Embed Size (px)
Citation preview

Cellular Signalling 24 (2012) 1821–1830
Contents lists available at SciVerse ScienceDirect
Cellular Signalling
j ourna l homepage: www.e lsev ie r .com/ locate /ce l l s ig
Regulation of SLC26A3 activity by NHERF4 PDZ-mediated interaction
Ji Hyun Lee a, Joo Hyun Nam b, Joonhee Park a,c, Dong-Won Kang d, Joo Young Kim a,Min Goo Lee a, Jae Seok Yoon d,⁎a Department of Pharmacology and Brain Korea 21 Project for Medical Science, Yonsei University College of Medicine, Seoul 120‐752, Republic of Koreab Department of Physiology, Dongguk University College of Medicine, Kyungju 780‐714, Republic of Koreac Center for Cell Signaling & Drug Discovery Research, Division of Life and Pharmaceutical Sciences and College of Pharmacy, Ewha Womans University, Seoul 120‐750, Republic of Koread Department of Pharmacology, Kwandong University College of Medicine, Gangneung 210‐701, Republic of Korea
Abbreviations: SLC26, solute-linked carrier 26; NHEtory factor; STAS, sulphate transporters and anti-sigmagenital chloride diarrhoea; PDZ, PDS-95/Discs largtransferase; Y2H, yeast two-hybrid; FRAP, fluorescenceNHE3, sodium/proton exchanger 3; CFTR, cystic fibroprotein.⁎ Corresponding author at: Department of Pharmacolog
of Medicine, 522 Naegok-Dong, Gangneung 210‐701, Rep7494; fax: +82 33 641 1074.
E-mail address: [email protected] (J.S. Yoon).
0898-6568/$ – see front matter © 2012 Elsevier Inc. Alldoi:10.1016/j.cellsig.2012.05.010
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 January 2012Received in revised form 11 May 2012Accepted 13 May 2012Available online 22 May 2012
Keywords:NHERF4SLC26A3PDZ-based protein–protein interactionsChloride/bicarbonate exchangePhosphorylation
SLC26A3 functions as a chloride/bicarbonate anion exchanger expressed in the secretory epithelial cells in theintestine, pancreas, and salivary glands. SLC26A3 has a C-terminal class I PDZ binding motif that assembles regu-latory factors or other transporters by anchoring to various PDZ scaffold proteins. NHERF4 is an epithelial-enriched PDZ domain scaffold protein that has attracted attention because of its enriched tissue expression inthe intestine and kidney. In this study, we identified SLC26A3 as a novel binding transporter of NHERF4. We in-vestigated the functional role of NHERF4 in the regulation of SLC26A3 by using integrated biochemicaland physiological approaches. A direct protein–protein interaction was identified between the PDZ-binding motif of SLC26A3 and the third PDZ domain of NHERF4. Interaction with NHERF4 decreased the levelof SLC26A3 expression on the plasma membrane, which led to reduced SLC26A3 anion exchange activity. Nota-bly, interaction with NHERF4 induced rapid internalisation of SLC26A3 from the plasma membrane. TheSLC26A3–NHERF4 interaction was modulated by phosphorylation; serine 329 of NHERF4-PDZ3 played a criticalrole in modulating binding selectivity. Our findings suggest that NHERF4 is a novel modulator of luminal fluidityin the intestine by adjusting SLC26A3 expression and activity through a phosphorylation-dependentmechanism.
© 2012 Elsevier Inc. All rights reserved.
1. Introduction
Physiological control of luminal fluidity in the intestine is critical forhomeostasis and overall health [1]. When the SLC26A3 gene was firstdiscovered in 1993, it was considered a tumour suppressor gene incolon cancer, because it was significantly down-regulated in colonic ad-enomas [2,3]. It is nowwell known that SLC26A3 functions as a chloride/bicarbonate (Cl−/HCO3
−) anion exchanger and is amajormediator of lu-minal chloride reuptake in the apical membrane of secretory epithelialcells [3–6]. The highly conserved proximal part of the intracellular tailof SLC26 proteins, which is named STAS, is an important region for prop-er transport activity. Malfunction of SLC26A3 caused by various types ofmutations in the STAS region is responsible for CLD [7,8]. Furthermore,
RF, Na+/H+ exchanger regula--factor antagonists; CLD, con-e/ZO-1; GST, glutathione-S-recovery after photobleaching;sis transmembrane regulator
y, KwandongUniversity Collegeublic of Korea. Tel.: +82 33 649
rights reserved.
several SLC26 proteins, including SLC26A3, contain a C-terminal class IPDZ-binding motif [4]. Although it is known that the PDZ-bindingmotif itself is not essential for transport activity, it plays a role in the as-sembly of regulatory factors or other transporters by anchoring to vari-ous PDZ scaffold proteins [6,9,10].
The NHERF family of epithelial-enriched PDZ domain scaffold pro-teins are key modulators of epithelial cell functions. NHERF proteinsare located near the apical regions of epithelial cells and interact withclass I PDZ-binding motifs (‐X-S/T-X-Φ). NHERF3 and NHERF4 havefour PDZ domains, while NHERF1 and NHERF2 have two PDZ domainsat their N-terminus and an ERM-binding domain at their C-terminus[11–13]. Of the four NHERF family members, NHERF4 was identifiedmost recently by Y2H screening of a human intestinal cDNA library byusing the C-terminus of guanylyl cyclase C [14]. NHERF4 has overall se-quence and structural homologywith other NHERF proteins; NHERF3 isthe most closely related. Investigation of the potential roles of NHERF4has recently attracted attention because of its enriched tissue expres-sion in the intestine and kidney. NHERF4 is also termed the intestinal-and kidney-enriched PDZ protein (IKEPP) [12]. Previous studies haveshown that NHERF4 interacts with several membrane transporters, in-cluding multi-drug resistance-related protein 2 (MRP2), transient re-ceptor potential cation channel (TRPV) 5, TRPV6, organic cationtransporter 2 (OCTN2), and type IIa sodium/phosphate co-transporter

1822 J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
(Na/Pi IIa) [15–19], but its regulatory mechanism has not yet beenelucidated.
In the present study, we revealed that the gastric epithelial SLC26A3anion transporter is a novel binding partner of NHERF4. The physiologi-cal significance of SLC26A3 interacting with NHERF4 and the regulatorymechanisms of the NHERF4 interaction were investigated. Our findingssuggest that NHERF4 is a novel modulator of luminal fluidity in the in-testine by adjusting SLC26A3 expression and activity.
2. Materials and methods
2.1. Materials, solution, and cells
Standard HEPES-buffered solution A contained 145 mMNaCl, 5 mMKCl, 1 mM MgCl2, 1 mM CaCl2, 10 mM D-glucose, and 10 mM HEPES(pH 7.4 with NaOH). HCO3
−-buffered solution B contained 120 mMNaCl, 5 mM KCl, 1 mM MgCl2, 1 mM CaCl2, 10 mM D-glucose, 5 mMHEPES, and 25 mM NaHCO3 (pH 7.4 with NaOH). Chloride (Cl−)-freesolutions were prepared by replacing Cl− with gluconate. HeLa cellswere maintained in Dulbecco's modified Eagle's medium (DMEM) sup-plementedwith 10% foetal bovine serum, and HT-29 cells (KCLB 30038;KoreanCell Line Bank, Seoul, Korea)weremaintained in RPMI1640 con-taining 10% foetal bovine serum. The fluorescent pH probe, 2′,7′-bis(2-carboxyethyl)-5(6)-carboxyfluorescein (BCECF), was purchased fromMolecular Probes (Eugene, OR). Primary antibodies used for immuno-staining and immunoblotting were as follows: anti-Myc monoclonaland polyclonal antibodies, anti-HA monoclonal and polyclonal anti-bodies, anti-SLC26A3 (C-20) antibody (Santa Cruz Biotechnology, SantaCruz, CA), anti-NHERF4 antibody (Novus Biologicals, CO), and anti-Hisantibody (Calbiochem,Darmstadt, Germany). Fluorescein isothiocyanate(FITC)- and rhodamine-conjugated secondary antibodies were fromZymed (Carlsbad, CA). All other general chemicals including the small in-terfering RNA (siRNA) of NHERF4were purchased from Sigma (St. Louis,MO).
