Embed Size (px)
Citation preview

Eur. J . Biochem. 107, 297-301 (1980) 8 by FEBS 1980
Regulatory Effects of Iodide and Thiocyanate on Tyrosine Oxidation Catalyzed by Thyroid Peroxidase
Jean-Luc MTCHOT, Jeannine OSTY, and Jacques NUNEZ
Unite de Recherche sur la Glande Thyroide et la Regulation Hormonale, Institut National de la Santk et de la Recherche Medicale, Bicetre
(Received January 24, 1980)
The effects of iodide, thiocyanate and perchlorate, three anions with the same molecular size, on the oxidation of tyrosine to 3,3'-bityrosine by several peroxidases were evaluated at pH 8.8, i.e. in conditions in which iodide is not oxidized. The following results were obtained:
1. Iodide greatly stimulates the rate of bityrosine formation in the presence of thyroid per- oxidase. No effect was seen with horseradish peroxidase or lactoperoxidase. Maximal iodide effects were obtained with about 0.5 mM iodide and K, for iodide was equal to about 0.028 mM. These results suggest that thyroid peroxidase contains a simple class of regulatory binding sites for iodide.
2. SCN- mimics iodide effects; maximal stimulatory effects were seen with about 0.5 mM thiocyanate and K,,, for SCN- was equal to 0.1 mM. The effects of SCN- and those of iodide were not additive. These results suggest that SCN- binds to the same regulatory site as iodide but with a slightly lower affinity. No effect of SCN- was seen with horseradish peroxidase or lacto- peroxidase.
3. (2104, another anion with the same molecular size as iodide and SCN-, had neither an effect on the oxidation of tyrosine to bityrosine nor did it prevent the stimulatory effect of iodide on this reaction. Bromide was without effect on the same reaction.
Thyroid peroxidase [l, 21 is the enzyme which ca- talyzes, in the thyroid gland, both the iodination of several tyrosine residues of thyroglobulin and the coupling of some of them to thyroid hormones [3,4]. This enzyme is therefore unique in the sense it performs successively two reactions; previous results [5,6] have shown that the coupling reaction begins only when the rate of the iodination reaction levels off. Using this observation it has been possible to dissociate these two steps of thyroid hormone synthesis and to show that iodide, in addition to being a substrate for the iodination step, is also required for the coupling reac- tion to occur [6].
Iodide is also required [7 - 11 ] for another reac- tion catalyzed by the peroxidases, i.e. the oxidation of thioureylene derivatives which are widely used as antithyroid drugs. These and other data [lo- 121 sug- gest that thyroid peroxidase contains a regulatory site for iodide, i.e. a site distinct from the substrate site which is involved in the catalysis of the iodination reaction. To study further this hypothetical site the
Etzzymes. Thyroid peroxidase (EC 1.1 1.1.8); lactoperoxidase and horseradish peroxidase (EC 3.1 3.1.7); glucose oxidase (EC 1.1.3.4).
effect of iodide was evaluated on the oxidation of Nu-acetyltyrosinamide to its bis derivative. This reac- tion occurs at an alkaline pH [10,13]: in these con- ditions no oxidation products of iodide can be de- tected; in addition tyrosine iodination could not be measured. Thus the possible regulatory role of iodide may be demonstrated without interference with the substrate site.
MATERIALS AND METHODS
Muterials
Lactoperoxidase and N"-acetyltyrosinamide were purchased from Sigma Chemical Co. ; horseradish per- oxidase from Boehringer (Mannheim) ; guaiacol and iodide were from Prolabo and thiocyanate from Riedel de Haen. Hog thyroid peroxidase was prepared as previously described [2].
Methods
The oxidation of N"-acetyltyrosinamide to 3,3'-bis- (Nu-acetyltyrosinamide) was followed by spectro- fluorimetry using Perkin Elmer MPF 43A equipment.
