Embed Size (px)
Citation preview

Research ArticleInflammation and ER Stress Downregulate BDH2 Expressionand Dysregulate Intracellular Iron in Macrophages
Susu M Zughaier1 Brandon B Stauffer2 and Nael A McCarty2
1 Department of Microbiology and Immunology Veterans Affairs Medical Center (VAMC (151-I)) Emory University School ofMedicine and Childrenrsquos Healthcare of Atlanta 1670 Clairmont Road Atlanta GA 30033 USA
2 Emory-Childrenrsquos Center for Cystic Fibrosis and Airways Disease Research Division of Pulmonology AllergyImmunologyCystic Fibrosis and Sleep Department of Pediatrics Emory University School of Medicine and Childrenrsquos Healthcare of AtlantaAtlanta GA 30322 USA
Correspondence should be addressed to Susu M Zughaier szughaiemoryedu
Received 2 September 2014 Revised 13 November 2014 Accepted 13 November 2014 Published 1 December 2014
Academic Editor Xiao-Feng Yang
Copyright copy 2014 Susu M Zughaier et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Macrophages play a very important role in host defense and in iron homeostasis by engulfing senescent red blood cells and recyclingiron Hepcidin is the master iron regulating hormone that limits dietary iron absorption from the gut and limits iron egress frommacrophages Upon infection macrophages retain iron to limit its bioavailability which limits bacterial growth Recently a shortchain butyrate dehydrogenase type 2 (BDH2) protein was reported to contain an iron responsive element and to mediate cellulariron trafficking by catalyzing the synthesis of themammalian siderophore that binds labile iron therefore BDH2 plays a crucial rolein intracellular iron homeostasis However BDH2 expression and regulation in macrophages have not yet been described Here weshow that LPS-induced inflammation combined with ER stress led to massive BDH2 downregulation increased the expression ofER stress markers upregulated hepcidin expression downregulated ferroportin expression caused iron retention in macrophagesand dysregulated cytokine release from macrophages We also show that ER stress combined with inflammation synergisticallyupregulated the expression of the iron carrier protein NGAL and the stress-inducible heme degrading enzyme heme oxygenase-1(HO-1) leading to iron liberation This is the first report to show that inflammation and ER stress downregulate the expression ofBDH2 in human THP-1 macrophages
1 Introduction
Iron is an essential nutrient required for numerous cell func-tions and in order to avoid metal toxicity iron metabolismis tightly regulated [1] Altered iron homeostasis and dys-regulated cellular iron metabolism are associated with manychronic diseases including diabetes mellitus cardiovasculardiseases arthritis and neurodegenerative diseases Alterediron homeostasis in chronic inflammation is characterized bymisdistribution of iron that is manifested in hypoferremiacommonly called anemia of chronic inflammation and ironretention in macrophages and the reticuloendothelial system[2 3]
Macrophages play a central role in iron homeostasis byuptake of damaged and senescent red blood cells and recycle
large amounts of iron daily [4 5] Macrophages also play avery important role in host defense by detecting invadingpathogens and responding in order to clear infection Sinceiron is essential for bacterial growth macrophages retainiron to limit its extracellular bioavailability which therebylimits bacterial growthThis response to infection is called theiron-limiting innate immune defense mechanism Hepcidinis the master iron regulating hormone that limits iron egressfrom macrophages [6] Hepcidin binds to ferroportin theonly known iron exporter leading to its internalization anddegradation and consequent iron retention [7] Thereforecellular iron homeostasis and iron egress via the hepcidin-ferroportin axis are very tightly regulated during infectionand inflammation [8ndash11]
Hindawi Publishing CorporationJournal of Immunology ResearchVolume 2014 Article ID 140728 16 pageshttpdxdoiorg1011552014140728
2 Journal of Immunology Research
Iron is a highly reactive metal thus to reduce metaltoxicity iron is maintained bound to hemoproteins and iniron-sulfur clusters or safely stored in ferritin cages [1]Free intracellular iron or poorly liganded iron is toxicsince it catalyzes the production of reactive oxygen species(ROS) Therefore macrophages detoxify labile iron by mul-tiple mechanisms Recently the presence of a mammaliansiderophore an iron-chelating small molecule was reportedand the iron-binding moiety of this molecule was identifiedas 25-dihydroxybenzoic acid (25DHBA) [12] This mam-malian siderophore captures free cytosolic iron and thusprotects the cells from oxidant stress [12] The enzyme thatcatalyzes the synthesis of 25DHBA is a member of the short-chain dehydrogenasereductase family (DHRS6) [13] called3-hydroxybutyrate dehydrogenase type 2 (BDH2) [12] Inhi-bition of BDH2 expression inmurine pro-B lymphocytic cellsled to the depletion of the mammalian siderophore and theaccumulation of labile ironThephysiological role of BDH2 insensing cellular iron was further confirmed by the discoverythat the BDH2 enzyme contains an iron responsive elementthat interacts with iron regulatory proteins that controlposttranscriptional gene expression [14] Recently Zughaieret al reported that BDH2 expression in macrophages issuppressed upon bacterial infection suggesting an importantrole for BDH2 in iron-limiting innate immunity [15] Morerecently the Devireddy group further confirmed the role ofBDH2 in cellular iron homeostasis and showed that depletionof the endogenous siderophore 25-DHBA in a mouse modelresulted in microcytic anemia and iron overload in thespleen [16 17] They also reported that BDH2 and ferritin-H synergistically regulate intracellular iron levels in an invitro experimental cell culture model [18] Therefore BDH2is required for intracellular iron homeostasis [12] Howeverthe role of BDH2 in iron-limiting innate immune defenses inmacrophages is not known
Macrophages engulf senescent RBCs and recycle hemo-globin thus they express heme oxygenase-1 (HO-1) thecatalytic enzyme that degrades heme consequently liberatingiron and producing carbon monoxide [19] HO-1 is highlyinduced in response to inflammation oxidative stress andhypoxia [19 20] due to its cytoprotective effects and anti-inflammatory activity [21 22] Therefore HO-1 as a stress-inducible protein is an important player in intracellulariron homeostasis Further neutrophil gelatinase-associatedlipocalin (NGAL) [23] functions as an iron carrier proteinthat shuttles bound iron across tissues by internalizationwith a membrane-localized receptor [24 25] and also playsa critical physiological role in intracellular iron homeostasisby trafficking the iron-bound siderophore 25DHBA [12] andiron-catecholamine complexes [26] NGAL plays a crucialrole in iron-limiting innate immune defenses since it limitsiron availability by also scavenging bacterial siderophores andthereby exerts an antibacterial function [27ndash29]
Intracellular iron homeostasis is disrupted by endoplas-mic reticulum (ER) stress [30] ER stress induced upon cellu-lar perturbations is regulated by activation of the unfoldedprotein response to restore homeostatic cellular functions[30] Control of ER stress is required for cell survival anduncontrolled ER stress leads to cell death [31 32] ER
stress alters cellular iron homeostasis by inducing hepcidinexpression [33 34] Altered iron homeostasis and ER stressare associated with chronic neurodegenerative diseases suchas Huntingtonrsquos disease and Alzheimerrsquos disease [35 36] It isnot known whether ER stress also affects BDH2 expressionThe goal of this studywas to investigate whether ER stress andinflammation modulate BDH2 expression in macrophages
Here we show that LPS-induced inflammation combinedwith chemically induced ER stress led to BDH2 downregula-tion increased the expression of ER stress markers includingCHOP upregulated hepcidin expression downregulated fer-roportin expression caused iron retention in macrophagesand dysregulated cytokine release from macrophages Fur-ther we show that ER stress combined with inflammationdramatically upregulated NGAL expression as well as theinducible hemeoxygenase-1 enzyme inmacrophagesWe alsoshow that vitamin D treatment prior to LPS restored BDH2expression This is the first report to show that ER stressand inflammation downregulate the expression of BDH2 inhuman THP-1 macrophages The results suggest that BDH2plays an important role in modulating iron-limiting innateimmune defenses
2 Materials and Methods
21 Reagents RPMI 1640 medium Dulbeccorsquos modifiedEagle medium (D-MEM) HBSS fetal bovine serum (FBS)penicillinstreptomycin sodium pyruvate and nonessentialamino acids were obtained fromCellgroMediatech (HerdonVA) Magnetic beads for a panel of human cytokines andchemokines were from Invitrogen (Carlsbad CA) Tuni-camycin calcein-AM and thiazolyl blue tetrazoliumbromidewere purchased from Sigma Aldrich (St Louis MO) Lipo-polysaccharide [37] from Neisseria meningitidis serogroup Bwas purified and quantified as previously described [38]
22 Cell Culture and Macrophage Stimulation THP-1macrophage-like cells were grown in RPMI 1640 mediumsupplemented with 10 FBS 50 120583gmL penicillin and50 IUmL of streptomycin HEK293 cells were grown inD-MEM supplemented with 10 FBS 50 120583gmL penicillinand 50 IUmL of streptomycin Freshly grown cells wereharvested and adjusted to 1 million cellsmL and thentransferred into 12-well tissue culture plates at 2mLwellTo induce ER stress cells were treated with tunicamycin(10 120583gmL in DMSO) and incubated overnight at 37∘CControl cells were treated with DMSO only and incubatedsimilarly Following overnight exposure to tunicamycin cellswere exposed to LPS (40 ngmL) to induce inflammationand incubated for 6 hr at 37∘C Cells were centrifugedand supernatants were saved at minus20∘C for cytokine meas-urements Harvested cells were washed with PBS and thenplaced in RLT buffer (Qiagen Hilden Germany) containing1 120573-mercaptoethanol passed over QiaShredder columnsand the resulting lysates were saved at minus80∘C for mRNAextraction
Journal of Immunology Research 3
23 RNA Isolation Quantitative Real-Time PCR and GeneExpression Analysis RNA was isolated using RNeasy Minikits (Qiagen) following the manufacturerrsquos instructions aspreviously described [39] Briefly cell lysates saved in RLTbuffer were mixed in 70 ethanol and then passed overRNeasy columns Columns were washed and treated with10 120583L of RNase-free DNase (Qiagen) for 15min at roomtemperature prior to RNA extraction followed by addi-tional washing and centrifugation RNA was eluted in 35 120583Lof RNase-free water and then was reverse transcribed tocDNA using QuantiTect Reverse Transcription kit (Qia-gen) following the manufacturerrsquos instructions Relative geneexpression was determined by quantitative RT-PCR per-formed on resulting cDNA using SYBR Green (PromegaMadison WI) following the manufacturerrsquos instructionsThemRNA level was calculated in reference to 120573-actin and foldchange gene expression was calculated in reference tocontrols using the ΔΔCT method Results were normalizedto unstimulated DMSO-treated cells which were used ascontrols for basal gene expression level The followingprimers were used for qRT-PCR reactions human hepci-din 51015840-GACCAGTGGCTCTGTTTTCC-31015840 and 51015840-CAC-ATCCCACACTTTGATCG-31015840 human BDH2 51015840-CAGCGT-CAAAGGAGTTGTGA-31015840 and 51015840-TGGCGTATCAACTGT-TCCTG-31015840 human NGAL 51015840-ATGACATGAACCTGCTCG-ATA-31015840 and 51015840-TCATAGTCGTTCATTATCTTC-31015840 human120573-actin 51015840-TCTTCCAGCCTTCCTTCCT-31015840 and 51015840-AGC-ACTGTGTTGGCGTACAG-31015840 Primers for ER stress mark-ers used in this study are as follows human hsXBP-1 51015840-TTC-CGGAGCTGGGTATCTCA-31015840 and 51015840-GAACCCCCGTAT-CCACAGTC-31015840 human GRP78 51015840-ACGGCAGCTGCT-ATTGCTTA-31015840 and 51015840-TCCTGACATCTTTGCCCGTC-31015840human CHOP 51015840-GACCTGCAAGAGGTCCTGTC-31015840 and51015840-GCAGGGTCAAGAGTGGTGAA-31015840 Ferroportin (HsSLC40A1 1 SG) transferrin receptor (Hs TFRC 1 SG) andheme oxygenase (Hs HMOX1 1 SG) QuantiTect primerswere purchased from Qiagen
24 Western Blot Analysis Freshly grown THP-1 cells wereharvested and adjusted to 1 million cellsmL and 8mL of cellswas transferred into 6-well tissue culture plates To induceER stress cells were treated with tunicamycin (10 120583gmL) andincubated overnight at 37∘C Control cells were treated withDMSO only and incubated similarly Following overnightincubation some cells were exposed to LPS (40 ngmL)to induce inflammation and incubated for 6 hr at 37∘CHarvested THP-1 cells were lysed in RIPA buffer (BostonBioProducts Boston MA) and centrifuged to remove nucleiand debris and the protein concentration of the supernatantwas measured using a BCA protein assay (ThermoWalthamMA) Equal amounts of protein were removed from eachmacrophage treatment group mixed with 6X Laemlli bufferand boiled for 15 minutes For untransfected HEK lysate andthe BDH2 transfected HEK positive control groups (OrigeneTechnologies Rockville MD) 50 ng of protein was loadedto adjust for overexpression Samples were loaded onto a12 MiniProtean TGX gel (BioRad Hercules CA) elec-trophoresed and transferred onto nitrocellulose membranes
(BioRad Hercules CA) To detect BDH2 membranes wereincubated with anti-BDH2 antibody (TrueMAB antibodyclone 2G1 from Origene) To detect actin membranes wereincubated with a mouse anti-120573 actin antibody (Sigma)In both cases membranes were incubated with an HRP-conjugated anti-mouse secondary antibody (Cell SignalingDanvers MA) incubated in Amersham western blot detec-tion reagent (GE Healthcare Cleveland OH) and visualizedwith HyBlot CL autoradiography film (Denville ScientificMetuchen NJ)
25 Cytokine and Chemokine Measurements Cytokines andchemokines released from THP-1 macrophages treated withtunicamycin alone LPS alone or both tunicamycin and LPSas described above were measured using Luminex magneticbeads (Invitrogen) following the manufacturerrsquos instructions
26 Measurement of Iron Retention inMacrophages Intracel-lular accumulation of labile iron in macrophages was mea-sured using the well-established calcein-AM method [40]Freshly grown THP-1 cells were treated with tunicamycin(10 120583gmL) and incubated overnight prior to stimulationwithLPS (40 ngmL) for 6 hr Control cells were treated withDMSO only Harvested cells were resuspended in RPMI1640 medium supplemented with 10 FBS and 05120583Mcalcein-AM was added and incubated for 15min at 37∘CCalcein-loaded cells were washed twice with PBS to removeextracellular calcein-AM and resuspended in HBSS 100120583Laliquots containing 025 million cells were transferred intoquadruplicate wells in black 96-well plates After a 20minincubation at 37∘C fluorescence was determined at 485 nm(excitation) and 535 nm (emission) wavelengths using a Bio-Tek Synergy 2 Instrument (Winooski VT)
27 Cellular Viability Measurement Using the ColorimetricMTT Reduction Assay The tetrazolium salt dye is cleavedby live and metabolically active cells but not by dead cellstherefore the MTT reduction assay was used to measurethe viability of THP-1 cells treated with tunicamycin withor without LPS [41 42] Briefly freshly grown THP-1 cellsadjusted to 1 million cellsmL were treated with tunicamycindoses ranging from 05 to 10 120583gmL and incubated overnightfollowed in some cases by 6 hr of exposure to LPS Controlcells treated with DMSO only were coincubated under thesame experimental conditions Five mg of thiazolyl bluetetrazolium bromide was dissolved in 1mL of PBS and 15 120583Lwas added to 150120583L of treated THP-1 cells whichwere furtherincubated at 37∘C for one hour Cells were centrifuged toremove extracellular dye and then lysed in 150 120583L of DMSOto dissolve the intracellular dye The absorbance of reduceddye was measured at 591 nm wavelength using a Bio-Tekspectrophotometer
28 Statistical Analysis Mean values plusmn SD of three inde-pendent experiments or two in few cases were calculatedwith Microsoft Excel Statistical analysis was performed withPrism v50 software using one-way ANOVA followed byBonferroni multiple comparisons post hoc analysis 119875 values
4 Journal of Immunology Research
37
25
50
75
(kD
)(k
D)
(kD
)
25
50
75
37
37
25
50
(i)
(a)
(b)
(c)
(ii)
(ii)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge
NS
05
06
07
08
09
1
11
0 1 25 5
BDH
2 m
RNA
relat
ive f
old
chan
ge
Vehi
cle
Tuni
cam
ycin
LPS
Tun+
LPS
HEK
(minus)
HEK
+BD
H2
120573-A
ctin
Ant
i-BD
H2
Ant
i-BD
H2
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowast
Tunicamycin dose (120583gmL)
1 2 3 4 5 6
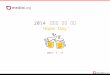
Figure 1 ER stress and inflammation downregulate BDH2 expression in macrophages (a)Western blot analysis of BDH2 protein expressionin THP-1 macrophage-like cells (1 times 106 cellsmL) treated with tunicamycin (10120583gmL) or vehicle (DMSO) overnight prior to LPS exposure(40 ngmL) for 6 hr BDH2 protein was detected with monoclonal anti-BDH2 TrueMAB antibody clone 2G Lane 1 vehicle-treated THP-1cells 2 tunicamycin treated 3 LPS treated 4 tunicamycin and LPS treated 5 untransfected HEK293 cells as negative control 6 BDH2transfected HEK293 cells as positive control (BDH2 positive control contains a FLAG tag and 10-fold higher concentration was loaded inpanel (i) compared to panel (ii) representing two independent experiments) lowastnonspecific band (b) THP-1 macrophages were treated withtunicamycin (10120583gmL) as above and RNAwas extracted and BDH2 gene expression was assessed by quantitative RT-PCR normalized to thatof120573-actin (c) THP-1 cells treatedwith tunicamycin doses 1 25 or 5120583gmL and incubated overnightThe fold change inBDH2gene expressionwas calculated in reference to DMSO treated control THP-1 cells using the ΔΔCTmethod and presented here normalized to vehicle (DMSO)treatment control Error bars represent the SD from the mean of nine readouts generated from three independent experiments lowast119875 valueswere calculated in reference to vehicle (DMSO) treatment control using one-way ANOVA followed by Bonferroni multiple comparisons posthoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005119875 gt 005 were not significant