2.2. Plasmid vectors
The coding region of SLC26A3 cDNA was amplified by PCR from acDNA library of the human colon cell line, T84, and subcloned intothe pEGFP-N1 (Invitrogen, Carlsbad, CA) and pCMV-Myc vectors(Clontech, Palo Alto, CA). The SLC26A3 cDNA sequence was verifiedby nucleotide sequencing, and was found to be identical to a registeredsequence (GenBank accession NM_000111). For deletions of theSLC26A3 C-terminal PDZ-bindingmotif, oligonucleotide-directedmuta-genesis was performed. pcDNA3.1-HA-NHERF4was a kind gift from Dr.Sharon L. Milgram (NHLBI, NIH, MD). S329A- and S329D-NHERF4 weregenerated by oligonucleotide-directed mutagenesis. For the GSTpulldown analysis, the SLC26A3 C-terminal 43 amino acid sequencewas added to GST by using the pGEX-KGvector (AmershamBiosciences,Sweden). 6× His-tagged-truncated NHERF4-PDZ pulldown plasmidvectors were also a kind gift from Dr. Sharon L. Milgram.
2.3. Yeast two-hybrid analysis
The L40 yeast strain harbouring the reporter genes HIS3 and LacZunder the control of upstream LexA DNA-binding domain was used inthe assay. To semi-quantify the interaction, HIS3 activity was deter-mined by the percentage of yeast colonies growingon histidine-freeme-dium. cDNA sequences containing the carboxyl-terminal ~50 aminoacids of CFTR, SLC4A7, SLC26A3, SLC26A6, SLC26A7, and SLC26A9 wereamplified by PCR, and subcloned into the pBHA construct (a bait vectorcontaining the LexA DNA-binding domain). The pGAD10 (a prey vector,Clontech) constructs containing PDZ domains of full-length NHERF4were generated.
2.4. Immunoprecipitation and immunoblotting
HeLa cells and mouse intestine tissue were lysed with lysis buffer(1% NP-40, 150 mM NaCl, 1 mM EDTA, 50 mM Tris–HCl, pH 7.4) con-taining a complete protease inhibitor cocktail (Roche Applied Science,Mannheim, Germany). After lysis, cell or tissue debris was removedby centrifugation. Cleared lysates were mixed with the appropriate an-tibodies and incubated overnight at 4 °C. Immune complexes were col-lected by incubation for 2 h at 4 °Cwith protein A or G plus agarose, andthenwashed four timeswith lysis buffer for electrophoresis. The immu-noprecipitates and cell or tissue lysateswere suspended in 2× SDS sam-ple buffer, and then separated by SDS-PAGE. The separated proteinswere transferred to nitrocellulose membranes, and the membraneswere blocked by incubation for 1 h in a solution containing 5% nonfatdry milk in phosphate-buffered saline (PBS) containing 0.1% Tween20. The membranes were then incubated with the appropriate primaryand secondary antibodies. Enhanced chemiluminescence solution wasused to detect protein bands. Use of mouse intestine tissue for thisstudywas approved by the Committee for the Care andUse of Laborato-ry Animals at Yonsei University College of Medicine.
2.5. Immunocytochemistry
Immunostaining of SLC26A3- and NHERF4-transfected HeLa cellswas performed as previously described [20]. Briefly, approximately36 h after transfection, cells were fixed and permeabilised by incuba-tion in cold methanol for 10 min at−20 °C, and then stained with theprimary antibodies and the fluorescently labelled secondary anti-bodies. Anti-Myc and anti-HA were the primary antibodies used forstaining SLC26A3 and NHERF4, respectively. FITC- and rhodamine-labelled secondary antibodies were used for fluorescence imaging ofSLC26A3 and NHERF4, respectively. Fluorescent images were obtainedwith a Zeiss LSM 510 confocal microscope. The fluorescence of FITCwas excited at 488 nm by an argon laser, and the fluorescence emittedwas detected with a 505–530-nm band-pass filter. Fluorescence of rho-damine was excited at 543 nm by a He–Ne laser, and emitted fluores-cence was detected with a 560–615-nm band-pass filter.
2.6. GST pulldown analysis
GST fusion SLC26A3 C-terminal 43 amino acid peptides (Ct43aa)were expressed in Escherichia coli BL21 cells, and purified withglutathione-sepharose (Amersham Biosciences, Sweden). Four 6×His-tagged NHERF4 PDZ domains were also expressed in E. coli BL21cells and purified with Ni-NTA agarose beads (Qiagen, Germany).Glutathione-sepharose binding to the GST-fusion SLC26A3-Ct43aawas performed by incubating with 20 μg of each of the four purifiedNHERF4 PDZ-domain proteins overnight with gentle agitation at 4 °C.The glutathione-sepharose was pelleted and washed three times withice-cold lysis buffer prior to re-suspension in 2× SDS sample bufferand SDS-PAGE. Immunoblots for 6× His-tagged proteins were per-formed to detect 6× His-tagged NHERF4 PDZ domains.
2.7. Measurement of intracellular pH and Cl−/HCO3− exchange
Measurement of intracellular pH (pHi) and Cl−/HCO3− exchange of
SLC26A3 was performed as previously described [20,21]. Briefly, mea-surements of pHi in HeLa cells transiently transfected with SLC26A3were performed using the pH-sensitive fluorescent probe BCECFwith co-transfection of the trans-gene marker pEGFP-N1 (Invitrogen).A cluster of cells showing green fluorescent protein (GFP) fluorescencewas loaded with BCECF, and the pHi was monitored. To minimise varia-tions from transfection efficiency, cells showing GFP fluorescence of80%–120% levels of that of the control cells transfected with pEGFP-N1were chosen tomeasure the anion exchange in each set of experiments.Measurements of pHi in HT-29 epithelial monolayers transfected with

1823J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
scrambled or NHERF4-specific siRNAwere performed by using BCECF ina double perfusion chamber. After dye loading, the cells were perfusedwith a HCO3
−-buffered solution B, and BCECF fluorescencewas recordedat the excitation wavelengths of 490 and 440 nm at a resolution of 2/sby using a recording setup (Delta Ram; PTI Inc. Birmingham, NJ). The490:440 ratios were calibrated by perfusion cells with solution con-taining 145 mMKCl, 10 mMHEPES, and 5 μMnigericin with the pH ad-justed to 6.2–7.6. Cl−i/HCO3
−o exchange activities were estimated from
the initial rate of pHi increase as a result of Cl−o removal in the HCO3−-
containing buffer (25 mM HCO3− with 5% CO2 gassing). The intrinsic
buffer capacity (βi) was calculated by measuring the pHi in responseto 5–40 mM NH4Cl pulses in Na-free solution. The βi of HeLa cells was15.21 (±1.62) mM/pH unit at pHi 7.2, and this value was not signifi-cantly different for cells transfected with WT-SLC26A3 or SLC26A3-ΔETKF.
2.8. Fluorescence recovery after photobleaching
The FRAP analysis was performed in HeLa cells. Approximately36 h after transfection of GFP-SLC26A3 with or without NHERF4 plas-mids, cells were perfused with solution A at room temperature. The488-nm laser line of the argon laser was used for GFP excitation,and the emitted fluorescence was detected using a 505–530-nmband-pass filter. The regions of interest (ROI) on the plasma mem-brane of cells were photobleached at full laser power, and the recov-ery of fluorescence was monitored by scanning the cells over 10-sintervals. The scanning was performed with lower laser power ofabout 10%–15% of full laser power, and the intensity was recordedusing a recording setup for confocal microscopy (Zeiss LSM 700). Per-cent fluorescence (%) was calculated by comparing the intensity ratioin the ROI within the bleached area before the photobleaching at eachtime interval. The recovery rates were estimated from percent fluo-rescence when the percent fluorescence reached a plateau (~5 minafter photobleaching). Diffusional mobility was estimated from theinitial rate of the percent fluorescence increase after photobleaching(Δpercent fluorescence/min).
2.9. Surface biotinylation and endocytic assay
Surface biotinylation of SLC26A3 was performed as previously de-scribed [20]. Briefly, transfected cells were washed with ice-cold PBS,and then incubated with another ice-cold PBS solution containing0.1 mM CaCl2 and 1 mMMgCl2 for 5 min. Cells were then biotinylatedusing EZ-Link sulfo-NHS-SS-biotin (Pierce, Rockford, IL; 0.5 mg/mL)for 30 min in the dark at 4 °C. After washing away free biotin with bo-vine serum albumin-containing (1% w/v) PBS, cells were lysed withlysis buffer. The lysates were centrifuged for 10 min (13,000 g), andthe pellet was discarded. Avidin solution (Streptavidin beads, Pierce,50 μL) was added to the supernatant (300 μg of protein in 300 μL oflysis buffer), and the mixture was incubated overnight with gentleagitation. Avidin-bound complexes were pelleted (13,000 g), washedthree times with lysis buffer, and immunoblotted.
