Embed Size (px)
Citation preview

JOURNAL OF FERMENTATION AND BIOENOINEERING Vol. 84, No. 4, 324-329. 1997
Regeneration of NADPH by Cactus Chloroplasts: Coupling Reaction with P450 Monooxygenase
MASAYUKI HARA,*‘,5 HIDE0 0HKAWA,2 MIDORI NARAT0,3 MAKOTO SHIRAI,3 YASUO ASADA,’ ISA0 KARUBE,4 AND JUN MIYAKE’s5
National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology (AZST), Ministry of International Trade and Industry (MITI), 1-I Higashi, Tsukuba, Ibaraki 305,’ Department of Biological and
Environmental Science, Faculty of Agriculture, Kobe University, Rokkodai-cho-I, Nada-ku, Kobe, Hyogo 657,2 Division of Biotechnology, Ibaraki University, Ami, Inashiki, Ibaraki 300-03,3 Research Center
for Advanced Science and Technology, University of Tokyo, 4-6-l Komaba, Meguro-ku, Tokyo 153,4 and National Institute for Advanced and Interdisciplinary Research,
AIST, MITI, l-l-4 Higashi, Tsukuba, Ibaraki 305,5 Japan
Received 26 May 1997/Accepted 17 July 1997
A stable photoreaction system was developed using cactus chloroplasts coupled with the fused enzyme between rat cytochrome P4501Al (CYPlAl) and yeast NADPH-cytochrome P450 oxidoreductase expressed in yeast microsomes. The fused enzyme catalyzed o-deethylation of ‘I-ethoxycoumarin to 7-hydroxycoumarin using molecular oxygen and NADPH produced by photosystems I and II. Cactus chloroplast were found to constitute a good material for use as a NADPH-regenerating system in bioreactors. For NADPH formation, cactus chloroplasts showed twice as much activity as those of spinach when the same chlorophyll concentration was used at 30°C, as well as being more thermostable. The optimal temperature for NADPH formation with cactus chloroplasts was 42S°C, whereas for spinach it ranged from 20 to 30°C. Cactus chloroplasts can thus be applied in bioreactor NADPH-regenerating systems using oxido-reductive enzymes like P45Os.
[Key words: NADPH, chloroplast, cactus, redox enzyme, P450 monooxygenase]
Bioreactors for redox enzyme reactions have not been well studied due to the difficulty of supplying redox power. Redox power in biochemical reactions is supplied as NADPH or NADH, but these substances are expensive to use in industrial applications. Photosynthetic reac- tions can supply such redox power under illumination. In chloroplasts, photosystem II has the ability to split water molecules to form molecular oxygen, while pho- tosystem I reduces NADP+ to form NADPH.
P450 monooxygenase is an example of a redox enzyme that needs redox power. In eucaryotic eels, microsomal cytochrome P450 (P450) monooxygenase (consisting of P450 and NADPH-P450 oxidoreductase) is involved in the oxidative metabolism of xenobiotics, including drugs, food additives, and environmental chemicals. A number of P450 species with broad, overlapping substrate speci- ficities catalyze different oxidative reactions of lipophilic substrates in the presence of O2 and NADPH (1). Cyto- chrome P45Os and related enzymes have attracted increas- ing attention because of their potential pharmacological, medical, and diagnostic applications. Human cytochrome P450 species expressed in microorganisms or cultured animal cells, for example, have been used for testing the effects of xenobiotics on human metabolism.
