Embed Size (px)
Citation preview

Biochem. J. (1973) 133, 701-708Printed in Great Britain
Reduced Nicotinamide-Adenine Dinucleotide-Nitrite Reductase fromAzotobacter chroococcum
By J. M. VEGA, M. G. GUERRERO, E. LEADBETTER* and M. LOSADADepartamento de Bioquimica, Facultad de Ciencias y Consejo Superior de Investigaciones
Cientificas, Universidad de Sevilla, Seville, Spain(Received 12 February 1973)
1. The assimilatory nitrite reductase of the N2-fixing bacterium Azotobacter chroococcumwas prepared in a soluble form from cells grown aerobically with nitrate as the nitrogensource, and some of its properties have been studied. 2. The enzyme is a FAD-dependentmetalloprotein (mol.wt. about 67000), which stoicheiometrically catalyses the directreduction ofnitrite to NH3 withNADH as the electron donor. 3. NADH-nitrite reductasecan exist in two either active or inactive interconvertible forms. Inactivation in vitro can beachieved by preincubation withNADH. Nitrite can specifically protect the enzyme againstthis inactivation and reverse the process once it has occurred. 4. A. chroococcum nitritereductase is an adaptive enzyme whose formation depends on the presence of eithernitrate or nitrite in the nutrient solution. 5. Tungstate inhibits growth of the micro-organism very efficiently, by competition with molybdate, when nitrate is the nitrogensource, but does not interfere when nitrite or NH3 is substituted for nitrate. The additionof tungstate to the culture media results in the loss of nitrate reductase activity but doesnot affect nitrite reductase.
The enzyme involved in the assimilatory reductionof nitrite to NH3 in algae and higher plants has beenthoroughly characterized in recent years and classi-fied as ferredoxin-nitrite reductase (Beevers & Hage-man, 1969; Hewitt, 1970; Kessler, 1971; Losada,1972). Nitrite reductase purified to homogeneityfrom green cells of different sources (Chlorella,spinach and squash leaves) contains 2 atoms of non-haem iron/molecule of 63000 daltons and does notseem to be a flavoprotein (Losada & Paneque, 1971;Cardenas et al., 1972a,b; Zumft, 1972). On the otherhand, the enzyme isolated from the nitrate-assimilat-ing fungi Neurospora crassa (Nason et al., 1954;Nicholas et al., 1960) and Torulopsis nitratophila(Rivas et al., 1973) has been characterized as anNAD(P)H-nitrite reductase which specifically re-quires FAD and some metal component(s).By contrast with nitrite reductases from the plant
kingdom, information on nitrite reductase from bac-teria of the assimilatory nitrate-reducing type is veryscanty (Nason, 1962; Takahashi et al., 1963; Hewitt& Nicholas, 1964). Spencer et al. (1957) found in ex-tracts of Azotobacter vinelandii a soluble nitrite andhydroxylamine reductase system, which utilized re-duced nicotinamide nucleotides as electron donorsand required added flavin nucleotides for maximalactivity. Inhibitor studies indicated that the systemhad an essential metal component. The product of the
* Present address: Department of Biology, AmherstCollege, Amherst, Mass. 01002, U.S.A.
Vol. 133
reduction of nitrite by the extracts was identified asNH3, whereas that of hydroxylamine reduction wasnot established. Both nitrite reductase and hydroxyl-amine reductase were adaptive enzymes, their forma-tion being stimulated by nitrate. Escherichia colistrain Bn grown in deep standing cultures with nitrateas the sole source of nitrogen has been shown to con-tain at least three nitrite reductases that reducednitrite (and hydroxylamine) to NH3 (Lazzarini &Atkinson, 1961), but only the enzyme specific forNADH appears to be responsible for physiologicalnitrite reduction (Kemp & Atkinson, 1966). Zarowny& Sanwal (1963) have also observed a NADH-specific nitrite reductase in extracts of E. coli K-12grown with nitrate as nitrogen source. Prakash &Sadana (1972) have obtained from Achromobacterfisheri grown on nitrate under low 02 partial pressurea homogenous haemoprotein that catalyses the reduc-tion of nitrite (and hydroxylamine) to NH3 and havepointed out that any biosynthetic function of theenzyme appears fortuitous.The present paper describes the preparation and
characterization ofa soluble NADH-nitrite reductaseof the assimilatory type from Azotobacter chroococ-cum cells aerobically grown on nitrate as the nitrogensource. The enzyme is a FAD-dependent metallo-protein (mol.wt. about 67000) of adaptive nature thatstoicheiometrically catalyses the direct reduction ofnitrite to NH3 and exists in either active or inactiveinterconvertible forms.