The endocytic assay of SLC26A3was performed based on a previous-ly described method [22]. Briefly, transfected cells were biotinylatedusing EZ-Link sulfo-NHS-SS-biotin for 30 min in the dark at 4 °C. Subse-quently, to induce endocytosis, the cells werewarmed to 37 °C for 5, 15,and 30min, and the remaining disulphide bonds on sulfo-NHS-SS-biotinylated proteins were reduced with 50 mM glutathione (GSH)added to the PBS solution six times for a total of 90 min at 4 °C. At thispoint in the protocol, biotinylated proteins reside within the endosomalcompartment. Subsequently, the cells were lysed with lysis buffer. Thelysates were centrifuged for 10 min (13,000 g), and the pellet was dis-carded. Avidin solution (50 μL, Streptavidin beads, Pierce) was addedto the supernatant (300 μg of protein in 300 μL of lysis buffer), and themixture was incubated overnight with gentle agitation. Avidin-bound
complexes were pelleted (13,000 g), washed three times with lysisbuffer, and immunoblotted.
2.10. Statistical analysis
The results of multiple experiments are presented as the means±SEM. Statistical analysis was performed with either the paired or un-paired two-tailed Student's t-tests using statistical software (GraphPad,San Diego, CA). Pb0.05 was considered statistically significant.
3. Results
3.1. SLC26A3 acts as a novel binding transporter of NHERF4 by aPDZ-mediated interaction
To screen binding gastric epithelial transporters of NHERF4, weused Y2H analysis. We selected key gastric epithelial transporterswith a class I PDZ-binding motif as candidates: CFTR, SLC4A7,SLC26A3, SLC26A6, SLC26A7, and SLC26A9. C-terminal PDZ-bindingmotifs in the transporters were used as bait, and PDZ domains offull-length NHERF4 were used as the prey. The Y2H analysis revealedthat NHERF4 interactedwith SLC26A3 and SLC26A6with high intensity,and with SLC26A9 and CFTR with low intensity. Among the bindingtransporters, we focussed on the binding between SLC26A3 andNHERF4 in the present study.
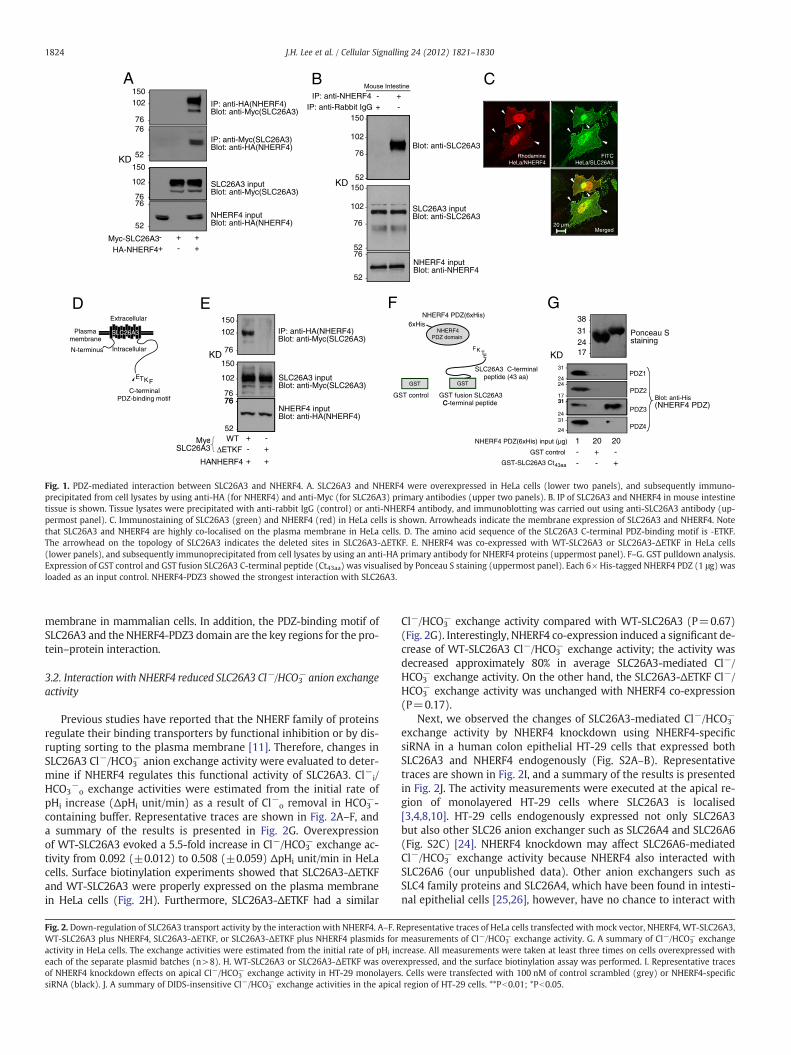
Although Y2H analysis is a powerfulmolecular tool to study protein–protein interactions, this technique has an inherent limitation that theinteractions are based on the yeast system. Therefore, SLC26A3 andNHERF4 interactions in mammalian cell-line systems were confirmedby IP. Myc-tagged wild-type (WT) SLC26A3 and HA-tagged NHERF4were overexpressed in HeLa cells, followed by IPwith antibodies againstthe SLC26A3 N-terminal Myc-epitope or the NHERF4 N-terminal HA-epitope (Fig. 1A).WT-SLC26A3wasobserved in the immunoprecipitateswith anti-HA antibody. In a converse experiment, NHERF4was detectedin the immunoprecipitateswith anti-Myc antibody. The protein–proteininteraction between endogenous SLC26A3 andNHERF4 in themouse in-testine tissuewas also observed (Fig. 1B and Fig. S1). An equal amount oftissue lysates (500 μg)was precipitatedwith anti-rabbit IgG (control) oranti-NHERF4 antibody, and only SLC26A3was observed in the immuno-precipitates with anti-NHERF4 antibody. We expected that the bandsaround 70 and 110 kDa were non-specific bands detected in mouse in-testine tissue (Fig. S1). The intracellular localisation of WT-SLC26A3and NHERF4 was examined using immunocytochemistry in HeLa cellsthat heterologously expressed SLC26A3 and NHERF4 (Fig. 1C). NHERF4and SLC26A3 co-localised to the plasma membrane region. To verifythat the protein–protein interaction between SLC26A3 and NHERF4 isPDZ-dependent, we generated deletions of the C-terminal PDZ-bindingmotif of SLC26A3 (SLC26A3-ΔETKF) (Fig. 1D) and examined its IP withNHERF4 in HeLa cells (Fig. 1E). Total levels of SLC26A3-ΔETKF expres-sion were similar with the levels of WT-SLC26A3 expression. The IP ex-periment revealed that SLC26A3-ΔETKF did not interact with NHERF4.
