Embed Size (px)
Citation preview

The Plant Cell, Vol. 7, 797-807, July 1995 Q 1995 American Society of Plant Physiologists
Regulation of Photosynthesis in C3 and C4 Plants: A Molecular Appmach
Robert T. Furbank and Wllliam C. Taylor’ Commonwealth Scientific and Industrial Research Organization, Division of Plant Industry, GPO Box 1600, and Cooperative Research Centre for Plant Science, Australian National University, GPO Box 475, Canberra 2601, Australia
INTRODUCTION
Most plants use the C3 pathway of photosynthesis, also called the photosynthetic carbon reduction cycle (PCR), shown in Fig- ure 1A. C3 plants have a single chloroplast type that performs all of the reactions that convert light energy into the chem- ical energy that is used to fix COp and to synthesize the re- duced carbon compounds upon which all life depends. Ribulose-i ,5-bisphosphate carboxylaseloxygenase (Rubisco) catalyzes primary carbon fixation, in which a fivecarbon sugar phosphate, ribulose-l,5-bisphosphate (RuBP), and COp are converted to two molecules of the threecarbon compound 3-phosphoglycerate (hence the name C3). Phosphoglycerate is then phosphorylated and reduced by the products of the light reactions of photosynthesis (ATP and NADPH) to produce triose phosphate (TP). TP can be exported from the chloroplast via the chloroplast envelope phosphate (Pi) transporter to the cytosol and used in the synthesis of sucrose, which is then translocated throughout the plant (see Sonnewald et al., 1994), or it can be retained within the chloroplast for starch synthe- sis or recycling to RuBP. Rubisco also catalyzes the fixation of Op in a process known as photorespiration, which competes directly with fixation of COp. At air levels of Coa, for every three COp molecules fixed by Rubisco to form 3-phosphoglyc- erate, approximately one O2 molecule is fixed, producing Sphosphoglycerate and 3-phosphoglycolate (Figure 1A). Be- cause 3-phosphoglycolate cannot be used in the PCR cycle, it must be recycled to phosphoglycerate via the photorespira- tory pathway, expending ATP and NADPH. This competition between 0 2 and COp and the energy costs associated with recycling phosphoglycolate largely determine the efficiency of C3 photosynthesis in air (Hatch, 1988; Woodrow and Berry, 1988).
The C4 pathway is a complexadaptation of the C3 pathway that overcomes the limitation of photorespiration and is found in a diverse collection of species, many of which grow in hot climates with sporadic rainfall. The C4 pathway effectively suppresses photorespiration by elevating the C02 concentra- tion at the site of Rubisco using a biochemical C02 pump. C4 plants have two chloroplast types, each found in a specialized cell type. Leaves of C4 plants show extensive vascularization,
To whom correspondence should be addressed.
with a ring of bundle sheath (8) cells surrounding each vein and an outer ring of mesophyll (M) cells surrounding the bun- dle sheath. The development of this socalled Kranz anatomy and the cell-specific compartmentalization of C4 enzymes are important features of C4 photosynthesis (Hatch, 1988, and references therein). C02 fixation in these plants is a two-step process. Atmospheric C02 is initially fixed in the cytosol of M cells by phosphoenolpyruvate carboxylase (PEPC) to form the four-carbon dicarboxylic acid oxaloacetate (hence the name C4), which is converted to malate or aspartate (Figure 16). These C4 acids then diffuse into the inner ring of B cells, where they are decarboxylated in the chloroplasts. The COp produced is then refixed by Rubisco. The mechanism of decar- boxylation in B chloroplasts varies among the three different C4 types. We confine our discussion to the most extensively studied type, the NADP-malic enzyme (ME) type, which is named for its B cell decarboxylating activity.
The key feature of C4 photosynthesis is the compartmen- talization of activities into two specialized cell and chloroplast types. Rubisco and the C3 PCR cycle are found in the inner ring of B cells. These cells are separated from the mesophyll cells and from the air in the intercellular spaces by a lamella that is highly resistant to the diffusion of C02 (Hatch, 1988). Thus, by virtue of this two-stage Coe fixation pathway, the mesophyll-located C4 cycle acts as a biochemical CO2 pump to increase the concentration of COp in the bundle sheath an estimated 10-fold over atmospheric concentrations. The net result is that the oxygenase activity of Rubisco is effectively suppressed and the PCR cycle operates more efficiently. C4 plants show higher rates of photosynthesis at high light inten- sities and high temperatures due to the increased efficiency of the PCR cycle (Hatch, 1988). In favorable environments, C4 plants outperform C3 plants, making them the most produc- tive crops and the worst weeds. Maize, sugarcane, sorghum, and amaranth are examples of C4 crops, and nutgrass (Cyperus mtundus), crabgrass (Dighria sanquinalis), and barn- yard grass (Echinochloa crusgalli) are some of the worst C4 weeds.
Although the pathways of C3 and C4 photosynthesis are well established and the properties of individual enzymes are reasonably well understood, there remain severa1 areas of

798 The Plant Cell
A
Cyl b6f
ADP ATP
CHLOROPLAST
STARCH i SUCROSE
B
Figure 1. Simplified Schemes of the C3 and C4 Photosynthetic Pathways.
(A) C3 pathway (photosynthetic carbon reduction or PCR cycle), em- phasizing the enzymes that have been manipulated using molecular genetic techniques. Carbon initially fixed by Rubisco is phosphorylated and reduced by the products of the light reactions (ATP and NADPH). The reduced three-carbon sugar-phosphate (triose phosphate, TP) can be either exported from the chloroplast for sucrose synthesis via the chloroplast envelope Pi transporter (Pi TRANS) or retained for starch synthesis or recycling to ribulose bisphosphate, the COn acceptor for the Rubisco enzyme. CA, carbonic anhydrase, catalyzes HC03 .-+ Coe; RUBSCO, ribulose-l,5-bisphosphate carboxylase-oxygenase, catalyzes ribulose bisphosphate + COn -, 2X Sphosphoglycerate or ribulose bisphosphate + O2 -, Sphosphoglycerate + phosphoglyco- late; GAPDH, glyceraldehyde Sphosphate dehydrogenase, catalyzes 1,Sdiphosphoglycerate + NADPH - glyceraldehyde Sphosphate + NADP+; FBPase, fructose l,Sbisphosphatase, catalyzes fructose bisphosphate -t fructose Sphosphate + Pi; PRK, phosphoribulokinase, catalyzes ribuloseSphosphate + ATP- ribulose bisphosphate + ADI? (B) C4 pathway, simplified to describe only the NADP-malic enzyme type, which transports malate from the mesophyll to the bundle sheath chloroplasts. Compartmentation of enzymes belween the two cell types is shown. COn (in the form of bicarbonate) is fixed by the enzyme PEPC to form the C4 acid oxaloacetate (OAA), which is reduced by NADPH from the light reactions to form malate (MAL), the C, acid that is transported to the bundle sheath cells. Malate is decarboxylated
relative ignorance. One concerns the process of assembly of thel photosynthetic apparatus during chloroplast development. How are the synthesis and assembly of these multimeric pro- tein complexes regulated? Another concerns the ways the net activity of each pathway is controlled as the plant encounters a range of environmental conditions each day and through- out its life. We show how molecular genetic techniques are contributing to a new understanding of these issues.
