Embed Size (px)
Citation preview

Research ArticlePlant Natural Product Formononetin Protects RatCardiomyocyte H9c2 Cells against Oxygen GlucoseDeprivation and Reoxygenation via Inhibiting ROSFormation and Promoting GSK-3120573 Phosphorylation
Yuanyuan Cheng1 Zhengyuan Xia2 Yifan Han3 and Jianhui Rong1
1School of Chinese Medicine Li Ka Shing Faculty of Medicine The University of Hong Kong 10 Sassoon Road Pokfulam Hong Kong2Department of Anesthesiology Li Ka Shing Faculty of Medicine The University of Hong Kong 10 Sassoon RoadPokfulam Hong Kong3Department of Applied Biology and Chemical Technology Hong Kong Polytechnic University Hung Hom Hong Kong
Correspondence should be addressed to Jianhui Rong jronghkucchkuhk
Received 23 October 2015 Revised 2 December 2015 Accepted 6 December 2015
Academic Editor Kota V Ramana
Copyright copy 2016 Yuanyuan Cheng et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
The opening of mitochondrial permeability transition pore (mPTP) is a major cause of cell death in ischemia reperfusion injuryBased on our pilot experiments plant natural product formononetin enhanced the survival of rat cardiomyocyte H9c2 cellsduring oxygen glucose deprivation (OGD) and reoxygenation For mechanistic studies we focused on two major cellular factorsnamely reactive oxygen species (ROS) and glycogen synthase kinase 3120573 (GSK-3120573) in the regulation of mPTP opening We foundthat formononetin suppressed the formation of ROS and superoxide in a concentration-dependent manner Formononetin alsorescued OGDreoxygenation-induced loss of mitochondrial membrane integrity Further studies suggested that formononetininduced Akt activation and GSK-3120573 (Ser9) phosphorylation thereby reducing GSK-3120573 activity towards mPTP opening PI3Kand PKC inhibitors abolished the effects of formononetin on mPTP opening and GSK-3120573 phosphorylation Immunoprecipitationexperiments further revealed that formononetin increased the binding of phosphor-GSK-3120573 to adenine nucleotide translocase(ANT) while it disrupted the complex of ANT with cyclophilin D Moreover immunofluorescence revealed that phospho-GSK-3120573 (Ser9) was mainly deposited in the space between mitochondria and cell nucleus Collectively these results indicated thatformononetin protected cardiomyocytes from OGDreoxygenation injury via inhibiting ROS formation and promoting GSK-3120573phosphorylation
1 Introduction
Acute myocardial infarction (AMI) is a leading cause ofhuman death worldwide [1] Timely reperfusion effectivelyreduces short-term mortality in early-reperfusion phase [2]One of the on-going issues is that reperfusion itself leads toadditional injury causing all forms of cell death and con-tractile dysfunction of surviving cells [3] The mitochondrialpermeability transition pore (mPTP) is implicated in thepathogenesis of myocardial ischemia-reperfusion injury [4]Ischemia induces the close of mPTP whereas reperfusion
promotes the mPTP opening [5] Thus control of mPTPopening at the early reperfusion is important to protect theheart against reperfusion injury [6] Among various regula-tors of mPTP opening glycogen synthase kinase 3120573 (GSK-3120573) is a constitutively active serine or threonine proteinkinase [7] GSK-3120573 stimulates themPTPopening and therebyinduces mitochondrial dysfunctions in myocardial ischemiareperfusion [8] The enzymatic activity of GSK-3120573 is regu-lated by phosphorylation Phosphorylation at tyrosine 216increases the activity whereas phosphorylation at serine 9significantly decreases the enzymatic activity of GSK-3120573
Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2016 Article ID 2060874 11 pageshttpdxdoiorg10115520162060874
2 Oxidative Medicine and Cellular Longevity
HO O
OOCH3
(a)
0
20
40
60
80
100
120
140
Cel
l via
bilit
y (
)
0 5 10 30 Formononetin (120583M)
NormalOGDR
lowastlowast lowastlowast lowastlowast
(b)
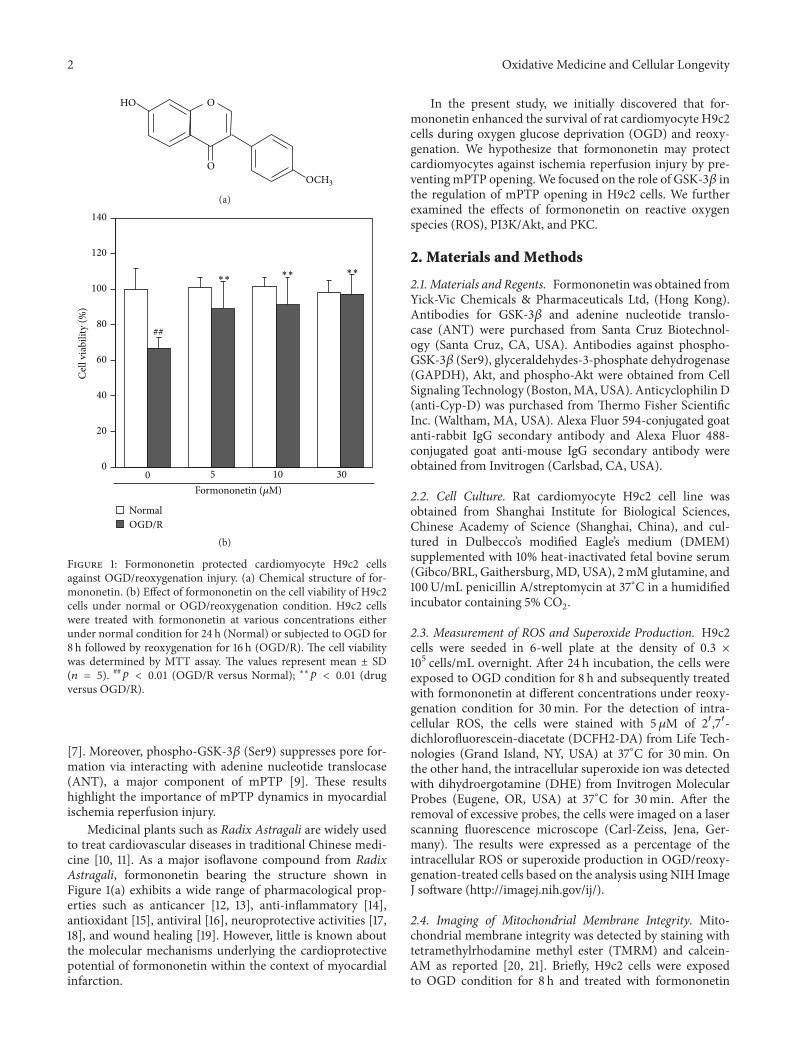
Figure 1 Formononetin protected cardiomyocyte H9c2 cellsagainst OGDreoxygenation injury (a) Chemical structure of for-mononetin (b) Effect of formononetin on the cell viability of H9c2cells under normal or OGDreoxygenation condition H9c2 cellswere treated with formononetin at various concentrations eitherunder normal condition for 24 h (Normal) or subjected to OGD for8 h followed by reoxygenation for 16 h (OGDR) The cell viabilitywas determined by MTT assay The values represent mean plusmn SD(119899 = 5) 119901 lt 001 (OGDR versus Normal) lowastlowast119901 lt 001 (drugversus OGDR)
[7] Moreover phospho-GSK-3120573 (Ser9) suppresses pore for-mation via interacting with adenine nucleotide translocase(ANT) a major component of mPTP [9] These resultshighlight the importance of mPTP dynamics in myocardialischemia reperfusion injury
Medicinal plants such as Radix Astragali are widely usedto treat cardiovascular diseases in traditional Chinese medi-cine [10 11] As a major isoflavone compound from RadixAstragali formononetin bearing the structure shown inFigure 1(a) exhibits a wide range of pharmacological prop-erties such as anticancer [12 13] anti-inflammatory [14]antioxidant [15] antiviral [16] neuroprotective activities [1718] and wound healing [19] However little is known aboutthe molecular mechanisms underlying the cardioprotectivepotential of formononetin within the context of myocardialinfarction
In the present study we initially discovered that for-mononetin enhanced the survival of rat cardiomyocyte H9c2cells during oxygen glucose deprivation (OGD) and reoxy-genation We hypothesize that formononetin may protectcardiomyocytes against ischemia reperfusion injury by pre-venting mPTP openingWe focused on the role of GSK-3120573 inthe regulation of mPTP opening in H9c2 cells We furtherexamined the effects of formononetin on reactive oxygenspecies (ROS) PI3KAkt and PKC
2 Materials and Methods
21 Materials and Regents Formononetin was obtained fromYick-Vic Chemicals amp Pharmaceuticals Ltd (Hong Kong)Antibodies for GSK-3120573 and adenine nucleotide translo-case (ANT) were purchased from Santa Cruz Biotechnol-ogy (Santa Cruz CA USA) Antibodies against phospho-GSK-3120573 (Ser9) glyceraldehydes-3-phosphate dehydrogenase(GAPDH) Akt and phospho-Akt were obtained from CellSignaling Technology (BostonMAUSA) AnticyclophilinD(anti-Cyp-D) was purchased from Thermo Fisher ScientificInc (Waltham MA USA) Alexa Fluor 594-conjugated goatanti-rabbit IgG secondary antibody and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody wereobtained from Invitrogen (Carlsbad CA USA)
22 Cell Culture Rat cardiomyocyte H9c2 cell line wasobtained from Shanghai Institute for Biological SciencesChinese Academy of Science (Shanghai China) and cul-tured in Dulbeccorsquos modified Eaglersquos medium (DMEM)supplemented with 10 heat-inactivated fetal bovine serum(GibcoBRL Gaithersburg MD USA) 2mM glutamine and100UmL penicillin Astreptomycin at 37∘C in a humidifiedincubator containing 5 CO
2
23 Measurement of ROS and Superoxide Production H9c2cells were seeded in 6-well plate at the density of 03 times105 cellsmL overnight After 24 h incubation the cells wereexposed to OGD condition for 8 h and subsequently treatedwith formononetin at different concentrations under reoxy-genation condition for 30min For the detection of intra-cellular ROS the cells were stained with 5120583M of 2101584071015840-dichlorofluorescein-diacetate (DCFH2-DA) from Life Tech-nologies (Grand Island NY USA) at 37∘C for 30min Onthe other hand the intracellular superoxide ion was detectedwith dihydroergotamine (DHE) from Invitrogen MolecularProbes (Eugene OR USA) at 37∘C for 30min After theremoval of excessive probes the cells were imaged on a laserscanning fluorescence microscope (Carl-Zeiss Jena Ger-many) The results were expressed as a percentage of theintracellular ROS or superoxide production in OGDreoxy-genation-treated cells based on the analysis using NIH ImageJ software (httpimagejnihgovij)
24 Imaging of Mitochondrial Membrane Integrity Mito-chondrial membrane integrity was detected by staining withtetramethylrhodamine methyl ester (TMRM) and calcein-AM as reported [20 21] Briefly H9c2 cells were exposedto OGD condition for 8 h and treated with formononetin
Oxidative Medicine and Cellular Longevity 3
(5 to 30 120583M) under reoxygenation condition for another20min H9c2 cells were incubated with 250 nM TMRM inmedium for 15min For calcein-AM staining the H9c2 cellswere loaded with 1120583M calcein-AM in the presences of 8mMcobalt chloride for 30min After the removal of excessive dyeby three washes with PBS the cell monolayer was examinedand imaged under a fluorescence microscope (Carl-ZeissJena Germany) through a 560 nm long-path filter
25 Immunoprecipitation Immunoprecipitation (IP) wasperformed as previously described [9] In brief 500 120583g oftotal cellular proteins from different treatment groups wasincubated with 1120583g primary antibodies against ANT andphospho-GSK-3120573 for 1 h at 4∘C The mixture was incubatedwith 20120583L of protein AG PLUS-agarose slurry (Santa CruzCA USA) at 4∘C overnight The samples were subsequentlycentrifuged at 2500 rpm for 5min at 4∘C The precipitationswere recovered and washed with PBS buffer for 3 timesFinally the pellets were dissolved in 60 120583L of 1x electrophore-sis sample buffer and boiled for 5min Thirty microliters ofeach sample was analyzed by Western blotting
26 Western Blot Analysis The cellular proteins were extractedwith ice-cold RIPA buffer according to the manufacturerrsquosinstructions (Sigma-Aldrich St Louis MO USA) In brief30 120583g of the cellular proteins was resolved in 10 SDS-PAGEand subsequently transferred onto polyvinylidene difluo-ride (PVDF) membrane (EMD Millipore Billerica MAUSA) Following the incubation in 5 BSA in TBST (50mMTris-Cl 150mM NaCl 01 Tween-20 pH 76) at roomtemperature for 2 h the membranes were probed with theprimary antibodies against the indicated proteins (1 1000dilution) at 4∘C overnight After three washes with TBSTbuffer the membranes were incubated with horseradishperoxidase- (HRP-) conjugated secondary antibody (1 1000dilution) at 4∘C for 3 h and visualized by enhanced chemi-luminescence (ECL) reaction reagents (GE Healthcare Upp-sala Sweden)
27 Cell Immunofluorescence Imaging H9c2 cells were seededon a glass slide at the density of 05 times 104 cellsslide After24 h incubation the cells were exposed to OGD conditionfor 8 h and treated with 30 120583M formononetin under reoxy-genation condition for another 20 minutes At the end oftreatment H9c2 cells were fixed with 4 paraformaldehydefor 30 minutes permeabilized with 05 Triton X-100 for20 minutes and blocked with 5 normal goat serum inPBS for 2 h at room temperature Phospho-GSK-3120573 (Ser9)and mitochondrial membrane protein Tom20 were probedwith specific primary antibodies overnight at 4∘C After 5washes with PBS the slides were incubated in the secondaryantibodies (ie Alexa Fluor 594-conjugated goat anti-rabbitIgG secondary antibody andAlexa Fluor 488-conjugated goatanti-mouse IgG secondary antibody) for 90min at roomtemperature The cell nuclei were stained with DAPI Afterthe removal of excessive fluorescence reagents the cells wereimaged on a Zeiss fluorescence microscopy (Carl-Zeiss JenaGermany)
28 Statistical Analysis Theresults were presented asmeansplusmnSD from three independent experiments The data werecompared by one-way analysis of variance (ANOVA) fol-lowed by least significant difference (LSD) post hoc testusing IBM software SPSS version 200 (Amonk NY USA)Differences with 119901 lt 005 were considered as statisticallysignificant
3 Results
31 Formononetin Enhanced the Survival of H9c2 Cells againstOGDReoxygenation Challenge The effect of formononetinon the viability of H9c2 cells was measured by standardcolorimetric MTT assay Under normal conditions for-mononetin at the concentration of 30 120583M did not affect cellviability over a period of 48 h As shown in Figure 1(b)however the cell viability was reduced to 667 plusmn 613after 8 h of OGD and 16 h of reoxygenation Interestinglyformononetin markedly increased the cell viability againstOGDreoxygenation-induced injury in a concentration-dependent manner
32 Inhibitory Effect of Formononetin on ROS and SuperoxideProduction The effect of formononetin on the intracellularredox status was determined by detecting the intracellu-lar H2O2and superoxide ion Upon the introduction into
the cells the nonpolar probe DCFH2-DA was convertedinto the polar intermediate DCFH by cellular esterasesand further oxidized to highly fluorescent product 2101584071015840-dichlorofluorescein (DCF) by the intracellular H
2O2and
other types of ROS On the other hand superoxide anionreacts with DHE generating a red fluorescent product 2-hydroxyethidium (EOH) As shown in Figures 2(a) and 2(b)OGDreoxygenation dramatically increased the generationof H2O2and superoxide ion in H9c2 cells Based on the
fluorescence intensity we estimated thatOGDreoxygenationincreased H
2O2level by 17-fold and also enhanced super-
oxide ion formation by 16-fold relative to the untreatedcontrols However formononetin attenuated the effects ofOGDreoxygenation on the generation ofH
2O2and superox-
ide ion in a concentration-dependentmanner Formononetinat 30 120583M reduced OGDreoxygenation-induced productionof H2O2and superoxide ion down to 13-fold and 12-fold
respectively
33 Effect of Formononetin on Mitochondrial MembraneIntegrity To determine the effect of formononetin on mito-chondrial membrane integrity a cell-permeant and cationicdye TMRM was used to localize active mitochondria Asshown in Figure 3(a) after exposure to OGD for 8 h andreoxygenation for 20min the capability of mitochondria tosequester TMRM was markedly diminished to 742 plusmn 22relative to untreated control cells Formononetin preservedthe sequestration of TMRM by active mitochondria in aconcentration-dependent manner After the treatment with30 120583M formononetin H9c2 cells showed the recovery ofTMRM fluorescence to 979 plusmn 50 relative to untreatedcontrol cells Calcein-AM staining showed similar results asshown in Figure 3(b)