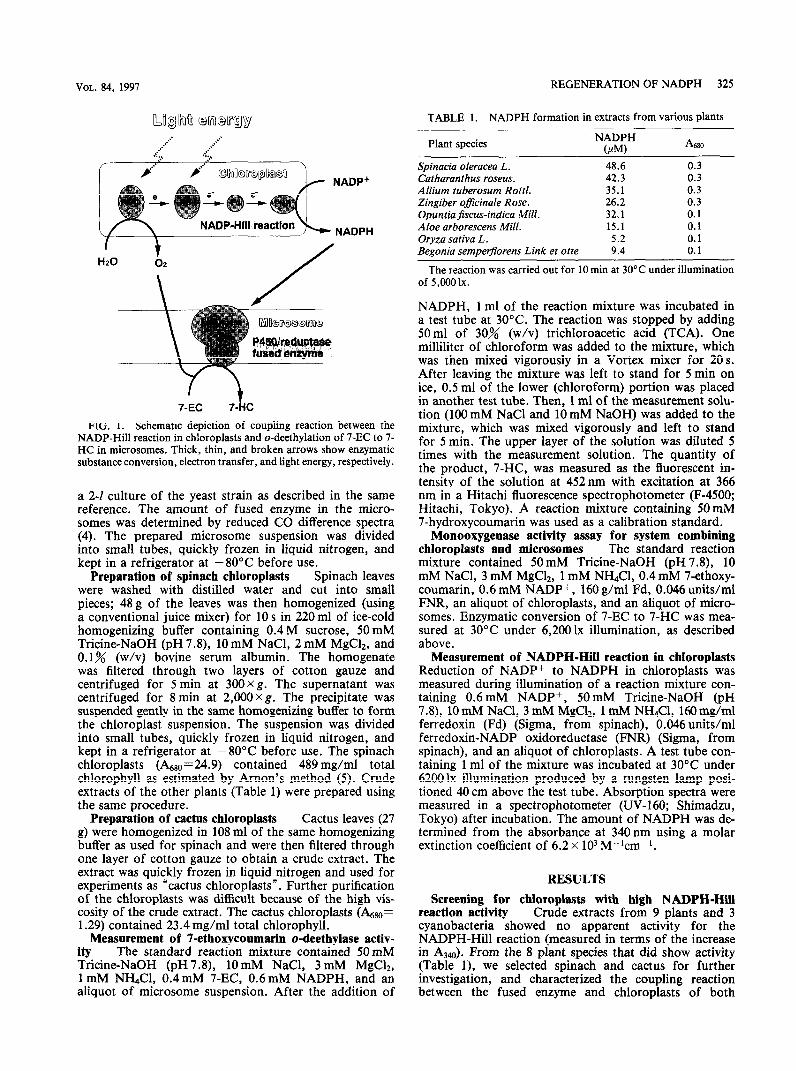
In a previous study we showed that an appropriate combination of photosystem and redox enzyme could work for the light-driven P450 monooxygenase reaction (2). This coupling reaction is depicted schematically in Fig. 1. However, because of the instability of the spinach chloroplasts used in the study, the activity and duration of the reaction were limited. As a next step, we looked for alternatives to spinach (Spinacia oleracea L.) chloroplasts and tried to measure the NADPH-Hill
* Corresponding author.
reaction using crude extracts of 17 other plants and 3 cyanobacterial species.
For further applications, the need for improved stabil- ity of the photosystem requires that photosynthetic organ- isms be screened to identify biological materials that would work better at higher temperature. In this re- search we examined 8 plant species and found that cactus (Opuntia j&us-indica Mill) chloroplasts constitute a promising biological material for constructing a photo- bioreactor with redox enzymes because these chloroplasts exhibited highly stable enzymatic activity for regenerat- ing NADPH from NADP+ under illumination. The characteristics of cactus chloroplasts and the coupling reaction with the P45O/P450 reductase fused enzyme are described in this report.
MATERIALS AND METHODS
Materials The plants (listed in Table 1) were pur- chased at a local vegetable store and a gardening shop. Ferredoxins (Fd) from spinach, ferredoxin-NADP ox- idoreductase (FNR), 7-ethoxycoumarin (7-EC), and 7- hydroxycoumarin (7-HC) were purchased from Sigma (USA). Nicotinamide adeninedinucleotide in oxidized (NADP+) and reduced (NADPH) forms was purchased from Boehringer Mannheim (Germany) and Oriental Yeast (Tokyo), respectively.
Preparation of yeast microsomes The expression plasmid pAFCR1 for the fused enzyme between rat cytochrome P450 and yeast NADPH-cytochrome P450 oxidoreductase or plasmid pAMR2 for the simultaneous expression of rat cytochrome P450 and yeast NADPH- cytochrome P450 oxidoreductase were transformed into the Saccharomyces cerevisiae strain AH22 as previously described (3). A microsome fraction was prepared from
324
VOL. 84, 1997 REGENERATION OF NADPH 325
NADP+
NADPH I
Hz0 02
I
7-EC A 7- C
FIG. I. Schematic depiction of coupling reaction between the NADP-Hill reaction in chloroplasts and o-deethylation of 7-EC to 7- HC in microsomes. Thick, thin, and broken arrows show enzymatic substance conversion, electron transfer, and light energy, respectively.
a 2-1 culture of the yeast strain as described in the same reference. The amount of fused enzyme in the micro- somes was determined by reduced CO difference spectra (4). The prepared microsome suspension was divided into small tubes, quickly frozen in liquid nitrogen, and kept in a refrigerator at -80°C before use.
