Embed Size (px)
Citation preview

Regional Accumulations of T Cells, Macrophages,and Smooth Muscle Cells in theHuman Atherosclerotic Plaque
Lena Jonasson, Jan Holm, Omar Skalli, Goran Bondjers, and Goran K. Hansson
The cellular composition of human atherosclerotic plaques was analyzed by im-munologic techniques. Plaques were removed from the internal carotid artery duringsurgery, and a panel of monoclonal antibodies was used to identify cell types. Macro-phages stained by Anti-Leu-M3 were found throughout the plaque, particularly in thelipid core region, where 60% of the cells reacted with this antibody. T cells expressingthe T3 antigen were most abundant in the fibrous cap, where they constituted 20% ofthe cell population. T cells were also isolated from the plaque and detected by arosetting test; many of these T cells were activated, as indicated by the expression ofHLA-DR. Other types of leukocytes were uncommon in the plaque. An antibody to theintermediate filament protein, desmin, was used as a marker for smooth muscle cellssince some, but not all, vascular smooth muscle cells contain this protein. The des-min-positive cells were uncommon in the nonatherosclerotic intima but were morenumerous in the plaque. In conclusion, atherosclerotic plaques are heterogeneouswith respect to cellular composition. The smooth muscle cell dominates in the fibrouscap, which also contains many T cells; the lipid core is dominated by macrophages.We suggest that interactions between smooth muscle cells and blood-borne cells areimportant in the pathogenesis of atherosclerosis.(Arteriosclerosis 6:131-138, March/April 1986)
Atherosclerosis is characterized by intimal cell prolif-eration, lipid accumulation, and connective tissue de-
position. Most of our present knowledge of the pathogene-sis of the disease come from studies on experimentalanimals. For instance, it has been found that mechanicalinjury to the intima leads to proliferation and migration ofmedial smooth muscle cells, which form an intimal thicken-ing with some of the features of the human disease.1"4 Ithas also been shown that cholesterol feeding results in
From the Arterial Biology Group, Departments of Histology andMedicine, and Departments of Clinicai Chemistry and Surgery,University of Goteborg, Goteborg, Sweden, and the Departmentof Pathology, University of Geneva, Switzerland.
This work was supported by grants from the Swedish MedicalResearch Council (projects 6816 and 4531), the Swedish NationalAssociation against Heart and Chest Disease, the University ofGoteborg, the Magnus Bergvall Foundation, and the Swedish So-ciety for Medical Sciences.
A preliminary report on this study was presented at the AnnualMeeting of the Federations of the American Societies for Experi-mental Biology in Saint Louis, Missouri, in 1984, and has beenpublished in abstract form (Fed Proc 1984;43:786).
Address for reprints: Dr. Goran K. Hansson, Department ofClinical Chemistry, University of Goteborg, Sahlgren's Hospital,S-41345, Goteborg, Sweden.
Received May 22, 1985; revision accepted August 27, 1985.
deposition of cholesterol in foam cells of intimal thicken-ings resembling those of human atherosclerosis.5"8 Theselesions differ significantly from those induced by catheterdamage. The former are composed predominantly of ma-crophage-like foam cells,910 and the latter, of typicalsmooth muscle cells together with connective tissue fi-bers.3' 4
The human fibrofatty atherosclerotic plaque bears allthese features. Morphologic studies of human plaqueshave demonstrated that they are composed of severaltypes of cells. Most of the cells contain myofibrils, suggest-ing that they are of smooth muscle origin. Other cells beara resemblance to macrophages, and are assumed to bederived from monocytes. This evaluation is, however, rath-er subjective and it has been very difficult to determine therelative contribution of the different cell types.
The development of the monoclonal antibody technol-ogy11 has provided tools for identification of cell typesusing antibodies directed against cell lineage-specific anti-gens.11"14 In this communication, we present an analysis ofthe human atherosclerotic plaque using a battery of suchantibodies together with polyclonal antibodies against in-termediate filaments. Our data show that the different re-gions of the plaque differ significantly in their cellular com-position. The selective accumulation of T cells andmacrophages in such regions may be important in thepathogenesis of the disease.
131
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from

132 ARTERIOSCLEROSIS VOL 6, No 2, MARCH/APRIL 1986
Methods
Antibodies
The characteristics of the antibodies used in the studyare shown in Table 1. The cell-type specific mouse mono-clonal antibodies, Anti-Leu-2a, Anti-Leu-3a + b, Anti-Leu-4, Anti-Leu-7, and Anti-Leu-M3, were purchased from Bee-ton Dickinson, Mountain View, California. Dako-pan-B wasobtained from Dako, Copenhagen, Denmark. Rabbit anti-vimentin was prepared by immunization of rabbits withvimentin that had been purified from bovine endothelialcells by preparative SDS electrophoresis. Rabbit antides-min, raised against chicken gizzard desmin, was used asthe affinity-purified antibodies.15'16 Biotinylated horse anti-mouse IgG and avidin-biotin-peroxidase complexes werepurchased from Vector Lab, Burlingame, California. Rho-damine-conjugated swine antirabbit-IgG was obtainedfrom Dako. Immunobeads coated with rabbit antihumanimmunoglobulins were purchased from Bio-Rad Laborato-ries, Richmond, California.