4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
Relat
ive R
OS
leve
l fo
ld ch
ange
OGDR
0 5 10 30
0 0 10 30
OGDR
Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
lowast
mdash
(a)
0
05
1
15
2
25
Relat
ive s
uper
oxid
e lev
el fo
ld ch
ange
OGDR
10 30
OGDR
0 0
0 5 10 30Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
mdash
(b)
Figure 2 Effects of formononetin on ROS and superoxide production in cardiac H9c2 cells H9c2 cells were treated with variousconcentrations of formononetin under OGD and reoxygenation (OGDR) condition Accumulation of ROS and superoxide inH9c2 cells wasdetected by probes DCFH2-DA and DHE respectively (a) Representative images and quantification of relative ROS levels The intracellularROS level in H9c2 cells was calculated based on fluorescence intensity The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (b) Representative images and quantification ofrelative superoxide levels The intracellular superoxide level in H9c2 cells was calculated based on fluorescence intensity The fluorescenceimages were analyzed with NIH Image J software (httpimagejnihgovij) The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m
34 Effects of Formononetin on Akt and GSK-3120573 Phospho-rylation To discover the potential mechanisms we firstexamined the effects of formononetin on the phosphorylationof Akt and GSK-3120573 especially the phosphorylation of GSK-3120573 at Ser9 As shown in Figure 3(c) formononetin at theconcentrations (5ndash30120583M) induced the phosphorylation ofAkt and GSK-3120573 at Ser9 in a concentration-dependent man-ner We further verified the effects of formononetin on thephosphorylation of Akt and GSK-3120573 at Ser9 in the cells afterexposure to OGD and reoxygenation condition As shownin Figure 3(d) the levels of phospho-Akt and phospho-GSK-3120573 (Ser9) were suppressed under OGDreoxygenation condi-tion However formononetin increased Akt phosphorylationagainst OGDreoxygenation stimulation in a concentration-dependent manner Interestingly formononetin even at theconcentration of 5120583Meffectively reversed the loss of GSK-3120573phosphorylation against OGDreoxygenation challenge
35 Formononetin Activated Akt and PKC to PreserveMitochondrial Integrity To understand how formononetin
induces the phosphorylation of GSK-3120573 at Ser9 followingOGDreoxygenation challenge we treated the cells withformononetin in combination with PI3K inhibitor LY294002(LY) PKC inhibitor GF109203X (GF) and ERK 12 inhibitorPD98059 (PD) As shown in Figure 4(a) PI3K inhibitorLY and PKC inhibitor GF almost abolished the stimulatoryeffect of formononetin on the phosphorylation of GSK-3120573at Ser9 whereas ERK12 inhibitor did not show any activityagainst formononetin-induced formation of phospho-GSK-3120573 (Ser9) Supportively PI3K inhibitor LY and PKC inhibitorGF effectively antagonized the effects of formononetin onmitochondrial retention of TMRM and calcein-AM againstOGDreoxygenation damage (Figures 4(b) and 4(c))
36 Formononetin Stimulated the Binding of p-GSK-3120573 toANT in mPTP To verify the effect of formononetin on themPTP opening we examined the formation of proteincomplex between two main mPTP components ANT andCyp-D Following immunoprecipitation with anti-ANTantibody we analyzed the presence of ANT and Cyp-D by
Oxidative Medicine and Cellular Longevity 5
40 0 0 5 10 30
50
60
70
80
90
100
110
120
Relat
ive T
MRE
sequ
estr
atio
n
OGDR
Con
trol
OG
DR
mdashmdash
lowastlowast
lowast
FM (3
0120583
M) F
M (5
120583M
)
Formononetin (120583M)
(a)
160
140
120
100
80
60
40
20
0
Relat
ive c
alce
in fl
uore
scen
ce
OGDRmdash
lowastlowastlowast
lowastlowastlowast
Con
trol
OG
DR
mdashFM
(30120583
M)
FM (5
120583M
)
0 0 5 10 30Formononetin (120583M)
(b)
0 5 10 30
Fold
of i
nduc
tion
3
25
2
15
1
05
0
lowastlowast
lowastlowast lowastlowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
0
5120583M10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(c)
00 5 10 30
OGDR2
15
1
05
0
ControlOGDR
Treatment
Fold
of i
nduc
tion
mdash
lowastlowast
lowastlowast
lowastlowast
lowast lowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
5120583M
10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(d)
Figure 3 Effects of formononetin on mitochondrial membrane integrity and phosphorylation of Akt and GSK-3120573 (a) Fluorescence imagesand quantification of TMRM sequestration in mitochondria H9c2 cells were treated with formononetin (5 10 and 30 120583M) under normal orOGD and reoxygenation condition and stained with TMRM TMRMfluorescence intensity was determined by NIH image J software Resultsare presented as means plusmn SD (119899 = 3) 119901 lt 001 (OGDR versus normal) lowast119901 lt 005 lowastlowast119901 lt 001 (drug versus OGDR) Scale bar 10120583m(b) Fluorescence images and quantification of calcein sequestration in mitochondria H9c2 cells were treated similarly as described in (a)and stained with calcein-AM Calcein fluorescence intensity was determined by NIH image J software Results are presented as means plusmnSD (119899 = 3) 119901 lt 0001 (OGDR versus normal) lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (c) Western blot analysis andquantification of phosphorylation of GSK-3120573 and Akt in response to formononetin After the treatment with formononetin (5 10 and 30 120583M)under normal condition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blottingusing specific antibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt005 lowastlowast119901 lt 001 (drug versus control) (d) Western blot analysis and quantification of phosphorylation of GSK-3120573 and Akt in responseto formononetin under normal or OGDR condition After the treatment with formononetin (5 10 and 30 120583M) under normal or OGDRcondition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blotting using specificantibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt 005 lowastlowast119901 lt 001(drugs versus control and drugs versus OGDR)
6 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
GAPDH
Fold
of c
hang
e
30 303030Kinase inhibitor LY PDGFLYPDGF
TreatmentFormononetin (120583M)
lowast
lowastlowast
mdashmdashmdashmdash mdashmdash
30 303030Kinase inhibitor LY PDGFLYPDGF
Formononetin (120583M) mdashmdashmdashmdash mdashmdash
p-GSK-3120573
GSK-3120573
(a)
405060708090
100110120
OGDR
Mito
chon
dria
lTM
RE re
tent
ion
OGDR
lowast lowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdash
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash
mdash
mdash
30120583M 30120583M 30120583M
(b)
Relat
ive c
alce
in fl
uore
scen
ce
160140120100
80604020
0
OGDR
lowastlowastlowast lowastlowastlowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdashmdash
mdash
OGDR
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash30120583M 30120583M 30120583M
(c)
Figure 4 Roles of PI3K ERK12 and PKC in the regulation of GSK-3120573 phosphorylation and mitochondrial membrane integrity byformononetin (a) Western blot analysis and quantification of GSK-3120573 phosphorylation After the treatment with formononetin plusmn kinaseinhibitors H9c2 cells were lysed and analyzed for GSK-3120573 phosphorylation at Ser9 by Western blotting using specific antibodies Westernblots were determined by a densitometric method Results are presented as means plusmn SD (119899 = 3) 119901 lt 001 (drug versus OGDR)lowast119901 lt 005 lowastlowast119901 lt 001 (drug + inhibitor versus drug) (b and c) Fluorescence images and quantification of TMRMand calcein sequestration inmitochondria After the treatment with formononetin plusmn kinase inhibitors under OGD and reoxygenation condition H9c2 cells were stainedwith TMRM and calcein-AM and imaged under a fluorescence microscope (Carl-Zeiss Jena Germany) TMRM and calcein fluorescenceintensity were quantified by a densitometric method 119901 lt 0001 (drug versus OGDR) lowast119901 lt 005 (drug + inhibitor versus drug) LY PI3Kinhibitor LY294002 GF PKC inhibitor GF109203X PD ERK 12 inhibitor PD98059 Scale bar 10120583m
Western blotting with specific antibodies As shown in Fig-ure 5(a) OGDreoxygenation increased the formation ofANT-Cyp-D complex whereas formononetin diminishedOGDreoxygenation-induced formation of ANT-Cyp-D
complex On the other hand we performed immunopre-cipitation with anti-phospho-GSK-3120573 (Ser9) antibody andsubsequently analyzed the presence of ANT and phospho-GSK-3120573 (Ser9) by Western blotting with specific antibodies
Oxidative Medicine and Cellular Longevity 7
IB cyclophilin D
IB ANT
IP ANT OGDR
Formononetinmdashmdash
mdashmdashmdash
+ +
+
(a)
IB ANT
OGDRFormononetin
mdashmdash
mdashmdashmdash
+ +
+
IP p-GSK-3120573 (Ser9)
IB p-GSK-3120573 (Ser9)
(b)
Con
trol
OG
DR
PBS
OG
DR
form
onon
etin
p-GSK-3120573 (Ser9) Tom20 DAPI Merge
(c)
Figure 5 Effects of formononetin on the formation of ANT-Cyp-D complex and the binding of phospho-GSK-3120573 (Ser9) to ANT (a)Immunoprecipitation with antibody against ANT After the treatment with formononetin under OGD and reoxygenation condition H9c2cells were lysed for immunoprecipitation (IP) with antibody against ANT The pull-down materials were analyzed by immunoblotting (IB)with antibodies against ANT and Cyp-D (b) Immunoprecipitation with antibody against phospho-GSK-3120573 (Ser9) After the treatmentwith formononetin under OGD and reoxygenation condition H9c2 cells were lysed for immunoprecipitation (IP) with antibody againstphospho-GSK-3120573 (Ser9) The pull-down materials were analyzed by immunoblotting (IB) with antibodies against ANT and phospho-GSK-3120573 (Ser9) (c) Immunofluorescence imaging of the intracellular phospho-GSK-3120573 (Ser9) After the treatment with formononetin under OGDand reoxygenation condition H9c2 cells were probed with antibody against phospho-GSK-3120573 (Ser9) and mitochondrial membrane proteinTom20 and subsequently detected by fluorophore-labelled secondary antibodies phospho-GSK-3120573 (Ser9) (RED) and Tom20 (GREEN)respectively The cell nuclei were indicated by DAPI staining The images were captured under a fluorescence microscope (Carl-Zeiss JenaGermany) Scale bar 10 120583m
We found that formononetinmarkedly increased the bindingof phospho-GSK-3120573 (Ser9) to ANT (Figure 5(b))
The binding of phospho-GSK-3120573 (Ser9) tomitochondrialmembrane was further characterized by immunofluores-cence staining After 8 h ofOGDH9c2 cells were treatedwith30 120583M formononetin or equal volume of PBS under reoxy-genation condition for 20min The intracellular phospho-GSK-3120573 (Ser9) was sequentially stained by specific antibodyand Alexa Fluor 594-conjugated goat anti-rabbit IgG sec-ondary antibody Mitochondria were indicated by sequentialstaining of Tom20 with anti-Tom20 and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody DAPI
was used to visualize the cell nuclei As shown in Figure 5(c)formononetin dramatically increased the localization ofphospho-GSK-3120573 (Ser9) in mitochondria in close proximityto the cell nucleus
4 Discussion
Reperfusion of ischemic hearts is the main cause of theopening ofmPTP leading to the dysfunction ofmitochondria[22] The present study investigated that the effects of plantnatural product formononetin on the mPTP opening in anin vitro OGDreoxygenation-induced cell model of ischemia
8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
2 Oxidative Medicine and Cellular Longevity
HO O
OOCH3
(a)
0
20
40
60
80
100
120
140
Cel
l via
bilit
y (
)
0 5 10 30 Formononetin (120583M)
NormalOGDR
lowastlowast lowastlowast lowastlowast
(b)
Figure 1 Formononetin protected cardiomyocyte H9c2 cellsagainst OGDreoxygenation injury (a) Chemical structure of for-mononetin (b) Effect of formononetin on the cell viability of H9c2cells under normal or OGDreoxygenation condition H9c2 cellswere treated with formononetin at various concentrations eitherunder normal condition for 24 h (Normal) or subjected to OGD for8 h followed by reoxygenation for 16 h (OGDR) The cell viabilitywas determined by MTT assay The values represent mean plusmn SD(119899 = 5) 119901 lt 001 (OGDR versus Normal) lowastlowast119901 lt 001 (drugversus OGDR)
[7] Moreover phospho-GSK-3120573 (Ser9) suppresses pore for-mation via interacting with adenine nucleotide translocase(ANT) a major component of mPTP [9] These resultshighlight the importance of mPTP dynamics in myocardialischemia reperfusion injury
Medicinal plants such as Radix Astragali are widely usedto treat cardiovascular diseases in traditional Chinese medi-cine [10 11] As a major isoflavone compound from RadixAstragali formononetin bearing the structure shown inFigure 1(a) exhibits a wide range of pharmacological prop-erties such as anticancer [12 13] anti-inflammatory [14]antioxidant [15] antiviral [16] neuroprotective activities [1718] and wound healing [19] However little is known aboutthe molecular mechanisms underlying the cardioprotectivepotential of formononetin within the context of myocardialinfarction
In the present study we initially discovered that for-mononetin enhanced the survival of rat cardiomyocyte H9c2cells during oxygen glucose deprivation (OGD) and reoxy-genation We hypothesize that formononetin may protectcardiomyocytes against ischemia reperfusion injury by pre-venting mPTP openingWe focused on the role of GSK-3120573 inthe regulation of mPTP opening in H9c2 cells We furtherexamined the effects of formononetin on reactive oxygenspecies (ROS) PI3KAkt and PKC
2 Materials and Methods