A combination of various amino acid sequences at the bindingpocket region of the PDZ domain is a crucial factor defining bindingselectivity [23]. NHERF4 possesses four PDZ domains, and each PDZconsists of different amino acid sequences [12]. Hence, each of thefour NHERF4 PDZ domains likely has different binding selectivity tothe SLC26A3 PDZ-binding motif. To identify a specific NHERF4 PDZ do-main that interacts with SLC26A3, we performed a GST pulldown assay.GST-fused SLC26A3 C-terminal 43 amino acids (SLC26A3-Ct43) and fourtruncated 6×His-taggedNHERF4 PDZdomains (PDZ158–130, PDZ2165–235,PDZ3271–346, and PDZ4410–482) were used for the assay (Fig. 1F). The thirdPDZ of NHERF4 (NHERF4-PDZ3) showed the strongest interaction withSLC26A3-Ct43 (Fig. 1G). Although the fourth PDZ of NHERF4 showedsome interaction, it was much weaker compared with NHERF4-PDZ3.These results indicate that the SLC26A3–NHERF4 interaction is PDZ-mediated, and the proteins localise at cell surface regions of the plasma
76 -
52 -
150 -
102 -
76 -
150 -
IP: anti-HA(NHERF4)Blot: anti-Myc(SLC26A3)
IP: anti-Myc(SLC26A3)Blot: anti-HA(NHERF4)
KD
A
- + ++ - +
76 -
52 -
102 -
76 -
SLC26A3 inputBlot: anti-Myc(SLC26A3)
NHERF4 inputBlot: anti-HA(NHERF4)
Myc-SLC26A3HA-NHERF4
B
52 -
150 -
102 -
76 -
- ++ -
IP: anti-NHERF4IP: anti-Rabbit IgG
Blot: anti-SLC26A3
Mouse IntestineC
Rhodamine HeLa/NHERF4
FITC HeLa/SLC26A3
-
52 -
150 -
102 -
76 -
52 -
76 -
KD
SLC26A3 inputBlot: anti-SLC26A3
NHERF4 inputBlot: anti-NHERF4
20 µmMerged
N-terminus
ETKFC-terminal
PDZ-binding motif
Plasma membrane
SLC26A3
Extracellular
Intracellular
D
150 -
102 -
76 -
150 -
102 -
76 -
76 -
KD
E
IP: anti-HA(NHERF4)Blot: anti-Myc(SLC26A3)
SLC26A3 inputBlot: anti-Myc(SLC26A3)
76 -
52 -
Myc-SLC26A3
HA-NHERF4
WT+-
++
-+
ΔETKF
NHERF4 inputBlot: anti-HA(NHERF4)
F
GST
6xHisNHERF4
PDZ domain
SLC26A3 C-terminal peptide (43 aa)
ETKF
GST control GST fusion SLC26A3 C
NHERF4 PDZ(6xHis)
GST
38 -
31 -
24 -
17 -
PDZ2
PDZ131 -
24 -24 -
17 -31 -
KD
Blot: anti-His (NHERF4 PDZ)
Ponceau S staining
G
C-terminal peptidePDZ3
PDZ4
31 -
24 -31 -
24 -
NHERF4 PDZ(6xHis) input (µg)
GST control
GST-SLC26A3 Ct43aa
1 20 20- + -- - +
Fig. 1. PDZ-mediated interaction between SLC26A3 and NHERF4. A. SLC26A3 and NHERF4 were overexpressed in HeLa cells (lower two panels), and subsequently immuno-precipitated from cell lysates by using anti-HA (for NHERF4) and anti-Myc (for SLC26A3) primary antibodies (upper two panels). B. IP of SLC26A3 and NHERF4 in mouse intestinetissue is shown. Tissue lysates were precipitated with anti-rabbit IgG (control) or anti-NHERF4 antibody, and immunoblotting was carried out using anti-SLC26A3 antibody (up-permost panel). C. Immunostaining of SLC26A3 (green) and NHERF4 (red) in HeLa cells is shown. Arrowheads indicate the membrane expression of SLC26A3 and NHERF4. Notethat SLC26A3 and NHERF4 are highly co-localised on the plasma membrane in HeLa cells. D. The amino acid sequence of the SLC26A3 C-terminal PDZ-binding motif is ‐ETKF.The arrowhead on the topology of SLC26A3 indicates the deleted sites in SLC26A3-ΔETKF. E. NHERF4 was co-expressed with WT-SLC26A3 or SLC26A3-ΔETKF in HeLa cells(lower panels), and subsequently immunoprecipitated from cell lysates by using an anti-HA primary antibody for NHERF4 proteins (uppermost panel). F–G. GST pulldown analysis.Expression of GST control and GST fusion SLC26A3 C-terminal peptide (Ct43aa) was visualised by Ponceau S staining (uppermost panel). Each 6× His-tagged NHERF4 PDZ (1 μg) wasloaded as an input control. NHERF4-PDZ3 showed the strongest interaction with SLC26A3.
1824 J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
membrane in mammalian cells. In addition, the PDZ-binding motif ofSLC26A3 and the NHERF4-PDZ3 domain are the key regions for the pro-tein–protein interaction.
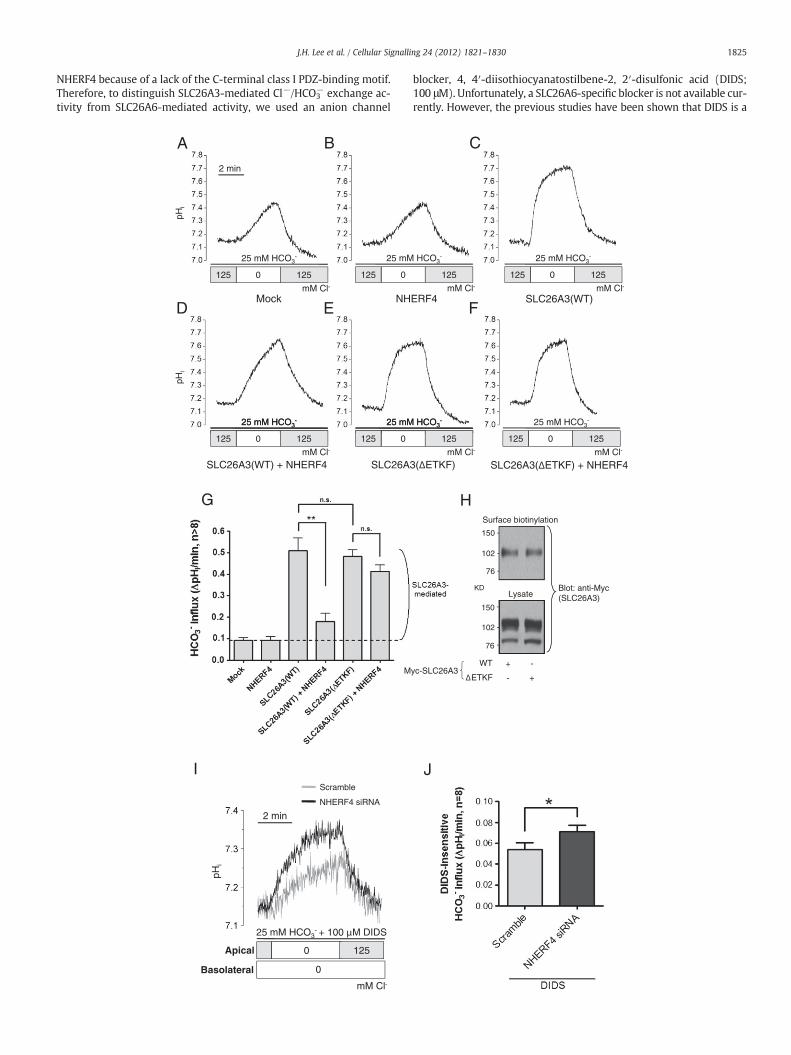
3.2. Interaction with NHERF4 reduced SLC26A3 Cl−/HCO3− anion exchange
activity
Previous studies have reported that the NHERF family of proteinsregulate their binding transporters by functional inhibition or by dis-rupting sorting to the plasma membrane [11]. Therefore, changes inSLC26A3 Cl−/HCO3
− anion exchange activity were evaluated to deter-mine if NHERF4 regulates this functional activity of SLC26A3. Cl−i/HCO3
−o exchange activities were estimated from the initial rate of
pHi increase (ΔpHi unit/min) as a result of Cl−o removal in HCO3−-
containing buffer. Representative traces are shown in Fig. 2A–F, anda summary of the results is presented in Fig. 2G. Overexpressionof WT-SLC26A3 evoked a 5.5-fold increase in Cl−/HCO3
− exchange ac-tivity from 0.092 (±0.012) to 0.508 (±0.059) ΔpHi unit/min in HeLacells. Surface biotinylation experiments showed that SLC26A3-ΔETKFand WT-SLC26A3 were properly expressed on the plasma membranein HeLa cells (Fig. 2H). Furthermore, SLC26A3-ΔETKF had a similar
Fig. 2. Down-regulation of SLC26A3 transport activity by the interaction with NHERF4. A–F. RWT-SLC26A3 plus NHERF4, SLC26A3-ΔETKF, or SLC26A3-ΔETKF plus NHERF4 plasmids foractivity in HeLa cells. The exchange activities were estimated from the initial rate of pHi ineach of the separate plasmid batches (n>8). H. WT-SLC26A3 or SLC26A3-ΔETKF was overof NHERF4 knockdown effects on apical Cl−/HCO3
− exchange activity in HT-29 monolayerssiRNA (black). J. A summary of DIDS-insensitive Cl−/HCO3
− exchange activities in the apica
Cl−/HCO3− exchange activity compared with WT-SLC26A3 (P=0.67)
(Fig. 2G). Interestingly, NHERF4 co-expression induced a significant de-crease of WT-SLC26A3 Cl−/HCO3
− exchange activity; the activity wasdecreased approximately 80% in average SLC26A3-mediated Cl−/HCO3
− exchange activity. On the other hand, the SLC26A3-ΔETKF Cl−/HCO3
− exchange activity was unchanged with NHERF4 co-expression(P=0.17).