Figure 1 . Schematic representation of an atherosclerotic plaqueshowing the location of the regions where cells were counted: theintima adjacent to the plaque (Region I), the "shoulder" region (II),the fibrous cap (III), and the necrotic core region (IV).
Arterial Tissue
Atherosclerotic plaques were obtained from the internalcarotid artery of 16 patients who were between 57 and 75years old and who did not have any other systemic dis-eases. The patients were operated on because of transi-tory ischemic attacks, and plaques were obtained duringendarterectomy. Nonatherosclerotic arterial tissue wasobtained from the aortic base during coronary bypass sur-gery (five patients, 53 to 69 years old), and from the uterineartery during hysterectomy (four patients, 26 to 48 yearsold). The operative procedures were done according to thehospital routines, and informed consent was obtained fromall patients taking part in this study. The biopsies wereimmersed in ice-cold Hanks buffered salt solution (HBSS),were transported to the laboratory, and were processedwithin 20 minutes from the time of excision.
Quantltatlon of Positive Cells
The cells were counted at x 200 magnification (x 20objective, x 10 eyepieces) in the following regions of theplaque: the intima adjacent to the plaque (Region I); the"shoulder region" in the periphery of the plaque (II); thefibrous cap (III); and the necrotic core region (IV)17'18 (Fig-ure 1). Region I was identified as the visual field of thick-ened intima immediately outside the edge of the plaque.Region II was defined as the field in the plaque immediate-ly proximal to the edge. In Region III, the field at the "sum-mit" of the plaque was counted, and in Region IV, the fielddirectly below Field 3 and immediately under the borderbetween the cap and core was counted. All immunoreac-tive, nucleated cells were counted in these fields, and thetotal number of cells was determined by counting all Htx-positive nuclei.
Immunocytochemlstry
The tissue samples were snap-frozen in n-hexane thathad been chilled with liquid nitrogen. The 8 ^m cryostatsections were fixed with 95% ethanol, were rinsed in phos-phate-buffered saline (PBS, 150 mM NaCI, 20 mM phos-phate buffer, pH 7.4) and were preincubated with 2% nor-mal serum. They were then incubated with either mousemonoclonal or rabbit polyclonal antibodies, ail diluted inPBS with 4% bovine serum albumin (Sigma, St. Louis,Missouri). After repeated rinsing with PBS, the sectionsthat had been reacted with monoclonal antibodies wereincubated in a similar fashion with biotinylated second-step antibodies. This was followed by incubation with 0.3%hydrogen peroxide in methanol to eliminate endogenousperoxidase activity. The sections were rinsed repeatedlyand were incubated with the avidin-biotin-peroxidase com-plex. The peroxidase was visualized by incubation withdiaminobenzidine (1 mg/ml) and hydrogen peroxide(0.01%) in PBS. The cells were counterstained withMayer's hemalun to visualize the nuclei and were mount-ed. The sections that had reacted with rabbit antibodieswere incubated with rhodamine-conjugated second-stepantibodies and were then mounted. After examination,they were demounted, stained with Mayer's Hemalun, andfinally mounted again. All antibodies were used at optimaldilutions determined by chessboard titrations. In controlslides, the first-step antibody was omitted.
Hlstochemlstry
Fresh cryostat sections were incubated for 40 minutes ina substrate solution for detection of alpha-naphthyl acetateesterase, as described.19 Then they were counterstainedwith Mayer's hemalun. Ethanol-fixed cryostat sectionswere incubated with 1 mg/ml diaminobenzidine and 0.01%hydrogen peroxide for visualization of endogenous peroxi-dase activity.20 The positive cells were counted as de-scribed above.
Isolation of Cells
The isolation procedure was a modification of the meth-od used by Haley et al.21 Parts of the plaques were immedi-ately cut into small pieces with fine scissors, were washedtwice with HBSS by low-speed centrifugation, and weredigested for 60 minutes at 37° C in a shaking water bath, in900 IU/ml collagenase (Cl. histolyticum type I, Sigma), 125U/ml elastase (Sigma), 1.0 mg/ml soybean trypsin inhibitor(Sigma), and 5.0 mg/ml bovine serum albumin in calcium-and magnesium-free HBSS. The digested material wasfiltered through a 150 mesh nylon filter, and cells wereharvested by low-speed centrifugation. Approximately 2 x10s cells were obtained per gram of tissue. They werewashed twice with HBSS and subjected to the receptorassays described below. All glassware used for these ex-periments were siliconized.
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from

HUMAN ATHEROSCLEROTIC PLAQUE Jonasson et al. 133
Fc Receptor Assay
A washed concentrate of sheep erythrocytes was incu-bated with antisheep erythrocytes (SBL, Stockholm, Swe-den) for 60 minutes at room temperature.22 The coatederythrocytes were washed twice with PBS and suspendedto a concentration of 1%. A mixture of 500 ^l of a suspen-sion of the isolated plaque cells (1 to 2 x 10s cells/ml), 200fi\ fetal bovine serum (Flow Lab, Irvine, Scotland), and 10drops of the erythrocyte suspension was centrifuged at 40g for 5 minutes, and the loose pellet was incubated for 25minutes at 37° C. The pellet was resuspended, and dropsof cells were air-dried onto microscope slides and werefixed with ice-cold acetone. Rosetted cells and the totalnumber of cells were counted in these droplets.
