Embed Size (px)
Citation preview
Regulacija ekspresije gena
kod prokariota
Bakterije
• Jednoćelijski organizmi koji nemaju jedro i druge organele.
• Geni u najvećem broju slučajeva ne poseduju introne i većina gena organizovana je u operone.
• Operon - grupa gena čiji proteinski produkti imaju srodnu funkciju ili učestvuju u istom metaboličkom procesu, pri čemu se prepisuju sa istog promotora i zajednički su regulisani.
• Policistronska iRNK - prepisana je sa operona i nosi informaciju za sintezu većeg broja proteina.
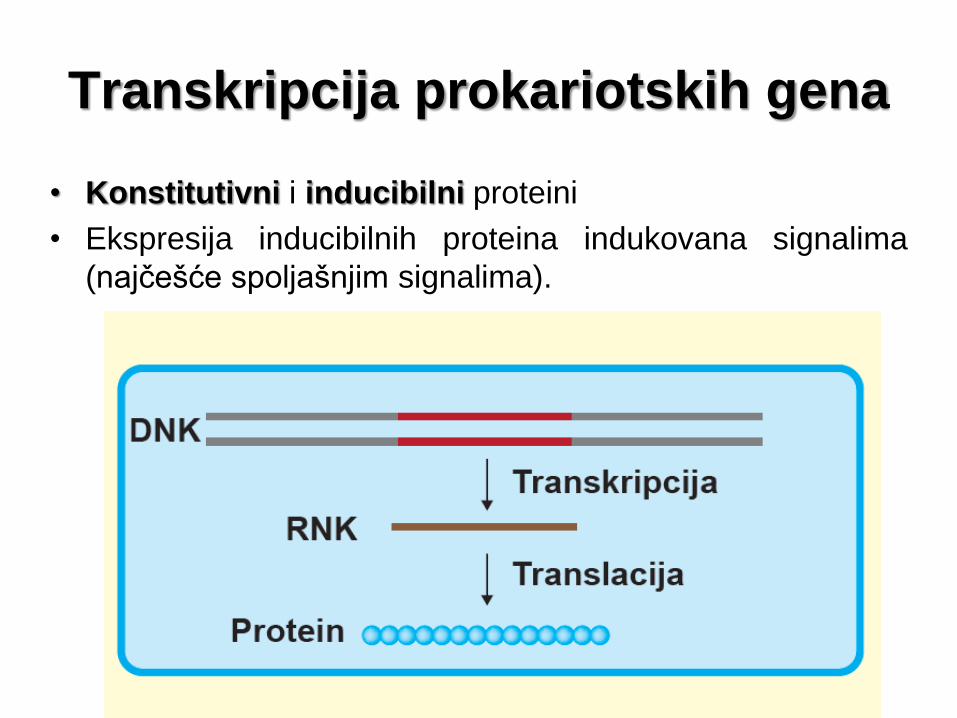
Transkripcija prokariotskih gena
• Konstitutivni i inducibilni proteini
• Ekspresija inducibilnih proteina indukovana signalima
(najčešće spoljašnjim signalima).
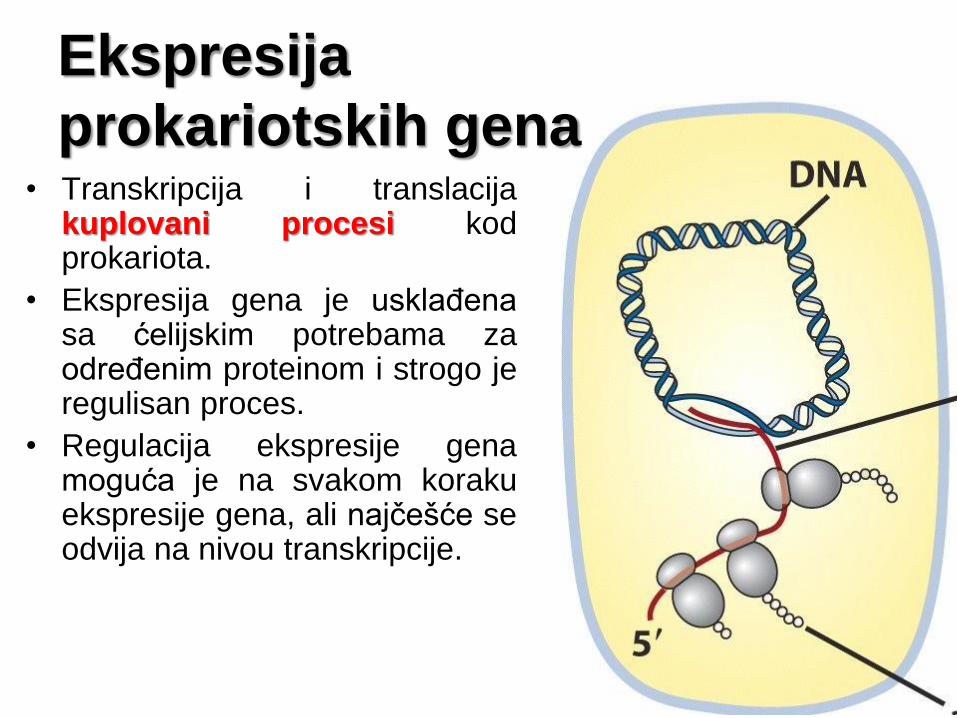
Ekspresija
prokariotskih gena • Transkripcija i translacija
kuplovani procesi kod prokariota.
• Ekspresija gena je usklađena sa ćelijskim potrebama za određenim proteinom i strogo je regulisan proces.
• Regulacija ekspresije gena moguća je na svakom koraku ekspresije gena, ali najčešće se odvija na nivou transkripcije.

Regulacija ekspresije gena
• Izbor promotora glavni mehanizam regulacije
ekspresije gena i kod prokariota i kod eukariota.
RNK polimeraza
Promotor
Sigma faktor

Regulatorni proteini
• Proces ekspresije gena kod prokariota regulisan je posebnom klasom proteina - regulatornim proteinima transkripcije.
• Oni predstavljaju „vezu” gena sa signalima koji kontrolišu proces transkripcije.
• Kada se bakterije kultivišu u in vitro uslovima spoljašnji signali su …..
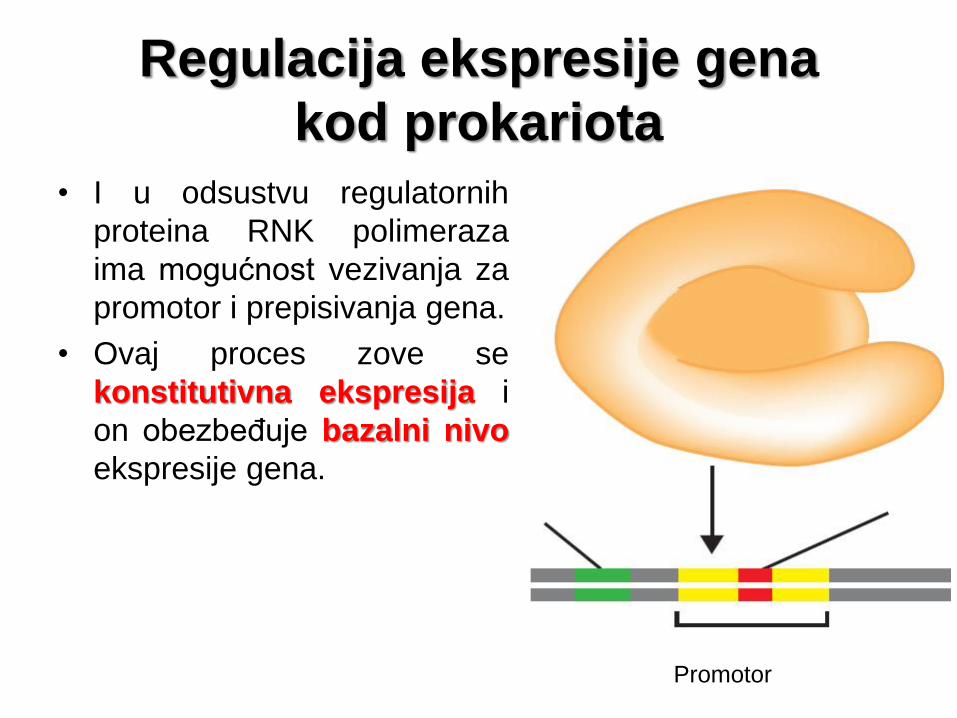
Regulacija ekspresije gena
kod prokariota
• I u odsustvu regulatornih
proteina RNK polimeraza
ima mogućnost vezivanja za
promotor i prepisivanja gena.
• Ovaj proces zove se
konstitutivna ekspresija i
on obezbeđuje bazalni nivo
ekspresije gena.
Promotor

Regulatorni proteini transkripcije
• Aktivatori transkripcije – proteini koji imaju pozitivan
efekat na ekspresiju gena.
• Represori transkripcije – proteini koji imaju negativan
efekat na ekspresiju gena, odnosno na njenu redukciju
ili potpunu blokadu.

