vesuvius using a combination of histochemical and molecular methods. Long bone samples with variable degrees of chromatic and morphological alterations consistent with exposure to temperatures of about 300 °C were taken from four specimens. Using histochemical stains, bone cryostat sections from three individuals revealed DNA within osteocyte lacunae but only for one sample the DNA was suitable for PCR amplification obtained, namely from reactions with the primer pair for X and Y amelogenin (AMEL) loci. The relative sequence differed from the homologous trait of AMELX deposited in GenBank for six bases, probably due to degradation processes following death. Our data are indicative that archaeological bones exposed to high temperatures of about 300 °C should be considered for DNA analysis, given the favorable conditions of corpse burial and skeleton preservation, such as those that occurred for the 79 AD victims in Herculaneum and Pompeii.
Key words: Ancient DNA, DNA amplification, archaeological bones, 79 AD eruption of Vesuvius, Herculaneum
Received: 15.02.2017 Accepted/Published Online: 08.05.2017 Final Version: 24.08.2017
Research Article

GUARINO et al. / Turk J Biol
641
aDNA preservation due to burial conditions was also shown by Bailey et al. (1999), who extracted DNA from a young Barbary macaque from the Sarno Baths in Pompeii. More recently, analysis of mitochondrial and nuclear DNA fragments was performed on five equids discovered in Pompeii within the House of the Chaste Lovers and on an equid found in the archaeological site of Herculaneum, thus allowing the taxonomical status of these equids to be tested (Di Bernardo et al., 2004a, 2004b). Human and equine Pompeian remains were also successfully investigated to prove the presence of ancient DNA within the osteocytic lacunae by means of histochemical methods (Guarino et al., 2000, 2006). Finally, a new molecular study on the thirteen skeletons found in the House of Polybius in Pompeii revealed that six individuals were genetically closely related (Di Bernardo et al., 2009).
Except for the above-mentioned work by Di Bernardo et al. (2004b), to our knowledge only one other study, largely preliminary, concerning aDNA analysis from 79 AD victims at Herculaneum has been carried out (Geraci et al., 2002). That study showed greater difficulty in recovering and analyzing DNA from Herculaneum skeletal remains when compared with molecular analyses of Pompeii bone remains, likely due to exposure to higher thermal stress in the former case (Mastrolorenzo et al., 2001a, 2010).
The present paper takes its cue from a bioanthropological and volcanological multidisciplinary study (Mastrolorenzo et al., 2010) aiming to further investigate the cause of death in Pompeii and the other Roman towns buried by the 79 AD Vesuvius Plinian eruption (Mastrolorenzo et al., 2001a). Using histochemical techniques, Mastrolorenzo et al. (2010) showed the presence of DNA within osteocytic lacunae of bone samples experimentally heated up to 300 °C. Here we test the possibility of extracting and amplifying aDNA from human bone remains of Herculaneum victims showing macro- and microscopic evidence of exposure to temperatures of about 300 °C, using a combination of histological and molecular methods.
2. Materials and methods2.1. SamplingThe study of the human skeletal remains that formed the focus of this work was approved by the Ethics Committee for Biomedical Activities of the University of Naples, Hospital University of Naples Federico II (Protocol 154/10, 9.08.10). The Archaeological Superintendency of Pompeii granted permission for field investigations and the study of the human skeletal materials unearthed by one of the authors (PP) in the 1997–1999 excavations of the waterfront chambers at the Herculaneum archaeological area (Province of Naples, southern Italy) (Figure 1).
We analyzed several human skeletal remains unearthed from the volcanic surge deposit on the ancient beach of the Herculaneum archeological site.