HOW GENE REGULATION CONTROLS C3 AND Ca ENZYME ACTlVlTlES
The rapid assembly of the photosynthetic apparatus is crucial for the transition from heterotrophic to photoautotrophic growth in newly germinated seedlings. Rapid assembly is made pos- sible by high levels of the cytosolic and chloroplastic mRNAs encoding photosynthetic proteins. High levels of nuclear- encoded mRNAs result from a combination of environmental and developmental controls on gene transcription. The small gene families coding for the small subunit (SSU) of Rubisco (the RbcS genes) and the chlorophyll alb apoproteins of the photosystem II light-hawesting complex (the Cab genes, which have recently been renamed the Lhcb genes) have served as paradigms for these studies. Light, acting through phytochrome and blue light photoreceptors, induces high rates of transcrip- tion of these genes (Tobin and Silverthorne, 1985; Thompson and White, 1991). cis-Acting DNA sequences responsible for light-induced transcription have been identified by fusing non- coding regions of the RbcS and Cab genes to reporter genes and measuring their activities in transgenic plants (Gilmartin et al., 1990). The signal transduction pathway from the pho- toreceptors to the transcription factors that activate transcription of Cab and RbcS genes is currently being dissected by mo- lecular genetic approaches, primarily in Arabidopsis (Deng, 1994). Light also acts on the transcription of Cab and RbcS genes by setting a circadian clock that has a major effect on the transcription of these genes and probably a lesser effect on the transcription of other nuclear genes (Giuliano et al., 1988; Nagy et al., 1988; Taylor, 1989a).
Although developmental regulation has been less well stud- ied than light regulation, its existence is evident from the restriction of Cab and RbcS gene expression to chlorenchy-
in the bundle sheath cells, where the C02 released is fixed by the PCR cycle in much the same way as in C3 plants. The threecarbon compound pyruvate (PYR) diffuses back to the mesophyll, where it is phosphorylated by ATP to regenerate the carbon acceptor phos- phoenolpyruvate (PEP). PEPC, phosphoenolpyruvate carboxylase, catalyzes phosphoenolpyruvate + HC03- - oxaloacetate; MDH, ma- late dehydrogenase. catalyzes oxaloacetate + NADPH - malate + NADP; NADPME, NADP-malic enzyme, catalyzes malate + NADP -, pyruvate + NADPH; PWK, pyruvate orthophosphate dikinase, cata- lyzes pyruvate + ATP + Pi -, phosphoenolpyruvate + AMP + PPi.

Regulation of Photosynthesis 799
mous leaf cells in C3 plants (Muller et al., 1980; Eckes et al., 1985). A signal from the developing chloroplast is required for high-leve1 transcription of Cab and RbcS genes, even when light signals are fully activated (Oelmuller, 1989; Taylor, 1989b). This chloroplast signal may be at least partially responsible for spatial regulation of nuclear genes coding for chloroplast proteins.
Three main features distinguish C4 plants from C3 plants: (1) the differentiation of the two cell and chloroplast types, (2) the presence of an additional set of genes, and (3) a mecha- nism regulating the cell-specific expression of these additional genes. Very little is known about how the differentiation of the two cell types is regulated. The position of cells relative to de- veloping veins seems to determine their fate, with those in close association with the vein becoming B cells and those at least one cell removed from the vein becoming M cells (Nelson and Langdale, 1992).
Genes encoding C4 enzymes are, in most cases, members of small gene families. Other members of each gene family code for isozymic forms that perform nonphotosynthetic func- tions; these non-C4 genes are usually expressed at low levels in a wide range of cells. C4 enzymes are found at high levels in either B or M cells. Therefore, genes encoding C4 enzymes must have evolved new programs of gene expression.
Two experimental approaches have been used to study the mechanisms regulating the cell-specific expression of C4 genes. One is the use of gene and antibody probes to localize mRNAs and proteins at various stages in leaf development. These studies have shown that light induces high-leve1 expres- sion of C4 genes, whereas cell-specific expression of some C4 genes is controlled by a light-independent developmental program. However, for other genes, light controls both high- level expression and cell specificity. For instance, Sheen and Bogorad (1985) found that transcripts for both RbcS and rbcL (a chloroplast gene that codes for the Rubisco large subunit, LSU) are present in both B and M cells of etiolated maize leaves. Light is necessary to induce high levels of both transcripts and to suppress their accumulation in M cells. In contrast, light does not affect the spatial pattern of C4 mRNA accumulation in amaranth leaves (Wang et al., 1993). However, RbcS and rbcL transcripts and proteins can be detected in both cell types at early stages of development but then become restricted to B cells (Wang et al., 1992). Accumulation of transcripts encod- ing C4 enzymes appears to be cell specific in both plants (Nelson and Langdale, 1989), although Wang et al. (1992) detected transcripts coding for PEPC in both cell types of amaranth. However, as they pointed out, it is possible that the B cell transcripts encode a nonphotosynthetic isoform.
A complementary approach isto identify cis-acting DNA se- quences responsible for cell-specific expression and to use these sequences to unravel the mechanisms that control high- level, cell-specific expression. Until recently, the lack of an ef- ficient, stable transformation system for any C4 plant has slowed progress using this approach. Therefore, one strategy has been to study the expression of C4 gene promoters in
transgenic C3 plants to identify sequences responsible for high-level, organ-specific expression.
F? Westhoff and colleagues have used this strategy to de- termine how the C4 members of the gene family encoding PEPC differ from members coding for nonphotosynthetic iso- forms. Sequence comparisons indicated that in the C4 dicot Haveria frinervia, these Ppc genes fall into four subfamilies (Hermans and Westhoff, 1990). The PpcA subfamily codes for the C4 isoform, and its members are expressed at high lev- els in M cells. The C3 species E pringlei also has PpcA genes, but these are expressed at low levels in leaves, roots, and stems. The PpcA genes of these two species are more similar to one another than either is to Ppc genes of other subfami- lies within the same species. Stockhaus et al. (1995) determined that the E frinervia PpcA gene has unique cis-acting sequences that are at least partially responsible for its high-leve1 expres- sion in leaves by comparing the expression in transgenic tobacco of gene fusions in which 5' regions from E frinervia and E pringlei PpcA genes were fused to the P-glucuronidase A(gusA) reporter gene. Sequences between -2118 and -500 of the E frinervia PpcA promoter conferred high-leve1 reporter gene expression in tobacco leaves, primarily in the palisade parenchyma, whereas promoter sequences from the E prin- g1ei PpcA conferred lower level gusA expression in roots and stems and very low expression in leaves. Whether the preferem tia1 expression of the E frinervia PpcA gene in tobacco palisade parenchyma is due to M cell-specific control remains to be seen.
Similar experiments by M. Matsuoka and colleagues re- vealed that cis-acting sequences from the 5' regions of the maize RbcS, Pepc, and Ppdk (which codes for M cell pyruvate orthophosphate dikinase, PPdK) genes confer light-regulated reporter gene expression in transgenic rice plants (Matsuoka et al., 1993,1994). In all three cases, leaf M cells preferentially expressed the reporter gene, despite the fact that RbcS is B cell specific in maize. Although rice leaves have B cells, these contain only a few chloroplasts, which may account for the rel- atively greater expression in M cells. Nove1 cis-acting sequences at the 5' ends of some C4 genes are therefore an important component of the mechanism that controls high-leve1 leaf-specific and light-regulated expression of these genes.