701

J. M. VEGA, M. G. GUERRERO, E. LEADBETTER AND M. LOSADA
Materials and MethodsCell cultureA. chroococcum (strain A.T.C.C. 4412, from Sala-
manca University's culture collection) was grownunder air with vigorous shaking at 27°C on a syn-thetic medium containing (mmol/litre): mannitol, 39;KNO3, 8; MgSO4,7H20, 0.8; CaCl2, 2H20, 0.34;FeSO4,7H20, 0.19; NaCl, 3.4; Na2MoO4, 0.01;disodium EDTA, 0.02; potassium phosphate buffer,pH7.3, 10. When NH3 or NH4NO3 replaced KNO3as the source ofnitrogen, total concentration ofnitro-gen was also maintained at 8mM. Since nitrite wastoxic at concentrations higher than 1 mm, care wastaken not to exceed this limit when NaNO2 orNaNO2 plus (NH4)2S04 was the nitrogen source. Inexperiments involving the effect of tungstate, molyb-date was omitted from the standard nutrient solutionand tungstate was added as indicated; the cells usedfor inoculation in these experiments were grown onnitrate media lacking added molybdate.
Cell-free extractsCells (lOg fresh wt.) were harvested in the exponen-
tial phase of growth by low-speed centrifugation,washed with 50mM-potassium phosphate buffer,pH 7.0, and resuspended in 3ml of the same buffer.The crude extract was prepared by breaking the cellsat 2°C for 2min in a vibration homogenizer (Buhler)with 100ml of glass beads (0.1mm diam.). The bro-ken material was extracted with 50ml of thephosphate buffer and after centrifugation for 20minat 40000g the supernatant was used as enzyme sourcefor the preparation of nitrite reductase.
Nitrite reductaseSince the crude extract possessed high NADH oxi-
dase activity, it was centrifuged at 2°C in a L2-50BSpinco ultracentrifuge for 3h at 120000g and theresulting supernatant, which was practically free ofoxidase activity, was used as the nitrite reductasepreparation. The enzyme was unusually unstablewhen purification was attempted.
Enzyme assaysTwo different methods were used for the assay of
nitrite reductase activity. NADH-nitrite reductasewas measured by following the nitrite-dependent oxi-dation ofNADH. The reaction was carried out underair at room temperature and absorbance at 340nmwas recorded with a Beckman DK-2A spectrophoto-meter, or exceptionally (for the measurement of theK. value for NADH) fluorescence at 450nm was re-corded with an Aminco-Bowman 4-8202 spectro-photofluorimeter. The reaction mixture included, in afinal volume of 2ml: potassium phosphate buffer,pH7.0. 150,mol; NADH, 0.3,umol; NaNO2,
2,umol; an appropriate amount of enzyme pre-paration. Although high-speed centrifugation prac-tically removed theNADH oxidase activity originallypresent in the crude extracts, NADH-nitrite reduc-tase activity was always corrected for the rate ofNADH oxidation in the absence of nitrite. ReducedMethyl Viologen-nitrite reductase was determined bynitrite disappearance (Ramirez et al., 1966), but withpotassium phosphate buffer, pH 7.0, instead of Tris-HC1 buffer. One unit of activity corresponds to theoxidation of 1 ,tmol ofNADH (or the stoicheiometricreduction of ijmol of nitrite)/min.
Nitrate reductase activity was measured by colori-metrically following the appearance of nitrite. Thereaction mixture was incubated at 30°C for 2min andcontained, in a final volume of 1 ml: potassium phos-phate buffer, pH7.0, 100,umol; KNO3, 10,umol;Methyl Viologen, 0.15,umol; Na2S204 in 0.1ml of95mM-NaHCO3, 0.8mg; an appropriate amountof enzyme preparation. The reaction was stopped byvigorously shaking the assay mixture. Enzyme activityunits are expressed as jumol of nitrite formed/min.
Analytical methodsGrowth was determined by measuring absorbance
changes at 660nm. Nitrite was measured by themethod of Snell & Snell (1949), and nitrate withnitrate reductase from spinach leaves as described byRelimpio et al. (1972). NH3 was determined by treat-ment with Nessler's reagent after diffusion andabsorption of the gas in 5mM-H2SO4 in Conwayunits (Conway, 1957). Protein was measured by themethod of Lowry et al. (1951) with ovalbumin as astandard.The molecular weight of NADH-nitrite reductase
was estimated by sucrose-density-gradient centri-fugation (Martin & Ames, 1961). Samples (0.2ml) ofnitrite reductase and several other proteins ofknownmolecular weight (catalase, 240000; glucose 6-phos-phate dehydrogenase, 110000; serum albumin,67000) were layered on the top of a 3.4ml linear5-20% (w/v) sucrose gradient layered over 0.1 5ml of50% (w/v) sucrose. Sucrose was dissolved in 50mM-potassium phosphate buffer, pH 7.0. Centrifugationwas carried out at 45000 rev./min for 11 h at 2°C in aSpinco L2-50B ultracentrifuge with a SW-56 Ti rotor.Gradients were fractioned from the bottom of thetube with a Densi-Flow apparatus from BuchlerInstruments Inc., Fort Lee, N.J., U.S.A. Fractions(3 drops) were collected with a LKB fraction col-lector. Protein and enzyme activities were then deter-mined in samples of each fraction.