21 Materials and Regents Formononetin was obtained fromYick-Vic Chemicals amp Pharmaceuticals Ltd (Hong Kong)Antibodies for GSK-3120573 and adenine nucleotide translo-case (ANT) were purchased from Santa Cruz Biotechnol-ogy (Santa Cruz CA USA) Antibodies against phospho-GSK-3120573 (Ser9) glyceraldehydes-3-phosphate dehydrogenase(GAPDH) Akt and phospho-Akt were obtained from CellSignaling Technology (BostonMAUSA) AnticyclophilinD(anti-Cyp-D) was purchased from Thermo Fisher ScientificInc (Waltham MA USA) Alexa Fluor 594-conjugated goatanti-rabbit IgG secondary antibody and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody wereobtained from Invitrogen (Carlsbad CA USA)
22 Cell Culture Rat cardiomyocyte H9c2 cell line wasobtained from Shanghai Institute for Biological SciencesChinese Academy of Science (Shanghai China) and cul-tured in Dulbeccorsquos modified Eaglersquos medium (DMEM)supplemented with 10 heat-inactivated fetal bovine serum(GibcoBRL Gaithersburg MD USA) 2mM glutamine and100UmL penicillin Astreptomycin at 37∘C in a humidifiedincubator containing 5 CO
2
23 Measurement of ROS and Superoxide Production H9c2cells were seeded in 6-well plate at the density of 03 times105 cellsmL overnight After 24 h incubation the cells wereexposed to OGD condition for 8 h and subsequently treatedwith formononetin at different concentrations under reoxy-genation condition for 30min For the detection of intra-cellular ROS the cells were stained with 5120583M of 2101584071015840-dichlorofluorescein-diacetate (DCFH2-DA) from Life Tech-nologies (Grand Island NY USA) at 37∘C for 30min Onthe other hand the intracellular superoxide ion was detectedwith dihydroergotamine (DHE) from Invitrogen MolecularProbes (Eugene OR USA) at 37∘C for 30min After theremoval of excessive probes the cells were imaged on a laserscanning fluorescence microscope (Carl-Zeiss Jena Ger-many) The results were expressed as a percentage of theintracellular ROS or superoxide production in OGDreoxy-genation-treated cells based on the analysis using NIH ImageJ software (httpimagejnihgovij)
24 Imaging of Mitochondrial Membrane Integrity Mito-chondrial membrane integrity was detected by staining withtetramethylrhodamine methyl ester (TMRM) and calcein-AM as reported [20 21] Briefly H9c2 cells were exposedto OGD condition for 8 h and treated with formononetin
Oxidative Medicine and Cellular Longevity 3
(5 to 30 120583M) under reoxygenation condition for another20min H9c2 cells were incubated with 250 nM TMRM inmedium for 15min For calcein-AM staining the H9c2 cellswere loaded with 1120583M calcein-AM in the presences of 8mMcobalt chloride for 30min After the removal of excessive dyeby three washes with PBS the cell monolayer was examinedand imaged under a fluorescence microscope (Carl-ZeissJena Germany) through a 560 nm long-path filter
25 Immunoprecipitation Immunoprecipitation (IP) wasperformed as previously described [9] In brief 500 120583g oftotal cellular proteins from different treatment groups wasincubated with 1120583g primary antibodies against ANT andphospho-GSK-3120573 for 1 h at 4∘C The mixture was incubatedwith 20120583L of protein AG PLUS-agarose slurry (Santa CruzCA USA) at 4∘C overnight The samples were subsequentlycentrifuged at 2500 rpm for 5min at 4∘C The precipitationswere recovered and washed with PBS buffer for 3 timesFinally the pellets were dissolved in 60 120583L of 1x electrophore-sis sample buffer and boiled for 5min Thirty microliters ofeach sample was analyzed by Western blotting
26 Western Blot Analysis The cellular proteins were extractedwith ice-cold RIPA buffer according to the manufacturerrsquosinstructions (Sigma-Aldrich St Louis MO USA) In brief30 120583g of the cellular proteins was resolved in 10 SDS-PAGEand subsequently transferred onto polyvinylidene difluo-ride (PVDF) membrane (EMD Millipore Billerica MAUSA) Following the incubation in 5 BSA in TBST (50mMTris-Cl 150mM NaCl 01 Tween-20 pH 76) at roomtemperature for 2 h the membranes were probed with theprimary antibodies against the indicated proteins (1 1000dilution) at 4∘C overnight After three washes with TBSTbuffer the membranes were incubated with horseradishperoxidase- (HRP-) conjugated secondary antibody (1 1000dilution) at 4∘C for 3 h and visualized by enhanced chemi-luminescence (ECL) reaction reagents (GE Healthcare Upp-sala Sweden)
27 Cell Immunofluorescence Imaging H9c2 cells were seededon a glass slide at the density of 05 times 104 cellsslide After24 h incubation the cells were exposed to OGD conditionfor 8 h and treated with 30 120583M formononetin under reoxy-genation condition for another 20 minutes At the end oftreatment H9c2 cells were fixed with 4 paraformaldehydefor 30 minutes permeabilized with 05 Triton X-100 for20 minutes and blocked with 5 normal goat serum inPBS for 2 h at room temperature Phospho-GSK-3120573 (Ser9)and mitochondrial membrane protein Tom20 were probedwith specific primary antibodies overnight at 4∘C After 5washes with PBS the slides were incubated in the secondaryantibodies (ie Alexa Fluor 594-conjugated goat anti-rabbitIgG secondary antibody andAlexa Fluor 488-conjugated goatanti-mouse IgG secondary antibody) for 90min at roomtemperature The cell nuclei were stained with DAPI Afterthe removal of excessive fluorescence reagents the cells wereimaged on a Zeiss fluorescence microscopy (Carl-Zeiss JenaGermany)
28 Statistical Analysis Theresults were presented asmeansplusmnSD from three independent experiments The data werecompared by one-way analysis of variance (ANOVA) fol-lowed by least significant difference (LSD) post hoc testusing IBM software SPSS version 200 (Amonk NY USA)Differences with 119901 lt 005 were considered as statisticallysignificant
3 Results
31 Formononetin Enhanced the Survival of H9c2 Cells againstOGDReoxygenation Challenge The effect of formononetinon the viability of H9c2 cells was measured by standardcolorimetric MTT assay Under normal conditions for-mononetin at the concentration of 30 120583M did not affect cellviability over a period of 48 h As shown in Figure 1(b)however the cell viability was reduced to 667 plusmn 613after 8 h of OGD and 16 h of reoxygenation Interestinglyformononetin markedly increased the cell viability againstOGDreoxygenation-induced injury in a concentration-dependent manner
32 Inhibitory Effect of Formononetin on ROS and SuperoxideProduction The effect of formononetin on the intracellularredox status was determined by detecting the intracellu-lar H2O2and superoxide ion Upon the introduction into
the cells the nonpolar probe DCFH2-DA was convertedinto the polar intermediate DCFH by cellular esterasesand further oxidized to highly fluorescent product 2101584071015840-dichlorofluorescein (DCF) by the intracellular H
2O2and
other types of ROS On the other hand superoxide anionreacts with DHE generating a red fluorescent product 2-hydroxyethidium (EOH) As shown in Figures 2(a) and 2(b)OGDreoxygenation dramatically increased the generationof H2O2and superoxide ion in H9c2 cells Based on the
fluorescence intensity we estimated thatOGDreoxygenationincreased H
2O2level by 17-fold and also enhanced super-
oxide ion formation by 16-fold relative to the untreatedcontrols However formononetin attenuated the effects ofOGDreoxygenation on the generation ofH
2O2and superox-
ide ion in a concentration-dependentmanner Formononetinat 30 120583M reduced OGDreoxygenation-induced productionof H2O2and superoxide ion down to 13-fold and 12-fold
respectively
33 Effect of Formononetin on Mitochondrial MembraneIntegrity To determine the effect of formononetin on mito-chondrial membrane integrity a cell-permeant and cationicdye TMRM was used to localize active mitochondria Asshown in Figure 3(a) after exposure to OGD for 8 h andreoxygenation for 20min the capability of mitochondria tosequester TMRM was markedly diminished to 742 plusmn 22relative to untreated control cells Formononetin preservedthe sequestration of TMRM by active mitochondria in aconcentration-dependent manner After the treatment with30 120583M formononetin H9c2 cells showed the recovery ofTMRM fluorescence to 979 plusmn 50 relative to untreatedcontrol cells Calcein-AM staining showed similar results asshown in Figure 3(b)
4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
Relat
ive R
OS
leve
l fo
ld ch
ange
OGDR
0 5 10 30
0 0 10 30
OGDR
Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
lowast
mdash
(a)
0
05
1
15
2
25
Relat
ive s
uper
oxid
e lev
el fo
ld ch
ange
OGDR
10 30
OGDR
0 0
0 5 10 30Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
mdash
(b)
Figure 2 Effects of formononetin on ROS and superoxide production in cardiac H9c2 cells H9c2 cells were treated with variousconcentrations of formononetin under OGD and reoxygenation (OGDR) condition Accumulation of ROS and superoxide inH9c2 cells wasdetected by probes DCFH2-DA and DHE respectively (a) Representative images and quantification of relative ROS levels The intracellularROS level in H9c2 cells was calculated based on fluorescence intensity The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (b) Representative images and quantification ofrelative superoxide levels The intracellular superoxide level in H9c2 cells was calculated based on fluorescence intensity The fluorescenceimages were analyzed with NIH Image J software (httpimagejnihgovij) The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m
34 Effects of Formononetin on Akt and GSK-3120573 Phospho-rylation To discover the potential mechanisms we firstexamined the effects of formononetin on the phosphorylationof Akt and GSK-3120573 especially the phosphorylation of GSK-3120573 at Ser9 As shown in Figure 3(c) formononetin at theconcentrations (5ndash30120583M) induced the phosphorylation ofAkt and GSK-3120573 at Ser9 in a concentration-dependent man-ner We further verified the effects of formononetin on thephosphorylation of Akt and GSK-3120573 at Ser9 in the cells afterexposure to OGD and reoxygenation condition As shownin Figure 3(d) the levels of phospho-Akt and phospho-GSK-3120573 (Ser9) were suppressed under OGDreoxygenation condi-tion However formononetin increased Akt phosphorylationagainst OGDreoxygenation stimulation in a concentration-dependent manner Interestingly formononetin even at theconcentration of 5120583Meffectively reversed the loss of GSK-3120573phosphorylation against OGDreoxygenation challenge
35 Formononetin Activated Akt and PKC to PreserveMitochondrial Integrity To understand how formononetin
induces the phosphorylation of GSK-3120573 at Ser9 followingOGDreoxygenation challenge we treated the cells withformononetin in combination with PI3K inhibitor LY294002(LY) PKC inhibitor GF109203X (GF) and ERK 12 inhibitorPD98059 (PD) As shown in Figure 4(a) PI3K inhibitorLY and PKC inhibitor GF almost abolished the stimulatoryeffect of formononetin on the phosphorylation of GSK-3120573at Ser9 whereas ERK12 inhibitor did not show any activityagainst formononetin-induced formation of phospho-GSK-3120573 (Ser9) Supportively PI3K inhibitor LY and PKC inhibitorGF effectively antagonized the effects of formononetin onmitochondrial retention of TMRM and calcein-AM againstOGDreoxygenation damage (Figures 4(b) and 4(c))
36 Formononetin Stimulated the Binding of p-GSK-3120573 toANT in mPTP To verify the effect of formononetin on themPTP opening we examined the formation of proteincomplex between two main mPTP components ANT andCyp-D Following immunoprecipitation with anti-ANTantibody we analyzed the presence of ANT and Cyp-D by
Oxidative Medicine and Cellular Longevity 5
40 0 0 5 10 30
50
60
70
80
90
100
110
120
Relat
ive T
MRE
sequ
estr
atio
n
OGDR
Con
trol
OG
DR
mdashmdash
lowastlowast
lowast
FM (3
0120583
M) F
M (5
120583M
)
Formononetin (120583M)
(a)
160
140
120
100
80
60
40
20
0
Relat
ive c
alce
in fl
uore
scen
ce
OGDRmdash
lowastlowastlowast
lowastlowastlowast
Con
trol
OG
DR
mdashFM
(30120583
M)
FM (5
120583M
)
0 0 5 10 30Formononetin (120583M)
(b)
0 5 10 30
Fold
of i
nduc
tion
3
25
2
15
1
05
0
lowastlowast
lowastlowast lowastlowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
0
5120583M10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(c)
00 5 10 30
OGDR2
15
1
05
0
ControlOGDR
Treatment
Fold
of i
nduc
tion
mdash
lowastlowast
lowastlowast
lowastlowast
lowast lowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
5120583M
10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(d)
Figure 3 Effects of formononetin on mitochondrial membrane integrity and phosphorylation of Akt and GSK-3120573 (a) Fluorescence imagesand quantification of TMRM sequestration in mitochondria H9c2 cells were treated with formononetin (5 10 and 30 120583M) under normal orOGD and reoxygenation condition and stained with TMRM TMRMfluorescence intensity was determined by NIH image J software Resultsare presented as means plusmn SD (119899 = 3) 119901 lt 001 (OGDR versus normal) lowast119901 lt 005 lowastlowast119901 lt 001 (drug versus OGDR) Scale bar 10120583m(b) Fluorescence images and quantification of calcein sequestration in mitochondria H9c2 cells were treated similarly as described in (a)and stained with calcein-AM Calcein fluorescence intensity was determined by NIH image J software Results are presented as means plusmnSD (119899 = 3) 119901 lt 0001 (OGDR versus normal) lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (c) Western blot analysis andquantification of phosphorylation of GSK-3120573 and Akt in response to formononetin After the treatment with formononetin (5 10 and 30 120583M)under normal condition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blottingusing specific antibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt005 lowastlowast119901 lt 001 (drug versus control) (d) Western blot analysis and quantification of phosphorylation of GSK-3120573 and Akt in responseto formononetin under normal or OGDR condition After the treatment with formononetin (5 10 and 30 120583M) under normal or OGDRcondition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blotting using specificantibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt 005 lowastlowast119901 lt 001(drugs versus control and drugs versus OGDR)