Transient expression studies of C4 gene promoter con- structs introduced into leaf cells of C4 plants have also provided clues about cell-specific regulation. Schaffner and Sheen (1992) showed that B'sequences from a maize C4 Aspc gene that are not present in a closely related n0n-G gene confer high-leve1 light-regulated expression in maize leaf pro- toplasts. Using microprojectile bombardment of maize leaf sections, Bansal and Bogorad (1993) have identified separate sites in the upstream region of a maize Cab gene that control light responses and cell specificity. Although Cab expression is not strictly cell specific in most C4 plants, this particular gene is expressed primarily in M cells. This cell preference appears to be due to the combination of enhanced expres- sion in M cells and suppression of B cell activity. Transient

800 The Plant Cell
expression experiments (T. Nelson, personal communication) have shown that 5’sequences of RbcS genes from F: trinervia (C4) and F: pringlei (C3) specify different expression patterns in E frherVia leaves. These results imply that, for at least some C4 genes, the cis-acting sequences controlling cell specific- ity are located at the 5’end of the gene and are found only in the genes from C4 species.
To study further the mechanisms controlling C4 gene ex- pression and the regulation of C4 enzyme activities (see following sections), we have developed an Agrobacterium- mediated transformation system for the C4 dicot Flaveria bi- dentis (Chitty et al., 1994). This system is reasonably fast and efficient, giving transformed plants 15 to 20 weeks after ex- plant cocultivation, and it has already provided some interesting insights. In E bidentis, two genes code for chloroplastic forms of ME, one of which, MeA, encodes the C4 form, which is ex- pressed at high levels and in a light-regulated fashion in 6 cells (J.S. Marshall, J.D. Stubbs, and W.C. Taylor, unpublished data). E bidenfis plants stably transformed with a series of 5‘MeA sequences fused to gusA show very low GUS activity in leaves but high levels in meristems and moderately high levels in stems and anthers (J.S. Marshall, J.A. Chitty, J.D. Stubbs, and W.C. Taylor, unpublished data). The longest of these constructs had 2.2 kb of 5‘ noncoding sequence, whereas the shortest had 0.39 kb. However, when 5.8 kb of sequence at the 3’ end of MeA was added to the longest S’construct, high-level leaf expression was attained. Preliminary analysis of primary trans- genic plants suggests that this expression occurs primarily in 6 cells.
In Flaveria species, PPdK is encoded by a single gene, Wk (E. Rosche and I? Westhoff, personal communication; C.J. Chastain, M. Matsuoka, and W.C. Taylor, unpublished data). Pdk encodes two different isoforms, a prevalent chloroplast form located in M cells and a presumably nonchloroplast form, the transcript of which is found in all organs at very low levels. Primary transformants with 1.5 kb of Wk 5‘ sequence fused to gusA show high levels of GUS activity in leaves (W.C. Taylor, J.A. Chitty, and E. Rosche, unpublished observations). Tran- scription of the mRNA encoding the nonchloroplast form of PPdK is driven by a promoter located in the large 6-kb intron of the Wk gene; similar results have been obtained with the maize Wk gene, which also encodes both a C4 isoform and a nonchloroplast form (Glackin and Grula, 1990; Sheen, 1991).
These studies show that environmental cues and develop- mental programs that use positive and negative regulatory mechanisms control the accumulation of mRNAs coding for photosynthetic proteins. This regulation affects the timing of protein synthesis and its cellular localization, but it does not always directly control the quantities of proteins. Regulation of protein quantity also occurs at the stage of assembly of mul- timeric complexes in the chloroplast. For example, when synthesis of LSU is inhibited, unassembled SSU polypeptides are rapidly degraded so that stoichiometric amounts of both Rubisco subunits accumulate (Schmidt and Mishkind, 1983). The assembly process is poorly understood, especially its quantitative aspects. How the quantity of each enzyme is
determined as the C3 and C4 pathways are assembled during chloroplast development is unknown. Regulation of enzyme quantity is also important in the mature leaf cell, because part of the plant’s response to environmental changes can be to change absolute amounts of enzymes as well as their activities.
METABOLIC ENGINEERING OF PHOTOSYNTHESIS
The response of photosynthetic rate to environmental parameters has been well characterized for a wide range of plants (for review, see Woodrow and Berry, 1988). From the kinetic characteristics of Rubisco (Figure l), it has been possible to construct comprehensive models of photosynthe- sis that accurately predict the response of carbon fixation in C3 plants to particular ambient C02 concentrations (von Caemmerer and Farquhar, 1981). In the case of C3 plants in air and saturating light, it is now clear that the photosynthetic rate is largely, although not entirely, governed by the amount and kinetic characteristics of Rubisco. The remaining fractional control of photosynthetic rate is distributed among the other enzymes, and the degree of limitation by each step presum- ably varies with environmental conditions, such as light regimes, in a manner that is difficult to assess (Figure 2). This kind of regulation occursvia covalent modification and allosteric effects on enzyme activity and in the short term (that is, hours) does not involve changes in the amount of enzyme.
Figure 1 illustrates the complexity of regulation in the pho- tosynthetic system, showing potentially regulatory enzymes in both the C3 and C4 pathways. In C4 plants, determining which enzymes control flux is further complicated by the ad- ditional complexity of cell specialization. Superimposed upon the regulation of the PCR cycle enzymes is regulation of the enzymes involved in the C02 concentrating mechanism of the C4 pathway.
From in vitro studies, we know that the activities of a num- ber of enzymes in the PCR cycle are capable of responding to changes in light and could potentially limit photosynthetic flux. This response occurs either indirectly, via changes in the stromal pH and Mg2+ concentration, both of which increase on illumination, or directly, by reduction/oxidation of the en- zyme via the thioredoxin system, a signal transduction pathway responsive to the redox state of photosystem I (Figure 3; Buchanan, 1991). In the case of Rubisco, activation is medi- ated by a specific activating protein (Rubisco activase; Salvucci et al., 1985), which senses chloroplast energy status. Such a complex set of light-responsive regulatory mechanisms is necessary for two reasons. First, a crude on/off switch is re- quired to prevent “futile cycles” occurring between respiratory and catabolic processes and the photosynthetic pathway, which share the same biochemical intermediates. Second, the ma- jor environmental variable that plants experience is short-term change in light intensity (light flecks, for example, are com- mon in many closed canopy crops, forests, and grasslands). It appears that a multienzyme control mechanism has evolved

Regulation of Photosynthesis 801
O 100% 200% Wild
< TYPe antisense overexpression
EXTRACTABLE ENNME ACTlVlTY
Figure 2. The Response of Photosynthetic Carbon Fixation to Changes in Enzyme Activity.
Enzyme activity can be reduced to below wild-type levels by antisense suppression or elevated to above wild-type levels by overexpression. The effect on photosynthetic flux depends on whether the enzyme (a) is essentially “nonlimiting” over a range of enzyme activities, (b) has control over flux but ‘%o-limits” along with the activity of other enzymes, or (c) is classically “limiting.”
for conservation of metabolites within the PCR cycle and for intrinsic stability of the photosynthetic system during such transients, when the photosynthetic flux can change 10-fold or more in seconds (see Woodrow and Berry, 1988; Geiger and Servaites, 1994).