Antibody-Mediated Phagocytosis
The procedure was a modification of the technique usedby Fowler et al.9 Isolated cells were incubated with coatederythrocytes as described for the Fc receptor assay. Incu-bation was for 60 minutes at 37° C, and the cells that hadingested erythrocytes were counted in the microscope.
E Receptor Assay
Tcell-specific E receptors (sheep erythrocyte receptors)were detected with AET-(32-aminoethylisothiouronium-hydrobromide)-treated sheep erythrocytes at 10% inHBSS.23 Eight drops of this suspension were incubatedwith 500 jil of isolated plaque cells (1 to 2 x 105 cells/ml)and 200 n\ fetal bovine serum, for 5 minutes at 37° C. Themixture was then centrifugated at 40 g for 5 minutes andwas incubated as a loose pellet at 4° C overnight. The cells
were resuspended to the initial volume, the nuclei werestained with Giemsa, and the total number of cells and therosetted cells were counted.
Cell Surface IgG Assay
B cells were labeled for surface immunoglobulin by theuse of Immunobeads. These are micron-sized hydrophilicpolyacrylamide beads with covalently bound purified anti-human immunoglobulin antibodies.24 Isolated plaque cellswere incubated in HBSS at 37° C for 30 minutes to removeadsorbed immunoglobulins. 100 /il of isolated cells (1 to 2x 105 cells/ml) were then mixed with 50 n\ Immunobeads.The mixture was centrifugated at 150 g for 30 minutes andincubated overnight at 4° C. The cells were resuspendedand the nuclei were stained. Rosetted cells were countedusing phase contrast microscopy.
Results
The general appearance of the plaque as it is shown inFigure 1 formed the basis for the analysis of the data. Asillustrated, the plaque was subdivided into the intima sur-rounding the plaque (I), the "shoulder region" or peripheryof the plaque, which is histologically a part of the fibrouscap (II), the fibrous cap on the central part of the lesion (III),and the lipid-rich core region (IV). This subdivision of theplaque is analogous to that used in our studies of experi-mental lesions in cholesterol-fed rabbits.17'18 It does not,however, take into account findings such as rupture orthrombosis. Therefore, no such areas were included in theanalysis.
nc *Unc v v ^ v ^ v ^ v - . i v^\
vFigure 2. Leu-M3+ cells are seen as cells with dark-stained cytoplasm in the intima adjacent to the plaque (in), in the "shoulder" (sh), inthe fibrous cap (fc) and in the necrotic core region (nc). Note the accumulation of Leu-M3+ cells around the amorphous central part of thecore region. Fragments of the media (m) are also seen in this slide (Immunoperoxidase and Htx, x 30, bar = 100
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from
134 ARTERIOSCLEROSIS VOL 6, No 2, MARCH/APRIL 1986
Table 1. Mouse Monoclonal Antibodies
Antibody
Anti-Leu-M3Anti-Leu-7
Anti-Leu-4Anti-Leu-2aAnti-Leu-3a + bDako-pan-BOKIai
Cell type
Monocytes/macrophagesLarge granular lymphocytesNK/K cellsT-cellsT cytotoxic/suppressor cellsT helper/lnducer cellsB cellsDR+ cells
Clone
MOP9HNK-1
SK7SK1SK3 + SK4To15
Antigen
NCNC
T3T8T4NCHLA-DR
Ref
1451
2725, 26, 2725, 26, 27—33
NC = not characterized.
The cell density differed markedly between regions andwas 117 ± 8 cells per mm2 (mean ± SE) in Region I, 203± 11 in Region 11,91 ± 8 in Region III, and 205 ± 18 inRegion IV.
Serial sections of plaques were incubated with a batteryof antibodies to identify specific cell types (Table 1). Themonoclonal antibodies were directed against cell type-specific cell surface antigens, and the polyclonal onesagainst intermediate filament proteins.