The skeletons belonged to some of the 300 human victims instantly killed by the 79 AD eruption’s hot first surge (Mastrolorenzo et al., 2001). Skeletal remains were carefully handled by one of the authors (PP) and stored in hermetically sealed boxes until laboratory analyses. In particular, in order to examine bone samples that were most likely exposed to temperatures up to 300 °C at the 79 AD first pyroclastic surge emplacement, we selected four long bones with colors ranging from bright to dark reddish brown (about 300 °C) and showing bone macro- and microfractures typically induced by heat exposure (Shipman et al., 1984; Holden et al., 1995; Mastrolorenzo et al., 2010) (Table 1). Sex and age at death were assessed according to standard diagnostic procedure (Ferembach et al., 1980; Buikstra and Ubelaker, 1994). From each of the selected samples, a cross slice (about 400 mg and 4 mm thick) was taken through the midshaft using a hacksaw, also recording the condition of the bone according to Stiner et al. (1995) as follows: heavy, when the bone could be sectioned without cracking; friable, when the bone cracked and crumbled owing to cutting. These slices were used for both histological and molecular analyses (see below). 2.2. Sample pretreatment and contamination controlIn order to eliminate contaminating DNA from the bone surface, the bone slices were first scraped with a brush to remove the outer layers, subsequently immersed in 5.0% sodium hypochlorite for about 15 min, and finally exposed to UV light for 10 min at a distance of 20 cm (Cipollaro et al., 1998). Sample pretreatment as well as the successive laboratory steps (DNA extraction and PCR) were performed by a single operator (LPP) in a completely sterile room with standard precautions, including the use of workstations decontaminated by means of UV irradiation and bleach, ultrapure reagents, sterile disposables, and full-body personal protective clothing (Kemp and Smith, 2005; Gamba et al., 2011). PCR amplification did not take place in the same room as pre-PCR handling such as DNA extraction and PCR setup (Gilbert et al., 2005a; Elsner et al., 2015).
The bone slices were decalcified in 0.5 M ethylenediamine tetraacetic acid (EDTA), pH 7.4, for 7–13 days depending on their size and hardness. All the decalcified slices were transversally cut into two parts and used for the histological analysis and DNA extraction as reported below.2.3. Histological analysisDecalcified bone slices were embedded in autoclaved Tissue-Tek O.C.T. compound (Sakura, USA), frozen at –22 °C, and cross-sectioned by means of a cryostat. About 20 nonserial sections, 10 µm thick, were collected on slides and stained with Mayer’s hemalum or 4’,6’-diamidino-2-phenylindole (DAPI, Sigma) in order to explore the

GUARINO et al. / Turk J Biol
642
persistence of nuclei within osteocyte lacunae according to Guarino et al. (2000, 2006). The stained sections were observed under a Leica Motic BA340 transmitted light microscope or under a Leica DM100 fluorescence microscope, both equipped with a Nikon Coolpix 5000 digital camera.
2.4. DNA extraction and authenticityDecalcified bone slices were minced into very small pieces by means of forceps (method A) and incubated at 37 °C overnight in lysis buffer (Tris-HCl 50 mM, NaCl 100 mM, EDTA 10 mM, sodium dodecyl sulfate (SDS) 1%, proteinase K 1%) (Sokolov, 2000). After addition of
Figure 1. A) Location of Herculaneum archaeological site. B) The suburban area and the 12 chambers on the seashore. C) and D) Taphonomic context of skeletal remains analyzed in the present study: human victims within chambers 12 (C) and 10 (D).

GUARINO et al. / Turk J Biol
643
1/10 v/v of a KCl saturated solution and centrifugation at 14,000 rpm for 10 min, DNA was extracted with phenol-chloroform-isoamyl alcohol (25:24:1). DNA was precipitated in absolute ethanol (EtOH), exsiccated, and dissolved in 20 µL of 1X TE (Tris 10 mM, EDTA 1 mM). For DNA extraction we also used about 100 serial cryostat sections (see Section 2.3), collected in 500 µL of 70% EtOH (method B) and stored until DNA extraction, which was performed as above reported.
In addition, we used the laboratory operator’s (LPP) DNA previously extracted from his blood cells in a distinct laboratory. Negative controls (both reagents and products, for both methods A and B) were used in each step of molecular analysis. To verify the aDNA authenticity we performed multiple extractions and PCR in two different laboratories, each one with its own instruments and reagents (Gilbert et al., 2005a; Elsner et al., 2015).
All extracted DNAs were visualized with agarose gel (2%) electrophoresis using ethidium bromide (5 µg/mL). Where Maillard products (see O’Rourke et al., 2000) were visible, DNA was eluted from a gel trait ranging from about 50 bp to 1000 bp, using the ‘squeeze and freeze’ method (Tautz and Renz, 1983).