6 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
GAPDH
Fold
of c
hang
e
30 303030Kinase inhibitor LY PDGFLYPDGF
TreatmentFormononetin (120583M)
lowast
lowastlowast
mdashmdashmdashmdash mdashmdash
30 303030Kinase inhibitor LY PDGFLYPDGF
Formononetin (120583M) mdashmdashmdashmdash mdashmdash
p-GSK-3120573
GSK-3120573
(a)
405060708090
100110120
OGDR
Mito
chon
dria
lTM
RE re
tent
ion
OGDR
lowast lowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdash
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash
mdash
mdash
30120583M 30120583M 30120583M
(b)
Relat
ive c
alce
in fl
uore
scen
ce
160140120100
80604020
0
OGDR
lowastlowastlowast lowastlowastlowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdashmdash
mdash
OGDR
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash30120583M 30120583M 30120583M
(c)
Figure 4 Roles of PI3K ERK12 and PKC in the regulation of GSK-3120573 phosphorylation and mitochondrial membrane integrity byformononetin (a) Western blot analysis and quantification of GSK-3120573 phosphorylation After the treatment with formononetin plusmn kinaseinhibitors H9c2 cells were lysed and analyzed for GSK-3120573 phosphorylation at Ser9 by Western blotting using specific antibodies Westernblots were determined by a densitometric method Results are presented as means plusmn SD (119899 = 3) 119901 lt 001 (drug versus OGDR)lowast119901 lt 005 lowastlowast119901 lt 001 (drug + inhibitor versus drug) (b and c) Fluorescence images and quantification of TMRMand calcein sequestration inmitochondria After the treatment with formononetin plusmn kinase inhibitors under OGD and reoxygenation condition H9c2 cells were stainedwith TMRM and calcein-AM and imaged under a fluorescence microscope (Carl-Zeiss Jena Germany) TMRM and calcein fluorescenceintensity were quantified by a densitometric method 119901 lt 0001 (drug versus OGDR) lowast119901 lt 005 (drug + inhibitor versus drug) LY PI3Kinhibitor LY294002 GF PKC inhibitor GF109203X PD ERK 12 inhibitor PD98059 Scale bar 10120583m
Western blotting with specific antibodies As shown in Fig-ure 5(a) OGDreoxygenation increased the formation ofANT-Cyp-D complex whereas formononetin diminishedOGDreoxygenation-induced formation of ANT-Cyp-D
complex On the other hand we performed immunopre-cipitation with anti-phospho-GSK-3120573 (Ser9) antibody andsubsequently analyzed the presence of ANT and phospho-GSK-3120573 (Ser9) by Western blotting with specific antibodies
Oxidative Medicine and Cellular Longevity 7
IB cyclophilin D
IB ANT
IP ANT OGDR
Formononetinmdashmdash
mdashmdashmdash
+ +
+
(a)
IB ANT
OGDRFormononetin
mdashmdash
mdashmdashmdash
+ +
+
IP p-GSK-3120573 (Ser9)
IB p-GSK-3120573 (Ser9)
(b)
Con
trol
OG
DR
PBS
OG
DR
form
onon
etin
p-GSK-3120573 (Ser9) Tom20 DAPI Merge
(c)
Figure 5 Effects of formononetin on the formation of ANT-Cyp-D complex and the binding of phospho-GSK-3120573 (Ser9) to ANT (a)Immunoprecipitation with antibody against ANT After the treatment with formononetin under OGD and reoxygenation condition H9c2cells were lysed for immunoprecipitation (IP) with antibody against ANT The pull-down materials were analyzed by immunoblotting (IB)with antibodies against ANT and Cyp-D (b) Immunoprecipitation with antibody against phospho-GSK-3120573 (Ser9) After the treatmentwith formononetin under OGD and reoxygenation condition H9c2 cells were lysed for immunoprecipitation (IP) with antibody againstphospho-GSK-3120573 (Ser9) The pull-down materials were analyzed by immunoblotting (IB) with antibodies against ANT and phospho-GSK-3120573 (Ser9) (c) Immunofluorescence imaging of the intracellular phospho-GSK-3120573 (Ser9) After the treatment with formononetin under OGDand reoxygenation condition H9c2 cells were probed with antibody against phospho-GSK-3120573 (Ser9) and mitochondrial membrane proteinTom20 and subsequently detected by fluorophore-labelled secondary antibodies phospho-GSK-3120573 (Ser9) (RED) and Tom20 (GREEN)respectively The cell nuclei were indicated by DAPI staining The images were captured under a fluorescence microscope (Carl-Zeiss JenaGermany) Scale bar 10 120583m
We found that formononetinmarkedly increased the bindingof phospho-GSK-3120573 (Ser9) to ANT (Figure 5(b))
The binding of phospho-GSK-3120573 (Ser9) tomitochondrialmembrane was further characterized by immunofluores-cence staining After 8 h ofOGDH9c2 cells were treatedwith30 120583M formononetin or equal volume of PBS under reoxy-genation condition for 20min The intracellular phospho-GSK-3120573 (Ser9) was sequentially stained by specific antibodyand Alexa Fluor 594-conjugated goat anti-rabbit IgG sec-ondary antibody Mitochondria were indicated by sequentialstaining of Tom20 with anti-Tom20 and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody DAPI
was used to visualize the cell nuclei As shown in Figure 5(c)formononetin dramatically increased the localization ofphospho-GSK-3120573 (Ser9) in mitochondria in close proximityto the cell nucleus
4 Discussion
Reperfusion of ischemic hearts is the main cause of theopening ofmPTP leading to the dysfunction ofmitochondria[22] The present study investigated that the effects of plantnatural product formononetin on the mPTP opening in anin vitro OGDreoxygenation-induced cell model of ischemia
8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 3
(5 to 30 120583M) under reoxygenation condition for another20min H9c2 cells were incubated with 250 nM TMRM inmedium for 15min For calcein-AM staining the H9c2 cellswere loaded with 1120583M calcein-AM in the presences of 8mMcobalt chloride for 30min After the removal of excessive dyeby three washes with PBS the cell monolayer was examinedand imaged under a fluorescence microscope (Carl-ZeissJena Germany) through a 560 nm long-path filter
25 Immunoprecipitation Immunoprecipitation (IP) wasperformed as previously described [9] In brief 500 120583g oftotal cellular proteins from different treatment groups wasincubated with 1120583g primary antibodies against ANT andphospho-GSK-3120573 for 1 h at 4∘C The mixture was incubatedwith 20120583L of protein AG PLUS-agarose slurry (Santa CruzCA USA) at 4∘C overnight The samples were subsequentlycentrifuged at 2500 rpm for 5min at 4∘C The precipitationswere recovered and washed with PBS buffer for 3 timesFinally the pellets were dissolved in 60 120583L of 1x electrophore-sis sample buffer and boiled for 5min Thirty microliters ofeach sample was analyzed by Western blotting
26 Western Blot Analysis The cellular proteins were extractedwith ice-cold RIPA buffer according to the manufacturerrsquosinstructions (Sigma-Aldrich St Louis MO USA) In brief30 120583g of the cellular proteins was resolved in 10 SDS-PAGEand subsequently transferred onto polyvinylidene difluo-ride (PVDF) membrane (EMD Millipore Billerica MAUSA) Following the incubation in 5 BSA in TBST (50mMTris-Cl 150mM NaCl 01 Tween-20 pH 76) at roomtemperature for 2 h the membranes were probed with theprimary antibodies against the indicated proteins (1 1000dilution) at 4∘C overnight After three washes with TBSTbuffer the membranes were incubated with horseradishperoxidase- (HRP-) conjugated secondary antibody (1 1000dilution) at 4∘C for 3 h and visualized by enhanced chemi-luminescence (ECL) reaction reagents (GE Healthcare Upp-sala Sweden)
27 Cell Immunofluorescence Imaging H9c2 cells were seededon a glass slide at the density of 05 times 104 cellsslide After24 h incubation the cells were exposed to OGD conditionfor 8 h and treated with 30 120583M formononetin under reoxy-genation condition for another 20 minutes At the end oftreatment H9c2 cells were fixed with 4 paraformaldehydefor 30 minutes permeabilized with 05 Triton X-100 for20 minutes and blocked with 5 normal goat serum inPBS for 2 h at room temperature Phospho-GSK-3120573 (Ser9)and mitochondrial membrane protein Tom20 were probedwith specific primary antibodies overnight at 4∘C After 5washes with PBS the slides were incubated in the secondaryantibodies (ie Alexa Fluor 594-conjugated goat anti-rabbitIgG secondary antibody andAlexa Fluor 488-conjugated goatanti-mouse IgG secondary antibody) for 90min at roomtemperature The cell nuclei were stained with DAPI Afterthe removal of excessive fluorescence reagents the cells wereimaged on a Zeiss fluorescence microscopy (Carl-Zeiss JenaGermany)
28 Statistical Analysis Theresults were presented asmeansplusmnSD from three independent experiments The data werecompared by one-way analysis of variance (ANOVA) fol-lowed by least significant difference (LSD) post hoc testusing IBM software SPSS version 200 (Amonk NY USA)Differences with 119901 lt 005 were considered as statisticallysignificant
3 Results
31 Formononetin Enhanced the Survival of H9c2 Cells againstOGDReoxygenation Challenge The effect of formononetinon the viability of H9c2 cells was measured by standardcolorimetric MTT assay Under normal conditions for-mononetin at the concentration of 30 120583M did not affect cellviability over a period of 48 h As shown in Figure 1(b)however the cell viability was reduced to 667 plusmn 613after 8 h of OGD and 16 h of reoxygenation Interestinglyformononetin markedly increased the cell viability againstOGDreoxygenation-induced injury in a concentration-dependent manner
32 Inhibitory Effect of Formononetin on ROS and SuperoxideProduction The effect of formononetin on the intracellularredox status was determined by detecting the intracellu-lar H2O2and superoxide ion Upon the introduction into
the cells the nonpolar probe DCFH2-DA was convertedinto the polar intermediate DCFH by cellular esterasesand further oxidized to highly fluorescent product 2101584071015840-dichlorofluorescein (DCF) by the intracellular H
2O2and
other types of ROS On the other hand superoxide anionreacts with DHE generating a red fluorescent product 2-hydroxyethidium (EOH) As shown in Figures 2(a) and 2(b)OGDreoxygenation dramatically increased the generationof H2O2and superoxide ion in H9c2 cells Based on the
fluorescence intensity we estimated thatOGDreoxygenationincreased H
2O2level by 17-fold and also enhanced super-
oxide ion formation by 16-fold relative to the untreatedcontrols However formononetin attenuated the effects ofOGDreoxygenation on the generation ofH
2O2and superox-
ide ion in a concentration-dependentmanner Formononetinat 30 120583M reduced OGDreoxygenation-induced productionof H2O2and superoxide ion down to 13-fold and 12-fold
respectively
33 Effect of Formononetin on Mitochondrial MembraneIntegrity To determine the effect of formononetin on mito-chondrial membrane integrity a cell-permeant and cationicdye TMRM was used to localize active mitochondria Asshown in Figure 3(a) after exposure to OGD for 8 h andreoxygenation for 20min the capability of mitochondria tosequester TMRM was markedly diminished to 742 plusmn 22relative to untreated control cells Formononetin preservedthe sequestration of TMRM by active mitochondria in aconcentration-dependent manner After the treatment with30 120583M formononetin H9c2 cells showed the recovery ofTMRM fluorescence to 979 plusmn 50 relative to untreatedcontrol cells Calcein-AM staining showed similar results asshown in Figure 3(b)
4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
Relat
ive R
OS
leve
l fo
ld ch
ange
OGDR
0 5 10 30
0 0 10 30
OGDR
Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
lowast
mdash
(a)
0
05
1
15
2
25
Relat
ive s
uper
oxid
e lev
el fo
ld ch
ange
OGDR
10 30
OGDR
0 0
0 5 10 30Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
mdash
(b)
Figure 2 Effects of formononetin on ROS and superoxide production in cardiac H9c2 cells H9c2 cells were treated with variousconcentrations of formononetin under OGD and reoxygenation (OGDR) condition Accumulation of ROS and superoxide inH9c2 cells wasdetected by probes DCFH2-DA and DHE respectively (a) Representative images and quantification of relative ROS levels The intracellularROS level in H9c2 cells was calculated based on fluorescence intensity The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (b) Representative images and quantification ofrelative superoxide levels The intracellular superoxide level in H9c2 cells was calculated based on fluorescence intensity The fluorescenceimages were analyzed with NIH Image J software (httpimagejnihgovij) The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m
34 Effects of Formononetin on Akt and GSK-3120573 Phospho-rylation To discover the potential mechanisms we firstexamined the effects of formononetin on the phosphorylationof Akt and GSK-3120573 especially the phosphorylation of GSK-3120573 at Ser9 As shown in Figure 3(c) formononetin at theconcentrations (5ndash30120583M) induced the phosphorylation ofAkt and GSK-3120573 at Ser9 in a concentration-dependent man-ner We further verified the effects of formononetin on thephosphorylation of Akt and GSK-3120573 at Ser9 in the cells afterexposure to OGD and reoxygenation condition As shownin Figure 3(d) the levels of phospho-Akt and phospho-GSK-3120573 (Ser9) were suppressed under OGDreoxygenation condi-tion However formononetin increased Akt phosphorylationagainst OGDreoxygenation stimulation in a concentration-dependent manner Interestingly formononetin even at theconcentration of 5120583Meffectively reversed the loss of GSK-3120573phosphorylation against OGDreoxygenation challenge
35 Formononetin Activated Akt and PKC to PreserveMitochondrial Integrity To understand how formononetin