Although we have amassed a great deal of information on the regulatory properties of enzymes in vitro, evidence for their individual contributions to controlling photosynthetic flux in vivo is largely circumstantial. This is because it is often difficult to extrapolate regulatory properties and kinetic characteris- tics of enzymes in vitro to the cellular environment of the intact plant. Estimation of inhibitor/activator concentrations and sub- strate levels within cellular compartments in vivo and the effect of the high protein concentration present in cells can become insurmountable problems for such studies (see Ashton, 1982). Recombinant DNA technology and plant genetic transforma- tion have provided us with excellent tools to get around some of these obstacles. Using the techniques of gene suppression and overexpression, it is possible to alter the amount of a single
enzyme in a transgenic plant, thus generating a series of mu- tants with a range of enzyme activities from below to above wild-type levels. Depending on the importance of this enzyme in determining photosynthetic rate, the phenotypic effects of changes in the leve1 of expression may vary widely (see Knight and Gray, 1992). Figure 2 shows the expected response of pho- tosynthetic rate to varying enzyme levels in three hypothetical cases: (1) in which the enzyme is present in considerable ex- cess; (2) in which the enzyme is ‘%o-limiting:’ along with other enzymes in the pathway; and (3) in which the enzyme is
A Thloredoxln medlated enzyme activation
B Phosphorylatlon/dephosphorylatlon regulatlon of PPdK
c Phosphorylatlonldephosphorylatlon regulatlon of PEP carboxylase
Klnase
E -/T* E-P “iflactivated“ ATP “Activated (Mal sensitive) 4 (Mal insensitive)
Phosphatase Figure 3. Three Examples of Enzyme Regulation by Covalent Modification.
(A) Regulation by the thioredoxin system. An enzyme is activated when a disulfide bridge on the protein is reduced by the regulatory protein thioredoxin-m (ThR-m). Thioredoxin’s redox state is determined by the redox state of ferredoxin (Fd), the terminal electron acceptor of pho- tosystem I, and consequently is responsive to the rate of electron transport and light intensity. (E) Phosphorylation regulation of PPdK. PDRP, the PPdK regulatory protein, catalyzes both the dephosphorylation of inactive enzyme (E-P) to active enzyme (E) and the reverse reaction. The relative rates of these reactions are believed to respond to the energy status of the chloroplast. (C) Phosphorylation regulation of PEPC. In contrast to PPdK, PEPC is activated when phosphorylated by a protein kinase and becomes insensitive to the metabolic inhibitor malate. The “inactivated dephos- phorylated form still has activity but is sensitive to inhibition by malate.

802 The Plant Cell
“limiting” for photosynthetic flux. A quantitative assessment of an enzyme’s role in each class can be made using control analysis (Kacser and Burns, 1973). This mathematical treat- ment of biochemical regulation assigns a “control coefficient” between O and 1, with O indicating no control over flux by the enzyme and 1 indicating that the enzyme is limiting. Results from studies on diverse organisms indicate that control of flux through a pathway is frequentl9 shared between a number of steps.
Metabolic engineering can be used to control not only the quantity of enzyme present in a transgenic plant but also its “quality.” Both site-directed mutagenesis and expression of het- erologous enzymes allow the significance of the regulatory properties of an enzyme to be investigated in vivo. Removing the amino acids responsible for light regulation of an enzyme, for example, is a powerful tool in assessing the importance of this property to the plant. The following sections outline some of the approaches used to alter enzyme quantity and quality; a unifying theme throughout is the role of individual enzymes in controlling photosynthetic rate in vivo.
Antisense Suppression of Key Photosynthetic Enzymes
C3 Pathway
By far the most widely used approach to the genetic manipu- lation of photosynthesis to date has been the use of antisense RNA technology to produce transgenic plants with reduced levels of key photosynthetic enzymes. “Metabolic engineering” of photosynthesis was first reported by Rodermel et al. (1988), who transformed tobacco with a full-length antisense RNA con- struct targeted to the SSU of Rubisco, using a constitutive promoter. These transformants showed a substantial reduc- tion in RbcS transcript levels as well as in Rubisco protein levels and enzyme activity (Rodermel et al., 1988; Quick et al., 1991). However, Rubisco activity could be reduced by up to 40% before even a marginal effect on photosynthesis would be ob- served. In contrast, similar experiments by Hudson et al. (1992) resulted in a range of phenotypes in which both photosynthe- sis and growth were adversely affected. Careful analysis of these plants has revealed that when plants are grown at high light and at atmospheric COn concentrations, Rubisco activ- ity exerts a high degree of control over photosynthetic carbon flux and under these conditions can be considered a limiting enzyme (Hudson et al., 1992). The apparent inconsistencies between these observations appear to relate mostly to the light intensities used for growth of transformants, that is, environ- ment cabinet illumination in the former case (Quick et al., 1991) and full sunlight in the latter (Hudson et al., 1992). These transgenic plants have far wider uses than in the study of pho- tosynthetic flux. For example, because Rubisco is also the major protein present in leaves, it is extremely important in the nitrogen relations and nutritive value of higher plants, mak- ing these transformants valuable in studying nitrogen use and
allocation (Masle et al., 1993). In addition, these plants have also been used to study the relationship between photosynths sis and growth and regulation of stomatal aperture (Evans et al., 1994).
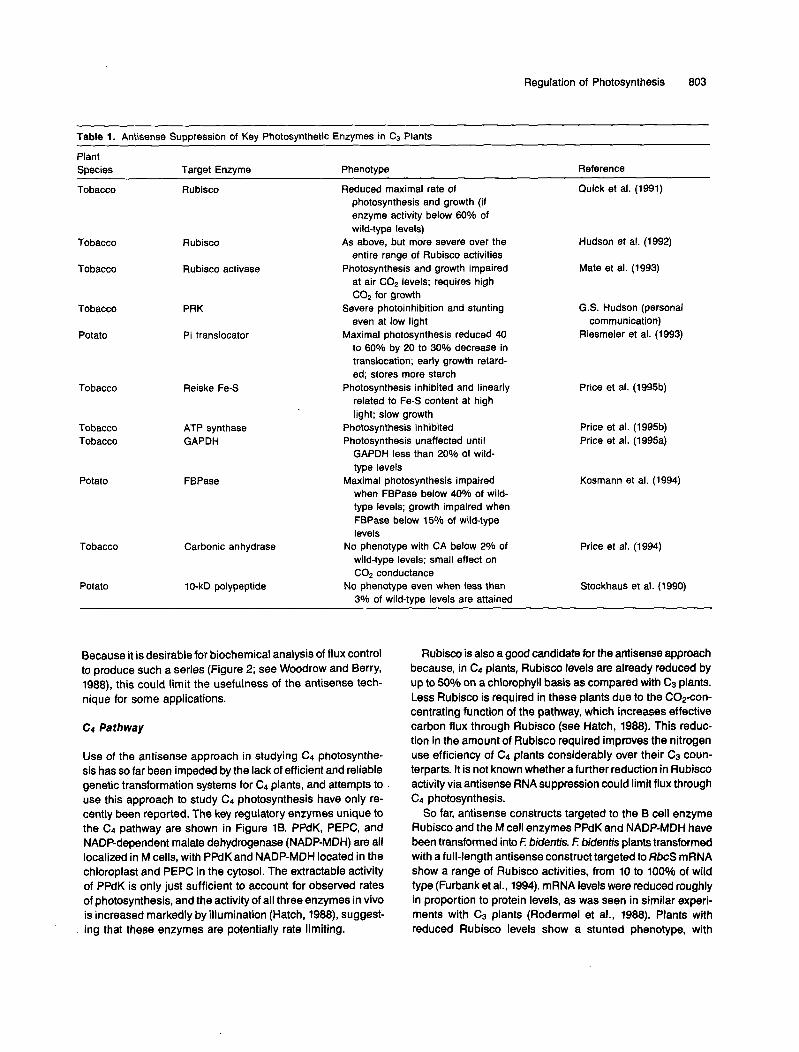
The list of photosynthetic enzymes whose expression has been reduced in C3 plants is steadily growing, as indicated in Table 1. In most cases, soluble stromal enzymes of pho- tosynthetic carbon metabolism have been targeted: Rubisco, Rubisco activase, fructose-1,Sbisphosphatase (FBPase), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), phos- phoribulokinase (PRK), and carbonic anhydrase. More recently, membrane proteins, such as translocators (chloroplast Pi trans- locator) and thylakoid membrane proteins (Reiske Fe-S center, ATPase, and the 10-kD polypeptide of photosystem I I (PSII]), have also been manipulated. As discussed earlier, the range of phenotypes observed in these experiments can vary enor- mously. Many enzymes, such as FBPase (Kosmann et al., 1994), GAPDH (Price et al., 1995a), and the Pi translocator (Riesmeier et ai., 1993), seem to fall into the intermediate or “co-limiting” category. For example, a 70% decrease in FBPase activity resulted in only a 20% decrease in maximum photosyn- thetic rate (Kosmann et al., 1994). For some enzymes, such as the thylakoid PSll 10-kD polypeptide (whose function is un- known; Stockhaus et al., 1990) and chloroplastic carbonic anhydrase (Price et al., 1994), little or no phenotype is evident when their levels are reduced by antisense suppression, sug- gesting that these proteins fall into the “nonlimiting” category.