DNA concentration and purity were assessed using a NanoDrop 1000 (Thermo Scientific, USA), taking into considerations the methodological limits of this procedure (Rohland and Hofreiter, 2007). 2.5. PCR amplification and sequencingIn order to minimize the possibility of amplifying segments of foreign bacterial DNA we performed PCR reactions for specific regions of the single-copy nuclear gene of amelogenin (AMEL) X (106 bp) and Y (112 bp), useful for determining the sex of human remains (Cipollaro et al., 1998). Primers used for PCR amplification of AMEL were as follows (Mannucci et al., 1994): 5’ CCCTGGGCTCTGTAAAGAATAGTG 3’ (forward) and 5’ TCAGAGCTTAAACTGGGAAGCTG 3’ (reverse). PCR amplification was conducted in a total volume of 25 µL, using the Thermal Cycler GeneAmp PCR System 3400 (Applied Biosystem, USA) with the following conditions:
initial denaturation at 95 °C for 5 min followed by 36 cycles comprising 94 °C for 45 s (denaturation), 58 °C for 45 s (primer annealing), and 72 °C for 1 min (extension), with a final elongation step at 72 °C for 7 min. After purification with the Wizard SV Gel and PCR Clean-Up System kit (Promega, USA), PCR products were reamplified using the same conditions described above.
Amplicons were sequenced on an automated sequencer ABI 377 (Applied Biosystems) using the BIGDYE TERMINATOR v3.1 kit (ABI, USA). The sequences obtained were imported into BioEdit 7.0.5.3 (Hall, 1999) and aligned with ClustalW (Thomson et al., 1994).
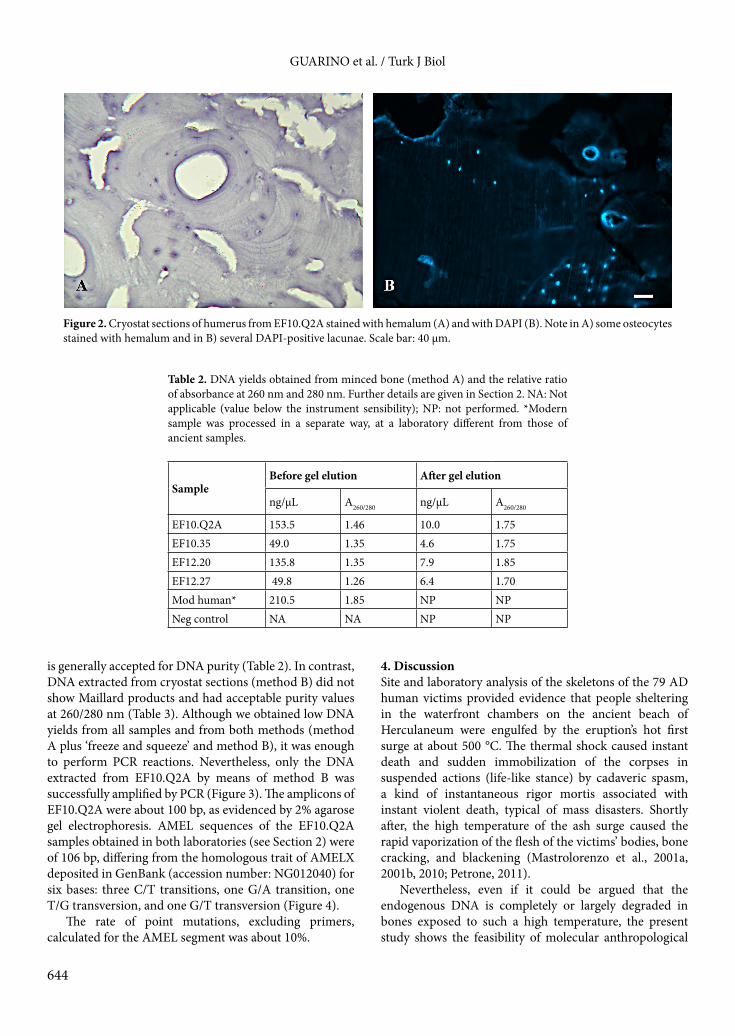
3. Results3.1. Histological analysisAll bone samples showed a good state of histological preservation (Figure 2). Analysis of the bone sections showed large areas with typical microstructure of human bone, albeit partly broken up owing to microcracking, typical of exposure to high temperatures close to 300 °C (Mastrolorenzo et al., 2010) and enhanced by the decalcification process. Signs of diagenesis due to microbial activity such as destructive microscopic foci, MDF, or other forms of bioerosion (Jans et al., 2004; White and Booth, 2014) were not found. Several osteocytes stained with hemalum as well as several DAPI-positive lacunae were observed in bone sections from EF10.Q2A, EF12.27, and EF10.35 (Figure 2), but not from EF12.20. 3.2. Molecular analysisDNA extractions and multiple PCR gave consistent results within and between the dedicated laboratories where the experimental procedures were carried out. Agarose electrophoretic gel of DNA from decalcified and fragmented Herculaneum bones (method A) showed an evident green band of Maillard products close to the front buffer. These products sensibly affected spectrophotometric quantifications of extracted DNA, overvaluing it (Table 2). In fact, after elution from gel bands by means of ‘freeze and squeeze’, for all samples the DNA values were lower and with a ratio of absorbance at 260/280 nm close to 1.8, which
Table 1. Human skeletal remains used in the present work.