3 Results
31 BDH2 Expression inMacrophages Is Downregulated by ERStress and Inflammation Macrophages play a very importantrole in host defense and in iron homeostasis [1] RecentlyBDH2 was reported to play a crucial role in intracellulariron trafficking by catalyzing the synthesis of 25-DHBA [12]Further ER stress has been shown to dysregulate cellulariron homeostasis [30] Zughaier et al reported that BDH2expression in macrophages is downregulated upon bacterialinfection [15] However BDH2 expression and regulationin macrophages during ER stress and inflammation havenot yet been described To investigate BDH2 expression in
macrophages and its regulation by ER stress and inflamma-tion we employed human macrophage-like monocytic cells(THP-1) treated with tunicamycin overnight prior to LPSexposure for 6 hr to induce ER stress and inflammationrespectively Here we show that BDH2 protein and geneexpression in macrophages was downregulated by ER stressand inflammation (Figure 1) Reduced BDH2 protein levelswere observed in THP-1 cells after treatment with LPS andtunicamycin as compared to cells treated with DMSO alone(Figure 1(a)) Consistent with this observation BDH2 geneexpression was significantly downregulated when THP-1macrophages were treated with tunicamycin and LPS eitheralone or in combination (Figure 1(b)) Treatment with tuni-camycin alone reduced BDH2 mRNA expression in a dose-dependent manner (Figure 1(c)) Further we examined theactivation of ER stress markers and as expected overnight
Journal of Immunology Research 5
02468
101214161820
No treatment Tunicamycin LPS
CHO
P m
RNA
relat
ive f
old
chan
geNS
NS
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
(a)
0
1
2
3
4
5
6
7
8
No treatment Tunicamycin LPS
hsXB
P m
RNA
relat
ive f
old
chan
ge
NS
NS
lowast
lowast
lowastlowast
lowastlowast
Tunicamycin +LPS
(b)
0
10
20
30
40
50
60
70
No treatment Tunicamycin LPS
NSGRP
78 m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
(c)
Figure 2 ER stress markers are induced in macrophages by tunicamycin THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr RNA was extracted and ER stress markers CHOP (a) hsXBP (b)and GRP78 (c) gene expressions were assessed by quantitative RT-PCR normalized to 120573-actin The fold change in CHOP and hsXBP geneexpression was calculated in reference to DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to notreatment control Error bars represent the SD from the mean of three independent experiments lowast119875 values were calculated in reference to notreatment (DMSO only) control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotatedlowastlowastlowast
119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
tunicamycin treatment significantly induced ER stress mark-ers CHOP hsXBP-1 GRP78 (Figure 2) and ATF6 (data notshown) In contrast 6 hr LPS treatment alone did not upreg-ulate ER stress markers in THP-1 cells (Figure 2) as docu-mented previously [43] Cellular viability was assessed usingtheMTT reduction assay [41] which showed that the doses oftunicamycin and LPS used here did not reduce cellular via-bility (see Supplementary Figure 1A in Supplementary Mate-rial available online at httpdxdoiorg1011552014140728)These data indicate that ER stress and inflammation down-regulate BDH2 expression in THP-1 macrophages
32 Cellular Iron Homeostasis Is Dysregulated by ER Stressand Inflammation in Macrophages ER stress controls ironhomeostasis via the induction of hepcidin the master iron
regulating hormone [2] Hepcidin binds to the only knowncellular iron exporter ferroportin and leads to its internal-ization and degradation consequently inhibiting iron egressfrom macrophages [7] The dysregulation of the hepcidin-ferroportin axis leads to iron retention in macrophagesand increases the cytosolic labile iron pool which is highlyreactive and causes oxidative stress [3] Therefore detox-ifying labile iron is essential to maintaining cellular ironhomeostasis To investigate intracellular iron status in tuni-camycin and LPS treated THP-1 macrophages we determinedthe expression of hepcidin and ferroportin The data showthat tunicamycin-induced ER stress as well as LPS-inducedinflammation led to significant upregulation of hepcidin geneexpression in THP-1 macrophages (Figure 3(a)) In contrasttunicamycin and LPS treatment led to a dramatic reduction
6 Journal of Immunology Research
0
2
4
6
8
10
12
14
16
18
No treatment Tunicamycin LPS
NSNS
Tunicamycin +LPS
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastH
epci
din
mRN
A re
lativ
e fol
d ch
ange
(a)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
FPN
1 m
RNA
relat
ive f
old
chan
ge
NS
NS NS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(b)
Figure 3 ER stress and inflammation dysregulate cellular iron homeostasis THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr ((a) (b)) Hepcidin and ferroportin (FPN1) gene expression wasassessed by quantitative RT-PCR normalized to 120573-actin The fold change in hepcidin and FPN1 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to no treatment control Error bars representthe SD from the mean of three independent experiments 119875 values were calculated in reference to vehicle (DMSO) treatment control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
400000
500000
600000
700000
800000
900000
1000000
No treatment Tunicamycin LPS
NS
Calc
ein-
AM
fluo
resc
ence
(AU
)
lowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Tunicamycin +LPS
Figure 4 ER stress and inflammation induce iron retention in macrophages THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr and iron retention in macrophages was determined using thecalcein-AMfluorescent probemethod [40]DMSO treated cells were incubated simultaneously andused as controls Calcein-AMfluorescencewasmeasured by excitation at 488 nmand emission at 528 nmwavelength (see Section 2) Calcein-AMfluorescence is quenched upon bindingiron and is therefore inversely correlated with intracellular iron accumulation AU arbitrary units Error bars represent the SD from themeanof triplicate reading and data are representative of two independent experiments 119875 values were calculated in reference to vehicle (DMSO)treatment control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001lowastlowast
119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
in ferroportin expression in macrophages (Figure 3(b)) Fer-roportin allows iron egress from macrophages therefore itsdegradation leads to iron retention We measured labile ironaccumulation in macrophages using calcein-AM [40] andfound that tunicamycin and LPS treatment led to significant
iron retention (Figure 4 and Supplementary Figure 1B)A synergistic effect on iron retention was observed whenmacrophages were cotreated with tunicamycin and LPS Theresults suggest that chemically induced ER stress and inflam-mation alter intracellular iron homeostasis in macrophages
Journal of Immunology Research 7
0
500
1000
1500
2000
2500
3000
3500
4000
No treatment Tunicamycin LPS
NG
AL
mRN
A re
lativ
e fol
d ch
ange
123 fold1 fold
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
5 25 05 0
NG
AL
mRN
A re
lativ
e fol
d ch
ange
Tunicamycin
NS
Tunicamycin + LPS
Tunicamycin doses (120583gmL)
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowastlowast
(b)
Figure 5 NGAL gene expression is massively upregulated by ER stress (a) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr (b) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycindoses (5 25 or 05 120583gmL) as in panel (a) or tunicamycin followed by LPS (40 ngmL) RNA was extracted and NGAL gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actinThe fold change in NGAL gene expression was calculated in reference to DMSOtreated control THP-1 cells using theΔΔCTmethod and presented here normalized to vehicle (DMSO) treatment control Error bars representthe SD from the mean of three independent experiments lowast119875 values were calculated in reference to no treatment (DMSO only) control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
33 ER Stress and Inflammation Dramatically Induce NGALGene Expression NGAL plays a dual role in physiologiccellular iron homeostasis and in host defense We recentlyreported that LPS induced NGAL secretion from peripheralmonocytes [44] ER stress is also reported to highly induceNGAL expression [45] However the impact of ER stresscombined with inflammation on NGAL expression is not yetknown To this end we determined NGAL gene expressionin tunicamycin and LPS treated macrophages As expectedLPS treatment led to significant upregulation of NGALgene expression (sim12-fold) but tunicamycin treatment led toa dramatic sim1500-fold increase in NGAL gene expression(Figure 5(a)) Tunicamycin induced NGAL gene expressionin a dose-dependent manner (Figure 5(b)) A synergisticeffect was observed when THP1 cells were cotreated withtunicamycin and LPS which resulted in an approximatelysim3000-fold increase in NGAL gene expression (Figure 5(a))This synergistic effect was tunicamycin dose-dependent(Figure 5(b)) Tunicamycin-induced ER stress led to sim100-fold greater increase in NGAL gene expression comparedto LPS The synergistic cross-talk between tunicamycin-induced ER stress and LPS-induced inflammation led to sim200-fold increase in NGAL gene expression compared toLPS alone These data suggest that chemically induced ERstress dramatically increases NGAL expression more thaninflammation but when combined a synergistic upregulationof NGAL is observed
34 ER Stress and Inflammation Induce Heme Oxygenase-1 (HO-1) Gene Expression HO-1 is a stress-induced anti-inflammatory [19 21 22] cytoprotective enzyme that cat-alyzes the degradation of heme and consequently liber-ates iron and carbon monoxide [20] The released carbonmonoxide suppresses hepcidin expression therefore HO-1plays a role in regulating iron homeostasis [46] Our datademonstrate that ER stress and inflammation dysregulateintracellular iron homeostasis in macrophagesTherefore weinvestigated the impact of ER stress and inflammation onHO-1 expression in macrophages as an anti-inflammatoryimmune response Here we show that HO-1 gene expressionis significantly upregulated in THP-1 cells treated with tuni-camycin or LPS alone (Figure 6) However the combinationof tunicamycin andLPS treatment synergistically upregulatedHO-1 expression in THP-1 cells (Figure 6) suggesting a potentresponse to combat cellular stress
35 Vitamin D Restores BDH2 Expression in LPS-StimulatedMacrophages In this report we show that tunicamycin-induced ER stress and LPS-induced inflammation downreg-ulate BDH2 expression inmacrophages and alter intracellulariron homeostasis Vitamin D possesses immune modulatoryactivity and exerts an anti-inflammatory effect [47ndash49]Zughaier et al found that vitamin D plays an important rolein regulating the iron-hepcidin-ferroportin axis in mono-cytes by restoring hepcidin and ferroportin gene expression
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Immunology Research
Iron is a highly reactive metal thus to reduce metaltoxicity iron is maintained bound to hemoproteins and iniron-sulfur clusters or safely stored in ferritin cages [1]Free intracellular iron or poorly liganded iron is toxicsince it catalyzes the production of reactive oxygen species(ROS) Therefore macrophages detoxify labile iron by mul-tiple mechanisms Recently the presence of a mammaliansiderophore an iron-chelating small molecule was reportedand the iron-binding moiety of this molecule was identifiedas 25-dihydroxybenzoic acid (25DHBA) [12] This mam-malian siderophore captures free cytosolic iron and thusprotects the cells from oxidant stress [12] The enzyme thatcatalyzes the synthesis of 25DHBA is a member of the short-chain dehydrogenasereductase family (DHRS6) [13] called3-hydroxybutyrate dehydrogenase type 2 (BDH2) [12] Inhi-bition of BDH2 expression inmurine pro-B lymphocytic cellsled to the depletion of the mammalian siderophore and theaccumulation of labile ironThephysiological role of BDH2 insensing cellular iron was further confirmed by the discoverythat the BDH2 enzyme contains an iron responsive elementthat interacts with iron regulatory proteins that controlposttranscriptional gene expression [14] Recently Zughaieret al reported that BDH2 expression in macrophages issuppressed upon bacterial infection suggesting an importantrole for BDH2 in iron-limiting innate immunity [15] Morerecently the Devireddy group further confirmed the role ofBDH2 in cellular iron homeostasis and showed that depletionof the endogenous siderophore 25-DHBA in a mouse modelresulted in microcytic anemia and iron overload in thespleen [16 17] They also reported that BDH2 and ferritin-H synergistically regulate intracellular iron levels in an invitro experimental cell culture model [18] Therefore BDH2is required for intracellular iron homeostasis [12] Howeverthe role of BDH2 in iron-limiting innate immune defenses inmacrophages is not known
Macrophages engulf senescent RBCs and recycle hemo-globin thus they express heme oxygenase-1 (HO-1) thecatalytic enzyme that degrades heme consequently liberatingiron and producing carbon monoxide [19] HO-1 is highlyinduced in response to inflammation oxidative stress andhypoxia [19 20] due to its cytoprotective effects and anti-inflammatory activity [21 22] Therefore HO-1 as a stress-inducible protein is an important player in intracellulariron homeostasis Further neutrophil gelatinase-associatedlipocalin (NGAL) [23] functions as an iron carrier proteinthat shuttles bound iron across tissues by internalizationwith a membrane-localized receptor [24 25] and also playsa critical physiological role in intracellular iron homeostasisby trafficking the iron-bound siderophore 25DHBA [12] andiron-catecholamine complexes [26] NGAL plays a crucialrole in iron-limiting innate immune defenses since it limitsiron availability by also scavenging bacterial siderophores andthereby exerts an antibacterial function [27ndash29]
Intracellular iron homeostasis is disrupted by endoplas-mic reticulum (ER) stress [30] ER stress induced upon cellu-lar perturbations is regulated by activation of the unfoldedprotein response to restore homeostatic cellular functions[30] Control of ER stress is required for cell survival anduncontrolled ER stress leads to cell death [31 32] ER
stress alters cellular iron homeostasis by inducing hepcidinexpression [33 34] Altered iron homeostasis and ER stressare associated with chronic neurodegenerative diseases suchas Huntingtonrsquos disease and Alzheimerrsquos disease [35 36] It isnot known whether ER stress also affects BDH2 expressionThe goal of this studywas to investigate whether ER stress andinflammation modulate BDH2 expression in macrophages
Here we show that LPS-induced inflammation combinedwith chemically induced ER stress led to BDH2 downregula-tion increased the expression of ER stress markers includingCHOP upregulated hepcidin expression downregulated fer-roportin expression caused iron retention in macrophagesand dysregulated cytokine release from macrophages Fur-ther we show that ER stress combined with inflammationdramatically upregulated NGAL expression as well as theinducible hemeoxygenase-1 enzyme inmacrophagesWe alsoshow that vitamin D treatment prior to LPS restored BDH2expression This is the first report to show that ER stressand inflammation downregulate the expression of BDH2 inhuman THP-1 macrophages The results suggest that BDH2plays an important role in modulating iron-limiting innateimmune defenses
2 Materials and Methods
21 Reagents RPMI 1640 medium Dulbeccorsquos modifiedEagle medium (D-MEM) HBSS fetal bovine serum (FBS)penicillinstreptomycin sodium pyruvate and nonessentialamino acids were obtained fromCellgroMediatech (HerdonVA) Magnetic beads for a panel of human cytokines andchemokines were from Invitrogen (Carlsbad CA) Tuni-camycin calcein-AM and thiazolyl blue tetrazoliumbromidewere purchased from Sigma Aldrich (St Louis MO) Lipo-polysaccharide [37] from Neisseria meningitidis serogroup Bwas purified and quantified as previously described [38]
22 Cell Culture and Macrophage Stimulation THP-1macrophage-like cells were grown in RPMI 1640 mediumsupplemented with 10 FBS 50 120583gmL penicillin and50 IUmL of streptomycin HEK293 cells were grown inD-MEM supplemented with 10 FBS 50 120583gmL penicillinand 50 IUmL of streptomycin Freshly grown cells wereharvested and adjusted to 1 million cellsmL and thentransferred into 12-well tissue culture plates at 2mLwellTo induce ER stress cells were treated with tunicamycin(10 120583gmL in DMSO) and incubated overnight at 37∘CControl cells were treated with DMSO only and incubatedsimilarly Following overnight exposure to tunicamycin cellswere exposed to LPS (40 ngmL) to induce inflammationand incubated for 6 hr at 37∘C Cells were centrifugedand supernatants were saved at minus20∘C for cytokine meas-urements Harvested cells were washed with PBS and thenplaced in RLT buffer (Qiagen Hilden Germany) containing1 120573-mercaptoethanol passed over QiaShredder columnsand the resulting lysates were saved at minus80∘C for mRNAextraction
Journal of Immunology Research 3
23 RNA Isolation Quantitative Real-Time PCR and GeneExpression Analysis RNA was isolated using RNeasy Minikits (Qiagen) following the manufacturerrsquos instructions aspreviously described [39] Briefly cell lysates saved in RLTbuffer were mixed in 70 ethanol and then passed overRNeasy columns Columns were washed and treated with10 120583L of RNase-free DNase (Qiagen) for 15min at roomtemperature prior to RNA extraction followed by addi-tional washing and centrifugation RNA was eluted in 35 120583Lof RNase-free water and then was reverse transcribed tocDNA using QuantiTect Reverse Transcription kit (Qia-gen) following the manufacturerrsquos instructions Relative geneexpression was determined by quantitative RT-PCR per-formed on resulting cDNA using SYBR Green (PromegaMadison WI) following the manufacturerrsquos instructionsThemRNA level was calculated in reference to 120573-actin and foldchange gene expression was calculated in reference tocontrols using the ΔΔCT method Results were normalizedto unstimulated DMSO-treated cells which were used ascontrols for basal gene expression level The followingprimers were used for qRT-PCR reactions human hepci-din 51015840-GACCAGTGGCTCTGTTTTCC-31015840 and 51015840-CAC-ATCCCACACTTTGATCG-31015840 human BDH2 51015840-CAGCGT-CAAAGGAGTTGTGA-31015840 and 51015840-TGGCGTATCAACTGT-TCCTG-31015840 human NGAL 51015840-ATGACATGAACCTGCTCG-ATA-31015840 and 51015840-TCATAGTCGTTCATTATCTTC-31015840 human120573-actin 51015840-TCTTCCAGCCTTCCTTCCT-31015840 and 51015840-AGC-ACTGTGTTGGCGTACAG-31015840 Primers for ER stress mark-ers used in this study are as follows human hsXBP-1 51015840-TTC-CGGAGCTGGGTATCTCA-31015840 and 51015840-GAACCCCCGTAT-CCACAGTC-31015840 human GRP78 51015840-ACGGCAGCTGCT-ATTGCTTA-31015840 and 51015840-TCCTGACATCTTTGCCCGTC-31015840human CHOP 51015840-GACCTGCAAGAGGTCCTGTC-31015840 and51015840-GCAGGGTCAAGAGTGGTGAA-31015840 Ferroportin (HsSLC40A1 1 SG) transferrin receptor (Hs TFRC 1 SG) andheme oxygenase (Hs HMOX1 1 SG) QuantiTect primerswere purchased from Qiagen