Preparation of spinach chloroplasts Spinach leaves were washed with distilled water and cut into small pieces; 48 g of the leaves was then homogenized (using a conventional juice mixer) for 10 s in 220 ml of ice-cold homogenizing buffer containing 0.4 M sucrose, 50 mM Tricine-NaOH (pH 7.8), 10 mM NaCl, 2 mM MgC&, and 0.1% (w/v) bovine serum albumin, The homogenate was filtered through two layers of cotton gauze and centrifuged for 5 min at 300 X g. The supernatant was centrifuged for 8 min at 2,OOOxg. The precipitate was suspended gently in the same homogenizing buffer to form the chloroplast suspension. The suspension was divided into small tubes, quickly frozen in liquid nitrogen, and kept in a refrigerator at -80°C before use. The spinach chloroplasts (k8,, = 24.9) contained 489 mg/ml total chlorophyll as estimated by Arnon’s method (5). Crude extracts of the other plants (Table 1) were prepared using the same procedure.
Preparation of cactus chloroplasts Cactus leaves (27 g) were homogenized in 108 ml of the same homogenizing buffer as used for spinach and were then filtered through one layer of cotton gauze to obtain a crude extract. The extract was quickly frozen in liquid nitrogen and used for experiments as “cactus chloroplasts” . Further purification of the chloroplasts was difficult because of the high vis- cosity of the crude extract. The cactus chloroplasts (&*= 1.29) contained 23.4 mg/ml total chlorophyll.
Measurement of 7-ethoxycoumarin o-deethylase activ- ity The standard reaction mixture contained 50mM Tricine-NaOH (pH 7.8), 1OmM NaCl, 3 mM MgC12, 1 mM NH&l, 0.4mM 7-EC, 0.6mM NADPH, and an aliquot of microsome suspension. After the addition of
TABLE 1. NADPH formation in extracts from various plants
Plant species NADPH
WI) A680
Spinacia oleracea L. 48.6 0.3 Catharanthus roseus. 42.3 0.3 AIlium tuberosum Rottl. 35.1 0.3 Zingiber oficinale Rose. 26.2 0.3 OpuntiajTscus-indica Mill. 32.1 0.1 A toe arborescens Mill. 15.1 0.1 Oryza sativa L . 5.2 0.1 Begonia semperforens Link et otte 9.4 0.1
The reaction was carried out for 10 min at 30°C under illumination of 5,000 lx.
NADPH, 1 ml of the reaction mixture was incubated in a test tube at 30°C. The reaction was stopped by adding 50ml of 30% (w/v) trichloroacetic acid (TCA). One milliliter of chloroform was added to the mixture, which was then mixed vigorously in a Vortex mixer for 20s. After leaving the mixture was left to stand for 5 min on ice, 0.5 ml of the lower (chloroform) portion was placed in another test tube. Then, 1 ml of the measurement solu- tion (100 mM NaCl and 10 mM NaOH) was added to the mixture, which was mixed vigorously and left to stand for 5 min. The upper layer of the solution was diluted 5 times with the measurement solution. The quantity of the product, 7-HC, was measured as the fluorescent in- tensity of the solution at 452 nm with excitation at 366 nm in a Hitachi fluorescence spectrophotometer (F-4500; Hitachi, Tokyo). A reaction mixture containing 50mM 7-hydroxycoumarin was used as a calibration standard.
Monooxygenase activity assay for system combining chloroplasts and microsomes The standard reaction mixture contained 50 mM Tricine-NaOH (pH 7.8), 10 mM NaCI, 3 mM MgClz, 1 mM NH&l, 0.4mM ‘I-ethoxy- coumarin, 0.6mM NADP+, 160 g/ml Fd, 0.046 units/ml FNR, an aliquot of chloroplasts, and an aliquot of micro- somes. Enzymatic conversion of 7-EC to 7-HC was mea- sured at 30°C under 6,2001x illumination, as described above.