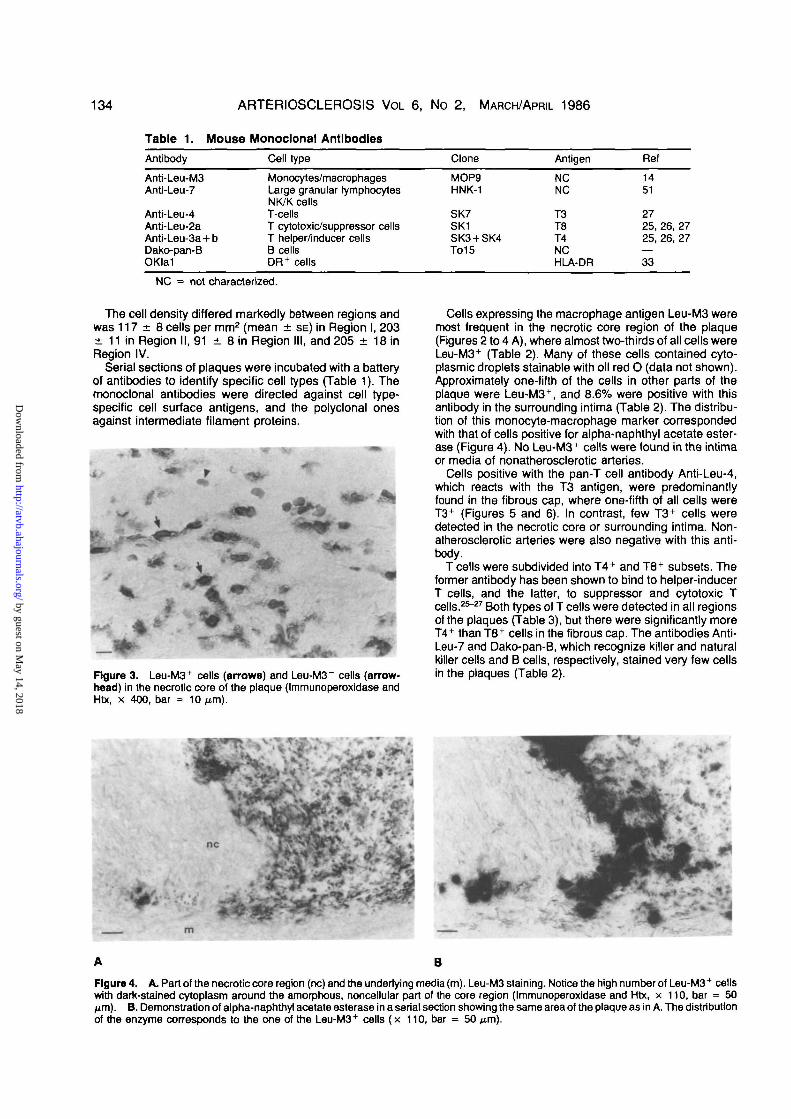
Figure 3. Leu-M3+ cells (arrows) and Leu-M3 cells (arrow-head) in the necrotic core of the plaque (Immunoperoxidase andHtx, x 400, bar = 10
Cells expressing the macrophage antigen Leu-M3 weremost frequent in the necrotic core region of the plaque(Figures 2 to 4 A), where almost two-thirds of all cells wereLeu-M3+ (Table 2). Many of these cells contained cyto-plasmic droplets stainable with oil red O (data not shown).Approximately one-fifth of the cells in other parts of theplaque were Leu-M3+, and 8.6% were positive with thisantibody in the surrounding intima (Table 2). The distribu-tion of this monocyte-macrophage marker correspondedwith that of cells positive for alpha-naphthyl acetate ester-ase (Figure 4). No Leu-M3+ cells were found in the intimaor media of nonatherosclerotic arteries.
Cells positive with the pan-T cell antibody Anti-Leu-4,which reacts with the T3 antigen, were predominantlyfound in the fibrous cap, where one-fifth of all cells wereT3+ (Figures 5 and 6). In contrast, few T3+ cells weredetected in the necrotic core or surrounding intima. Non-atherosclerotic arteries were also negative with this anti-body.
T cells were subdivided into T4+ and T8+ subsets. Theformer antibody has been shown to bind to helper-inducerT cells, and the latter, to suppressor and cytotoxic Tcells.25"27 Both types of T cells were detected in all regionsof the plaques (Table 3), but there were significantly moreT4+ than T8+ cells in the fibrous cap. The antibodies Anti-Leu-7 and Dako-pan-B, which recognize killer and naturalkiller cells and B cells, respectively, stained very few cellsin the plaques (Table 2).
A B
Figure 4. A. Part of the necrotic core region (nc) and the underiying media (m). Leu-M3 staining. Notice the high number of Leu-M3+ cellswith dark-stained cytoplasm around the amorphous, noncellular part of the core region (Immunoperoxidase and Htx, x 110, bar = 50jim). B. Demonstration of alpha-naphthyl acetate esterase in a serial section showing the same area of the plaque as in A. The distributionof the enzyme corresponds to the one of the Leu-M3+ cells (x 110, bar = 50 )
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from

HUMAN ATHEROSCLEROTIC PLAQUE Jonasson et al. 135
Table 2. Cellular Composition of the Plaque
Cell type Intima Shoulder Fibrous cap Necrotic core
MacrophagesT cellsNK/K cellsB cellsGranulocytesDesmin +
8.6±1.5a'b'c
7.7±2.5f'90.1 ±0.10.5±0.405.8±0.7iw
18.2±3.4ad
21.6±3.2('h
0.4 ±0.30.6 ±0.30.1 ±0.1
20.7±2.5l'm-n
23.9±3.7tJ'e
17.5±2.49J
0.5 ±0.30.9 ±0.40.4 ±0.2
61.2±2.1k'm'°
60.3±5.1cd-e
9.2±2.3h'!
0.1 ±0.10.1 ±0.10
28.5±2.4''n'°
Values are % of total number of cells, mean ± SE. Values having the same superscript are significantly different(p < 0.05).
fc
\ i n
Figure 5. T3+ cells are seen as cells with dark-stained cytoplasm primarily in the "shoulder" (sh) and in the fibrous cap (fc). Few positivecells are seen in the surrounding intima (in) and in the necrotic core region (nc). The larger dark spots In the core region are calciumdeposits. Part of the media (m) is seen (Immunoperoxidase and Htx, x 30, bar = 100 )
Figure 6. T3+ cells (arrows) and T3~ cells (arrowhead) in the"shoulder" region (Immunoperoxidase and Htx, x 255, bar = 20
Granulocytes were identified by histochemical stainingfor endogenous peroxidase in combination with character-istic nuclear structure. They usually appeared subendo-thelialty in the cap and shoulder regions and in low fre-quencies (Table 2).