The interpretation of the phenotypic effects of an antisense construct on photosynthesis and growth of the transformed plants is not always straightforward. The importance of growth conditions in influencing phenotype should not be underesti- mated, as demonstrated by the effect of light intensity during growth on the interpretation of the Rubisco antisense experi- ments described previously. It also appears that in many instances, the relationship between steady state mRNA lev- els and protein levels in transgenic plants expressing antisense RNA may not be simple. Attempting to reduce levels of the 23- and 33-kD polypeptides of the PSll oxygen-evolving complex and the Reiske Fe-S center of the cytochrome bdf complex, Palomares et ai. (1993) recently reported up to a 90% reduction in target mRNA levels in transgenic plants without a discernible effect on protein levels or phenotype. In contrast, Price et al. (1995b) produced slow-growing transgenic tobacco with reduced Reiske Fe-S and electron transport rates; how- ever, a 93 to 94% reduction in message level gave only a 60 to 86% reduction in protein. The phenotype was often unsta- ble and could be ameliorated by high growth irradiance, suggesting that low mRNA levels are not limiting under some conditions or that the endogenous sense transcript can “swamp” out the antisense mRNA at high irradiance. Similar nonlinear relationships of protein level to transcript level have also been observed in plants transformed with an antisense gene targeted to the chloroplast envelope Pi translocator (Riesmeier et al., 1993). If this proves to be a widespread phenomenon, it may be difficult in some cases to produce a series of mutants with a wide range of enzyme activities.
Regulation of Photosynthesis 803
Table 1. Antisense Suppression of Key Photosynthetic Enzymes in C3 Plants
Plant Species Target Enzyme Phenotype Reference
Tobacco Rubisco Reduced maximal rate of Quick et al. (1991) photosynthesis and growth (if enzyme activity below 60% of wild-type levels)
entire range of Rubisco activities
at air C o e levels; requires high COp for growth
even at low light communication)
to 60% by 20 to 30% decrease in translocation; early growth retard- ed; stores more starch
related to Fe-S content at high light; slow growth
Tobacco Rubisco As above, but more severe over the Hudson et al. (1992)
Tobacco Rubisco activase Photosynthesis and growth impaired Mate et al. (1993)
Tobacco PRK Severe photoinhibition and stunting G.S. Hudson (personal
Potato Pi translocator Maximal photosynthesis reduced 40 Riesmeier et al. (1993)
Tobacco Reiske Fe-S Photosynthesis inhibited and linearly Price et al. (1995b)
Tobacco ATP synthase Photosynthesis inhibited Price et al. (1995b) Tobacco GAPDH Photosynthesis unaffected until Price et al. (1995a)
GAPDH less than 20% of wild- type levels
Potato FBPase Maximal photosynthesis impaired Kosmann et al. (1994) when FBPase below 40% of wild- type levels; growth impaired when FBPase below 15% of wild-type levels
No phenotype with CA below 2% of wild-type levels; small effect on COp conductance
3% of wild-type levels are attained
Carbonic anhydrase Price et al. (1994)
Potato 10-kD polypeptide No phenotype even when less than Stockhaus et al. (1990)
Tobacco
Because it is desirable for biochemical analysis of flux control to produce such a series (Figure 2; see Woodrow and Berry, 1988), this could limit the usefulness of the antisense tech- nique for some applications.
C4 Pathway
Use of the antisense approach in studying C4 photosynthe- sis has so far been impeded by the lack of efficient and reliable genetic transformation systems for Cq plants, and attempts to use this approach to study C4 photosynthesis have only re- cently been reported. The key regulatory enzymes unique to the C4 pathway are shown in Figure 1B. PPdK, PEPC, and NADP-dependent malate dehydrogenase (NADP-MDH) are all localized in M cells, with PPdK and NADP-MDH located in the chloroplast and PEPC in the cytosol. The extractable activity of PPdK is only just sufficient to account for observed rates of photosynthesis, and the activity of all three enzymes in vivo is increased markedly by illumination (Hatch, 1988), suggest- ing that these enzymes are potentially rate limiting.
Rubisco is also a good candidate for the antisense approach because, in C4 plants, Rubisco levels are already reduced by up to 50% on a chlorophyll basis as compared with C3 plants. Less Rubisco is required in these plants due to the C02-con- centrating function of the pathway, which increases effective carbon flux through Rubisco (see Hatch, 1988). This reduc- tion in the amount of Rubisco required improves the nitrogen use efficiency of C4 plants considerably over their C3 coun- terparts. It is not known whether a further reduction in Rubisco activity via antisense RNA suppression could limit flux through C4 photosynthesis.
So far, antisense constructs targeted to the B cell enzyme Rubisco and the M cell enzymes PPdK and NADP-MDH have been transformed into E bidentis. F: bidentis plants transformed with a full-length antisense construct targeted to RbcS mRNA show a range of Rubisco activities, from 10 to 1000/0 of wild type (Furbank et al., 1994). mRNA levels were reduced roughly in proportion to protein levels, as was seen in similar experi- ments with C3 plants (Rodermel et al., 1988). Plants with reduced Rubisco levels show a stunted phenotype, with

804 The Plant Cell
proportionally reduced photosynthetic capacity at high light and over a range of C02 concentrations. Interestingly, there appears to be a strong regulatory mechanism linking Rubisco activity to the rate of the mesophyll C4 cycle. Although the lev- els of mesophyll enzymes were unaffected in the transformants, down-regulation of the mesophyll C4 cycle turnover was ob- served in intact leaves, presumably via down-regulation of enzyme activity. The biochemical basis for such a mechanism is currently unknown, and further physiological and biochem- ical analysis of the progeny of these transformants is under way. These plants will be a valuable tool for understanding photosynthetic regulation in C4 plants because, unlike the CB case, the role of Rubisco activity in determining photosynthetic rate in C4 plants under varying environmental conditions has not been extensively modeled.