Measurement of NADPH-Hill reaction in chloroplasts Reduction of NADP+ to NADPH in chloroplasts was measured during illumination of a reaction mixture con- taining 0.6 mM NADP+, 50 mM Tricine-NaOH (pH 7.8), 10 mM NaCl, 3 mM MgC12, 1 mM N&Cl, 160 mg/ml ferredoxin (Fd) (Sigma, from spinach), 0.046 units/ml ferredoxin-NADP oxidoreductase (FNR) (Sigma, from spinach), and an aliquot of chloroplasts. A test tube con- taining 1 ml of the mixture was incubated at 30°C under 62001x illumination produced by a tungsten lamp posi- tioned 40 cm above the test tube. Absorption spectra were measured in a spectrophotometer (UV-160; Shimadzu, Tokyo) after incubation. The amount of NADPH was de- termined from the absorbance at 340 nm using a molar extinction coefficient of 6.2 x lo3 M-lcm-l.
RESULTS
Screening for chloroplasts with high NADPH-Hill reaction activity Crude extracts from 9 plants and 3 cyanobacteria showed no apparent activity for the NADPH-Hill reaction (measured in terms of the increase in A3&. From the 8 plant species that did show activity (Table l), we selected spinach and cactus for further investigation, and characterized the coupling reaction between the fused enzyme and chloroplasts of both
326 HARA ET AL. J. FERMENT. BIOENG.,
s 100
=t 90
B 60
40
20
-I 0 2 4 6 a IO 12
Chlorophyll (&g/ml)
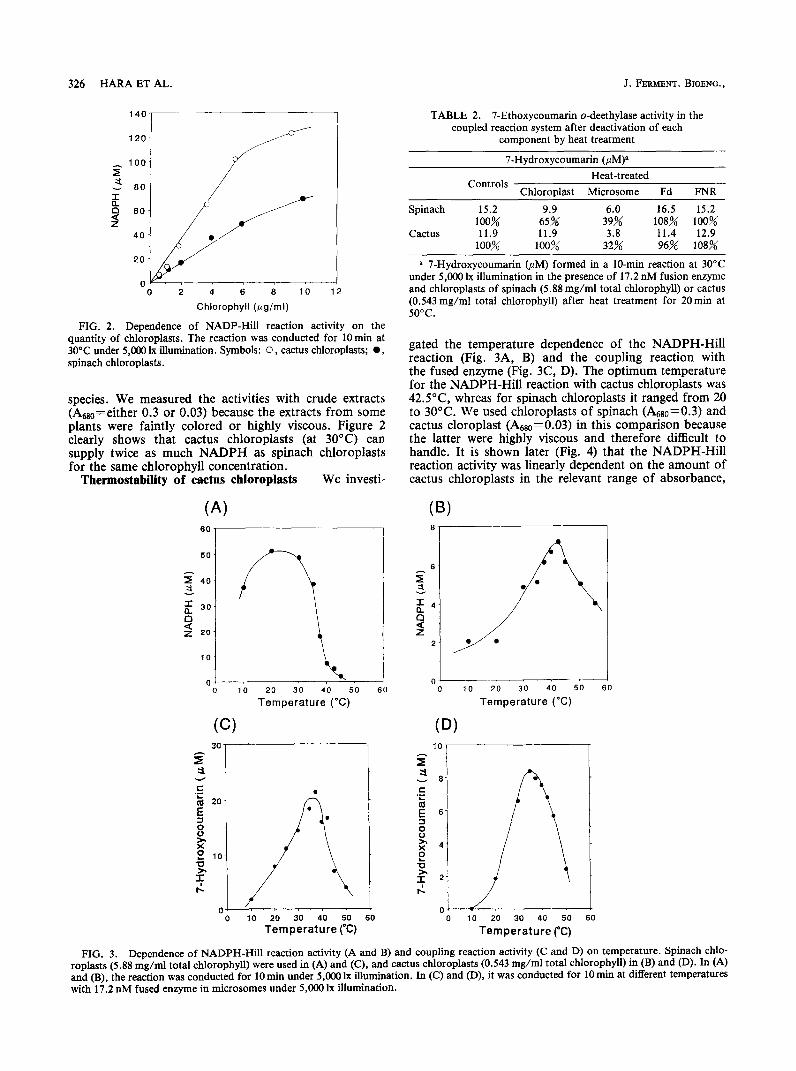
FIG. 2. Dependence of NADP-Hill reaction activity on the quantity of chloroplasts. The reaction was conducted for 10 min at 30°C under 5,000 lx illumination. Symbols: 0, cactus chloroplasts; l , spinach chloroplasts.
species. We measured the activities with crude extracts (AaO=either 0.3 or 0.03) because the extracts from some plants were faintly colored or highly viscous. Figure 2 clearly shows that cactus chloroplasts (at 30°C) can supply twice as much NADPH as spinach chloroplasts for the same chlorophyll concentration.