We did not have access to any marker for the entiresmooth muscle cell population, but we used antidesmin toidentify smooth muscle cells expressing this muscle-spe-cific intermediate filament protein.282® Desmin-positivecells were rare in the surrounding intima, but increased inthe shoulder region (Table 2). Both in the fibrous cap and inthe necrotic core, all nonleukocytic cells appeared to bedesmin-positive. The frequency of desmin-positive cellswas only 7% of the nonleukocytic cells in the surroundingintima, but was much higher in the plaque. Of the nonleu-kocytic cells, 35% were desmin+ in the shoulder regionand 100% were desmin+ in the core region and in thecentral fibrous cap. Antivimentin stained 98% of the cells inthe plaque and was detected both in leukocytic cells and incells assumed to be smooth muscle cells.
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from

136 ARTERIOSCLEROSIS VOL 6, No 2, MARCH/APRIL 1986
Table 3.
T cell type
T4 +
T8 +
T4+/T8 +
T Cell Subsets
FibrousIntima Shoulder cap
2.8±1.1 9.0 + 2.2 12.1 ±2.93.6±1.0 6.6±1.7 4.7±1.2
0.7 1.4 2.6
Values are % of total number
Table 4. Characterization
Experiment Phagocytosis
12
2025
Necroticcore
5.6 ±1.83.8 ±1.0
1.5
of cells, mean ± SE.
of Isolated Cells
Fc-receptore
2638
E-receptors
55
Values are % of total number of cells.
Cells were also identified after isolation from plaques byenzyme digestion (Table 4). On the average, 32% of thesecells expressed Fc receptors, which are present on mono-cytes, macrophages, B cells, granulocytes, and certain Tand killer cells.30'31 Approximately 23% of all isolated cellsshowed a capacity for antibody-mediated phagocytosis,which is characteristic of monocytes, macrophages, andgranulocytes. No B cells were detected by immunobeadsreacting with their cell surface IgG (data not shown). Ereceptors characteristic for T cells were detected on 5% ofthe cells. Approximately 60% of E receptor-positive cellsexpressed the class II transplantation antigen HLA-DR,which was detected by a monoclonal antibody. In vitro,HLA-DR appears on T cells after activation by antigen orlectins,32-33 and our data therefore suggest that many ofthe T cells in the plaques were in an activated state.
Discussion
Many uncertainties remain about the composition of thehuman atherosclerotic plaque. One reason is that morpho-logic techniques alone cannot be used to identify the cellsin the plaque with any certainty. Even at the electron micro-scopic level, discrimination between large lymphocytes,macrophages, and even some smooth muscle cells is verydifficult. We have therefore used an alternative approachand applied the monoclonal antibody technique to identifycells in the plaques. The rationale for this approach is that itis possible to prepare monoclonal antibodies against cellsurface antigens that appear on the surface of only onespecific cell type. This has turned out to be a fruitful strate-gy in studies of cell differentiation and cellular interactionsin the immune system. 12-14'34
In the present study, we have applied well-definedmonoclonal antibodies to leukocytes for cell-type identifi-cation in sections of human atherosclerotic plaques, as-suming that these cells would maintain their cell-surfaceantigens in the tissue. Our data suggest that this is indeedthe case. For instance, the distribution of Leu-M3+ cellscorresponds to that of the histochemical macrophagemarker, alpha-naphtyl acetate esterase; also, the numbersof different subpopulations of T cells (T4+ and T8+ cells)add up to approximately the same number of cells as thatstained by the pan-T cell marker T3. There was, unfortu-nately, no antibody or other marker available to us whichwould recognize the entire smooth muscle cell population.The intermediate filament protein, desmin, appears only inmuscle cells,28'a and all plaque cells that contain desmin
can therefore be identified as smooth muscle cells. How-ever, many vascular smooth muscle cells lack desmin andhave intermediate filaments formed only by vimentin.15'M
We were unable to recognize such desmin", vimentin+
smooth muscle cells, since leukocytes also containvimentin.
The three-dimensional shape of the different types ofcells creates another problem in the interpretation of thedata. Fusiform cells will appear in several sections, andthere is a risk that the frequency of such cells is overesti-mated, and the frequency of round cells is underestimated.We have reduced this bias by counting only cells whosenuclei are seen in the section. We may, however, still haveoverestimated the frequency of smooth muscle cells, sincetheir nuclei are also more fusiform than those of leuko-cytes. One should be aware of these problems when inter-preting the present results.