Analysis of F: bidentis transformants containing other pho- tosynthetic antisense constructs is still in its early stages. NADP-MDH has been reduced by as much as 60°/o from wild- type levels (S.J. Trevanion, R.T. Furbank, and A.R. Ashton, unpublished observations); in these plants, there is a commen- surate reduction in the light-saturated rate of photosynthesis but only a slight effect on growth.
In the case of PPdK, antisense transformants show up to a 90% reduction in both enzyme activity and transcript levels (Furbank et al., 1994). Transformants containing a full-length antisense construct fall into two groups: in one, enzyme ac- tivities range from 10 to 20% of wild type; in the other, enzyme activities are normal. No transformants showed an intermedi- ate level of PPdK, although some plants transformed with an 0.8-kb antisense construct corresponding to the 5’ end of the Wk cDNA show a reduction in Wk transcript levels but no reduction in extractable enzyme activity. The phenotype of plants with 20% or less PPdK activity is severe. Regenerants are incapable of photo-autotrophic growth in air and can be maintained only on exogenous sucrose or in an atmosphere containing lO/o COn. Plants deprived of exogenous sucrose show rapid and severe photoinhibition in air, even at very low light intensity. One explanation for the all-or-none nature of the phenotype in these experiments could be that the rela- tionship between transcript level and protein level is nonlinear or possibly even sigmoidal. Thus, when the transcript level drops below a critical threshold, a catastrophic falloff in pro- tein levels may occur.
Site-Directed Mutagenesis and “Overexpression”
High-leve1 expression of a native or heterologous protein to increase flux through a pathway, divert flux, or change the regulatory properties of an enzyme is a powerful tool for un- derstanding regulation of photosynthesis. So far, these techniques have been more commonly applied to the path- ways of sucrose and starch biosynthesis than to photosynthesis per se (Sonnewald et al., 1994; Stitt, 1994). A classic example
I
of this approach is the expression of a bacterial ADP-glucose pyrophosphorylase in plants to “short circuit” regulation of this step and increase starch accumulation (see Martin and Smith, 1995, this issue). Site-directed mutagenesis to alter the kinetic properties of Rubisco has been moderately successful in vitro; however, the heteropolymeric nature of the protein and the fact that LSU is chloroplast encoded have prevented expression of the mutated holoenzyme in vivo. It is also generally accepted that overexpression of genes in plants is more difficult than antisense suppression due to cosuppression (Napoli et al., 1990; van der Krol et ai., 1990) and the possible inactivation of the overexpressed protein by endogenous regulatory sys- tems (for example, Sonnewald et al., 1994).
In the past two decades, it has become apparent that a ma- jor form of enzyme regulation involves covalent modification of specific amino acid residues either by the formation of di- sulfide bridges or by protein kinase-mediated phosphorylation (Figure 3). This type of regulation in plants responds directly to light (via the thioredoxin system) or to cellular energy reserves such as ATI? With sequences of more photosynthetic enzymes appearing in genetic databases almost daily, this area of en- zyme regulation is particularly amenable to study by expression of recombinant protein both in vitro and in vivo. For example, NADP-MDH is a light-activated enzyme in the C4 pathway (Figure 1) that undergoes reductive activation in the light via photosynthetic electron transfer and the thioredoxin system (Buchanan, 1991). Reduction of a disulfide bridge between two cysteine residues in NADP-MDH by reduced thioredoxin activates the enzyme (Figure 3). NADP-MDH protein is homol- ogous to the NAD-dependent nonphotosynthetic form of the enzyme, but it also has C- and N-terminal extensions that have been implicated in conferring the unique regulatory properties of the photosynthetic enzyme. Using site-directed mutagenesis, the pairs of cysteine residues responsible for redox activation at both the N and C termini of the sorghum enzyme have been identified (see lssakidis et al., 1994). Mu- tant forms of this enzyme that are not inactivated by oxidation and thus should not be inactivated in the dark have been pro- duced. Flaveria and tobacco have been transformed with these constructs to examine the role of light activation of this enzyme in vivo (S.J. Trevanion, A.R. Ashton, and I. Issakidis, personal communication).
Two other key enzymes in C4 photosynthesis are regulated by covalent modification: PPdK and PEPC (Figures 1 and 3). PEPC, the primary CO,-fixing enzyme of the C4 pathway, is “activated in vivo when phosphorylated by a specific protein kinase (for review, see Budde and Chollet, 1988). This activa- tion does not result in an increase in the V,, of the enzyme but manifests itself by a decrease in sensitivity to the inhibitor malate. In the sorghum protein, phosphorylation of a serine residue is responsible for this activation. Substitution of this serine in a recombinant enzyme expressed in Escherichia coli prevents this activation (Duff et ai., 1993). This region of the amino acid sequence appears to be highly conserved among PEPC enzymes from many sources, suggesting that this

Regulation of Photosynthesis 805
regulatory mechanism may be universal. So far, no transfor- mation experiments have been performed with a recombinant enzyme in higher plants; therefore, it is not possible to assess the importance of this regulatory mechanism in vivo. The sig- na1 transduction chain that controls the activity of the protein serine kinase in response to environmental conditions remains somewhat of a mystery, as does the identity of the kinase itself.
PPdK, which is responsible for regenerating PEP, the ac- ceptor for COp fixation in the mesophyll chloroplast, has long been recognized as a light-activated enzyme (Figure 1; reviewed in Hatch, 1988). It is now known that, in contrast to PEPC, this enzyme is inactivated by phosphorylation of a threo- nine residue (Ashton and Hatch, 1983; Ashton et al., 1984). This residue is not the site that is phosphorylated during catal- ysis, which is a nearby histidine residue. The enzyme that catalyzes regulatory phosphorylation of PPdK, the PPdK regulatory protein, is unusual in that it uses ADP rather than ATP as the phosphate donor. In addition, this same enzyme catalyzes the removal of the phosphate group, reactivating PPdK by phosphorolysis rather than by hydrolysis (Figure 3; Ashton et ai., 1984). It is not clear how the activity of the PPdK regulatory protein itself is controlled, although high pyruvate levels appear to block inactivation (Burnell et ai., 1986). The regulatory protein is not abundant in leaves, and purification to homogeneity has proven elusive (Smith et al., 1994), as has the isolation of its gene.
The gene coding for PPdK has now been cloned from a num- ber of species (see previous discussion). Preliminary work has been done expressing the native enzyme from maize in E. coh (A.R. Ashton and R.T. Furbank, unpublished observations) and from representatives of the Haveria genus (J.N. Burnell, per- sonal communication). By mutagenesis of the regulatory threonine and expression of the recombinant protein in E. Coli and higher plants, it should be possible to examine the regula- tory significance of phosphorylation.
One of the greatest challenges in the area of photosynthe- sis is to use the large body of information available in the literature on the enzymes of the pathway and their regulation to understand and perhaps improve photosynthetic perfor- mance in whole plants. As discussed earlier, the major barrier to this research has been the difficulty of performing biochem- ical manipulations in vivo in a precise and interpretable fashion. Early indications suggest that metabolic engineering, coupled with more traditional biochemical and physiological ap- proaches, may provide the means to achieve this aim.
ACKNOWLEDGMENTS
We thank our colleagues who communicated work in progress and apologize to those whose contributions to the field were omitted due to space limitations. We are grateful to Tony Ashton for helpful com- ments on the manuscript.
REFERENCES
Ashton, A.R. (1982). A role for ribulose-l,5-bisphosphate carboxylase as a metabolite buffer. FEBS Lett. 145, 1-7.