Regulatorne sekvence
• Aktivatori i represori transkripcije vezuju se za molekul DNK na način koji je specifičan za sekvencu.
• Sekvence koje ovi proteini prepoznaju i za koje se vezuju u molekulu DNK nazivaju se regulatorne sekvence.
• Regulatorni proteini sa suprotnim efektom na transkripciju kod prokariota poseduju različite regulatorne sekvence. One se nalaze u blizini ili u samom promotoru.
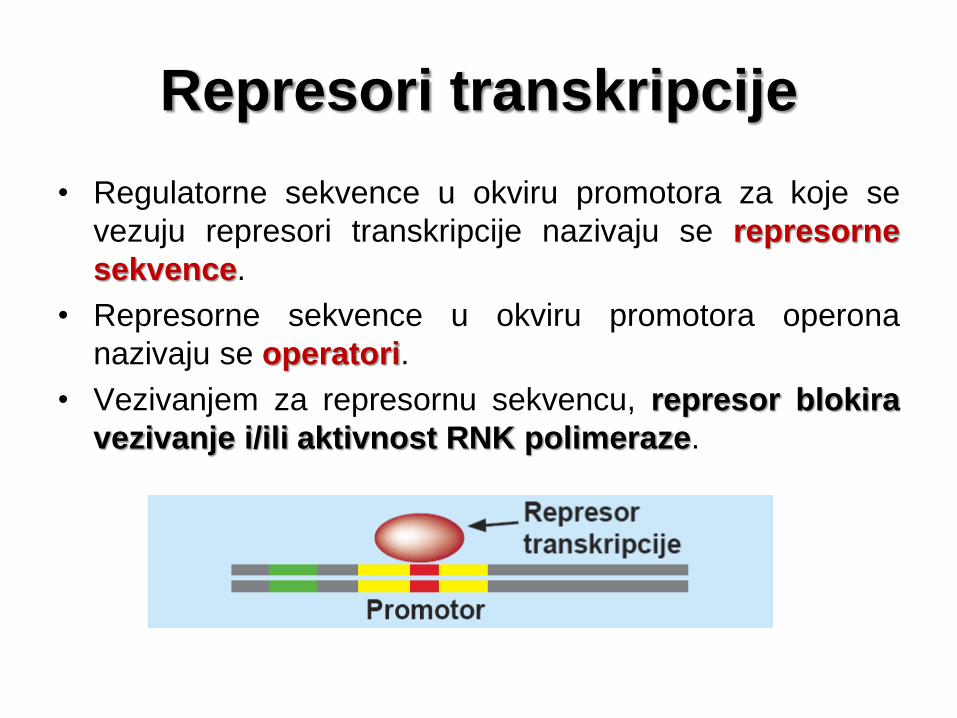
Represori transkripcije
• Regulatorne sekvence u okviru promotora za koje se
vezuju represori transkripcije nazivaju se represorne
sekvence.
• Represorne sekvence u okviru promotora operona
nazivaju se operatori.
• Vezivanjem za represornu sekvencu, represor blokira
vezivanje i/ili aktivnost RNK polimeraze.
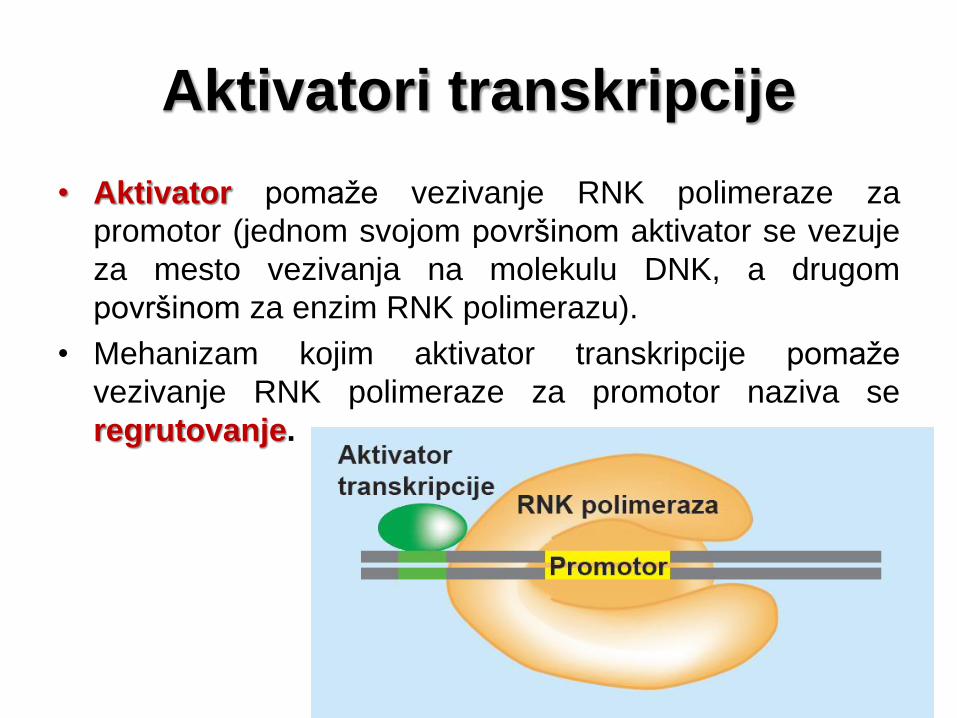
Aktivatori transkripcije
• Aktivator pomaže vezivanje RNK polimeraze za
promotor (jednom svojom površinom aktivator se vezuje
za mesto vezivanja na molekulu DNK, a drugom
površinom za enzim RNK polimerazu).
• Mehanizam kojim aktivator transkripcije pomaže
vezivanje RNK polimeraze za promotor naziva se
regrutovanje.
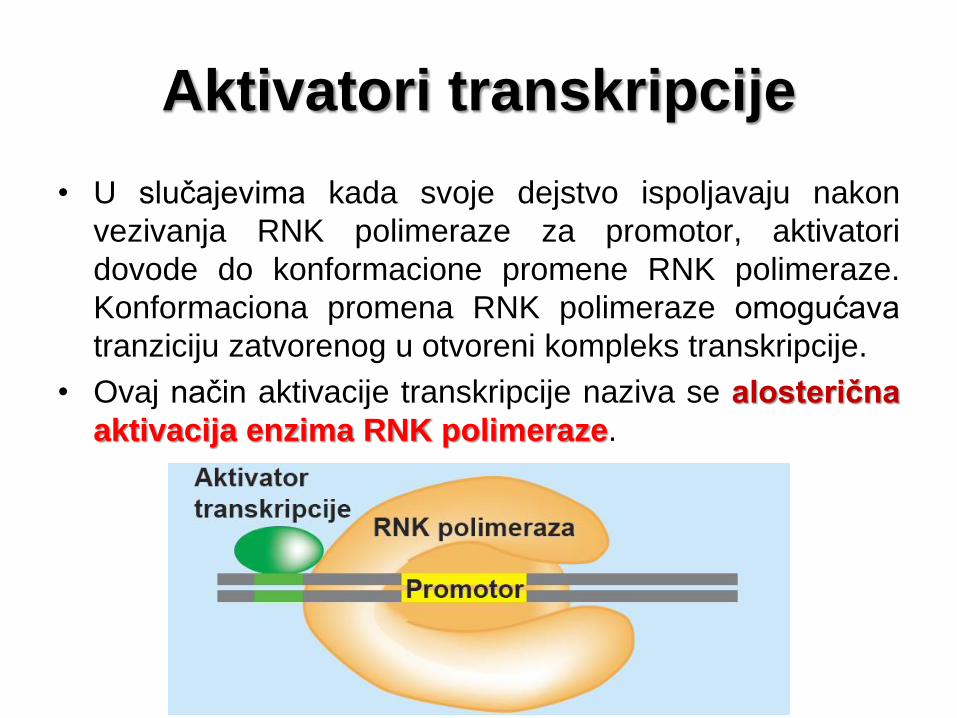
Aktivatori transkripcije
• U slučajevima kada svoje dejstvo ispoljavaju nakon
vezivanja RNK polimeraze za promotor, aktivatori
dovode do konformacione promene RNK polimeraze.
Konformaciona promena RNK polimeraze omogućava
tranziciju zatvorenog u otvoreni kompleks transkripcije.
• Ovaj način aktivacije transkripcije naziva se alosterična
aktivacija enzima RNK polimeraze.
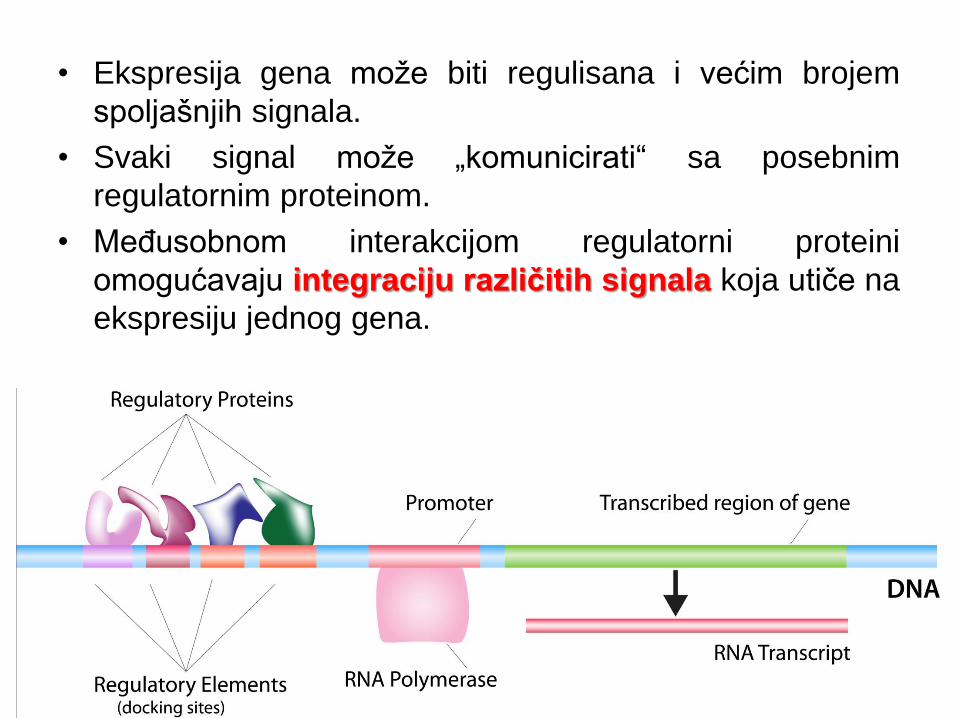
• Ekspresija gena može biti regulisana i većim brojem
spoljašnjih signala.
• Svaki signal može „komunicirati“ sa posebnim
regulatornim proteinom.
• Međusobnom interakcijom regulatorni proteini
omogućavaju integraciju različitih signala koja utiče na
ekspresiju jednog gena.
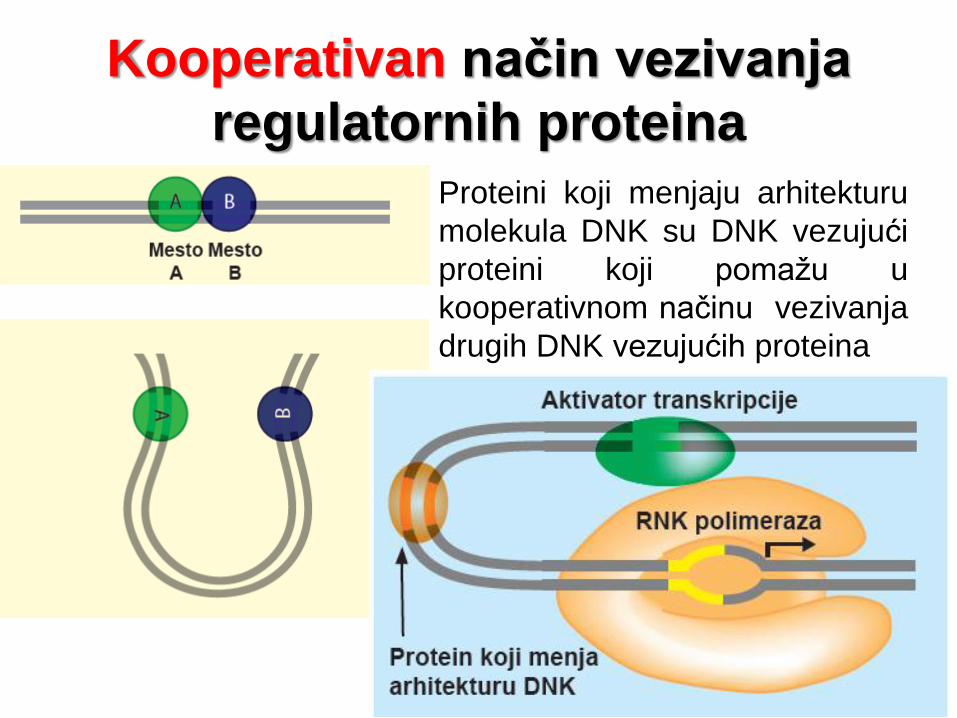
Kooperativan način vezivanja
regulatornih proteina
Proteini koji menjaju arhitekturu
molekula DNK su DNK vezujući
proteini koji pomažu u
kooperativnom načinu vezivanja
drugih DNK vezujućih proteina

Regulatorni proteini transkripcije
• Pored alosterične promene enzima RNK polimeraze i regulatorni proteini mogu menjati svoju konformaciju.
• Tipičan prokariotski regulatorni protein javlja se u dve konformacije: jedna koja omogućava i druga koja blokira njegovo vezivanje za molekul DNK.
• Konformacija regulatornih proteina zavisi od interakcije sa signalom kojim je i regulisana njegova aktivnost i ekspresija gena.

Laktozni operon
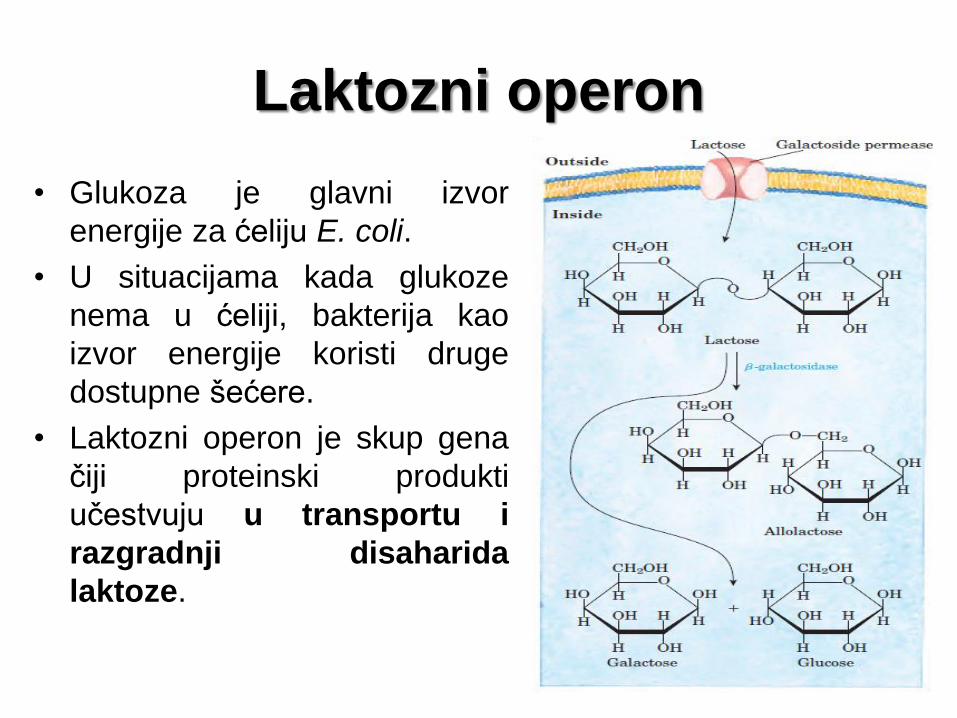
Laktozni operon
• Glukoza je glavni izvor
energije za ćeliju E. coli.
• U situacijama kada glukoze
nema u ćeliji, bakterija kao
izvor energije koristi druge
dostupne šećere.
• Laktozni operon je skup gena
čiji proteinski produkti
učestvuju u transportu i
razgradnji disaharida
laktoze.
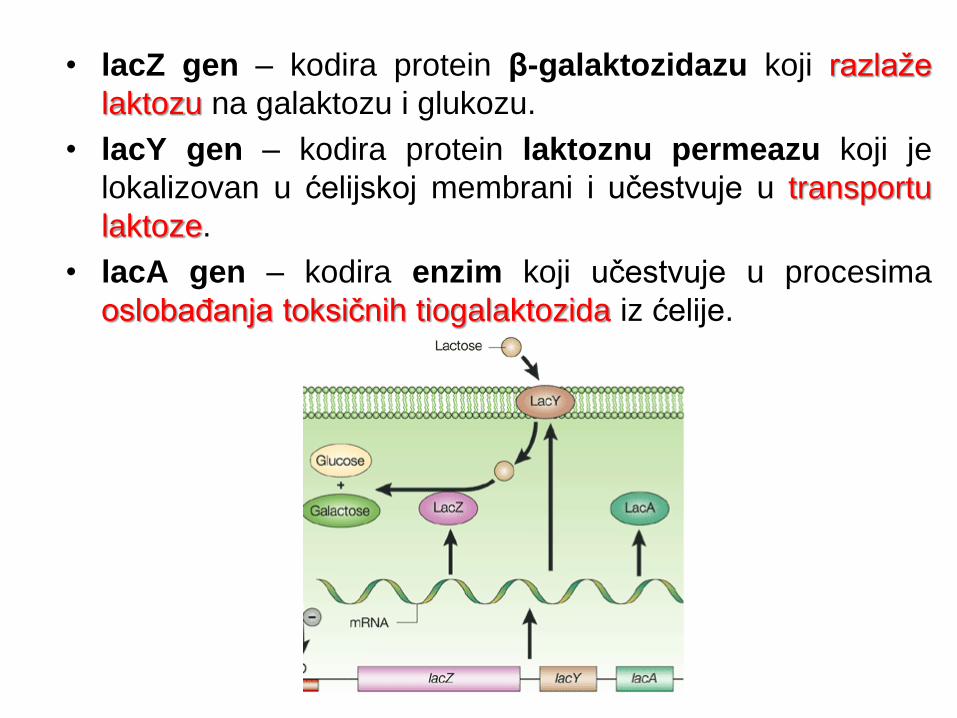
• lacZ gen – kodira protein β-galaktozidazu koji razlaže
laktozu na galaktozu i glukozu.
• lacY gen – kodira protein laktoznu permeazu koji je
lokalizovan u ćelijskoj membrani i učestvuje u transportu
laktoze.
• lacA gen – kodira enzim koji učestvuje u procesima
oslobađanja toksičnih tiogalaktozida iz ćelije.

Laktozni operon
• Bazalni nivo ekspresije operona registrovan je u
bakterijskim ćelijama u kojima je prisutna i laktoza i
glukoza.
• Lac operon eksprimira se u visokom nivou kada je u
ćeliji prisutna laktoza, a odsustvuje glukoza.