Next, we observed the changes of SLC26A3-mediated Cl−/HCO3−
exchange activity by NHERF4 knockdown using NHERF4-specificsiRNA in a human colon epithelial HT-29 cells that expressed bothSLC26A3 and NHERF4 endogenously (Fig. S2A–B). Representativetraces are shown in Fig. 2I, and a summary of the results is presentedin Fig. 2J. The activity measurements were executed at the apical re-gion of monolayered HT-29 cells where SLC26A3 is localised[3,4,8,10]. HT-29 cells endogenously expressed not only SLC26A3but also other SLC26 anion exchanger such as SLC26A4 and SLC26A6(Fig. S2C) [24]. NHERF4 knockdown may affect SLC26A6-mediatedCl−/HCO3
− exchange activity because NHERF4 also interacted withSLC26A6 (our unpublished data). Other anion exchangers such asSLC4 family proteins and SLC26A4, which have been found in intesti-nal epithelial cells [25,26], however, have no chance to interact with
epresentative traces of HeLa cells transfected with mock vector, NHERF4, WT-SLC26A3,measurements of Cl−/HCO3
− exchange activity. G. A summary of Cl−/HCO3− exchange
crease. All measurements were taken at least three times on cells overexpressed withexpressed, and the surface biotinylation assay was performed. I. Representative traces. Cells were transfected with 100 nM of control scrambled (grey) or NHERF4-specificl region of HT-29 cells. **Pb0.01; *Pb0.05.
1825J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
NHERF4 because of a lack of the C-terminal class I PDZ-binding motif.Therefore, to distinguish SLC26A3-mediated Cl−/HCO3
− exchange ac-tivity from SLC26A6-mediated activity, we used an anion channel
G
M
2 min
I
NHERF4 siRNA
Scramble
pHi
mM Cl-
25 mM HCO3- + 100 M DIDS
0 125
0
Apical
Basolateral
2 min
mM Cl-
25 mM HCO3-
125 0 125
25 mM
125 0
A
pHi
B
Mock NH
25 mM
125 0
mM Cl-
25 mM HCO3-
125 0 125
pHi
D E
blocker, 4, 4′-diisothiocyanatostilbene-2, 2′-disulfonic acid (DIDS;100 μM). Unfortunately, a SLC26A6-specific blocker is not available cur-rently. However, the previous studies have been shown that DIDS is a
Surface biotinylation
LysateKD Blot: anti-Myc
(SLC26A3)
H
150 -
102 -
76 -
150 -
+ -
- +
WT
ETKFyc-SLC26A3
102 -
76 -
J
mM Cl-
HCO3-
125
C
ERF4 SLC26A3(WT)mM Cl-
25 mM HCO3-
125 0 125
mM Cl-
25 mM HCO3-
125 0 125
mM Cl-
HCO3-
125
F

1826 J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
potent blocker of SLC26A6 but is not sensitive to SLC26A3 at low doses(Fig. S2D) [27–31]. NHERF4 knockdown evoked a significant increase inDIDS-insensitive Cl− /HCO3
− exchange activity from 0.052 (±0.006) to0.071 (±0.006) ΔpHi unit/min in apical region of HT-29 cells. Thesedata indicate that the PDZ-mediated NHERF4 interaction negativelyregulates SLC26A3-mediated Cl−/HCO3
− exchange activity.
3.3. NHERF4 decreased the level of SLC26A3 expression on the plasmamembrane through rapid internalisation
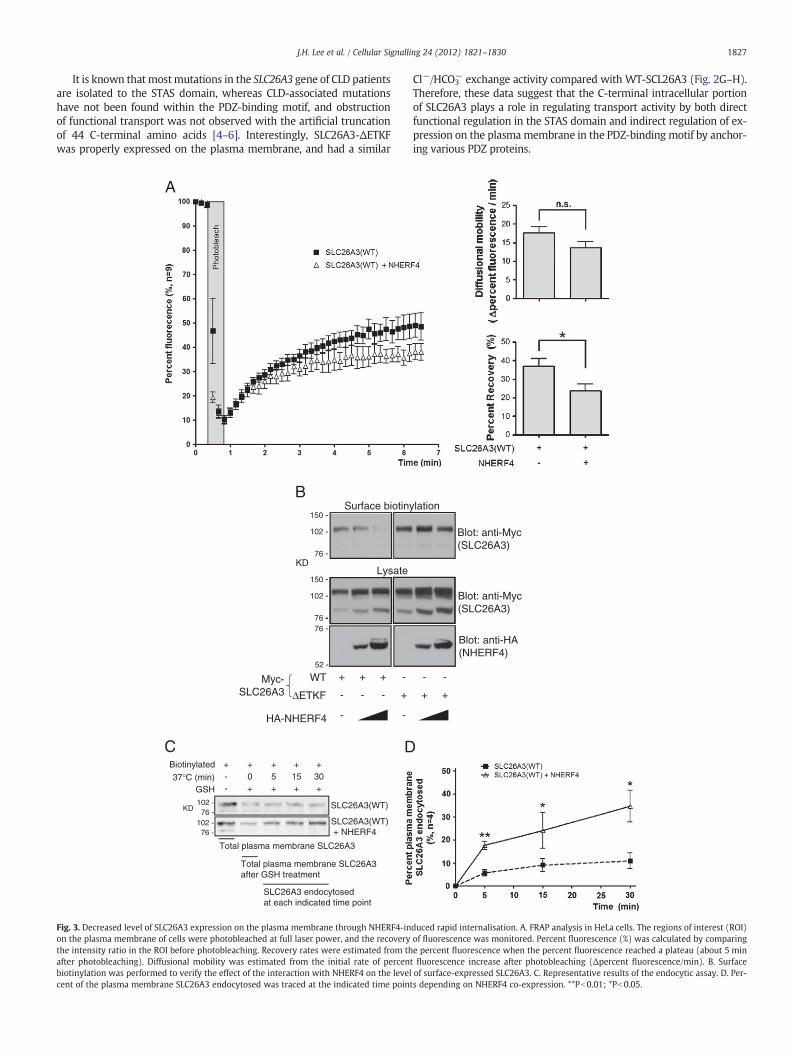
Next, we investigated whether NHERF4 alters the level of SLC26A3expression on the plasma membrane. Down-regulation of transporteractivity can be induced not only by direct inhibition of transport func-tional activity but also by disturbance of stable expression on the plas-ma membrane. FRAP analysis revealed that the interaction withNHERF4 decreased approximately 13% of the recovery rate of SLC26A3on the plasma membrane after photobleaching (i.e. from 37.03%(±3.99) to 23.75% (±3.48)) (Fig. 3A). On the other hand, the differencein SLC26A3 diffusional mobility (Δpercent fluorescence/min) on theplasma membrane was unaffected by the NHERF4 interaction(P=0.12). These data suggest that the NHERF4 interaction does not af-fect SLC26A3 mobility on the plasma membrane, but the level ofsurface-expressed SLC26A3 is altered. Therefore, we investigated thechange in the surface-expressed level of SLC26A3, which is dependenton the interaction with NHERF4, by using surface biotinylation. Surfacebiotinylation experiments revealed that NHERF4 significantly de-creased the surface-expression level of WT-SLC26A3 (Fig. 3B). Notably,the level of surface-expressed WT-SLC26A3 was inversely proportionalto the level of NHERF4 expression. On the other hand, NHERF4 did notaffect the level of surface-expressed SLC26A3-ΔETKF, which does notinteract with NHERF4.
The rate of internalisation of surface-expressed SLC26A3 that wasdependent on theNHERF4 interactionwas evaluatedusing an endocyticassay (Fig. 3C–D). Cells were surface-biotinylated at 4 °C after SLC26A3overexpression with or without NHERF4, and then, the cells were incu-bated at 37 °C for 5, 15, or 30 min to induce endocytosis. The level ofbiotin-labelled proteins resistant to GSH reduction that are endocyticproteins was used to determine the rate of internalisation. NHERF4co-expression induced significantly more rapid and quantitative inter-nalisation of surface-expressed SLC26A3 at time points within 30 min.The percentage of SLC26A3 internalisation was only 10.93% (±3.53),whereas NHERF4 co-expression led to 34.67% (±6.93) of SLC26A3internalisation at 30 min. These results indicate that the NHERF4 inter-action decreased the level of surface-expressed SLC26A3 by inducingrapid internalisation.