Ashton, A.R., and Hatch, M.D. (1983). Regulation of C4 photosynthe- sis: Regulation of pyruvate, Pi dikinase by ADP-dependent phosphorylation and dephosphorylation. Biochem. Biophys. Res. Commun. 115, 53-60.
Ashton, A.R., Burnell, J.N., and Hatch, M.D. (1984). Regulation of C, photosynthesis: lnactivation of pyruvate, Pi dikinase by ADP- dependent phosphorylation and activation by phosphorolysis. Arch. Biochem. Biophys. 230, 492-503.
Bansal, K.C., and Bogorad, L. (1993). Cell type-preferred expres- sion of maize cab-m: Repression in bundle sheath cells and enhancement in mesophyll cells. Proc. Natl. Acad. Sci. USA 90,
Buchanan, 6.6. (1991). Regulation of COn assimilation in oxygenic photosynthesis: The ferredoxinhhioredoxin system. Arch. Biochem. Biophys. 288, 1-8.
Budde, R.J.A., and Chollet, R. (1988). Regulation of enzyme activity in plants by reversible phosphorylation. Physiol. Plant. 72,435-439.
Burnell, J.N., Jenklns, C.L.D., and Hatch, M.D. (1986). Regulation of C4 photosynthesis: A role for pyruvate in regulating pyruvate, Pi dikinase activity in vivo. Aust. J. Plant Physiol. 13, 203-210.
Chitty, J.A., Furbank, R.T., Marshall, J.S., Chen, Z., andTaylor, W.C. (1994). Genetic transformation of the C, plant, flavefia bidentis. Plant J. 6, 949-956.
Deng, X.-W. (1994). Fresh view of light signal transduction in plants. Cell 76, 423-426.
Duff, S.M.G., Leplniec, L., Cretln, C., Andreo, C., Condon, S.A., Sarath, G., Vldal, J., Gadal, P., and Chollet, R. (1993). An en- gineered change in the L-malate sensitivity of a site directed mutant of sorghum phosphoenolpyruvate carboxylase: The effect of sequen- tia1 mutagenesis and S-carboxymethylation at position 8. Arch. Biochem. Biophys. 306, 272-276.
Eckes, P., Schell, J., and Willmitzer, L. (1985). Organ-specific ex- pression of three leafktem specific cDNAs from potato is regulated by light and correlated with chloroplast development. MOI. Gen. Ge- net. 199, 216-221.
Evans, J.R., von Caemmerer, S., Setchell, B.A., and Hudson, G.S. (1994). The relationship between COn transfer conductance and leaf anatomy in transgenic tobacco with a reduced content of Rubisco. Aust. J. Plant Physiol. 21, 475-495.
Furbank, R.T., Chltty, J.A., Taylor, W.C., and Jenkins, C.L.D. (1994). Genetic manipulation of C4 photosynthesis: Antisense of key photc- synthetic enzymes in Flaveria bidentis. Plant Physiol. 105, 23.
Gelger, D.R., and Servaites, J.C. (1994). Dynamics of self-regulation of photosynthetic carbon metabolism. Plant Physiol. Biochem. 32,
Gilmartin, P.M., Sarokin, L., Memellnk, J., and Chua, N.-H. (1990). Molecular light switches for plant genes. Plant Cell 2, 369-378.
Giuliano, G., Hoffman, N.E., Ko, K., Scolnik, P.A., andcashmore, A.R. (1988). A light-entrained circadian clock controls transcription of severa1 plant genes. EMBO J. 7, 3635-3642.
Glackin, C.A., and Grula, J.W. (1990). Organ-specific transcripts of different size and abundance derive from the same pyruvate
4057-4061.
173-183.

806 The Plant Cell
orthophosphate dikinase gene in maize. Proc. Natl. Acad. Sci. USA
Hatch, M.D. (1988). C4 photosynthesis: A unique blend of modified biochemistry, anatomy and ultrastructure. Biochim. Biophys. Acta
Hermans, J., and Westhoff, P. (1990). Analysis of expression and evolu- tionary relationships of phosphoenolpyruvate carboxylase genes in Flaveria trinervia (C,) and I? pringlei (C3). MOI. Gen. Genet. 224,
Hudson, G.S., Evans, J.R., von Caemmerer, S., Anrldsson, Y.B.C., and Andrews, T.J. (1992). Reduction of ribulose-1.5-bisphosphate carboxylaseloxygenase content by antisense RNA reduces pho- tosynthesis in transgenic tobacco plants. Plant Physiol. 98,294-302.
Issakidis, E., Saarlnen, M., Decottlgnles, P., Jacquot, J.-P., Cretin, C, Gadal, P., and Mlglnlac-Maslow, M. (1994). ldentification and characterization of the second regulatory disulphide bridge of recom- binant sorghum leaf NADP-malate dehydrogenase. J. Biol. Chem.
Kacser, H., and Burns, J.A. (1973). The control of flux. Symp. SOC. Exp. Biol. 27, 65-104.
Knight, J.S., and Gray, J.C. (1992). Transgenic plants as a tool for the study of photosynthetic carbon assimilation. In Trends in Pho- tosynthesis Research, J. Barber, M.G. Guerrero, and H. Medrano, eds (Andover, MA: Intercept), pp. 267-277.
Kosmann, J., Sonnewald, U., and Wlllmltzer, L. (1994). Reduction of the chloroplast fructose-1 ,I-bisphosphatase in transgenic potato plants impairs photosynthesis and plant growth. Plant J. 6,637-650.
Martln, C., and Smlth, A.M. (1995). Starch biosynthesis. Plant Cell 7, 971-985.
Masle, J., Hudson, G.S., and Badger, M.R. (1993). Effects of am- bient COp concentration on growth and nitrogen use in tobacco (Nicotiana tabacum) plants transformed with an antisense gene to the small subunit of ribulose-l,5-bisphosphate carboxylaseloxygen- ase. Plant Physiol. 103, 1075-1088.
Mate, C.J., Hudson, G.S., von Caemmerer, S., Evans, J.R., and Andrews, T.J. (1993). Reduction of ribulose bisphosphate carboxy- lase activase levels in tobacco (Nicotiana tabacum) by antisense RNA reduces ribulose bisphosphate carboxylase carbamylation and impairs photosynthesis. Plant Physiol. 102, 1119-1128.
Matsuoka, M., Tada, Y., Fujlmura, T., and Kano-Murakami, Y. (1993). Tissue-specific light-regulated expression directed by the promoter of a C4 gene, maize pyruvate, orthophosphate dikinase, in a C3 plant, rice. Proc. Natl. Acad. Sci. USA 90, 9586-9590.
Matsuoka, M., Kyozuka, J., Shlmamoto, K., and Kano-Murakami, Y. (1994). The promoters of two carboxylases in a C4 plant (maize) direct cell-specific, light-regulated expression in a C3 plant (rice). Plant J. 6, 311-319.
Muller, M., Viro, M., Balke, C., and Kloppstech, K. (1980). Poly- adenylated mRNA for the light-hawesting chlorophyll alb protein: Its presence in green and absence in chloroplast-free plant cells. Planta 148, 444-447.
67, 3004-3008.
895, 81-106.
459-468.
269, 3511-3517.
Nagy, F., Kay, S.A., and Chua, N.-H. (1988). A circadian clock regu- lates transcription of the wheat Ceb-7 gene. Genes Dev. 2,376-382.
Napoli, C., Lemleux, C., and Jorgensen, R. (1990). lntroduction of a chimeric chalcone synthase gene into petunia results in revers- ible co-suppression of homologous genes in trens. Plant Cell 2, 279-289.