Code Sex Bone Macroscopic appearance
EF10.Q2A F? Tibia dx Orange-reddish, heavyEF10.35 M Fibula sx Orange-reddish, heavyEF12.20 M Femur dx Orange-reddish, heavyEF12.27 M Humerus dx Orange-reddish, heavy
Sex was determined on the basis of anthropometric analysis. EF, Herculaneum remains; F?, probably a female.
GUARINO et al. / Turk J Biol
644
is generally accepted for DNA purity (Table 2). In contrast, DNA extracted from cryostat sections (method B) did not show Maillard products and had acceptable purity values at 260/280 nm (Table 3). Although we obtained low DNA yields from all samples and from both methods (method A plus ‘freeze and squeeze’ and method B), it was enough to perform PCR reactions. Nevertheless, only the DNA extracted from EF10.Q2A by means of method B was successfully amplified by PCR (Figure 3). The amplicons of EF10.Q2A were about 100 bp, as evidenced by 2% agarose gel electrophoresis. AMEL sequences of the EF10.Q2A samples obtained in both laboratories (see Section 2) were of 106 bp, differing from the homologous trait of AMELX deposited in GenBank (accession number: NG012040) for six bases: three C/T transitions, one G/A transition, one T/G transversion, and one G/T transversion (Figure 4).
The rate of point mutations, excluding primers, calculated for the AMEL segment was about 10%.
4. DiscussionSite and laboratory analysis of the skeletons of the 79 AD human victims provided evidence that people sheltering in the waterfront chambers on the ancient beach of Herculaneum were engulfed by the eruption’s hot first surge at about 500 °C. The thermal shock caused instant death and sudden immobilization of the corpses in suspended actions (life-like stance) by cadaveric spasm, a kind of instantaneous rigor mortis associated with instant violent death, typical of mass disasters. Shortly after, the high temperature of the ash surge caused the rapid vaporization of the flesh of the victims’ bodies, bone cracking, and blackening (Mastrolorenzo et al., 2001a, 2001b, 2010; Petrone, 2011).
Nevertheless, even if it could be argued that the endogenous DNA is completely or largely degraded in bones exposed to such a high temperature, the present study shows the feasibility of molecular anthropological
Figure 2. Cryostat sections of humerus from EF10.Q2A stained with hemalum (A) and with DAPI (B). Note in A) some osteocytes stained with hemalum and in B) several DAPI-positive lacunae. Scale bar: 40 µm.
Table 2. DNA yields obtained from minced bone (method A) and the relative ratio of absorbance at 260 nm and 280 nm. Further details are given in Section 2. NA: Not applicable (value below the instrument sensibility); NP: not performed. *Modern sample was processed in a separate way, at a laboratory different from those of ancient samples.