ResultsElectron donors and cofactors
Table 1 shows thatNADH was an effective electrondonor for the reduction of nitrite catalysed by A.
1973
702
NADH-NITRITE REDUCTASE FROM AZOTOBACTER CHROOCOCCUM
Table 1. Electron donors and cofactors for nitritereductasefrom A. chroococcum
In the experiments with reduced nicotinamide nucleo-tides as electron donors, the reaction mixture con-tained, inafinal volume of2ml: potassium phosphatebuffer, pH7.0, 150,umol; enzyme preparation, 0.4mg;NaNO2, 2,umol; NAD(P)H, 0.3,mol; where indi-cated FAD, 5nmol. NAD(P)H oxidation was fol-lowed by the change in absorbance at 340nm. Otherexperimental conditions were the same as in thestandard assay for NADH-nitrite reductase activity.In the experiments with dithionite as reductant thereaction mixture contained in a final volume of 2ml:potassium phosphate buffer, pH7.0, 150,umol;enzyme preparation, 2mg; sodium dithionite, 7.5mgin 0.3 ml of a 2.5 % (w/v) NaHCO3 solution; NaNO2,4,tmol; where indicated Methyl or Benzyl Viologen,1.5,umol; FAD or FMN, 1 .5,umol. The reaction wascolorimetrically followed by nitrite disappearance.Other experimental conditions were the same as in thestandard assay for Methyl Viologen-nitrite reductaseactivity.
chroococcum nitrite reductase. The reaction wasspecifically stimulated about twofold by the additionof FAD; FMN was without effect. NADPH was arelatively ineffective electron donor compared withNADH and the addition ofFAD gave no additionalstimulation. The reducing agent sodium dithionitewas also operative as electron donor for the enzymicreduction of nitrite. The reaction was stimulatedabout twofold by the addition of Methyl Viologen asan artificial electron carrier, but Benzyl Viologenand flavin nucleotides were without effect.
Enzymic reduction ofnitrite with NADHThe characterization of the reaction catalysed by
nitrite reductase with NADH as electron donor isshown in Table 2. The oxidation of NADH wasdependent on the presence of nitrite and no reactiontook place when the enzyme preparation was omittedor boiled for 5min. In the absence of nitrite, the addi-tion ofeither 1 mM-hydroxylamine or 1 mM-sulphite tothe reaction mixture did not bring about any oxidationof NADH, even in the presence of FAD.
Electron donorsand cofactors
NADHNADH, FADNADPHNADPH, FADS2042, Methyl ViologenS2042-, Benzyl ViologenS2042, FADS2042-, FMNS2042None
NAD(P)H oxidized ornitrite reduced
(nmol/min per mgof protein)
35770566661921201241201240
Table 2. Reduction ofnitrite by A. chroococcum nitritereductase with NADH as electron donor
Experimental conditions were the same as in thestandard assay for NADH-nitrite reductase activity.Enzyme preparation, 0.6mg.
SystemCompleteMinus NADHMinus NO2-Minus nitrite reductaseComplete, nitrite reductase
heated (Smin at 100°C)
NADH oxidized(nmol/min)
950800
0 0.1 0.3 0.5
/[NO2- (HM-
90
60 [
30
0 30 60 90
1/[NADHJ (mM-l)Fig. 1. Reciprocalplots ofthe effect of(a) nitrite and (b) NADH concentrations on A. chroococcum nitrite reductase
activity
Experimental conditions were as in the standard assay for NADH-nitrite reductase activity, except that nitrite(a) or NADH (b) were added as indicated. Enzyme preparation, 0.6 and 0.2mg respectively.Vol. 133
(a)30
;5laz; 1
(b)
r~~~~~~~
703
J. M. VEGA, M. G. GUERRERO, E. LEADBETTER AND M. LOSADA