298 Iodide and SCN- Effects on Tyrosine Oxidation by Thyroid Peroxidase
1 2 3 4 5 0 0
lVa- Acetyltyrosinamide (mM)
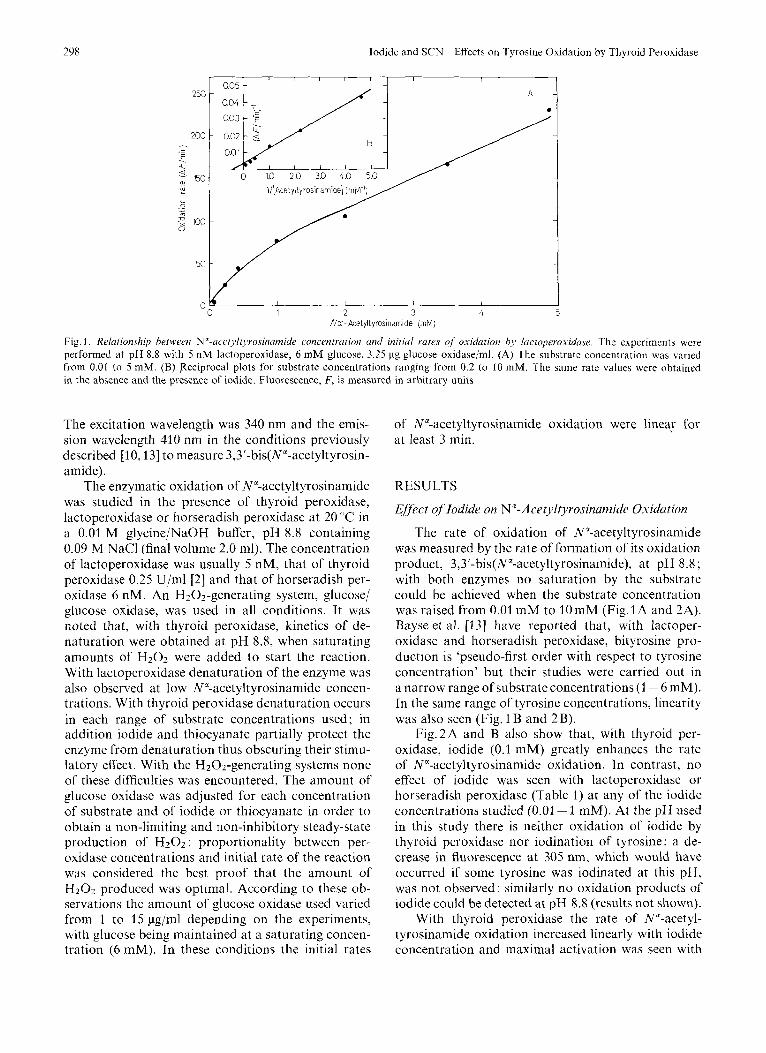
Fig. 1. Relutioizship bet+veen N~-ucetyl[~Jrosinamide concentrutioii und initial r u m of midation by lortoperoxiduse. The experiments were performed at pH 8.8 with 5 n M lactoperoxidase, 6 mM glucose, 3.25 pg glucose oxidase/ml. (A) The substrate concentration was varied from 0.01 to 5 niM. (B) Reciprocal plots for substrate concentrations ranging from 0.2 to 10 mM. The same rate values were obtained in the absence and the presence of iodide, Fluorescence, F, is measurcd in arbitrary units
The excitation wavelength was 340 nm and the emis- sion wavelength 410 nm in the conditions previously described [lo, 131 to measure 3,3'-bis(N"-acetyltyrosin- amide).
The enzymatic oxidation of N"-acetyltyrosinamide was studied in the presence of thyroid peroxidase, lactoperoxidase or horseradish peroxidase at 20 "C in a 0.01 M glycine/NaOH buffer, pH 8.8 containing 0.09 M NaCl (final volume 2.0 ml). The concentration of lactoperoxidase was usually 5 nM, that of thyroid peroxidase 0.25 U/ml [2] and that of horseradish per- oxidase 6 nM. An H20z-generating system, glucose/ glucose oxidase, was used in all conditions. It was noted that, with thyroid peroxidase, kinetics of de- naturation were obtained at pH 8.8, when saturating amounts of Hz02 were added to start the reaction. With lactoperoxidase denaturation of the enzyme was also observed at low Nu-acetyltyrosinamide concen- trations. With thyroid peroxidase denaturation occurs in each range of substrate concentrations used; in addition iodide and thiocyanate partially protect the enzyme from denaturation thus obscuring their stimu- latory effect. With the HaO2-generating systems none of these difficulties was encountered. The amount of glucose oxidase was adjusted for each concentration of substrate and of iodide or thiocyanate in order to obtain a non-limiting and non-inhibitory steady-state production of Hz02 : proportionality between per- oxidase concentrations and initial rate of the reaction was considered the best proof that the amount of HzOz produced was optimal. According to these ob- servations the amount of glucose oxidase used varied from 1 to 15 pg/ml depending on the experiments, with glucose being maintained at a saturating concen- tration (6 mM). In these conditions the initial rates
of N"-acetyltyrosinamide oxidation were linear for at least 3 min.