Thermostability of cactus chloroplasts We investi-
(A) 60
50
5 40
2 30
9 z 20
10
0 0 10 20 30 40 50
Temperature (“C)
0 10 20 30 40 50 60 0 10 20 30 40 50 60 Temperature f’C) Temperature CC)
60
(6) s
6
9 it4 2 Z
A . . 1~
2- // . .
TABLE 2. 7-Ethoxycoumarin o-deethylase activity in the coupled reaction system after deactivation of each
component by heat treatment
7-Hydroxycoumarin (,uM)B Heat-treated
Controls Chloroplast Microsome Fd FNR
Spinach 15.2 9.9 6.0 16.5 15.2 100% 65% 39% 108% 100%
Cactus 11.9 11.9 3.8 11.4 12.9 100% 100% 32% 96% 108%
a 7-Hydroxycoumarin (PM) formed in a IO-min reaction at 30°C under 5,000 lx illumination in the presence of 17.2 nM fusion enzyme and chloroplasts of spinach (5.88 mg/ml total chlorophyll) or cactus (0.543 mg/ml total chlorophyll) after heat treatment for 20 min at 50°C.
gated the temperature dependence of the NADPH-Hill reaction (Fig. 3A, B) and the coupling reaction with the fused enzyme (Fig. 3C, D). The optimum temperature for the NADPH-Hill reaction with cactus chloroplasts was 42.5”C, whreas for spinach chloroplasts it ranged from 20 to 30°C. We used chloroplasts of spinach (&80=0.3) and cactus cloroplast (Adso=0.03) in this comparison because the latter were highly viscous and therefore difficult to handle. It is shown later (Fig. 4) that the NADPH-Hill reaction activity was linearly dependent on the amount of cactus chloroplasts in the relevant range of absorbance,
“1 0 10 20 30 40 50
Temperature (“C)
PI
60
FIG. 3. Dependence of NADPH-Hill reaction activity (A and B) and coupling reaction activity (C and D) on temperature. Spinach chlo- roplasts (5.88 mg/ml total chlorophyll) were used in (A) and (C), and cactus chloroplasts (0.543 mg/ml total chlorophyll) in (B) and (D). In (A) and (B), the reaction was conducted for 10 min under 5,WO lx illumination. In (C) and (D), it was conducted for 10 min at different temperatures with 17.2 nM fused enzyme in microsomes under 5,0001x illumination.
VOL. 84, 1997 REGENERATION OF NADPH 327
I 10000 20000 30000
Light intensity (lx)
0 IO 20 30 40 50
Chlorophyll (kg/ml)
Enzyme (nM)
Light intensity (lx)
w
20.