Regulatorni proteini
laktoznog operona
• Ekspresija laktoznog operona regulisana je sa dva
regulatorna proteina (aktivatorom i represorom
transkripcije).
• Oba regulatorna proteina su DNK vezujući proteini a
njihova aktivnost koordinisana je različitim signalima.
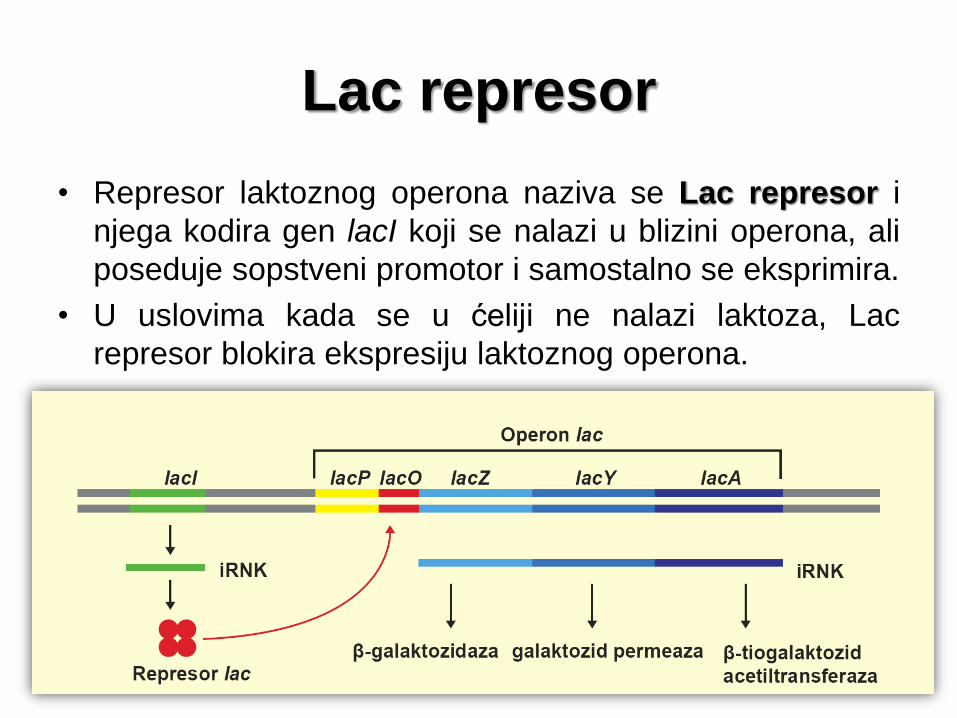
Lac represor
• Represor laktoznog operona naziva se Lac represor i
njega kodira gen lacI koji se nalazi u blizini operona, ali
poseduje sopstveni promotor i samostalno se eksprimira.
• U uslovima kada se u ćeliji ne nalazi laktoza, Lac
represor blokira ekspresiju laktoznog operona.
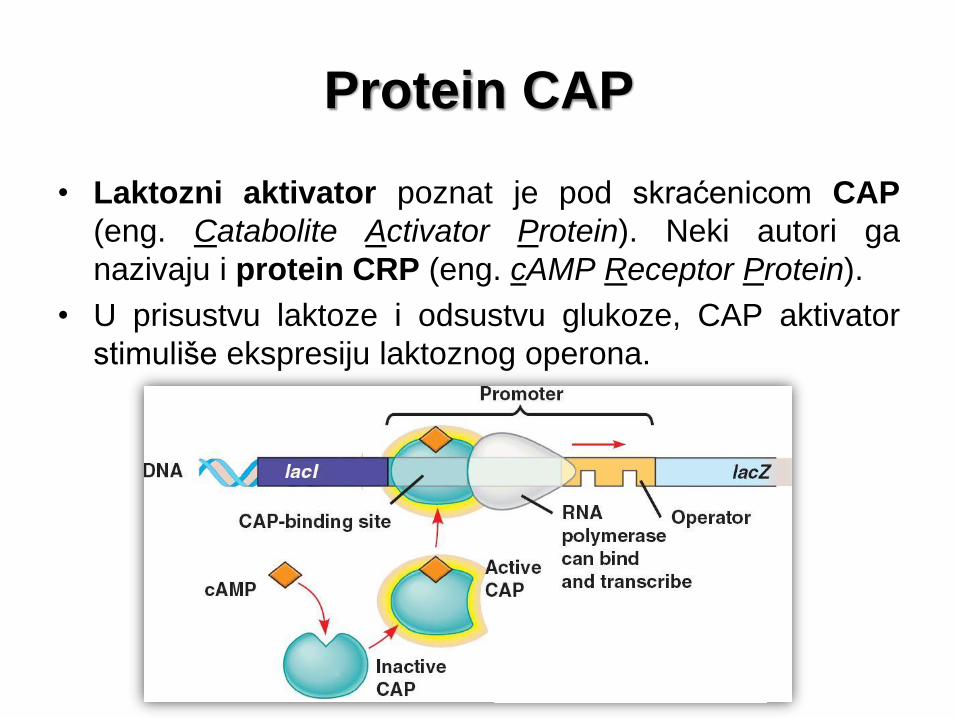
Protein CAP
• Laktozni aktivator poznat je pod skraćenicom CAP
(eng. Catabolite Activator Protein). Neki autori ga
nazivaju i protein CRP (eng. cAMP Receptor Protein).
• U prisustvu laktoze i odsustvu glukoze, CAP aktivator
stimuliše ekspresiju laktoznog operona.
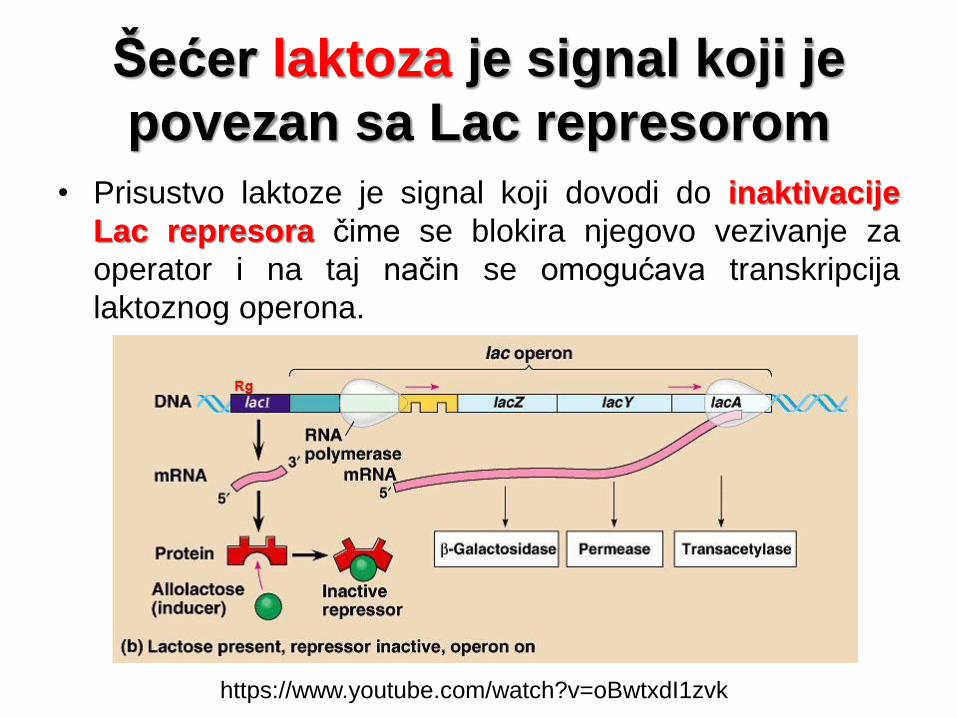
Šećer laktoza je signal koji je
povezan sa Lac represorom
• Prisustvo laktoze je signal koji dovodi do inaktivacije
Lac represora čime se blokira njegovo vezivanje za
operator i na taj način se omogućava transkripcija
laktoznog operona.
https://www.youtube.com/watch?v=oBwtxdI1zvk

Protein CAP aktivator transkripcije
• Kada u ćeliji nema glukoze dolazi do aktivacije proteina
CAP. Mesto vezivanja aktivatora CAP nalazi se 60 bp
uzvodno od mesta starta transkripcije.
• Aktivator se u formi dimera vezuje za svoje mesto
vezivanja i regrutuje enzim RNK polimerazu.

• Samo vezivanje CAP aktivatora za CAP mesto na molekulu DNK nije dovoljno za njegov pozitivan efekat na ekspresiju laktoznog operona.
• Pored domena za vezivanje za molekula DNK, aktivatori poseduju i tzv. aktivirajući domen.
• Aktivirajući domen omogućava kontakt (protein-protein interakciju) aktivatora i enzima RNK polimeraze.
• Zahvaljujući aktivirajućem domenu aktivator ima sposobnost regrutovanja RNK polimeraze.
• Ispitivanjem mutiranih RNK polimeraza utvrđeno je C-terminalni domen α subjedinice RNK polimeraze koji se naziva CTD odgovoran za kontakt sa aktivatorom.
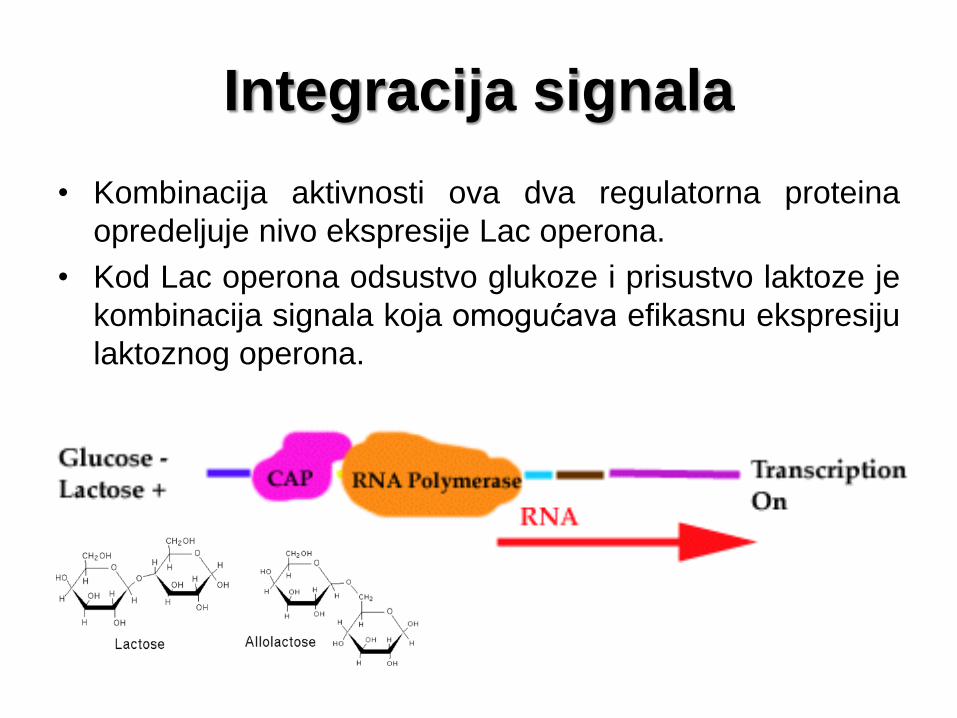
Integracija signala
• Kombinacija aktivnosti ova dva regulatorna proteina
opredeljuje nivo ekspresije Lac operona.
• Kod Lac operona odsustvo glukoze i prisustvo laktoze je
kombinacija signala koja omogućava efikasnu ekspresiju
laktoznog operona.