induces the phosphorylation of GSK-3120573 at Ser9 followingOGDreoxygenation challenge we treated the cells withformononetin in combination with PI3K inhibitor LY294002(LY) PKC inhibitor GF109203X (GF) and ERK 12 inhibitorPD98059 (PD) As shown in Figure 4(a) PI3K inhibitorLY and PKC inhibitor GF almost abolished the stimulatoryeffect of formononetin on the phosphorylation of GSK-3120573at Ser9 whereas ERK12 inhibitor did not show any activityagainst formononetin-induced formation of phospho-GSK-3120573 (Ser9) Supportively PI3K inhibitor LY and PKC inhibitorGF effectively antagonized the effects of formononetin onmitochondrial retention of TMRM and calcein-AM againstOGDreoxygenation damage (Figures 4(b) and 4(c))
36 Formononetin Stimulated the Binding of p-GSK-3120573 toANT in mPTP To verify the effect of formononetin on themPTP opening we examined the formation of proteincomplex between two main mPTP components ANT andCyp-D Following immunoprecipitation with anti-ANTantibody we analyzed the presence of ANT and Cyp-D by
Oxidative Medicine and Cellular Longevity 5
40 0 0 5 10 30
50
60
70
80
90
100
110
120
Relat
ive T
MRE
sequ
estr
atio
n
OGDR
Con
trol
OG
DR
mdashmdash
lowastlowast
lowast
FM (3
0120583
M) F
M (5
120583M
)
Formononetin (120583M)
(a)
160
140
120
100
80
60
40
20
0
Relat
ive c
alce
in fl
uore
scen
ce
OGDRmdash
lowastlowastlowast
lowastlowastlowast
Con
trol
OG
DR
mdashFM
(30120583
M)
FM (5
120583M
)
0 0 5 10 30Formononetin (120583M)
(b)
0 5 10 30
Fold
of i
nduc
tion
3
25
2
15
1
05
0
lowastlowast
lowastlowast lowastlowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
0
5120583M10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(c)
00 5 10 30
OGDR2
15
1
05
0
ControlOGDR
Treatment
Fold
of i
nduc
tion
mdash
lowastlowast
lowastlowast
lowastlowast
lowast lowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
5120583M
10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(d)
Figure 3 Effects of formononetin on mitochondrial membrane integrity and phosphorylation of Akt and GSK-3120573 (a) Fluorescence imagesand quantification of TMRM sequestration in mitochondria H9c2 cells were treated with formononetin (5 10 and 30 120583M) under normal orOGD and reoxygenation condition and stained with TMRM TMRMfluorescence intensity was determined by NIH image J software Resultsare presented as means plusmn SD (119899 = 3) 119901 lt 001 (OGDR versus normal) lowast119901 lt 005 lowastlowast119901 lt 001 (drug versus OGDR) Scale bar 10120583m(b) Fluorescence images and quantification of calcein sequestration in mitochondria H9c2 cells were treated similarly as described in (a)and stained with calcein-AM Calcein fluorescence intensity was determined by NIH image J software Results are presented as means plusmnSD (119899 = 3) 119901 lt 0001 (OGDR versus normal) lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (c) Western blot analysis andquantification of phosphorylation of GSK-3120573 and Akt in response to formononetin After the treatment with formononetin (5 10 and 30 120583M)under normal condition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blottingusing specific antibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt005 lowastlowast119901 lt 001 (drug versus control) (d) Western blot analysis and quantification of phosphorylation of GSK-3120573 and Akt in responseto formononetin under normal or OGDR condition After the treatment with formononetin (5 10 and 30 120583M) under normal or OGDRcondition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blotting using specificantibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt 005 lowastlowast119901 lt 001(drugs versus control and drugs versus OGDR)
6 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
GAPDH
Fold
of c
hang
e
30 303030Kinase inhibitor LY PDGFLYPDGF
TreatmentFormononetin (120583M)
lowast
lowastlowast
mdashmdashmdashmdash mdashmdash
30 303030Kinase inhibitor LY PDGFLYPDGF
Formononetin (120583M) mdashmdashmdashmdash mdashmdash
p-GSK-3120573
GSK-3120573
(a)
405060708090
100110120
OGDR
Mito
chon
dria
lTM
RE re
tent
ion
OGDR
lowast lowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdash
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash
mdash
mdash
30120583M 30120583M 30120583M
(b)
Relat
ive c
alce
in fl
uore
scen
ce
160140120100
80604020
0
OGDR
lowastlowastlowast lowastlowastlowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdashmdash
mdash
OGDR
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash30120583M 30120583M 30120583M
(c)
Figure 4 Roles of PI3K ERK12 and PKC in the regulation of GSK-3120573 phosphorylation and mitochondrial membrane integrity byformononetin (a) Western blot analysis and quantification of GSK-3120573 phosphorylation After the treatment with formononetin plusmn kinaseinhibitors H9c2 cells were lysed and analyzed for GSK-3120573 phosphorylation at Ser9 by Western blotting using specific antibodies Westernblots were determined by a densitometric method Results are presented as means plusmn SD (119899 = 3) 119901 lt 001 (drug versus OGDR)lowast119901 lt 005 lowastlowast119901 lt 001 (drug + inhibitor versus drug) (b and c) Fluorescence images and quantification of TMRMand calcein sequestration inmitochondria After the treatment with formononetin plusmn kinase inhibitors under OGD and reoxygenation condition H9c2 cells were stainedwith TMRM and calcein-AM and imaged under a fluorescence microscope (Carl-Zeiss Jena Germany) TMRM and calcein fluorescenceintensity were quantified by a densitometric method 119901 lt 0001 (drug versus OGDR) lowast119901 lt 005 (drug + inhibitor versus drug) LY PI3Kinhibitor LY294002 GF PKC inhibitor GF109203X PD ERK 12 inhibitor PD98059 Scale bar 10120583m
Western blotting with specific antibodies As shown in Fig-ure 5(a) OGDreoxygenation increased the formation ofANT-Cyp-D complex whereas formononetin diminishedOGDreoxygenation-induced formation of ANT-Cyp-D
complex On the other hand we performed immunopre-cipitation with anti-phospho-GSK-3120573 (Ser9) antibody andsubsequently analyzed the presence of ANT and phospho-GSK-3120573 (Ser9) by Western blotting with specific antibodies
Oxidative Medicine and Cellular Longevity 7
IB cyclophilin D
IB ANT
IP ANT OGDR
Formononetinmdashmdash
mdashmdashmdash
+ +
+
(a)
IB ANT
OGDRFormononetin
mdashmdash
mdashmdashmdash
+ +
+
IP p-GSK-3120573 (Ser9)
IB p-GSK-3120573 (Ser9)
(b)
Con
trol
OG
DR
PBS
OG
DR
form
onon
etin
p-GSK-3120573 (Ser9) Tom20 DAPI Merge
(c)
Figure 5 Effects of formononetin on the formation of ANT-Cyp-D complex and the binding of phospho-GSK-3120573 (Ser9) to ANT (a)Immunoprecipitation with antibody against ANT After the treatment with formononetin under OGD and reoxygenation condition H9c2cells were lysed for immunoprecipitation (IP) with antibody against ANT The pull-down materials were analyzed by immunoblotting (IB)with antibodies against ANT and Cyp-D (b) Immunoprecipitation with antibody against phospho-GSK-3120573 (Ser9) After the treatmentwith formononetin under OGD and reoxygenation condition H9c2 cells were lysed for immunoprecipitation (IP) with antibody againstphospho-GSK-3120573 (Ser9) The pull-down materials were analyzed by immunoblotting (IB) with antibodies against ANT and phospho-GSK-3120573 (Ser9) (c) Immunofluorescence imaging of the intracellular phospho-GSK-3120573 (Ser9) After the treatment with formononetin under OGDand reoxygenation condition H9c2 cells were probed with antibody against phospho-GSK-3120573 (Ser9) and mitochondrial membrane proteinTom20 and subsequently detected by fluorophore-labelled secondary antibodies phospho-GSK-3120573 (Ser9) (RED) and Tom20 (GREEN)respectively The cell nuclei were indicated by DAPI staining The images were captured under a fluorescence microscope (Carl-Zeiss JenaGermany) Scale bar 10 120583m
We found that formononetinmarkedly increased the bindingof phospho-GSK-3120573 (Ser9) to ANT (Figure 5(b))
The binding of phospho-GSK-3120573 (Ser9) tomitochondrialmembrane was further characterized by immunofluores-cence staining After 8 h ofOGDH9c2 cells were treatedwith30 120583M formononetin or equal volume of PBS under reoxy-genation condition for 20min The intracellular phospho-GSK-3120573 (Ser9) was sequentially stained by specific antibodyand Alexa Fluor 594-conjugated goat anti-rabbit IgG sec-ondary antibody Mitochondria were indicated by sequentialstaining of Tom20 with anti-Tom20 and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody DAPI
was used to visualize the cell nuclei As shown in Figure 5(c)formononetin dramatically increased the localization ofphospho-GSK-3120573 (Ser9) in mitochondria in close proximityto the cell nucleus
4 Discussion
Reperfusion of ischemic hearts is the main cause of theopening ofmPTP leading to the dysfunction ofmitochondria[22] The present study investigated that the effects of plantnatural product formononetin on the mPTP opening in anin vitro OGDreoxygenation-induced cell model of ischemia
8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
Relat
ive R
OS
leve
l fo
ld ch
ange
OGDR
0 5 10 30
0 0 10 30
OGDR
Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
lowast
mdash
(a)
0
05
1
15
2
25
Relat
ive s
uper
oxid
e lev
el fo
ld ch
ange
OGDR
10 30
OGDR
0 0
0 5 10 30Formononetin (120583M)
Formononetin (120583M)
lowastlowastlowast
mdash
(b)
Figure 2 Effects of formononetin on ROS and superoxide production in cardiac H9c2 cells H9c2 cells were treated with variousconcentrations of formononetin under OGD and reoxygenation (OGDR) condition Accumulation of ROS and superoxide inH9c2 cells wasdetected by probes DCFH2-DA and DHE respectively (a) Representative images and quantification of relative ROS levels The intracellularROS level in H9c2 cells was calculated based on fluorescence intensity The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (b) Representative images and quantification ofrelative superoxide levels The intracellular superoxide level in H9c2 cells was calculated based on fluorescence intensity The fluorescenceimages were analyzed with NIH Image J software (httpimagejnihgovij) The results were expressed as means plusmn SD (119899 = 3) 119901 lt 0001(OGDR versus Normal) lowast119901 lt 005 lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m
34 Effects of Formononetin on Akt and GSK-3120573 Phospho-rylation To discover the potential mechanisms we firstexamined the effects of formononetin on the phosphorylationof Akt and GSK-3120573 especially the phosphorylation of GSK-3120573 at Ser9 As shown in Figure 3(c) formononetin at theconcentrations (5ndash30120583M) induced the phosphorylation ofAkt and GSK-3120573 at Ser9 in a concentration-dependent man-ner We further verified the effects of formononetin on thephosphorylation of Akt and GSK-3120573 at Ser9 in the cells afterexposure to OGD and reoxygenation condition As shownin Figure 3(d) the levels of phospho-Akt and phospho-GSK-3120573 (Ser9) were suppressed under OGDreoxygenation condi-tion However formononetin increased Akt phosphorylationagainst OGDreoxygenation stimulation in a concentration-dependent manner Interestingly formononetin even at theconcentration of 5120583Meffectively reversed the loss of GSK-3120573phosphorylation against OGDreoxygenation challenge
35 Formononetin Activated Akt and PKC to PreserveMitochondrial Integrity To understand how formononetin
induces the phosphorylation of GSK-3120573 at Ser9 followingOGDreoxygenation challenge we treated the cells withformononetin in combination with PI3K inhibitor LY294002(LY) PKC inhibitor GF109203X (GF) and ERK 12 inhibitorPD98059 (PD) As shown in Figure 4(a) PI3K inhibitorLY and PKC inhibitor GF almost abolished the stimulatoryeffect of formononetin on the phosphorylation of GSK-3120573at Ser9 whereas ERK12 inhibitor did not show any activityagainst formononetin-induced formation of phospho-GSK-3120573 (Ser9) Supportively PI3K inhibitor LY and PKC inhibitorGF effectively antagonized the effects of formononetin onmitochondrial retention of TMRM and calcein-AM againstOGDreoxygenation damage (Figures 4(b) and 4(c))
36 Formononetin Stimulated the Binding of p-GSK-3120573 toANT in mPTP To verify the effect of formononetin on themPTP opening we examined the formation of proteincomplex between two main mPTP components ANT andCyp-D Following immunoprecipitation with anti-ANTantibody we analyzed the presence of ANT and Cyp-D by
Oxidative Medicine and Cellular Longevity 5
40 0 0 5 10 30
50
60
70
80
90
100
110
120
Relat
ive T
MRE
sequ
estr
atio
n
OGDR
Con
trol
OG
DR
mdashmdash
lowastlowast
lowast
FM (3
0120583
M) F
M (5
120583M
)
Formononetin (120583M)
(a)
160
140
120
100
80
60
40
20
0
Relat
ive c
alce
in fl
uore
scen
ce
OGDRmdash
lowastlowastlowast
lowastlowastlowast
Con
trol
OG
DR
mdashFM
(30120583
M)
FM (5
120583M
)
0 0 5 10 30Formononetin (120583M)
(b)
0 5 10 30
Fold
of i
nduc
tion
3
25
2
15
1
05
0
lowastlowast
lowastlowast lowastlowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
0
5120583M10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(c)
00 5 10 30
OGDR2
15
1
05
0
ControlOGDR
Treatment
Fold
of i
nduc
tion
mdash
lowastlowast
lowastlowast
lowastlowast
lowast lowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
5120583M
10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(d)