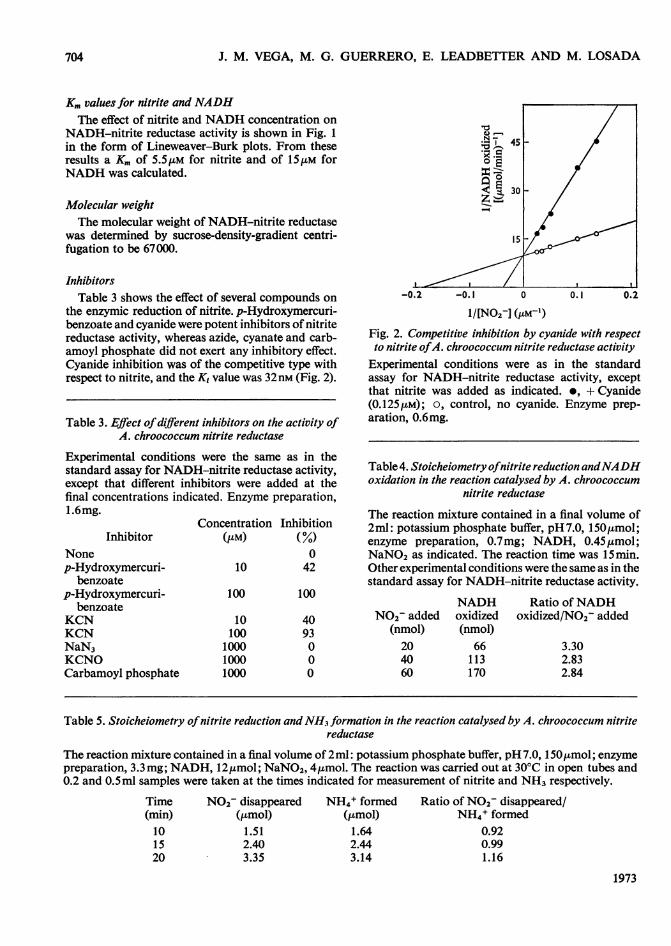
Km values for nitrite and NADHThe effect of nitrite and NADH concentration on
NADH-nitrite reductase activity is shown in Fig. 1in the form of Lineweaver-Burk plots. From theseresults a Km of 5.5 tM for nitrite and of 15 tM forNADH was calculated.
Molecular weightThe molecular weight of NADH-nitrite reductase
was determined by sucrose-density-gradient centri-fugation to be 67000.
InhibitorsTable 3 shows the effect of several compounds on
the enzymic reduction of nitrite. p-Hydroxymercuri-benzoate and cyanide were potent inhibitors of nitritereductase activity, whereas azide, cyanate and carb-amoyl phosphate did not exert any inhibitory effect.Cyanide inhibition was of the competitive type withrespect to nitrite, and the K, value was 32nM (Fig. 2).
Table 3. Effect ofdifferent inhibitors on the activity ofA. chroococcum nitrite reductase
Experimental conditions were the same as in thestandard assay for NADH-nitrite reductase activity,except that different inhibitors were added at thefinal concentrations indicated. Enzyme preparation,1.6mg.
InhibitorNonep-Hydroxymercuri-
benzoatep-Hydroxymercuri-
benzoateKCNKCNNaN3KCNOCarbamoyl phosphate
Concentration Inh(VM)
10
100 I
10100
100010001000
-0.2
1/[NO2-] (PM-')Fig. 2. Competitive inhibition by cyanide with respectto nitrite ofA. chroococcum nitrite reductase activity
Experimental conditions were as in the standardassay for NADH-nitrite reductase activity, exceptthat nitrite was added as indicated. *, + Cyanide(0.125[tm); O, control, no cyanide. Enzyme prep-aration, 0.6mg.
Table 4. Stoicheiometry ofnitrite reduction andNADHoxidation in the reaction catalysed by A. chroococcum
nitrite reductase
.ibition The reaction mixture contained in a final volume ofibition 2ml: potassium phosphate buffer, pH7.0, 150,umol;(Y/o) enzyme preparation, 0.7mg; NADH, 0.45,.tmol;0 NaNO2 as indicated. The reaction time was 15min.
42 Other experimental conditions were the same as in thestandard assay for NADH-nitrite reductase activity.
100
4093000
NADHNO2- added oxidized
(nmol) (nmol)20 6640 11360 170
Ratio ofNADHoxidized/NO2- added
3.302.832.84
Table 5. Stoicheiometry ofnitrite reduction and NH3 formation in the reaction catalysed by A. chroococcum nitritereductase
The reaction mixture contained in a final volume of 2ml: potassium phosphate buffer, pH 7.0, 150,umol; enzymepreparation, 3.3mg; NADH, 12,umol; NaNO2, 4,umol. The reaction was carried out at 30°C in open tubes and0.2 and 0.5 ml samples were taken at the times indicated for measurement of nitrite and NH3 respectively.
N02- disappeared(,umol)1.512.403.35
NH4+ formed(,tmol)1.642.443.14
Ratio of N02- disappeared/NH4+ formed
0.920.991.16
Time(min)101520
1973
704
NADH-NITRITE REDUCTASE FROM AZOTOBACTER CHROOCOCCUM
Stoicheiometry ofNADH oxidation, nitrite reductionand NH3 formationAs shown in Table 4, the reduction of nitrite by
nitrite reductase was concomitant with the oxidationof stoicheiometric amounts of NADH: 3 mol of thenucleotide was oxidized/mol of nitrite reduced whenthe reaction was complete. The product of the en-zymic reduction of nitrite was identified as NH3 andthe stoicheiometry of nitrite reduction and NH3formation was established: 1 mol of nitrite was con-sumed/mol of NH3 formed (Table 5).