24 Western Blot Analysis Freshly grown THP-1 cells wereharvested and adjusted to 1 million cellsmL and 8mL of cellswas transferred into 6-well tissue culture plates To induceER stress cells were treated with tunicamycin (10 120583gmL) andincubated overnight at 37∘C Control cells were treated withDMSO only and incubated similarly Following overnightincubation some cells were exposed to LPS (40 ngmL)to induce inflammation and incubated for 6 hr at 37∘CHarvested THP-1 cells were lysed in RIPA buffer (BostonBioProducts Boston MA) and centrifuged to remove nucleiand debris and the protein concentration of the supernatantwas measured using a BCA protein assay (ThermoWalthamMA) Equal amounts of protein were removed from eachmacrophage treatment group mixed with 6X Laemlli bufferand boiled for 15 minutes For untransfected HEK lysate andthe BDH2 transfected HEK positive control groups (OrigeneTechnologies Rockville MD) 50 ng of protein was loadedto adjust for overexpression Samples were loaded onto a12 MiniProtean TGX gel (BioRad Hercules CA) elec-trophoresed and transferred onto nitrocellulose membranes
(BioRad Hercules CA) To detect BDH2 membranes wereincubated with anti-BDH2 antibody (TrueMAB antibodyclone 2G1 from Origene) To detect actin membranes wereincubated with a mouse anti-120573 actin antibody (Sigma)In both cases membranes were incubated with an HRP-conjugated anti-mouse secondary antibody (Cell SignalingDanvers MA) incubated in Amersham western blot detec-tion reagent (GE Healthcare Cleveland OH) and visualizedwith HyBlot CL autoradiography film (Denville ScientificMetuchen NJ)
25 Cytokine and Chemokine Measurements Cytokines andchemokines released from THP-1 macrophages treated withtunicamycin alone LPS alone or both tunicamycin and LPSas described above were measured using Luminex magneticbeads (Invitrogen) following the manufacturerrsquos instructions
26 Measurement of Iron Retention inMacrophages Intracel-lular accumulation of labile iron in macrophages was mea-sured using the well-established calcein-AM method [40]Freshly grown THP-1 cells were treated with tunicamycin(10 120583gmL) and incubated overnight prior to stimulationwithLPS (40 ngmL) for 6 hr Control cells were treated withDMSO only Harvested cells were resuspended in RPMI1640 medium supplemented with 10 FBS and 05120583Mcalcein-AM was added and incubated for 15min at 37∘CCalcein-loaded cells were washed twice with PBS to removeextracellular calcein-AM and resuspended in HBSS 100120583Laliquots containing 025 million cells were transferred intoquadruplicate wells in black 96-well plates After a 20minincubation at 37∘C fluorescence was determined at 485 nm(excitation) and 535 nm (emission) wavelengths using a Bio-Tek Synergy 2 Instrument (Winooski VT)
27 Cellular Viability Measurement Using the ColorimetricMTT Reduction Assay The tetrazolium salt dye is cleavedby live and metabolically active cells but not by dead cellstherefore the MTT reduction assay was used to measurethe viability of THP-1 cells treated with tunicamycin withor without LPS [41 42] Briefly freshly grown THP-1 cellsadjusted to 1 million cellsmL were treated with tunicamycindoses ranging from 05 to 10 120583gmL and incubated overnightfollowed in some cases by 6 hr of exposure to LPS Controlcells treated with DMSO only were coincubated under thesame experimental conditions Five mg of thiazolyl bluetetrazolium bromide was dissolved in 1mL of PBS and 15 120583Lwas added to 150120583L of treated THP-1 cells whichwere furtherincubated at 37∘C for one hour Cells were centrifuged toremove extracellular dye and then lysed in 150 120583L of DMSOto dissolve the intracellular dye The absorbance of reduceddye was measured at 591 nm wavelength using a Bio-Tekspectrophotometer
28 Statistical Analysis Mean values plusmn SD of three inde-pendent experiments or two in few cases were calculatedwith Microsoft Excel Statistical analysis was performed withPrism v50 software using one-way ANOVA followed byBonferroni multiple comparisons post hoc analysis 119875 values
4 Journal of Immunology Research
37
25
50
75
(kD
)(k
D)
(kD
)
25
50
75
37
37
25
50
(i)
(a)
(b)
(c)
(ii)
(ii)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge
NS
05
06
07
08
09
1
11
0 1 25 5
BDH
2 m
RNA
relat
ive f
old
chan
ge
Vehi
cle
Tuni
cam
ycin
LPS
Tun+
LPS
HEK
(minus)
HEK
+BD
H2
120573-A
ctin
Ant
i-BD
H2
Ant
i-BD
H2
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowast
Tunicamycin dose (120583gmL)
1 2 3 4 5 6
Figure 1 ER stress and inflammation downregulate BDH2 expression in macrophages (a)Western blot analysis of BDH2 protein expressionin THP-1 macrophage-like cells (1 times 106 cellsmL) treated with tunicamycin (10120583gmL) or vehicle (DMSO) overnight prior to LPS exposure(40 ngmL) for 6 hr BDH2 protein was detected with monoclonal anti-BDH2 TrueMAB antibody clone 2G Lane 1 vehicle-treated THP-1cells 2 tunicamycin treated 3 LPS treated 4 tunicamycin and LPS treated 5 untransfected HEK293 cells as negative control 6 BDH2transfected HEK293 cells as positive control (BDH2 positive control contains a FLAG tag and 10-fold higher concentration was loaded inpanel (i) compared to panel (ii) representing two independent experiments) lowastnonspecific band (b) THP-1 macrophages were treated withtunicamycin (10120583gmL) as above and RNAwas extracted and BDH2 gene expression was assessed by quantitative RT-PCR normalized to thatof120573-actin (c) THP-1 cells treatedwith tunicamycin doses 1 25 or 5120583gmL and incubated overnightThe fold change inBDH2gene expressionwas calculated in reference to DMSO treated control THP-1 cells using the ΔΔCTmethod and presented here normalized to vehicle (DMSO)treatment control Error bars represent the SD from the mean of nine readouts generated from three independent experiments lowast119875 valueswere calculated in reference to vehicle (DMSO) treatment control using one-way ANOVA followed by Bonferroni multiple comparisons posthoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005119875 gt 005 were not significant
3 Results
31 BDH2 Expression inMacrophages Is Downregulated by ERStress and Inflammation Macrophages play a very importantrole in host defense and in iron homeostasis [1] RecentlyBDH2 was reported to play a crucial role in intracellulariron trafficking by catalyzing the synthesis of 25-DHBA [12]Further ER stress has been shown to dysregulate cellulariron homeostasis [30] Zughaier et al reported that BDH2expression in macrophages is downregulated upon bacterialinfection [15] However BDH2 expression and regulationin macrophages during ER stress and inflammation havenot yet been described To investigate BDH2 expression in
macrophages and its regulation by ER stress and inflamma-tion we employed human macrophage-like monocytic cells(THP-1) treated with tunicamycin overnight prior to LPSexposure for 6 hr to induce ER stress and inflammationrespectively Here we show that BDH2 protein and geneexpression in macrophages was downregulated by ER stressand inflammation (Figure 1) Reduced BDH2 protein levelswere observed in THP-1 cells after treatment with LPS andtunicamycin as compared to cells treated with DMSO alone(Figure 1(a)) Consistent with this observation BDH2 geneexpression was significantly downregulated when THP-1macrophages were treated with tunicamycin and LPS eitheralone or in combination (Figure 1(b)) Treatment with tuni-camycin alone reduced BDH2 mRNA expression in a dose-dependent manner (Figure 1(c)) Further we examined theactivation of ER stress markers and as expected overnight
Journal of Immunology Research 5
02468
101214161820
No treatment Tunicamycin LPS
CHO
P m
RNA
relat
ive f
old
chan
geNS
NS
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
(a)
0
1
2
3
4
5
6
7
8
No treatment Tunicamycin LPS
hsXB
P m
RNA
relat
ive f
old
chan
ge
NS
NS
lowast
lowast
lowastlowast
lowastlowast
Tunicamycin +LPS
(b)
0
10
20
30
40
50
60
70
No treatment Tunicamycin LPS
NSGRP
78 m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
(c)
Figure 2 ER stress markers are induced in macrophages by tunicamycin THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr RNA was extracted and ER stress markers CHOP (a) hsXBP (b)and GRP78 (c) gene expressions were assessed by quantitative RT-PCR normalized to 120573-actin The fold change in CHOP and hsXBP geneexpression was calculated in reference to DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to notreatment control Error bars represent the SD from the mean of three independent experiments lowast119875 values were calculated in reference to notreatment (DMSO only) control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotatedlowastlowastlowast
119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
tunicamycin treatment significantly induced ER stress mark-ers CHOP hsXBP-1 GRP78 (Figure 2) and ATF6 (data notshown) In contrast 6 hr LPS treatment alone did not upreg-ulate ER stress markers in THP-1 cells (Figure 2) as docu-mented previously [43] Cellular viability was assessed usingtheMTT reduction assay [41] which showed that the doses oftunicamycin and LPS used here did not reduce cellular via-bility (see Supplementary Figure 1A in Supplementary Mate-rial available online at httpdxdoiorg1011552014140728)These data indicate that ER stress and inflammation down-regulate BDH2 expression in THP-1 macrophages
32 Cellular Iron Homeostasis Is Dysregulated by ER Stressand Inflammation in Macrophages ER stress controls ironhomeostasis via the induction of hepcidin the master iron
regulating hormone [2] Hepcidin binds to the only knowncellular iron exporter ferroportin and leads to its internal-ization and degradation consequently inhibiting iron egressfrom macrophages [7] The dysregulation of the hepcidin-ferroportin axis leads to iron retention in macrophagesand increases the cytosolic labile iron pool which is highlyreactive and causes oxidative stress [3] Therefore detox-ifying labile iron is essential to maintaining cellular ironhomeostasis To investigate intracellular iron status in tuni-camycin and LPS treated THP-1 macrophages we determinedthe expression of hepcidin and ferroportin The data showthat tunicamycin-induced ER stress as well as LPS-inducedinflammation led to significant upregulation of hepcidin geneexpression in THP-1 macrophages (Figure 3(a)) In contrasttunicamycin and LPS treatment led to a dramatic reduction
6 Journal of Immunology Research
0
2
4
6
8
10
12
14
16
18
No treatment Tunicamycin LPS
NSNS
Tunicamycin +LPS
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastH
epci
din
mRN
A re
lativ
e fol
d ch
ange
(a)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
FPN
1 m
RNA
relat
ive f
old
chan
ge
NS
NS NS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(b)
Figure 3 ER stress and inflammation dysregulate cellular iron homeostasis THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr ((a) (b)) Hepcidin and ferroportin (FPN1) gene expression wasassessed by quantitative RT-PCR normalized to 120573-actin The fold change in hepcidin and FPN1 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to no treatment control Error bars representthe SD from the mean of three independent experiments 119875 values were calculated in reference to vehicle (DMSO) treatment control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
400000
500000
600000
700000
800000
900000
1000000
No treatment Tunicamycin LPS
NS
Calc
ein-
AM
fluo
resc
ence
(AU
)
lowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Tunicamycin +LPS
Figure 4 ER stress and inflammation induce iron retention in macrophages THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr and iron retention in macrophages was determined using thecalcein-AMfluorescent probemethod [40]DMSO treated cells were incubated simultaneously andused as controls Calcein-AMfluorescencewasmeasured by excitation at 488 nmand emission at 528 nmwavelength (see Section 2) Calcein-AMfluorescence is quenched upon bindingiron and is therefore inversely correlated with intracellular iron accumulation AU arbitrary units Error bars represent the SD from themeanof triplicate reading and data are representative of two independent experiments 119875 values were calculated in reference to vehicle (DMSO)treatment control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001lowastlowast
119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
in ferroportin expression in macrophages (Figure 3(b)) Fer-roportin allows iron egress from macrophages therefore itsdegradation leads to iron retention We measured labile ironaccumulation in macrophages using calcein-AM [40] andfound that tunicamycin and LPS treatment led to significant
iron retention (Figure 4 and Supplementary Figure 1B)A synergistic effect on iron retention was observed whenmacrophages were cotreated with tunicamycin and LPS Theresults suggest that chemically induced ER stress and inflam-mation alter intracellular iron homeostasis in macrophages
Journal of Immunology Research 7
0
500
1000
1500
2000
2500
3000
3500
4000
No treatment Tunicamycin LPS
NG
AL
mRN
A re
lativ
e fol
d ch
ange
123 fold1 fold
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
5 25 05 0
NG
AL
mRN
A re
lativ
e fol
d ch
ange
Tunicamycin
NS
Tunicamycin + LPS
Tunicamycin doses (120583gmL)
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowastlowast
(b)
Figure 5 NGAL gene expression is massively upregulated by ER stress (a) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr (b) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycindoses (5 25 or 05 120583gmL) as in panel (a) or tunicamycin followed by LPS (40 ngmL) RNA was extracted and NGAL gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actinThe fold change in NGAL gene expression was calculated in reference to DMSOtreated control THP-1 cells using theΔΔCTmethod and presented here normalized to vehicle (DMSO) treatment control Error bars representthe SD from the mean of three independent experiments lowast119875 values were calculated in reference to no treatment (DMSO only) control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
33 ER Stress and Inflammation Dramatically Induce NGALGene Expression NGAL plays a dual role in physiologiccellular iron homeostasis and in host defense We recentlyreported that LPS induced NGAL secretion from peripheralmonocytes [44] ER stress is also reported to highly induceNGAL expression [45] However the impact of ER stresscombined with inflammation on NGAL expression is not yetknown To this end we determined NGAL gene expressionin tunicamycin and LPS treated macrophages As expectedLPS treatment led to significant upregulation of NGALgene expression (sim12-fold) but tunicamycin treatment led toa dramatic sim1500-fold increase in NGAL gene expression(Figure 5(a)) Tunicamycin induced NGAL gene expressionin a dose-dependent manner (Figure 5(b)) A synergisticeffect was observed when THP1 cells were cotreated withtunicamycin and LPS which resulted in an approximatelysim3000-fold increase in NGAL gene expression (Figure 5(a))This synergistic effect was tunicamycin dose-dependent(Figure 5(b)) Tunicamycin-induced ER stress led to sim100-fold greater increase in NGAL gene expression comparedto LPS The synergistic cross-talk between tunicamycin-induced ER stress and LPS-induced inflammation led to sim200-fold increase in NGAL gene expression compared toLPS alone These data suggest that chemically induced ERstress dramatically increases NGAL expression more thaninflammation but when combined a synergistic upregulationof NGAL is observed
34 ER Stress and Inflammation Induce Heme Oxygenase-1 (HO-1) Gene Expression HO-1 is a stress-induced anti-inflammatory [19 21 22] cytoprotective enzyme that cat-alyzes the degradation of heme and consequently liber-ates iron and carbon monoxide [20] The released carbonmonoxide suppresses hepcidin expression therefore HO-1plays a role in regulating iron homeostasis [46] Our datademonstrate that ER stress and inflammation dysregulateintracellular iron homeostasis in macrophagesTherefore weinvestigated the impact of ER stress and inflammation onHO-1 expression in macrophages as an anti-inflammatoryimmune response Here we show that HO-1 gene expressionis significantly upregulated in THP-1 cells treated with tuni-camycin or LPS alone (Figure 6) However the combinationof tunicamycin andLPS treatment synergistically upregulatedHO-1 expression in THP-1 cells (Figure 6) suggesting a potentresponse to combat cellular stress
35 Vitamin D Restores BDH2 Expression in LPS-StimulatedMacrophages In this report we show that tunicamycin-induced ER stress and LPS-induced inflammation downreg-ulate BDH2 expression inmacrophages and alter intracellulariron homeostasis Vitamin D possesses immune modulatoryactivity and exerts an anti-inflammatory effect [47ndash49]Zughaier et al found that vitamin D plays an important rolein regulating the iron-hepcidin-ferroportin axis in mono-cytes by restoring hepcidin and ferroportin gene expression
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 3