0.0 0.1 0.2 0.3 0.4 0.5 0.6 f
Chlorophyll (kg/ml)
(F)
4
0 100 200 300
Enzyme (nM)
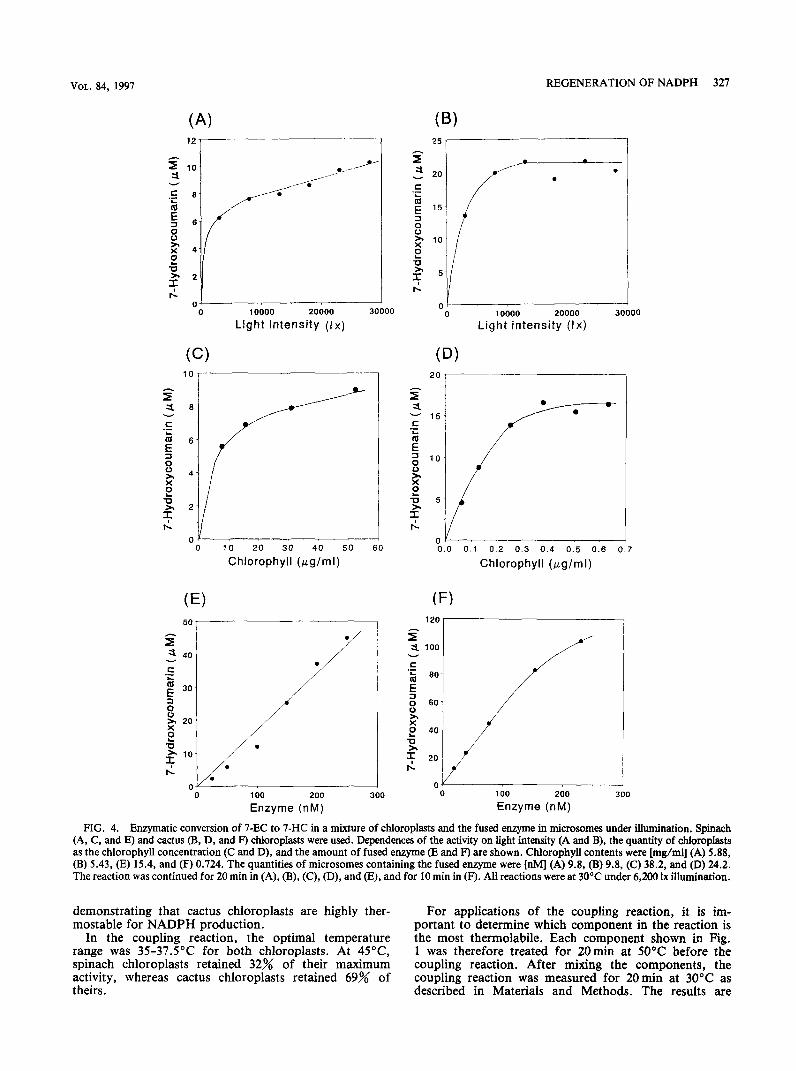
FIG. 4. Enzymatic conversion of 7-EC to 7-HC in a mixture of chloroplasts and the fused enzyme in microsomes under illnmination. Spinach (A, C, and E) and cactus (B, D, and F) chioroplasts were used. Dependences of the activity on light intensity (A and B), the quantity of chloroplasts as the chlorophyll concentration (C and D), and the amount of fused enzyme (E and F) are shown. Chlorophyll contents were [mg/ml] (A) 5.88, (B) 5.43, (E) 15.4, and (F) 0.724. The quantities of microsomes containing the fused enzyme were [nM] (A) 9.8, (B) 9.8, (C) 38.2, and (D) 24.2. The reaction was continued for 20 min in (A), (B), (C), (D), and (E), and for 10 min in (F). All reactions were at 30°C under 6,200 lx illumination.
demonstrating that cactus chloroplasts are highly ther- mostable for NADPH production.
In the coupling reaction, the optimal temperature range was 35-37.5”C for both chloroplasts. At 45°C spinach chloroplasts retained 32% of their maximum activity, whereas cactus chloroplasts retained 69% of theirs.
For applications of the coupling reaction, it is im- portant to determine which component in the reaction is the most thermolabile. Each component shown in Fig. 1 was therefore treated for 20min at 5O’C before the coupling reaction. After mixing the components, the coupling reaction was measured for 20min at 30°C as described in Materials and Methods. The results are

328 HARA ET AL. J. FERMENT. BIOENG.,
shown in Table 2. The fused enzyme in the microsomes was the most labile to heat treatment, and the spinach chloroplasts were found to be less thermostable than their cactus counterparts. Neither cactus chloroplasts, Fd nor FNR was deactivated significantly by treatment for 20min at 50°C. These results indicate that cactus chloroplasts are a promising biological material for an NADPH-regenerating system.
Coupling reaction with P450 monooxygenase We also studied the 7-ethoxycoumarin o-deethylase activity in the coupling reaction in more detail. Enzymatic con- version of 7-EC to 7-HC by the fused enzyme proceeded almost linearly for 20min in the presence of 0.6 mM NADPH (data not shown) as follows:
7-EC+NADPH+02 (fused enzyme) - 7-HCtNADP+ tCH3CHOtH20
The formation of NADPH from NADP+ in the presence of cactus or spinach chloroplasts was almost linear for 30 min under illumination (data not shown) according to the following equation:
4NADP+ + 2Hz0 + 4NADPH + O2 (chloroplasts/light)
Under illumination and in the presence of chloroplasts from cactus or spinach, the coupling reaction with the fused enzyme (Fig. 1) proceeded linearly for 30min at 30°C (data not shown).