Our data are compatible with the theory that the majorityof cells in the fibrous cap are derived from smooth musclecells, since they do not react with any of the markers formacrophages or leukocytes. Furthermore, a substantialproportion of these cells contained the muscle-specific in-termediate filament protein desmin, and ultrastructuralstudies in other laboratories have shown that many of thecells in these regions contain myofilaments and densebodies.35
The frequency of desmin-positive cells was only 6% inthe intima surrounding the plaque, but was 21% in theshoulder region and 58% in the fibrous cap. The data fromthe fibrous cap and core regions suggest that virtually allsmooth muscle cells in this region contain desmin. Gab-biani et al.15 have shown that the smooth muscle cells thatproliferate in the intima after a balloon injury are predomi-nantly vimentin+ , desmin", while desmin+ cells are fre-quent in a long-standing intimal lesion in which prolifera-tion is low. If this is also the case in humans, our data couldsuggest that proliferating, desmin" cells appear mainly inthe surrounding intima and shoulder region, whereas thecentral fibrous cap contains mainly stationary, desmin+
cells.The data presented in this paper indicate that a large
proportion of the cells of the carotid plaque are derivedfrom the blood. These cells have characteristic distribu-tions in the different regions of the plaque. Leu-M3+ ma-crophages constitute almost two-thirds of the cells in thecore region. A fairly large proportion (approximately 25%)of the cells in the fibrous cap and the shoulder region arealso positive for this monocyte-macrophage marker. Simi-larly, 32% of the cells isolated from the plaques had Fcreceptors, which are present on macrophages and someleukocytes,30'31 and 23% were capable of antibody-medi-
cated phagocytosis. The detection of macrophages inplaques by these techniques agrees with recent studiesusing other monoclonal antibodies.36-38
It is likely that macrophages enter the plaque as mono-cytes, and entry of monocytes into early lesions has beenobserved in several experimental models.818'39 In thecholesterol-fed rabbit, monocytes preferentially stick toareas where there is endothelial cell injury,18 and we haveshown that this is dependent on an interaction between theFc receptor of the monocyte and IgG deposited on cyto-skeletal, intermediate filaments of the injured cell.1940
Circumstantial evidence from several laboratories sug-gests that macrophages are involved in the elimination ofcholesterol from plaques,839 and we have recently ob-served that macrophage-derived foam cells of humanplaques express the apolipoprotein E gene (G Bondjers et
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from

HUMAN ATHEROSCLEROTIC PLAQUE Jonasson et al. 137
al., unpublished data). Finally, macrophages producegrowth factors for mesenchymal cells and for T cells, whichcould be important for the further progression of the le-sion."1-43
The most surprising finding in the plaque was the highfrequency of T lymphocytes, which were virtually absentfrom normal human arteries. They were particularly com-mon in the fibrous cap where one-fifth of all cells ex-pressed trie T cell-receptor-associated protein T3.
The conclusion that T cells are present in the plaque wasconfirmed by an independent test. Of the cells isolatedfrom the plaque after collagenase digestion, 5% ex-pressed the E receptor, which is a unique T cell function.23
The proportion of E cells in the isolates was lower than onemight have expected from the staining data with antibodiesto T3 on sections, but this may be due to enrichment ofother cell types during the isolation procedure, or to de-struction of some E receptors during enzymatic digestion.
The mechanisms for recruitment of T cells to plaquesare unclear. Soluble mediators produced by other celltypes could play a role; in particular, lnterieukin-1 pro-duced by macrophages has been shown to attract lympho-cytes/2 However, lnterleukin-1 is reportedly at least aseffective as a B cell chemoattractant as it is for T cells.4*The selective accumulation of T cells observed in theplaque must therefore be due either to a selective recruit-ment of T cells by another chemotactic mechanism, or to aselective trapping of T cells once they are in the plaque.The latter could be caused by tissue antigens to whichclonal T cells react. Such a stimulation of specific T cells byantigens would lead to an activation and a proliferation ofthe T cells; a substantial proportion of the T cells in theplaque were indeed activated, as reflected in their expres-sion of HLA-DR.
The products of activated lymphocytes may induce adifferentiation or phenotypic modification of many cells.The best known example of this is the ability of gammainterferon, which is a product of activated T cells, to inducethe expression of class II MHC antigens (in humans, HLA-DR, DQ, and DP) on target cells, which may then serve asantigen-presenting cells.45 We have recently reported46
that smooth muscle cells in atherosclerotic plaques (butnot in normal arteries) express HLA-DR. Taken togetherwith the present observation of a high frequency of activat-ed T cells in the plaque, this suggests that interferon re-leased from such cells can induce a phenotypic change innearby smooth muscle cells, which is characterized byexpression of HLA-DR.
Lymphocytes may also modulate local lipoprotein me-tabolism and cell proliferation. Conditioned media from ac-tivated lymphocytes increase LDL uptake by fibroblasts,47
but inhibit uptake of both native and malondialdehyde-modified LDL by macrophages.48 This makes it possiblethat lymphokines are involved in the control of cellularresponse to LDL influx in the developing plaque. Similarly,a lymphokine released by activated T cells induces prolif-eration of quiescent mesenchymal cells,49 and this couldbe another mechanism for T cell modulation of vascularcell reactions.
In summary, this study of human atheroscleroticplaques has shown that these lesions are heterogeneouswith respect to cellular composition. Macrophages, T lym-phocytes, and different types of smooth muscle cells ap-pear in different numbers in different regions of the plaque.It seems reasonable to suggest that interactions betweenthese cell types are important for the progression of thedisease.
AcknowledgmentsWe thank Tore Schersten, Kent Lundholm, and Fredrik Lund-
gren for providing surgical material and Elisabeth Lagerstedt forher skilled technical assistance.