23 RNA Isolation Quantitative Real-Time PCR and GeneExpression Analysis RNA was isolated using RNeasy Minikits (Qiagen) following the manufacturerrsquos instructions aspreviously described [39] Briefly cell lysates saved in RLTbuffer were mixed in 70 ethanol and then passed overRNeasy columns Columns were washed and treated with10 120583L of RNase-free DNase (Qiagen) for 15min at roomtemperature prior to RNA extraction followed by addi-tional washing and centrifugation RNA was eluted in 35 120583Lof RNase-free water and then was reverse transcribed tocDNA using QuantiTect Reverse Transcription kit (Qia-gen) following the manufacturerrsquos instructions Relative geneexpression was determined by quantitative RT-PCR per-formed on resulting cDNA using SYBR Green (PromegaMadison WI) following the manufacturerrsquos instructionsThemRNA level was calculated in reference to 120573-actin and foldchange gene expression was calculated in reference tocontrols using the ΔΔCT method Results were normalizedto unstimulated DMSO-treated cells which were used ascontrols for basal gene expression level The followingprimers were used for qRT-PCR reactions human hepci-din 51015840-GACCAGTGGCTCTGTTTTCC-31015840 and 51015840-CAC-ATCCCACACTTTGATCG-31015840 human BDH2 51015840-CAGCGT-CAAAGGAGTTGTGA-31015840 and 51015840-TGGCGTATCAACTGT-TCCTG-31015840 human NGAL 51015840-ATGACATGAACCTGCTCG-ATA-31015840 and 51015840-TCATAGTCGTTCATTATCTTC-31015840 human120573-actin 51015840-TCTTCCAGCCTTCCTTCCT-31015840 and 51015840-AGC-ACTGTGTTGGCGTACAG-31015840 Primers for ER stress mark-ers used in this study are as follows human hsXBP-1 51015840-TTC-CGGAGCTGGGTATCTCA-31015840 and 51015840-GAACCCCCGTAT-CCACAGTC-31015840 human GRP78 51015840-ACGGCAGCTGCT-ATTGCTTA-31015840 and 51015840-TCCTGACATCTTTGCCCGTC-31015840human CHOP 51015840-GACCTGCAAGAGGTCCTGTC-31015840 and51015840-GCAGGGTCAAGAGTGGTGAA-31015840 Ferroportin (HsSLC40A1 1 SG) transferrin receptor (Hs TFRC 1 SG) andheme oxygenase (Hs HMOX1 1 SG) QuantiTect primerswere purchased from Qiagen
24 Western Blot Analysis Freshly grown THP-1 cells wereharvested and adjusted to 1 million cellsmL and 8mL of cellswas transferred into 6-well tissue culture plates To induceER stress cells were treated with tunicamycin (10 120583gmL) andincubated overnight at 37∘C Control cells were treated withDMSO only and incubated similarly Following overnightincubation some cells were exposed to LPS (40 ngmL)to induce inflammation and incubated for 6 hr at 37∘CHarvested THP-1 cells were lysed in RIPA buffer (BostonBioProducts Boston MA) and centrifuged to remove nucleiand debris and the protein concentration of the supernatantwas measured using a BCA protein assay (ThermoWalthamMA) Equal amounts of protein were removed from eachmacrophage treatment group mixed with 6X Laemlli bufferand boiled for 15 minutes For untransfected HEK lysate andthe BDH2 transfected HEK positive control groups (OrigeneTechnologies Rockville MD) 50 ng of protein was loadedto adjust for overexpression Samples were loaded onto a12 MiniProtean TGX gel (BioRad Hercules CA) elec-trophoresed and transferred onto nitrocellulose membranes
(BioRad Hercules CA) To detect BDH2 membranes wereincubated with anti-BDH2 antibody (TrueMAB antibodyclone 2G1 from Origene) To detect actin membranes wereincubated with a mouse anti-120573 actin antibody (Sigma)In both cases membranes were incubated with an HRP-conjugated anti-mouse secondary antibody (Cell SignalingDanvers MA) incubated in Amersham western blot detec-tion reagent (GE Healthcare Cleveland OH) and visualizedwith HyBlot CL autoradiography film (Denville ScientificMetuchen NJ)
25 Cytokine and Chemokine Measurements Cytokines andchemokines released from THP-1 macrophages treated withtunicamycin alone LPS alone or both tunicamycin and LPSas described above were measured using Luminex magneticbeads (Invitrogen) following the manufacturerrsquos instructions
26 Measurement of Iron Retention inMacrophages Intracel-lular accumulation of labile iron in macrophages was mea-sured using the well-established calcein-AM method [40]Freshly grown THP-1 cells were treated with tunicamycin(10 120583gmL) and incubated overnight prior to stimulationwithLPS (40 ngmL) for 6 hr Control cells were treated withDMSO only Harvested cells were resuspended in RPMI1640 medium supplemented with 10 FBS and 05120583Mcalcein-AM was added and incubated for 15min at 37∘CCalcein-loaded cells were washed twice with PBS to removeextracellular calcein-AM and resuspended in HBSS 100120583Laliquots containing 025 million cells were transferred intoquadruplicate wells in black 96-well plates After a 20minincubation at 37∘C fluorescence was determined at 485 nm(excitation) and 535 nm (emission) wavelengths using a Bio-Tek Synergy 2 Instrument (Winooski VT)
27 Cellular Viability Measurement Using the ColorimetricMTT Reduction Assay The tetrazolium salt dye is cleavedby live and metabolically active cells but not by dead cellstherefore the MTT reduction assay was used to measurethe viability of THP-1 cells treated with tunicamycin withor without LPS [41 42] Briefly freshly grown THP-1 cellsadjusted to 1 million cellsmL were treated with tunicamycindoses ranging from 05 to 10 120583gmL and incubated overnightfollowed in some cases by 6 hr of exposure to LPS Controlcells treated with DMSO only were coincubated under thesame experimental conditions Five mg of thiazolyl bluetetrazolium bromide was dissolved in 1mL of PBS and 15 120583Lwas added to 150120583L of treated THP-1 cells whichwere furtherincubated at 37∘C for one hour Cells were centrifuged toremove extracellular dye and then lysed in 150 120583L of DMSOto dissolve the intracellular dye The absorbance of reduceddye was measured at 591 nm wavelength using a Bio-Tekspectrophotometer
28 Statistical Analysis Mean values plusmn SD of three inde-pendent experiments or two in few cases were calculatedwith Microsoft Excel Statistical analysis was performed withPrism v50 software using one-way ANOVA followed byBonferroni multiple comparisons post hoc analysis 119875 values
4 Journal of Immunology Research
37
25
50
75
(kD
)(k
D)
(kD
)
25
50
75
37
37
25
50
(i)
(a)
(b)
(c)
(ii)
(ii)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge
NS
05
06
07
08
09
1
11
0 1 25 5
BDH
2 m
RNA
relat
ive f
old
chan
ge
Vehi
cle
Tuni
cam
ycin
LPS
Tun+
LPS
HEK
(minus)
HEK
+BD
H2
120573-A
ctin
Ant
i-BD
H2
Ant
i-BD
H2
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowast
Tunicamycin dose (120583gmL)
1 2 3 4 5 6
Figure 1 ER stress and inflammation downregulate BDH2 expression in macrophages (a)Western blot analysis of BDH2 protein expressionin THP-1 macrophage-like cells (1 times 106 cellsmL) treated with tunicamycin (10120583gmL) or vehicle (DMSO) overnight prior to LPS exposure(40 ngmL) for 6 hr BDH2 protein was detected with monoclonal anti-BDH2 TrueMAB antibody clone 2G Lane 1 vehicle-treated THP-1cells 2 tunicamycin treated 3 LPS treated 4 tunicamycin and LPS treated 5 untransfected HEK293 cells as negative control 6 BDH2transfected HEK293 cells as positive control (BDH2 positive control contains a FLAG tag and 10-fold higher concentration was loaded inpanel (i) compared to panel (ii) representing two independent experiments) lowastnonspecific band (b) THP-1 macrophages were treated withtunicamycin (10120583gmL) as above and RNAwas extracted and BDH2 gene expression was assessed by quantitative RT-PCR normalized to thatof120573-actin (c) THP-1 cells treatedwith tunicamycin doses 1 25 or 5120583gmL and incubated overnightThe fold change inBDH2gene expressionwas calculated in reference to DMSO treated control THP-1 cells using the ΔΔCTmethod and presented here normalized to vehicle (DMSO)treatment control Error bars represent the SD from the mean of nine readouts generated from three independent experiments lowast119875 valueswere calculated in reference to vehicle (DMSO) treatment control using one-way ANOVA followed by Bonferroni multiple comparisons posthoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005119875 gt 005 were not significant
3 Results
31 BDH2 Expression inMacrophages Is Downregulated by ERStress and Inflammation Macrophages play a very importantrole in host defense and in iron homeostasis [1] RecentlyBDH2 was reported to play a crucial role in intracellulariron trafficking by catalyzing the synthesis of 25-DHBA [12]Further ER stress has been shown to dysregulate cellulariron homeostasis [30] Zughaier et al reported that BDH2expression in macrophages is downregulated upon bacterialinfection [15] However BDH2 expression and regulationin macrophages during ER stress and inflammation havenot yet been described To investigate BDH2 expression in
macrophages and its regulation by ER stress and inflamma-tion we employed human macrophage-like monocytic cells(THP-1) treated with tunicamycin overnight prior to LPSexposure for 6 hr to induce ER stress and inflammationrespectively Here we show that BDH2 protein and geneexpression in macrophages was downregulated by ER stressand inflammation (Figure 1) Reduced BDH2 protein levelswere observed in THP-1 cells after treatment with LPS andtunicamycin as compared to cells treated with DMSO alone(Figure 1(a)) Consistent with this observation BDH2 geneexpression was significantly downregulated when THP-1macrophages were treated with tunicamycin and LPS eitheralone or in combination (Figure 1(b)) Treatment with tuni-camycin alone reduced BDH2 mRNA expression in a dose-dependent manner (Figure 1(c)) Further we examined theactivation of ER stress markers and as expected overnight
Journal of Immunology Research 5
02468
101214161820
No treatment Tunicamycin LPS
CHO
P m
RNA
relat
ive f
old
chan
geNS
NS
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
(a)
0
1
2
3
4
5
6
7
8
No treatment Tunicamycin LPS
hsXB
P m
RNA
relat
ive f
old
chan
ge
NS
NS
lowast
lowast
lowastlowast
lowastlowast
Tunicamycin +LPS
(b)
0
10
20
30
40
50
60
70
No treatment Tunicamycin LPS
NSGRP
78 m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
(c)
Figure 2 ER stress markers are induced in macrophages by tunicamycin THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr RNA was extracted and ER stress markers CHOP (a) hsXBP (b)and GRP78 (c) gene expressions were assessed by quantitative RT-PCR normalized to 120573-actin The fold change in CHOP and hsXBP geneexpression was calculated in reference to DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to notreatment control Error bars represent the SD from the mean of three independent experiments lowast119875 values were calculated in reference to notreatment (DMSO only) control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotatedlowastlowastlowast
119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
tunicamycin treatment significantly induced ER stress mark-ers CHOP hsXBP-1 GRP78 (Figure 2) and ATF6 (data notshown) In contrast 6 hr LPS treatment alone did not upreg-ulate ER stress markers in THP-1 cells (Figure 2) as docu-mented previously [43] Cellular viability was assessed usingtheMTT reduction assay [41] which showed that the doses oftunicamycin and LPS used here did not reduce cellular via-bility (see Supplementary Figure 1A in Supplementary Mate-rial available online at httpdxdoiorg1011552014140728)These data indicate that ER stress and inflammation down-regulate BDH2 expression in THP-1 macrophages
32 Cellular Iron Homeostasis Is Dysregulated by ER Stressand Inflammation in Macrophages ER stress controls ironhomeostasis via the induction of hepcidin the master iron
regulating hormone [2] Hepcidin binds to the only knowncellular iron exporter ferroportin and leads to its internal-ization and degradation consequently inhibiting iron egressfrom macrophages [7] The dysregulation of the hepcidin-ferroportin axis leads to iron retention in macrophagesand increases the cytosolic labile iron pool which is highlyreactive and causes oxidative stress [3] Therefore detox-ifying labile iron is essential to maintaining cellular ironhomeostasis To investigate intracellular iron status in tuni-camycin and LPS treated THP-1 macrophages we determinedthe expression of hepcidin and ferroportin The data showthat tunicamycin-induced ER stress as well as LPS-inducedinflammation led to significant upregulation of hepcidin geneexpression in THP-1 macrophages (Figure 3(a)) In contrasttunicamycin and LPS treatment led to a dramatic reduction
6 Journal of Immunology Research
0
2
4
6
8
10
12
14
16
18
No treatment Tunicamycin LPS
NSNS
Tunicamycin +LPS
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastH
epci
din
mRN
A re
lativ
e fol
d ch
ange
(a)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
FPN
1 m
RNA
relat
ive f
old
chan
ge
NS
NS NS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(b)
Figure 3 ER stress and inflammation dysregulate cellular iron homeostasis THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr ((a) (b)) Hepcidin and ferroportin (FPN1) gene expression wasassessed by quantitative RT-PCR normalized to 120573-actin The fold change in hepcidin and FPN1 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to no treatment control Error bars representthe SD from the mean of three independent experiments 119875 values were calculated in reference to vehicle (DMSO) treatment control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
400000
500000
600000
700000
800000
900000
1000000
No treatment Tunicamycin LPS
NS
Calc
ein-
AM
fluo
resc
ence
(AU
)
lowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Tunicamycin +LPS
Figure 4 ER stress and inflammation induce iron retention in macrophages THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr and iron retention in macrophages was determined using thecalcein-AMfluorescent probemethod [40]DMSO treated cells were incubated simultaneously andused as controls Calcein-AMfluorescencewasmeasured by excitation at 488 nmand emission at 528 nmwavelength (see Section 2) Calcein-AMfluorescence is quenched upon bindingiron and is therefore inversely correlated with intracellular iron accumulation AU arbitrary units Error bars represent the SD from themeanof triplicate reading and data are representative of two independent experiments 119875 values were calculated in reference to vehicle (DMSO)treatment control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001lowastlowast
119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
in ferroportin expression in macrophages (Figure 3(b)) Fer-roportin allows iron egress from macrophages therefore itsdegradation leads to iron retention We measured labile ironaccumulation in macrophages using calcein-AM [40] andfound that tunicamycin and LPS treatment led to significant
iron retention (Figure 4 and Supplementary Figure 1B)A synergistic effect on iron retention was observed whenmacrophages were cotreated with tunicamycin and LPS Theresults suggest that chemically induced ER stress and inflam-mation alter intracellular iron homeostasis in macrophages
Journal of Immunology Research 7
0
500
1000
1500
2000
2500
3000
3500
4000
No treatment Tunicamycin LPS
NG
AL
mRN
A re
lativ
e fol
d ch
ange
123 fold1 fold
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
5 25 05 0
NG
AL
mRN
A re
lativ
e fol
d ch
ange
Tunicamycin
NS
Tunicamycin + LPS
Tunicamycin doses (120583gmL)
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowastlowast
(b)
Figure 5 NGAL gene expression is massively upregulated by ER stress (a) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr (b) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycindoses (5 25 or 05 120583gmL) as in panel (a) or tunicamycin followed by LPS (40 ngmL) RNA was extracted and NGAL gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actinThe fold change in NGAL gene expression was calculated in reference to DMSOtreated control THP-1 cells using theΔΔCTmethod and presented here normalized to vehicle (DMSO) treatment control Error bars representthe SD from the mean of three independent experiments lowast119875 values were calculated in reference to no treatment (DMSO only) control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
33 ER Stress and Inflammation Dramatically Induce NGALGene Expression NGAL plays a dual role in physiologiccellular iron homeostasis and in host defense We recentlyreported that LPS induced NGAL secretion from peripheralmonocytes [44] ER stress is also reported to highly induceNGAL expression [45] However the impact of ER stresscombined with inflammation on NGAL expression is not yetknown To this end we determined NGAL gene expressionin tunicamycin and LPS treated macrophages As expectedLPS treatment led to significant upregulation of NGALgene expression (sim12-fold) but tunicamycin treatment led toa dramatic sim1500-fold increase in NGAL gene expression(Figure 5(a)) Tunicamycin induced NGAL gene expressionin a dose-dependent manner (Figure 5(b)) A synergisticeffect was observed when THP1 cells were cotreated withtunicamycin and LPS which resulted in an approximatelysim3000-fold increase in NGAL gene expression (Figure 5(a))This synergistic effect was tunicamycin dose-dependent(Figure 5(b)) Tunicamycin-induced ER stress led to sim100-fold greater increase in NGAL gene expression comparedto LPS The synergistic cross-talk between tunicamycin-induced ER stress and LPS-induced inflammation led to sim200-fold increase in NGAL gene expression compared toLPS alone These data suggest that chemically induced ERstress dramatically increases NGAL expression more thaninflammation but when combined a synergistic upregulationof NGAL is observed
34 ER Stress and Inflammation Induce Heme Oxygenase-1 (HO-1) Gene Expression HO-1 is a stress-induced anti-inflammatory [19 21 22] cytoprotective enzyme that cat-alyzes the degradation of heme and consequently liber-ates iron and carbon monoxide [20] The released carbonmonoxide suppresses hepcidin expression therefore HO-1plays a role in regulating iron homeostasis [46] Our datademonstrate that ER stress and inflammation dysregulateintracellular iron homeostasis in macrophagesTherefore weinvestigated the impact of ER stress and inflammation onHO-1 expression in macrophages as an anti-inflammatoryimmune response Here we show that HO-1 gene expressionis significantly upregulated in THP-1 cells treated with tuni-camycin or LPS alone (Figure 6) However the combinationof tunicamycin andLPS treatment synergistically upregulatedHO-1 expression in THP-1 cells (Figure 6) suggesting a potentresponse to combat cellular stress