SampleBefore gel elution After gel elution
ng/µL A260/280 ng/µL A260/280
EF10.Q2A 153.5 1.46 10.0 1.75EF10.35 49.0 1.35 4.6 1.75EF12.20 135.8 1.35 7.9 1.85EF12.27 49.8 1.26 6.4 1.70Mod human* 210.5 1.85 NP NPNeg control NA NA NP NP

GUARINO et al. / Turk J Biol
645
studies on those bones from the 79 AD victims that show evidence of exposure to lower temperatures of about 300 °C. This temperature confirms the taphonomic evidence, indicating that heat effects locally might show some variability, mostly related to the density of corpses sheltered within each chamber: the fewer the people per chamber, the greater the effects of high temperature on victims (i.e. heat-induced skull fractures, teeth fractures, bone fractures and blackening, spine extension, hands and feet hyperflexion) and vice versa (Mastrolorenzo et al., 2001a, 2001b). In general, the heat of the ash was sufficient to vaporize most of the organic matter. Hence, the initial violent vaporization caused a rapid drop in ash temperature, as confirmed by paleomagnetic data (Incoronato et al., 1998). This could also explain why the most evident effects of the heat are limited to teeth and those parts of the skeleton least protected by fat and tissue, a process that is indeed particularly evident in less crowded chambers.
Our experimental evidence shows that archaeological bones clearly altered by high temperatures deserve to be considered for DNA analysis, reinforcing what was already pointed out by other studies (Zhang and Wu, 2005; Ottoni et al., 2009). Interestingly, the study of Ottoni et al. (2009) showed that in moderately heated (up to 140 °C) archaeological bone, DNA preservation is not related to the presence of intact collagen fibrils. A possible alternative explanation is that moderate heating inactivates nucleases and PCR inhibitors, destroys cell walls, and partially degrades collagens. As a consequence, there is both a greater ability of the bone crystals to bind DNA fragments and more DNA fragments available for absorption, resulting in greater preservation of DNA
(Ottoni et al., 2009). The crucial role of the hydroxyapatite crystals in DNA preservation within bone tissue has been shown by several studies (Gotherstrom et al., 2002; Salamon et al., 2005) and bone recrystallization begins when bone is exposed to temperatures equal to or higher than 600 °C (Holden et al., 1995; Quatrehomme et al., 1998), as recently detected in some bones of the 79 AD victims unearthed in Oplontis, a suburban village closed to Pompeii (Mastrolorenzo et al., 2010).
To date, a wide variety of methods (Rohland and Hofreiter, 2007) have been developed to solve the difficulties associated with recovery and analysis of aDNA, such as low amounts of DNA obtained, degraded templates, PCR inhibitors, etc. In this study we successfully applied for the first time a method that allows DNA extraction from osteocytes, placing cryostat sections of decalcified bone in 70% EtOH and then in proteinase K plus SDS digestion solution. This procedure offers a double advantage. First, PCR inhibition byproducts of sugar reduction such as Maillard products (O’Rourke et al., 2000) were not observed. Probably the low thickness of the sections (12 µm) allows the removal of most of the PCR inhibitors during their storage in ethanol prior to the next steps for DNA extraction. Interestingly, Maillard products were observed in the gel electrophoresis of genomic DNA
Table 3. DNA yields obtained from cryostat sections (method B) and the relative ratio of absorbance at 260 nm and 280 nm. Further details are given in Section 2.
Sample ng/µL A260/280
EF10.Q2A 17.5 1.73EF10.35 4.5 1.74EF12.20 6.5 1.70EF12.27 6.9 1.78
Figure 3. The 2% agarose gel electrophoresis of amplicons from method B (cryostat sections). From left to right, lane 1: EF10.Q2A; lane 2: EF10.35; lane 3: EF12.27; lane 4: EF12.20; lane 5: modern human; 6: blank; lane L: ladder 100 bp (New England Biolabs Inc., UK).
Figure 4. Comparison and alignment by ClustalW of the canonical AMELX and EF10.Q2A DNA sequences. C→T and G→A transitions are evidenced by dark gray (light blue); T→G and A→T transversions in light gray (yellow). In EF10.Q2A lowercase letters indicate primer base pairs for AMEL.

GUARINO et al. / Turk J Biol
646
obtained by means of the mincing procedure (method A). Furthermore, it should be noted that the EF10.Q2A sample also did not give amplification products using method A after gel elution. This is probably because PCR inhibitors were not totally removed. In addition, although the EF10.35, EF12.20, and EF12.27 samples gave measurable DNA by method B (cryostat method), they were not successfully amplified, probably because their DNA was more fragmented than that of EF10.Q2A.
Second, the cryostat method allows to select the bone samples to be processed for molecular analysis on the basis of DNA detection in the osteocytic lacunae by histochemical staining.