Interconvertible active and inactive forms of nitritereductase
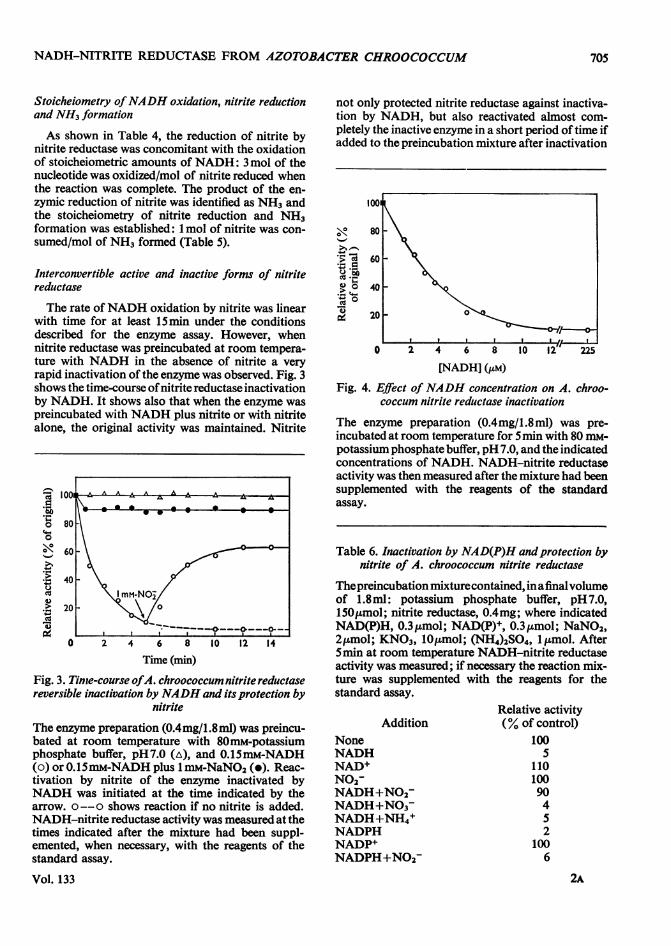
The rate ofNADH oxidation by nitrite was linearwith time for at least 15min under the conditionsdescribed for the enzyme assay. However, whennitrite reductase was preincubated at room tempera-ture with NADH in the absence of nitrite a veryrapid inactivation of the enzyme was observed. Fig. 3shows the time-course ofnitrite reductase inactivationby NADH. It shows also that when the enzyme waspreincubated with NADH plus nitrite or with nitritealone, the original activity was maintained. Nitrite
*E0._-0
:.t._
cE
0 2 4 6 8 10 12 14Time (min)
Fig. 3. Tinte-course ofA. chroococcum nitrite reductasereversible inactivation by NADH and its protection by
nitrite
The enzyme preparation (0.4mg/1.8ml) was preincu-bated at room temperature with 80mM-potassiumphosphate buffer, pH7.0 (A), and 0.15mM-NADH(o) or 0.15mM-NADH plus 1 mM-NaNO2 (e). Reac-tivation by nitrite of the enzyme inactivated byNADH was initiated at the time indicated by thearrow. o --o shows reaction if no nitrite is added.NADH-nitrite reductase activity was measured at thetimes indicated after the mixture had been suppl-emented, when necessary, with the reagents of thestandard assay.
Vol. 133
not only protected nitrite reductase against inactiva-tion by NADH, but also reactivated almost com-pletely the inactive enzyme in a short period of time ifadded to the preincubation mixture after inactivation
II-I
. t4
A,-4o4 t-
*4. ocd
[NADHI (uM)
Fig. 4. Effect ofNADH concentration on A. chroo-coccum nitrite reductase inactivation
The enzyme preparation (0.4mg/1.8ml) was pre-incubated at room temperature for 5min with 80 mm-potassium phosphate buffer, pH 7.0, and the indicatedconcentrations of NADH. NADH-nitrite reductaseactivity was then measured after the mixture had beensupplemented with the reagents of the standardassay.
Table 6. Inactivation by NAD(P)H andprotection bynitrite of A. chroococcum nitrite reductase
Thepreincubation mixturecontained, inafinalvolumeof 1.8ml: potassium phosphate buffer, pH7.0,150,umol; nitrite reductase, 0.4mg; where indicatedNAD(P)H, 0.3,mol; NAD(P)+, 0.3,umol; NaNO2,2,umol; KNO3, 10,umol; (NH4)2SO4, lumol. After5min at room temperature NADH-nitrite reductaseactivity was measured; if necessary the reaction mix-ture was supplemented with the reagents for thestandard assay.