RESULTS
Ejject of Iodide on N"-Acetyltyl-osinumicl'e Osidcztinn
The rate of oxidation of N"-acetyltyrosinamide was measured by the rate of formation of its oxidation product, 3,3'-bis(N"-acetyltyrosinamide), at pH 8.8 ; with both enzymes no saturation by the substrate could be achieved when the substrate concentration was raised from 0.01 mM to 10mM (Fig.lA and 2A). Bayse et al. [I31 have reported that, with lactoper- oxidase and horseradish peroxidase, bityrosine pro- duction is 'pseudo-first order with respect to tyrosine concentration' but their studies were carried out in a narrow range of substrate concentrations (1 - 6 mM). In the same range of tyrosine concentrations, linearity was also seen (Fig. 1 B and 2 B).
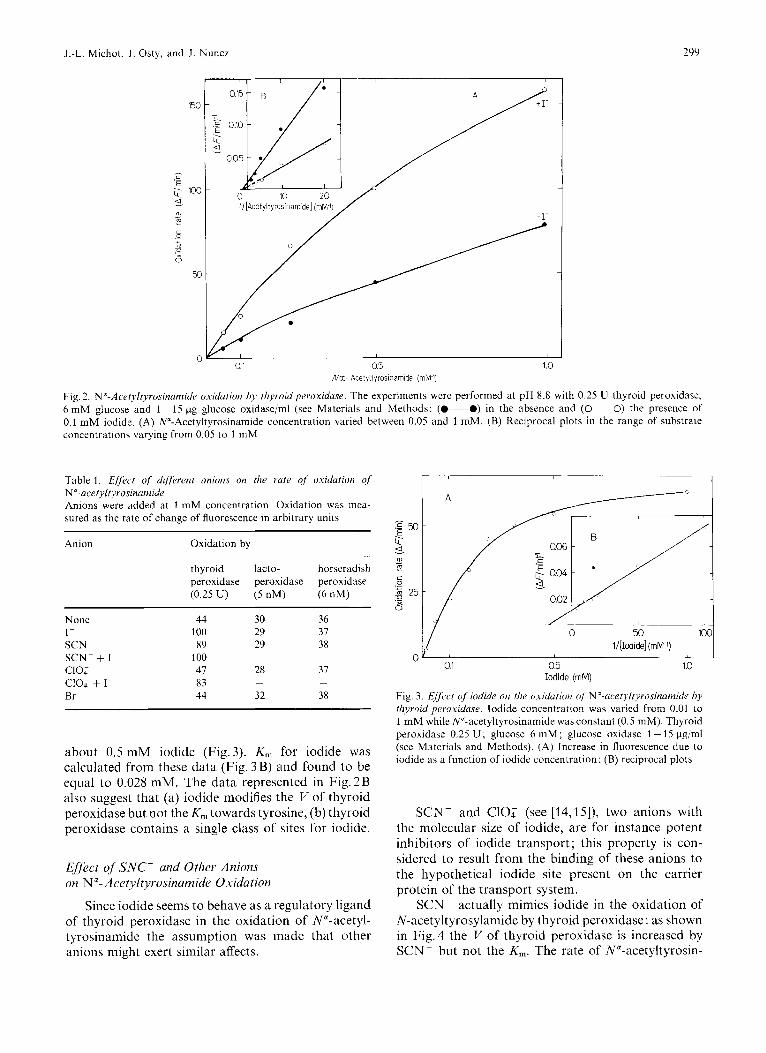
Fig.2A and B also show that, with thyroid per- oxidase, iodide (0.1 mM) greatly enhances the rate of N"-acetyltyrosinamide oxidation. In contrast, no effect of iodide was seen with lactoperoxidase or horseradish peroxidase (Table 1) at any of the iodide concentrations studied (0.01 - 1 mM). At the pH used in this study there is neither oxidation of iodide by thyroid peroxidase nor iodination of tyrosine: a de- crease in fluorescence at 305 nm, which would have occurred if some tyrosine was iodinated at this pH, was not observed; similarly no oxidation products of iodide could be detected at pH 8.8 (results not shown).