35 Vitamin D Restores BDH2 Expression in LPS-StimulatedMacrophages In this report we show that tunicamycin-induced ER stress and LPS-induced inflammation downreg-ulate BDH2 expression inmacrophages and alter intracellulariron homeostasis Vitamin D possesses immune modulatoryactivity and exerts an anti-inflammatory effect [47ndash49]Zughaier et al found that vitamin D plays an important rolein regulating the iron-hepcidin-ferroportin axis in mono-cytes by restoring hepcidin and ferroportin gene expression
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
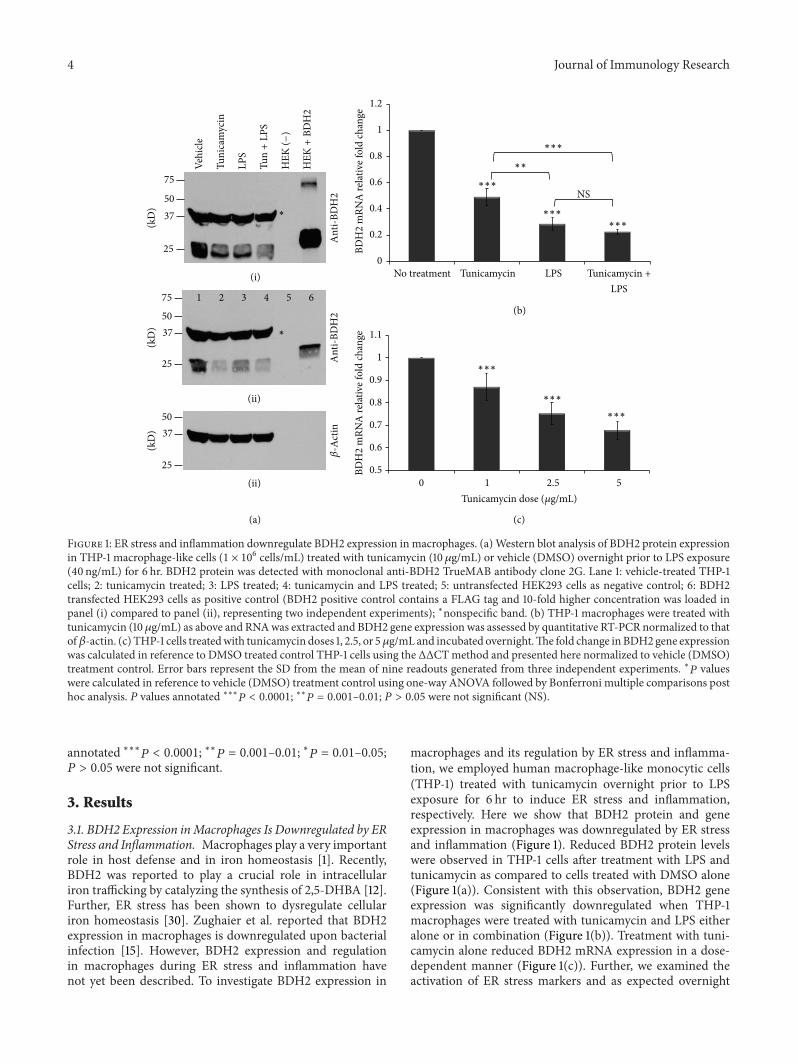
4 Journal of Immunology Research
37
25
50
75
(kD
)(k
D)
(kD
)
25
50
75
37
37
25
50
(i)
(a)
(b)
(c)
(ii)
(ii)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge
NS
05
06
07
08
09
1
11
0 1 25 5
BDH
2 m
RNA
relat
ive f
old
chan
ge
Vehi
cle
Tuni
cam
ycin
LPS
Tun+
LPS
HEK
(minus)
HEK
+BD
H2
120573-A
ctin
Ant
i-BD
H2
Ant
i-BD
H2
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowast
Tunicamycin dose (120583gmL)
1 2 3 4 5 6
Figure 1 ER stress and inflammation downregulate BDH2 expression in macrophages (a)Western blot analysis of BDH2 protein expressionin THP-1 macrophage-like cells (1 times 106 cellsmL) treated with tunicamycin (10120583gmL) or vehicle (DMSO) overnight prior to LPS exposure(40 ngmL) for 6 hr BDH2 protein was detected with monoclonal anti-BDH2 TrueMAB antibody clone 2G Lane 1 vehicle-treated THP-1cells 2 tunicamycin treated 3 LPS treated 4 tunicamycin and LPS treated 5 untransfected HEK293 cells as negative control 6 BDH2transfected HEK293 cells as positive control (BDH2 positive control contains a FLAG tag and 10-fold higher concentration was loaded inpanel (i) compared to panel (ii) representing two independent experiments) lowastnonspecific band (b) THP-1 macrophages were treated withtunicamycin (10120583gmL) as above and RNAwas extracted and BDH2 gene expression was assessed by quantitative RT-PCR normalized to thatof120573-actin (c) THP-1 cells treatedwith tunicamycin doses 1 25 or 5120583gmL and incubated overnightThe fold change inBDH2gene expressionwas calculated in reference to DMSO treated control THP-1 cells using the ΔΔCTmethod and presented here normalized to vehicle (DMSO)treatment control Error bars represent the SD from the mean of nine readouts generated from three independent experiments lowast119875 valueswere calculated in reference to vehicle (DMSO) treatment control using one-way ANOVA followed by Bonferroni multiple comparisons posthoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005119875 gt 005 were not significant
3 Results
31 BDH2 Expression inMacrophages Is Downregulated by ERStress and Inflammation Macrophages play a very importantrole in host defense and in iron homeostasis [1] RecentlyBDH2 was reported to play a crucial role in intracellulariron trafficking by catalyzing the synthesis of 25-DHBA [12]Further ER stress has been shown to dysregulate cellulariron homeostasis [30] Zughaier et al reported that BDH2expression in macrophages is downregulated upon bacterialinfection [15] However BDH2 expression and regulationin macrophages during ER stress and inflammation havenot yet been described To investigate BDH2 expression in
macrophages and its regulation by ER stress and inflamma-tion we employed human macrophage-like monocytic cells(THP-1) treated with tunicamycin overnight prior to LPSexposure for 6 hr to induce ER stress and inflammationrespectively Here we show that BDH2 protein and geneexpression in macrophages was downregulated by ER stressand inflammation (Figure 1) Reduced BDH2 protein levelswere observed in THP-1 cells after treatment with LPS andtunicamycin as compared to cells treated with DMSO alone(Figure 1(a)) Consistent with this observation BDH2 geneexpression was significantly downregulated when THP-1macrophages were treated with tunicamycin and LPS eitheralone or in combination (Figure 1(b)) Treatment with tuni-camycin alone reduced BDH2 mRNA expression in a dose-dependent manner (Figure 1(c)) Further we examined theactivation of ER stress markers and as expected overnight
Journal of Immunology Research 5
02468
101214161820
No treatment Tunicamycin LPS
CHO
P m
RNA
relat
ive f
old
chan
geNS
NS
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
(a)
0
1
2
3
4
5
6
7
8
No treatment Tunicamycin LPS
hsXB
P m
RNA
relat
ive f
old
chan
ge
NS
NS
lowast
lowast
lowastlowast
lowastlowast
Tunicamycin +LPS
(b)
0
10
20
30
40
50
60
70
No treatment Tunicamycin LPS
NSGRP
78 m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
(c)
Figure 2 ER stress markers are induced in macrophages by tunicamycin THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr RNA was extracted and ER stress markers CHOP (a) hsXBP (b)and GRP78 (c) gene expressions were assessed by quantitative RT-PCR normalized to 120573-actin The fold change in CHOP and hsXBP geneexpression was calculated in reference to DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to notreatment control Error bars represent the SD from the mean of three independent experiments lowast119875 values were calculated in reference to notreatment (DMSO only) control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotatedlowastlowastlowast
119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
tunicamycin treatment significantly induced ER stress mark-ers CHOP hsXBP-1 GRP78 (Figure 2) and ATF6 (data notshown) In contrast 6 hr LPS treatment alone did not upreg-ulate ER stress markers in THP-1 cells (Figure 2) as docu-mented previously [43] Cellular viability was assessed usingtheMTT reduction assay [41] which showed that the doses oftunicamycin and LPS used here did not reduce cellular via-bility (see Supplementary Figure 1A in Supplementary Mate-rial available online at httpdxdoiorg1011552014140728)These data indicate that ER stress and inflammation down-regulate BDH2 expression in THP-1 macrophages
32 Cellular Iron Homeostasis Is Dysregulated by ER Stressand Inflammation in Macrophages ER stress controls ironhomeostasis via the induction of hepcidin the master iron
regulating hormone [2] Hepcidin binds to the only knowncellular iron exporter ferroportin and leads to its internal-ization and degradation consequently inhibiting iron egressfrom macrophages [7] The dysregulation of the hepcidin-ferroportin axis leads to iron retention in macrophagesand increases the cytosolic labile iron pool which is highlyreactive and causes oxidative stress [3] Therefore detox-ifying labile iron is essential to maintaining cellular ironhomeostasis To investigate intracellular iron status in tuni-camycin and LPS treated THP-1 macrophages we determinedthe expression of hepcidin and ferroportin The data showthat tunicamycin-induced ER stress as well as LPS-inducedinflammation led to significant upregulation of hepcidin geneexpression in THP-1 macrophages (Figure 3(a)) In contrasttunicamycin and LPS treatment led to a dramatic reduction
6 Journal of Immunology Research
0
2
4
6
8
10
12
14
16
18
No treatment Tunicamycin LPS
NSNS
Tunicamycin +LPS
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastH
epci
din
mRN
A re
lativ
e fol
d ch
ange
(a)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
FPN
1 m
RNA
relat
ive f
old
chan
ge
NS
NS NS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(b)
Figure 3 ER stress and inflammation dysregulate cellular iron homeostasis THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr ((a) (b)) Hepcidin and ferroportin (FPN1) gene expression wasassessed by quantitative RT-PCR normalized to 120573-actin The fold change in hepcidin and FPN1 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to no treatment control Error bars representthe SD from the mean of three independent experiments 119875 values were calculated in reference to vehicle (DMSO) treatment control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
400000
500000
600000
700000
800000
900000
1000000
No treatment Tunicamycin LPS
NS
Calc
ein-
AM
fluo
resc
ence
(AU
)
lowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Tunicamycin +LPS
Figure 4 ER stress and inflammation induce iron retention in macrophages THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr and iron retention in macrophages was determined using thecalcein-AMfluorescent probemethod [40]DMSO treated cells were incubated simultaneously andused as controls Calcein-AMfluorescencewasmeasured by excitation at 488 nmand emission at 528 nmwavelength (see Section 2) Calcein-AMfluorescence is quenched upon bindingiron and is therefore inversely correlated with intracellular iron accumulation AU arbitrary units Error bars represent the SD from themeanof triplicate reading and data are representative of two independent experiments 119875 values were calculated in reference to vehicle (DMSO)treatment control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001lowastlowast
119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
in ferroportin expression in macrophages (Figure 3(b)) Fer-roportin allows iron egress from macrophages therefore itsdegradation leads to iron retention We measured labile ironaccumulation in macrophages using calcein-AM [40] andfound that tunicamycin and LPS treatment led to significant
iron retention (Figure 4 and Supplementary Figure 1B)A synergistic effect on iron retention was observed whenmacrophages were cotreated with tunicamycin and LPS Theresults suggest that chemically induced ER stress and inflam-mation alter intracellular iron homeostasis in macrophages
Journal of Immunology Research 7
0
500
1000
1500
2000
2500
3000
3500
4000
No treatment Tunicamycin LPS
NG
AL
mRN
A re
lativ
e fol
d ch
ange
123 fold1 fold
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
5 25 05 0
NG
AL
mRN
A re
lativ
e fol
d ch
ange
Tunicamycin
NS
Tunicamycin + LPS
Tunicamycin doses (120583gmL)
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowastlowast
(b)
Figure 5 NGAL gene expression is massively upregulated by ER stress (a) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr (b) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycindoses (5 25 or 05 120583gmL) as in panel (a) or tunicamycin followed by LPS (40 ngmL) RNA was extracted and NGAL gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actinThe fold change in NGAL gene expression was calculated in reference to DMSOtreated control THP-1 cells using theΔΔCTmethod and presented here normalized to vehicle (DMSO) treatment control Error bars representthe SD from the mean of three independent experiments lowast119875 values were calculated in reference to no treatment (DMSO only) control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
33 ER Stress and Inflammation Dramatically Induce NGALGene Expression NGAL plays a dual role in physiologiccellular iron homeostasis and in host defense We recentlyreported that LPS induced NGAL secretion from peripheralmonocytes [44] ER stress is also reported to highly induceNGAL expression [45] However the impact of ER stresscombined with inflammation on NGAL expression is not yetknown To this end we determined NGAL gene expressionin tunicamycin and LPS treated macrophages As expectedLPS treatment led to significant upregulation of NGALgene expression (sim12-fold) but tunicamycin treatment led toa dramatic sim1500-fold increase in NGAL gene expression(Figure 5(a)) Tunicamycin induced NGAL gene expressionin a dose-dependent manner (Figure 5(b)) A synergisticeffect was observed when THP1 cells were cotreated withtunicamycin and LPS which resulted in an approximatelysim3000-fold increase in NGAL gene expression (Figure 5(a))This synergistic effect was tunicamycin dose-dependent(Figure 5(b)) Tunicamycin-induced ER stress led to sim100-fold greater increase in NGAL gene expression comparedto LPS The synergistic cross-talk between tunicamycin-induced ER stress and LPS-induced inflammation led to sim200-fold increase in NGAL gene expression compared toLPS alone These data suggest that chemically induced ERstress dramatically increases NGAL expression more thaninflammation but when combined a synergistic upregulationof NGAL is observed
34 ER Stress and Inflammation Induce Heme Oxygenase-1 (HO-1) Gene Expression HO-1 is a stress-induced anti-inflammatory [19 21 22] cytoprotective enzyme that cat-alyzes the degradation of heme and consequently liber-ates iron and carbon monoxide [20] The released carbonmonoxide suppresses hepcidin expression therefore HO-1plays a role in regulating iron homeostasis [46] Our datademonstrate that ER stress and inflammation dysregulateintracellular iron homeostasis in macrophagesTherefore weinvestigated the impact of ER stress and inflammation onHO-1 expression in macrophages as an anti-inflammatoryimmune response Here we show that HO-1 gene expressionis significantly upregulated in THP-1 cells treated with tuni-camycin or LPS alone (Figure 6) However the combinationof tunicamycin andLPS treatment synergistically upregulatedHO-1 expression in THP-1 cells (Figure 6) suggesting a potentresponse to combat cellular stress
35 Vitamin D Restores BDH2 Expression in LPS-StimulatedMacrophages In this report we show that tunicamycin-induced ER stress and LPS-induced inflammation downreg-ulate BDH2 expression inmacrophages and alter intracellulariron homeostasis Vitamin D possesses immune modulatoryactivity and exerts an anti-inflammatory effect [47ndash49]Zughaier et al found that vitamin D plays an important rolein regulating the iron-hepcidin-ferroportin axis in mono-cytes by restoring hepcidin and ferroportin gene expression
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
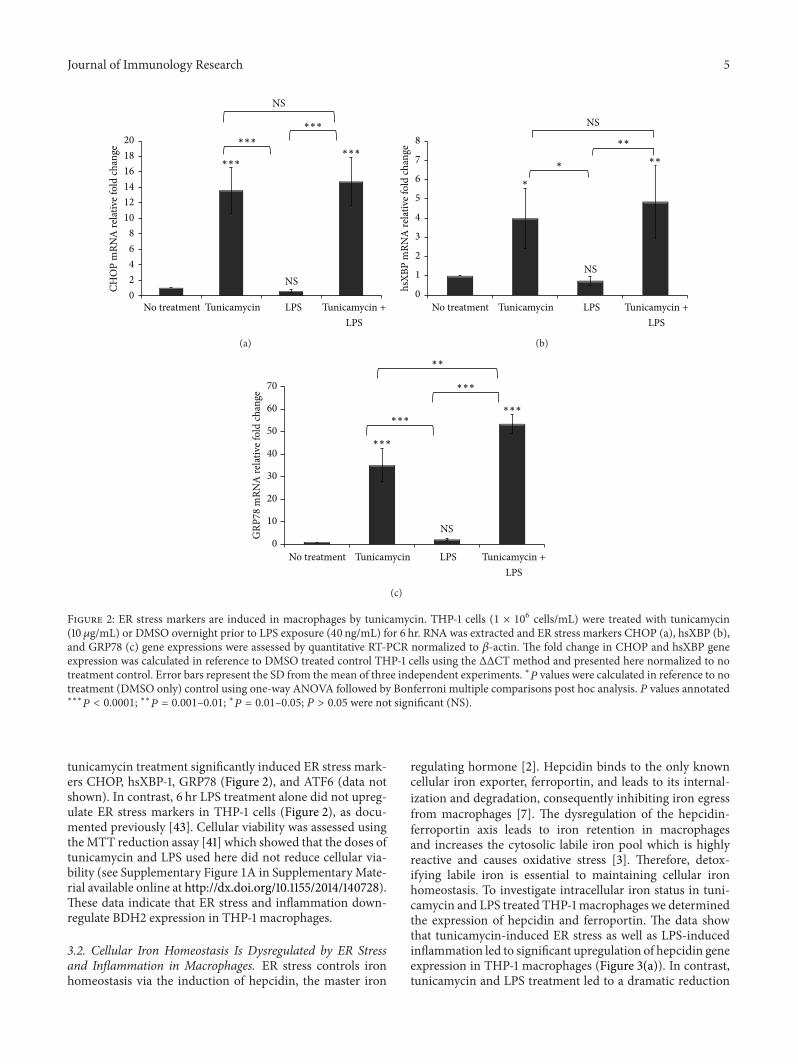
Journal of Immunology Research 5
02468
101214161820
No treatment Tunicamycin LPS
CHO
P m
RNA
relat
ive f
old
chan
geNS
NS
Tunicamycin +LPS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
(a)
0
1
2
3
4
5
6
7
8
No treatment Tunicamycin LPS
hsXB
P m
RNA
relat
ive f
old
chan
ge
NS
NS
lowast
lowast
lowastlowast
lowastlowast
Tunicamycin +LPS
(b)
0
10
20
30
40
50
60
70
No treatment Tunicamycin LPS
NSGRP
78 m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
(c)
Figure 2 ER stress markers are induced in macrophages by tunicamycin THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr RNA was extracted and ER stress markers CHOP (a) hsXBP (b)and GRP78 (c) gene expressions were assessed by quantitative RT-PCR normalized to 120573-actin The fold change in CHOP and hsXBP geneexpression was calculated in reference to DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to notreatment control Error bars represent the SD from the mean of three independent experiments lowast119875 values were calculated in reference to notreatment (DMSO only) control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotatedlowastlowastlowast
119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
tunicamycin treatment significantly induced ER stress mark-ers CHOP hsXBP-1 GRP78 (Figure 2) and ATF6 (data notshown) In contrast 6 hr LPS treatment alone did not upreg-ulate ER stress markers in THP-1 cells (Figure 2) as docu-mented previously [43] Cellular viability was assessed usingtheMTT reduction assay [41] which showed that the doses oftunicamycin and LPS used here did not reduce cellular via-bility (see Supplementary Figure 1A in Supplementary Mate-rial available online at httpdxdoiorg1011552014140728)These data indicate that ER stress and inflammation down-regulate BDH2 expression in THP-1 macrophages
32 Cellular Iron Homeostasis Is Dysregulated by ER Stressand Inflammation in Macrophages ER stress controls ironhomeostasis via the induction of hepcidin the master iron
regulating hormone [2] Hepcidin binds to the only knowncellular iron exporter ferroportin and leads to its internal-ization and degradation consequently inhibiting iron egressfrom macrophages [7] The dysregulation of the hepcidin-ferroportin axis leads to iron retention in macrophagesand increases the cytosolic labile iron pool which is highlyreactive and causes oxidative stress [3] Therefore detox-ifying labile iron is essential to maintaining cellular ironhomeostasis To investigate intracellular iron status in tuni-camycin and LPS treated THP-1 macrophages we determinedthe expression of hepcidin and ferroportin The data showthat tunicamycin-induced ER stress as well as LPS-inducedinflammation led to significant upregulation of hepcidin geneexpression in THP-1 macrophages (Figure 3(a)) In contrasttunicamycin and LPS treatment led to a dramatic reduction
6 Journal of Immunology Research
0
2
4
6
8
10
12
14
16
18
No treatment Tunicamycin LPS
NSNS
Tunicamycin +LPS
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastH
epci
din
mRN
A re
lativ
e fol
d ch
ange
(a)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
FPN
1 m
RNA
relat
ive f
old
chan
ge
NS
NS NS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(b)
Figure 3 ER stress and inflammation dysregulate cellular iron homeostasis THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr ((a) (b)) Hepcidin and ferroportin (FPN1) gene expression wasassessed by quantitative RT-PCR normalized to 120573-actin The fold change in hepcidin and FPN1 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to no treatment control Error bars representthe SD from the mean of three independent experiments 119875 values were calculated in reference to vehicle (DMSO) treatment control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
400000
500000
600000
700000
800000
900000
1000000
No treatment Tunicamycin LPS
NS
Calc
ein-
AM
fluo
resc
ence
(AU
)
lowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Tunicamycin +LPS
Figure 4 ER stress and inflammation induce iron retention in macrophages THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr and iron retention in macrophages was determined using thecalcein-AMfluorescent probemethod [40]DMSO treated cells were incubated simultaneously andused as controls Calcein-AMfluorescencewasmeasured by excitation at 488 nmand emission at 528 nmwavelength (see Section 2) Calcein-AMfluorescence is quenched upon bindingiron and is therefore inversely correlated with intracellular iron accumulation AU arbitrary units Error bars represent the SD from themeanof triplicate reading and data are representative of two independent experiments 119875 values were calculated in reference to vehicle (DMSO)treatment control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001lowastlowast
119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