In our study, only for the EF10.Q2A sample were yields and quality of aDNA suitable for direct amplification and sequencing of short single-locus sequences, such as AMEL. The length (106 bp) of the obtained amplicons was consistent with the target AMEL fragment and their nucleotide sequence differed from that deposited in GenBank for six bases. This finding allowed us to attribute the EF10.Q2A sample to a female, solving the uncertain sex determination based on anthropometric analysis. On the other hand, if the EF10.Q2A sample was of a male, the direct sequencing of AMEL would have produced unreadable chromatograms owing to the presence of two sequences of different length, 106 bp (AMELX) and 112 bp (AMELY). The difference in six bases of our trait of sequence AMELX from that deposited in GenBank is mainly due to pyrimidine transitions. This kind of point mutations is commonly observed in nuclear sequences of aDNA, because they are supposed to be more prone than purines to damages by oxidation (Hansen et al., 2001; Gilbert et al., 2003, 2005b; Binladen et al., 2006). Furthermore, even if referred to one datum, our AMELX sequence displays a rate of point mutations threefold that observed in nuclear genes of aDNA by Binladen et al. (2006). This higher
rate could be explained as a consequence of the changes induced by the high temperatures, also considering the diagenetic alterations that occurred postmortem. Further studies on Herculaneum skeletal remains or other archaeological bones with similar heat alteration could support our hypothesis.
It is generally accepted by aDNA researchers that the criteria list of Cooper and Poinar (2000) as the basis to test the authenticity and reliability of data (Gilbert et al., 2005a) should be used. In our study we did not consider it necessary to clone amplicons because our chromatograms did not show any ambiguous nucleotide positions. On the other hand, as showed by Winters et al. (2011) in their study on aDNA, the consensus sequence of clones does not differ from the direct sequence.
To conclude, the main goal of the present paper was to demonstrate PCR amplifiability of DNA recovered from archaeological skeletal remains of the victims killed by the first hot pyroclastic surge at Herculaneum. The results achieved in the present study are the first successful attempt to analyze Herculaneum archaeological human bones at the molecular level by means of amplification of single-copy gene sequences. Our data, although based only on a positive sample, indicate that even ancient bones showing signs of thermal exposure up to 300 °C, as in the case of the 79 AD victims skeletons, should be considered for DNA analysis. This opens up the chance to carry out systematic studies about the genetic characteristics of the ancient population of Herculaneum, as could also be done for that of Pompeii.
AcknowledgmentsWe thank the Superintendence of Pompeii for granting a permit for field investigation and for the study of the human skeletal material. We also thank Mark Walters for reviewing the English version of the manuscript.
References
Bailey JF, Henneberg M, Colson IB, Ciarallo A, Hedges RE, Sykes B (1999). Monkey business in Pompeii- unique find of a juvenile barbary macaque skeleton in Pompeii identified using osteology and ancient DNA techniques. Mol Biol Evol 16: 1410-1414.
Binladen J, Wiuf C, Gilbert TM, Bunce M, Barnett R, Larson G, Greenwood AD, Haile J, Ho YW, Hansen AJ et al. (2006). Assessing the fidelity of ancient DNA sequences amplified from nuclear genes. Genetics 172: 733-74.
Brown T, Brown K (2011). Biomolecular Archaeology: An Introduction. Oxford, UK: John Wiley and Sons.
Buikstra JE, Ubelaker DH (1994). Standards for Data Collection from Human Skeletal Remains. Research Series No. 44. Fayetteville, AR, USA: Arkansas Archaeological Survey.
Cipollaro M, Di Bernardo G, Forte A, Galano G, De Masi L, Galderisi U, Angelini, F, Cascino A (1999). Histological analysis and ancient DNA amplification results on human bone remains found in Caius Iulius Polybius house in Pompeii. Croatian Med J 40: 392-397.
Cipollaro M, Di Bernardo G, Galano G, Galderisi U, Guarino FM, Angelini F, Cascino A (1998). Ancient DNA in human bone remains from Pompeii archeological site. Biochem Biophys Res Comm 247: 901-904.
Cooper A, Poinar H (2000). Ancient DNA: do it right or not at all. Science 289: 1139.
Di Bernardo G, Del Gaudio S, Galderisi U, Cascino A, Cipollaro M (2009). Ancient DNA and family relationships in a Pompeian house. Ann Hum Gen 73: 429-437.