With thyroid peroxidase the rate of N"-acetyl- tyrosinamide oxidation increased linearly with iodide concentration and maximal activation was seen with
J.-L. Michot, J . Osty, and J . Nunez 299
Fig. 2. N'-Acel),ityrosinumide oxidation by thyroid peroxiduse. The experiments were performed at pH 8.8 with 0.25 U thyroid peroxidase, 6 m M glucose and 1 ~ 15 pg glucose oxidasejml (see Materials and Methods; (0 -0) in the absence and ( 0 0 ) the presence of 0.1 mM iodide. (A) N"-Acetyltyrosinamide concentration varied between 0.05 and 1 mM. (B) Reciprocal plots in the range of substrate concentrations varying from 0.05 to 1 mM
Table 1. Effect of' different unions on the rule qf oxidutiun qf N"-acef~ltyrosinumide Anions were added at 1 m M concentration. Oxidation was mea- sured as the rate of change of fluorescence in arbitrary units
Anion Oxidation by
thyroid lacto- horseradish peroxidase peroxidase peroxidase (0.25 U) ( 5 nM) (6 nM)
None 44 30 36 1- 100 29 37 SCN 89 29 3x SCN- + I - 100 cloy 47 28 37 (2104 + I - 83 Br 44 32 38
- -
- -
about 0.5 mM iodide (Fig.3). K, for iodide was calculated from these data (Fig. 3B) and found to be equal to 0.028 mM. The data represented in Fig.2B also suggest that (a) iodide modifies the I/ of thyroid peroxidase but not the K , towards tyrosine, (b) thyroid peroxidase contains a single class of sites for iodide.
Ejyect o j SNC- and Other Anions on Nu-Acetyltyrosinamide Oxidation
Since iodide seems to behave as a regulatory ligand of thyroid peroxidase in the oxidation of N"-acetyl- tyrosinainide the assumption was made that other anions might exert similar affects.
" 0.1 0.5
Iodide (rnM) 10
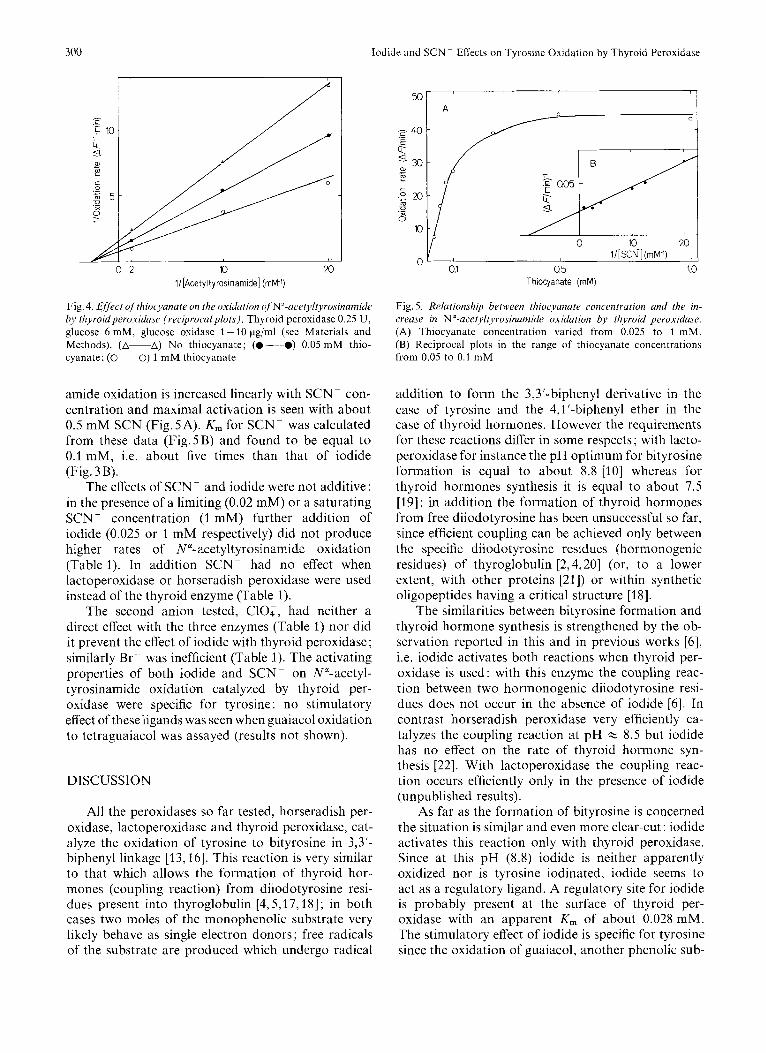
Fig. 3. fifiect of iodide on the o.iidutioti of N"-ucetyltyrosinumiu'L. hy thyroid peroxiduse. Iodide concentration was varied from 0.01 to 1 m M while Nu-acetyllyrosinamide was constant (0.5 mM). Thyroid peroxidase 0.25 U; glucose 6 m M ; glucose oxidase 1-15 pg/ml (see Materials and Methods). (A) Increase in fluorescence due to iodide as a function of iodide concentration; (B) reciprocal plots