The 7-HC formation rate in the mixed reaction system increased with light intensity up to 8,000 lx illumination, and then became saturated at 10,0001x (Fig. 4A, B). Smaller quantities of cactus chloroplasts (as compared with spinach chloroplasts) were needed for saturation in the coupling reactions shown in Figs. 4C and 4D.
The NADP-Hill reaction rate required to reduce NADP+ to NADPH varied with the chloroplast prepara- tion batch, but it was always higher than the rate of enzymatic conversion of 7-EC to 7-HC in the mixed reac- tion with the fused enzyme, regardless of the chloroplast preparation. The coupling reaction also depended on the concentration of the fused enzyme (Fig. 4E, F). The reaction was optimal at a pH around 7 (neutral) for both kinds of chloroplasts (data not shown).
The coupling reaction showed Michaelis-Menten satu- ration kinetics with substrate 7-EC. The apparent K,,, and V,, values of the mixed system for 7-EC with cac- tus chloroplasts were 120mM and 29.6mol 7-HC/mol P450/min, respectively. Km and V,, values with spinach chloroplasts were similar: 230mM and 26.4mol 7-HC/ mol P450/min, respectively. These K,,, values were higher than that for the fused enzyme in the presence of excess NADPH (Km= 19 mM).
DISCUSSION
Photosynthetic regeneration of NADPH Our in- terest has focused on the efficient regeneration of NADPH from NADP+ to drive redox enzymes. This is a key technology for the application of redox enzymes in the development of light energy-dependent bioreactors. The important question is how to supply redox power to the redox enzymes. We have been studying the applica- tion of a photosynthetic apparatus to supply the redox power for this purpose. This research has shown that cactus chloroplasts constitute a promising supply of re- dox power as NADPH, because they show higher activ- ity in the NADPH-Hill reaction at 30°C and have higher thermostability than spinach chloroplasts. The difference
in the thermostability of the NADPH-Hill reaction ob- served between spinach and cactus probably derives from the different metabolic pathways for photosynthe- sis in the two plants. Spinach is a Cs plant whereas cactus is a CAM plant. The latter is thought to be the adapted to higher temperatures that occur in deserts.
Alternative methods An alternative strategy for supplying redox power to redox enzymes is to use elec- trochemical reduction of enzymes or redox carriers by means of electrodes. Previous reports show that NADP+ is reduced to NADPH by a poly-L-lysine modified elec- trode (6), and that a 45O/P450 reductase fused enzyme can be driven by an electrochemically generated redu- ctant [cobalt(II)sepulchrate trichloride] (7). These results have interesting implications for coupling reactions with many redox enzymes, although efficient regeneration of large amounts of NADPH still seems to be difficult because of technical problems (e.g., deactivation of the enzyme by peroxide generated by electrolysis).
Chloroplasts and redox enzymes Spinach chloro- plasts have already been applied to coupling with hydrog- enase, which is another redox enzyme (8). An example of redox enzyme use that we are very much interested in is the application of P450 monooxygenases in a biore- actor that decomposed environmental chemicals. We have proposed combining cytochrome P450 monooxygenase and the photosynthetic electron transfer systems of chlo- roplasts in reference (2), and have described several features of the mixed reaction consisting of fused en- zyme activity and the NADPH-Hill reaction of isolated chloroplasts. The two reactions were successfully cou- pled in the mixture. However, before fusion enzymes can be used in industrial applications, their thermolabil- ity must be improved.
Another approach to the same problem involves the use of living cells expressing P450 enzyme. Reports show that fused enzymes can be transported to the chloro- plasts of transgenic tobacco plants and have enhanced monooxyganase activity through coupling with photosyn- thetic systems in chloroplasts (9), and that cytochrome P450 and related enzymes can be expressed in tobacco plants (10).
Future applications Applications of spinach chlo- roplasts already reported include biological devices, such as biological photocells (ll), herbicide sensors (12), and bioreactors to produce NADPH (13). In these applica- tions, chloroplasts were either an aqueous suspension in a buffer or in an immobilized state. The mechanism of photosynthetic electron transfer in spinach chloroplasts has been well-characterized biochemically, whereas the mechanism involved in cactus chloroplasts is not yet as well understood. However, we have clearly shown here that cactus chloroplasts are a promising supplier of NADPH in applications, such as these, and that they can replace spinach chloroplasts in many of them.