AdditionNoneNADHNAD+NO2-NADH+NO2-NADH+NO3-NADH+NH4+NADPHNADP+NADPH+NO2-
Relative activity(% of control)
1005
11010090452
1006
2A
705

J. M. VEGA, M. G. GUERRERO, E. LEADBETTER AND M. LOSADA
Table 7. Effect ofthe inorganic nitrogen source on thecellular activity ofnitrite reductase in A. chroococcum
Cells were grown on media with the indicated nitrogensources. After 12h cell-free extracts were prepared andcentrifuged at high speed and NADH-nitrite reduc-tase activity was measured in the resulting superna-tants. Other experimental conditions are describedin the Materials and Methods section.
Nitrogen sourceN2N2 +(NH4)2SO4N2+KNO3N2+NH4NO3N2+NaNO2N2+NaNO2 +(NH4)2SO4
Nitrite reductase(munits/mg ofprotein)
00
12.79.87.3
12.4
Table 8. Effect oftungstate on the enzymic activities ofthe nitrate-reducing system in A. chroococcum
Cells were grown on nitrate media lacking addedmolybdate and containing the indicated amounts oftungstate. After 14h cell-free extracts were preparedand centrifuged at high speed and the activities ofnitrate reductase and nitrite reductase were measuredin the resulting supematants. Other experimentalconditions are described in the Materials andMethods section.
Enzyme activity(munits/mg of protein)
WO42- added(CM)
010
1001000
Nitratereductase
23.03.33.12.3
NADH-nitritereductase
20352525
by NADH had taken place. Fig. 4 shows nitrite re-ductase inactivation as a function of NADH con-centration: total loss of activity was observed afterpreincubation with 1OMm-NADH for 5min.OnlyNADH was effective in promoting the inacti-
vation process; NAD+ was inert. NADPH was also avery effective inactivator, but nitrite only exhibitedits protective effect against inactivation by NAD(P)Hwhen NADH was the reducing agent; preincubationwith NADPH in the presence of nitrite resulted intotal inactivation. Protection by nitrite was specific,nitrate and NH3 being ineffective in this respect(Table 6).
Induction of nitrite reductaseThe nitrite reductase activities of cell-free extracts
from A. chroococcum were greatly influenced by thenature of the inorganic nitrogen source in the culturemedium. As shown in Table 7, the cellular nitritereductase activities were very low when the sourceof nitrogen was exclusively air (i.e., 80% N2 gas) orNH3. In contrast, the nitrite reductase content washigh in cells grown aerobically with nitrate or nitriteeither with or without NH3. When growing on mediacontaining a mixture of different forms of inorganicnitrogen, the cells could utilize nitrate or nitrite evenin the presence of NH3.
Effect of tungstate on nitrate reduction
In the absence ofadded molybdenum, the additionof tungstate (0.1 mM) to the culture medium com-pletely prevented the aerobic growth of A.chroococcum when nitrate served as the sole nitrogensource, but this inhibition did not occur when inor-ganic nitrogen was supplied as nitrite or NH3, even at
tungstate concentrations that had totally suppressedgrowth with nitrate. The molybdenum requirementfor growth was particularly marked when N2 was thesole nitrogen source, and in this case the addition ofonly 1 ,uM-tungstate to the nutrient solution withoutmolybdenum was enough to prevent growth com-pletely. The activities of nitrate reductase and nitritereductase in A. chroococcum cells as a function of thetungstate concentration are shown in Table 8. Thenitrate reductase specific activity of the cell-free ex-tracts decreased markedly as the concentration oftungstate in the growth medium increased, whereasthe nitrite reductase activity remained constant.
Discussion
Nitrite reductase from A. chroococcum closelyresembles that from nitrate-assimilating fungi (Nasonet al., 1954; Nicholas et al., 1960; Rivas et al., 1973)with respect toNAD(P)H as electron donor andFADas cofactor. It thus differs essentially from the ferre-doxin-nitrite reductase of green plants, which cannotuse nicotinamide nucleotides as electron donors andseems not to be a flavoprotein (Losada & Paneque,1971; Cardenas et al., 1972a,b; Zumft, 1972).The marked inhibition by p-hydroxymercuriben-
zoate of A. chroococcum NADH-nitrite reductaseindicates the involvement of thiol groups in enzymeactivity, and the sensitivity to cyanide, a metal-binding reagent, suggests that a metal component isessential. Since the inhibition by cyanide was com-petitive with nitrite, it seems likely that this chelatingagent inhibits by reversibly binding at an essentialsite on the enzyme. On the other hand, azide at a con-centration as high as 1nM did not show any inhibitory
1973
706

NADH-NITRITE REDUCTASE FROM AZOTOBACTER CHROOCOCCUM 707
effect. Therefore with respect to these metal inhibitorsboth NAD(P)H-nitrite reductase (Nason et al.,1954; Nicholas et al., 1960; Rivas et al., 1973) andferredoxin-nitrite reductase (Losada & Paneque,1971; Cardenas et al., 1972a,b; Zumft, 1972) behavesimilarly.