References1. Baumgartner HR. Eine neue Methode zur Erzeugung von
Thromben durch gezielte Uberdehnung der Gefasswand. ZGes Exp Med 1963;167:227-236
2. Bjdrkerud S, Bond|ers G. Repair responses and tissue lipidafter experimental injury to the artery. Ann NY Acad Sd1976:275:180-198
3. Ross R, Glomset JA. Atherosclerosis and the arterialsmooth muscle cell. Science 1973;180:1332-1339
4. Ross R, Glomset JA. The pathogenesis of atherosclerosis.N Ertgl J Med 1976295:369-420
5. Bondjers G, Bjdrkerud S. Arterial repair and atherosclerosisafter mechanical Injury. VI. Cholesterol elimination in vitrofrom experimental atherosclerotic lesions. Exp Mol Pathol1977;26:341-347
6. Gerrtty RG, Nalto HK, Richardson M, Schwartz CJ. DietaryInduced atherogenesis in swine. Morphology of the intima inprelesion stages. Am J Pathol 1979;95:775-785
7. Poole JCF, Florey HW. Changes in the endothelium of theaorta and the behaviour of macrophages in experimental ath-eroma of rabbits. J Pathol Bacteriol 1958:75:245-252
8. Gerrlty RG. The role of the monocyte in atherogenesis. I.Transition of blood-bome monocytes into foam cells in fattylesions. Am J Pathol 1981:103:181-190
9. Fowler S, Shlo H, Haley NJ. Characterization of lipid-ladenaortic cells from cholesterol-fed rabbits. IV. Investigation ofmacrophage-like properties of aortic cell populations. LabInvest 1979;41:372-378
10. Schaffner T, Taylor K, Bartuccl EJ, et al. Arterial foam cellswith distinctive immuno-morphologic and histochemical fea-tures of macrophages. Am J Pathol 1980;100:57-78
11. Kdtiler G, Mllsteln C. Continuous cultures of fused cellssecreting antibody of predefined specificity. Nature 1975;256:495-497
12. Bluestone JA, Hodes RJ. Cell-surface molecules involvedin T cell functions. Immunology Today 1983;4:256
13. Coffman RL. Surface antigen expression and immunoglobu-lin gene rearrangement during mouse pre-B cell develop-ment. Immunological Rev 1982;69:5-12
14. Dlmltrlu-Bona A, Burmester GR, Waters SJ, WinchesterRJ. Human mononuclear phagocyte differentiation antigens.I. Patterns of antigenic expression on the surface of humanmonocytes and macrophages defined by monoclonal anti-bodies. J Immunol 1983;130:145-152
15. Gabblanl G, Rungger-Brflndle E, de Chastonay C, FrankeWW. Vimentin-containing smooth muscle cells In aortic inti-mal thickenings after endothelial injury. Lab Invest 1982;47:265-269
16. Kocher O, Skalll O, Bloom W, Gabblanl G. Cytoskeleton ofrat aortic smooth muscle cells. Normal conditions and experi-mental intimal thickening. Lab Invest 1984;50:645-651
17. Bondjers G, Brattsand R, Bylock A, Hansson GK, BJor-kemd S. Endothelial integrity and atherogenesis in rabbitswith moderate hypercholesterolemia. Artery 1977;3:395-408
18. Hansson GK, Bondjers G. Endothelial proliferation and ath-erogenesis in rabbits with moderate hypercholesterolemia.Artery 1980; 7:316-329
19. Hansson GK, Bjornheden T, Bylock A, Bondjers G. Fc-dependent binding of monocytes to areas with endothelialinjury in the rabbit aorta. Exp Mol Pathol 1981;34:264-280
20. Graham RC, Kamovsky MJ. The early stages of absorptionof injected horseradish peroxidase in the proximal tubules ofmouse kidney: Ultrastructural cytochemistry by a new tech-nique. J Histochem Cytochem 1966;14:291-302
21. Haley NJ, Shlo H, Fowler S. Characterization of lipid-ladenaortic cells from cholesterol-fed rabbits. I. Resolution of aorticcell populations by metrizamide density gradient centrifuga-tion. Lab Invest 1977;37:287-296
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from

138 ARTERIOSCLEROSIS VOL 6, No 2, MARCH/APRIL 1986
22. Bianco D, Patrick R, Nussenzweig V. A population of lym-phocytes bearing a membrane receptor for antigen-antibody-complement complexes. J Exp Med 1970; 132:702-707
23. Kaplan E, Clark C. Improved rosetting assay for detection ofhuman T-lymphocytes. J Immunol Methods 1974;5:131-137
24. Ammann AJ, Borg D, Kondo C, Wara DW. Quantitation of Bcells in peripheral blood by polyacrylamide beads coated withanti-human chain antibody. J Immunol Methods 1977;17:365-371
25. Engleman EG, Benike CJ, Glickman E, Evans RL. Anti-bodies to membrane structure that distinguish suppressor/cy-totoxic and helper T lymphocyte subpopulations block themixed leukocyte reaction in man. J Exp Med 1981 ;153:193-198
26. Kotzin BL, Benike CJ, Engleman EG. Induction of immuno-globulin-secreting cells in the allogeneic mixed leucocyte re-action: regulation by helper and suppressor lymphocyte sub-sets in man. J Immunol 1981 ;127:931-935
27. Ledbetter JA, Evans RL, Lipinski M, Cummingham-Run-dles C, Good RA, Herzenberg LA. Evolutionary conserva-tion of surface molecules that distinguish T-lymphocyte help-er/inducer and cytotoxic/suppressor subpopulations inmouse and man. J Exp Med 1981 ;153:310-323
28. Lazarides E. Intermediate filaments: A chemically hetero-genous, developmental^ regulated class of proteins. AnnRev Biochem 1982;51:219-250