3.4. Phosphorylation of NHERF4 regulates the interaction with SLC26A3
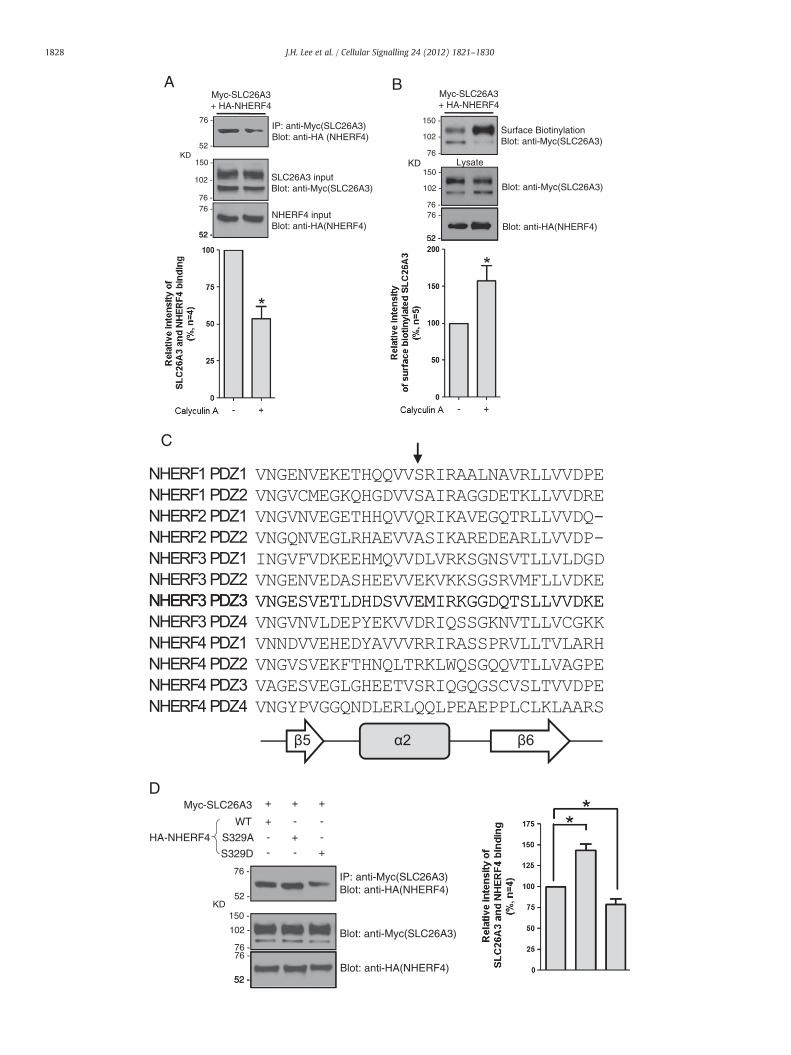
It has been suggested that NHERF PDZ proteins are involved in dy-namic roles such as signal transduction by clustering of associated pro-teins as well as in static roles as a scaffold at certain cell surface regions[11]. For example, PDZ phosphorylation of NHERF3 regulates the re-cruitment of scavenger receptor class B type I (SRBI) [32]. We investi-gated whether phosphorylation modulates the SLC26A3–NHERF4interaction. After administration of a cell-permeable phosphatase inhibi-tor, calyculin A (10 nM for 30 min), we measured the intensity of theSLC26A3–NHERF4 interaction by IP and the level of surface-expressedSLC26A3 by surface biotinylation inHeLa cells (Fig. 4A–B). The IP exper-iment revealed that calyculin A-induced phosphorylation decreased theintensity of the SLC26A3–NHERF4 interaction by approximately 46%and increased the level of surface-expressed SLC26A3 by 1.6 folds.
Recently, Weinman et al. reported that serine 77 of NHERF1(NHERF1-S77) regulated its binding selectivity to the target PDZ bind-ing motif by phosphorylation [33–35]. NHERF1-S77 is located on thesurface of the second α-helix in NHERF1-PDZ1 where the target PDZbinding motif interacts. Interestingly, multiple sequence alignments
within all PDZ domains of the NHERF family revealed that serine 329of NHERF4-PDZ3 (NHERF4-S329) is located on the secondα-helix sim-ilar to NHERF1-S77 (Fig. 4C). However, NHERF4-PDZ1, PDZ2, and PDZ4have different amino acid residues at this site: arginine, arginine, andglutamine, respectively. Interestingly, we identified NHERF4-PDZ3 tobe the key domain to interactwith SLC26A3 (Fig. 1G). To verifywhetherphosphorylation at NHERF4-S329 indeed defines binding selectivity ofthe SLC26A3–NHERF4 interaction, two substitutions were generatedby the oligonucleotide-directed mutagenesis method: substitution toalanine (S329A) and to aspartate (S329D). The S329A substitution abol-ishes phosphorylation of the serine residue, whereas S329D mimicsphosphorylation at that site [33,34,36]. IP experiments revealed thatthe phospho-abolishing mutation (S329A) increased the intensity ofthe SLC26A3 interaction, whereas the phospho-mimicking mutation(S329D) showed a significant decrease in the intensity of the SLC26A3interaction (Fig. 4D). These data indicate that the SLC26A3–NHERF4 inter-action is inhibited by phosphorylation, and NHERF4-S329, in particular, isthe residue that modulates the binding selectivity by phosphorylation.
4. Discussion
Using a PDZ-mediated interaction, SLC26A3 recruits various regula-tory proteins such as E3KARP (sodium/proton exchanger 3 protein ki-nase A regulatory protein) with other transporters including SLC26A6,NHE3, and CFTR [5,6,37]. Although a C-terminal PDZ-binding motif ofSLC26A3 is not directly necessary for transport activity, PDZ scaffoldproteins modulate its expression and transport activity by assemblingregulatory factors and other transporters [6,9,10]. In this study, ourdata indicate that NHERF4 also regulates SLC26A3 by a PDZ-mediateddirect protein–protein interaction. Interaction with NHERF4 reducedthe half-time of SLC26A3 expression on the plasmamembrane becauseof rapid induction of internalisation of surface-expressed SLC26A3.Therefore, interaction with NHERF4 led to a reduction of SLC26A3Cl−/HCO3
− exchange activity. On the other hand, data in Figs. 2G and3B using SLC26A3-ΔETKF, which does not interact with NHERF4,demonstrate that both the expression level on the plasmamembraneand the Cl−/HCO3
− exchange activity of SLC26A3-ΔETKF were not dif-ferent in the presence or absence of NHERF4.
The NHERF family of proteins are well known as epithelial-enriched PDZdomain scaffold proteins thatmodulate epithelial cell func-tions by co-ordination of regulatory factors, second messenger cascades,and various transporters [11,12]. However, the functional roles and reg-ulatory mechanisms of the most recently identified NHERF4 still remainmostly unknown. In the present study, we determined that NHERF4interacted with several gastric epithelial transporters including CFTR,SLC26A3, SLC26A6, and SLC26A9, but not with SLC4A7 and SLC26A7 (in-teraction intensity: SLC26A3=SLC26A6>CFTR=SLC26A9, data notshown) by using Y2H screening. These Y2H data are consistent withour hypothesis that NHERF4 would not interact with SLC4A7 orSLC26A7, because it is known that NHERF4 is expressed in the apicalmembrane region of epithelial cells, whereas SLC4A7 and SLC26A7 arebasolateral transporters [11,12,38,39]. However, contrary to Y2H results,IP experiments in HeLa cells revealed that NHERF4 did not interact withCFTR or SLC26A9 (data not shown).We verified that SLC26A3 interactedwith NHERF4 by both IP experiments in HeLa cells (Fig. 1A) and a GSTpulldown analysis (Fig. 1G). Our remaining questions are whetherNHERF4 regulates SLC26A6 and whether NHERF4 plays a role of a scaf-fold protein for the SLC26A3 and SLC26A6 protein complex. Gastric ep-ithelial transporters such as CFTR and SLC26 family including SLC26A3and SLC26A6 form a macromolecular complex by binding to PDZ scaf-fold proteins. CFTR easily activates SLC26A3 and SLC26A6, and viceversa in the complex [5]. However, our unpublished data showed thatNHERF4 may be not a PDZ scaffold protein for the complex includingCFTR (data now shown), but NHERF4 may regulate the SLC26 familyprotein complex by phosphorylation dependent mechanism.