SCN- and C104 (see [14,15]), two anions with the molecular size of iodide, are for instance potent inhibitors of iodide transport; this property is con- sidered to result from the binding of these anions to the hypothetical iodide site present on the carrier protein of the transport system.
SCN- actually mimics iodide in the oxidation of N-acetyltyrosylamide by thyroid peroxidase : as shown in Fig. 4 the V' of thyroid peroxidase is increascd by SCN- but not the K,. The rate of N"-acetyltyrosin-
Iodide and SCN ~ Effects on Tyrosine Oxidation by Thyroid Peroxidase 300
I ._ E 10 li 9
P
5 5
0,
aJ c
c 0 ._ 0
-
A 0 2 10 m
I / [Acetylt yrosinamide] (mM-1)
Fig. 4. Effect qftkiocyanate on the oxidation of'N'-ucetyltyrosinami~e by thyroidperoxidase (recipruculpluts). Thyroid peroxidase 0.25 U, glucose 6 mM, glucose oxidase 1 - 10 &ml (see Materials and Methods). (A-A) No thiocyanate; (0 ----a) 0.05 mM thio- cyanate; (0- 0) 1 mM thiocyanate
amide oxidation is increased linearly with SCN- con- centration and maximal activation is seen with about 0.5 mM SCN (Fig. 5A). K, for SCN- was calculated from these data (Fig.SB) and found to be equal to 0.1 mM, i.e. about five times than that of iodide (Fig. 3B).
The effects of SCN- and iodide were not additive : in the presence of a limiting (0.02 mM) or a saturating SCN- concentration (1 mM) further addition of iodide (0.025 or 1 mM respectively) did not produce higher rates of N"-acetyltyrosinamide oxidation (Table 1). In addition SCN- had no effect when lactoperoxidase or horseradish peroxidase were used instead of the thyroid enzyme (Table 1).
The second anion tested, ClOZ, had neither a direct effect with the three enzymes (Table 1) nor did it prevent the effect of iodide with thyroid peroxidase ; similarly Br- was inefficient (Table 1). The activating properties of both iodide and SCN- on N"-acetyl- tyrosinamide oxidation catalyzed by thyroid per- oxidase were specific for tyrosine: no stimulatory effect of these ligands was seen when guaiacol oxidation to tetraguaiacol was assayed (results not shown).