Laktozni operon
• Nakon ulaska u ćeliju, laktoza prelazi u formu
alolaktoze.
• Konverzija laktoze u alolaktozu katalizovana je
enzimom čiji se gen nalazi u okviru Lac operona - β-
galaktozidaza.
• Ovaj enzim je prisutna u ćeliji zahvaljujući bazičnom
nivou ekspresije Lac operona.
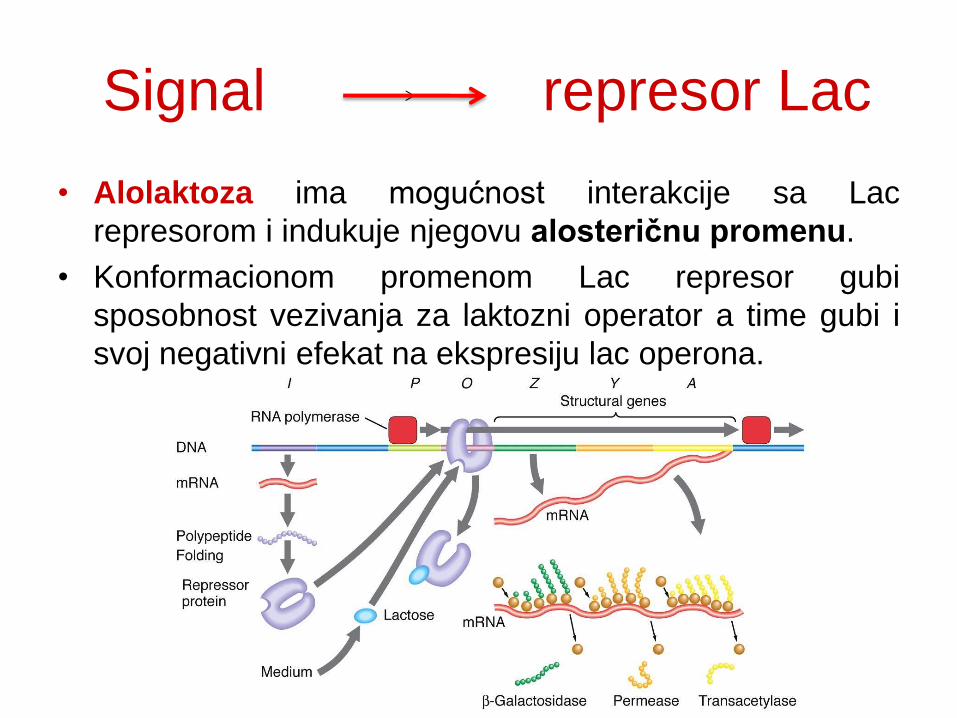
Signal represor Lac
• Alolaktoza ima mogućnost interakcije sa Lac
represorom i indukuje njegovu alosteričnu promenu.
• Konformacionom promenom Lac represor gubi
sposobnost vezivanja za laktozni operator a time gubi i
svoj negativni efekat na ekspresiju lac operona.
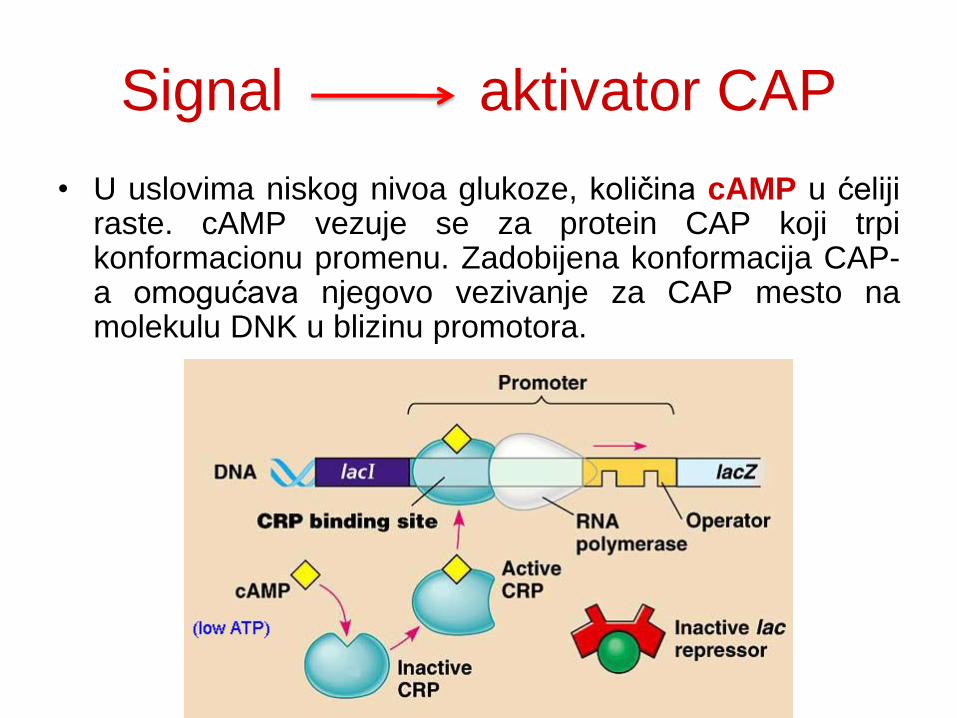
Signal aktivator CAP
• U uslovima niskog nivoa glukoze, količina cAMP u ćeliji raste. cAMP vezuje se za protein CAP koji trpi konformacionu promenu. Zadobijena konformacija CAP-a omogućava njegovo vezivanje za CAP mesto na molekulu DNK u blizinu promotora.
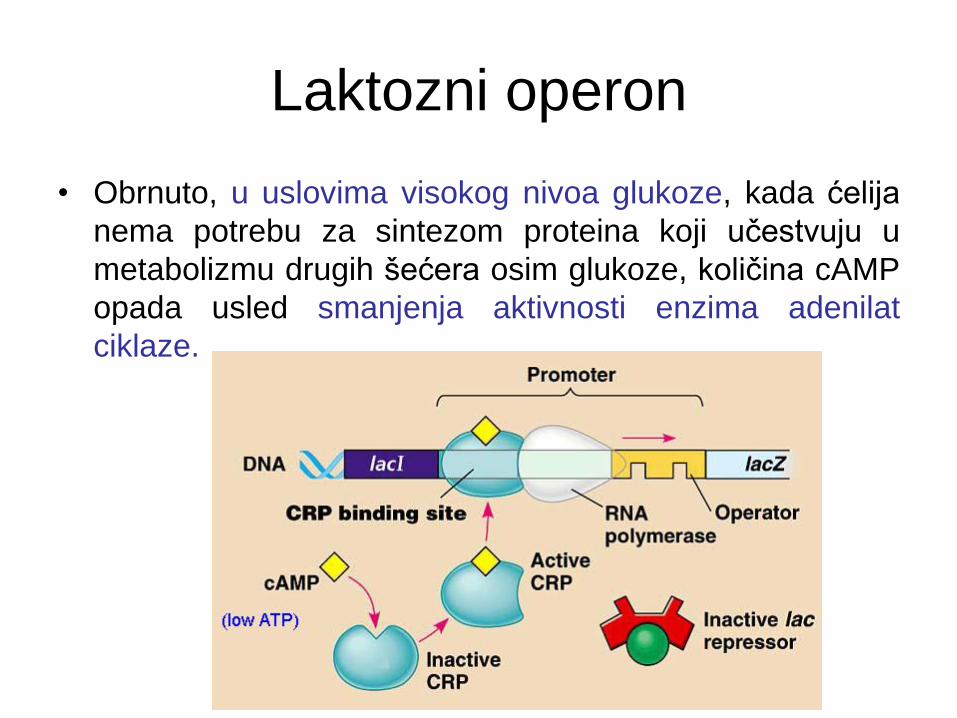
Laktozni operon
• Obrnuto, u uslovima visokog nivoa glukoze, kada ćelija
nema potrebu za sintezom proteina koji učestvuju u
metabolizmu drugih šećera osim glukoze, količina cAMP
opada usled smanjenja aktivnosti enzima adenilat
ciklaze.
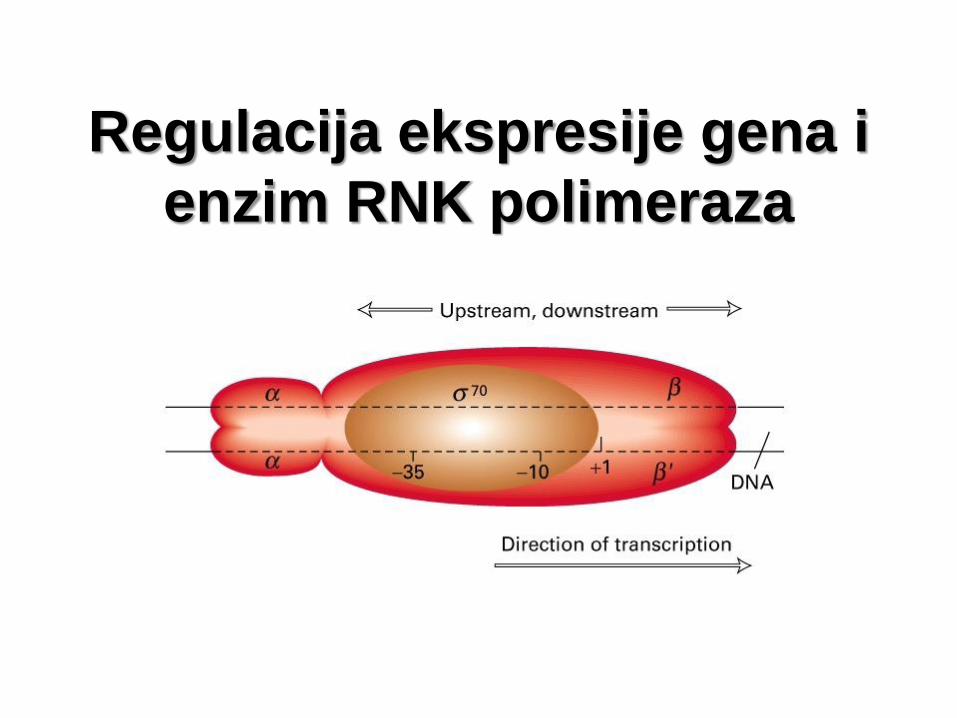
Regulacija ekspresije gena i
enzim RNK polimeraza
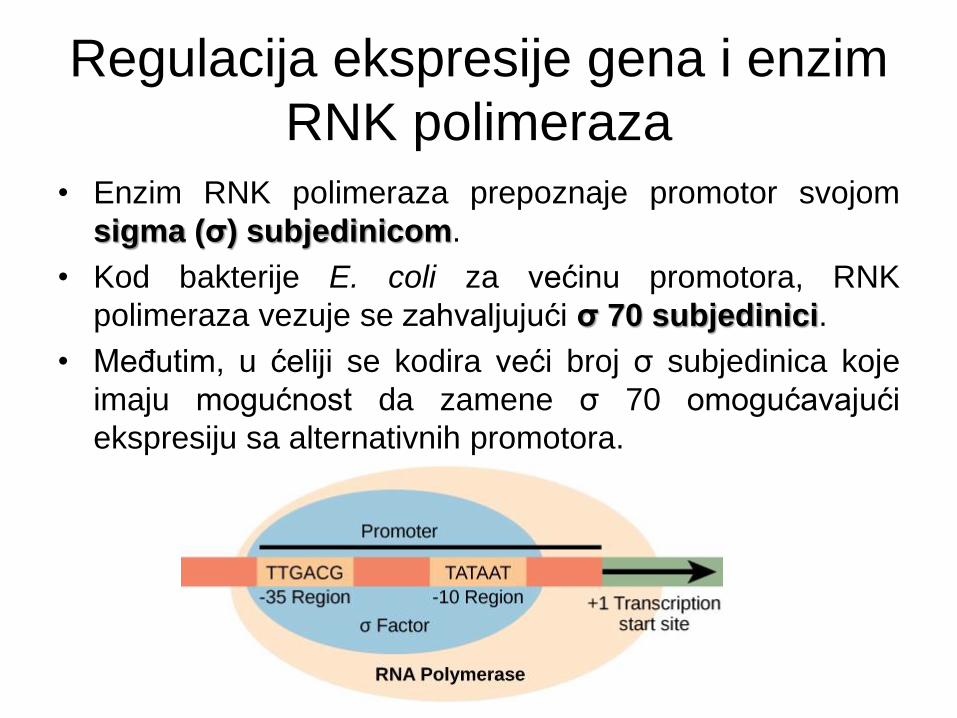
Regulacija ekspresije gena i enzim
RNK polimeraza
• Enzim RNK polimeraza prepoznaje promotor svojom
sigma (σ) subjedinicom.
• Kod bakterije E. coli za većinu promotora, RNK
polimeraza vezuje se zahvaljujući σ 70 subjedinici.
• Međutim, u ćeliji se kodira veći broj σ subjedinica koje
imaju mogućnost da zamene σ 70 omogućavajući
ekspresiju sa alternativnih promotora.
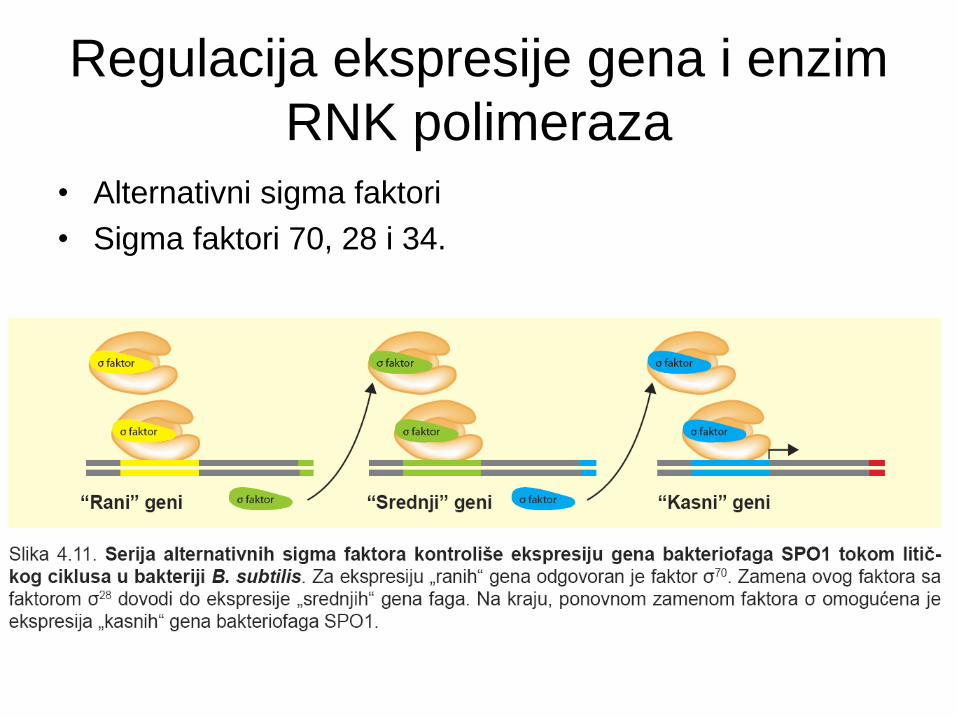
Regulacija ekspresije gena i enzim
RNK polimeraza
• Alternativni sigma faktori
• Sigma faktori 70, 28 i 34.

Alosterične promene enzima RNK
polimeraze ili molekula DNK
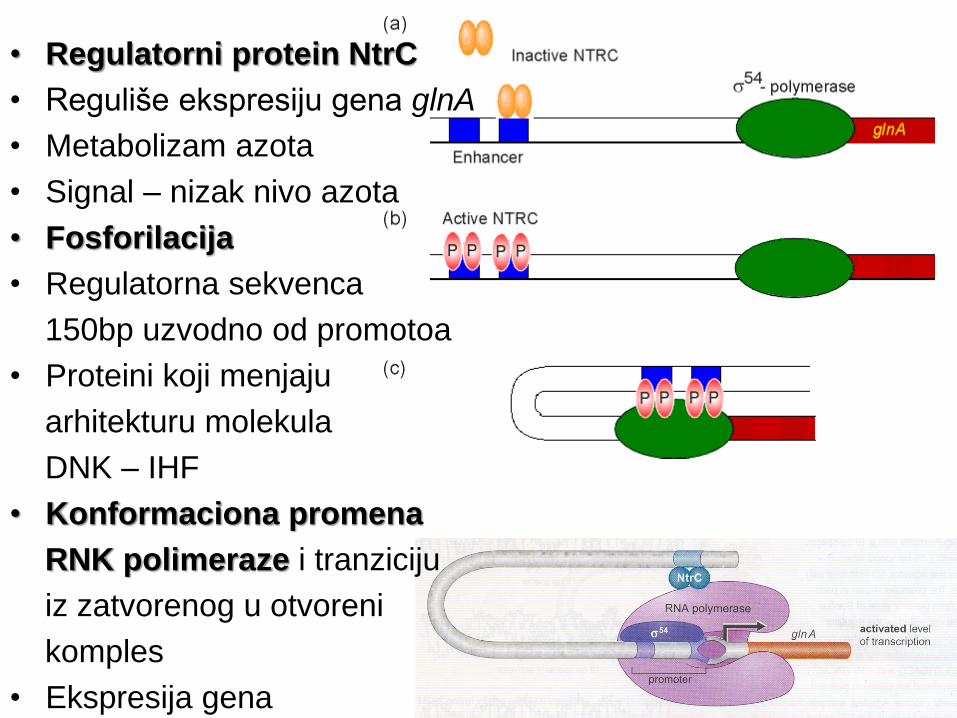
• Regulatorni protein NtrC
• Reguliše ekspresiju gena glnA
• Metabolizam azota
• Signal – nizak nivo azota
• Fosforilacija
• Regulatorna sekvenca
150bp uzvodno od promotoa
• Proteini koji menjaju
arhitekturu molekula
DNK – IHF
• Konformaciona promena
RNK polimeraze i tranziciju
iz zatvorenog u otvoreni
komples
• Ekspresija gena
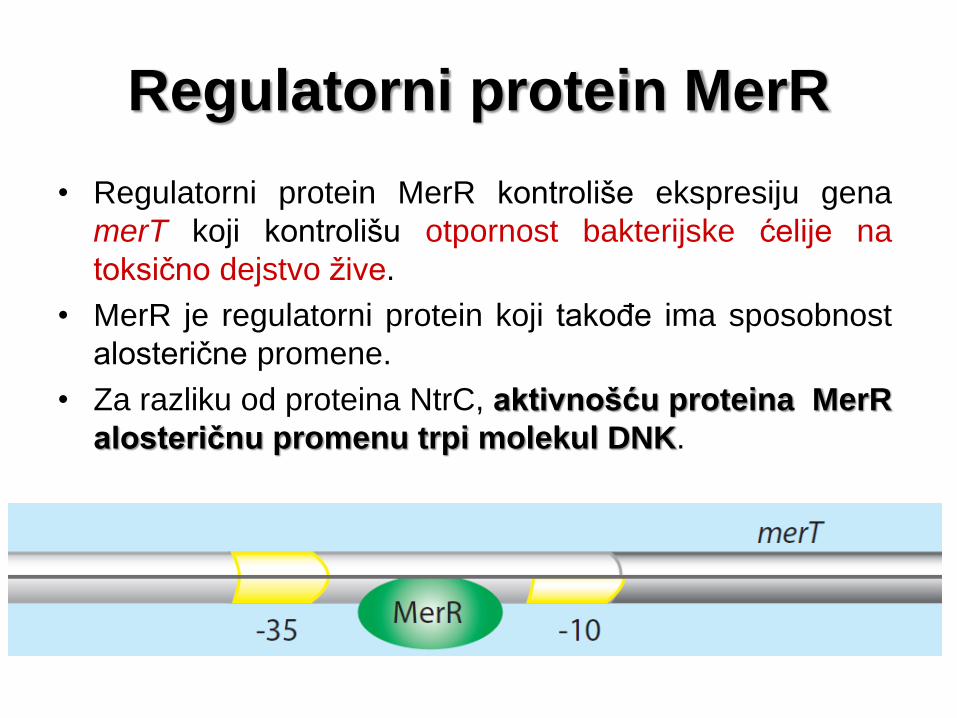
Regulatorni protein MerR
• Regulatorni protein MerR kontroliše ekspresiju gena
merT koji kontrolišu otpornost bakterijske ćelije na
toksično dejstvo žive.
• MerR je regulatorni protein koji takođe ima sposobnost
alosterične promene.
• Za razliku od proteina NtrC, aktivnošću proteina MerR
alosteričnu promenu trpi molekul DNK.