1827J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
It is known that mostmutations in the SLC26A3 gene of CLD patientsare isolated to the STAS domain, whereas CLD-associated mutationshave not been found within the PDZ-binding motif, and obstructionof functional transport was not observed with the artificial truncationof 44 C-terminal amino acids [4–6]. Interestingly, SLC26A3-ΔETKFwas properly expressed on the plasma membrane, and had a similar
A
B150 -
102 -
76 -
Surface biotiny
150 -
102 -
76 -
KDLysate
-
+
-
+
-
-
+
+
-
-
ΔETKF
HA-NHERF4
WTMyc-SLC26A3
76 -
52 -
-
C
+5+
+++-30150-++++
GSH37°C (min)
Biotinylated
SLC26A3(WT)
SLC26A3(WT)+ NHERF4
Total plasma membrane SLC26A3
102 -76 -
102 -76 -
KD
D
Total plasma membrane SLC26A3 after GSH treatment
SLC26A3 endocytosed at each indicated time point
Fig. 3. Decreased level of SLC26A3 expression on the plasma membrane through NHERF4-inon the plasma membrane of cells were photobleached at full laser power, and the recoverythe intensity ratio in the ROI before photobleaching. Recovery rates were estimated from thafter photobleaching). Diffusional mobility was estimated from the initial rate of percenbiotinylation was performed to verify the effect of the interaction with NHERF4 on the levelcent of the plasma membrane SLC26A3 endocytosed was traced at the indicated time poin
Cl−/HCO3− exchange activity compared with WT-SCL26A3 (Fig. 2G–H).
Therefore, these data suggest that the C-terminal intracellular portionof SLC26A3 plays a role in regulating transport activity by both directfunctional regulation in the STAS domain and indirect regulation of ex-pression on the plasma membrane in the PDZ-binding motif by anchor-ing various PDZ proteins.
lation
Blot: anti-Myc(SLC26A3)
Blot: anti-Myc(SLC26A3)
++
--
Blot: anti-HA(NHERF4)
duced rapid internalisation. A. FRAP analysis in HeLa cells. The regions of interest (ROI)of fluorescence was monitored. Percent fluorescence (%) was calculated by comparinge percent fluorescence when the percent fluorescence reached a plateau (about 5 mint fluorescence increase after photobleaching (Δpercent fluorescence/min). B. Surfaceof surface-expressed SLC26A3. C. Representative results of the endocytic assay. D. Per-ts depending on NHERF4 co-expression. **Pb0.01; *Pb0.05.
AMyc-SLC26A3+ HA-NHERF4
NHERF4 inputBlot: anti-HA(NHERF4)
SLC26A3 inputBlot: anti-Myc(SLC26A3)
IP: anti-Myc(SLC26A3)Blot: anti-HA (NHERF4)
KD
102 -
76 -
150 -
76 -
52 -
76 -
52 -
B
102 -
76 -
150 -
KD Lysate
102 -
76 -
150 -
76 -
52 -
Myc-SLC26A3+ HA-NHERF4
Surface BiotinylationBlot: anti-Myc(SLC26A3)
Blot: anti-Myc(SLC26A3)
Blot: anti-HA(NHERF4)52 - 52 -
76 -
52 -
150 -
102 -
76 -76 -
52 -
WT
HA-NHERF4 S329A
S329D
Myc-SLC26A3
--+
-+-+--
+++
KD
Blot: anti-HA(NHERF4)
Blot: anti-Myc(SLC26A3)
IP: anti-Myc(SLC26A3)Blot: anti-HA(NHERF4)
D
52 -
C
1828 J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830

1829J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
Recently, Zachos et al. showed that NHE3was a new binding partnerof NHERF4, and this interaction reduced NHE3 activity in an intracellu-lar calcium-dependent manner [40]. They reported that (1) the NHE3–NHERF4 complex was localised in the Rab11-positive recycling endo-some and (2) elevated intracellular calcium levels disrupted theinteraction between NHE3 and NHERF4, which increased NHE3 expres-sion on the plasma membrane and stimulated NHE3 activity. Similarly,we demonstrated that the NHERF4 interaction decreased SLC26A3 ac-tivity by reducing expression on the plasma membrane. However, inthe present study, we identified a different regulatory mechanism ofNHERF4. The data in Fig. 3A–D show that the interaction with NHERF4affected the level of surface-expressed SLC26A3 as a result of NHERF4-induced rapid internalisation of SLC26A3 from the plasma membrane.Furthermore, our data indicate that the SLC26A3–NHERF4 interactionwasmodulated by phosphorylation; the phosphorylation state of serine329 in NHERF4-PDZ3 modulated the binding selectivity (Fig. 4A–D).
Another key remaining question is whether NHERF4 recruits otherregulatory factors to modulate SLC26A3 expression on the plasmamembrane. We hypothesised that NHERF4 directly recruits regulato-ry factors through its own PDZ domains or indirectly by anchoringother NHERF proteins. The candidates for direct recruitment are severalendocytic Rab small guanosine triphosphatases and soluble NSF attach-ment protein receptor (SNARE) proteins [41–43]. We screened a fewregulatory factors by IP. Unfortunately, the screening results showedthat NHERF4 did not interact with Rab11 among the candidates (datanot shown). On the other hand, NHERF family proteins have theirown PDZ-binding motif at the C-terminus. Therefore, they can formintermolecular interactions and head-to-tail intramolecular interac-tions using their own PDZ domains and PDZ-binding motif [11,44]. In-terestingly, our unpublished data showed that deletion of the PDZ-binding motif of NHERF4 (NHERF4-ΔSDLL) tended to increase thelevel of surface-expressed SLC26A3 compared to WT-NHERF4 (datanot shown), which suggests that NHERF4may form a NHERF family pro-tein complex and recruit various regulatory proteins either directly or in-directly. In addition, theNHERF family protein complexmay interactwiththe cytoskeleton using the ERM-binding domain at the C-terminus ofNHERF1 or NHERF2.
Our findings suggest that NHERF4 is a modulator of luminal fluidityin the intestine by adjusting SLC26A3 expression and activity. SLC26A3has a crucial role in luminal chloride re-uptake in the apical membraneof the ileal and colonic surface epithelium [3–6]. Indeed, in this study,we showed not only the interaction between endogenous SLC26A3andNHERF4 proteins inmouse intestine tissue (Fig. 1B) but also the en-hancement of DIDS-insensitive Cl−/HCO3
− exchange activity byNHERF4knockdown in human colon epithelial HT-29 cells. We predict that re-sults from this study can be used for pharmacological applications toregulate luminal fluidity and diseases related to the intestine and kid-ney. For example, a short chemical peptide, which is identical to theSLC26A3 PDZ-binding motif sequence, could be used as a competitorof the SLC26A3–NHERF4 interaction. Specific chemical molecules thatinduce phosphorylation or dephosphorylation of NHERF4-PDZ3 couldalso be potential candidates.
In conclusion, this study clearly shows that a PDZ-mediated interac-tion with NHERF4 down-regulates SLC26A3 activity through inducingrapid internalisation of surface-expressed SLC26A3. The SLC26A3–NHERF4 interaction is dependent on phosphorylation,which is primarilymodulated by S329 in NHERF4-PDZ3. Our findings suggest that regula-tion of the SLC26A3–NHERF4 interaction through a phosphorylation-
Fig. 4. Regulation of the SLC26A3–NHERF4 interaction by phosphorylation. A. After SLC26A3inhibitor, calyculin A (10 nM for 30 min), was administered. Subsequently, IP was used to eoverexpressed in HeLa cells for 36 h. After inhibition of newly synthesised SLC26A3 and NH30 min) was administered. Surface biotinylation was performed using the cell lysates. C. Amsoftware. The arrow indicates a residue on the surface of the second α-helix of the PDZ domanner as NHERF1-S77. The predicted secondary structure of the second α-helix in the PDexpressed with WT-SLC26A3 in HeLa cells and immunoprecipitated from cell lysates by usi
dependentmechanism plays a critical role in themodulation of luminalfluidity in the intestine.
Acknowledgements
We thank Dr. Kyung Hwan Kim for helpful discussions and editorialassistance, and Hee Wook Lee and Yonsei-Carl Zeiss Advance ImagingCenter, Yonsei University College of Medicine for technical assistance.We also thank Dr. Sharon L. Milgram for the kind gift of the NHERF4plasmid constructs. This work was supported by the National ResearchFoundation of Korea (NRF) Grant funded by the Korean Government(MEST) (NRF-2010-0024820).
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.cellsig.2012.05.010.
References
[1] K.E. Barrett, S.J. Keely, Annual Review of Physiology 62 (2000) 535–572.[2] C.W. Schweinfest, K.W. Henderson, S. Suster, N. Kondoh, T.S. Papas, Proceedings
of the National Academy of Sciences of the United States of America 90 (9)(1993) 4166–4170.
[3] P.A. Dawson, D. Markovich, Current Medicinal Chemistry 12 (4) (2005) 385–396.[4] D.B. Mount, M.F. Romero, Pflügers Archiv 447 (5) (2004) 710–721.[5] M.R. Dorwart, N. Shcheynikov, D. Yang, S. Muallem, Physiology (Bethesda, Md.)