DISCUSSION
All the peroxidases so far tested, horseradish per- oxidase, lactoperoxidase and thyroid peroxidase, cat- alyze the oxidation of tyrosine to bityrosine in 3,3'- biphenyl linkage [13,16]. This reaction is very similar to that which allows the formation of thyroid hor- mones (coupling reaction) from diiodotyrosine resi- dues present into thyroglobulin [4, S,17,18]; in both cases two moles of the monophenolic substrate very likely behave as single electron donors; free radicals of the substrate are produced which undergo radical
0.1 0.5 Thiocyanate (mM)
1 .o
Fig. 5 . Relationship between thiucyanate roncentrution and tl7e in- crease in N"-acetyltyrosinumide oxidation by thyroid perosidase. (A) Thiocyanate concentration varied from 0.025 to 1 mM. (B) Reciprocal plots in the range of thiocyanate concentrations from 0.05 to 0.1 mM
addition to form the 3,3'-biphenyl derivative in the case of tyrosine and the 4,1'-biphenyl ether in the case of thyroid hormones. However the requirements for these reactions differ in some respects; with lacto- peroxidase for instance the pH optimum for bityrosine formation is equal to about 8.8 [lo] whereas for thyroid hormones synthesis it is equal to about 7.5 [19]; in addition the formation of thyroid hormones from free diiodotyrosine has been unsuccessful so far, since efficient coupling can be achieved only between the specific diiodotyrosine residues (hormonogenic residues) of thyroglobulin [2,4,20] (or, to a lower extent, with other proteins [21]) or within synthetic oligopeptides having a critical structure [18].
The similarities between bityrosine formation and thyroid hormone synthesis is strengthened by the ob- servation reported in this and in previous works [6], i.e. iodide activates both reactions when thyroid per- oxidase is used: with this enzyme the coupling reac- tion between two hormonogenic diiodotyrosine resi- dues does not occur in the absence of iodide [6]. In contrast horseradish peroxidase very efficiently ca- talyzes the coupling reaction at pH z 8.5 but iodide has no effect on the rate of thyroid hormone syn- thesis [22]. With lactoperoxidase the coupling reac- tion occurs efficiently only in the presence of iodide (unpublished results).
As far as the formation of bityrosine is concerned the situation is similar and even more clear-cut: iodide activates this reaction only with thyroid peroxidase. Since at this pH (8.8) iodide is neither apparently oxidized nor is tyrosine iodinated, iodide seems to act as a regulatory ligand. A regulatory site for iodide is probably present at the surface of thyroid per- oxidase with an apparent K , of about 0.028 mM. The stimulatory effect of iodide is specific for tyrosine since the oxidation of guaiacol, another phenolic sub-

J.-L. Michot, J. Osty, and J. Nunez 301
strate of thyroid peroxidase, is not stimulated by iodide.
lodide has also been shown to play an important role in the mode of action of the antithyroid drugs [7 - 121 ; in fact many of these compounds cannot be oxidized by peroxidases in the absence of iodide. Iodide is bound to lactoperoxidase with a dissociation constant of 0.02 mM and is not consumed during the oxidation of the antithyroid drugs or various goitro- gens [lo]. However this effect of iodide is seen not only with thyroid peroxidase (unpublished results) but also with lactoperoxidase [lo- 121. Recent results [12] show that the effect of iodide actually depends on the drug used: with the antithyroid drugs of the thiourea group no effect of iodide was seen with lacto- peroxidase whereas the oxidation of methylthiouracil and thiobarbiturate is markedly stimulated by iodide. The assumption was made therefore that the rate of drug oxidation depends on the geometry of the ternary complex formed by iodide, the drug and the enzyme. This conclusion might also explain why iodide has a clear effect on the oxidation of tyrosine by thyroid peroxidase but not on that of guaiacol. Similarly iodide should modify the properties of lactoperoxidase in the oxidation of some antithyroid drugs but not in that of tyrosine.
The mechanism of action of iodide in the activation of thyroid peroxidase was further analyzed by using two other anions with the same molecular size as iodide. SCN- and C104 are both used as inhibitors of iodide transport in the thyroid gland [16,17]. SCN- differs however in several respects from C104 since in addition to inhibiting iodide transport it is a com- petitive substrate for thyroid peroxidase at pH z 7.0. Actually SCN inhibits tyrosine iodination and iodide oxidation at pH 7.0 with all the peroxidases so far tested (unpublished results).