Regulatorni protein MerR
• Mesto vezivanja ovog aktivatora nalazi se unutar samog
promotora i to između konsenzusnih sekvenci -10 i -35.
• U uslovima odsustva žive, protein MerR vezan je za
svoje vezujuće mesto u okviru promotora gena merT.
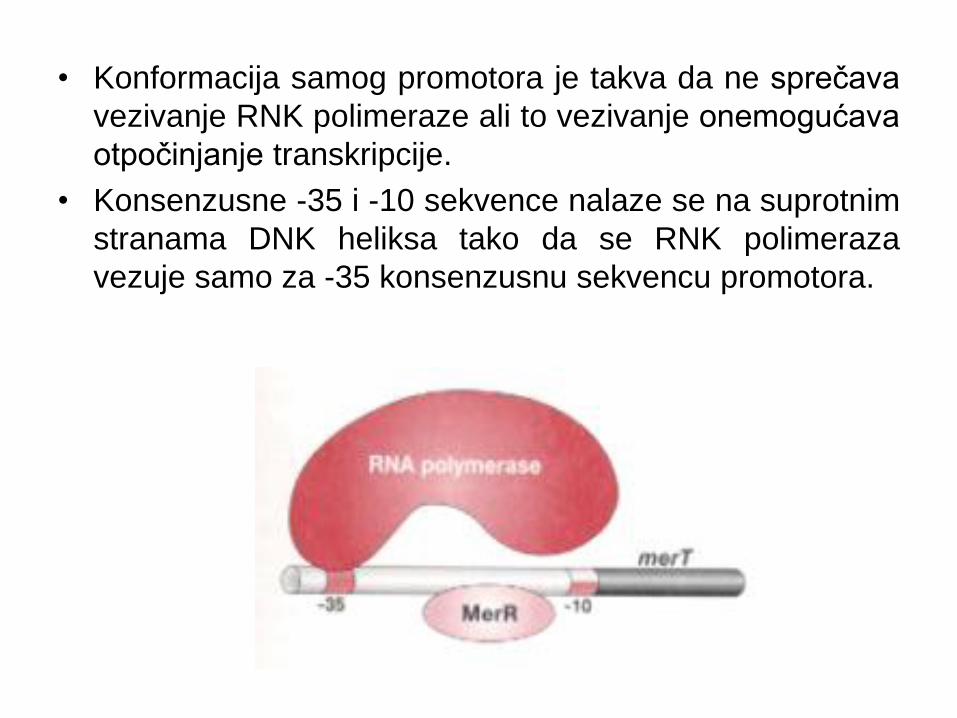
• Konformacija samog promotora je takva da ne sprečava
vezivanje RNK polimeraze ali to vezivanje onemogućava
otpočinjanje transkripcije.
• Konsenzusne -35 i -10 sekvence nalaze se na suprotnim
stranama DNK heliksa tako da se RNK polimeraza
vezuje samo za -35 konsenzusnu sekvencu promotora.
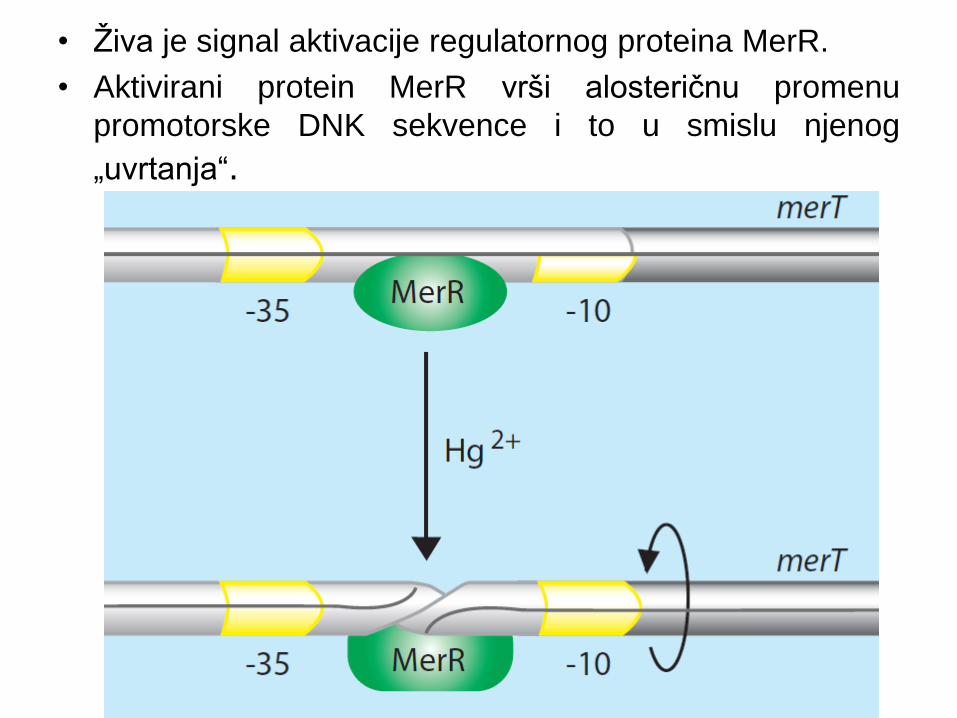
• Živa je signal aktivacije regulatornog proteina MerR.
• Aktivirani protein MerR vrši alosteričnu promenu
promotorske DNK sekvence i to u smislu njenog
„uvrtanja“.
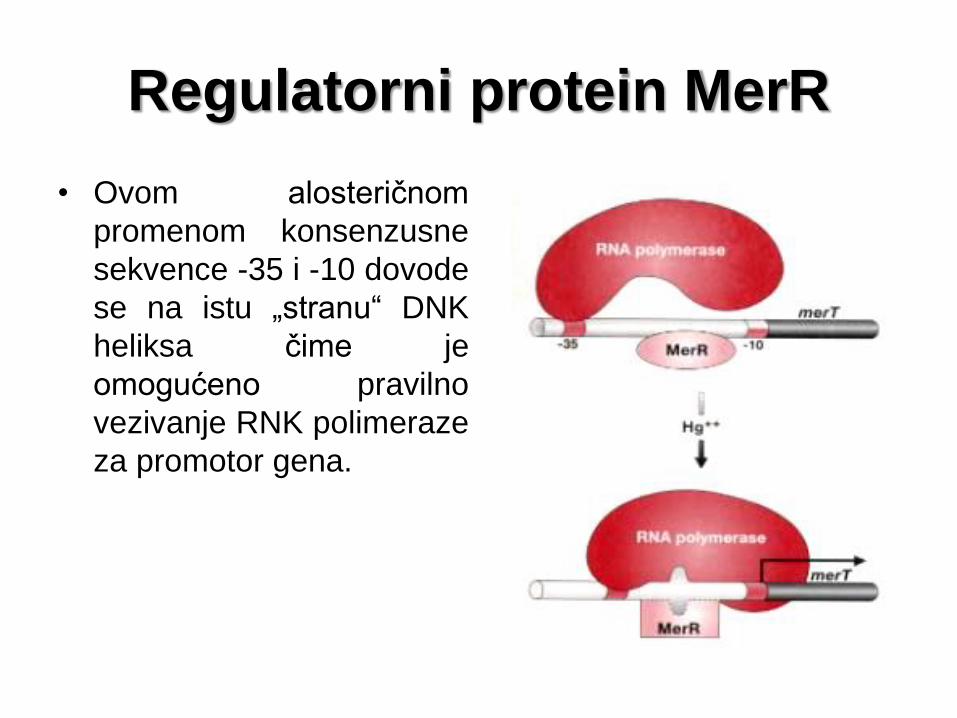
Regulatorni protein MerR
• Ovom alosteričnom
promenom konsenzusne
sekvence -35 i -10 dovode
se na istu „stranu“ DNK
heliksa čime je
omogućeno pravilno
vezivanje RNK polimeraze
za promotor gena.
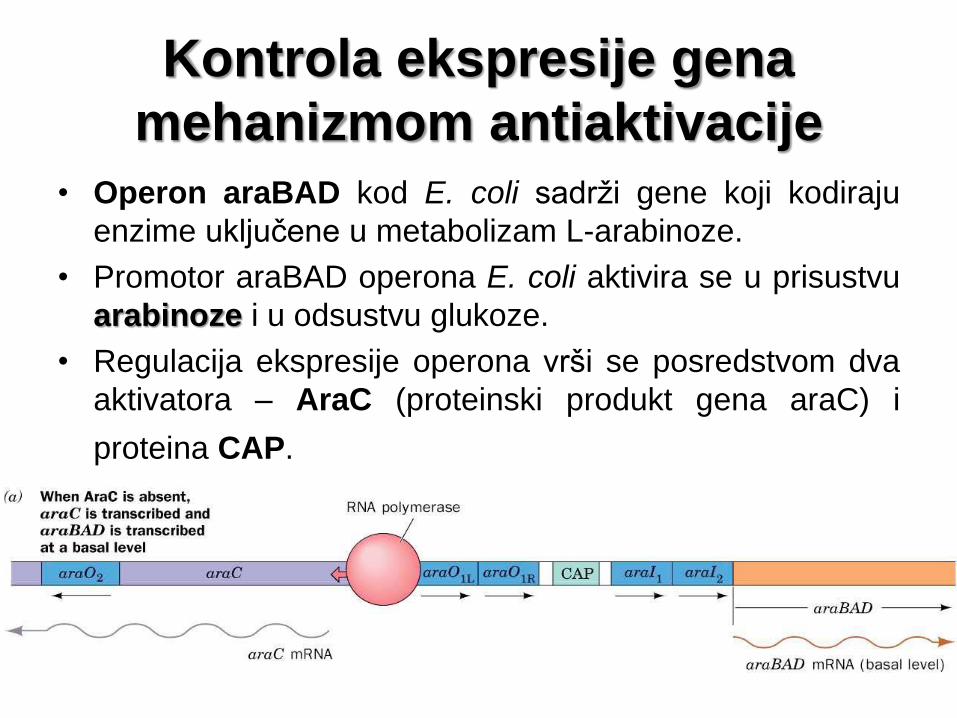
Kontrola ekspresije gena
mehanizmom antiaktivacije
• Operon araBAD kod E. coli sadrži gene koji kodiraju
enzime uključene u metabolizam L-arabinoze.
• Promotor araBAD operona E. coli aktivira se u prisustvu
arabinoze i u odsustvu glukoze.
• Regulacija ekspresije operona vrši se posredstvom dva
aktivatora – AraC (proteinski produkt gena araC) i
proteina CAP.
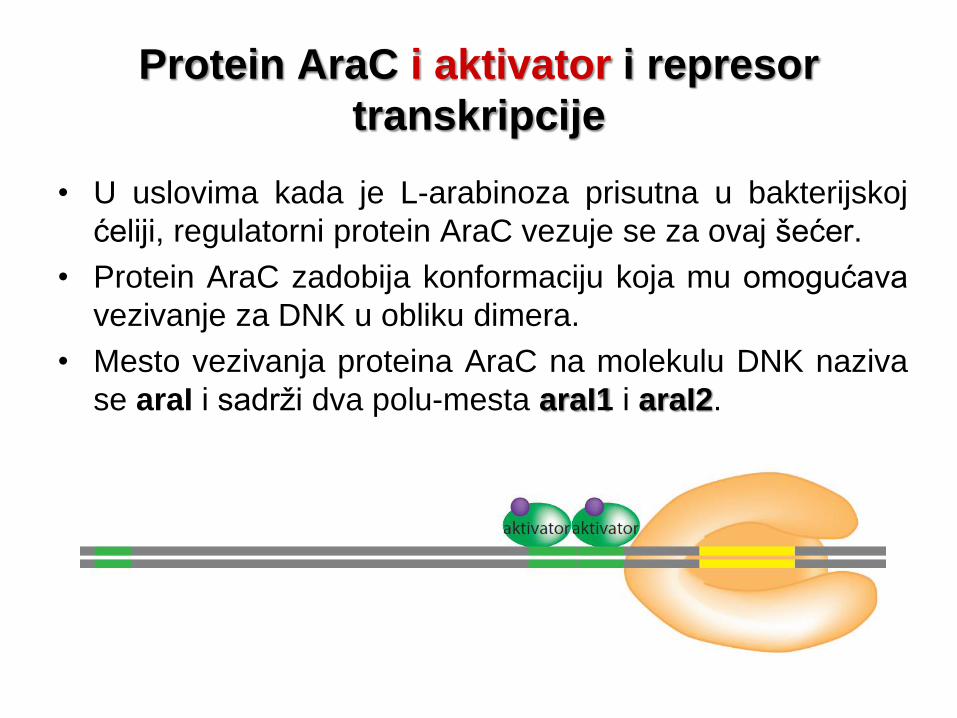
Protein AraC i aktivator i represor
transkripcije
• U uslovima kada je L-arabinoza prisutna u bakterijskoj
ćeliji, regulatorni protein AraC vezuje se za ovaj šećer.
• Protein AraC zadobija konformaciju koja mu omogućava
vezivanje za DNK u obliku dimera.
• Mesto vezivanja proteina AraC na molekulu DNK naziva
se araI i sadrži dva polu-mesta araI1 i araI2.
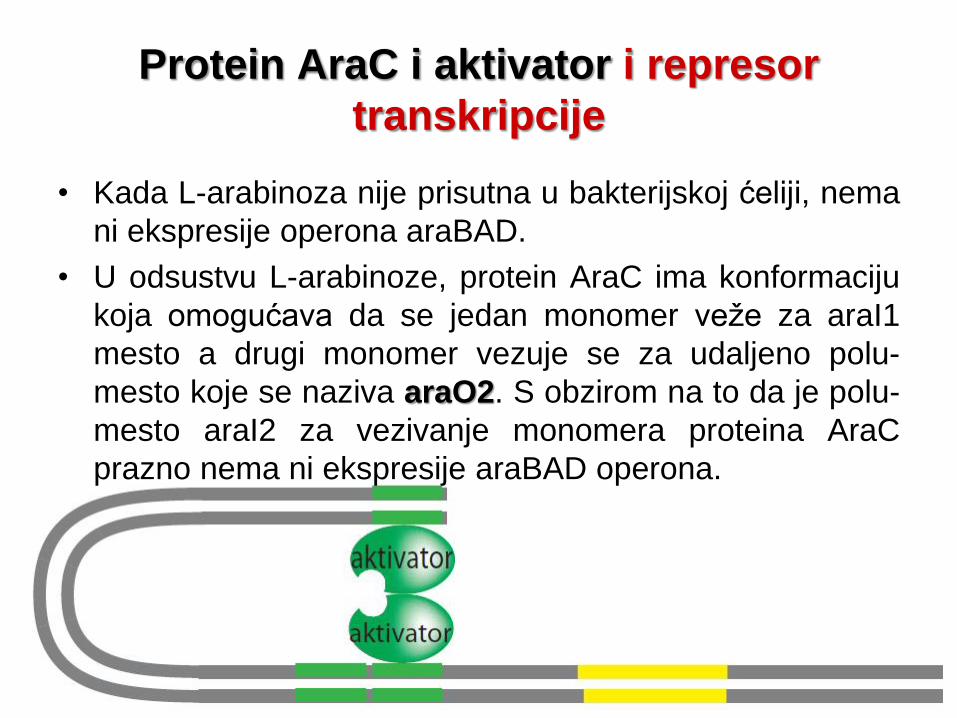
Protein AraC i aktivator i represor
transkripcije
• Kada L-arabinoza nije prisutna u bakterijskoj ćeliji, nema
ni ekspresije operona araBAD.
• U odsustvu L-arabinoze, protein AraC ima konformaciju
koja omogućava da se jedan monomer veže za araI1
mesto a drugi monomer vezuje se za udaljeno polu-
mesto koje se naziva araO2. S obzirom na to da je polu-
mesto araI2 za vezivanje monomera proteina AraC
prazno nema ni ekspresije araBAD operona.