Nelson, T., and Langdale, J.A. (1989). Patterns of leaf development in C4 plants. Plant Cell 1, 3-13.
Nelson, T., and Langdale, J.A. (1992). Developmental genetics of C, photosynthesis. Annu. Rev. Plant Physiol. Plant MOI. Biol. 43,2547.
Oelmuller, R. (1989). Photooxidative destruction of chloroplasts and its effect on nuclear gene expression and extraplastidic enzyme lev- els. Photochem. Photobiol. 49, 229-239.
Palomares, R., Herrman, R.G., and Oelmuller, R. (1993). Antisense RNA for components associated with the oxygen evolving complex and the Rieske ironlsulfur protein of the tobacco thylakoid mem- brane suppresses accumulation of mRNA but not protein. Planta
Prlce, G.D., von Caemmerer, S., Evans, J.R., Yu, J.-W., Lloyd, J., Oja, V., Kell, P., Harrlson, K., Gallagher, A., and Badger, M.R. (1994). Specific reduction of the chloroplast carbonic anhydrase ac- tivity by antisense RNA in transgenic tobacco plants has a minor effect on photosynthetic COp assimilation. Planta 193, 331-340.
Prlce, G.D., Evans, J.R., von Caemmerer, S., RI, J.-W., and Badger, M.R. (1995a). Specific reduction of chloroplast glyceraldehyde-3- phosphate dehydrogenase activity by antisense RNA reduces C02 assimilation via a reduction in RuBP regeneration in transgenic tobacco plants. Planta 195, 369-378.
Prlce, G.D., Yu, J.-W., von Caemmerer, S., Evans, J.R., Chow, W.S., Anderson, J.M., Hurry, V., and Badger, M.R. (1995b). Studying the central roles of the chloroplast cytochrome B6/f and the ATP synthase complexes in transgenic tobacco: Transformation with an- tisense RNAdirected against the Reiske FeS and the ATPd nuclear encoded polypeptides. Aust. J. Plant Physiol., in press.
Quick, W.P., Schurr, U., Schelbe, R., Schultze, E.-D., Rodermel, S.R., Bogorad, L., and Stltt, M. (1991). Decreased ribulose-1s-bis- phosphate carboxylaseoxygenase in transgenic tobacco transformed with “antisense” rbcS. I. lmpact on photosynthesis in ambient growth conditions. Planta 183, 542-554.
Riesmeier, J.W., Flugge, U.4, Schulz, B., Heineke, D., Heldt, H.W., Wlllmltzer, L., and Frommer, W.B. (1993). Antisense repression of the chloroplast triose phosphate translocator affects carbon par- titioning in transgenic potato plants. Proc. Natl. Acad. Sci. USA 90,
Rodermel, S.R., Abbott, M.S., and Bogorad, L. (1988). Nuclear- organelle interactions: Nuclear antisense gene inhibits ribulose bisphosphate carboxylase enzyme levels in transformed tobacco plants. Cell 55, 673-684.
Salvuccl, M.E., Portis, A.R., Jr., and Ogren, W.L. (1985). Asoluble chloroplast protein catalyzes ribulose-bisphosphate carboxylasel oxygenase activation in vivo. Photosynthesis Res. 7, 193-201.
Schaffner, A.R., and Sheen, J. (1992). Maize C4 photosynthesis in- volves differential regulation of phosphoenolpyruvate carboxylase genes. Plant J. 2, 221-232.
Schmidt, G.W., and Mlshklnd, M.L. (1983). Rapid degradation of un- assembled ribulose-l,5-bisphosphate carboxylase small subunits in chloroplasts. Proc. Natl. Acad. Sci. USA 80, 2632-2636.
Sheen, J. (1991). Molecular mechanisms underlying the differential expression of maize pyruvate, orthophosphate dikinase genes. Plant Cell 3, 225-245.
Sheen, JrY, and Bogorad, L. (1985). Differential expression of the ribulose bisphosphate carboxylase large subunit gene in bundle sheath and mesophyll cells of developing maize leaves is influenced by light. Plant Physiol. 79, 1072-1076.
190, 305-312.
6160-6164.

Regulation of Photosynthesis 807
Smlth, C.M., Duff, S.M.G., and Chollet, R. (1994). Partia1 purifica- tion and characterization of maize-leaf pyruvate, Pi dikinase regulatory protein: A low abundance mesophyll chloroplast stromal protein. Arch. Biochem. Biophys. 308, 200-206.
Sonnewald, U., Lerchl, J., Zrenner, I?., and Frommer, W. (1994). Manipulation of sink-source relations in transgenic plants. Plant Cell Environ. 17, 649-658.
Stltt, M. (1994). Manipulation of carbohydrate partitioning. Curr. Opin. Biotechnol. 5, 137-143.
Stockhaus, J., Hofer, M., Renger, G., Westoff, P., Wydrzynski, T., and Willmltzer, L. (1990). Anti-sense RNA efficiently inhibits for- mation of the 10 kd polypeptide of photosystem II in transgenic potato plants: Analysis of the role of the 10 kd protein. EM60 J. 9,3013-3021.
Stockhaus, J., Poetsch, W,, Stelnmuller, K., and Westhoff, P. (1995). Evolution of the C., phosphoenolpyruvate carboxylase promoter of the C4 dicot F/averia frinervia: An expression analysis in the C3 plant tobacco. MOI. Gen. Genet. 245, 286-293.
Taylor, W.C. (1989a). Transcriptional regulation by a circadian rhythm. Plant Cell 1, 259-264.
Taylor, W.C. (1989b). Regulatory interactions between nuclear and plastid genomes. Annu. Rev. Plant Physiol. Plant MOI. Biol. 40, 21 1-233.
Thompson, W.F., and Whlte, M.J. (1991). Physiological and molecu- lar studies of light-regulated nuclear genes in higher plants. Annu. Rev. Plant Physiol. Plant MOI. Biol. 42, 423-466.
Tobln, E.M., and Sllverthorne, J. (1985). Light regulation of gene ex- pression in higher plants. Annu. Rev. Plant Physiol. 36, 569-593.
van der Krol, A.R., Yur, L.A., Beld, M., MOI, J.N.M., and Stultje, A.R. (1990). Flavonoid genes in petunia: Addition of a limited num- ber of gene copies may lead to a suppression of gene expression. Plant Cell 2, 291-299.
von Caemmerer, S., and Faquhar, G.D. (1981). Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 153, 376-387.
Wang, J.-L., Klesslg, D.F., and Berry, J.O. (1992). Regulation of C4 gene expression in developing amaranth leaves. Plant Cell 4,
Wang, J.-L., Long, J.J., Hotchklss, T., and Berry, J.O. (1993). C4 photosynthetic gene expression in light- and dark-grown amaranth cotyledons. Plant Physiol. 102, 1085-1093.
Woodrow, I.E., and Berry, J.A. (1988). Enzymatic regulation of pho- tosynthetic COp fixation in C3 plants. Annu. Rev. Plant Physiol. Plant MOI. Biol. 39, 533-594.
173-184.

DOI 10.1105/tpc.7.7.797 1995;7;797-807Plant Cell
R. T. Furbank and W. C. TaylorRegulation of Photosynthesis in C3 and C4 Plants: A Molecular Approach.
This information is current as of May 25, 2018
Permissions 8X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw153229
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists





