At pH 8.8 SCN- behaves in the opposite direction and mimics very efficiently the stimulatory effect of iodide on tyrosine oxidation (Fig.4 and 5 ) with an apparent K,,, of 0.1 mM. In contrast C104 has no effect on tyrosine oxidation and does not prevent the stimulatory effect of iodide in this reaction. It is interesting to note that SCN-, but not Clog, also mimics efficiently the iodide effect on the coupling reaction catalyzed by thyroid peroxidase (unpublished rcsults). Actually SCN- and iodide might bind to the
same regulatory site at the surface of the enzyme as suggested by the similarity in their effects and because the stimulation of tyrosine oxidation produced by the ligands is not additive. The reaction of tyrosine oxidation to 3,3’-bityrosine appears to be a good model for the coupling reaction: in both cases iodide (and SCN-) stimulates the reaction and this effect is specific for the thyroid enzyme.
REFERENCES
1. Coval, M. L. & Taurog, A. (1967) J . Biol. Cliem. 242, 5510-
2.
3.
4.
5.
6.
7.
8.
9. 10.
11 .
12.
13.
14. 15.
16.
17.
18.
19.
20.
21. 22.
5523.
(Paris) 54, 483 - 492.
J . Biockem. 51, 329-336.
Biorliem. Biopkys. 139,221 -229.
Pommier, J., De Prailaunk, S. & Nunez, J. (1972) Biochirnie
Deme, D., Fimiani, E., Pommier, J. & Nunez, J. (,1975) Eur.
Taurog, A,, Lothrop, M. L. & Eastabrook, R . W. (1970) Arch.
Pommier, J., Dkme, D. & Nunez, J . (1973) Eur. J . Biochem. 37,406-414.
Deme, D., Pommier, J. & Nunez, J. (1976) Eur. J . Biochem. 70, 435 - 440.
Maloof, F. & Soodak, M. (1965) in Current Topics in Thyroid Research (Cassano, C. & Andreoli, M., eds) pp. 277-290, Academic Press, New York.
Morris, D. R. & Hager, L. P. (1966) J . B i d . Cl7em. 241,
Taurog, A. (1976) Endocrinology, Y8, 1031 -1046. Michot, J. L., Nunez, J., Johnson, M. L., Irace, G . & Edel-
hoch, H. (1 978) J . Biol. Cliem. 254, 2205 - 2209. Davidson, R., Soodak. M. , Strout, H. V., Neary, J. T., Naka-
mum, C. & Maloof, F. (1979) Endocrinology, 104, 919-924. Edelhoch, H., Irace, G. , Johnson, M. L., Michot, J. L. &
Nunez. J. (1980) J . Bid . Chrm. 254, 11822-11830. Bayse, G. S., Michaels, A. W. & Morrison, M. (1972) Bioc/iim.
Bio}~hys. A(,tu, 284, 34-42. Wolff, J. (1964) Physiof. Rev. 44, 45-90. Wolff, J . & Maurey, J . R. (1963) Biocliim. Bioplzys. Actu. 69,
Gross, A. J . & Sizer, I. W. (1959) J . Biol. Chem. 234, 1611-
Lamas, L., Dorris, M. L. & Taurog, A. (1972) Endorrimhgy,
Cahnrnann, H. J., Pommier, J. & Nune7, J . (1977) Proi.. Nai/
Deme, D., Pommier, J . & Nunez, J. (1978) Biochim. Biophys.
Deme, D., Gavaret, J. M., Pomniier, J . & Nunez, J. (1976)
Taurog, A. (1970) Rec,ent Prog. Horm. Re.7. 26, 189-241. Virion, A, , Pommier, J . & Nunez, J . (1979) Eur. J . Biocliem.
3582- 3589.
58 - 67.
1614.
90, 1417-1426.
Acutl. Sci. U . S . A . 74, 5333-5335.
Actu, 540, 73-82.
Eur, J . Biocheni. 70, 7- 13.
102. 549 - 554.
J.-L. Michot, J. Osty, and J . Nunez, Unit6 de Kecherches sur la Glande Thyroide et la Regulation Hormonale (Unite 96 de I’lnstitut National de la Sante et de la Recherche Medicale), HBpital-Hospice de Bicsfre, 78 Avenue du General-Leclercq, F-94270 Le Kremlin-Bicitre, Francc





