Figure 3 Effects of formononetin on mitochondrial membrane integrity and phosphorylation of Akt and GSK-3120573 (a) Fluorescence imagesand quantification of TMRM sequestration in mitochondria H9c2 cells were treated with formononetin (5 10 and 30 120583M) under normal orOGD and reoxygenation condition and stained with TMRM TMRMfluorescence intensity was determined by NIH image J software Resultsare presented as means plusmn SD (119899 = 3) 119901 lt 001 (OGDR versus normal) lowast119901 lt 005 lowastlowast119901 lt 001 (drug versus OGDR) Scale bar 10120583m(b) Fluorescence images and quantification of calcein sequestration in mitochondria H9c2 cells were treated similarly as described in (a)and stained with calcein-AM Calcein fluorescence intensity was determined by NIH image J software Results are presented as means plusmnSD (119899 = 3) 119901 lt 0001 (OGDR versus normal) lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (c) Western blot analysis andquantification of phosphorylation of GSK-3120573 and Akt in response to formononetin After the treatment with formononetin (5 10 and 30 120583M)under normal condition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blottingusing specific antibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt005 lowastlowast119901 lt 001 (drug versus control) (d) Western blot analysis and quantification of phosphorylation of GSK-3120573 and Akt in responseto formononetin under normal or OGDR condition After the treatment with formononetin (5 10 and 30 120583M) under normal or OGDRcondition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blotting using specificantibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt 005 lowastlowast119901 lt 001(drugs versus control and drugs versus OGDR)
6 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
GAPDH
Fold
of c
hang
e
30 303030Kinase inhibitor LY PDGFLYPDGF
TreatmentFormononetin (120583M)
lowast
lowastlowast
mdashmdashmdashmdash mdashmdash
30 303030Kinase inhibitor LY PDGFLYPDGF
Formononetin (120583M) mdashmdashmdashmdash mdashmdash
p-GSK-3120573
GSK-3120573
(a)
405060708090
100110120
OGDR
Mito
chon
dria
lTM
RE re
tent
ion
OGDR
lowast lowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdash
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash
mdash
mdash
30120583M 30120583M 30120583M
(b)
Relat
ive c
alce
in fl
uore
scen
ce
160140120100
80604020
0
OGDR
lowastlowastlowast lowastlowastlowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdashmdash
mdash
OGDR
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash30120583M 30120583M 30120583M
(c)
Figure 4 Roles of PI3K ERK12 and PKC in the regulation of GSK-3120573 phosphorylation and mitochondrial membrane integrity byformononetin (a) Western blot analysis and quantification of GSK-3120573 phosphorylation After the treatment with formononetin plusmn kinaseinhibitors H9c2 cells were lysed and analyzed for GSK-3120573 phosphorylation at Ser9 by Western blotting using specific antibodies Westernblots were determined by a densitometric method Results are presented as means plusmn SD (119899 = 3) 119901 lt 001 (drug versus OGDR)lowast119901 lt 005 lowastlowast119901 lt 001 (drug + inhibitor versus drug) (b and c) Fluorescence images and quantification of TMRMand calcein sequestration inmitochondria After the treatment with formononetin plusmn kinase inhibitors under OGD and reoxygenation condition H9c2 cells were stainedwith TMRM and calcein-AM and imaged under a fluorescence microscope (Carl-Zeiss Jena Germany) TMRM and calcein fluorescenceintensity were quantified by a densitometric method 119901 lt 0001 (drug versus OGDR) lowast119901 lt 005 (drug + inhibitor versus drug) LY PI3Kinhibitor LY294002 GF PKC inhibitor GF109203X PD ERK 12 inhibitor PD98059 Scale bar 10120583m
Western blotting with specific antibodies As shown in Fig-ure 5(a) OGDreoxygenation increased the formation ofANT-Cyp-D complex whereas formononetin diminishedOGDreoxygenation-induced formation of ANT-Cyp-D
complex On the other hand we performed immunopre-cipitation with anti-phospho-GSK-3120573 (Ser9) antibody andsubsequently analyzed the presence of ANT and phospho-GSK-3120573 (Ser9) by Western blotting with specific antibodies
Oxidative Medicine and Cellular Longevity 7
IB cyclophilin D
IB ANT
IP ANT OGDR
Formononetinmdashmdash
mdashmdashmdash
+ +
+
(a)
IB ANT
OGDRFormononetin
mdashmdash
mdashmdashmdash
+ +
+
IP p-GSK-3120573 (Ser9)
IB p-GSK-3120573 (Ser9)
(b)
Con
trol
OG
DR
PBS
OG
DR
form
onon
etin
p-GSK-3120573 (Ser9) Tom20 DAPI Merge
(c)
Figure 5 Effects of formononetin on the formation of ANT-Cyp-D complex and the binding of phospho-GSK-3120573 (Ser9) to ANT (a)Immunoprecipitation with antibody against ANT After the treatment with formononetin under OGD and reoxygenation condition H9c2cells were lysed for immunoprecipitation (IP) with antibody against ANT The pull-down materials were analyzed by immunoblotting (IB)with antibodies against ANT and Cyp-D (b) Immunoprecipitation with antibody against phospho-GSK-3120573 (Ser9) After the treatmentwith formononetin under OGD and reoxygenation condition H9c2 cells were lysed for immunoprecipitation (IP) with antibody againstphospho-GSK-3120573 (Ser9) The pull-down materials were analyzed by immunoblotting (IB) with antibodies against ANT and phospho-GSK-3120573 (Ser9) (c) Immunofluorescence imaging of the intracellular phospho-GSK-3120573 (Ser9) After the treatment with formononetin under OGDand reoxygenation condition H9c2 cells were probed with antibody against phospho-GSK-3120573 (Ser9) and mitochondrial membrane proteinTom20 and subsequently detected by fluorophore-labelled secondary antibodies phospho-GSK-3120573 (Ser9) (RED) and Tom20 (GREEN)respectively The cell nuclei were indicated by DAPI staining The images were captured under a fluorescence microscope (Carl-Zeiss JenaGermany) Scale bar 10 120583m
We found that formononetinmarkedly increased the bindingof phospho-GSK-3120573 (Ser9) to ANT (Figure 5(b))
The binding of phospho-GSK-3120573 (Ser9) tomitochondrialmembrane was further characterized by immunofluores-cence staining After 8 h ofOGDH9c2 cells were treatedwith30 120583M formononetin or equal volume of PBS under reoxy-genation condition for 20min The intracellular phospho-GSK-3120573 (Ser9) was sequentially stained by specific antibodyand Alexa Fluor 594-conjugated goat anti-rabbit IgG sec-ondary antibody Mitochondria were indicated by sequentialstaining of Tom20 with anti-Tom20 and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody DAPI
was used to visualize the cell nuclei As shown in Figure 5(c)formononetin dramatically increased the localization ofphospho-GSK-3120573 (Ser9) in mitochondria in close proximityto the cell nucleus
4 Discussion
Reperfusion of ischemic hearts is the main cause of theopening ofmPTP leading to the dysfunction ofmitochondria[22] The present study investigated that the effects of plantnatural product formononetin on the mPTP opening in anin vitro OGDreoxygenation-induced cell model of ischemia
8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 5
40 0 0 5 10 30
50
60
70
80
90
100
110
120
Relat
ive T
MRE
sequ
estr
atio
n
OGDR
Con
trol
OG
DR
mdashmdash
lowastlowast
lowast
FM (3
0120583
M) F
M (5
120583M
)
Formononetin (120583M)
(a)
160
140
120
100
80
60
40
20
0
Relat
ive c
alce
in fl
uore
scen
ce
OGDRmdash
lowastlowastlowast
lowastlowastlowast
Con
trol
OG
DR
mdashFM
(30120583
M)
FM (5
120583M
)
0 0 5 10 30Formononetin (120583M)
(b)
0 5 10 30
Fold
of i
nduc
tion
3
25
2
15
1
05
0
lowastlowast
lowastlowast lowastlowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
0
5120583M10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(c)
00 5 10 30
OGDR2
15
1
05
0
ControlOGDR
Treatment
Fold
of i
nduc
tion
mdash
lowastlowast
lowastlowast
lowastlowast
lowast lowastlowast
Formononetin (120583M)
p-AktAkt p-GSK-3120573GSK-3120573
5120583M
10120583M30120583M
p-Akt
Akt
p-GSK-3120573
GSK-3120573
GAPDH
(d)
Figure 3 Effects of formononetin on mitochondrial membrane integrity and phosphorylation of Akt and GSK-3120573 (a) Fluorescence imagesand quantification of TMRM sequestration in mitochondria H9c2 cells were treated with formononetin (5 10 and 30 120583M) under normal orOGD and reoxygenation condition and stained with TMRM TMRMfluorescence intensity was determined by NIH image J software Resultsare presented as means plusmn SD (119899 = 3) 119901 lt 001 (OGDR versus normal) lowast119901 lt 005 lowastlowast119901 lt 001 (drug versus OGDR) Scale bar 10120583m(b) Fluorescence images and quantification of calcein sequestration in mitochondria H9c2 cells were treated similarly as described in (a)and stained with calcein-AM Calcein fluorescence intensity was determined by NIH image J software Results are presented as means plusmnSD (119899 = 3) 119901 lt 0001 (OGDR versus normal) lowastlowastlowast119901 lt 0001 (drug versus OGDR) Scale bar 10120583m (c) Western blot analysis andquantification of phosphorylation of GSK-3120573 and Akt in response to formononetin After the treatment with formononetin (5 10 and 30 120583M)under normal condition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blottingusing specific antibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt005 lowastlowast119901 lt 001 (drug versus control) (d) Western blot analysis and quantification of phosphorylation of GSK-3120573 and Akt in responseto formononetin under normal or OGDR condition After the treatment with formononetin (5 10 and 30 120583M) under normal or OGDRcondition H9c2 cells were lysed and subsequently analyzed for phosphorylation of GSK-3120573 (Ser9) and Akt byWestern blotting using specificantibodies Western blots were quantified by a densitometric method Results are presented as means plusmn SD (119899 = 3) lowast119901 lt 005 lowastlowast119901 lt 001(drugs versus control and drugs versus OGDR)
6 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
GAPDH
Fold
of c
hang
e
30 303030Kinase inhibitor LY PDGFLYPDGF
TreatmentFormononetin (120583M)
lowast
lowastlowast
mdashmdashmdashmdash mdashmdash
30 303030Kinase inhibitor LY PDGFLYPDGF
Formononetin (120583M) mdashmdashmdashmdash mdashmdash
p-GSK-3120573
GSK-3120573
(a)
405060708090
100110120
OGDR
Mito
chon
dria
lTM
RE re
tent
ion
OGDR
lowast lowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdash
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash
mdash
mdash
30120583M 30120583M 30120583M
(b)
Relat
ive c
alce
in fl
uore
scen
ce
160140120100
80604020
0
OGDR
lowastlowastlowast lowastlowastlowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdashmdash
mdash
OGDR
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash30120583M 30120583M 30120583M
(c)
Figure 4 Roles of PI3K ERK12 and PKC in the regulation of GSK-3120573 phosphorylation and mitochondrial membrane integrity byformononetin (a) Western blot analysis and quantification of GSK-3120573 phosphorylation After the treatment with formononetin plusmn kinaseinhibitors H9c2 cells were lysed and analyzed for GSK-3120573 phosphorylation at Ser9 by Western blotting using specific antibodies Westernblots were determined by a densitometric method Results are presented as means plusmn SD (119899 = 3) 119901 lt 001 (drug versus OGDR)lowast119901 lt 005 lowastlowast119901 lt 001 (drug + inhibitor versus drug) (b and c) Fluorescence images and quantification of TMRMand calcein sequestration inmitochondria After the treatment with formononetin plusmn kinase inhibitors under OGD and reoxygenation condition H9c2 cells were stainedwith TMRM and calcein-AM and imaged under a fluorescence microscope (Carl-Zeiss Jena Germany) TMRM and calcein fluorescenceintensity were quantified by a densitometric method 119901 lt 0001 (drug versus OGDR) lowast119901 lt 005 (drug + inhibitor versus drug) LY PI3Kinhibitor LY294002 GF PKC inhibitor GF109203X PD ERK 12 inhibitor PD98059 Scale bar 10120583m
Western blotting with specific antibodies As shown in Fig-ure 5(a) OGDreoxygenation increased the formation ofANT-Cyp-D complex whereas formononetin diminishedOGDreoxygenation-induced formation of ANT-Cyp-D
complex On the other hand we performed immunopre-cipitation with anti-phospho-GSK-3120573 (Ser9) antibody andsubsequently analyzed the presence of ANT and phospho-GSK-3120573 (Ser9) by Western blotting with specific antibodies
Oxidative Medicine and Cellular Longevity 7
IB cyclophilin D
IB ANT
IP ANT OGDR
Formononetinmdashmdash
mdashmdashmdash
+ +
+
(a)
IB ANT
OGDRFormononetin
mdashmdash
mdashmdashmdash
+ +
+
IP p-GSK-3120573 (Ser9)
IB p-GSK-3120573 (Ser9)
(b)
Con
trol
OG
DR
PBS
OG
DR
form
onon
etin
p-GSK-3120573 (Ser9) Tom20 DAPI Merge
(c)
Figure 5 Effects of formononetin on the formation of ANT-Cyp-D complex and the binding of phospho-GSK-3120573 (Ser9) to ANT (a)Immunoprecipitation with antibody against ANT After the treatment with formononetin under OGD and reoxygenation condition H9c2cells were lysed for immunoprecipitation (IP) with antibody against ANT The pull-down materials were analyzed by immunoblotting (IB)with antibodies against ANT and Cyp-D (b) Immunoprecipitation with antibody against phospho-GSK-3120573 (Ser9) After the treatmentwith formononetin under OGD and reoxygenation condition H9c2 cells were lysed for immunoprecipitation (IP) with antibody againstphospho-GSK-3120573 (Ser9) The pull-down materials were analyzed by immunoblotting (IB) with antibodies against ANT and phospho-GSK-3120573 (Ser9) (c) Immunofluorescence imaging of the intracellular phospho-GSK-3120573 (Ser9) After the treatment with formononetin under OGDand reoxygenation condition H9c2 cells were probed with antibody against phospho-GSK-3120573 (Ser9) and mitochondrial membrane proteinTom20 and subsequently detected by fluorophore-labelled secondary antibodies phospho-GSK-3120573 (Ser9) (RED) and Tom20 (GREEN)respectively The cell nuclei were indicated by DAPI staining The images were captured under a fluorescence microscope (Carl-Zeiss JenaGermany) Scale bar 10 120583m
We found that formononetinmarkedly increased the bindingof phospho-GSK-3120573 (Ser9) to ANT (Figure 5(b))
The binding of phospho-GSK-3120573 (Ser9) tomitochondrialmembrane was further characterized by immunofluores-cence staining After 8 h ofOGDH9c2 cells were treatedwith30 120583M formononetin or equal volume of PBS under reoxy-genation condition for 20min The intracellular phospho-GSK-3120573 (Ser9) was sequentially stained by specific antibodyand Alexa Fluor 594-conjugated goat anti-rabbit IgG sec-ondary antibody Mitochondria were indicated by sequentialstaining of Tom20 with anti-Tom20 and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody DAPI