Regulacija ekspresije gena kod
prokariota nakon inicijacije
transkripcije
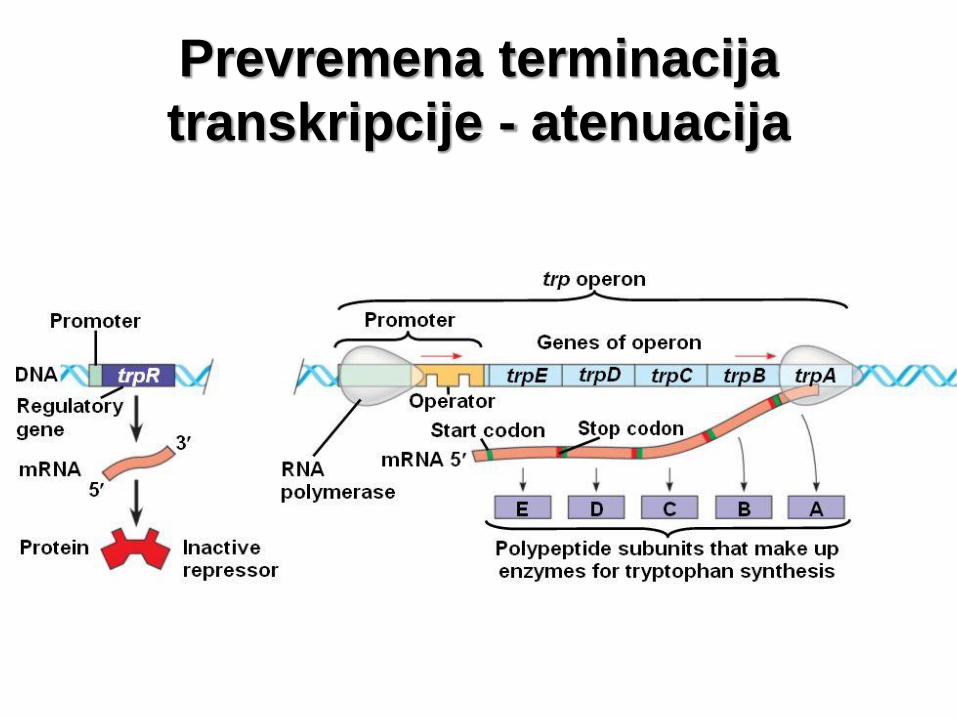
Prevremena terminacija
transkripcije - atenuacija
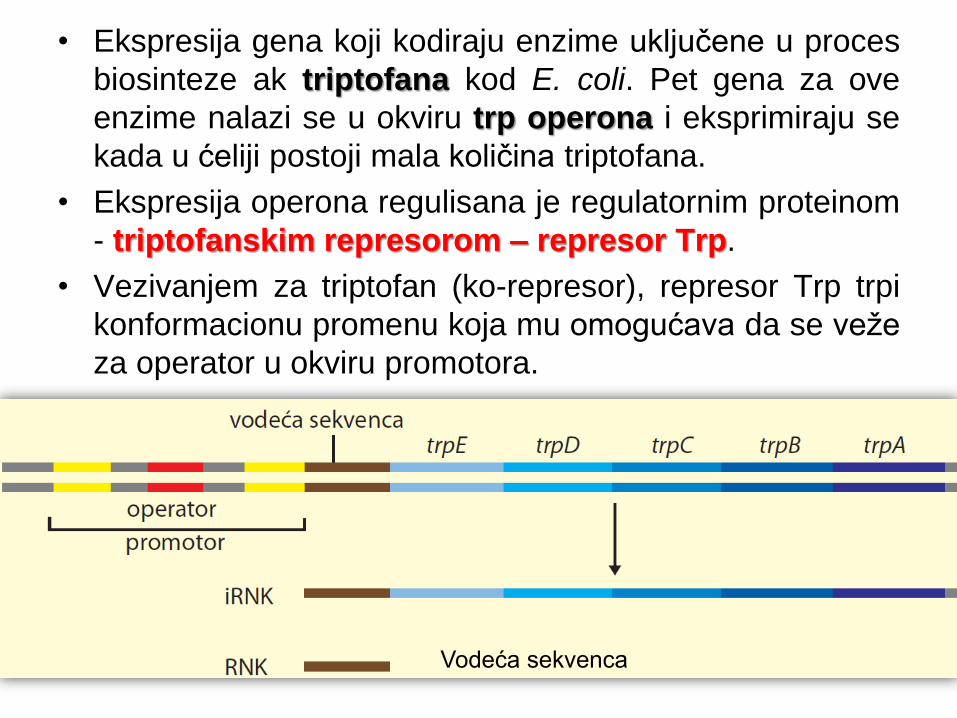
• Ekspresija gena koji kodiraju enzime uključene u proces
biosinteze ak triptofana kod E. coli. Pet gena za ove
enzime nalazi se u okviru trp operona i eksprimiraju se
kada u ćeliji postoji mala količina triptofana.
• Ekspresija operona regulisana je regulatornim proteinom
- triptofanskim represorom – represor Trp.
• Vezivanjem za triptofan (ko-represor), represor Trp trpi
konformacionu promenu koja mu omogućava da se veže
za operator u okviru promotora.
Vodeća sekvenca
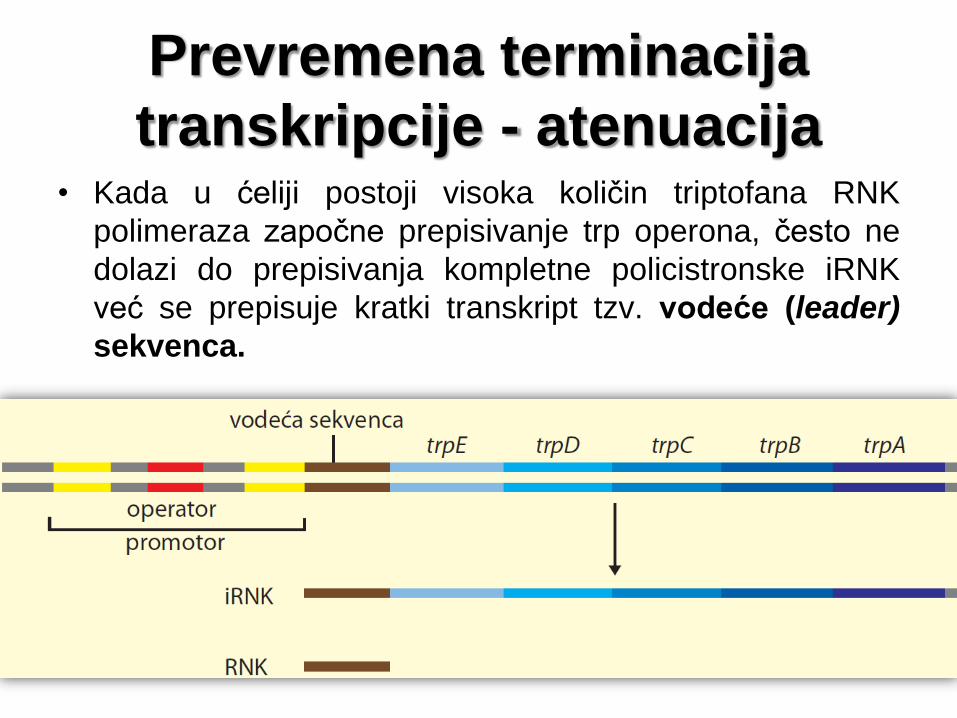
Prevremena terminacija
transkripcije - atenuacija • Kada u ćeliji postoji visoka količin triptofana RNK
polimeraza započne prepisivanje trp operona, često ne
dolazi do prepisivanja kompletne policistronske iRNK
već se prepisuje kratki transkript tzv. vodeće (leader)
sekvenca.

5' kraj policistronske iRNK trp operona
Vodeća sekvenca
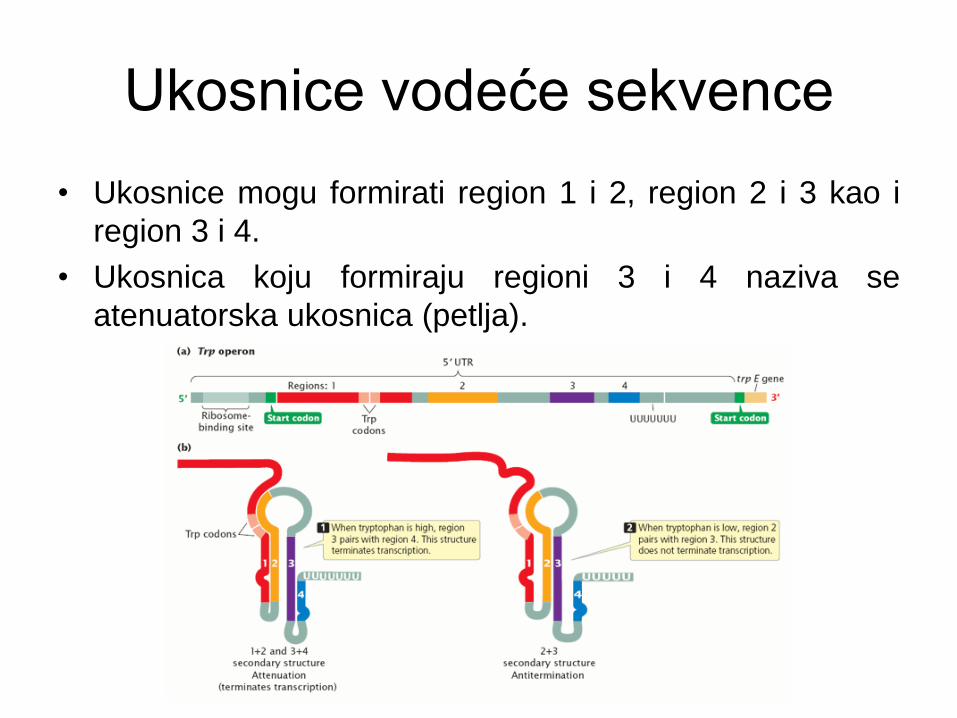
Ukosnice vodeće sekvence
• Ukosnice mogu formirati region 1 i 2, region 2 i 3 kao i
region 3 i 4.
• Ukosnica koju formiraju regioni 3 i 4 naziva se
atenuatorska ukosnica (petlja).
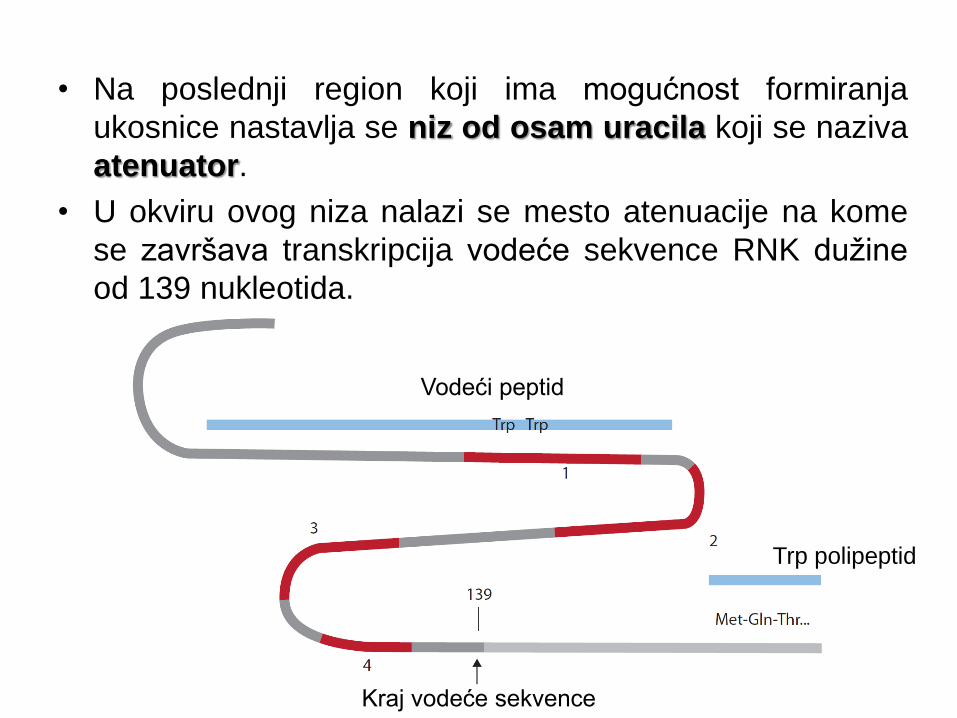
• Na poslednji region koji ima mogućnost formiranja
ukosnice nastavlja se niz od osam uracila koji se naziva
atenuator.
• U okviru ovog niza nalazi se mesto atenuacije na kome
se završava transkripcija vodeće sekvence RNK dužine
od 139 nukleotida.
Vodeći peptid
Trp polipeptid
Kraj vodeće sekvence