in ferroportin expression in macrophages (Figure 3(b)) Fer-roportin allows iron egress from macrophages therefore itsdegradation leads to iron retention We measured labile ironaccumulation in macrophages using calcein-AM [40] andfound that tunicamycin and LPS treatment led to significant
iron retention (Figure 4 and Supplementary Figure 1B)A synergistic effect on iron retention was observed whenmacrophages were cotreated with tunicamycin and LPS Theresults suggest that chemically induced ER stress and inflam-mation alter intracellular iron homeostasis in macrophages
Journal of Immunology Research 7
0
500
1000
1500
2000
2500
3000
3500
4000
No treatment Tunicamycin LPS
NG
AL
mRN
A re
lativ
e fol
d ch
ange
123 fold1 fold
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
5 25 05 0
NG
AL
mRN
A re
lativ
e fol
d ch
ange
Tunicamycin
NS
Tunicamycin + LPS
Tunicamycin doses (120583gmL)
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowastlowast
(b)
Figure 5 NGAL gene expression is massively upregulated by ER stress (a) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr (b) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycindoses (5 25 or 05 120583gmL) as in panel (a) or tunicamycin followed by LPS (40 ngmL) RNA was extracted and NGAL gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actinThe fold change in NGAL gene expression was calculated in reference to DMSOtreated control THP-1 cells using theΔΔCTmethod and presented here normalized to vehicle (DMSO) treatment control Error bars representthe SD from the mean of three independent experiments lowast119875 values were calculated in reference to no treatment (DMSO only) control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
33 ER Stress and Inflammation Dramatically Induce NGALGene Expression NGAL plays a dual role in physiologiccellular iron homeostasis and in host defense We recentlyreported that LPS induced NGAL secretion from peripheralmonocytes [44] ER stress is also reported to highly induceNGAL expression [45] However the impact of ER stresscombined with inflammation on NGAL expression is not yetknown To this end we determined NGAL gene expressionin tunicamycin and LPS treated macrophages As expectedLPS treatment led to significant upregulation of NGALgene expression (sim12-fold) but tunicamycin treatment led toa dramatic sim1500-fold increase in NGAL gene expression(Figure 5(a)) Tunicamycin induced NGAL gene expressionin a dose-dependent manner (Figure 5(b)) A synergisticeffect was observed when THP1 cells were cotreated withtunicamycin and LPS which resulted in an approximatelysim3000-fold increase in NGAL gene expression (Figure 5(a))This synergistic effect was tunicamycin dose-dependent(Figure 5(b)) Tunicamycin-induced ER stress led to sim100-fold greater increase in NGAL gene expression comparedto LPS The synergistic cross-talk between tunicamycin-induced ER stress and LPS-induced inflammation led to sim200-fold increase in NGAL gene expression compared toLPS alone These data suggest that chemically induced ERstress dramatically increases NGAL expression more thaninflammation but when combined a synergistic upregulationof NGAL is observed
34 ER Stress and Inflammation Induce Heme Oxygenase-1 (HO-1) Gene Expression HO-1 is a stress-induced anti-inflammatory [19 21 22] cytoprotective enzyme that cat-alyzes the degradation of heme and consequently liber-ates iron and carbon monoxide [20] The released carbonmonoxide suppresses hepcidin expression therefore HO-1plays a role in regulating iron homeostasis [46] Our datademonstrate that ER stress and inflammation dysregulateintracellular iron homeostasis in macrophagesTherefore weinvestigated the impact of ER stress and inflammation onHO-1 expression in macrophages as an anti-inflammatoryimmune response Here we show that HO-1 gene expressionis significantly upregulated in THP-1 cells treated with tuni-camycin or LPS alone (Figure 6) However the combinationof tunicamycin andLPS treatment synergistically upregulatedHO-1 expression in THP-1 cells (Figure 6) suggesting a potentresponse to combat cellular stress
35 Vitamin D Restores BDH2 Expression in LPS-StimulatedMacrophages In this report we show that tunicamycin-induced ER stress and LPS-induced inflammation downreg-ulate BDH2 expression inmacrophages and alter intracellulariron homeostasis Vitamin D possesses immune modulatoryactivity and exerts an anti-inflammatory effect [47ndash49]Zughaier et al found that vitamin D plays an important rolein regulating the iron-hepcidin-ferroportin axis in mono-cytes by restoring hepcidin and ferroportin gene expression
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
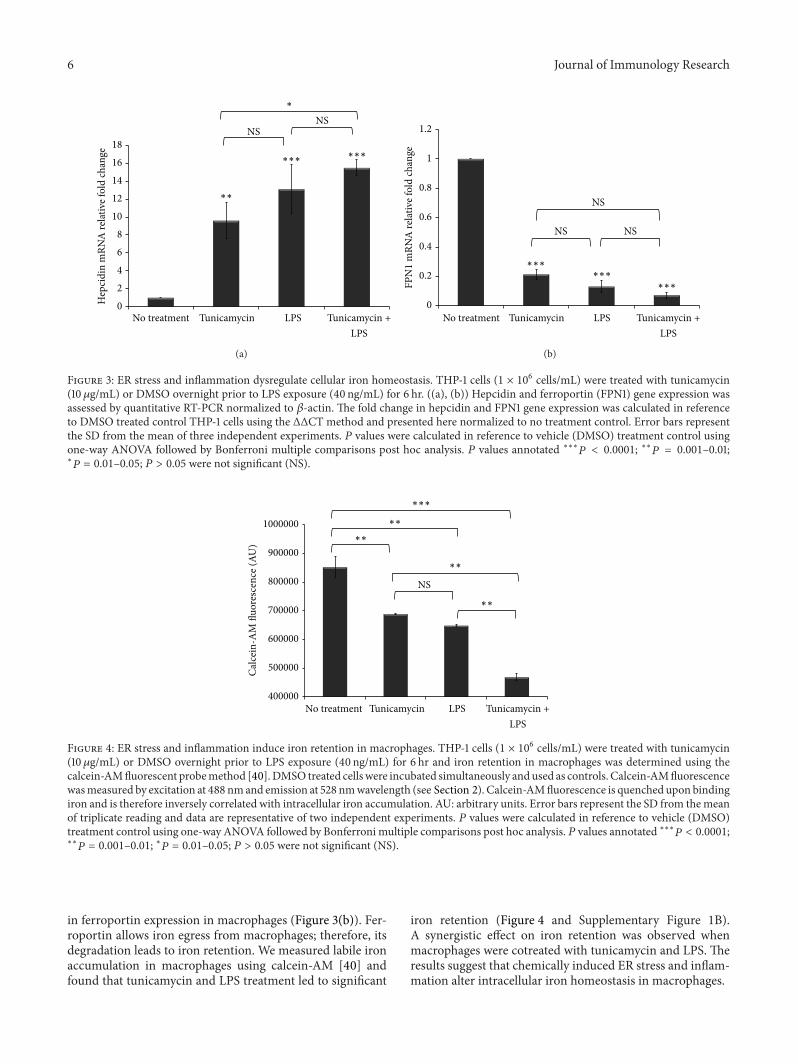
6 Journal of Immunology Research
0
2
4
6
8
10
12
14
16
18
No treatment Tunicamycin LPS
NSNS
Tunicamycin +LPS
lowastlowastlowastlowastlowastlowast
lowastlowast
lowastH
epci
din
mRN
A re
lativ
e fol
d ch
ange
(a)
0
02
04
06
08
1
12
No treatment Tunicamycin LPS
FPN
1 m
RNA
relat
ive f
old
chan
ge
NS
NS NS
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(b)
Figure 3 ER stress and inflammation dysregulate cellular iron homeostasis THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr ((a) (b)) Hepcidin and ferroportin (FPN1) gene expression wasassessed by quantitative RT-PCR normalized to 120573-actin The fold change in hepcidin and FPN1 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT method and presented here normalized to no treatment control Error bars representthe SD from the mean of three independent experiments 119875 values were calculated in reference to vehicle (DMSO) treatment control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
400000
500000
600000
700000
800000
900000
1000000
No treatment Tunicamycin LPS
NS
Calc
ein-
AM
fluo
resc
ence
(AU
)
lowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Tunicamycin +LPS
Figure 4 ER stress and inflammation induce iron retention in macrophages THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr and iron retention in macrophages was determined using thecalcein-AMfluorescent probemethod [40]DMSO treated cells were incubated simultaneously andused as controls Calcein-AMfluorescencewasmeasured by excitation at 488 nmand emission at 528 nmwavelength (see Section 2) Calcein-AMfluorescence is quenched upon bindingiron and is therefore inversely correlated with intracellular iron accumulation AU arbitrary units Error bars represent the SD from themeanof triplicate reading and data are representative of two independent experiments 119875 values were calculated in reference to vehicle (DMSO)treatment control using one-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001lowastlowast
119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
in ferroportin expression in macrophages (Figure 3(b)) Fer-roportin allows iron egress from macrophages therefore itsdegradation leads to iron retention We measured labile ironaccumulation in macrophages using calcein-AM [40] andfound that tunicamycin and LPS treatment led to significant
iron retention (Figure 4 and Supplementary Figure 1B)A synergistic effect on iron retention was observed whenmacrophages were cotreated with tunicamycin and LPS Theresults suggest that chemically induced ER stress and inflam-mation alter intracellular iron homeostasis in macrophages
Journal of Immunology Research 7
0
500
1000
1500
2000
2500
3000
3500
4000
No treatment Tunicamycin LPS
NG
AL
mRN
A re
lativ
e fol
d ch
ange
123 fold1 fold
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
5 25 05 0
NG
AL
mRN
A re
lativ
e fol
d ch
ange
Tunicamycin
NS
Tunicamycin + LPS
Tunicamycin doses (120583gmL)
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowastlowast
(b)
Figure 5 NGAL gene expression is massively upregulated by ER stress (a) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr (b) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycindoses (5 25 or 05 120583gmL) as in panel (a) or tunicamycin followed by LPS (40 ngmL) RNA was extracted and NGAL gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actinThe fold change in NGAL gene expression was calculated in reference to DMSOtreated control THP-1 cells using theΔΔCTmethod and presented here normalized to vehicle (DMSO) treatment control Error bars representthe SD from the mean of three independent experiments lowast119875 values were calculated in reference to no treatment (DMSO only) control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
33 ER Stress and Inflammation Dramatically Induce NGALGene Expression NGAL plays a dual role in physiologiccellular iron homeostasis and in host defense We recentlyreported that LPS induced NGAL secretion from peripheralmonocytes [44] ER stress is also reported to highly induceNGAL expression [45] However the impact of ER stresscombined with inflammation on NGAL expression is not yetknown To this end we determined NGAL gene expressionin tunicamycin and LPS treated macrophages As expectedLPS treatment led to significant upregulation of NGALgene expression (sim12-fold) but tunicamycin treatment led toa dramatic sim1500-fold increase in NGAL gene expression(Figure 5(a)) Tunicamycin induced NGAL gene expressionin a dose-dependent manner (Figure 5(b)) A synergisticeffect was observed when THP1 cells were cotreated withtunicamycin and LPS which resulted in an approximatelysim3000-fold increase in NGAL gene expression (Figure 5(a))This synergistic effect was tunicamycin dose-dependent(Figure 5(b)) Tunicamycin-induced ER stress led to sim100-fold greater increase in NGAL gene expression comparedto LPS The synergistic cross-talk between tunicamycin-induced ER stress and LPS-induced inflammation led to sim200-fold increase in NGAL gene expression compared toLPS alone These data suggest that chemically induced ERstress dramatically increases NGAL expression more thaninflammation but when combined a synergistic upregulationof NGAL is observed
34 ER Stress and Inflammation Induce Heme Oxygenase-1 (HO-1) Gene Expression HO-1 is a stress-induced anti-inflammatory [19 21 22] cytoprotective enzyme that cat-alyzes the degradation of heme and consequently liber-ates iron and carbon monoxide [20] The released carbonmonoxide suppresses hepcidin expression therefore HO-1plays a role in regulating iron homeostasis [46] Our datademonstrate that ER stress and inflammation dysregulateintracellular iron homeostasis in macrophagesTherefore weinvestigated the impact of ER stress and inflammation onHO-1 expression in macrophages as an anti-inflammatoryimmune response Here we show that HO-1 gene expressionis significantly upregulated in THP-1 cells treated with tuni-camycin or LPS alone (Figure 6) However the combinationof tunicamycin andLPS treatment synergistically upregulatedHO-1 expression in THP-1 cells (Figure 6) suggesting a potentresponse to combat cellular stress
35 Vitamin D Restores BDH2 Expression in LPS-StimulatedMacrophages In this report we show that tunicamycin-induced ER stress and LPS-induced inflammation downreg-ulate BDH2 expression inmacrophages and alter intracellulariron homeostasis Vitamin D possesses immune modulatoryactivity and exerts an anti-inflammatory effect [47ndash49]Zughaier et al found that vitamin D plays an important rolein regulating the iron-hepcidin-ferroportin axis in mono-cytes by restoring hepcidin and ferroportin gene expression
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
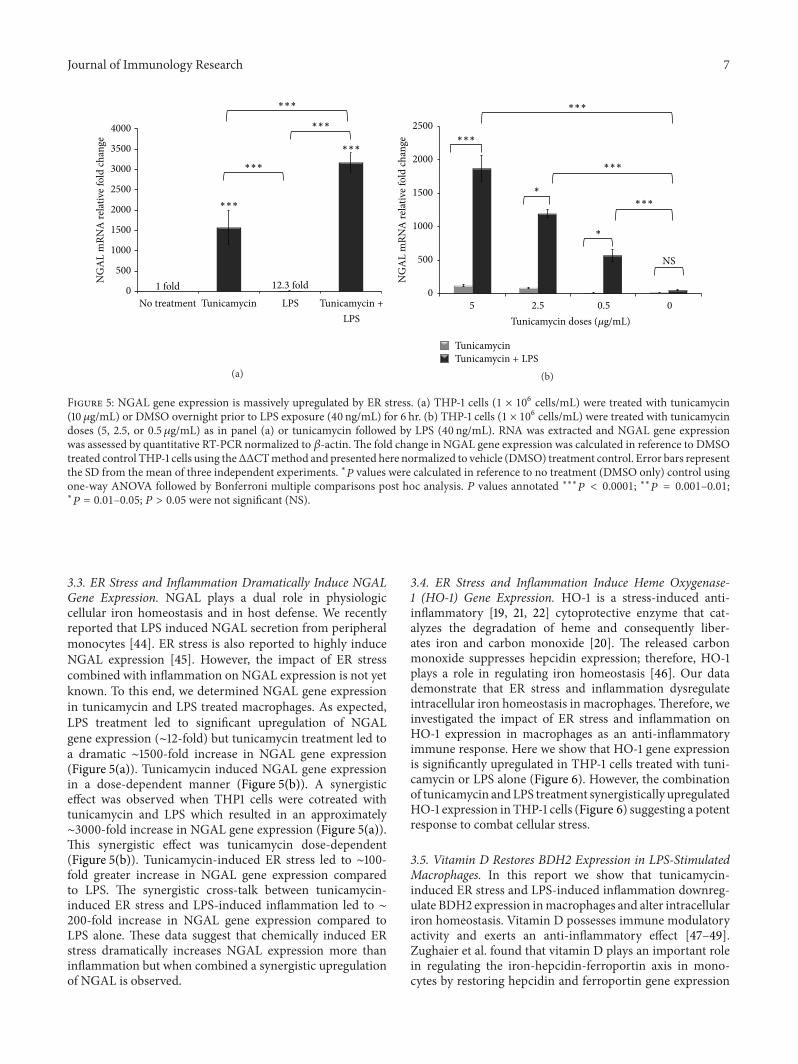
Journal of Immunology Research 7
0
500
1000
1500
2000
2500
3000
3500
4000
No treatment Tunicamycin LPS
NG
AL
mRN
A re
lativ
e fol
d ch
ange
123 fold1 fold
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
5 25 05 0
NG
AL
mRN
A re
lativ
e fol
d ch
ange
Tunicamycin
NS
Tunicamycin + LPS
Tunicamycin doses (120583gmL)
lowastlowastlowast
lowastlowastlowast
lowast
lowast
lowastlowastlowast
lowastlowastlowast
(b)
Figure 5 NGAL gene expression is massively upregulated by ER stress (a) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycin(10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr (b) THP-1 cells (1 times 106 cellsmL) were treated with tunicamycindoses (5 25 or 05 120583gmL) as in panel (a) or tunicamycin followed by LPS (40 ngmL) RNA was extracted and NGAL gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actinThe fold change in NGAL gene expression was calculated in reference to DMSOtreated control THP-1 cells using theΔΔCTmethod and presented here normalized to vehicle (DMSO) treatment control Error bars representthe SD from the mean of three independent experiments lowast119875 values were calculated in reference to no treatment (DMSO only) control usingone-way ANOVA followed by Bonferroni multiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001lowast
119875 = 001ndash005 119875 gt 005 were not significant (NS)
33 ER Stress and Inflammation Dramatically Induce NGALGene Expression NGAL plays a dual role in physiologiccellular iron homeostasis and in host defense We recentlyreported that LPS induced NGAL secretion from peripheralmonocytes [44] ER stress is also reported to highly induceNGAL expression [45] However the impact of ER stresscombined with inflammation on NGAL expression is not yetknown To this end we determined NGAL gene expressionin tunicamycin and LPS treated macrophages As expectedLPS treatment led to significant upregulation of NGALgene expression (sim12-fold) but tunicamycin treatment led toa dramatic sim1500-fold increase in NGAL gene expression(Figure 5(a)) Tunicamycin induced NGAL gene expressionin a dose-dependent manner (Figure 5(b)) A synergisticeffect was observed when THP1 cells were cotreated withtunicamycin and LPS which resulted in an approximatelysim3000-fold increase in NGAL gene expression (Figure 5(a))This synergistic effect was tunicamycin dose-dependent(Figure 5(b)) Tunicamycin-induced ER stress led to sim100-fold greater increase in NGAL gene expression comparedto LPS The synergistic cross-talk between tunicamycin-induced ER stress and LPS-induced inflammation led to sim200-fold increase in NGAL gene expression compared toLPS alone These data suggest that chemically induced ERstress dramatically increases NGAL expression more thaninflammation but when combined a synergistic upregulationof NGAL is observed
34 ER Stress and Inflammation Induce Heme Oxygenase-1 (HO-1) Gene Expression HO-1 is a stress-induced anti-inflammatory [19 21 22] cytoprotective enzyme that cat-alyzes the degradation of heme and consequently liber-ates iron and carbon monoxide [20] The released carbonmonoxide suppresses hepcidin expression therefore HO-1plays a role in regulating iron homeostasis [46] Our datademonstrate that ER stress and inflammation dysregulateintracellular iron homeostasis in macrophagesTherefore weinvestigated the impact of ER stress and inflammation onHO-1 expression in macrophages as an anti-inflammatoryimmune response Here we show that HO-1 gene expressionis significantly upregulated in THP-1 cells treated with tuni-camycin or LPS alone (Figure 6) However the combinationof tunicamycin andLPS treatment synergistically upregulatedHO-1 expression in THP-1 cells (Figure 6) suggesting a potentresponse to combat cellular stress
35 Vitamin D Restores BDH2 Expression in LPS-StimulatedMacrophages In this report we show that tunicamycin-induced ER stress and LPS-induced inflammation downreg-ulate BDH2 expression inmacrophages and alter intracellulariron homeostasis Vitamin D possesses immune modulatoryactivity and exerts an anti-inflammatory effect [47ndash49]Zughaier et al found that vitamin D plays an important rolein regulating the iron-hepcidin-ferroportin axis in mono-cytes by restoring hepcidin and ferroportin gene expression
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
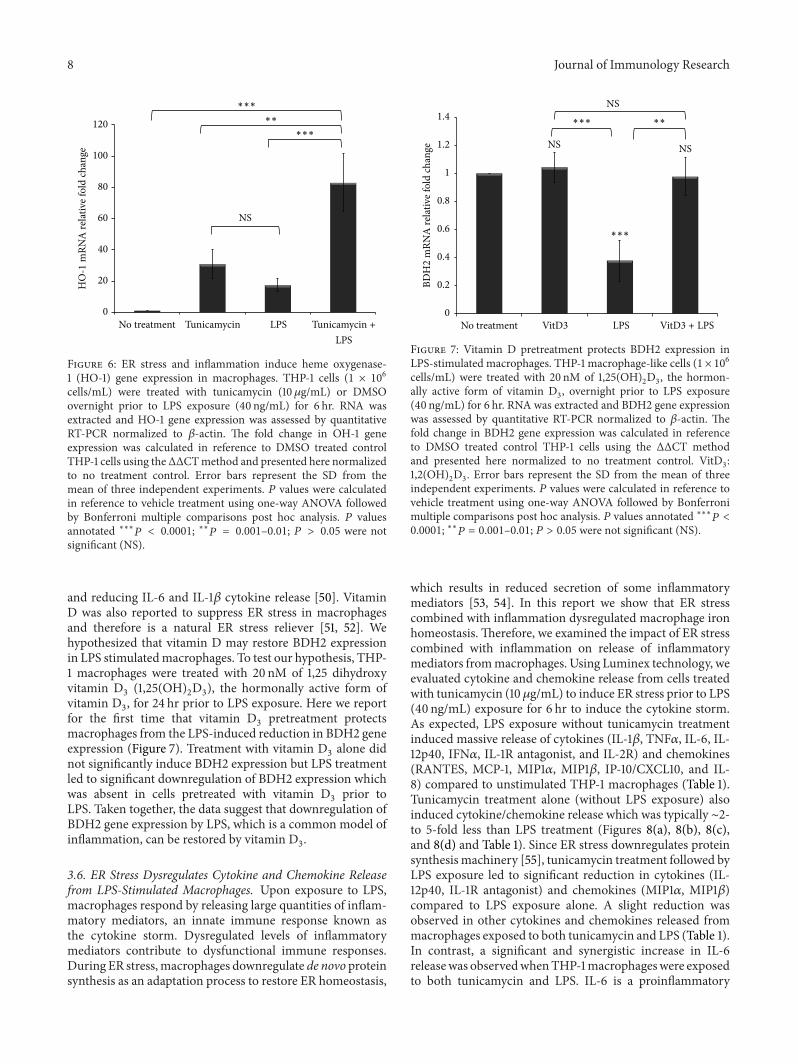
8 Journal of Immunology Research
0
20
40
60
80
100
120
No treatment Tunicamycin LPS
NS
HO
-1m
RNA
relat
ive f
old
chan
ge
lowastlowastlowast
lowastlowastlowast
lowastlowast
Tunicamycin +LPS
Figure 6 ER stress and inflammation induce heme oxygenase-1 (HO-1) gene expression in macrophages THP-1 cells (1 times 106cellsmL) were treated with tunicamycin (10120583gmL) or DMSOovernight prior to LPS exposure (40 ngmL) for 6 hr RNA wasextracted and HO-1 gene expression was assessed by quantitativeRT-PCR normalized to 120573-actin The fold change in OH-1 geneexpression was calculated in reference to DMSO treated controlTHP-1 cells using theΔΔCTmethod and presented here normalizedto no treatment control Error bars represent the SD from themean of three independent experiments 119875 values were calculatedin reference to vehicle treatment using one-way ANOVA followedby Bonferroni multiple comparisons post hoc analysis 119875 valuesannotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were notsignificant (NS)
and reducing IL-6 and IL-1120573 cytokine release [50] VitaminD was also reported to suppress ER stress in macrophagesand therefore is a natural ER stress reliever [51 52] Wehypothesized that vitamin D may restore BDH2 expressionin LPS stimulatedmacrophages To test our hypothesis THP-1 macrophages were treated with 20 nM of 125 dihydroxyvitamin D
3(125(OH)
2D3) the hormonally active form of
vitamin D3 for 24 hr prior to LPS exposure Here we report
for the first time that vitamin D3pretreatment protects
macrophages from the LPS-induced reduction in BDH2 geneexpression (Figure 7) Treatment with vitamin D
3alone did
not significantly induce BDH2 expression but LPS treatmentled to significant downregulation of BDH2 expression whichwas absent in cells pretreated with vitamin D
3prior to
LPS Taken together the data suggest that downregulation ofBDH2 gene expression by LPS which is a common model ofinflammation can be restored by vitamin D
3
36 ER Stress Dysregulates Cytokine and Chemokine Releasefrom LPS-Stimulated Macrophages Upon exposure to LPSmacrophages respond by releasing large quantities of inflam-matory mediators an innate immune response known asthe cytokine storm Dysregulated levels of inflammatorymediators contribute to dysfunctional immune responsesDuring ER stressmacrophages downregulate de novo proteinsynthesis as an adaptation process to restore ER homeostasis
0
02
04
06
08
1
12
14
No treatment VitD3 LPS
BDH
2 m
RNA
relat
ive f
old
chan
ge NS NS
NS
VitD3 + LPS
lowastlowastlowast
lowastlowastlowast lowastlowast