was used to visualize the cell nuclei As shown in Figure 5(c)formononetin dramatically increased the localization ofphospho-GSK-3120573 (Ser9) in mitochondria in close proximityto the cell nucleus
4 Discussion
Reperfusion of ischemic hearts is the main cause of theopening ofmPTP leading to the dysfunction ofmitochondria[22] The present study investigated that the effects of plantnatural product formononetin on the mPTP opening in anin vitro OGDreoxygenation-induced cell model of ischemia
8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
25
GAPDH
Fold
of c
hang
e
30 303030Kinase inhibitor LY PDGFLYPDGF
TreatmentFormononetin (120583M)
lowast
lowastlowast
mdashmdashmdashmdash mdashmdash
30 303030Kinase inhibitor LY PDGFLYPDGF
Formononetin (120583M) mdashmdashmdashmdash mdashmdash
p-GSK-3120573
GSK-3120573
(a)
405060708090
100110120
OGDR
Mito
chon
dria
lTM
RE re
tent
ion
OGDR
lowast lowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdash
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash
mdash
mdash
30120583M 30120583M 30120583M
(b)
Relat
ive c
alce
in fl
uore
scen
ce
160140120100
80604020
0
OGDR
lowastlowastlowast lowastlowastlowast
30 303030Kinase inhibitor PDGFLY
Formononetin (120583M) mdashmdash mdash mdashmdash
mdash
OGDR
Kinase inhibitor GFLYFormononetin
mdash mdashmdash mdash
mdash30120583M 30120583M 30120583M
(c)
Figure 4 Roles of PI3K ERK12 and PKC in the regulation of GSK-3120573 phosphorylation and mitochondrial membrane integrity byformononetin (a) Western blot analysis and quantification of GSK-3120573 phosphorylation After the treatment with formononetin plusmn kinaseinhibitors H9c2 cells were lysed and analyzed for GSK-3120573 phosphorylation at Ser9 by Western blotting using specific antibodies Westernblots were determined by a densitometric method Results are presented as means plusmn SD (119899 = 3) 119901 lt 001 (drug versus OGDR)lowast119901 lt 005 lowastlowast119901 lt 001 (drug + inhibitor versus drug) (b and c) Fluorescence images and quantification of TMRMand calcein sequestration inmitochondria After the treatment with formononetin plusmn kinase inhibitors under OGD and reoxygenation condition H9c2 cells were stainedwith TMRM and calcein-AM and imaged under a fluorescence microscope (Carl-Zeiss Jena Germany) TMRM and calcein fluorescenceintensity were quantified by a densitometric method 119901 lt 0001 (drug versus OGDR) lowast119901 lt 005 (drug + inhibitor versus drug) LY PI3Kinhibitor LY294002 GF PKC inhibitor GF109203X PD ERK 12 inhibitor PD98059 Scale bar 10120583m
Western blotting with specific antibodies As shown in Fig-ure 5(a) OGDreoxygenation increased the formation ofANT-Cyp-D complex whereas formononetin diminishedOGDreoxygenation-induced formation of ANT-Cyp-D
complex On the other hand we performed immunopre-cipitation with anti-phospho-GSK-3120573 (Ser9) antibody andsubsequently analyzed the presence of ANT and phospho-GSK-3120573 (Ser9) by Western blotting with specific antibodies
Oxidative Medicine and Cellular Longevity 7
IB cyclophilin D
IB ANT
IP ANT OGDR
Formononetinmdashmdash
mdashmdashmdash
+ +
+
(a)
IB ANT
OGDRFormononetin
mdashmdash
mdashmdashmdash
+ +
+
IP p-GSK-3120573 (Ser9)
IB p-GSK-3120573 (Ser9)
(b)
Con
trol
OG
DR
PBS
OG
DR
form
onon
etin
p-GSK-3120573 (Ser9) Tom20 DAPI Merge
(c)
Figure 5 Effects of formononetin on the formation of ANT-Cyp-D complex and the binding of phospho-GSK-3120573 (Ser9) to ANT (a)Immunoprecipitation with antibody against ANT After the treatment with formononetin under OGD and reoxygenation condition H9c2cells were lysed for immunoprecipitation (IP) with antibody against ANT The pull-down materials were analyzed by immunoblotting (IB)with antibodies against ANT and Cyp-D (b) Immunoprecipitation with antibody against phospho-GSK-3120573 (Ser9) After the treatmentwith formononetin under OGD and reoxygenation condition H9c2 cells were lysed for immunoprecipitation (IP) with antibody againstphospho-GSK-3120573 (Ser9) The pull-down materials were analyzed by immunoblotting (IB) with antibodies against ANT and phospho-GSK-3120573 (Ser9) (c) Immunofluorescence imaging of the intracellular phospho-GSK-3120573 (Ser9) After the treatment with formononetin under OGDand reoxygenation condition H9c2 cells were probed with antibody against phospho-GSK-3120573 (Ser9) and mitochondrial membrane proteinTom20 and subsequently detected by fluorophore-labelled secondary antibodies phospho-GSK-3120573 (Ser9) (RED) and Tom20 (GREEN)respectively The cell nuclei were indicated by DAPI staining The images were captured under a fluorescence microscope (Carl-Zeiss JenaGermany) Scale bar 10 120583m
We found that formononetinmarkedly increased the bindingof phospho-GSK-3120573 (Ser9) to ANT (Figure 5(b))
The binding of phospho-GSK-3120573 (Ser9) tomitochondrialmembrane was further characterized by immunofluores-cence staining After 8 h ofOGDH9c2 cells were treatedwith30 120583M formononetin or equal volume of PBS under reoxy-genation condition for 20min The intracellular phospho-GSK-3120573 (Ser9) was sequentially stained by specific antibodyand Alexa Fluor 594-conjugated goat anti-rabbit IgG sec-ondary antibody Mitochondria were indicated by sequentialstaining of Tom20 with anti-Tom20 and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody DAPI
was used to visualize the cell nuclei As shown in Figure 5(c)formononetin dramatically increased the localization ofphospho-GSK-3120573 (Ser9) in mitochondria in close proximityto the cell nucleus
4 Discussion
Reperfusion of ischemic hearts is the main cause of theopening ofmPTP leading to the dysfunction ofmitochondria[22] The present study investigated that the effects of plantnatural product formononetin on the mPTP opening in anin vitro OGDreoxygenation-induced cell model of ischemia
8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 7
IB cyclophilin D
IB ANT
IP ANT OGDR
Formononetinmdashmdash
mdashmdashmdash
+ +
+
(a)
IB ANT
OGDRFormononetin
mdashmdash
mdashmdashmdash
+ +
+
IP p-GSK-3120573 (Ser9)
IB p-GSK-3120573 (Ser9)
(b)
Con
trol
OG
DR
PBS
OG
DR
form
onon
etin
p-GSK-3120573 (Ser9) Tom20 DAPI Merge
(c)
Figure 5 Effects of formononetin on the formation of ANT-Cyp-D complex and the binding of phospho-GSK-3120573 (Ser9) to ANT (a)Immunoprecipitation with antibody against ANT After the treatment with formononetin under OGD and reoxygenation condition H9c2cells were lysed for immunoprecipitation (IP) with antibody against ANT The pull-down materials were analyzed by immunoblotting (IB)with antibodies against ANT and Cyp-D (b) Immunoprecipitation with antibody against phospho-GSK-3120573 (Ser9) After the treatmentwith formononetin under OGD and reoxygenation condition H9c2 cells were lysed for immunoprecipitation (IP) with antibody againstphospho-GSK-3120573 (Ser9) The pull-down materials were analyzed by immunoblotting (IB) with antibodies against ANT and phospho-GSK-3120573 (Ser9) (c) Immunofluorescence imaging of the intracellular phospho-GSK-3120573 (Ser9) After the treatment with formononetin under OGDand reoxygenation condition H9c2 cells were probed with antibody against phospho-GSK-3120573 (Ser9) and mitochondrial membrane proteinTom20 and subsequently detected by fluorophore-labelled secondary antibodies phospho-GSK-3120573 (Ser9) (RED) and Tom20 (GREEN)respectively The cell nuclei were indicated by DAPI staining The images were captured under a fluorescence microscope (Carl-Zeiss JenaGermany) Scale bar 10 120583m
We found that formononetinmarkedly increased the bindingof phospho-GSK-3120573 (Ser9) to ANT (Figure 5(b))
The binding of phospho-GSK-3120573 (Ser9) tomitochondrialmembrane was further characterized by immunofluores-cence staining After 8 h ofOGDH9c2 cells were treatedwith30 120583M formononetin or equal volume of PBS under reoxy-genation condition for 20min The intracellular phospho-GSK-3120573 (Ser9) was sequentially stained by specific antibodyand Alexa Fluor 594-conjugated goat anti-rabbit IgG sec-ondary antibody Mitochondria were indicated by sequentialstaining of Tom20 with anti-Tom20 and Alexa Fluor 488-conjugated goat anti-mouse IgG secondary antibody DAPI
was used to visualize the cell nuclei As shown in Figure 5(c)formononetin dramatically increased the localization ofphospho-GSK-3120573 (Ser9) in mitochondria in close proximityto the cell nucleus
4 Discussion
Reperfusion of ischemic hearts is the main cause of theopening ofmPTP leading to the dysfunction ofmitochondria[22] The present study investigated that the effects of plantnatural product formononetin on the mPTP opening in anin vitro OGDreoxygenation-induced cell model of ischemia
8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Oxidative Medicine and Cellular Longevity
reperfusion injury Following OGD challenge we mimickedreperfusion by incubating the cells with drugs under reoxy-genation condition as previously described [23] We discov-ered that formononetin could enhance the survival of H9c2cells and inhibit the mPTP opening through the potentialmechanisms involving the inhibition of ROS productionand Akt- and PKC-mediated phosphorylation of GSK-3120573 atSer9
Formononetin a major isoflavone from Radix Astragaliwas recently found to significantly reduce the infractedvolume and the brain water accumulation in a rat model ofischemia-reperfusion injury via the activation of PI3KAktsignaling pathway [17] Sulphonated formononetin alsoshowed cardioprotective effect on acute myocardial infarc-tion in rats possibly by regulating energy metabolism incardiac mitochondria [24] Inhibition of mPTP opening wasrecently suggested as a potential therapeutic target for thedevelopment of new cardioprotective drugs against myocar-dial infarction [25 26]These results stimulated us to explorewhether formononetin could exhibit cardioprotective effectsby regulating mPTP opening In this study we focused onthe effects of formononetin onOGDReoxygenation-induceddisruption of mitochondrial membrane integrity and theunderlying mechanisms We first verified that formononetincould effectively suppress the overproduction of ROS andsuperoxide ion afterOGDreoxygenation challengeWe char-acterized the effects of formononetin on the phosphorylationof GSK-3120573 (Ser9) Phosphorylation of GSK-3120573 at Ser9 causesthe reduction of GSK-3120573 activity [27] Inactivation of GSK-3120573 is known to modulate the mPTP opening conferringcardioprotection during ischemic preconditioning and post-conditioning [9 28]
In the present study we found that formononetinmarkedly induced the phosphorylation of GSK-3120573 at Ser9in a concentration-dependent manner Presumably for-mononetin could inactivate the enzymatic role of GSK-3120573 in cardiac H9c2 cells Importantly formononetin couldrecover the loss of phospho-GSK-3120573 (Ser9) formation againstOGDReoxygenation challenge To elucidate themechanismsbywhich formononetin stimulates the formation of phospho-GSK-3120573 (Ser9) we explored the roles of the PI3KAkt andPKCpathways It was previously demonstrated that PI3KAktand PKC directly phosphorylated GSK-3120573 at Ser9 [29]Another recent study reported that formononetin protectedcerebra against ischemiareperfusion injury via the activationof PI3KAkt signaling pathway [17] Indeed formononetininduced the activation of Akt in a concentration-dependentmanner Formononetin at the concentration of 30120583Machieved the most effective induction of Akt activation Onthe other hand formononetin at the concentration of 5120583Malready strongly promoted the formation of of phospho-GSK-3120573 (Ser9) Thus we employed specific kinase inhibitors toprofile the actions of Akt ERK12 and PKC on the phos-phorylation of GSK-3120573 at Ser9 Consequently we discoveredthat both Akt and PKC contributed to the phosphorylationof GSK-3120573 at Ser9 Importantly PI3K inhibitor LY294002and PKC inhibitor GF109203X not only antagonized thestimulatory effects of formononetin on the phosphorylationof GSK-3120573 at Ser9 but also attenuated the protective effects of
formononetin on mitochondrial membrane integrity Theseresults highlight that formononetinmaymaintain mitochon-drial membrane integrity via inducing the phosphorylationof GSK-3120573 at Ser9 through Akt- and PKC-dependent mecha-nisms We further demonstrated that formononetin inducedthe disassociation of ANT-Cyp-D complex and promotedthe binding of phospho-GSK-3120573 (Ser9) to ANT MoreoverGSK-3120573 is a multifunctional enzyme that regulates severalcellular functions and transcription factors [30] NuclearGSK-3120573 level is markedly increased prior to the activation ofthe caspase cascade leading to apoptosis [31] Interestinglyour immunofluorescence analysis suggested that phospho-GSK-3120573 (Ser9) might be deposited in mitochondria in closeproximity to the cell nucleus The deposition of p-GSK-3120573at the interface between mitochondria and cell nucleus maylimit the flux of ROS into cell nucleus and thereby suppresscell apoptosis