It has been frequently claimed that the reduction ofnitrite to NH3 proceeds by a series of two-electrontransfers, each catalysed by a different enzyme(Nason, 1962; Takahashi et al., 1963; Hewitt &Nicholas, 1964). However, present evidence obtainedwith ferredoxin-nitrite reductase from Chlorella cellsand spinach and squash leaves has demonstrated thatnitrite is completely reduced to NH3 with no obligatefree intermediates (Beevers & Hageman, 1969;Losada & Paneque, 1971; Losada, 1972). Similarconclusions have been reached for the bacterial en-zyme (Lazzarini & Atkinson, 1961; Kemp & Atkin-son, 1966; Prakash & Sadana, 1972). The results re-ported here corroborate this view and show that inA. chroococcum nitrite is also directly and stoicheio-metrically reduced to NH3 without the formation ofhydroxylamine as a free intermediate.With regard to the existence of two either active or
inactive interconvertible forms of A. chroococcumNADH-nitrite reductase, it is important to considertogether the relevant roles of the two enzymes of thenitrate-reducing system (nitrate reductase and nitritereductase) in the overall regulation of the assimilatorypathway of nitrate reduction in bacteria, fungi, algaeand higher plants.We have recently shown that NH3, the end-product
of this pathway, promotes in vivo the reversible inacti-vation of the second moiety of the NADH-nitratereductase complex in green algae (Losada et al.,1970; Herrera et al., 1972). Apparently, NH3 actsas an uncoupler of photophosphorylation thus lead-ing to an increase in the cellular concentrations ofNAD(P)H and ADP (Losada et al., 1973). Lack of02 leads to the same effect (Losada et al., 1973). Con-version in vitro of active Chlorella nitrate reductaseinto its inactive form depends on its reduction byNADH in the presence of ADP (Maldonado et al.,1973). The transformation in vitro is also reversibleand on reoxidation the enzyme again becomes active(Jetschmann etal., 1972; Moreno etal., 1972; Maldo-nado et al., 1973). The phenomenon seems to be ageneral property of plant nitrate reductase ratherthan a peculiarity of the enzyme from photosyntheticcells, since it has been also observed with the enzymesfrom the yeasts Hansenula anomala (Pichinoty &M6t6nier, 1966) and T. nitratophila (Rivas et al.,1973).In prokaryotic organisms, it has been reported that
the nitrate-reducing system of A. vinelandii and otherbacteria requires 02 for both the reduction of nitrateto nitrite and the reduction of nitrite to NH3(Takahashi et al., 1963). We have recently shown that
A. chroococcum nitrate reductase can exist in twointerconvertible either active or inactive forms(M. G. Guerrero, J. M. Vega, E. Leadbetter & M.Losada, unpublished work). The results presented inthe present paper demonstrate that A. chroococcumNADH-nitrite reductase can be inactivated by pre-incubation with NADH in the absence of nitrite andthat the latter substrate specifically prevents and re-verses such inactivation. These reversible inactivationprocesses seem again to be of general metabolicsignificance in bacteria and are probably related toredox changes in the enzyme proteins. Kemp &Atkinson (1966) had previously observed that activityof E. coli nitrite reductase in vitro was enhanced bypreincubation with nitrite and decreased by pre-incubation with NADH.
Nitrite reductase is absent from extracts of A.chroococcum cells grown on N2 or NH3 as sole nitro-gen source, but is formed by cells growing on nitrateor nitrite even in the presence of N2 or NH3, thusconfirming the inducible nature of the assimilatorybacterial enzyme (Spencer et al., 1957; Kemp &Atkinson, 1966).The experiments with tungstate as a competitive
inhibitor of molybdate have corroborated in A.chroococcum the results obtained previously with thisbacterium (M. G. Guerrero, J. M. Vega, E. Lead-better & M. Losada, unpublished work) and withChlorella fusca (Cardenas et al., 1971; Vega et al.,1971), that the site of molybdenum action in theassimilatory metabolic pathway leading from nitrateto NH3 is exclusively in the reduction of nitrate tonitrite and that molybdenum is an essential com-ponent of nitrate reductase but does not play any rolein the reaction catalysed by nitrite reductase.
This work was carried out with the aid of a researchgrant from the Instituto de Estudios Nucleares, Spain.We thank Professor A. Paneque, Professor E. Palacianand Professor J. Cdrdenas for helpful discussion and MissM. J. Perez de Le6n, Miss I. Fernandez, Mr. J. Morenoand Mr. J. F. Alonso for skilled technical assistance.