Atenuacija
• U okviru otvorenog okvira čitanja vodeće sekvence (u
regionu 1 koji je odgovoran za formiranje ukosnice)
nalaze se dva uzastopna kodona za triptofan.
• Njihova funkcija je da zaustave ribozom za vreme
translacije leader peptida.
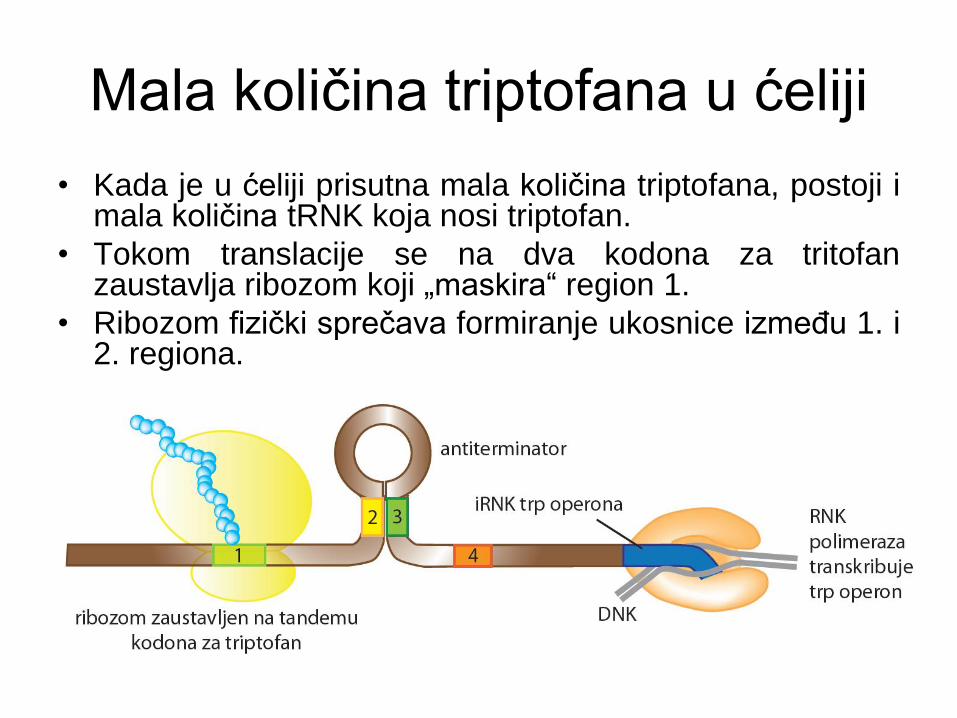
Mala količina triptofana u ćeliji
• Kada je u ćeliji prisutna mala količina triptofana, postoji i mala količina tRNK koja nosi triptofan.
• Tokom translacije se na dva kodona za tritofan zaustavlja ribozom koji „maskira“ region 1.
• Ribozom fizički sprečava formiranje ukosnice između 1. i 2. regiona.
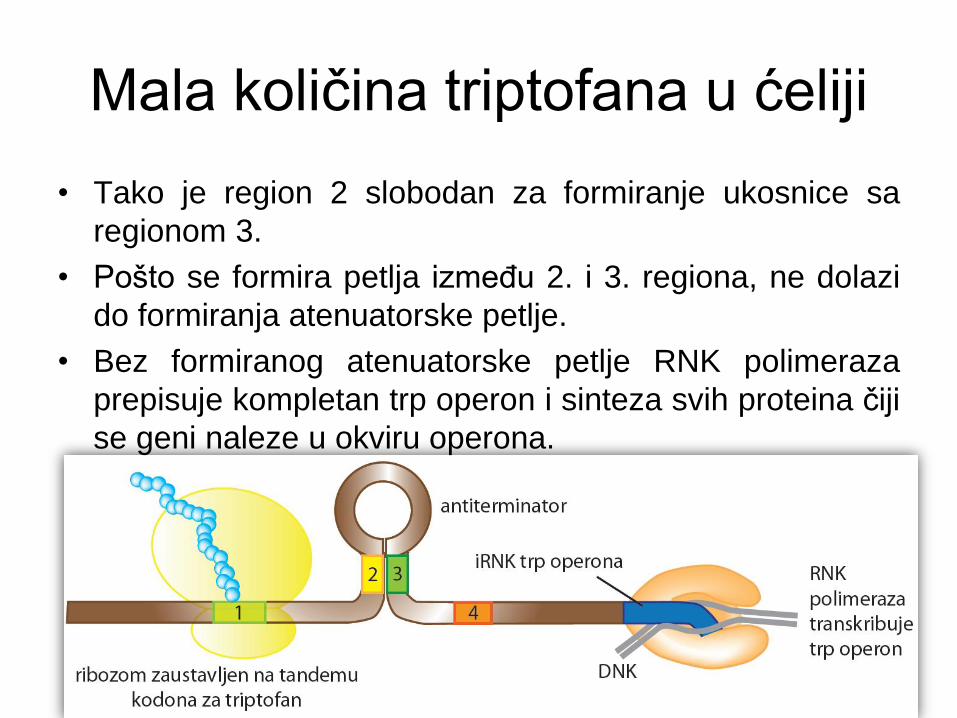
Mala količina triptofana u ćeliji
• Tako je region 2 slobodan za formiranje ukosnice sa
regionom 3.
• Pošto se formira petlja između 2. i 3. regiona, ne dolazi
do formiranja atenuatorske petlje.
• Bez formiranog atenuatorske petlje RNK polimeraza
prepisuje kompletan trp operon i sinteza svih proteina čiji
se geni naleze u okviru operona.
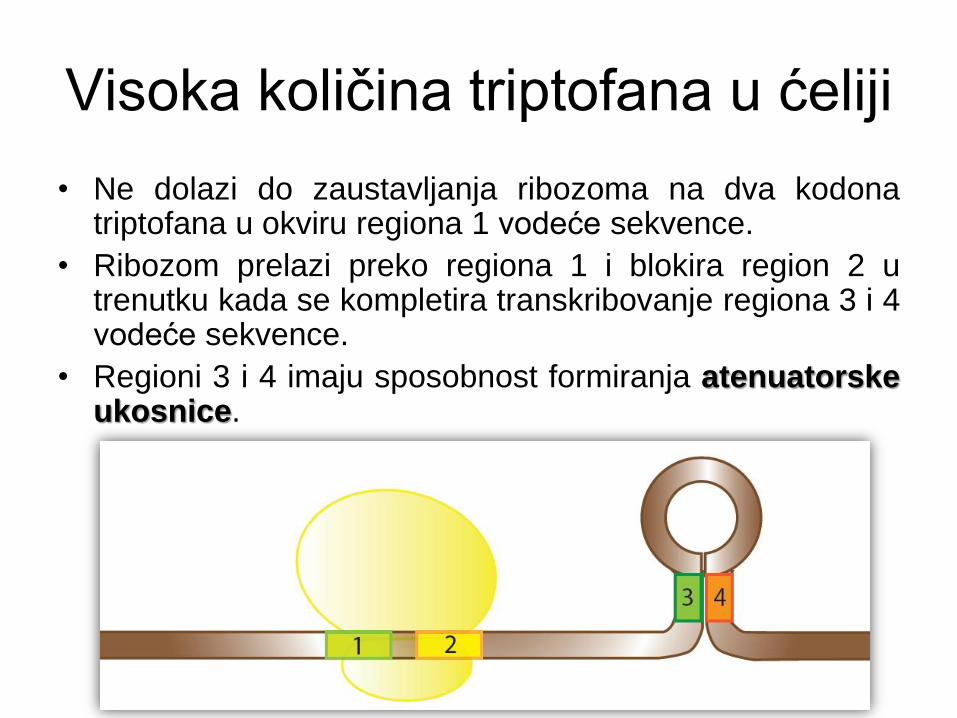
Visoka količina triptofana u ćeliji
• Ne dolazi do zaustavljanja ribozoma na dva kodona triptofana u okviru regiona 1 vodeće sekvence.
• Ribozom prelazi preko regiona 1 i blokira region 2 u trenutku kada se kompletira transkribovanje regiona 3 i 4 vodeće sekvence.
• Regioni 3 i 4 imaju sposobnost formiranja atenuatorske ukosnice.
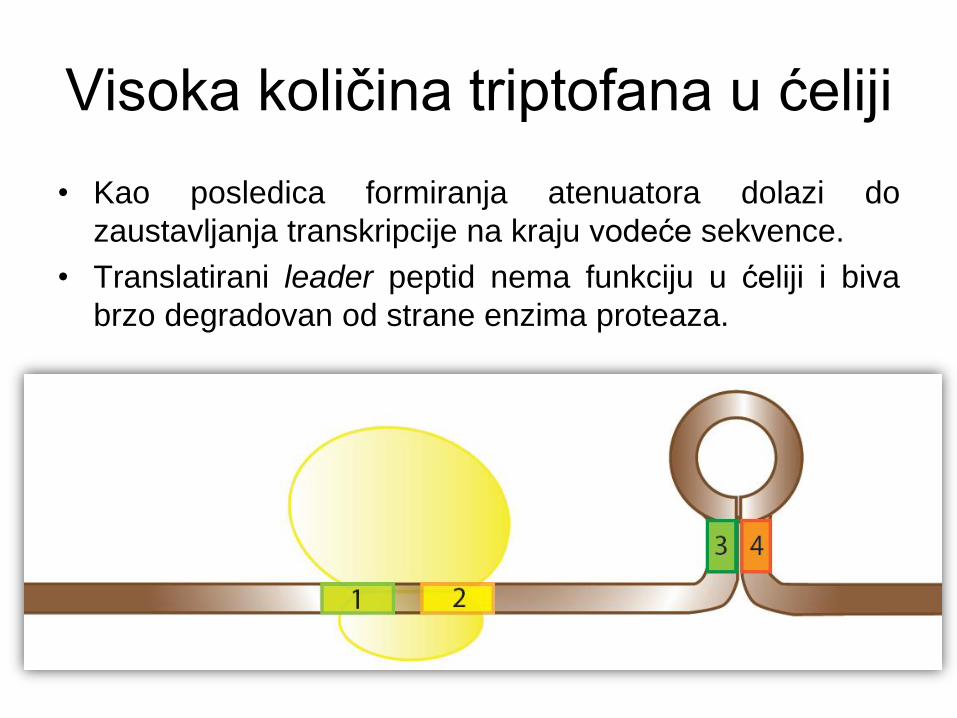
Visoka količina triptofana u ćeliji
• Kao posledica formiranja atenuatora dolazi do
zaustavljanja transkripcije na kraju vodeće sekvence.
• Translatirani leader peptid nema funkciju u ćeliji i biva
brzo degradovan od strane enzima proteaza.
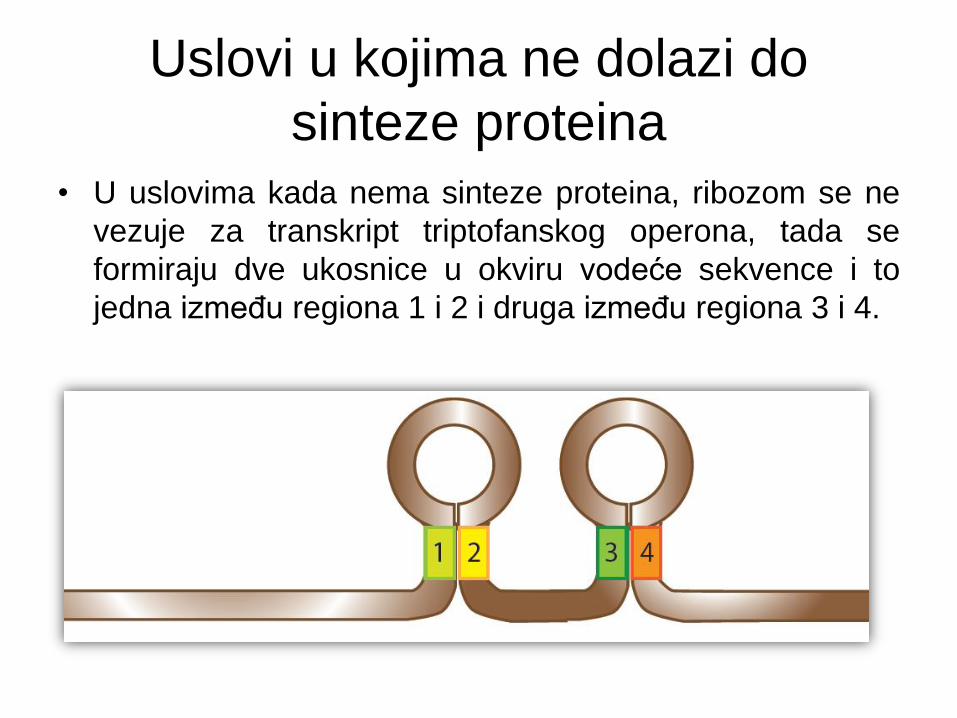
Uslovi u kojima ne dolazi do
sinteze proteina
• U uslovima kada nema sinteze proteina, ribozom se ne
vezuje za transkript triptofanskog operona, tada se
formiraju dve ukosnice u okviru vodeće sekvence i to
jedna između regiona 1 i 2 i druga između regiona 3 i 4.

Regulacija ekspresije gena
translacionom represijom

Regulacija ekspresije gena
translacionom represijom
• Regulacija ekspresije gena translacionom represijom podrazumeva postojanje neke vrste „represora“ koji blokira proces translacije.
• Primer ove vrste regulacije ekspresije gena su ribozomski proteini (r-proteini) koji su translacioni represori sopstvene sinteze.
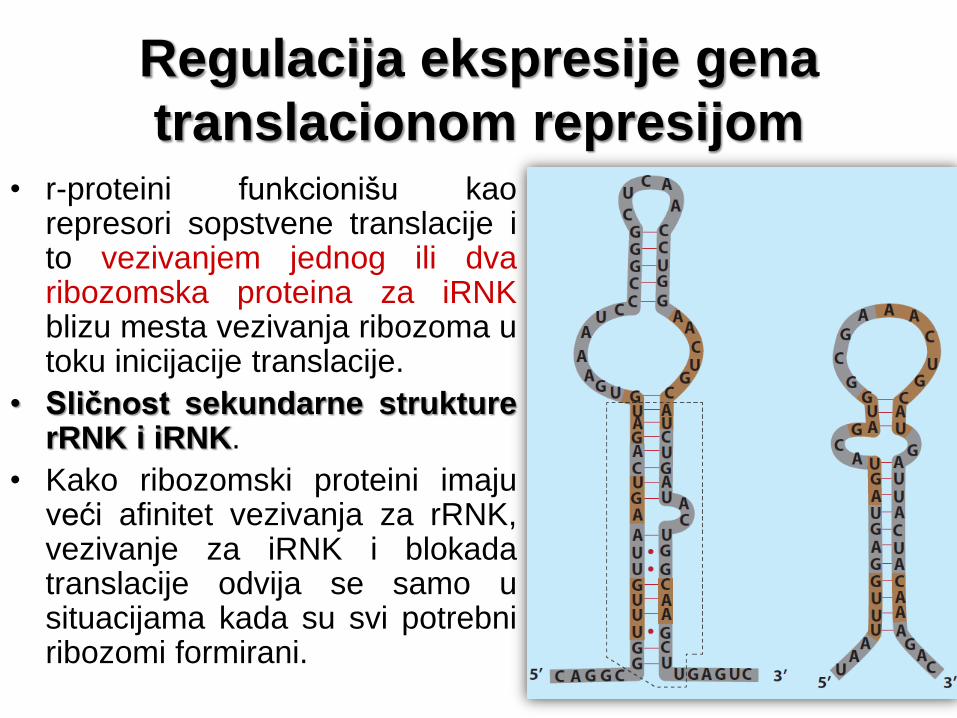
Regulacija ekspresije gena
translacionom represijom • r-proteini funkcionišu kao
represori sopstvene translacije i to vezivanjem jednog ili dva ribozomska proteina za iRNK blizu mesta vezivanja ribozoma u toku inicijacije translacije.
• Sličnost sekundarne strukture rRNK i iRNK.
• Kako ribozomski proteini imaju veći afinitet vezivanja za rRNK, vezivanje za iRNK i blokada translacije odvija se samo u situacijama kada su svi potrebni ribozomi formirani.
Uloga molekula RNK u
regulaciji ekspresije kod
prokariota