The mPTP opening may be regulated by posttranslo-cational modification of Cyp-D and physical interactionof Cyp-D with matrix proteins (eg p53 PPAR alpha)[32] It was recently reported that active GSK-3120573 couldphosphorylate Cyp-D and stimulate the opening of mPTP[33] Conversely the suppression of GSK-3120573 resulted in theinhibition of mPTP opening by attenuating the phospho-rylation of Cyp-D in murine tubular epithelial cells [34]In the present study we found that formononetin couldinduce GSK-3120573 phosphorylation at Ser9 causing the lossof GSK-3120573 enzyme activity Taken together it is possiblethat formononetin inhibited the phosphorylation of Cyp-Dcontributing to the inhibition of mPTP opening In additionto ANT Cyp-D interacts with several other proteins (egp53) to induce PTP opening and necrosis Tumor suppressorprotein p53 is accumulated in the mitochondrial matrix andinteracts with Cyp-D during ischemiareperfusion injuryConversely the reduction of p53 is protective during strokedue to the loss of Cyp-D and p53 complex [35] Oxidativestress could induce the formation of Cyp-D-PPAR120572 complexin cardiomyocytes while the disruption of Cyp-D-PPAR120572complex by metformin could protect cardiomyocytes againstoxidative stress-induced cell death [36] Moreover recentstudies provided new evidences to support the roles ofCyp-D and F1F0-ATPase in the control of mPTP open-ing F1F0-ATPase mainly affects mitochondrial ATP storage[37] Suppression of the F1F0-ATPase in the mitochondrialmatrix could conserve energy during the cardioprotectionintervention of Bcl-2 or GSK-3120573 inhibitors although GSK-3120573 inhibitors could not affect the activity of F1F0-ATPase[38] Others also demonstrated a central role for mitochon-drial GSK-3120573 but not F1F0-ATPase as target of PKC120585 tolimit IR damage [39] In the present study we discoveredthat formononetin could disrupt the interaction betweenANT and Cyp-D The disassociation of ANT from Cyp-D likely indicates that formononetin dismantles the mPTPcomplex via stimulating GSK-3120573 phosphorylation at Ser9and inhibiting the enzymatic activity of GSK-3120573 Furtherwork is required to clarify (1) whether formononetin affectsthe interactions of Cyp-D with p53 and PPAR120572 and (2)whether formononetin affects the activity of mitochondrialF1F0-ATPase
Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 9
OGDreoxygenation
PKC Akt
P
Cell death
Formononetin
Binding
Activating
mPTP
ROS
ROS
Mitochondria
Cyp-
D HKII
VDACANT
-P
ANT
GSK-3120573
p-GSK-3120573 (Ser9)
Figure 6 Proposed mechanism underlying the cardioprotectiveeffect of formononetin Formononetin activates PI3KAkt and PKCsignaling pathways and subsequently induces the phosphorylationof GSK-3120573 at Ser9 The phosphorylation of GSK-3120573 at Ser9 leadsto the inactivation of GSK-3120573 in the opening of mPTP On theother hand phospho-GSK-3120573 (Ser9) migrates towards mitochon-dria and binds to ANT causing the disassociation of the mPTPcomplex involving ANT Cyp-D VDAC and HKII Formononetinalso inhibits the overproduction of ROS and superoxide possi-bly reducing the oxidative damage of mitochondria in cardiomy-ocytes
In addition it was reported that the phosphorylationof GSK-3120573 at Ser9 suppressed mPTP opening via inter-acting with and phosphorylating VDAC [40] ActivatedGSK-3120573 also phosphorylates VDAC and thereby disruptsthe binding of hexokinase II to mitochondrial components[41] However the exact role of VDAC in the regulationof mPTP opening remains elusive The results from ourstudy defined that formononetin inhibited the activity ofGSK-3120573 by Ser9 phosphorylation and did not exclude theinteractions of GSK-3120573 and phospho-GSK-3120573 (Ser9) withVDAC
In summary the results from the present study sug-gest that formononetin may protect cardiomyocytes againstischemia reperfusion injury by inhibiting themPTP openingThe potential underlying mechanisms could be outlined inFigure 6 Formononetin may initiate its actions by suppress-ing the production of ROS and superoxide ion and activatingAkt and PKC pathways Upon activation Akt and PKCsubsequently phosphorylate GSK-3120573 at Ser9 leading to directinhibition of GSK-3120573 activity towards the mPTP openingand the disruption of the protein complex involving ANTand Cyp-D Ultimately formononetin attenuated the mPTPopening and maintained mitochondrial membrane integrityThus formononetin may serve as a useful lead compoundfor future development of new cardioprotective drugs againstacute myocardial infarction
Conflict of Interests
The authors declare that there is no conflict of interests
Authorsrsquo Contribution
Yuanyuan Cheng performed experiments analyzed data andwrote the paper Zhengyuan Xia and Yifan Han discussed thestudy and contributed reagents Jianhui Rong designed theresearch and wrote the paper
Acknowledgments
This work was supported by General Research Fund (HKU775812 M) from the Research Grants Council of Hong Kongand the Seed Fund for Basic Research Programme Universityof Hong Kong (to Jianhui Rong)
References
[1] D M Yellon and D J Hausenloy ldquoMyocardial reperfusioninjuryrdquo The New England Journal of Medicine vol 357 no 11pp 1074ndash1135 2007
[2] P Pagliaro FMoro F TullioM-G Perrelli andC Penna ldquoCar-dioprotective pathways during reperfusion focus on redoxsignaling and other modalities of cell signalingrdquo Antioxidantsamp Redox Signaling vol 14 no 5 pp 833ndash850 2011
[3] R S Whelan V Kaplinskiy and R N Kitsis ldquoCell death in thepathogenesis of heart disease mechanisms and significancerdquoAnnual Review of Physiology vol 72 pp 19ndash44 2009
[4] J N Weiss P Korge H M Honda and P Ping ldquoRole of themitochondrial permeability transition in myocardial diseaserdquoCirculation Research vol 93 no 4 pp 292ndash301 2003
[5] E J Griffiths and A P Halestrap ldquoMitochondrial non-specific pores remain closed during cardiac ischaemia but openupon reperfusionrdquo Biochemical Journal vol 307 no 1 pp 93ndash98 1995
[6] D J Hausenloy M R Duchen and D M Yellon ldquoInhibitingmitochondrial permeability transition pore opening at reperfu-sion protects against ischaemia-reperfusion injuryrdquo Cardiovas-cular Research vol 60 no 3 pp 617ndash625 2003
[7] R S Jope C J Yuskaitis and E Beurel ldquoGlycogen synthasekinase-3 (GSK3) inflammation diseases and therapeuticsrdquoNeurochemical Research vol 32 no 4-5 pp 577ndash595 2007
[8] TMiuraMNishihara and TMiki ldquoDrug development target-ing the glycogen synthase kinase-3120573 (GSK-3120573)-mediated signaltransduction pathway role of GSK-3120573 inmyocardial protectionagainst ischemiareperfusion injuryrdquo Journal of Pharmacologi-cal Sciences vol 109 no 2 pp 162ndash167 2009
[9] M Nishihara T Miura T Miki et al ldquoModulation of the mito-chondrial permeability transition pore complex in GSK-3120573-mediated myocardial protectionrdquo Journal of Molecular and Cel-lular Cardiology vol 43 no 5 pp 564ndash570 2007
[10] A Fugh-Berman ldquoHerbs and dietary supplements in the pre-vention and treatment of cardiovascular diseaserdquo PreventiveCardiology vol 3 no 1 pp 24ndash32 2000
10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Oxidative Medicine and Cellular Longevity
[11] S Lu K-J Chen Q-Y Yang and H-R Sun ldquoProgress in theresearch of Radix Astragali in treating chronic heart failureeffective ingredients dose-effect relationship and adverse reac-tionrdquo Chinese Journal of Integrative Medicine vol 17 no 6 pp473ndash477 2011
[12] K K-W Auyeung P-C Law and J K-S Ko ldquoNovel anti-angiogenic effects of formononetin in human colon cancer cellsand tumor xenograftrdquoOncology Reports vol 28 no 6 pp 2188ndash2194 2012
[13] Y-M Jin T-M Xu Y-H Zhao Y-CWang andM-H Cui ldquoInvitro and in vivo anti-cancer activity of formononetin onhumancervical cancer cell line HeLardquo Tumor Biology vol 35 no 3 pp2279ndash2284 2014
[14] Z Ma W Ji Q Fu and S Ma ldquoFormononetin inhibited theinflammation of LPS-induced acute lung injury in mice associ-atedwith induction of PPAR gamma expressionrdquo Inflammationvol 36 no 6 pp 1560ndash1566 2013
[15] W C Jia G Liu C D Zhang et al ldquoFormononetin attenu-ates hydrogen peroxide (H
2O2)-induced apoptosis and NF-120581B
activation in RGC-5 cellsrdquo European Review for Medical andPharmacological Sciences vol 18 no 15 pp 2191ndash2197 2014
[16] H Wang D Zhang M Ge Z Li J Jiang and Y Li ldquoFor-mononetin inhibits enterovirus 71 replication by regulatingCOX- 2PGE
2expressionrdquoVirology Journal vol 12 no 1 article
35 2015[17] K Liang Y Ye Y Wang J Zhang and C Li ldquoFormonone-
tin mediates neuroprotection against cerebral ischemiareper-fusion in rats via downregulation of the BaxBcl-2 ratio andupregulation PI3KAkt signaling pathwayrdquo Journal of the Neu-rological Sciences vol 344 no 1-2 pp 100ndash104 2014
[18] M Sun T Zhou L Zhou et al ldquoFormononetin protects neu-rons against hypoxia-induced cytotoxicity through upregula-tion of ADAM10 and sA120573PP120572rdquo Journal of Alzheimerrsquos Diseasevol 28 no 4 pp 795ndash808 2012
[19] J-E Huh D-W Nam Y-H Baek et al ldquoFormononetin accel-erates wound repair by the regulation of early growth responsefactor-1 transcription factor through the phosphorylation of theERK and p38 MAPK pathwaysrdquo International Immunopharma-cology vol 11 no 1 pp 46ndash54 2011
[20] R C Scaduto Jr and LW Grotyohann ldquoMeasurement of mito-chondrial membrane potential using fluorescent rhodaminederivativesrdquoBiophysical Journal vol 76 no 1 pp 469ndash477 1999
[21] A Prathapan V P Vineetha and K G Raghu ldquoProtective effectof Boerhaavia diffusa Lagainst mitochondrial dysfunction inangiotensin II induced hypertrophy in H9c2 cardiomyoblastcellsrdquo PLoS ONE vol 9 no 4 Article ID e96220 2014
[22] A P Halestrap S J Clarke and S A Javadov ldquoMitochon-drial permeability transition pore opening during myocar-dial reperfusionmdasha target for cardioprotectionrdquoCardiovascularResearch vol 61 no 3 pp 372ndash385 2004
[23] W-Y Wu W-Y Wang Y-L Ma et al ldquoSodium tanshinone IIAsilate inhibits oxygen-glucose deprivationrecovery-inducedcardiomyocyte apoptosis via suppression of the NF-120581BTNF-120572 pathwayrdquo British Journal of Pharmacology vol 169 no 5 pp1058ndash1071 2013
[24] S Zhang X Tang J Tian et al ldquoCardioprotective effect ofsulphonated formononetin on acute myocardial infarction inratsrdquo Basic amp Clinical Pharmacology amp Toxicology vol 108 no6 pp 390ndash395 2011
[25] D Morin R Assaly S Paradis and A Berdeaux ldquoInhibitionof mitochondrial membrane permeability as a putative phar-macological target for cardioprotectionrdquo Current MedicinalChemistry vol 16 no 33 pp 4382ndash4398 2009
[26] S Javadov and M Karmazyn ldquoMitochondrial permeabilitytransition pore opening as an endpoint to initiate cell death andas a putative target for cardioprotectionrdquo Cellular Physiologyand Biochemistry vol 20 no 1ndash4 pp 1ndash22 2007
[27] W M Blankesteijn V A M van de Schans P ter Horst andJ F M Smits ldquoThe WntfrizzledGSK-3120573 pathway a noveltherapeutic target for cardiac hypertrophyrdquo Trends in Pharma-cological Sciences vol 29 no 4 pp 175ndash180 2008
[28] L Gomez M Paillard H Thibault G Derumeaux and MOvize ldquoInhibition of GSK3120573 by postconditioning is required toprevent opening of the mitochondrial permeability transitionpore during reperfusionrdquo Circulation vol 117 no 21 pp 2761ndash2768 2008
[29] G Chanoit S Lee J Xi et al ldquoExogenous zinc protects car-diac cells from reperfusion injury by targeting mitochondrialpermeability transition pore through inactivation of glycogensynthase kinase-3120573rdquo American Journal of PhysiologymdashHeartand Circulatory Physiology vol 295 no 3 pp H1227ndashH12332008
[30] S Pedretti and E Raddatz ldquoSTAT3120572 interacts with nuclearGSK3120573 and cytoplasmic RISK pathway and stabilizes rhythmin the anoxic-reoxygenated embryonic heartrdquo Basic Research inCardiology vol 106 no 3 pp 355ndash369 2011
[31] G N Bijur and R S Jope ldquoProapoptotic stimuli induce nuclearaccumulation of glycogen synthase kinase-3120573rdquo The Journal ofBiological Chemistry vol 276 no 40 pp 37436ndash37442 2001
[32] S Javadov and A Kuznetsov ldquoMitochondrial permeabilitytransition and cell death the role of cyclophilin drdquo Frontiers inPhysiology vol 4 article 76 2013
[33] A Rasola M Sciacovelli F Chiara B Pantic W S Brusilowand P Bernardi ldquoActivation of mitochondrial ERK protectscancer cells from death through inhibition of the permeabilitytransitionrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 2 pp 726ndash731 2010
[34] H Bao Y Ge S Zhuang L D Dworkin Z Liu and R GongldquoInhibition of glycogen synthase kinase-3120573 prevents NSAID-induced acute kidney injuryrdquo Kidney International vol 81 no7 pp 662ndash673 2012
[35] A V Vaseva N D Marchenko K Ji S E Tsirka S Holzmannand U M Moll ldquoP53 opens the mitochondrial permeabilitytransition pore to trigger necrosisrdquoCell vol 149 no 7 pp 1536ndash1548 2012
[36] G Barreto-Torres J S Hernandez S Jang et al ldquoThe beneficialeffects of AMP kinase activation against oxidative stress areassociated with prevention of PPAR120572-cyclophilin D interactionin cardiomyocytesrdquoTheAmerican Journal of PhysiologymdashHeartandCirculatory Physiology vol 308 no 7 ppH749ndashH758 2015
[37] B Agarwal D F Stowe R K Dash Z J Bosnjak and A K SCamara ldquoMitochondrial targets for volatile anesthetics againstcardiac ischemia-reperfusion injuryrdquo Frontiers in Physiologyvol 5 article 341 2014
[38] C Steenbergen S Das J Su R Wong and E Murphy ldquoCar-dioprotection and altered mitochondrial adenine nucleotidetransportrdquo Basic Research in Cardiology vol 104 no 2 pp 149ndash156 2009
Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 11
[39] D K Bowles K K Maddali V C Dhulipala and D H Korz-ick ldquoPKC120575 mediates anti-proliferative pro-apoptic effects oftestosterone on coronary smooth musclerdquo American Journal ofPhysiologymdashCell Physiology vol 293 no 2 pp C805ndashC8132007
[40] S Javadov V Rajapurohitam A Kilic A Zeidan A Choi andM Karmazyn ldquoAnti-hypertrophic effect of NHE-1 inhibitioninvolves GSK-3120573-dependent attenuation of mitochondrial dys-functionrdquo Journal of Molecular and Cellular Cardiology vol 46no 6 pp 998ndash1007 2009
[41] J G Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3beta disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom





![Research Article 2-Heptyl-Formononetin Increases ...downloads.hindawi.com/journals/bmri/2013/926942.pdf · BioMed Research International decreasesbodyweightandfatmass[ ],lowerstheplasma](https://img.pdfslide.net/doc/110x75/5fcff57faf36410a6221c8df/research-article-2-heptyl-formononetin-increases-biomed-research-international.jpg)