GUARINO et al. / Turk J Biol
647
Di Bernardo G, Del Gaudio S, Galderisi U, Cipollaro M (2004a). 2000 Year-old ancient equids: an ancient-DNA lesson from Pompeii remains. J Exp Zool Part B 302: 550-556.
Di Bernardo G, Galderisi U, Del Gaudio S, D’Aniello A, Lanave C, De Robertis MT, Cascino A, Cipollaro M (2004b). Genetic characterization of Pompeii and Herculaneum Equidae buried by Vesuvius in 79 AD. J Cell Phys 199: 200-205.
Elsner J, Schibler J, Hofreiter M, Schlumbaum A (2015). Burial condition is the most important factor for mtDNA PCR amplification success in Palaeolithic equid remains from the Alpine foreland. Archaeol Anthropol Sci 7: 505-515.
Ferembach D, Schwidetzky I, Stloukal M (1980). Recommendations for age and sex diagnoses of skeletons. J Hum Evol 9: 517-549.
Gamba C, Fernandez E, Tirado M, Pastor F, Arroyo-Pardo E (2011). Ancient nuclear DNA and kinship analysis: the case of a medieval burial in San Esteban Church in Cuellar (Segovia, Central Spain). Am J Phys Anthropol 144: 485-491.
Geraci G, Del Gaudio R, Di Giaimo R (2002). Le analisi paleogenetiche. In: Petrone PP, Fedele F, editors. Vesuvio 79 A.D. Vita e morte ad Ercolano. Naples, Italy: Fridericiana Napoli, pp. 85-88 (in Italian).
Gilbert MTP, Bandelt HJ, Hofreiter M, Barnes I (2005a). Assessing ancient DNA studies. TRENDS Ecol Evol 20: 541-544.
Gilbert MTP, Hansen AJ, Willerslev E, Rudbeck L, Barnes I, Lynnerup N, Cooper A (2003). Characterization of genetic miscoding lesions caused by postmortem damage. Am J Hum Gen 72: 48-61.
Gilbert MTP, Rudeck L, Willerslev E, Hansen AJ, Smith C, Penkman KEH, Prangerberg K, Nielsen-March CM, Jans ME, Arthur P et al. (2005b). Biochemical and physical correlates of DNA contamination in archaeological human bones and teeth excavated at Matera, Italy. J Archaeol Sci 32: 785-793.
Gotherstrom A, Collins MJ, Angerbjorn A, Liden K (2002). Bone preservation and DNA amplification. Archaeometry 44: 395-404.
Guarino FM, Angelini F, Odierna G, Bianco MR, Di Bernardo G, Forte A, Cascino A, Cipollaro M (2000). Detection of DNA in ancient bones using histochemical methods. Biotech Histochem 75: 110-117.
Guarino FM, Angelini F, Vollono C, Orefice C (2006). Bone preservation in human remains from the Terme del Sarno at Pompeii using light microscopy and scanning electron microscopy. J Archaeol Sci 33: 513-520.
Hall TA (1999). BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41: 95-98.
Hansen A, Willerslev E, Wiuf C, Mourier T, Arctander P (2001). Statistical evidence for miscoding lesions in ancient DNA templates. Mol Biol Evol 18: 262-265.
Holden JL, Phakey PP, Clement JG (1995). Scanning electron microscope observations of heat-treated human bone. For Sci Int 74: 29-45.
Hoss M, Jaruga P, Zastawny TH, Dizdaroglu M, Pääbo S (1996). NA damage and DNA sequence retrieval from ancient tissues. Nucleic Acids Res 24: 1304-1307.
Incoronato A, Mastrolorenzo G, Pagano M, Spina G (1998). Determinazione delle temperature di deposizione, mediante procedure paleomagnetiche di piroclastiti del 79 A.D. ad Ercolano. In: D’amico C, Livadie A, editors. Le scienze della Terra e l’archeometria. Naples, Italy: Pàtron, pp 95-98 (in Italian).
Jans MM, Nielsen-Marsh CM, Smith CI, Collins H, Kars H (2004). Characterization of microbial attack on archaeological bone. J Archaeol Sci 31: 87-95.
Kaestle FA, Horsburgh KA (2002). Ancient DNA in anthropology: methods, applications, and ethics. Am J Phys Anthropol 35 (Suppl.): 92-130.