Mali molekuli RNK kod
prokariota • U bakterijama je detektovano prisustvo molekula RNK
koji po svojoj strukturi i funkciji ne pripadaju ni jednoj od tri najčešće grupe molekula RNK (iRNK, rRNK i tRNK).
• To su mali molekuli RNK (eng. small RNA, sRNA) veličine od 80 do 110 nukleotida.
• Nazivaju se i nekonvencionalne, funkcionalne ili regulatorne RNK.
• Samo je u genomu E. coli detektovano preko 100 gena koji kodiraju za sRNK.
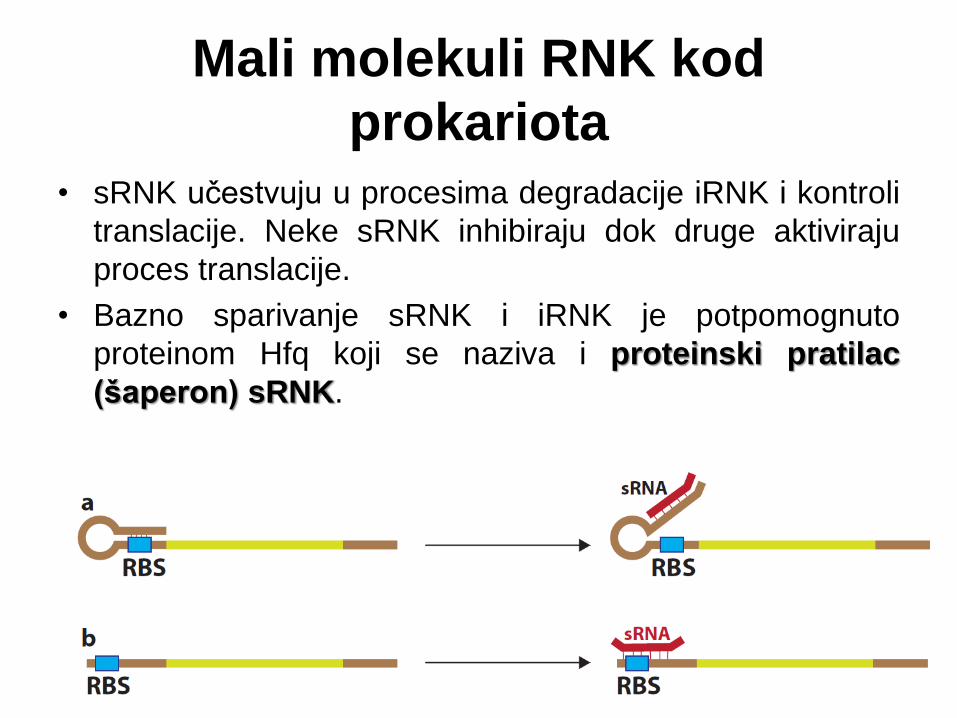
Mali molekuli RNK kod
prokariota
• sRNK učestvuju u procesima degradacije iRNK i kontroli
translacije. Neke sRNK inhibiraju dok druge aktiviraju
proces translacije.
• Bazno sparivanje sRNK i iRNK je potpomognuto
proteinom Hfq koji se naziva i proteinski pratilac
(šaperon) sRNK.
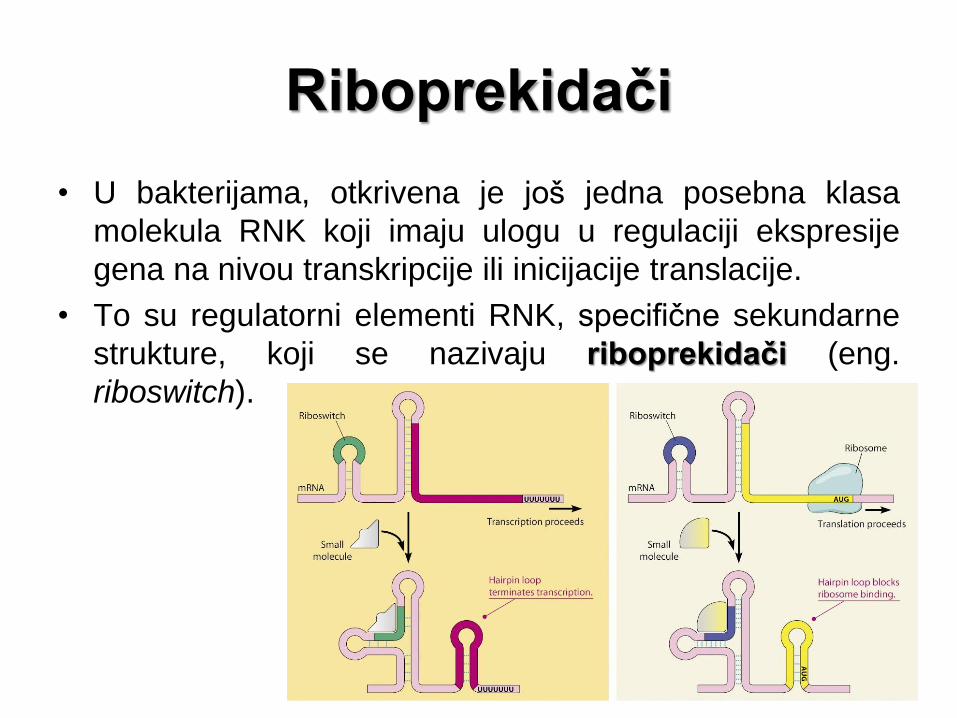
Riboprekidači
• U bakterijama, otkrivena je još jedna posebna klasa
molekula RNK koji imaju ulogu u regulaciji ekspresije
gena na nivou transkripcije ili inicijacije translacije.
• To su regulatorni elementi RNK, specifične sekundarne
strukture, koji se nazivaju riboprekidači (eng.
riboswitch).
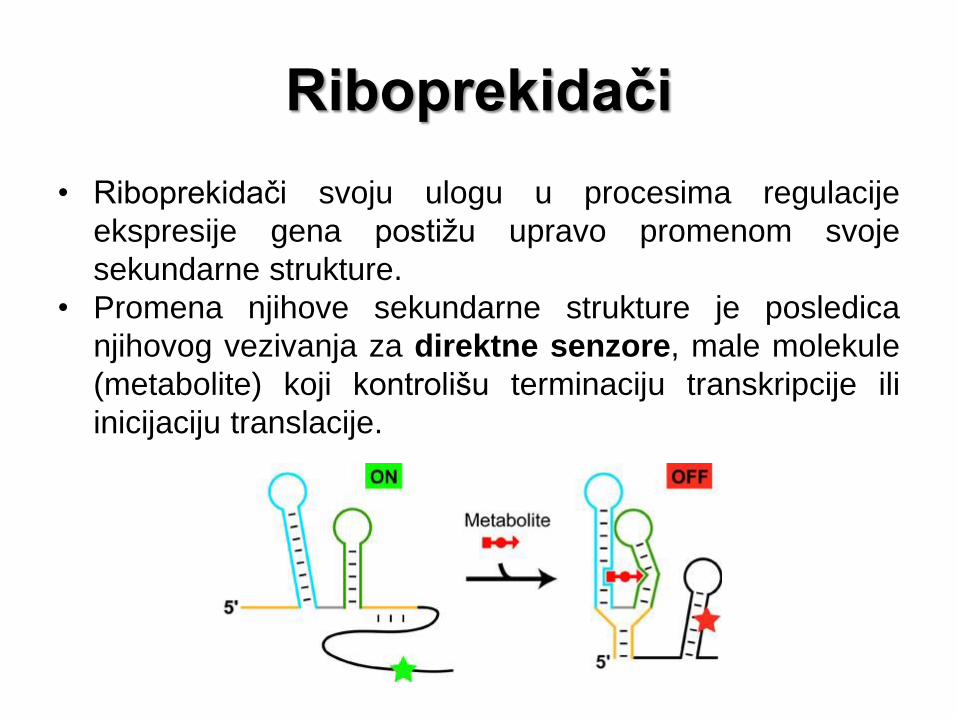
Riboprekidači
• Riboprekidači svoju ulogu u procesima regulacije
ekspresije gena postižu upravo promenom svoje
sekundarne strukture.
• Promena njihove sekundarne strukture je posledica
njihovog vezivanja za direktne senzore, male molekule
(metabolite) koji kontrolišu terminaciju transkripcije ili
inicijaciju translacije.
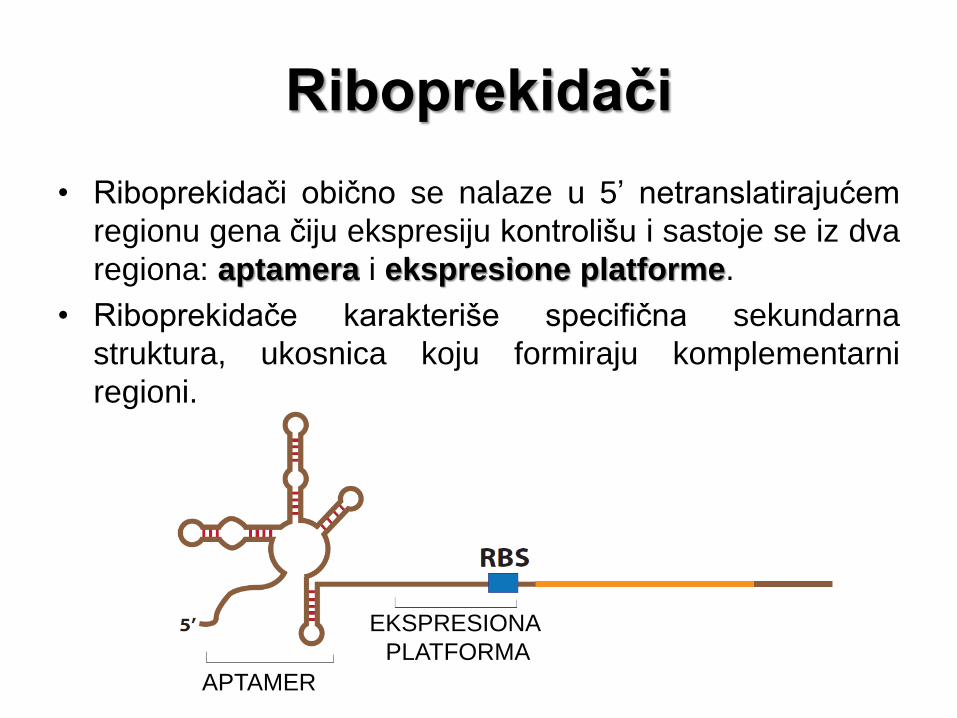
Riboprekidači
• Riboprekidači obično se nalaze u 5’ netranslatirajućem
regionu gena čiju ekspresiju kontrolišu i sastoje se iz dva
regiona: aptamera i ekspresione platforme.
• Riboprekidače karakteriše specifična sekundarna
struktura, ukosnica koju formiraju komplementarni
regioni.
APTAMER
EKSPRESIONA
PLATFORMA
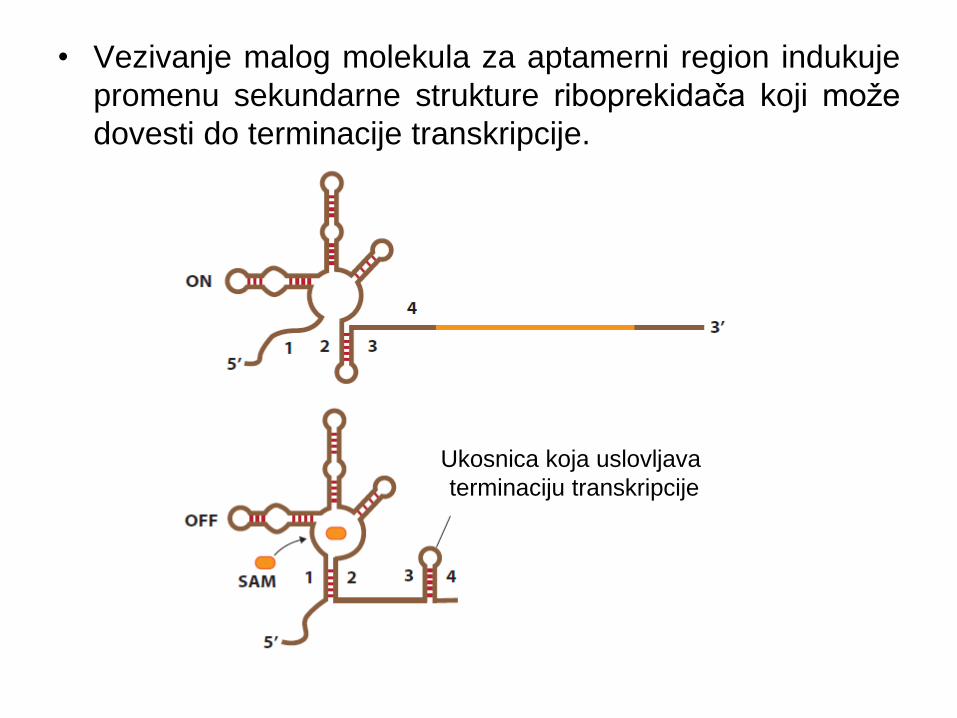
• Vezivanje malog molekula za aptamerni region indukuje
promenu sekundarne strukture riboprekidača koji može
dovesti do terminacije transkripcije.
Ukosnica koja uslovljava
terminaciju transkripcije

Uloga riboprekidača u inhibiciji
translacije
• U odsustvu senzornog molekula, riboprekidač poseduje
sekundarnu strukturu koja omogućava nesmetano
odvijanje translacije.
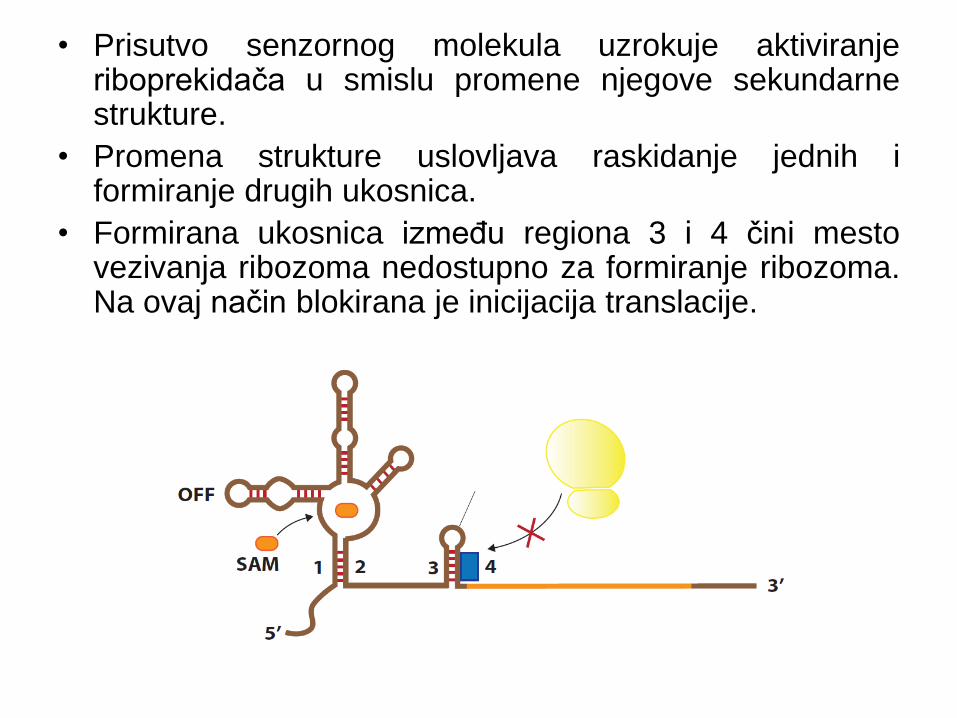
• Prisutvo senzornog molekula uzrokuje aktiviranje riboprekidača u smislu promene njegove sekundarne strukture.
• Promena strukture uslovljava raskidanje jednih i formiranje drugih ukosnica.
• Formirana ukosnica između regiona 3 i 4 čini mesto vezivanja ribozoma nedostupno za formiranje ribozoma. Na ovaj način blokirana je inicijacija translacije.