References
Beevers, L. & Hageman, R. H. (1969) Annu. Rev. PlantPhysiol. 20, 495-522
CArdenas, J., Rivas, J., Paneque, A. & Losada, M. (1971)Arch. Mikrobiol. 79, 367-376
CArdenas, J., Barea, J. L., Rivas, J. & Moreno, C. G.(1972a) FEBS Lett. 23, 131-135
CArdenas, J., Rivas, J. & Barea, J. L. (1972b) Rev. RealAcad. Cienc. Madrid 66, 565-577
Conway, E. J. (1957) Microdiffusion Analysis and Volu-metric Error, pp. 90-132, Crosby Lockwood, London
Herrera, J., Paneque, A., Maldonado, J. M&, Barea, J. L. &Losada, M. (1972) Biochem. Biophys. Res. Commun. 48,996-1003
Hewitt, E. J. (1970) in Nitrogen Nutrition of the Plant(Kirby E. A., ed.), pp. 78-103, University ofLeeds
Vol. 133

708 J. M. VEGA, M. G. GUERRERO, E. LEADBETTER AND M. LOSADA
Hewitt, E. J. & Nicholas, D. J. D. (1964) in ModernMethods of Plant Analysis (Linskens, H. F., Sanwal,B. D. & Tracey, M. V., eds.), vol. 7, pp. 67-169,Springer Verlag, Berlin
Jetschmann, K., Solomonson, L. P. & Vennesland, B.(1972) Biochim. Biophys. Acta 275, 276-278
Kemp, J. D. & Atkinson, D. E. (1966) J. Bacteriol. 92,628-634
Kessler, E. (1971) Forst. Botanik 33, 95-103Lazzarini, R. A. & Atkinson, D. E. (1961) J. Biol. Chem.
236, 3330-3335Losada, M. (1972) La Fotosintesis del Nitrc5geno Nitrico,
Real Academia de Ciencias, pp. 1-83, MadridLosada, M. & Paneque, A. (1971) Methods Enzymol. 23,
487-491Losada, M., Paneque, A., Aparicio, P. J., Vega, J. Ma,
CArdenas, J. & Herrera, J. (1970) Biochem. Biophys.Res. Commun. 38, 1009-1015
Losada, M., Herrera, J., Maldonado, J. M' &Paneque, A.(1973) Plant Sci. Lett. 1, 31-37
Lowry, 0. H., Rosebrough, M. J., Farr, A. L. & Randall,R. J. (1951) J. Biol. Chem. 193, 265-275
Maldonado, J. Ma, Herrera, J., Paneque, A. & Losada, M.(1973) Biochem. Biophys. Res. Commun. 51, 27-33
Martin, R. G. & Ames, B. N. (1961) J. Biol. Chem. 236,1372-1379
Moreno, C. G., Aparicio, P. J., Palacian, E. & Losada, M.(1972) FEBS Lett. 26, 11-14
Nason, A. (1962) Bacteriol. Rev. 26, 16-41Nason, A., Abraham, R. G. & Averbach, B. C. (1954)
Biochim. Biophys. Acta 15, 159-161Nicholas, D. J. D., Medina, A. & Jones, 0. T. G. (1960)
Biochim. Biophys. Acta 37, 468-476Pichinoty, F. & MWtEnier, G. (1966) Ann. Inst. Pasteur 111,282-313
Prakash, OM & Sadana, J. C. (1972) Arch. Biochem.Biophys. 148, 614-632.
Ramlrez, J. M., Del Campo, F. F., Paneque, A. & Losada,M. (1966) Biochim. Biophys. Acta 188, 58-71
Relimpio, A. M., Guerrero, M. G., Paneque, A. &Losada,M. (1972) Z. Pflanzenphysiol. 66, 290-293
Rivas, J., Guerrero, M. G., Paneque, A. & Losada, M.(1973) Plant Sci. Lett. 1, 105-113,
Snell, F. D. & Snell, C. T. (1949) Colorimetric Methods ofAnalysis, p. 804, D. van Nostrand Company, New York
Spencer, D., Takahashi, H. & Nason, A. (1957) J.Bacteriol. 73, 553-562
Takahashi, H., Taniguchi, S. & Egami, F. (1963) Comp.Biochem. 5, 91-202
Vega, J. Ma, Herrera, J., Aparicio, P. J., Paneque, A. &Losada, M. (1971) Plant Physiol. 48, 294-299
Zarowny, D. P. & Sanwal, B. D. (1963) Can. J. Microbiol.9, 531-539
Zumft, W. G. (1972) Biochim. Biophys. Acta 276, 363-375
1973





