Kemp BM, Smith DG (2005). Use of bleach to eliminate contaminating DNA from the surface of bones and teeth. For Sci Int 154: 53-61.
Lindahl T (1993). Instability and decay of the primary structure of DNA. Nature 362: 709-715.
Mannucci A, Sullivan K, Ivanov PL, Gill P (1994). Forensic application of a rapid quantitative DNA sex test by amplification of the X-Y homologous gene amelogenni. Int J Legal Med 106: 190-193.
Mastrolorenzo G, Petrone PP, Pagano M, Incoronato A, Baxter PJ, Canzanella A, Fattore L (2001a). Herculaneum victims of Vesuvius in AD 79. Nature 410: 769-770.
Mastrolorenzo G, Petrone PP, Pagano M, Incoronato A, Baxter PJ, Fattore L, Fergola L (2001b). The 79 AD Vesuvius Plinian eruption at Herculaneum and its impact on the people. In: Juvigne E, Raynal JP, editors. Tephras. Les dossiers de l’Archéo-Logis, n°1. Lausanne, France: CDERAD Goudet, pp. 183-189.
Mastrolorenzo G, Petrone P, Pappalardo L, Guarino FM (2010). Lethal thermal impact at periphery of pyroclastic surges: evidences at Pompeii. PLoS ONE 5 1-12 e11127.
O’Rourke DH, Hayes MG, Carlyle S (2000). Ancient DNA studies in physical anthropology. Ann Rev Anthropol 29: 217-242.
Ottoni C, Koon HEC, Collins MJ, Penkman KEH, Rickards O, Craig OE (2009). Preservation of ancient DNA in thermally damaged archaeological bone. Naturwissenschaften 96: 267-278.
Pääbo S (1989). Ancient DNA: extraction, characterization, molecular cloning, and enzymatic amplification. P Natl Acad Sci USA 86: 939-1943.
Petrone PP (2011). Human corpses as time capsules: new perspectives in the study of past mass disasters. J Anthropol Sci 89: 1-4.
Pruvost M, Schwartz R, Correia VB, Champlot S, Braguier S, Morel N, Jalvo Y, Grange T, Gelgi EM (2007). Freshly excavated fossil bones are best for amplification of ancient DNA. P Natl Acad Sci USA 104: 739-744.
Quatrehomme G, Bolla M., Muller M, Rocca JP, Grevin G, Bailet P, Ollier A (1998). Experimental single controller study of burned bones: contribution of scanning electron microscopy. J For Sci 43: 417-422.

GUARINO et al. / Turk J Biol
648
Rohland N, Hofreiter M (2007). Comparison and optimization of ancient DNA extraction. BioTechniques 42: 343-352.
Salamon M, Tuross N, Arensburg B, Weiner S (2005). Relatively well preserved DNA is present in the crystal aggregates of fossil bones. P Natl Acad Sci USA 102: 13783-13788.
Shipman P, Foster G, Schoeninger M (1984). Burnt bones and teeth: an experimental study of colour, morphology, crystal structure and shrinkage. J Archaeol Sci 11: 307-325.
Sokolov EP (2000). An improved method for DNA isolation from mucopolysaccharide-rich molluscan tissue. J Moll Study 66: 573-575.
Stiner MC, Kuhn S, Bor-Yosef O (1995). Differential burning, recrystallization, and fragmentation of archaeological bone. J Archaeol Sci 22: 223-237.
Tautz D, Renz M (1983). An optimized freeze-squeeze method for the recovery of DNA fragments from agarose gels. Anal Biochem 132: 14-19.
Thompson JD, Higgins DG, Gibson TJ (1994). Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acid Res 22: 4673-4680.
White L, Booth TJ (2014). The origin of bacteria responsible for bioerosion to the internal bone microstructure: results from experimentally-deposited pig carcasses. For Sci Int 239: 92-102.
Winters M, Barta JL, Monroe C, Kemp BM (2011). To clone or not to clone: methods analysis for retrieving consensus sequences in ancient DNA sample. PLoS One 6: e21247.
Zhang L, Wu Q (2005) Single gene retrieval from thermally degraded DNA. J Biosci 30: 599-604.





