Figure 7 Vitamin D pretreatment protects BDH2 expression inLPS-stimulated macrophages THP-1 macrophage-like cells (1 times 106cellsmL) were treated with 20 nM of 125(OH)
2
D3
the hormon-ally active form of vitamin D
3
overnight prior to LPS exposure(40 ngmL) for 6 hr RNA was extracted and BDH2 gene expressionwas assessed by quantitative RT-PCR normalized to 120573-actin Thefold change in BDH2 gene expression was calculated in referenceto DMSO treated control THP-1 cells using the ΔΔCT methodand presented here normalized to no treatment control VitD
3
12(OH)
2
D3
Error bars represent the SD from the mean of threeindependent experiments 119875 values were calculated in reference tovehicle treatment using one-way ANOVA followed by Bonferronimultiple comparisons post hoc analysis 119875 values annotated lowastlowastlowast119875 lt00001 lowastlowast119875 = 0001ndash001 119875 gt 005 were not significant (NS)
which results in reduced secretion of some inflammatorymediators [53 54] In this report we show that ER stresscombined with inflammation dysregulated macrophage ironhomeostasis Therefore we examined the impact of ER stresscombined with inflammation on release of inflammatorymediators frommacrophages Using Luminex technology weevaluated cytokine and chemokine release from cells treatedwith tunicamycin (10120583gmL) to induce ER stress prior to LPS(40 ngmL) exposure for 6 hr to induce the cytokine stormAs expected LPS exposure without tunicamycin treatmentinduced massive release of cytokines (IL-1120573 TNF120572 IL-6 IL-12p40 IFN120572 IL-1R antagonist and IL-2R) and chemokines(RANTES MCP-1 MIP1120572 MIP1120573 IP-10CXCL10 and IL-8) compared to unstimulated THP-1 macrophages (Table 1)Tunicamycin treatment alone (without LPS exposure) alsoinduced cytokinechemokine release which was typically sim2-to 5-fold less than LPS treatment (Figures 8(a) 8(b) 8(c)and 8(d) and Table 1) Since ER stress downregulates proteinsynthesis machinery [55] tunicamycin treatment followed byLPS exposure led to significant reduction in cytokines (IL-12p40 IL-1R antagonist) and chemokines (MIP1120572 MIP1120573)compared to LPS exposure alone A slight reduction wasobserved in other cytokines and chemokines released frommacrophages exposed to both tunicamycin and LPS (Table 1)In contrast a significant and synergistic increase in IL-6releasewas observedwhenTHP-1macrophageswere exposedto both tunicamycin and LPS IL-6 is a proinflammatory
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
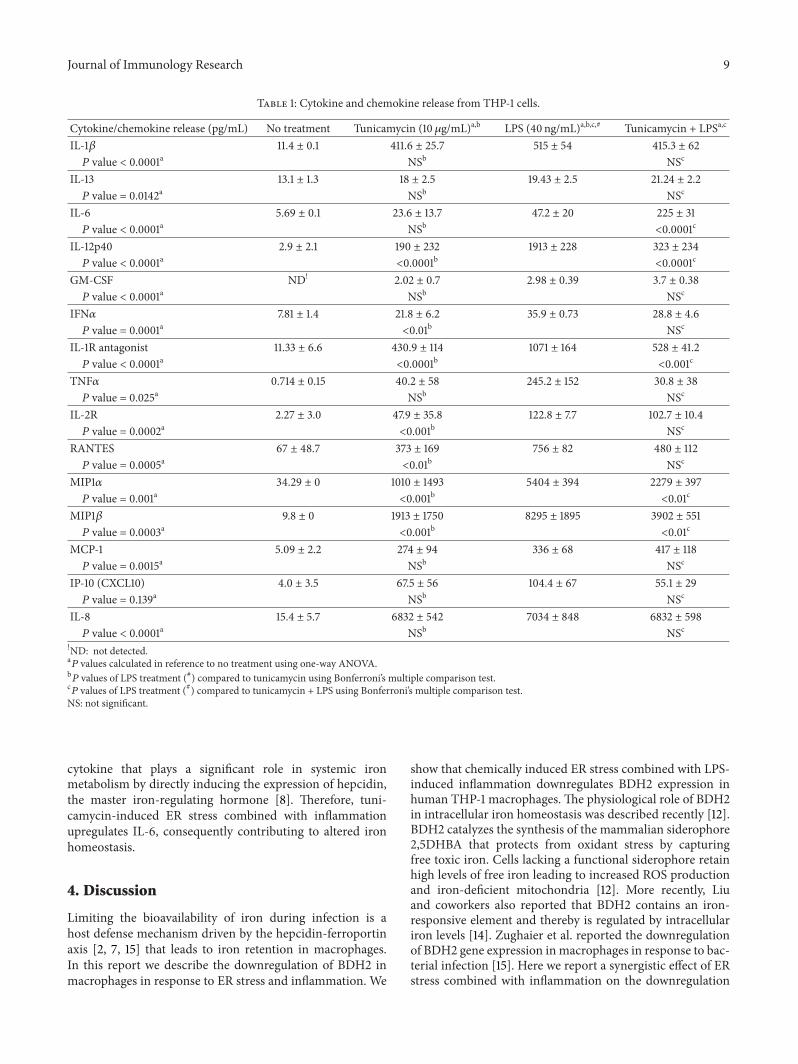
Journal of Immunology Research 9
Table 1 Cytokine and chemokine release from THP-1 cells
Cytokinechemokine release (pgmL) No treatment Tunicamycin (10 120583gmL)ab LPS (40 ngmL)abc Tunicamycin + LPSac
IL-1120573 114 plusmn 01 4116 plusmn 257 515 plusmn 54 4153 plusmn 62119875 value lt 00001a NSb NSc
IL-13 131 plusmn 13 18 plusmn 25 1943 plusmn 25 2124 plusmn 22119875 value = 00142a NSb NSc
IL-6 569 plusmn 01 236 plusmn 137 472 plusmn 20 225 plusmn 31119875 value lt 00001a NSb lt00001c
IL-12p40 29 plusmn 21 190 plusmn 232 1913 plusmn 228 323 plusmn 234119875 value lt 00001a lt00001b lt00001c
GM-CSF ND 202 plusmn 07 298 plusmn 039 37 plusmn 038119875 value lt 00001a NSb NSc
IFN120572 781 plusmn 14 218 plusmn 62 359 plusmn 073 288 plusmn 46119875 value = 00001a lt001b NSc
IL-1R antagonist 1133 plusmn 66 4309 plusmn 114 1071 plusmn 164 528 plusmn 412119875 value lt 00001a lt00001b lt0001c
TNF120572 0714 plusmn 015 402 plusmn 58 2452 plusmn 152 308 plusmn 38119875 value = 0025a NSb NSc
IL-2R 227 plusmn 30 479 plusmn 358 1228 plusmn 77 1027 plusmn 104119875 value = 00002a lt0001b NSc
RANTES 67 plusmn 487 373 plusmn 169 756 plusmn 82 480 plusmn 112119875 value = 00005a lt001b NSc
MIP1120572 3429 plusmn 0 1010 plusmn 1493 5404 plusmn 394 2279 plusmn 397119875 value = 0001a lt0001b lt001c
MIP1120573 98 plusmn 0 1913 plusmn 1750 8295 plusmn 1895 3902 plusmn 551119875 value = 00003a lt0001b lt001c
MCP-1 509 plusmn 22 274 plusmn 94 336 plusmn 68 417 plusmn 118119875 value = 00015a NSb NSc
IP-10 (CXCL10) 40 plusmn 35 675 plusmn 56 1044 plusmn 67 551 plusmn 29119875 value = 0139a NSb NSc
IL-8 154 plusmn 57 6832 plusmn 542 7034 plusmn 848 6832 plusmn 598119875 value lt 00001a NSb NSc
ND not detecteda119875 values calculated in reference to no treatment using one-way ANOVA
b119875 values of LPS treatment () compared to tunicamycin using Bonferronirsquos multiple comparison test
c119875 values of LPS treatment () compared to tunicamycin + LPS using Bonferronirsquos multiple comparison testNS not significant
cytokine that plays a significant role in systemic ironmetabolism by directly inducing the expression of hepcidinthe master iron-regulating hormone [8] Therefore tuni-camycin-induced ER stress combined with inflammationupregulates IL-6 consequently contributing to altered ironhomeostasis
4 Discussion
Limiting the bioavailability of iron during infection is ahost defense mechanism driven by the hepcidin-ferroportinaxis [2 7 15] that leads to iron retention in macrophagesIn this report we describe the downregulation of BDH2 inmacrophages in response to ER stress and inflammation We
show that chemically induced ER stress combined with LPS-induced inflammation downregulates BDH2 expression inhuman THP-1 macrophages The physiological role of BDH2in intracellular iron homeostasis was described recently [12]BDH2 catalyzes the synthesis of the mammalian siderophore25DHBA that protects from oxidant stress by capturingfree toxic iron Cells lacking a functional siderophore retainhigh levels of free iron leading to increased ROS productionand iron-deficient mitochondria [12] More recently Liuand coworkers also reported that BDH2 contains an iron-responsive element and thereby is regulated by intracellulariron levels [14] Zughaier et al reported the downregulationof BDH2 gene expression inmacrophages in response to bac-terial infection [15] Here we report a synergistic effect of ERstress combined with inflammation on the downregulation
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
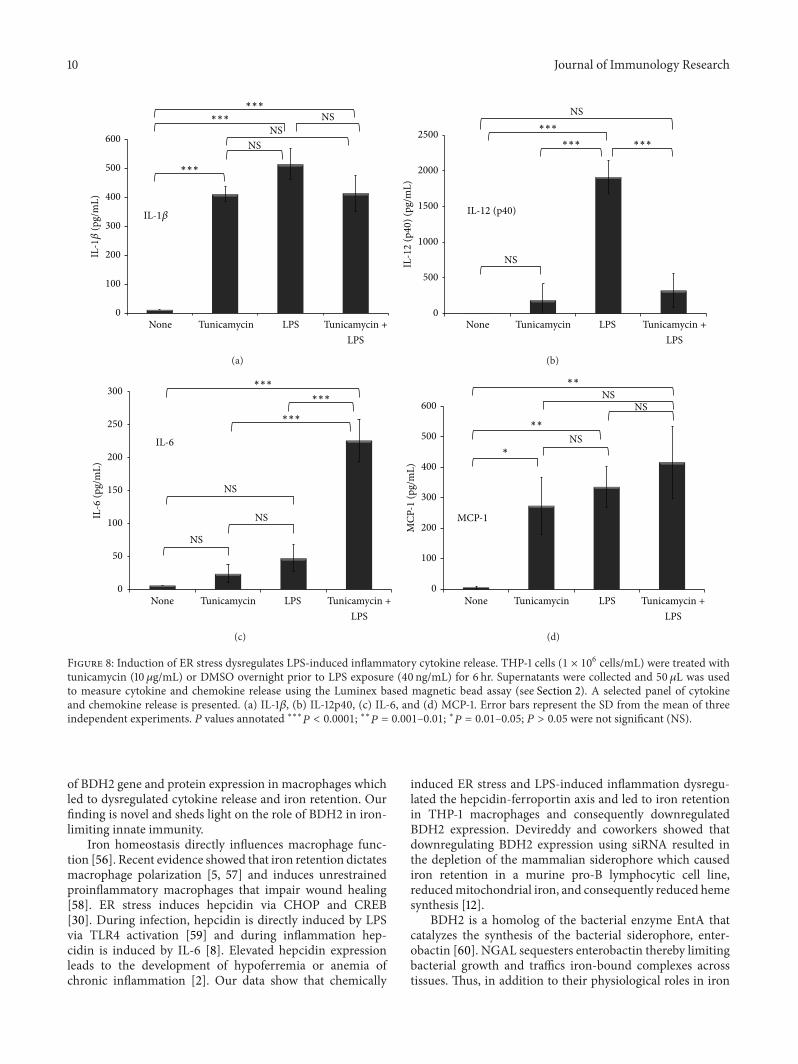
10 Journal of Immunology Research
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
IL-1120573
(pg
mL)
IL-1120573
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(a)
0
500
1000
1500
2000
2500
None Tunicamycin LPS
NS
NSIL-12
(p40
) (pg
mL)
IL-12 (p40)
lowastlowastlowast
lowastlowastlowast lowastlowastlowast
Tunicamycin +LPS
(b)
0
50
100
150
200
250
300
None Tunicamycin LPS
NS
NS
NSIL-6
(pg
mL)
IL-6
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Tunicamycin +LPS
(c)
0
100
200
300
400
500
600
None Tunicamycin LPS
NSNS
NS
lowastlowast
lowastlowast
lowast
MCP
-1(p
gm
L)
MCP-1
Tunicamycin +LPS
(d)
Figure 8 Induction of ER stress dysregulates LPS-induced inflammatory cytokine release THP-1 cells (1 times 106 cellsmL) were treated withtunicamycin (10120583gmL) or DMSO overnight prior to LPS exposure (40 ngmL) for 6 hr Supernatants were collected and 50 120583L was usedto measure cytokine and chemokine release using the Luminex based magnetic bead assay (see Section 2) A selected panel of cytokineand chemokine release is presented (a) IL-1120573 (b) IL-12p40 (c) IL-6 and (d) MCP-1 Error bars represent the SD from the mean of threeindependent experiments 119875 values annotated lowastlowastlowast119875 lt 00001 lowastlowast119875 = 0001ndash001 lowast119875 = 001ndash005 119875 gt 005 were not significant (NS)
of BDH2 gene and protein expression in macrophages whichled to dysregulated cytokine release and iron retention Ourfinding is novel and sheds light on the role of BDH2 in iron-limiting innate immunity
Iron homeostasis directly influences macrophage func-tion [56] Recent evidence showed that iron retention dictatesmacrophage polarization [5 57] and induces unrestrainedproinflammatory macrophages that impair wound healing[58] ER stress induces hepcidin via CHOP and CREB[30] During infection hepcidin is directly induced by LPSvia TLR4 activation [59] and during inflammation hep-cidin is induced by IL-6 [8] Elevated hepcidin expressionleads to the development of hypoferremia or anemia ofchronic inflammation [2] Our data show that chemically
induced ER stress and LPS-induced inflammation dysregu-lated the hepcidin-ferroportin axis and led to iron retentionin THP-1 macrophages and consequently downregulatedBDH2 expression Devireddy and coworkers showed thatdownregulating BDH2 expression using siRNA resulted inthe depletion of the mammalian siderophore which causediron retention in a murine pro-B lymphocytic cell linereducedmitochondrial iron and consequently reduced hemesynthesis [12]
BDH2 is a homolog of the bacterial enzyme EntA thatcatalyzes the synthesis of the bacterial siderophore enter-obactin [60] NGAL sequesters enterobactin thereby limitingbacterial growth and traffics iron-bound complexes acrosstissues Thus in addition to their physiological roles in iron
Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 11
Altered cellular iron homeostasis
Hepcidin
Detoxifyiron
BDH2
Upregulateshepcidin
DownregulatesBDH2
Controlsiron egress
ferroportin
LPS Tunicamycin
Iron retention
Macrophage functions
Iron recycling
Host defense
Synthesis ofmammaliansiderophore
ER stress and inflammation downregulate BDH2
times
25DHBA
Figure 9 Schematic presentation of BDH2 downregulation in macrophages BDH2 is the rate-limiting enzyme in the synthesis of themammalian siderophore 25DHBA that detoxifies and traffics intracellular labile iron Treatment of THP-1 macrophage-like monocytes withLPS or tunicamycin downregulates BDH2 expression leading to intracellular labile iron accumulation Hepcidin is the master iron regulatinghormone that binds to the only iron exporter ferroportin causing its internalization and degradation consequently preventing iron egressfrom macrophages LPS and tunicamycin treatment increases hepcidin expression and downregulates ferroportin causing iron retention inmacrophages The downregulation of BDH2 contributes to altered cellular iron homeostasis
homeostasis NGAL and BDH2 constitute the iron-limitingarm of innate immune defense Further NGAL is highlyinduced as an antioxidant response during oxidative stress[61] and in response to ER stress [45] Our data demonstratea synergistic effect of ER stress and inflammation on thedramatic upregulation of NGAL Elevated levels of NGALare associated with many chronic diseases like kidney dis-ease heart disease arthritis chronic obstructive pulmonarydisease and cancer [62ndash64] High levels of NGAL wereshown to deactivate macrophages and worsen outcomes ofpneumococcal pneumonia [65] We recently reported thatcirculating NGAL levels are elevated in subjects with cysticfibrosis compared to healthy donors and demonstrated thatperipheral monocytes secrete NGAL in response to LPSand to Pseudomonas aeruginosa infection [44] MoreoverNGAL expression is upregulated whereas BDH2 expressionis downregulated in macrophages upon bacterial infectionwith Neisseria gonorrhoeae [15] Altered iron homeostasismanifesting as hypoferremia is common across all thesediseases Taken together ER stress combined with inflam-mation downregulates BDH2 expression consequent to thedysregulation of hepcidin-ferroportin axis leading to ironretention in macrophages and dramatic NGAL upregulationwhich contribute to altered intracellular iron homeostasis(Figure 9)
ER stress has been shown to induce chronic inflammationin metabolic and chronic heart diseases [66 67] Tuni-camycin which inhibits protein glycosylation and triggers theunfolded protein response [68] combined with LPS syner-gistically increased IL-6 release frommacrophages Hepcidinexpression is directly induced by IL-6 Previous study showedthat treatment with anti-IL-6 antibody lowered hepcidin
levels which alleviated hypoferremia and restored iron home-ostasis [2 69] Amongmany inflammatorymediators IL-6 is apredictor of mortality in chronic kidney disease [70] in HIV[71] and in other chronic diseases associated with immuneactivation due to microbial translocation [72 73] Our resultssuggest that ER stress combined with inflammation increasesIL-6 that induces hepcidin expression leading to increasedintracellular iron which downregulates BDH2 expressionFurther NGAL possesses an immunomodulatory activityand exerts a suppressive effect on macrophages leading toreduced cytokine release [65 74] We observed a syner-gistic effect of ER stress combined with inflammation onthe dramatic upregulation of NGAL expression which maycontribute to dysregulated cytokine and chemokine responsein macrophages In this report we describe a mechanism bywhich ER stress combined with inflammation contributesto dysfunctional iron-limiting innate immune responses inmacrophages
Vitamin D possesses immunomodulatory activities andlowers circulating cytokines in vivo [47 48 75] Recentstudy demonstrated that vitamin D relieves ER stress inmacrophages [52] We reported that the hormonally activeform of vitamin D 125 dihydroxyvitamin D
3 reducedMCP-
1 release from macrophages treated with LPS or infectedwith Pseudomonas aeruginosa bacteria [47] More recentlyZughaier et al showed that 125 dihydroxyvitamin D
3treat-
ment reduced IL-6 release from LPS activated macrophagesand reduced hepcidin gene expression while upregulatingferroportin gene expression in a dose-dependent mannerthereby vitamin D restored the hepcidin-ferroportin axisin macrophages [50] Taken together our data show thatER stress and inflammation downregulate BDH2 expression
12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Journal of Immunology Research
O
Fe
Fe
O
O
OO
O
OOO
NN
N
N
O
OOO
O
O
O
OO
OO
OOOH
OH
OH
OO
OOO
O
OO
OH
H
HHN
HN
HO
HO
OH
OH
HN
NHNHN
H
Deferiprone Fe(III)-deferiprone
Enterobactin Fe(III)-enterobactin
Does NGAL bind Fe(III)-deferiprone
Scavenged iron chelates fit in NGALrsquos hydrophobic pocket
NGAL bound Fe(III)-enterobactin
Figure 10 A schematic presentation of proposed small molecule iron chelator acting similar to the mammalian siderophore 25-dihydroxybenzoic acid The DFP-iron complex shares structural resemblance to enterobactin the major bacterial siderophore that bindsiron with very high affinity As a defense mechanism the host upregulates NGAL expression to scavenge enterobactin which in turn limitsiron availability and inhibits bacterial growth DFP is an iron chelating small molecule drug that gains access to cells and scavenges freeintracellular iron and then exits as a DFP-iron complex Based on the structural resemblance to enterobactin-iron complexes the DFP-ironcomplex is expected to bind NGAL by fitting into the hydrophobic pocket [79] Therefore small molecule iron chelators may act as themammalian siderophore and redistribute iron retained in macrophages
and vitamin D treatment restored BDH2 expression in LPSactivated macrophages This is the first report to show thatvitaminD restores BDH2 expression It is not knownwhethervitaminDexerts direct or indirect regulatory effects onBDH2gene expression but it is worthy of further investigation
The important role of BDH2 in intracellular homeostasisrelies on the production of the mammalian siderophore 25-DHBA that binds free iron and thereby restores cellular ironhomeostasis [17] We propose that the synthetic iron chelat-ing drug Deferiprone (DFP 3-hydroxy-12-dimethylpyridin-4(1H)-one) [76 77] commonly used to treat iron overloaddisorders [78] may emulate the mammalian siderophore andredistribute iron retained in macrophages DFP is a smallmolecule that diffuses across the cellular membrane andscavenges free intracellular iron Three molecules of DFPmake a complexwith one ferric ironmolecule [76]ThisDFP-iron complex shares structural resemblance to enterobactinthe major bacterial siderophore that binds iron with veryhigh affinity (Figure 10) In response to infection and as adefense mechanism the host upregulates NGAL expressionto scavenge enterobactin which in turn limits iron availabilityand inhibits bacterial growth Our data show that NGALis dramatically upregulated in response to ER stress and
inflammation We hypothesize that upon downregulationof BDH2 expression in macrophages DFP may compensatefor the reduction in mammalian siderophore productionDFP gains access to cells and scavenges free intracellulariron and then exits as a DFP-iron complex Based on thestructural resemblance to enterobactin-iron complexes theDFP-iron complex is expected to bind NGAL by fitting intothe hydrophobic pocket [79] (Figure 10) NGAL-siderophorecomplexes are then excreted to urine Therefore DFP couldbe a potential therapeutic for altered cellular iron homeostasisthat lowers both intracellular iron and abnormal NGAL lev-els consequently restoring intracellular iron homeostasis inmacrophages Our hypothesis is under current investigation
5 Conclusion
Inflammation combined with ER stress led to massiveBDH2 downregulation and increased the expression ofER stress markers upregulated hepcidin expression down-regulated ferroportin expression caused iron retentionin macrophages and dysregulated cytokine release from
Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 13
macrophages ER stress combined with inflammation syn-ergistically upregulated the expression of the iron carrierprotein NGAL and the stress-inducible heme degradingenzyme heme oxygenase-1 (HO-1) leading to iron liberationHowever vitaminD treatment prior to LPS exposure restoredBDH2 expression BDH2 is expressed in many types ofcells [80] therefore downregulation of BDH2 may havedetrimental effects on cellular functions Downregulation ofBDH2 during infection is due to activation of hepcidin andintracellular iron retention indicating an important role forBDH2 in iron-limiting innate immunity and host defenseOur data suggest that continued ER stress combined withinflammation impacts on BDH2 expression and contributesto altered intracellular iron homeostasis
Abbreviations
ER stress Endoplasmic reticulum stressLPS LipopolysaccharideBDH2 Butyrate dehydrogenase type 2FPN1 FerroportinNGAL Neutrophil gelatinase-associated
lipocalinUPR Unfolded protein responseCHOP Transcription factor CEBP homolog
proteinHsXBP-1 X-box binding protein 1 spiced isoformGRP78 Glucose regulating protein 78ATF6 Activating transcription factor 6MTT Thiazolyl blue tetrazolium bromideDHBA Dihydroxybenzoic acidROS Reactive oxygen speciesDMSO DimethylsulfoxideIL-1120573 Interleukin 1 betaTNF120572 Tumor necrosis factor alphaIL-13 Interleukin 13IL-6 Interleukin 6IL-12(p40) Interleukin 12 subunit 40GM-CSF Granulocyte macrophage