ACKNOWLEDGMENTS
This work was supported in part by the Protein Molecular Assembly Project, AIST, MITI. We thank Dr. Hiromasa Imaishi, Mr. Masato Miyake, and Dr. Chikashi Nakamura for their invalua- ble advice on expression of the enzyme, and Mr. Tomoaki Ishizaki, Ms. Ritsuko Ohtsuka, Ms. Kinuko Koike, Ms. Ayako Saito, MS. Miki Okamura, and Ms. Hiromi Kuita for their technical assis- tance.

VOL. 84, 1997 REGENERATION OF NADPH 329
1.
2.
3.
4.
5.
6.
7.
REFERENCES
Kamatski, T.: Metabolism of xenobiotics, p. 141-158. In Omura, T., Ishimura, Y., and Fujii-Kuriyama, Y. (ed.), Cyto- chrome P-450. Kodansha, Tokyo (1993). Kim, Y.-S., Hara, M., Ikebukuro, K., Miyake, J., Obkawa, II., and Karube, I.: Photo-induced activation of cytochrome P450/reductase fusion enzyme coupled with spinach chloro- plasts. Biotechnol. Tech., 10, 717-720 (19%). Sakaki, T., Kominami, S., Takemori, S., Obkawa, H., Akiyoshi-SbIbata, M., and Yabusaki, Y.: Kinetic studies on a genetically engineered fused enzyme between rat cytochrome P450lAl and yeast NADPH-P450 reductase. Biochemistrv. 33. 4933-4939 (1994).
II I
Omura, T. and Sate, R.: The carbon monoxide-binding pig- ment of liver microsomes. J. Biol. Chem., 239, 2379-2385 (1964). Arnon, D. I.: Copper enzymes in isolated chloroplasts. Poly- phenol oxidase in Beta vulgaris. Plant Physiol., 24, l-15 (1949). Tanigucbi, I., Iwakhi, K., Nishiyama, K., and Matsubayasbi, T.: Electrochemical behavior of the enzyme reaction of chlo- rella ferredoxin-FNR-NADP+ system. Denki Kagaku, 63, 1191- 1192 (1995). Faulkner, K. M., Shet, M. S., Fisher, C. W., and Estabrook, R. W.: Electrocatalytically driven w-hydroxylation of fatty
8.
9.
10.
11.
12.
13.
acids using cytochrome P4504Al. Proc. Natl. Acad. Sci. USA, 92,7705-7709 (1995). Horiucbi, K., Saito, IL., and Okura, I.: Photoinduced hydro- gen evolution by irradiation of visible light with the system con- taining grana and hydrogenase from Alcaligenes eutrophus H16. Nihon Kagaku Kaishi, 10, 914-916 (19%). (in Japanese) Suginra. M.. Arima. D.. Yabusaki. Y.. and Ohkawa. H.: Elec- tron interaction betvveen’microsomal cytochrome P456 monoox- ygenase and photosynthetic electron transfer system in tobacco chloroplasts. Plant Cell Physiol., 38, ~28, 3aA12 (1997). Shiota, N., Nagasawa, A., Sakaki, T., Yabusaki, Y., and Ohkawa, H.: Herbicide-resistant tobacco plants expressing the fused enzyme between rat cytochrome P450lAl (CYPlAl) and yeast NADPH-cytochrome P450 oxidoreductase. Plant Physiol., 106, 17-23 (1994). Ochiai, H., Shibata, A., Sawa, Y., and Kate, T.: Properties of the chloroplast film electrode immobilized on an SnOrcoated glass plate. Photochem. Photobiol., 35, 149-155 (1982).- Tamnra, T., Sate, A., Hara, M., Asada, Y., and Miyake, J.: Herbicide assay by use of a simple photocell. Biotechnol. Tech., 8, 615-618 (1994). Karube, I., Itsuka, T., Kayano, H., Matsunaga, T., and Suzuki, S.: Photochemical system for reeeneratine NADPH from NADP with use of immobilized chloriplasts. Biotechnol. Bioeng., 22, 2655-2665 (1980).





