23 (2008) 104–114.[6] P. Hoglund, Novartis Foundation Symposium 273 (2006) 74–86 (discussion
86–90, 261–264).[7] S. Makela, J. Kere, C. Holmberg, P. Hoglund, Human Mutation 20 (6) (2002)
425–438.[8] J. Kere, H. Lohi, P. Hoglund, American Journal of Physiology 276 (1 Pt 1) (1999)
G7–G13.[9] S. Wedenoja, E. Pekansaari, P. Hoglund, S. Makela, C. Holmberg, J. Kere, Human
Mutation 32 (7) (2011) 715–722.[10] M.R. Dorwart, N. Shcheynikov, J.M. Baker, J.D. Forman-Kay, S. Muallem, P.J.
Thomas, Journal of Biological Chemistry 283 (13) (2008) 8711–8722.[11] T. Sugiura, T. Shimizu, A. Kijima, S. Minakata, Y. Kato, Journal of Pharmaceutical
Sciences 100 (9) (2011) 3620–3635.[12] W.R. Thelin, C.A. Hodson, S.L. Milgram, The Journal of Physiology 567 (Pt 1)
(2005) 13–19.[13] J.H. Lee, W. Richter, W. Namkung, K.H. Kim, E. Kim, M. Conti, M.G. Lee, Journal of
Biological Chemistry 282 (14) (2007) 10414–10422.[14] R.O. Scott, W.R. Thelin, S.L. Milgram, Journal of Biological Chemistry 277 (25)
(2002) 22934–22941.[15] M.A. Lanaspa, H. Giral, S.Y. Breusegem, N. Halaihel, G. Baile, J. Catalan, J.A.
Carrodeguas, N.P. Barry, M. Levi, V. Sorribas, American Journal of Physiology -Renal Physiology 292 (1) (2007) F230–F242.
[16] T. Hegedus, T. Sessler, R. Scott, W. Thelin, E. Bakos, A. Varadi, K. Szabo, L. Homolya,S.L. Milgram, B. Sarkadi, Biochemical and Biophysical Research Communications302 (3) (2003) 454–461.
[17] S.F. van de Graaf, J.G. Hoenderop, A.W. van der Kemp, S.M. Gisler, R.J. Bindels,Pflügers Archiv 452 (4) (2006) 407–417.
[18] H.J. Kim, D.K. Yang, I. So, Biochemical and Biophysical Research Communications361 (2) (2007) 433–438.
[19] C. Watanabe, Y. Kato, T. Sugiura, Y. Kubo, T. Wakayama, S. Iseki, A. Tsuji, DrugMetabolism and Disposition 34 (11) (2006) 1927–1934.
[20] J.S. Yoon, H.J. Park, S.Y. Yoo, W. Namkung, M.J. Jo, S.K. Koo, H.Y. Park, W.S. Lee, K.H.Kim, M.G. Lee, Journal of Medical Genetics 45 (7) (2008) 411–419.
[21] W. Namkung, K.H. Kim, M.G. Lee, Gastroenterology 129 (6) (2005) 1979–1990.[22] K. Weixel, N.A. Bradbury, Methods in Molecular Medicine 70 (2002) 323–340.[23] J.R. Chen, B.H. Chang, J.E. Allen, M.A. Stiffler, G. MacBeath, Nature Biotechnology
26 (9) (2008) 1041–1045.[24] Z. Wang, S. Petrovic, E. Mann, M. Soleimani, American Journal of Physiology -
Gastrointestinal and Liver Physiology 282 (3) (2002) G573–G579.[25] V.M. Rajendran, J. Black, T.A. Ardito, P. Sangan, S.L. Alper, C. Schweinfest, M.
Kashgarian, H.J. Binder, American Journal of Physiology - Gastrointestinal andLiver Physiology 279 (5) (2000) G931–G942.
and NHERF4 were overexpressed in HeLa cells for 36 h, a cell-permeable phosphatasexamine the cell lysates by using the anti-Myc antibody. B. SLC26A3 and NHERF4 wereERF4 by treatment with cycloheximide (50 μg/mL for 60 min), calyculin A (10 nM forino acid sequences of each NHERF family PDZ domain were aligned using Vector NTImain. A serine residue of NHERF4-PDZ3 is located on the second α-helix in the sameZ domains is shown (grey box, bottom). D. WT-, S329A-, or S329D-NHERF4 was over-ng anti-Myc antibody. *Pb0.05.

1830 J.H. Lee et al. / Cellular Signalling 24 (2012) 1821–1830
[26] A.N. Charney, R.W. Egnor, D. Henner, H. Rashid, N. Cassai, G.S. Sidhu, AmericanJournal of Physiology - Cell Physiology 286 (5) (2004) C1062–C1070.
[27] M.N. Chernova, L. Jiang, B.E. Shmukler, C.W. Schweinfest, P. Blanco, S.D. Freedman,A.K. Stewart, S.L. Alper, The Journal of Physiology 549 (Pt 1) (2003) 3–19.
[28] C. Barmeyer, J.H. Ye, S. Sidani, J. Geibel, H.J. Binder, V.M. Rajendran, PflügersArchiv 454 (3) (2007) 441–450.
[29] P. Hegyi, Z. Rakonczay Jr., L. Tiszlavicz, A. Varro, A. Toth, G. Racz, G. Varga, M.A. Gray,B.E. Argent, Novartis Foundation Symposium 273 (2006) 164–173 (discussion173–166, 261–164).
[30] A.K. Stewart, B.E. Shmukler, D.H. Vandorpe, F. Reimold, J.F. Heneghan, M. Nakakuki,A. Akhavein, S. Ko, H. Ishiguro, S.L. Alper. Am J Physiol Cell Physiol; 301(2):C289-303.
[31] R.W. Freel, M.Morozumi, M. Hatch, American Journal of Physiology - Gastrointestinaland Liver Physiology 297 (5) (2009) G918–G929.
[32] T. Nakamura, N. Shibata, T. Nishimoto-Shibata, D. Feng, M. Ikemoto, K. Motojima,O.N. Iso, K. Tsukamoto, M. Tsujimoto, H. Arai, Proceedings of the NationalAcademy of Sciences of the United States of America 102 (38) (2005)13404–13409.
[33] E.J. Weinman, R.S. Biswas, G. Peng, L. Shen, E.X. Turner CL, D. Steplock, S. Shenolikar,R. Cunningham, The Journal of Clinical Investigation 117 (11) (2007) 3412–3420.
[34] J.W. Voltz, M. Brush, S. Sikes, D. Steplock, E.J. Weinman, S. Shenolikar, Journal ofBiological Chemistry 282 (46) (2007) 33879–33887.
[35] E.J. Weinman, D. Steplock, Y. Zhang, R. Biswas, R.J. Bloch, S. Shenolikar, Journal ofBiological Chemistry 285 (33) (2010) 25134–25138.
[36] H.Y. Gee, S.H. Noh, B.L. Tang, K.H. Kim, M.G. Lee, Cell 146 (5) (2011) 746–760.[37] G. Lamprecht, A. Heil, S. Baisch, E. Lin-Wu, C.C. Yun, H. Kalbacher, M. Gregor, U.
Seidler, Biochemistry 41 (41) (2002) 12336–12342.[38] S. Petrovic, S. Barone, J. Xu, L. Conforti, L. Ma, M. Kujala, J. Kere, M. Soleimani,
American Journal of Physiology - Renal Physiology 286 (1) (2004) F161–F169.[39] T.H. Kwon, A. Pushkin,N. Abuladze, S. Nielsen, I. Kurtz, American Journal of Physiology
- Renal Physiology 278 (2) (2000) F327–F336.[40] N.C. Zachos, C. Hodson, O. Kovbasnjuk, X. Li, W.R. Thelin, B. Cha, S. Milgram, M.
Donowitz, Cellular Physiology and Biochemistry 22 (5–6) (2008) 693–704.[41] D.F. Markgraf, K. Peplowska, C. Ungermann, FEBS Letters 581 (11) (2007)
2125–2130.[42] H.Y. Gee, B.L. Tang, K.H. Kim, M.G. Lee, Journal of Biological Chemistry 285 (46)
(2010) 35519–35527.[43] F.Y. Teng, Y. Wang, B.L. Tang, Genome Biology 2 (11) (2001) (REVIEWS3012).[44] D.P. LaLonde, A. Bretscher, Biochemistry 48 (10) (2009) 2261–2271.





