29. Rungger-Brandle E, Gabbiani G. The role of cytoskeletaland cytocontractile elements in pathologic processes. Am JPathol 1983:110:359-392
30. Huber H, Polley M, Linscott WP, Fudenberg HH, Miiller-Eberhard HJ. Human monocytes: Distinct receptor sites forthe third component of complement and for immunoglobulinG. Science 1968:162:1281-1283
31. Parish CR. Separation and functional analysis of subpopula-tions of lymphocytes bearing complement and Fc-receptors.Transplant Rev 1975:25:98
32. Palacios R. Mechanism of T cell activation: role and function-al relationship of HLA-DR antigens and interleukins. ImmunolRev 1982:63:73-110
33. Reinherz EL, Kung PC, Pesando JM, Ritz J, Goldstein G,Schlossman SF. la determinants on human T-cell subsetsdefined by monoclonal antibody. Activation stimuli requiredfor expression. J Exp Med 1979:150:1472-1482
34. Kunz P, Goldstein G, Reinherz EL, Schlossman SF.Monoclonal antibodies defining distinctive human T cell sur-face antigens. Science 1979:205:347-356
35. Haust MD. Light and electron microscopy of human athero-sclerotic lesions. Adv Exp Med Biol 1978:104:33-59
36. Vedeler CA, Nyland H, Matre R. In situ characterization ofthe foam cells in early human atherosclerotic lesions. ActaPathol Microbiol Scand [C]1984;92:133-137
37. Aqel NM, Ball RY, Waldmann H, Mitchinson MJ. Monocyticorigin of foam in human atherosclerotic plaques. Atheroscle-rosis 1984:53:265-271
38. Klurfeld DM. Identification of foam cells in human atheroscle-rotic lesions as macrophages using monoclonal antibodies.Arch Pathol Lab Med 1985:109:445-449
39. Faggiotto A, Ross R. Studies of hypercholesterolemia in thenonhuman primate. II. Fatty streak conversion to fibrousplaque. Arteriosclerosis 1984:4:341-356
40. Hansson GK, Starkebaum GA, Benditt EP, Schwartz SM.Fc-mediated binding of IgG to vimentin-type intermediate fila-ments in vascular endothelial cells. Proc Natl Acad Sci USA1984; 81:3103-3107
41. Glenn KC, Ross R. Human monocyte-derived growth fac-tor(s) for mesenchymal cells: Activation of secretion by endo-toxin and concanavalin A. Cell 1981;25:603-611
42. Kampschmidt RF. The numerous postulated biologicalmanifestations of lnterleukin-1. J Leukocyte Biol 1984;36:341-355
43. Martin BM, Gimbrone MA, Unanue ER, Cotran RS. Stimu-lation of nonlymphoid mesenchymal cell proliferation by amacrophage-derived growth factor. J Immunol 1981 ;126:1510-1518
44. Miossec P, Yu C-L, Ziff M. Lymphocyte chemotactic activityof human lnterleukin-1. J Immunol 1984; 133:2007-2011
45. Pober JS, Collins T, Gimbrone MA, et al. Lymphocytesrecognize human vascular endothelial and dermal fibroblastla antigens induced by recombinant immune interferon. Na-ture 1983;305:726-729
46. Jonasson L, Holm J, Skalli O, Gabbiani G, Hansson GK.Expression of class II transplantation antigen on vascularsmooth muscle cells in human atherosclerosis. J Clin Invest1985;76:125-131
47. Hiramatsu K, Chait A, Bierman EL. The effects of human Tand B lymphocytes on low-density lipoprotein catabolism bycultured fibroblasts. Biochim Biophys Acta 1983,753:393-398
48. Fogelman AM, Seager J, Haberland ME, Hokom M, Tan-aka R, Edwards PA. Lymphocyte-conditioned medium pro-tects human monocyte-macrophages from cholesteryl esteraccumulation. Proc Natl Acad Sci, USA 1982;79:922-926
49. Wahl SM, Gately CL. Modulation of fibroblast growth by alymphokine of human T cell and continuous T cell line origin. JImmunol 1983;130:1226-1230
50. Hansson GK, Jonasson L, Holm J, Bondjers G. Cellularcomposition of the human atherosclerotic plaque [abstr]. FedProc 1984;43:786
51. Abo T, Balch CM. A differentation antigen of human NK andK cells identified by a monoclonal antibody. J Immunol1981:127:1024-1029
Index Terms:macrophages
atherosclerosis • desmin• smooth muscle cells •
immunocytochemistrylymphocytes
monoclonal antibodies
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from

L Jonasson, J Holm, O Skalli, G Bondjers and G K Hanssonatherosclerotic plaque.
Regional accumulations of T cells, macrophages, and smooth muscle cells in the human
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1986 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleArteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.6.2.1311986;6:131-138Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/6/2/131World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document.Permissions and Rights Question and AnswerFurther information about this process is available in theis being requested is located, click Request Permissions in the middle column of the Web page under Services.Clearance Center, not the Editorial Office. Once the online version of the published article for which permission
can be obtained via RightsLink, a service of the CopyrightArteriosclerosis, Thrombosis, and Vascular Biology Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on May 14, 2018
http://atvb.ahajournals.org/D
ownloaded from