colony-stimulating factorINF120572 Interferon alphaIL-1R antagonist Interleukin 1 receptor antagonistIL-2R Interleukin 2 receptorMIP1120572 Macrophage inflammatory protein 1
alphaMIP1120573 Macrophage inflammatory protein 1
betaMCP-1 Monocyte chemotactic proteinIP-10 Inducible small protein 10IL-8 Interleukin 8
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Susu M Zughaier conceived and designed the experimentsSusu M Zughaier and Brandon B Stauffer performed theexperiments Susu M Zughaier and Brandon B Stauffer ana-lyzed the data Susu M Zughaier and Nael A McCarty con-tributed reagentsmaterialsanalysis tools Susu M Zughaierwrote the paper Susu M Zughaier Brandon B Staufferand Nael A McCarty critically reviewed the paper Susu MZughaier is the principal investigator
Acknowledgments
This work was supported by an Emory-Egleston ChildrenrsquosResearch Center grant to Susu M Zughaier The authorsthank Dana Guentert for technical assistance
References
[1] JWang andK Pantopoulos ldquoRegulation of cellular ironmetab-olismrdquo Biochemical Journal vol 434 no 3 pp 365ndash381 2011
[2] T Ganz and E Nemeth ldquoHepcidin and disorders of ironmetab-olismrdquo Annual Review of Medicine vol 62 pp 347ndash360 2011
[3] P Lipinski R R Starzynski A Stys and M Stracilo ldquoIronhomeostasis a defense mechanism in oxidative stressrdquo PostepyBiochemii vol 56 pp 305ndash316 2010
[4] X Pan B Tamilselvam E J Hansen and S Daefler ldquoModula-tion of iron homeostasis in macrophages by bacterial intracel-lular pathogensrdquo BMCMicrobiology vol 10 article 64 2010
[5] S Recalcati M Locati A Marini et al ldquoDifferential regulationof iron homeostasis during human macrophage polarizedactivationrdquo European Journal of Immunology vol 40 no 3 pp824ndash835 2010
[6] T Ganz ldquoThe role of hepcidin in iron sequestration duringinfections and in the pathogenesis of anemia of chronic diseaserdquoThe Israel Medical Association Journal IMAJ vol 4 no 11 pp1043ndash1045 2002
[7] E Nemeth M S Tuttle J Powelson et al ldquoHepcidin regulatescellular iron efflux by binding to ferroportin and inducing itsinternalizationrdquo Science vol 306 no 5704 pp 2090ndash20932004
[8] E Nemeth S Rivera V Gabayan et al ldquoIL-6 mediates hypo-ferremia of inflammation by inducing the synthesis of theiron regulatory hormone hepcidinrdquo The Journal of ClinicalInvestigation vol 113 no 9 pp 1271ndash1276 2004
[9] E Nemeth E V Valore M Territo G Schiller A Lichtensteinand T Ganz ldquoHepcidin a putative mediator of anemia ofinflammation is a type II acute-phase proteinrdquo Blood vol 101no 7 pp 2461ndash2463 2003
[10] MTheurl ITheurl KHochegger et al ldquoKupffer cellsmodulateiron homeostasis inmice via regulation of hepcidin expressionrdquoJournal of Molecular Medicine vol 86 no 7 pp 825ndash835 2008
[11] I Theurl M Theurl M Seifert et al ldquoAutocrine formation ofhepcidin induces iron retention in human monocytesrdquo Bloodvol 111 no 4 pp 2392ndash2399 2008
[12] L R Devireddy D O Hart D H Goetz and M R GreenldquoA mammalian siderophore synthesized by an enzyme with abacterial homolog involved in enterobactin productionrdquo Cellvol 141 no 6 pp 1006ndash1017 2010
14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

14 Journal of Immunology Research
[13] K Guo P Lukacik E Papagrigoriou et al ldquoCharacteriza-tion of human DHRS6 an orphan short chain dehydro-genasereductase enzyme a novel cytosolic type 2 R-120573-hydroxybutyrate dehydrogenaserdquo The Journal of BiologicalChemistry vol 281 no 15 pp 10291ndash10297 2006
[14] Z Liu R Lanford S Mueller et al ldquoSiderophore-mediated irontrafficking in humans is regulated by ironrdquo Journal of MolecularMedicine vol 90 no 10 pp 1209ndash1221 2012
[15] S M Zughaier J L Kandler and W M Shafer ldquoNeisseriagonorrhoeae modulates iron-limiting innate immune defensesin macrophagesrdquo PLoS ONE vol 9 no 1 Article ID e876882014
[16] Z Liu S Reba W-D Chen et al ldquoRegulation of mammaliansiderophore 25-DHBA in the innate immune response toinfectionrdquo Journal of Experimental Medicine vol 211 no 6 pp1197ndash1213 2014
[17] Z Liu A Ciocea and L Devireddy ldquoEndogenous siderophore25-dihydroxybenzoic acid deficiency promotes anemia andsplenic iron overload in micerdquo Molecular and Cellular Biologyvol 34 no 13 pp 2533ndash2546 2014
[18] Z Liu K K Velpula and L Devireddy ldquo3-Hydroxybutyratedehydrogenase-2 and ferritin-H synergistically regulate intra-cellular ironrdquoThe FEBS Journal vol 281 no 10 pp 2410ndash24212014
[19] AAgarwal and S Bolisetty ldquoAdaptive responses to tissue injuryrole of heme oxygenase-1rdquoTransactions of the American Clinicaland Climatological Association vol 124 pp 111ndash122 2013
[20] TDHull S Bolisetty ACDealmeida et al ldquoHemeoxygenase-1 expression protects the heart from acute injury caused byinducible Cre recombinaserdquo Laboratory Investigation vol 93no 8 pp 868ndash879 2013
[21] P Mandal S Roychowdhury P-H Park B T Pratt T Rogerand L E Nagy ldquoAdiponectin and heme oxygenase-1 suppressTLR4MyD88-independent signaling in rat Kupffer cells and inmice after chronic ethanol exposurerdquo The Journal of Immunol-ogy vol 185 no 8 pp 4928ndash4937 2010
[22] P Mandal M T Pritchard and L E Nagy ldquoAnti-inflammatorypathways and alcoholic liver disease role of an adiponectininterleukin-10heme oxygenase-1 pathwayrdquo World Journal ofGastroenterology vol 16 no 11 pp 1330ndash1336 2010
[23] M Packiam H Wu S J Veit N Mavrogiorgos A E Jerseand R R Ingalls ldquoProtective role of Toll-like receptor 4 inexperimental gonococcal infection of female micerdquo MucosalImmunology vol 5 no 1 pp 19ndash29 2012
[24] J Mishra C Dent R Tarabishi et al ldquoNeutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renalinjury after cardiac surgeryrdquoThe Lancet vol 365 no 9466 pp1231ndash1238 2005
[25] S Chakraborty S Kaur S Guha and S K Batra ldquoThe multi-faceted roles of neutrophil gelatinase associated lipocalin(NGAL) in inflammation and cancerrdquo Biochimica et BiophysicaActa Reviews on Cancer vol 1826 no 1 pp 129ndash169 2012
[26] G Bao M Clifton T M Hoette et al ldquoIron traffics in circu-lation bound to a siderocalin (Ngal)mdashcatechol complexrdquoNatureChemical Biology vol 6 no 8 pp 602ndash609 2010
[27] D H Goetz M A Holmes N Borregaard M E Bluhm K NRaymond and R K Strong ldquoThe neutrophil lipocalin NGAL isa bacteriostatic agent that interferes with siderophore-mediatediron acquisitionrdquo Molecular Cell vol 10 no 5 pp 1033ndash10432002
[28] T H Flo K D Smith S Sato et al ldquoLipocalin 2 mediates aninnate immune response to bacterial infection by sequestratingironrdquo Nature vol 432 no 7019 pp 917ndash921 2004
[29] M E Peek A Bhatnagar N A McCarty and S M ZughaierldquoPyoverdine the Major Siderophore in Pseudomonas aerugi-nosa Evades NGAL Recognitionrdquo Interdisciplinary Perspectiveson Infectious Diseases vol 2012 Article ID 843509 10 pages2012
[30] M de Sousa S J Oliveira and J P Pinto ldquoER stress and ironhomeostasis a new frontier for theUPRrdquo Biochemistry ResearchInternational vol 2011 Article ID 896474 10 pages 2011
[31] J-J Yin Y-B Li Y Wang et al ldquoThe role of autophagy inendoplasmic reticulum stress-induced pancreatic 120573 cell deathrdquoAutophagy vol 8 no 2 pp 158ndash164 2012
[32] D H Suh M-K Kim H S Kim H H Chung and Y SSong ldquoUnfolded protein response to autophagy as a promisingdruggable target for anticancer therapyrdquoAnnals of the New YorkAcademy of Sciences vol 1271 no 1 pp 20ndash32 2012
[33] C Vecchi G Montosi K Zhang et al ldquoER stress controls ironmetabolism through induction of hepcidinrdquo Science vol 325no 5942 pp 877ndash880 2009
[34] S J Oliveira J P Pinto G Picarote et al ldquoER stress-induciblefactor CHOP affects the expression of hepcidin by modulatingCEBPalpha activityrdquo PLoS ONE vol 4 no 8 Article ID e66182009
[35] R Vidal B Caballero A Couve and C Hetz ldquoConvergingpathways in the occurrence of endoplasmic reticulum (ER)stress in Huntingtonrsquos diseaserdquoCurrentMolecularMedicine vol11 no 1 pp 1ndash12 2011
[36] S Matus L H Glimcher and C Hetz ldquoProtein folding stressin neurodegenerative diseases a glimpse into the ERrdquo CurrentOpinion in Cell Biology vol 23 no 2 pp 239ndash252 2011
[37] X Qu Z Zou Q Sun et al ldquoAutophagy gene-dependentclearance of apoptotic cells during embryonic developmentrdquoCell vol 128 no 5 pp 931ndash946 2007
[38] S M Zughaier Y-L Tzeng S M Zimmer A Datta RW Carl-son and D S Stephens ldquoNeisseria meningitidis lipooligosac-charide structure-dependent activation of the macrophageCD14Toll-like receptor 4 pathwayrdquo Infection and Immunity vol72 no 1 pp 371ndash380 2004
[39] S M Zughaier ldquoNeisseria meningitidis capsular polysaccha-rides induce inflammatory responses via TLR2 and TLR4-MD-2rdquo Journal of Leukocyte Biology vol 89 no 3 pp 469ndash480 2011
[40] O Kakhlon and Z I Cabantchik ldquoThe labile iron poolcharacterization measurement and participation in cellularprocessesrdquo Free Radical Biology amp Medicine vol 33 no 8 pp1037ndash1046 2002
[41] D Gerlier and N Thomasset ldquoUse of MTT colorimetric assayto measure cell activationrdquo Journal of Immunological Methodsvol 94 no 1-2 pp 57ndash63 1986
[42] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[43] Y Nakayama M Endo H Tsukano M Mori Y Oike andT Gotoh ldquoMolecular mechanisms of the LPS-induced non-apoptotic ER stress-CHOP pathwayrdquo The Journal of Biochem-istry vol 147 no 4 pp 471ndash483 2010
[44] S M Zughaier V Tangpricha T Leong A A Stecenko andN A McCarty ldquoPeripheral monocytes derived from patientswith cystic fibrosis and healthy donors secrete ngal in response
Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 15
to Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[45] N R Mahadevan J Rodvold G Almanza A F Perez M CWheeler and M Zanetti ldquoER stress drives Lipocalin 2 upregu-lation in prostate cancer cells in an NF-120581B-dependent mannerrdquoBMC Cancer vol 11 article 229 2011
[46] S Immenschuh E Baumgart-Vogt and S Mueller ldquoHemeoxygenase-1 and iron in liver inflammation a complex alliancerdquoCurrent Drug Targets vol 11 no 12 pp 1541ndash1550 2010
[47] J A Alvarez S M Zughaier J Law et al ldquoEffects of high-dosecholecalciferol on serum markers of inflammation and immu-nity in patients with early chronic kidney diseaserdquo EuropeanJournal of Clinical Nutrition vol 67 no 3 pp 264ndash269 2013
[48] R E Grossmann S M Zughaier S Liu R H Lyles and VTangpricha ldquoImpact of vitamin D supplementation on markersof inflammation in adults with cystic fibrosis hospitalizedfor a pulmonary exacerbationrdquo European Journal of ClinicalNutrition vol 66 no 9 pp 1072ndash1074 2012
[49] J A Alvarez J Law K E Coakley et al ldquoHigh-dose cholecalcif-erol reduces parathyroid hormone in patients with early chronickidney disease a pilot randomized double-blind placebo-controlled trialrdquoTheAmerican Journal of Clinical Nutrition vol96 no 3 pp 672ndash679 2012
[50] S M Zughaier J A Alvarez J H Sloan R J Konrad andV Tangpricha ldquoThe role of vitamin D in regulating the iron-hepcidin-ferroportin axis inmonocytesrdquo Journal of Clinical andTranslational Endocrinology vol 1 no 1 pp e19ndashe25 2014
[51] A E Riek J Oh and C Bernal-Mizrachi ldquo125(OH)2
vitaminD suppresses macrophage migration and reverses atherogeniccholesterol metabolism in type 2 diabetic patientsrdquoThe Journalof Steroid Biochemistry andMolecular Biology vol 136 no 1 pp309ndash312 2013
[52] A E Riek J Oh J E Sprague et al ldquoVitamin D suppressionof endoplasmic reticulum stress promotes an antiatherogenicmonocytemacrophage phenotype in type 2 diabetic patientsrdquoJournal of Biological Chemistry vol 287 no 46 pp 38482ndash38494 2012
[53] C G Leon R Tory J Jia O Sivak and K M WasanldquoDiscovery and development of toll-like receptor 4 (TLR4)antagonists a new paradigm for treating sepsis and otherdiseasesrdquo Pharmaceutical Research vol 25 no 8 pp 1751ndash17612008
[54] T M El-Achkar M Hosein and P C Dagher ldquoPathways ofrenal injury in systemic gram-negative sepsisrdquoEuropean Journalof Clinical Investigation vol 38 no 2 pp 39ndash44 2008
[55] S Deegan S Saveljeva A M Gorman and A Samali ldquoStress-induced self-cannibalism on the regulation of autophagy byendoplasmic reticulum stressrdquo Cellular and Molecular LifeSciences vol 70 no 14 pp 2425ndash2441 2013
[56] G Cairo S Recalcati A Mantovani and M Locati ldquoIrontrafficking andmetabolism inmacrophages contribution to thepolarized phenotyperdquo Trends in Immunology vol 32 no 6 pp241ndash247 2011
[57] G Corna L Campana E Pignatti et al ldquoPolarization dictatesiron handling by inflammatory and alternatively activatedmacrophagesrdquo Haematologica vol 95 no 11 pp 1814ndash18222010
[58] A Sindrilaru T Peters S Wieschalka et al ldquoAn unrestrainedproinflammatory M1 macrophage population induced by ironimpairs wound healing in humans and micerdquo The Journal ofClinical Investigation vol 121 no 3 pp 985ndash997 2011
[59] C Peyssonnaux A S Zinkernagel V Datta X Lauth R SJohnson and V Nizet ldquoTLR4-dependent hepcidin expressionby myeloid cells in response to bacterial pathogensrdquo Blood vol107 no 9 pp 3727ndash3732 2006
[60] R J Abergel M C Clifton J C Pizarro et al ldquoThe siderocalinenterobactin interaction a link between mammalian immunityand bacterial iron transportrdquo Journal of the American ChemicalSociety vol 130 no 34 pp 11524ndash11534 2008
[61] P Bahmani R Halabian M Rouhbakhsh et al ldquoNeutrophilgelatinase-associated lipocalin induces the expression of hemeoxygenase-1 and superoxide dismutase 1 2rdquo Cell Stress andChaperones vol 15 no 4 pp 395ndash403 2010
[62] A Yndestad L Landroslash T Ueland et al ldquoIncreased systemicand myocardial expression of neutrophil gelatinase-associatedlipocalin in clinical and experimental heart failurerdquo EuropeanHeart Journal vol 30 no 10 pp 1229ndash1236 2009
[63] H Zhang L Xu D Xiao et al ldquoUpregulation of neutrophilgelatinase-associated lipocalin in oesophageal squamous cellcarcinoma significant correlation with cell differentiation andtumour invasionrdquo Journal of Clinical Pathology vol 60 no 5pp 555ndash561 2007
[64] S M Zughaier V Tangpricha T Leong A A Stecenko and NA McCarty ldquoPeripheral monocytes derived from patients withcystic fibrosis and healthy donors secrete ngal in responseto Pseudomonas aeruginosa infectionrdquo Journal of InvestigativeMedicine vol 61 no 6 pp 1018ndash1025 2013
[65] J MWarszawska R Gawish O Sharif et al ldquoLipocalin 2 deac-tivates macrophages and worsens pneumococcal pneumoniaoutcomesrdquo Journal of Clinical Investigation vol 123 no 8 pp3363ndash3372 2013
[66] N Kawasaki R Asada A Saito S Kanemoto and K ImaizumildquoObesity-induced endoplasmic reticulum stress causes chronicinflammation in adipose tissuerdquo Scientific Reports vol 2 article799 2012
[67] T Gotoh M Endo and Y Oike ldquoEndoplasmic reticulumstress-related inflammation and cardiovascular diseasesrdquo Inter-national Journal of Inflammation vol 2011 Article ID 259462 8pages 2011
[68] H Zhang S Nakajima H Kato et al ldquoSelective potent block-ade of the IRE1 and ATF6 pathways by 4-phenylbutyric acidanaloguesrdquo British Journal of Pharmacology vol 170 no 4 pp822ndash834 2013
[69] F-D Wang D-B Zhou S-L Li et al ldquoEffect of recombi-nant human erythropoietin on hepcidin mRNA expression inpatients with multiple myelomardquo Zhongguo Shi Yan Xue Ye XueZa Zhi vol 19 no 2 pp 390ndash394 2011
[70] R Pecoits-Filho P Barany B Lindholm O Heimburger andP Stenvinkel ldquoInterleukin-6 is an independent predictor ofmortality in patients starting dialysis treatmentrdquo NephrologyDialysis Transplantation vol 17 no 9 pp 1684ndash1688 2002
[71] B McDonald S Moyo L Gabaitiri et al ldquoPersistently elevatedserum interleukin-6 predicts mortality among adults receivingcombination antiretroviral therapy in Botswana results from aclinical trialrdquo AIDS Research and Human Retroviruses vol 29no 7 pp 993ndash999 2013
[72] R P Andrie U M Becher R Frommold et al ldquoInterleukin-6is the strongest predictor of 30-day mortality in patients withcardiogenic shock due to myocardial infarctionrdquo Critical Carevol 16 no 4 article R152 2012
[73] K M Erlandson A A Allshouse C M Jankowski et alldquoAssociation of functional impairment with inflammation and
16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

16 Journal of Immunology Research
immune activation inHIV type 1-infected adults receiving effec-tive antiretroviral therapyrdquo The Journal of Infectious Diseasesvol 208 no 2 pp 249ndash259 2013
[74] J Zhang Y Wu Y Zhang D Leroith D A Bernlohr and XChen ldquoThe role of lipocalin 2 in the regulation of inflammationin adipocytes and macrophagesrdquoMolecular Endocrinology vol22 no 6 pp 1416ndash1426 2008
[75] B M Dixon T Barker T McKinnon et al ldquoPositive corre-lation between circulating cathelicidin antimicrobial peptide(hCAP18LL-37) and 25-hydroxyvitamin D levels in healthyadultsrdquo BMC Research Notes vol 5 article 575 2012
[76] V Wiwanitkit ldquoQuantum chemical analysis of the deferiprone-iron binding reactionrdquo International Journal of Nanomedicinevol 1 no 1 pp 111ndash113 2006
[77] V M Nurchi G Crisponi T Pivetta M Donatoni and MRemelli ldquoPotentiometric spectrophotometric and calorimetricstudy on iron(III) and copper(II) complexes with 12-dimethyl-3-hydroxy-4-pyridinonerdquo Journal of Inorganic Biochemistryvol 102 no 4 pp 684ndash692 2008
[78] G J Kontoghiorghes A Spyrou and A Kolnagou ldquoIron chela-tion therapy in hereditary hemochromatosis and thalassemiaintermedia regulatory and non regulatory mechanisms ofincreased iron absorptionrdquo Hemoglobin vol 34 no 3 pp 251ndash264 2010
[79] M A Fischbach H Lin D R Liu and C TWalsh ldquoHow path-ogenic bacteria evade mammalian sabotage in the battle forironrdquo Nature Chemical Biology vol 2 no 3 pp 132ndash138 2006
[80] H T Chang L K Olson and K A Schwartz ldquoKetolytic andglycolytic enzymatic expression profiles in malignant gliomasimplication for ketogenic diet therapyrdquo Nutrition andMetabolism vol 10 no 1 article 47 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom