Embed Size (px)
Citation preview

Research ArticleTrypanosoma Infection Rates in Glossina Species inMtito Andei Division Makueni County Kenya
Daniel Mutiso Nthiwa1 David O Odongo2 Horace Ochanda2
Samoel Khamadi3 and Bernard M Gichimu4
1Department of Biological Sciences Embu University College PO Box 6 Embu 60100 Kenya2School of Biological Sciences University of Nairobi PO Box 30197 Nairobi 00100 Kenya3Kenya Medical Research Institute Center for Virus Research PO Box 54840 Nairobi 00200 Kenya4Department of Agricultural Resource Management Embu University College PO Box 6 Embu 60100 Kenya
Correspondence should be addressed to Daniel Mutiso Nthiwa danielmutiso8gmailcom
Received 17 August 2015 Revised 14 October 2015 Accepted 15 October 2015
Academic Editor Jose F Silveira
Copyright copy 2015 Daniel Mutiso Nthiwa et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
AfricanAnimal Trypanosomiasis (AAT) transmitted cyclically by tsetse fly (Glossina spp) is amajor obstacle to livestock productionin the tropical parts of Africa The objective of this study was to determine the infection rates of trypanosomes in Glossina speciesin Mtito Andei Division Makueni County Kenya Tsetse fly species G longipennis and G pallidipes were trapped and DNA wasisolated from their dissected internal organs (proboscis salivary glands and midguts) The DNA was then subjected to a nestedPCR assay using internal transcribed spacer primers and individual trypanosome species were identified following agarose gelelectrophoresis Out of the 117 flies trapped in the area 39 (333) were teneral while 78 (67) were nonteneral G pallidipesconstituted the largest percentage of 58 while G longipennis were 42The overall trypanosomes infection rate in all nonteneralGlossina spp was 1153 with G longipennis recording the highest infection rate of 2308 while G pallidipes had an infectionrate of 577 T vivax was the most infectious (1026) compared to T congolense (128) Mean apparent densities were stronglypositively correlatedwith infection rates (r = 095) confirming the importance of this parameter as an indicator of AAT transmissionrisk
1 Introduction
AfricanAnimal Trypanosomiasis (AAT) is a serious livestockdisease caused by a Kinetoplastida protozoan parasite ofthe genus Trypanosoma and transmitted cyclically by tsetse(Glossina) and mechanically by other hematophagous fliessuch as Tabanus Haematopota Stomoxys and Chrysops [12] It is a major constraint of livestock production in sub-Saharan Africa as it prevents full use of the land to feedthe rapidly increasing human population [3] Subsequentlythis contributes to poverty hunger and underdevelopmentin the affected areas [4] This disease is therefore a majorimpediment to agricultural and economic advancement inthe endemic areas [5 6] AAT and its vectors occur in large
areas of sub-Saharan Africa and its occurrence parallels thatof the biological vector [7 8] The epidemiology and effectsof this disease on livestock especially cattle production aredetermined largely by the prevalence and distribution of thedisease and its vectors in the affected areas and also theproportion of infected flies in the tsetse population [9] Theecodistribution of tsetse is determined by climate presence ofvegetation water and presence of blood meals (humans andanimals) but tsetse are mostly found in rural areas [10]
The economic losses due to AAT are approximated tobe over 15 billion dollars annually [11] Tsetse infest about10 million km2 of fertile land in Africa spread across 36countries with approximately 7 million km2 of the infestedland being suitable for mixed agriculture if the disease was
Hindawi Publishing CorporationJournal of Parasitology ResearchVolume 2015 Article ID 607432 8 pageshttpdxdoiorg1011552015607432
2 Journal of Parasitology Research
controlled [11] Out of 165 million cattle found in Africa only10 million are found within the tsetse belt due to the diseaseconstraint and these are the lowest producing breeds [12]TheKenyan economy is largely agricultural-based with the sectoraccounting for about 24 of the Gross Domestic Product(GDP) and 70 of the employment [13] Only 7 of Kenyanland is suitable for crop production and a further 5 cansustain crops in years of adequate rains [13] The rest of landis arid and semiarid and constitutes the range lands which arenot suitable for crop production but rather good for livestockproduction [13 14]
Mtito Andei is located in Makueni County about 290 kmSouth East of Nairobi The area is comprised mainly of aridand semiarid lands and livestock rearing is the mainstay ofthis region [14] This area has a history of animal trypanoso-miasis as it is infested with G pallidipes G longipennisand G brevipalpis [14 15] The area was not inhabiteduntil the 1930s due to its low agricultural potential andheavy tsetse infestation Assessment of prevalence vectorhotspots and infection status can be instrumental in makingdecisions with regard to the formulations of suitable tsetseand trypanosomiasis control measures However there isno accurate information that exists in regard to the currentprevalence of trypanosomiasis inMtitoAndeiThis is becauseprevious data has been collected through microscopy onblood smears dissection and microhematocrit techniqueswhich are less sensitive [16 17] Further these methodscannot identify immature andmixed infections subsequentlyresulting to low detection levels [18] This study was aimedat providing additional information on the prevalence oftrypanosomes in Glossina species in Mtito Andei
2 Materials and Method
21 Description of the Study Site This study was carried outin Mtito Andei Division situated in Makueni County about290 kmSouth East ofNairobiThe study areawas divided intofive villages namely Nthunguni Iviani Kyusyani Ngiluniand Kamunyu (Figure 1) The altitude of the area varies from600m to 1100m above sea level The area is characterizedby marginal agricultural lands of Savannah grassland withmostly low lying gently eastward sloping plains occasionallybroken by seasonal and perennial rivers [20] The climate ofthis area is typically semiarid with low and unreliable rainfallaveraging 600mm annually and annual average temperatureof 23∘C [20] Rainfall is bimodal with long rains occurringfrom March to May and short rains from November orDecember to early January The highest mean temperaturesbetween 32∘C and 33∘C prevail in February to March [20]
22 Study Design A cross-sectional study using simplerandom sampling technique was conducted to determinethe infection rates of trypanosomes in Glossina species inMtito Andei Division Sample size was determined using theformula (119899 = 119885211987511987621198892) described in [21] where 119899 is theminimum sample size required 119885 is the confidence interval119875 is the expected prevalence in proportion from the study 119876is 100 minus 119875 and 119889 is the desired precision taken at 005
Ethiopia
Kenya Somalia
Mtito AndeiTanzania100 km
N
Kamunyu
Iviani
Nthunguni
KyusyaniNgiluni
0 10
(km)
Figure 1 Map of Mtito Andei showing the georeferenced trappingsites
Figure 2 A biconical trap baited with cow urine and acetone toenhance trapping efficiency
23 Collection and Identification of Tsetse Tsetse trappingwas done between April and May 2012 This period was cho-sen because it is a rainy season and therefore more abundantflies were expected [1] Trapping was done using 5 biconicaltraps in each village which were deployed near Mtito AndeiRiver and Athi River and in the nearby vegetation The trapswere set at georeferenced sites and baited with acetone andcow urine to increase trapping efficiency [22] The trapswere set at regular intervals of 100 meters along variousvegetation types and under shade to avoid undue flymortalitydue to heat (Figure 2) To prevent the ascent of ants on thepoles towards the collecting cages each pole was smearedwith grease All trapped flies were counted their sex was
Journal of Parasitology Research 3
determined and identified to species level using conventionalidentification keys before dissection [23]
24 Determination of Apparent Density (AD) Apparent den-sity (AD) is an estimation of fliesrsquo density and is given in termsof the number of flies caught per trap per day (119865119879119863) It wascalculated using the following formula
119865119879119863 =sum119865
119879 times 119863 (1)
where sum119865 is the number of total flies 119879 is the number offunctioning traps and119863 is the number of days forwhich trapswere operational [1]
The average AD for each trapping site was then calculatedto obtain the data on tsetse distribution in the area
25 Identification of Trypanosomes in Glossina spp
251 Tsetse Dissection The flies were immobilized usingethyl acetate placed in a cotton wool at the bottom ofa specimen tube Dissection was carried out on freshlyimmobilized flies as trypanosomes are less likely to be foundin dead dried flies [23] A total of 78 nonteneral flies weredissected from which midgut salivary glands and probosciswere removed and analyzed under a microscope for thepresence of trypanosomes Nonteneral flies were identified bythe presence of dark or brown colour on their abdomenwhichindicated the last blood meal [23] Freshly immobilized flieswere surface sterilized by brief immersion in 70 ethanolthen dry blotted on clean tissue paper and then dissectedin normal saline [22] To avoid cross contamination thedissecting instruments were sterilized by immersion in 3ndash5 (wv) sodium hypochlorite for approximately 2 minutesfollowed by extensive rinsing in distilled water and later finalimmersion in normal saline [22] Dissected organs (midgutproboscis and salivary glands) from each fly were transferredinto 15mL microcentrifuge tube containing 120120583L of tissuelysis buffer (from Qiagen DNeasy Blood and Tissue Kit) Thetissues were then stored at minus20∘C until further processing
252 Trypanosomesrsquo DNA Extraction DNA was extractedusing the Qiagen DNeasy blood and tissue kit and accordingto the manufacturerrsquos instructions Twenty (20) microliters(120583L) of proteinase K was added to the dissected tissues andincubated at 56∘C overnight in a thermocycler until the tissuewas completely digested The lysate was vortexed for 15 secand 200120583L of buffer AL was added to the sample followedby vortexing to mix the contents Two hundred (200) 120583L ofabsolute ethanol was added andmixed thoroughly by furthervortexing before pipetting themixture intoDNeasymini spincolumns placed in a 2mL collection tube The columns werecentrifuged at 8000 rpm for 1 minute and the flow throughdiscarded together with the collection tube The DNeasymini spin columns were placed in a new 2mL collectiontube and 500 120583L of wash buffer AW1 was added It was thencentrifuged at 8000 rpm for 1 minute and the flow throughtogether with the collection tube discardedThe spin columnwas again placed in a new collection tube and 500120583L of
Table 1 Sequence of primers
Primer ID Primer sequence (51015840-31015840)Outerprimers
ITS 1 GAT TAC GTC CCT GCC ATT TGITS 2 TTG TTC GCT ATC GGT CTT CC
Innerprimers
ITS 3 GGA AGC AAA AGT CGT AAC AAG GITS 4 TGT TTT CTT TTC CTC CGC
SSU18S ITS 1 58S ITS 2 LSU(1)
28S
Figure 3 Schematic diagram of rDNA showing ITS 1 and ITS 2annealing positions [19]
wash buffer AW2 was added The column was centrifuged at14000 rpm for 3minutesThe flow through and the collectiontubes were discarded and the spin columns placed in a clean15mL microcentrifuge tube followed by addition of 30 120583Lof elution buffer (AE buffer) The column was incubated atroom temperature for 1 minute and centrifuged for 1 minuteat 8000 rpm to elute the bound DNA DNA extracted fromsamples was quantified using a nanodrop spectrophotometer(Thermo Fisher Scientific Inc) and its purity was determinedby measuring the intensity of absorbance at wavelengths 260and 280 nanometers
253 PCR Amplification and DNA Analysis The PCRoligonucleotide primers used are shown in Table 1 Theprimers were synthesized by Bioneer Company andwere pro-vided by International Livestock Research Institute (ILRI)Kenya
These primers were designed to amplify the internaltranscribed (ITS) region of ribosomal DNA (rDNA) a regionwhich varies in size within trypanosome species except formembers of Trypanozoon and are capable of differentiatingtrypanosomes based on their amplified fragments ITS 1 andITS 2 (outer primers) amplify the conserved regions of rDNAsmall subunit (SSU) and large subunit (LSU) representedby black arrows in Figure 3 while ITS 3 and ITS 4 (innerprimers) amplify the noncoding spacer regions of rDNA andare represented by white arrows
All PCR amplifications were performed in 30 120583L reactionvolumes containing final concentrations of 1x InvitrogenPCR buffer 35mM MgCl
2(Promega) 02mM dNTPs mix
125U120583L Invitrogen Taq 100 ng120583L of each forward andreverse primer and 5 120583L of trypanosome DNA as templateA set of purified genomic DNA of T brucei brucei (Tbb) Tcongolense Kilifi (Tck) and T congolense Savannah (Tcs)(strain IL1180) were included as positive controls during allPCR assays and nuclease free water as negative control Theouter primers (ITS 1 and 2) were used in the first roundof reaction The cycling conditions were as follows 1 cycleof 95∘C for 7 minutes as initial denaturation followed by 35cycles of 94∘C for 1 minute 55∘C for 1 minute and 72∘C for 2minutes The final extension was done at 72∘C for 5 minutes
4 Journal of Parasitology ResearchEa
ch sp
ecie
s (
)
Kamunyu Ngiluni Kyusyani Iviani NthunguniTrapping sites
G pallidipesG longipennis
0
20
40
60
80
100
120
Figure 4 Distribution of Glossina species captured from study sitesin Mtito Andei Division
The PCR was carried out on a Bio-Rad thermocycler In thesecond round of reaction the outer primers were substitutedwith inner primers (ITS 3 and 4) and 5 120583L of round one PCRproduct was used as DNA template The cycling conditionsfor the second round of reaction were as described for theprimary amplification To analyze the amplicons 10120583L ofthe PCR product was resolved in a 2 agarose gel at 80volts for 45 minutes and the gel was visualised under UVtransilluminator following ethidium bromide staining
26 Data Analysis The infectious trypanosomes were iden-tified by comparing the molecular sizes of their DNA frag-ments with the documented band sizes of trypanosomespecies [19]The data was then used to calculate the infectionrates and prevalence of the trypanosomes on Glossina sppConventional data on vector distribution apparent densitiesand infection rates from the five sites was subjected to one-way analysis of variance (ANOVA) using SPSS version 160software at 95 confidence interval Chi-square test was doneto determine the association of infection rates with flyrsquos sexand vector species A correlation analysis was also done toestablishwhether there was a significant relationship betweeninfection rates and apparent densities
3 Results
31 Vector Distribution Out of the 117 flies trapped in thearea G pallidipes constituted the largest percentage of 58whileG longipennis had 42 Among the trapped tsetse flies39 (333) were teneral while 78 (67) were nonteneral InKamunyu area G longipennis was the most prevalent specieswhile G pallidipes dominated Ngiluni Kyusyani Iviani andNthunguni (Figure 4)
Apart from Glossina spp the area was also inhabitedwith mechanical flies such as Tabanus spp and Stomoxys sppTabanids were highly predominant over Stomoxines in all thetrapping sites with an overall fly trap density (FTD) of 011while Stomoxys spp had FTD of 002 Of the total fly catchGlossina spp were the most prevalent in all the tapping sites
FliesGlossina spp Tabanus spp Stomoxys spp
0
10
20
30
40
50
60
70
80
90
100
()
Figure 5 Relative distribution of total fly catch
Trapping sitesKamunyu Kyusyani Iviani Ngiluni Nthunguni
0
01
02
03
04
05
06
Mea
n ap
pare
nt d
ensit
ies
Figure 6 Comparison of mean apparent densities within the fivetrapping sites
while Stomoxys spp constituted the lowest percentage andwere recorded only in Kyusyani area (Figure 5)
32 Apparent Densities (AD) There was no significant dif-ference in the apparent densities (AD) of Glossina speciescaptured across the five trapping sites which ranged from 01to 05 depending on the locality and averaged 03 flies per trapper day (Figure 6)
33 Identification of Trypanosomes in Glossina spp Theexpected band sizes of trypanosomes on amplification usingITS primers are shown in Table 2 Molecular identification oftrypanosomes T vivax and T congolense Forest in Glossinaspp was successful as their DNA amplicons matched theexpected sizes of 611 bp and 1513 bp respectively (Table 2)The amplified fragments for positive controls of T bruceibrucei T congolense Kilifi and T congolense Savannah werealso within the expected sizes (Table 2)
Figure 7 shows a sample of electrophoresis productof successfully amplified DNA fragments of trypanosomespecies T vivax (samples 1 and 5) and the positive controlsT brucei brucei (Tbb) T congolense Kilifi (Tck) and Tcongolense Savannah (Tcs) ldquoMrdquo represents 100 base pairmolecular marker (Promega) while 6 is the negative controlSamples 2 3 and 4 were negative for trypanosomes
34 Infection Rates Out of 78 flies analyzed using nestedPCR and gel electrophoresis the prevalence of trypanosomes
Journal of Parasitology Research 5
Table 2 Documented band sizes of trypanosome species [19]
Trypanosome species Expected band sizesin base pairs (bp)
T congolense forest 1513T congolense Kilifi 1422T congolense Savannah 1413T congolense Tsavo 954T brucei 1207ndash1224T vivax 611
15 kb1kb
500bp
minusVe M Tbb Tck Tcs 1 2 3 4 5 6
Figure 7 An image of an agarose gel showing PCR amplified DNAfragments using ITS nested PCR primers
infection in G pallidipes and G longipennis was 577 and2308 respectively (Table 3)The overall infection rate in allGlossina spp was recorded as 1153 There was a significant(119901 lt 005) difference in the trypanosomes infection rateamong Glossina species Infections with T vivax were themost prevalent (1026) in both species of tsetse comparedto T congolense with a prevalence of 128 (Table 3)
There was significant (119901 lt 005) difference in infectionrates among the trapping sites with Kamunyu recording thehighest rate of 641 followed by Kyusyani at 256 Ngiluniand Iviani recorded similar infection rates of 128 whileNthunguni had an infection rate of 0
A correlation analysis between apparent densities andinfection rates showed a strong positive relationship with acorrelation coefficient of 095 For instance Kamunyu arearecorded the highest apparent density as well as the highestinfection rates
There was a significant difference (119901 lt 005) in T vivaxand T congolense infections amongmale and female flies Forboth T vivax and T congolense the infections were higher inmales than in females with an overall prevalence of 1395and 857 respectively (Table 4)
4 Discussion
41 Vector Distribution in Mtito Andei This study found thatthere was variation in the distribution of G pallidipes andG longipennis in the area G longipennis was the only fuscagroup of flies trapped inMtitoAndei and is known to occur inother Eastern Africa countries such as Uganda Tanzania andEthiopia [2 24]This species is found in drier areasmore thanany other tsetse of fusca group because the pupariumcontainslarge water reserves and a low permeability of its pupalmembranes which helps it to survive harsh environmentalconditions [25 26] The typical habitats for this fly are dry
thorn bush riverline thickets near acacia woodlands and theresting sites are preferably shady sites of tree trunks logsand underside of branches of multistemmed trees [24] MtitoAndei provided all these ecological conditions hence suitablefor this fly
G pallidipes is a morsitans group of flies which is widelydistributed in East Afica and is a major vector of animaltrypanosomiasis [27] It was mostly found in areas close toMtitoAndei River andAthi River as it prefers dense evergreenvegetation heavier shade and humid habitats which areclose to riverline thickets [22] It is also present in otherEastern African countries such as Uganda Ethiopia andSomalia [2 28] Previous studies have shown thatG pallidipesmight be a vector of Trypanosoma b rhodesiense causingHuman African Trypanosomiasis (HAT) in Western Kenyaand Busoga District of Uganda as this parasite has beenisolated from the vector [24 27]
Among the mechanical vectors found in Mtito AndeiTabanids were highly predominant over Stomoxines in allthe trapping sites with the latter being recorded only inKyusyani area This finding corroborates the report detailedin [1] that the species composition of mechanical flies isstrongly dependent on site and substantial differences inenvironmental and climatic conditions of the sampled areaSimilar observations were also made in other areas suchas Brazil [29] The presence of mechanical flies in the areacannot be underestimated as they have been shown tomechanically transmit T vivax and T evansi [30] and canbe responsible for seasonal epidemic patterns in low tsetsedensity areas [1 31]
42 Apparent Densities of Glossina spp This study did notshow significant difference in the apparent densities (AD)of Glossina species captured across the five trapping sitesDespite the low mean AD recorded for tsetse in this area (03flies per trap per day) it is possible that tsetse serve both asbiological transmitters and reservoirs of the parasite while themechanical vectors amplify it during high density periodsOther than amplifying the parasite the mechanical flies canintroduce it to nonendemic areas through immigration
43 Trypanosome Infection Rates This study recorded lowoverall trypanosome infection rates of 1153 Similar resultswere obtained in the Kenyan Coast in a study that reportedtrypanosome infection rates of 1262 724 950 786and 797 in Ukunda Diani Muhaka Shimba hills andMwalewa respectively [32] A comparable study inMouhounRiver Burkina Faso reported overall infection rates of 10 in1423 dissected flies screened for trypanosome infections [33]Further infection rates for T congolense and T vivax wererecorded at 76 and 225 respectively Previous studieshave also showed low prevalence of trypanosome infections(about 10) in the field caught flies and associated this withinteractions of various suppressive factors in the generallysusceptible fly population [34] In addition there are a num-ber of barriers to both establishment and maturation processof trypanosomes and thus only a small proportion of theseinfections reach maturity [24] Further these trypanosomes
6 Journal of Parasitology Research
Table 3 Trypanosome infection rates in Glossina species
Glossina species Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
G pallidipes 49 2 (385) 1 (192) 3 (577) 52G longipennis 20 6 (2308) 0 (0) 6 (2308) 26Total 69 8 (1026) 1 (128) 9 (1153) 78
Table 4 Comparison of infection rates in male and female flies
Sex Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
Male 37 5 (1163) 1 (233) 6 (1395) 43Female 32 3 (857) 0 (0) 3 (857) 35Total 69 8 (1026) 1 (128) 9 (1153) 78
may lack differentiation trigger or may be inhibited by thevectorrsquos immune response [35 36]
Theoverall trypanosomes infection rates inG longipenniswere significantly higher than in G pallidipes However inCoastal Kenya higher infection rates were reported in Gpallidipes (57) than in G longipennis (02) [37] Thedifference in infection rates in the two tsetse species inMtito Andei could be due to variation in feeding preferencesenvironmental factors and host range differences [11 33]Thenumber of parasites available to infect tsetse parasite infec-tivity to tsetse and the strain or subspecies have also beenfound to affect infection rates in tsetse [38] Moreover thenutritional status of the tsetse at the time of infective bloodmeal can also affect their ability to acquire trypanosomeinfections [9] In addition low vectorial capacity has alsobeen reported and is attributed to higher levels of attacinexpression in the proventriculus and midgut [39 40] Thiskind of trait has been reported in G pallidipes and it couldexplain disparity of infection rates between G pallidipes andG longipennis [34] Within species infection rates were alsovariable and this was attributed to individual host factorsFor instance the vulnerability of flies to T brucei infectionswas shown to be due to maternally inherited features whichare associatedwith the presence of intracellular rickettsia-likeorganisms (RLOs) [41] Tsetse carrying these RLOs and othersimultaneous infections such as bacteria fungi and virus inthemidgutweremore likely to be infectedwith trypanosomesthan those without [41ndash43]
T vivax infection was more prevalent in all trappingsites than T congolense Similar results were reported in theLuangwa Valley Zambia [44] However in Cote drsquoIvoire Tcongolense was found to be more infectious (90) comparedto T simiae and T vivax in 50 of 139 microscopically positiveflies that were further analyzed using species specific primersfor T congolense subtypes T simiae and T vivax [45]The higher T vivax infections can be ascribed to shortdevelopmental lifecycle of about 10 days which is entirelycompleted within the proboscis For T congolense and Tbrucei development period has been estimated to be 14days and 30 days respectively [46] Further compared toproboscis tsetse midgut is a hostile environment which
contains midgut lectins which are able to kill trypanosomesin vivo [47] Midgut also has proteolytic digestive enzymespotent antimicrobial substances [34] prophenoloxidase cas-cade and immune molecules such as agglutinins whichhelp in expression of immune effector genes and mediatingcommunication between the gut and the fat body contentswhen microbes are encountered [48] The presence of allthese immune molecules can prevent establishment andmaturation of T congolense and T brucei as these parasitesuse midgut in part of their lifecycles hence the low infectionsrates are detected It is also known that T congolense subtypeparasites are generally low with low parasitaemia comparedto those of T vivax [38] It is therefore possible that suchscanty parasites are rarely picked up by the flies during bloodmeals which may account for the inability of the parasite toget established in the arthropod vector
This study revealed more infection rates in males thanin females In Nigeria higher infection rates in males thanfemales were also reported [49] Some studies suggest thatfemale flies should have higher infection rates than males asthey live longer thanmales and thus they have higher chancesof getting infection [50] However this relationship has notbeen established In contrast other studies explain that moremalesmay be infected than females as they are involved in sexactivities and competition than females [51] Whether moremales or females are infected in field tsetse populations thisstudy found no statististical difference with variation in sex
Apparent densities were strongly positively correlatedwith infection rates (119903 = 095) A similar observationwas reported in Mouhoun River basin Burkina Faso whereinfection rates positively correlated with apparent densities(119903 = 097) [1] This observation validates this parameter as anindicator of AAT transmission risk
5 Conclusion
This study attests that tsetse plays a major role in maintainingthe trypanosome parasite and concludes that sustainablereduction of tsetse from Mtito Andei Division can greatlyreduce the prevalence of AATThis study also showed that thehost factors are equally important in epidemiology of AAT
Journal of Parasitology Research 7
as higher trypanosome infection rates were recorded in Glongipennis than G pallidipes but the sex of the vector is lessimportant Further the study showed that apparent density ofthe vector is a good indicator of AAT transmission risk Thestudy therefore affirms the importance of designing controlstrategies targeting the biological vector (tsetse) mechanicalvectors (Tabanus and Stomoxys spp) and the trypanosomeparasites
6 Recommendations
(1) Since both T congolense and T vivax are pathogenicin cattle and other ruminants with the latter causingacute hemorrhagic syndrome [52] it would be impor-tant to conduct large scale integrated tsetse control inMtitoAndei by application of vector control strategiescoupled with curative treatment of livestock as theseanimals could act as source of reinfection to tsetse
(2) Surveillance studies to determine trypanosomeprevalence in livestock should be conducted in thisarea for effective trypanosomiasis control programs
(3) Entomological surveys should be conducted at dif-ferent seasons to understand seasonal dynamics ofthe vectors and the associated trypanosomiasis riskData on seasonal variations of potential vectors canbe integrated into epidemiologicalmodels to facilitatebetter understanding of the relative importance ofcyclical and mechanical vectors in Mtito Andei
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] N Kone E K NrsquoGoran I Sidibe A W Kombassere and JBouyer ldquoSpatio-temporal distribution of tsetse and other bitingflies in the Mouhoun River basin Burkina Fasordquo Medical andVeterinary Entomology vol 25 no 2 pp 156ndash168 2011
[2] G Cecchi M Paone R A Herrero M J B Vreysen and R CMattioli ldquoDeveloping a continental atlas of the distribution andtrypanosomal infection of tsetse flies (Glossina species)rdquo Para-sites amp Vectors vol 8 article 284 2015
[3] L J Morrison and A MacLeod ldquoAfrican trypanosomiasisrdquoParasite Immunology vol 33 no 8 pp 421ndash422 2011
[4] G Cecchi R C Mattioli J Slingenbergh and S De la RocqueldquoLand cover and tsetse fly distributions in sub-Saharan AfricardquoMedical and Veterinary Entomology vol 22 no 4 pp 364ndash3732008
[5] P Solano S J Torr and M J Lehane ldquoIs vector control neededto eliminate gambiense human African trypanosomiasisrdquoFrontiers in Cellular and Infection Microbiology vol 3 no 1article 33 2013
[6] A P M Shaw G Cecchi G RWWint R C Mattioli and T PRobinson ldquoMapping the economic benefits to livestock keepersfrom intervening against bovine trypanosomosis in EasternAfricardquo Preventive Veterinary Medicine vol 113 no 2 pp 197ndash210 2014
[7] J S Beadell C Hyseni P P Abila et al ldquoPhylogeography andpopulation structure of Glossina fuscipes fuscipes in Ugandaimplications for control of tsetserdquo PLoS Neglected TropicalDiseases vol 4 no 3 article e636 2010
[8] K Bardosh CWaiswa and S CWelburn ldquoConflict of interestuse of pyrethroids and amidines against tsetse and ticks inzoonotic sleeping sickness endemic areas of Ugandardquo Parasitesand Vectors vol 6 article 204 2013
[9] C Kubi J Van denAbbeele R DeDeken TMarcotty P Dornyand P Van Den Bossche ldquoThe effect of starvation on the sus-ceptibility of teneral and non-teneral tsetse flies to trypanosomeinfectionrdquoMedical andVeterinary Entomology vol 20 no 4 pp388ndash392 2006
[10] V Lutje J Seixas and A Kennedy ldquoChemotherapy for second-stage human African trypanosomiasisrdquo Cochrane Database ofSystematic Reviews vol 8 Article ID CD006201 2010
[11] M Bitew Y Amide T Zenebe and H Degefu ldquoTrypanosomeinfection rate inGlossina pallidipes andGlossina fuscipes fuscipesin Gojeb Valley Southwest Ethiopiardquo Global Veterinaria vol 6no 2 pp 131ndash135 2011
[12] R A Oluwafemi A A Ilemobade and E A O Laseinde ldquoTheimpact of African animal trypanosomiasis and tsetse on thelivelihood and well being of cattle and their owners in theBICOT study area ofNigeriardquo Scientific Research andEssays vol2 no 9 pp 380ndash383 2007
[13] R K Oniangrsquoo ldquoEnhancing peoplersquos nutritional status throughrevitalisation of Agriculture and related activitiesrdquo AfricanJournal of Food Agriculture Nutrition and Development vol 1no 1 pp 43ndash50 2001
[14] G L Mugunieri P Irungu and J M Omiti ldquoPerformance ofcommunity-based animal health workers in the delivery of live-stock health servicesrdquo Tropical Animal Health and Productionvol 36 no 6 pp 523ndash535 2004
[15] C A Spinage African EcologyThe Tsetse fly I Africarsquos Bane andBenefice Springer-Verlag Berlin Germany 2012
[16] P Solano J F Michel T Lefrancois et al ldquoPolymerase chainreaction as a diagnosis tool for detecting trypanosomes in nat-urally infected cattle in Burkina Fasordquo Veterinary Parasitologyvol 86 no 2 pp 95ndash103 1999
[17] T Tran G Napier T Rowan et al ldquoDevelopment and eval-uation of an ITS1 lsquoTouchdownrsquo PCR for assessment of drugefficacy against animal African trypanosomosisrdquo VeterinaryParasitology vol 202 no 3-4 pp 164ndash170 2014
[18] S M Thumbi F A McOdimba R O Mosi and J O JungrsquoaldquoComparative evaluation of three PCR base diagnostic assaysfor the detection of pathogenic trypanosomes in cattle bloodrdquoParasites and Vectors vol 1 article 46 2008
[19] A Cox A Tilley F McOdimba et al ldquoA PCR based assay fordetection and differentiation of African trypanosome species inbloodrdquoExperimental Parasitology vol 111 no 1 pp 24ndash29 2005
[20] L Brownhill E M Njuguna and M Miruka Strategy forMainstreaming Gender Analysis in the KARIMcGill UniversityProject Enhancing Ecologically Resilient Food Security ThroughInnovative Farming Systems in the Semi AridMidlands of KenyaKenya Agricultural Research Institute (KARI) McGill Univer-sity 3rd edition 2013
[21] M Thrusfield Veterinary Epidemiology Blackwell ScienceOxford UK 3rd edition 2005
[22] I Malele L Craske C Knight et al ldquoThe use of specific andgeneric primers to identify trypanosome infections of wildtsetse flies in Tanzania by PCRrdquo Infection Genetics and Evolu-tion vol 3 no 4 pp 271ndash279 2003
8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 Journal of Parasitology Research
controlled [11] Out of 165 million cattle found in Africa only10 million are found within the tsetse belt due to the diseaseconstraint and these are the lowest producing breeds [12]TheKenyan economy is largely agricultural-based with the sectoraccounting for about 24 of the Gross Domestic Product(GDP) and 70 of the employment [13] Only 7 of Kenyanland is suitable for crop production and a further 5 cansustain crops in years of adequate rains [13] The rest of landis arid and semiarid and constitutes the range lands which arenot suitable for crop production but rather good for livestockproduction [13 14]
Mtito Andei is located in Makueni County about 290 kmSouth East of Nairobi The area is comprised mainly of aridand semiarid lands and livestock rearing is the mainstay ofthis region [14] This area has a history of animal trypanoso-miasis as it is infested with G pallidipes G longipennisand G brevipalpis [14 15] The area was not inhabiteduntil the 1930s due to its low agricultural potential andheavy tsetse infestation Assessment of prevalence vectorhotspots and infection status can be instrumental in makingdecisions with regard to the formulations of suitable tsetseand trypanosomiasis control measures However there isno accurate information that exists in regard to the currentprevalence of trypanosomiasis inMtitoAndeiThis is becauseprevious data has been collected through microscopy onblood smears dissection and microhematocrit techniqueswhich are less sensitive [16 17] Further these methodscannot identify immature andmixed infections subsequentlyresulting to low detection levels [18] This study was aimedat providing additional information on the prevalence oftrypanosomes in Glossina species in Mtito Andei
2 Materials and Method
21 Description of the Study Site This study was carried outin Mtito Andei Division situated in Makueni County about290 kmSouth East ofNairobiThe study areawas divided intofive villages namely Nthunguni Iviani Kyusyani Ngiluniand Kamunyu (Figure 1) The altitude of the area varies from600m to 1100m above sea level The area is characterizedby marginal agricultural lands of Savannah grassland withmostly low lying gently eastward sloping plains occasionallybroken by seasonal and perennial rivers [20] The climate ofthis area is typically semiarid with low and unreliable rainfallaveraging 600mm annually and annual average temperatureof 23∘C [20] Rainfall is bimodal with long rains occurringfrom March to May and short rains from November orDecember to early January The highest mean temperaturesbetween 32∘C and 33∘C prevail in February to March [20]
22 Study Design A cross-sectional study using simplerandom sampling technique was conducted to determinethe infection rates of trypanosomes in Glossina species inMtito Andei Division Sample size was determined using theformula (119899 = 119885211987511987621198892) described in [21] where 119899 is theminimum sample size required 119885 is the confidence interval119875 is the expected prevalence in proportion from the study 119876is 100 minus 119875 and 119889 is the desired precision taken at 005
Ethiopia
Kenya Somalia
Mtito AndeiTanzania100 km
N
Kamunyu
Iviani
Nthunguni
KyusyaniNgiluni
0 10
(km)
Figure 1 Map of Mtito Andei showing the georeferenced trappingsites
Figure 2 A biconical trap baited with cow urine and acetone toenhance trapping efficiency
23 Collection and Identification of Tsetse Tsetse trappingwas done between April and May 2012 This period was cho-sen because it is a rainy season and therefore more abundantflies were expected [1] Trapping was done using 5 biconicaltraps in each village which were deployed near Mtito AndeiRiver and Athi River and in the nearby vegetation The trapswere set at georeferenced sites and baited with acetone andcow urine to increase trapping efficiency [22] The trapswere set at regular intervals of 100 meters along variousvegetation types and under shade to avoid undue flymortalitydue to heat (Figure 2) To prevent the ascent of ants on thepoles towards the collecting cages each pole was smearedwith grease All trapped flies were counted their sex was
Journal of Parasitology Research 3
determined and identified to species level using conventionalidentification keys before dissection [23]
24 Determination of Apparent Density (AD) Apparent den-sity (AD) is an estimation of fliesrsquo density and is given in termsof the number of flies caught per trap per day (119865119879119863) It wascalculated using the following formula
119865119879119863 =sum119865
119879 times 119863 (1)
where sum119865 is the number of total flies 119879 is the number offunctioning traps and119863 is the number of days forwhich trapswere operational [1]
The average AD for each trapping site was then calculatedto obtain the data on tsetse distribution in the area
25 Identification of Trypanosomes in Glossina spp
251 Tsetse Dissection The flies were immobilized usingethyl acetate placed in a cotton wool at the bottom ofa specimen tube Dissection was carried out on freshlyimmobilized flies as trypanosomes are less likely to be foundin dead dried flies [23] A total of 78 nonteneral flies weredissected from which midgut salivary glands and probosciswere removed and analyzed under a microscope for thepresence of trypanosomes Nonteneral flies were identified bythe presence of dark or brown colour on their abdomenwhichindicated the last blood meal [23] Freshly immobilized flieswere surface sterilized by brief immersion in 70 ethanolthen dry blotted on clean tissue paper and then dissectedin normal saline [22] To avoid cross contamination thedissecting instruments were sterilized by immersion in 3ndash5 (wv) sodium hypochlorite for approximately 2 minutesfollowed by extensive rinsing in distilled water and later finalimmersion in normal saline [22] Dissected organs (midgutproboscis and salivary glands) from each fly were transferredinto 15mL microcentrifuge tube containing 120120583L of tissuelysis buffer (from Qiagen DNeasy Blood and Tissue Kit) Thetissues were then stored at minus20∘C until further processing
252 Trypanosomesrsquo DNA Extraction DNA was extractedusing the Qiagen DNeasy blood and tissue kit and accordingto the manufacturerrsquos instructions Twenty (20) microliters(120583L) of proteinase K was added to the dissected tissues andincubated at 56∘C overnight in a thermocycler until the tissuewas completely digested The lysate was vortexed for 15 secand 200120583L of buffer AL was added to the sample followedby vortexing to mix the contents Two hundred (200) 120583L ofabsolute ethanol was added andmixed thoroughly by furthervortexing before pipetting themixture intoDNeasymini spincolumns placed in a 2mL collection tube The columns werecentrifuged at 8000 rpm for 1 minute and the flow throughdiscarded together with the collection tube The DNeasymini spin columns were placed in a new 2mL collectiontube and 500 120583L of wash buffer AW1 was added It was thencentrifuged at 8000 rpm for 1 minute and the flow throughtogether with the collection tube discardedThe spin columnwas again placed in a new collection tube and 500120583L of
Table 1 Sequence of primers
Primer ID Primer sequence (51015840-31015840)Outerprimers
ITS 1 GAT TAC GTC CCT GCC ATT TGITS 2 TTG TTC GCT ATC GGT CTT CC
Innerprimers
ITS 3 GGA AGC AAA AGT CGT AAC AAG GITS 4 TGT TTT CTT TTC CTC CGC
SSU18S ITS 1 58S ITS 2 LSU(1)
28S
Figure 3 Schematic diagram of rDNA showing ITS 1 and ITS 2annealing positions [19]
wash buffer AW2 was added The column was centrifuged at14000 rpm for 3minutesThe flow through and the collectiontubes were discarded and the spin columns placed in a clean15mL microcentrifuge tube followed by addition of 30 120583Lof elution buffer (AE buffer) The column was incubated atroom temperature for 1 minute and centrifuged for 1 minuteat 8000 rpm to elute the bound DNA DNA extracted fromsamples was quantified using a nanodrop spectrophotometer(Thermo Fisher Scientific Inc) and its purity was determinedby measuring the intensity of absorbance at wavelengths 260and 280 nanometers
253 PCR Amplification and DNA Analysis The PCRoligonucleotide primers used are shown in Table 1 Theprimers were synthesized by Bioneer Company andwere pro-vided by International Livestock Research Institute (ILRI)Kenya
These primers were designed to amplify the internaltranscribed (ITS) region of ribosomal DNA (rDNA) a regionwhich varies in size within trypanosome species except formembers of Trypanozoon and are capable of differentiatingtrypanosomes based on their amplified fragments ITS 1 andITS 2 (outer primers) amplify the conserved regions of rDNAsmall subunit (SSU) and large subunit (LSU) representedby black arrows in Figure 3 while ITS 3 and ITS 4 (innerprimers) amplify the noncoding spacer regions of rDNA andare represented by white arrows
All PCR amplifications were performed in 30 120583L reactionvolumes containing final concentrations of 1x InvitrogenPCR buffer 35mM MgCl
2(Promega) 02mM dNTPs mix
125U120583L Invitrogen Taq 100 ng120583L of each forward andreverse primer and 5 120583L of trypanosome DNA as templateA set of purified genomic DNA of T brucei brucei (Tbb) Tcongolense Kilifi (Tck) and T congolense Savannah (Tcs)(strain IL1180) were included as positive controls during allPCR assays and nuclease free water as negative control Theouter primers (ITS 1 and 2) were used in the first roundof reaction The cycling conditions were as follows 1 cycleof 95∘C for 7 minutes as initial denaturation followed by 35cycles of 94∘C for 1 minute 55∘C for 1 minute and 72∘C for 2minutes The final extension was done at 72∘C for 5 minutes
4 Journal of Parasitology ResearchEa
ch sp
ecie
s (
)
Kamunyu Ngiluni Kyusyani Iviani NthunguniTrapping sites
G pallidipesG longipennis
0
20
40
60
80
100
120
Figure 4 Distribution of Glossina species captured from study sitesin Mtito Andei Division
The PCR was carried out on a Bio-Rad thermocycler In thesecond round of reaction the outer primers were substitutedwith inner primers (ITS 3 and 4) and 5 120583L of round one PCRproduct was used as DNA template The cycling conditionsfor the second round of reaction were as described for theprimary amplification To analyze the amplicons 10120583L ofthe PCR product was resolved in a 2 agarose gel at 80volts for 45 minutes and the gel was visualised under UVtransilluminator following ethidium bromide staining
26 Data Analysis The infectious trypanosomes were iden-tified by comparing the molecular sizes of their DNA frag-ments with the documented band sizes of trypanosomespecies [19]The data was then used to calculate the infectionrates and prevalence of the trypanosomes on Glossina sppConventional data on vector distribution apparent densitiesand infection rates from the five sites was subjected to one-way analysis of variance (ANOVA) using SPSS version 160software at 95 confidence interval Chi-square test was doneto determine the association of infection rates with flyrsquos sexand vector species A correlation analysis was also done toestablishwhether there was a significant relationship betweeninfection rates and apparent densities
3 Results
31 Vector Distribution Out of the 117 flies trapped in thearea G pallidipes constituted the largest percentage of 58whileG longipennis had 42 Among the trapped tsetse flies39 (333) were teneral while 78 (67) were nonteneral InKamunyu area G longipennis was the most prevalent specieswhile G pallidipes dominated Ngiluni Kyusyani Iviani andNthunguni (Figure 4)
Apart from Glossina spp the area was also inhabitedwith mechanical flies such as Tabanus spp and Stomoxys sppTabanids were highly predominant over Stomoxines in all thetrapping sites with an overall fly trap density (FTD) of 011while Stomoxys spp had FTD of 002 Of the total fly catchGlossina spp were the most prevalent in all the tapping sites
FliesGlossina spp Tabanus spp Stomoxys spp
0
10
20
30
40
50
60
70
80
90
100
()
Figure 5 Relative distribution of total fly catch
Trapping sitesKamunyu Kyusyani Iviani Ngiluni Nthunguni
0
01
02
03
04
05
06
Mea
n ap
pare
nt d
ensit
ies
Figure 6 Comparison of mean apparent densities within the fivetrapping sites
while Stomoxys spp constituted the lowest percentage andwere recorded only in Kyusyani area (Figure 5)
32 Apparent Densities (AD) There was no significant dif-ference in the apparent densities (AD) of Glossina speciescaptured across the five trapping sites which ranged from 01to 05 depending on the locality and averaged 03 flies per trapper day (Figure 6)
33 Identification of Trypanosomes in Glossina spp Theexpected band sizes of trypanosomes on amplification usingITS primers are shown in Table 2 Molecular identification oftrypanosomes T vivax and T congolense Forest in Glossinaspp was successful as their DNA amplicons matched theexpected sizes of 611 bp and 1513 bp respectively (Table 2)The amplified fragments for positive controls of T bruceibrucei T congolense Kilifi and T congolense Savannah werealso within the expected sizes (Table 2)
Figure 7 shows a sample of electrophoresis productof successfully amplified DNA fragments of trypanosomespecies T vivax (samples 1 and 5) and the positive controlsT brucei brucei (Tbb) T congolense Kilifi (Tck) and Tcongolense Savannah (Tcs) ldquoMrdquo represents 100 base pairmolecular marker (Promega) while 6 is the negative controlSamples 2 3 and 4 were negative for trypanosomes
34 Infection Rates Out of 78 flies analyzed using nestedPCR and gel electrophoresis the prevalence of trypanosomes
Journal of Parasitology Research 5
Table 2 Documented band sizes of trypanosome species [19]
Trypanosome species Expected band sizesin base pairs (bp)
T congolense forest 1513T congolense Kilifi 1422T congolense Savannah 1413T congolense Tsavo 954T brucei 1207ndash1224T vivax 611
15 kb1kb
500bp
minusVe M Tbb Tck Tcs 1 2 3 4 5 6
Figure 7 An image of an agarose gel showing PCR amplified DNAfragments using ITS nested PCR primers
infection in G pallidipes and G longipennis was 577 and2308 respectively (Table 3)The overall infection rate in allGlossina spp was recorded as 1153 There was a significant(119901 lt 005) difference in the trypanosomes infection rateamong Glossina species Infections with T vivax were themost prevalent (1026) in both species of tsetse comparedto T congolense with a prevalence of 128 (Table 3)
There was significant (119901 lt 005) difference in infectionrates among the trapping sites with Kamunyu recording thehighest rate of 641 followed by Kyusyani at 256 Ngiluniand Iviani recorded similar infection rates of 128 whileNthunguni had an infection rate of 0
A correlation analysis between apparent densities andinfection rates showed a strong positive relationship with acorrelation coefficient of 095 For instance Kamunyu arearecorded the highest apparent density as well as the highestinfection rates
There was a significant difference (119901 lt 005) in T vivaxand T congolense infections amongmale and female flies Forboth T vivax and T congolense the infections were higher inmales than in females with an overall prevalence of 1395and 857 respectively (Table 4)
4 Discussion
41 Vector Distribution in Mtito Andei This study found thatthere was variation in the distribution of G pallidipes andG longipennis in the area G longipennis was the only fuscagroup of flies trapped inMtitoAndei and is known to occur inother Eastern Africa countries such as Uganda Tanzania andEthiopia [2 24]This species is found in drier areasmore thanany other tsetse of fusca group because the pupariumcontainslarge water reserves and a low permeability of its pupalmembranes which helps it to survive harsh environmentalconditions [25 26] The typical habitats for this fly are dry
thorn bush riverline thickets near acacia woodlands and theresting sites are preferably shady sites of tree trunks logsand underside of branches of multistemmed trees [24] MtitoAndei provided all these ecological conditions hence suitablefor this fly
G pallidipes is a morsitans group of flies which is widelydistributed in East Afica and is a major vector of animaltrypanosomiasis [27] It was mostly found in areas close toMtitoAndei River andAthi River as it prefers dense evergreenvegetation heavier shade and humid habitats which areclose to riverline thickets [22] It is also present in otherEastern African countries such as Uganda Ethiopia andSomalia [2 28] Previous studies have shown thatG pallidipesmight be a vector of Trypanosoma b rhodesiense causingHuman African Trypanosomiasis (HAT) in Western Kenyaand Busoga District of Uganda as this parasite has beenisolated from the vector [24 27]
Among the mechanical vectors found in Mtito AndeiTabanids were highly predominant over Stomoxines in allthe trapping sites with the latter being recorded only inKyusyani area This finding corroborates the report detailedin [1] that the species composition of mechanical flies isstrongly dependent on site and substantial differences inenvironmental and climatic conditions of the sampled areaSimilar observations were also made in other areas suchas Brazil [29] The presence of mechanical flies in the areacannot be underestimated as they have been shown tomechanically transmit T vivax and T evansi [30] and canbe responsible for seasonal epidemic patterns in low tsetsedensity areas [1 31]
42 Apparent Densities of Glossina spp This study did notshow significant difference in the apparent densities (AD)of Glossina species captured across the five trapping sitesDespite the low mean AD recorded for tsetse in this area (03flies per trap per day) it is possible that tsetse serve both asbiological transmitters and reservoirs of the parasite while themechanical vectors amplify it during high density periodsOther than amplifying the parasite the mechanical flies canintroduce it to nonendemic areas through immigration
43 Trypanosome Infection Rates This study recorded lowoverall trypanosome infection rates of 1153 Similar resultswere obtained in the Kenyan Coast in a study that reportedtrypanosome infection rates of 1262 724 950 786and 797 in Ukunda Diani Muhaka Shimba hills andMwalewa respectively [32] A comparable study inMouhounRiver Burkina Faso reported overall infection rates of 10 in1423 dissected flies screened for trypanosome infections [33]Further infection rates for T congolense and T vivax wererecorded at 76 and 225 respectively Previous studieshave also showed low prevalence of trypanosome infections(about 10) in the field caught flies and associated this withinteractions of various suppressive factors in the generallysusceptible fly population [34] In addition there are a num-ber of barriers to both establishment and maturation processof trypanosomes and thus only a small proportion of theseinfections reach maturity [24] Further these trypanosomes
6 Journal of Parasitology Research
Table 3 Trypanosome infection rates in Glossina species
Glossina species Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
G pallidipes 49 2 (385) 1 (192) 3 (577) 52G longipennis 20 6 (2308) 0 (0) 6 (2308) 26Total 69 8 (1026) 1 (128) 9 (1153) 78
Table 4 Comparison of infection rates in male and female flies
Sex Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
Male 37 5 (1163) 1 (233) 6 (1395) 43Female 32 3 (857) 0 (0) 3 (857) 35Total 69 8 (1026) 1 (128) 9 (1153) 78
may lack differentiation trigger or may be inhibited by thevectorrsquos immune response [35 36]
Theoverall trypanosomes infection rates inG longipenniswere significantly higher than in G pallidipes However inCoastal Kenya higher infection rates were reported in Gpallidipes (57) than in G longipennis (02) [37] Thedifference in infection rates in the two tsetse species inMtito Andei could be due to variation in feeding preferencesenvironmental factors and host range differences [11 33]Thenumber of parasites available to infect tsetse parasite infec-tivity to tsetse and the strain or subspecies have also beenfound to affect infection rates in tsetse [38] Moreover thenutritional status of the tsetse at the time of infective bloodmeal can also affect their ability to acquire trypanosomeinfections [9] In addition low vectorial capacity has alsobeen reported and is attributed to higher levels of attacinexpression in the proventriculus and midgut [39 40] Thiskind of trait has been reported in G pallidipes and it couldexplain disparity of infection rates between G pallidipes andG longipennis [34] Within species infection rates were alsovariable and this was attributed to individual host factorsFor instance the vulnerability of flies to T brucei infectionswas shown to be due to maternally inherited features whichare associatedwith the presence of intracellular rickettsia-likeorganisms (RLOs) [41] Tsetse carrying these RLOs and othersimultaneous infections such as bacteria fungi and virus inthemidgutweremore likely to be infectedwith trypanosomesthan those without [41ndash43]
T vivax infection was more prevalent in all trappingsites than T congolense Similar results were reported in theLuangwa Valley Zambia [44] However in Cote drsquoIvoire Tcongolense was found to be more infectious (90) comparedto T simiae and T vivax in 50 of 139 microscopically positiveflies that were further analyzed using species specific primersfor T congolense subtypes T simiae and T vivax [45]The higher T vivax infections can be ascribed to shortdevelopmental lifecycle of about 10 days which is entirelycompleted within the proboscis For T congolense and Tbrucei development period has been estimated to be 14days and 30 days respectively [46] Further compared toproboscis tsetse midgut is a hostile environment which
contains midgut lectins which are able to kill trypanosomesin vivo [47] Midgut also has proteolytic digestive enzymespotent antimicrobial substances [34] prophenoloxidase cas-cade and immune molecules such as agglutinins whichhelp in expression of immune effector genes and mediatingcommunication between the gut and the fat body contentswhen microbes are encountered [48] The presence of allthese immune molecules can prevent establishment andmaturation of T congolense and T brucei as these parasitesuse midgut in part of their lifecycles hence the low infectionsrates are detected It is also known that T congolense subtypeparasites are generally low with low parasitaemia comparedto those of T vivax [38] It is therefore possible that suchscanty parasites are rarely picked up by the flies during bloodmeals which may account for the inability of the parasite toget established in the arthropod vector
This study revealed more infection rates in males thanin females In Nigeria higher infection rates in males thanfemales were also reported [49] Some studies suggest thatfemale flies should have higher infection rates than males asthey live longer thanmales and thus they have higher chancesof getting infection [50] However this relationship has notbeen established In contrast other studies explain that moremalesmay be infected than females as they are involved in sexactivities and competition than females [51] Whether moremales or females are infected in field tsetse populations thisstudy found no statististical difference with variation in sex
Apparent densities were strongly positively correlatedwith infection rates (119903 = 095) A similar observationwas reported in Mouhoun River basin Burkina Faso whereinfection rates positively correlated with apparent densities(119903 = 097) [1] This observation validates this parameter as anindicator of AAT transmission risk
5 Conclusion
This study attests that tsetse plays a major role in maintainingthe trypanosome parasite and concludes that sustainablereduction of tsetse from Mtito Andei Division can greatlyreduce the prevalence of AATThis study also showed that thehost factors are equally important in epidemiology of AAT
Journal of Parasitology Research 7
as higher trypanosome infection rates were recorded in Glongipennis than G pallidipes but the sex of the vector is lessimportant Further the study showed that apparent density ofthe vector is a good indicator of AAT transmission risk Thestudy therefore affirms the importance of designing controlstrategies targeting the biological vector (tsetse) mechanicalvectors (Tabanus and Stomoxys spp) and the trypanosomeparasites
6 Recommendations
(1) Since both T congolense and T vivax are pathogenicin cattle and other ruminants with the latter causingacute hemorrhagic syndrome [52] it would be impor-tant to conduct large scale integrated tsetse control inMtitoAndei by application of vector control strategiescoupled with curative treatment of livestock as theseanimals could act as source of reinfection to tsetse
(2) Surveillance studies to determine trypanosomeprevalence in livestock should be conducted in thisarea for effective trypanosomiasis control programs
(3) Entomological surveys should be conducted at dif-ferent seasons to understand seasonal dynamics ofthe vectors and the associated trypanosomiasis riskData on seasonal variations of potential vectors canbe integrated into epidemiologicalmodels to facilitatebetter understanding of the relative importance ofcyclical and mechanical vectors in Mtito Andei
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] N Kone E K NrsquoGoran I Sidibe A W Kombassere and JBouyer ldquoSpatio-temporal distribution of tsetse and other bitingflies in the Mouhoun River basin Burkina Fasordquo Medical andVeterinary Entomology vol 25 no 2 pp 156ndash168 2011
[2] G Cecchi M Paone R A Herrero M J B Vreysen and R CMattioli ldquoDeveloping a continental atlas of the distribution andtrypanosomal infection of tsetse flies (Glossina species)rdquo Para-sites amp Vectors vol 8 article 284 2015
[3] L J Morrison and A MacLeod ldquoAfrican trypanosomiasisrdquoParasite Immunology vol 33 no 8 pp 421ndash422 2011
[4] G Cecchi R C Mattioli J Slingenbergh and S De la RocqueldquoLand cover and tsetse fly distributions in sub-Saharan AfricardquoMedical and Veterinary Entomology vol 22 no 4 pp 364ndash3732008
[5] P Solano S J Torr and M J Lehane ldquoIs vector control neededto eliminate gambiense human African trypanosomiasisrdquoFrontiers in Cellular and Infection Microbiology vol 3 no 1article 33 2013
[6] A P M Shaw G Cecchi G RWWint R C Mattioli and T PRobinson ldquoMapping the economic benefits to livestock keepersfrom intervening against bovine trypanosomosis in EasternAfricardquo Preventive Veterinary Medicine vol 113 no 2 pp 197ndash210 2014
[7] J S Beadell C Hyseni P P Abila et al ldquoPhylogeography andpopulation structure of Glossina fuscipes fuscipes in Ugandaimplications for control of tsetserdquo PLoS Neglected TropicalDiseases vol 4 no 3 article e636 2010
[8] K Bardosh CWaiswa and S CWelburn ldquoConflict of interestuse of pyrethroids and amidines against tsetse and ticks inzoonotic sleeping sickness endemic areas of Ugandardquo Parasitesand Vectors vol 6 article 204 2013
[9] C Kubi J Van denAbbeele R DeDeken TMarcotty P Dornyand P Van Den Bossche ldquoThe effect of starvation on the sus-ceptibility of teneral and non-teneral tsetse flies to trypanosomeinfectionrdquoMedical andVeterinary Entomology vol 20 no 4 pp388ndash392 2006
[10] V Lutje J Seixas and A Kennedy ldquoChemotherapy for second-stage human African trypanosomiasisrdquo Cochrane Database ofSystematic Reviews vol 8 Article ID CD006201 2010
[11] M Bitew Y Amide T Zenebe and H Degefu ldquoTrypanosomeinfection rate inGlossina pallidipes andGlossina fuscipes fuscipesin Gojeb Valley Southwest Ethiopiardquo Global Veterinaria vol 6no 2 pp 131ndash135 2011
[12] R A Oluwafemi A A Ilemobade and E A O Laseinde ldquoTheimpact of African animal trypanosomiasis and tsetse on thelivelihood and well being of cattle and their owners in theBICOT study area ofNigeriardquo Scientific Research andEssays vol2 no 9 pp 380ndash383 2007
[13] R K Oniangrsquoo ldquoEnhancing peoplersquos nutritional status throughrevitalisation of Agriculture and related activitiesrdquo AfricanJournal of Food Agriculture Nutrition and Development vol 1no 1 pp 43ndash50 2001
[14] G L Mugunieri P Irungu and J M Omiti ldquoPerformance ofcommunity-based animal health workers in the delivery of live-stock health servicesrdquo Tropical Animal Health and Productionvol 36 no 6 pp 523ndash535 2004
[15] C A Spinage African EcologyThe Tsetse fly I Africarsquos Bane andBenefice Springer-Verlag Berlin Germany 2012
[16] P Solano J F Michel T Lefrancois et al ldquoPolymerase chainreaction as a diagnosis tool for detecting trypanosomes in nat-urally infected cattle in Burkina Fasordquo Veterinary Parasitologyvol 86 no 2 pp 95ndash103 1999
[17] T Tran G Napier T Rowan et al ldquoDevelopment and eval-uation of an ITS1 lsquoTouchdownrsquo PCR for assessment of drugefficacy against animal African trypanosomosisrdquo VeterinaryParasitology vol 202 no 3-4 pp 164ndash170 2014
[18] S M Thumbi F A McOdimba R O Mosi and J O JungrsquoaldquoComparative evaluation of three PCR base diagnostic assaysfor the detection of pathogenic trypanosomes in cattle bloodrdquoParasites and Vectors vol 1 article 46 2008
[19] A Cox A Tilley F McOdimba et al ldquoA PCR based assay fordetection and differentiation of African trypanosome species inbloodrdquoExperimental Parasitology vol 111 no 1 pp 24ndash29 2005
[20] L Brownhill E M Njuguna and M Miruka Strategy forMainstreaming Gender Analysis in the KARIMcGill UniversityProject Enhancing Ecologically Resilient Food Security ThroughInnovative Farming Systems in the Semi AridMidlands of KenyaKenya Agricultural Research Institute (KARI) McGill Univer-sity 3rd edition 2013
[21] M Thrusfield Veterinary Epidemiology Blackwell ScienceOxford UK 3rd edition 2005
[22] I Malele L Craske C Knight et al ldquoThe use of specific andgeneric primers to identify trypanosome infections of wildtsetse flies in Tanzania by PCRrdquo Infection Genetics and Evolu-tion vol 3 no 4 pp 271ndash279 2003
8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Journal of Parasitology Research 3
determined and identified to species level using conventionalidentification keys before dissection [23]
24 Determination of Apparent Density (AD) Apparent den-sity (AD) is an estimation of fliesrsquo density and is given in termsof the number of flies caught per trap per day (119865119879119863) It wascalculated using the following formula
119865119879119863 =sum119865
119879 times 119863 (1)
where sum119865 is the number of total flies 119879 is the number offunctioning traps and119863 is the number of days forwhich trapswere operational [1]
The average AD for each trapping site was then calculatedto obtain the data on tsetse distribution in the area
25 Identification of Trypanosomes in Glossina spp
251 Tsetse Dissection The flies were immobilized usingethyl acetate placed in a cotton wool at the bottom ofa specimen tube Dissection was carried out on freshlyimmobilized flies as trypanosomes are less likely to be foundin dead dried flies [23] A total of 78 nonteneral flies weredissected from which midgut salivary glands and probosciswere removed and analyzed under a microscope for thepresence of trypanosomes Nonteneral flies were identified bythe presence of dark or brown colour on their abdomenwhichindicated the last blood meal [23] Freshly immobilized flieswere surface sterilized by brief immersion in 70 ethanolthen dry blotted on clean tissue paper and then dissectedin normal saline [22] To avoid cross contamination thedissecting instruments were sterilized by immersion in 3ndash5 (wv) sodium hypochlorite for approximately 2 minutesfollowed by extensive rinsing in distilled water and later finalimmersion in normal saline [22] Dissected organs (midgutproboscis and salivary glands) from each fly were transferredinto 15mL microcentrifuge tube containing 120120583L of tissuelysis buffer (from Qiagen DNeasy Blood and Tissue Kit) Thetissues were then stored at minus20∘C until further processing
252 Trypanosomesrsquo DNA Extraction DNA was extractedusing the Qiagen DNeasy blood and tissue kit and accordingto the manufacturerrsquos instructions Twenty (20) microliters(120583L) of proteinase K was added to the dissected tissues andincubated at 56∘C overnight in a thermocycler until the tissuewas completely digested The lysate was vortexed for 15 secand 200120583L of buffer AL was added to the sample followedby vortexing to mix the contents Two hundred (200) 120583L ofabsolute ethanol was added andmixed thoroughly by furthervortexing before pipetting themixture intoDNeasymini spincolumns placed in a 2mL collection tube The columns werecentrifuged at 8000 rpm for 1 minute and the flow throughdiscarded together with the collection tube The DNeasymini spin columns were placed in a new 2mL collectiontube and 500 120583L of wash buffer AW1 was added It was thencentrifuged at 8000 rpm for 1 minute and the flow throughtogether with the collection tube discardedThe spin columnwas again placed in a new collection tube and 500120583L of
Table 1 Sequence of primers
Primer ID Primer sequence (51015840-31015840)Outerprimers
ITS 1 GAT TAC GTC CCT GCC ATT TGITS 2 TTG TTC GCT ATC GGT CTT CC
Innerprimers
ITS 3 GGA AGC AAA AGT CGT AAC AAG GITS 4 TGT TTT CTT TTC CTC CGC
SSU18S ITS 1 58S ITS 2 LSU(1)
28S
Figure 3 Schematic diagram of rDNA showing ITS 1 and ITS 2annealing positions [19]
wash buffer AW2 was added The column was centrifuged at14000 rpm for 3minutesThe flow through and the collectiontubes were discarded and the spin columns placed in a clean15mL microcentrifuge tube followed by addition of 30 120583Lof elution buffer (AE buffer) The column was incubated atroom temperature for 1 minute and centrifuged for 1 minuteat 8000 rpm to elute the bound DNA DNA extracted fromsamples was quantified using a nanodrop spectrophotometer(Thermo Fisher Scientific Inc) and its purity was determinedby measuring the intensity of absorbance at wavelengths 260and 280 nanometers
253 PCR Amplification and DNA Analysis The PCRoligonucleotide primers used are shown in Table 1 Theprimers were synthesized by Bioneer Company andwere pro-vided by International Livestock Research Institute (ILRI)Kenya
These primers were designed to amplify the internaltranscribed (ITS) region of ribosomal DNA (rDNA) a regionwhich varies in size within trypanosome species except formembers of Trypanozoon and are capable of differentiatingtrypanosomes based on their amplified fragments ITS 1 andITS 2 (outer primers) amplify the conserved regions of rDNAsmall subunit (SSU) and large subunit (LSU) representedby black arrows in Figure 3 while ITS 3 and ITS 4 (innerprimers) amplify the noncoding spacer regions of rDNA andare represented by white arrows
All PCR amplifications were performed in 30 120583L reactionvolumes containing final concentrations of 1x InvitrogenPCR buffer 35mM MgCl
2(Promega) 02mM dNTPs mix
125U120583L Invitrogen Taq 100 ng120583L of each forward andreverse primer and 5 120583L of trypanosome DNA as templateA set of purified genomic DNA of T brucei brucei (Tbb) Tcongolense Kilifi (Tck) and T congolense Savannah (Tcs)(strain IL1180) were included as positive controls during allPCR assays and nuclease free water as negative control Theouter primers (ITS 1 and 2) were used in the first roundof reaction The cycling conditions were as follows 1 cycleof 95∘C for 7 minutes as initial denaturation followed by 35cycles of 94∘C for 1 minute 55∘C for 1 minute and 72∘C for 2minutes The final extension was done at 72∘C for 5 minutes
4 Journal of Parasitology ResearchEa
ch sp
ecie
s (
)
Kamunyu Ngiluni Kyusyani Iviani NthunguniTrapping sites
G pallidipesG longipennis
0
20
40
60
80
100
120
Figure 4 Distribution of Glossina species captured from study sitesin Mtito Andei Division
The PCR was carried out on a Bio-Rad thermocycler In thesecond round of reaction the outer primers were substitutedwith inner primers (ITS 3 and 4) and 5 120583L of round one PCRproduct was used as DNA template The cycling conditionsfor the second round of reaction were as described for theprimary amplification To analyze the amplicons 10120583L ofthe PCR product was resolved in a 2 agarose gel at 80volts for 45 minutes and the gel was visualised under UVtransilluminator following ethidium bromide staining
26 Data Analysis The infectious trypanosomes were iden-tified by comparing the molecular sizes of their DNA frag-ments with the documented band sizes of trypanosomespecies [19]The data was then used to calculate the infectionrates and prevalence of the trypanosomes on Glossina sppConventional data on vector distribution apparent densitiesand infection rates from the five sites was subjected to one-way analysis of variance (ANOVA) using SPSS version 160software at 95 confidence interval Chi-square test was doneto determine the association of infection rates with flyrsquos sexand vector species A correlation analysis was also done toestablishwhether there was a significant relationship betweeninfection rates and apparent densities
3 Results
31 Vector Distribution Out of the 117 flies trapped in thearea G pallidipes constituted the largest percentage of 58whileG longipennis had 42 Among the trapped tsetse flies39 (333) were teneral while 78 (67) were nonteneral InKamunyu area G longipennis was the most prevalent specieswhile G pallidipes dominated Ngiluni Kyusyani Iviani andNthunguni (Figure 4)
Apart from Glossina spp the area was also inhabitedwith mechanical flies such as Tabanus spp and Stomoxys sppTabanids were highly predominant over Stomoxines in all thetrapping sites with an overall fly trap density (FTD) of 011while Stomoxys spp had FTD of 002 Of the total fly catchGlossina spp were the most prevalent in all the tapping sites
FliesGlossina spp Tabanus spp Stomoxys spp
0
10
20
30
40
50
60
70
80
90
100
()
Figure 5 Relative distribution of total fly catch
Trapping sitesKamunyu Kyusyani Iviani Ngiluni Nthunguni
0
01
02
03
04
05
06
Mea
n ap
pare
nt d
ensit
ies
Figure 6 Comparison of mean apparent densities within the fivetrapping sites
while Stomoxys spp constituted the lowest percentage andwere recorded only in Kyusyani area (Figure 5)
32 Apparent Densities (AD) There was no significant dif-ference in the apparent densities (AD) of Glossina speciescaptured across the five trapping sites which ranged from 01to 05 depending on the locality and averaged 03 flies per trapper day (Figure 6)
33 Identification of Trypanosomes in Glossina spp Theexpected band sizes of trypanosomes on amplification usingITS primers are shown in Table 2 Molecular identification oftrypanosomes T vivax and T congolense Forest in Glossinaspp was successful as their DNA amplicons matched theexpected sizes of 611 bp and 1513 bp respectively (Table 2)The amplified fragments for positive controls of T bruceibrucei T congolense Kilifi and T congolense Savannah werealso within the expected sizes (Table 2)
Figure 7 shows a sample of electrophoresis productof successfully amplified DNA fragments of trypanosomespecies T vivax (samples 1 and 5) and the positive controlsT brucei brucei (Tbb) T congolense Kilifi (Tck) and Tcongolense Savannah (Tcs) ldquoMrdquo represents 100 base pairmolecular marker (Promega) while 6 is the negative controlSamples 2 3 and 4 were negative for trypanosomes
34 Infection Rates Out of 78 flies analyzed using nestedPCR and gel electrophoresis the prevalence of trypanosomes
Journal of Parasitology Research 5
Table 2 Documented band sizes of trypanosome species [19]
Trypanosome species Expected band sizesin base pairs (bp)
T congolense forest 1513T congolense Kilifi 1422T congolense Savannah 1413T congolense Tsavo 954T brucei 1207ndash1224T vivax 611
15 kb1kb
500bp
minusVe M Tbb Tck Tcs 1 2 3 4 5 6
Figure 7 An image of an agarose gel showing PCR amplified DNAfragments using ITS nested PCR primers
infection in G pallidipes and G longipennis was 577 and2308 respectively (Table 3)The overall infection rate in allGlossina spp was recorded as 1153 There was a significant(119901 lt 005) difference in the trypanosomes infection rateamong Glossina species Infections with T vivax were themost prevalent (1026) in both species of tsetse comparedto T congolense with a prevalence of 128 (Table 3)
There was significant (119901 lt 005) difference in infectionrates among the trapping sites with Kamunyu recording thehighest rate of 641 followed by Kyusyani at 256 Ngiluniand Iviani recorded similar infection rates of 128 whileNthunguni had an infection rate of 0
A correlation analysis between apparent densities andinfection rates showed a strong positive relationship with acorrelation coefficient of 095 For instance Kamunyu arearecorded the highest apparent density as well as the highestinfection rates
There was a significant difference (119901 lt 005) in T vivaxand T congolense infections amongmale and female flies Forboth T vivax and T congolense the infections were higher inmales than in females with an overall prevalence of 1395and 857 respectively (Table 4)
4 Discussion
41 Vector Distribution in Mtito Andei This study found thatthere was variation in the distribution of G pallidipes andG longipennis in the area G longipennis was the only fuscagroup of flies trapped inMtitoAndei and is known to occur inother Eastern Africa countries such as Uganda Tanzania andEthiopia [2 24]This species is found in drier areasmore thanany other tsetse of fusca group because the pupariumcontainslarge water reserves and a low permeability of its pupalmembranes which helps it to survive harsh environmentalconditions [25 26] The typical habitats for this fly are dry
thorn bush riverline thickets near acacia woodlands and theresting sites are preferably shady sites of tree trunks logsand underside of branches of multistemmed trees [24] MtitoAndei provided all these ecological conditions hence suitablefor this fly
G pallidipes is a morsitans group of flies which is widelydistributed in East Afica and is a major vector of animaltrypanosomiasis [27] It was mostly found in areas close toMtitoAndei River andAthi River as it prefers dense evergreenvegetation heavier shade and humid habitats which areclose to riverline thickets [22] It is also present in otherEastern African countries such as Uganda Ethiopia andSomalia [2 28] Previous studies have shown thatG pallidipesmight be a vector of Trypanosoma b rhodesiense causingHuman African Trypanosomiasis (HAT) in Western Kenyaand Busoga District of Uganda as this parasite has beenisolated from the vector [24 27]
Among the mechanical vectors found in Mtito AndeiTabanids were highly predominant over Stomoxines in allthe trapping sites with the latter being recorded only inKyusyani area This finding corroborates the report detailedin [1] that the species composition of mechanical flies isstrongly dependent on site and substantial differences inenvironmental and climatic conditions of the sampled areaSimilar observations were also made in other areas suchas Brazil [29] The presence of mechanical flies in the areacannot be underestimated as they have been shown tomechanically transmit T vivax and T evansi [30] and canbe responsible for seasonal epidemic patterns in low tsetsedensity areas [1 31]
42 Apparent Densities of Glossina spp This study did notshow significant difference in the apparent densities (AD)of Glossina species captured across the five trapping sitesDespite the low mean AD recorded for tsetse in this area (03flies per trap per day) it is possible that tsetse serve both asbiological transmitters and reservoirs of the parasite while themechanical vectors amplify it during high density periodsOther than amplifying the parasite the mechanical flies canintroduce it to nonendemic areas through immigration
43 Trypanosome Infection Rates This study recorded lowoverall trypanosome infection rates of 1153 Similar resultswere obtained in the Kenyan Coast in a study that reportedtrypanosome infection rates of 1262 724 950 786and 797 in Ukunda Diani Muhaka Shimba hills andMwalewa respectively [32] A comparable study inMouhounRiver Burkina Faso reported overall infection rates of 10 in1423 dissected flies screened for trypanosome infections [33]Further infection rates for T congolense and T vivax wererecorded at 76 and 225 respectively Previous studieshave also showed low prevalence of trypanosome infections(about 10) in the field caught flies and associated this withinteractions of various suppressive factors in the generallysusceptible fly population [34] In addition there are a num-ber of barriers to both establishment and maturation processof trypanosomes and thus only a small proportion of theseinfections reach maturity [24] Further these trypanosomes
6 Journal of Parasitology Research
Table 3 Trypanosome infection rates in Glossina species
Glossina species Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
G pallidipes 49 2 (385) 1 (192) 3 (577) 52G longipennis 20 6 (2308) 0 (0) 6 (2308) 26Total 69 8 (1026) 1 (128) 9 (1153) 78
Table 4 Comparison of infection rates in male and female flies
Sex Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
Male 37 5 (1163) 1 (233) 6 (1395) 43Female 32 3 (857) 0 (0) 3 (857) 35Total 69 8 (1026) 1 (128) 9 (1153) 78
may lack differentiation trigger or may be inhibited by thevectorrsquos immune response [35 36]
Theoverall trypanosomes infection rates inG longipenniswere significantly higher than in G pallidipes However inCoastal Kenya higher infection rates were reported in Gpallidipes (57) than in G longipennis (02) [37] Thedifference in infection rates in the two tsetse species inMtito Andei could be due to variation in feeding preferencesenvironmental factors and host range differences [11 33]Thenumber of parasites available to infect tsetse parasite infec-tivity to tsetse and the strain or subspecies have also beenfound to affect infection rates in tsetse [38] Moreover thenutritional status of the tsetse at the time of infective bloodmeal can also affect their ability to acquire trypanosomeinfections [9] In addition low vectorial capacity has alsobeen reported and is attributed to higher levels of attacinexpression in the proventriculus and midgut [39 40] Thiskind of trait has been reported in G pallidipes and it couldexplain disparity of infection rates between G pallidipes andG longipennis [34] Within species infection rates were alsovariable and this was attributed to individual host factorsFor instance the vulnerability of flies to T brucei infectionswas shown to be due to maternally inherited features whichare associatedwith the presence of intracellular rickettsia-likeorganisms (RLOs) [41] Tsetse carrying these RLOs and othersimultaneous infections such as bacteria fungi and virus inthemidgutweremore likely to be infectedwith trypanosomesthan those without [41ndash43]
T vivax infection was more prevalent in all trappingsites than T congolense Similar results were reported in theLuangwa Valley Zambia [44] However in Cote drsquoIvoire Tcongolense was found to be more infectious (90) comparedto T simiae and T vivax in 50 of 139 microscopically positiveflies that were further analyzed using species specific primersfor T congolense subtypes T simiae and T vivax [45]The higher T vivax infections can be ascribed to shortdevelopmental lifecycle of about 10 days which is entirelycompleted within the proboscis For T congolense and Tbrucei development period has been estimated to be 14days and 30 days respectively [46] Further compared toproboscis tsetse midgut is a hostile environment which
contains midgut lectins which are able to kill trypanosomesin vivo [47] Midgut also has proteolytic digestive enzymespotent antimicrobial substances [34] prophenoloxidase cas-cade and immune molecules such as agglutinins whichhelp in expression of immune effector genes and mediatingcommunication between the gut and the fat body contentswhen microbes are encountered [48] The presence of allthese immune molecules can prevent establishment andmaturation of T congolense and T brucei as these parasitesuse midgut in part of their lifecycles hence the low infectionsrates are detected It is also known that T congolense subtypeparasites are generally low with low parasitaemia comparedto those of T vivax [38] It is therefore possible that suchscanty parasites are rarely picked up by the flies during bloodmeals which may account for the inability of the parasite toget established in the arthropod vector
This study revealed more infection rates in males thanin females In Nigeria higher infection rates in males thanfemales were also reported [49] Some studies suggest thatfemale flies should have higher infection rates than males asthey live longer thanmales and thus they have higher chancesof getting infection [50] However this relationship has notbeen established In contrast other studies explain that moremalesmay be infected than females as they are involved in sexactivities and competition than females [51] Whether moremales or females are infected in field tsetse populations thisstudy found no statististical difference with variation in sex
Apparent densities were strongly positively correlatedwith infection rates (119903 = 095) A similar observationwas reported in Mouhoun River basin Burkina Faso whereinfection rates positively correlated with apparent densities(119903 = 097) [1] This observation validates this parameter as anindicator of AAT transmission risk
5 Conclusion
This study attests that tsetse plays a major role in maintainingthe trypanosome parasite and concludes that sustainablereduction of tsetse from Mtito Andei Division can greatlyreduce the prevalence of AATThis study also showed that thehost factors are equally important in epidemiology of AAT
Journal of Parasitology Research 7
as higher trypanosome infection rates were recorded in Glongipennis than G pallidipes but the sex of the vector is lessimportant Further the study showed that apparent density ofthe vector is a good indicator of AAT transmission risk Thestudy therefore affirms the importance of designing controlstrategies targeting the biological vector (tsetse) mechanicalvectors (Tabanus and Stomoxys spp) and the trypanosomeparasites
6 Recommendations
(1) Since both T congolense and T vivax are pathogenicin cattle and other ruminants with the latter causingacute hemorrhagic syndrome [52] it would be impor-tant to conduct large scale integrated tsetse control inMtitoAndei by application of vector control strategiescoupled with curative treatment of livestock as theseanimals could act as source of reinfection to tsetse
(2) Surveillance studies to determine trypanosomeprevalence in livestock should be conducted in thisarea for effective trypanosomiasis control programs
(3) Entomological surveys should be conducted at dif-ferent seasons to understand seasonal dynamics ofthe vectors and the associated trypanosomiasis riskData on seasonal variations of potential vectors canbe integrated into epidemiologicalmodels to facilitatebetter understanding of the relative importance ofcyclical and mechanical vectors in Mtito Andei
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] N Kone E K NrsquoGoran I Sidibe A W Kombassere and JBouyer ldquoSpatio-temporal distribution of tsetse and other bitingflies in the Mouhoun River basin Burkina Fasordquo Medical andVeterinary Entomology vol 25 no 2 pp 156ndash168 2011
[2] G Cecchi M Paone R A Herrero M J B Vreysen and R CMattioli ldquoDeveloping a continental atlas of the distribution andtrypanosomal infection of tsetse flies (Glossina species)rdquo Para-sites amp Vectors vol 8 article 284 2015
[3] L J Morrison and A MacLeod ldquoAfrican trypanosomiasisrdquoParasite Immunology vol 33 no 8 pp 421ndash422 2011
[4] G Cecchi R C Mattioli J Slingenbergh and S De la RocqueldquoLand cover and tsetse fly distributions in sub-Saharan AfricardquoMedical and Veterinary Entomology vol 22 no 4 pp 364ndash3732008
[5] P Solano S J Torr and M J Lehane ldquoIs vector control neededto eliminate gambiense human African trypanosomiasisrdquoFrontiers in Cellular and Infection Microbiology vol 3 no 1article 33 2013
[6] A P M Shaw G Cecchi G RWWint R C Mattioli and T PRobinson ldquoMapping the economic benefits to livestock keepersfrom intervening against bovine trypanosomosis in EasternAfricardquo Preventive Veterinary Medicine vol 113 no 2 pp 197ndash210 2014
[7] J S Beadell C Hyseni P P Abila et al ldquoPhylogeography andpopulation structure of Glossina fuscipes fuscipes in Ugandaimplications for control of tsetserdquo PLoS Neglected TropicalDiseases vol 4 no 3 article e636 2010
[8] K Bardosh CWaiswa and S CWelburn ldquoConflict of interestuse of pyrethroids and amidines against tsetse and ticks inzoonotic sleeping sickness endemic areas of Ugandardquo Parasitesand Vectors vol 6 article 204 2013
[9] C Kubi J Van denAbbeele R DeDeken TMarcotty P Dornyand P Van Den Bossche ldquoThe effect of starvation on the sus-ceptibility of teneral and non-teneral tsetse flies to trypanosomeinfectionrdquoMedical andVeterinary Entomology vol 20 no 4 pp388ndash392 2006
[10] V Lutje J Seixas and A Kennedy ldquoChemotherapy for second-stage human African trypanosomiasisrdquo Cochrane Database ofSystematic Reviews vol 8 Article ID CD006201 2010
[11] M Bitew Y Amide T Zenebe and H Degefu ldquoTrypanosomeinfection rate inGlossina pallidipes andGlossina fuscipes fuscipesin Gojeb Valley Southwest Ethiopiardquo Global Veterinaria vol 6no 2 pp 131ndash135 2011
[12] R A Oluwafemi A A Ilemobade and E A O Laseinde ldquoTheimpact of African animal trypanosomiasis and tsetse on thelivelihood and well being of cattle and their owners in theBICOT study area ofNigeriardquo Scientific Research andEssays vol2 no 9 pp 380ndash383 2007
[13] R K Oniangrsquoo ldquoEnhancing peoplersquos nutritional status throughrevitalisation of Agriculture and related activitiesrdquo AfricanJournal of Food Agriculture Nutrition and Development vol 1no 1 pp 43ndash50 2001
[14] G L Mugunieri P Irungu and J M Omiti ldquoPerformance ofcommunity-based animal health workers in the delivery of live-stock health servicesrdquo Tropical Animal Health and Productionvol 36 no 6 pp 523ndash535 2004
[15] C A Spinage African EcologyThe Tsetse fly I Africarsquos Bane andBenefice Springer-Verlag Berlin Germany 2012
[16] P Solano J F Michel T Lefrancois et al ldquoPolymerase chainreaction as a diagnosis tool for detecting trypanosomes in nat-urally infected cattle in Burkina Fasordquo Veterinary Parasitologyvol 86 no 2 pp 95ndash103 1999
[17] T Tran G Napier T Rowan et al ldquoDevelopment and eval-uation of an ITS1 lsquoTouchdownrsquo PCR for assessment of drugefficacy against animal African trypanosomosisrdquo VeterinaryParasitology vol 202 no 3-4 pp 164ndash170 2014
[18] S M Thumbi F A McOdimba R O Mosi and J O JungrsquoaldquoComparative evaluation of three PCR base diagnostic assaysfor the detection of pathogenic trypanosomes in cattle bloodrdquoParasites and Vectors vol 1 article 46 2008
[19] A Cox A Tilley F McOdimba et al ldquoA PCR based assay fordetection and differentiation of African trypanosome species inbloodrdquoExperimental Parasitology vol 111 no 1 pp 24ndash29 2005
[20] L Brownhill E M Njuguna and M Miruka Strategy forMainstreaming Gender Analysis in the KARIMcGill UniversityProject Enhancing Ecologically Resilient Food Security ThroughInnovative Farming Systems in the Semi AridMidlands of KenyaKenya Agricultural Research Institute (KARI) McGill Univer-sity 3rd edition 2013
[21] M Thrusfield Veterinary Epidemiology Blackwell ScienceOxford UK 3rd edition 2005
[22] I Malele L Craske C Knight et al ldquoThe use of specific andgeneric primers to identify trypanosome infections of wildtsetse flies in Tanzania by PCRrdquo Infection Genetics and Evolu-tion vol 3 no 4 pp 271ndash279 2003
8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 Journal of Parasitology ResearchEa
ch sp
ecie
s (
)
Kamunyu Ngiluni Kyusyani Iviani NthunguniTrapping sites
G pallidipesG longipennis
0
20
40
60
80
100
120
Figure 4 Distribution of Glossina species captured from study sitesin Mtito Andei Division
The PCR was carried out on a Bio-Rad thermocycler In thesecond round of reaction the outer primers were substitutedwith inner primers (ITS 3 and 4) and 5 120583L of round one PCRproduct was used as DNA template The cycling conditionsfor the second round of reaction were as described for theprimary amplification To analyze the amplicons 10120583L ofthe PCR product was resolved in a 2 agarose gel at 80volts for 45 minutes and the gel was visualised under UVtransilluminator following ethidium bromide staining
26 Data Analysis The infectious trypanosomes were iden-tified by comparing the molecular sizes of their DNA frag-ments with the documented band sizes of trypanosomespecies [19]The data was then used to calculate the infectionrates and prevalence of the trypanosomes on Glossina sppConventional data on vector distribution apparent densitiesand infection rates from the five sites was subjected to one-way analysis of variance (ANOVA) using SPSS version 160software at 95 confidence interval Chi-square test was doneto determine the association of infection rates with flyrsquos sexand vector species A correlation analysis was also done toestablishwhether there was a significant relationship betweeninfection rates and apparent densities
3 Results
31 Vector Distribution Out of the 117 flies trapped in thearea G pallidipes constituted the largest percentage of 58whileG longipennis had 42 Among the trapped tsetse flies39 (333) were teneral while 78 (67) were nonteneral InKamunyu area G longipennis was the most prevalent specieswhile G pallidipes dominated Ngiluni Kyusyani Iviani andNthunguni (Figure 4)
Apart from Glossina spp the area was also inhabitedwith mechanical flies such as Tabanus spp and Stomoxys sppTabanids were highly predominant over Stomoxines in all thetrapping sites with an overall fly trap density (FTD) of 011while Stomoxys spp had FTD of 002 Of the total fly catchGlossina spp were the most prevalent in all the tapping sites
FliesGlossina spp Tabanus spp Stomoxys spp
0
10
20
30
40
50
60
70
80
90
100
()
Figure 5 Relative distribution of total fly catch
Trapping sitesKamunyu Kyusyani Iviani Ngiluni Nthunguni
0
01
02
03
04
05
06
Mea
n ap
pare
nt d
ensit
ies
Figure 6 Comparison of mean apparent densities within the fivetrapping sites
while Stomoxys spp constituted the lowest percentage andwere recorded only in Kyusyani area (Figure 5)
32 Apparent Densities (AD) There was no significant dif-ference in the apparent densities (AD) of Glossina speciescaptured across the five trapping sites which ranged from 01to 05 depending on the locality and averaged 03 flies per trapper day (Figure 6)
33 Identification of Trypanosomes in Glossina spp Theexpected band sizes of trypanosomes on amplification usingITS primers are shown in Table 2 Molecular identification oftrypanosomes T vivax and T congolense Forest in Glossinaspp was successful as their DNA amplicons matched theexpected sizes of 611 bp and 1513 bp respectively (Table 2)The amplified fragments for positive controls of T bruceibrucei T congolense Kilifi and T congolense Savannah werealso within the expected sizes (Table 2)
Figure 7 shows a sample of electrophoresis productof successfully amplified DNA fragments of trypanosomespecies T vivax (samples 1 and 5) and the positive controlsT brucei brucei (Tbb) T congolense Kilifi (Tck) and Tcongolense Savannah (Tcs) ldquoMrdquo represents 100 base pairmolecular marker (Promega) while 6 is the negative controlSamples 2 3 and 4 were negative for trypanosomes
34 Infection Rates Out of 78 flies analyzed using nestedPCR and gel electrophoresis the prevalence of trypanosomes
Journal of Parasitology Research 5
Table 2 Documented band sizes of trypanosome species [19]
Trypanosome species Expected band sizesin base pairs (bp)
T congolense forest 1513T congolense Kilifi 1422T congolense Savannah 1413T congolense Tsavo 954T brucei 1207ndash1224T vivax 611
15 kb1kb
500bp
minusVe M Tbb Tck Tcs 1 2 3 4 5 6
Figure 7 An image of an agarose gel showing PCR amplified DNAfragments using ITS nested PCR primers
infection in G pallidipes and G longipennis was 577 and2308 respectively (Table 3)The overall infection rate in allGlossina spp was recorded as 1153 There was a significant(119901 lt 005) difference in the trypanosomes infection rateamong Glossina species Infections with T vivax were themost prevalent (1026) in both species of tsetse comparedto T congolense with a prevalence of 128 (Table 3)
There was significant (119901 lt 005) difference in infectionrates among the trapping sites with Kamunyu recording thehighest rate of 641 followed by Kyusyani at 256 Ngiluniand Iviani recorded similar infection rates of 128 whileNthunguni had an infection rate of 0
A correlation analysis between apparent densities andinfection rates showed a strong positive relationship with acorrelation coefficient of 095 For instance Kamunyu arearecorded the highest apparent density as well as the highestinfection rates
There was a significant difference (119901 lt 005) in T vivaxand T congolense infections amongmale and female flies Forboth T vivax and T congolense the infections were higher inmales than in females with an overall prevalence of 1395and 857 respectively (Table 4)
4 Discussion
41 Vector Distribution in Mtito Andei This study found thatthere was variation in the distribution of G pallidipes andG longipennis in the area G longipennis was the only fuscagroup of flies trapped inMtitoAndei and is known to occur inother Eastern Africa countries such as Uganda Tanzania andEthiopia [2 24]This species is found in drier areasmore thanany other tsetse of fusca group because the pupariumcontainslarge water reserves and a low permeability of its pupalmembranes which helps it to survive harsh environmentalconditions [25 26] The typical habitats for this fly are dry
thorn bush riverline thickets near acacia woodlands and theresting sites are preferably shady sites of tree trunks logsand underside of branches of multistemmed trees [24] MtitoAndei provided all these ecological conditions hence suitablefor this fly
G pallidipes is a morsitans group of flies which is widelydistributed in East Afica and is a major vector of animaltrypanosomiasis [27] It was mostly found in areas close toMtitoAndei River andAthi River as it prefers dense evergreenvegetation heavier shade and humid habitats which areclose to riverline thickets [22] It is also present in otherEastern African countries such as Uganda Ethiopia andSomalia [2 28] Previous studies have shown thatG pallidipesmight be a vector of Trypanosoma b rhodesiense causingHuman African Trypanosomiasis (HAT) in Western Kenyaand Busoga District of Uganda as this parasite has beenisolated from the vector [24 27]
Among the mechanical vectors found in Mtito AndeiTabanids were highly predominant over Stomoxines in allthe trapping sites with the latter being recorded only inKyusyani area This finding corroborates the report detailedin [1] that the species composition of mechanical flies isstrongly dependent on site and substantial differences inenvironmental and climatic conditions of the sampled areaSimilar observations were also made in other areas suchas Brazil [29] The presence of mechanical flies in the areacannot be underestimated as they have been shown tomechanically transmit T vivax and T evansi [30] and canbe responsible for seasonal epidemic patterns in low tsetsedensity areas [1 31]
42 Apparent Densities of Glossina spp This study did notshow significant difference in the apparent densities (AD)of Glossina species captured across the five trapping sitesDespite the low mean AD recorded for tsetse in this area (03flies per trap per day) it is possible that tsetse serve both asbiological transmitters and reservoirs of the parasite while themechanical vectors amplify it during high density periodsOther than amplifying the parasite the mechanical flies canintroduce it to nonendemic areas through immigration
43 Trypanosome Infection Rates This study recorded lowoverall trypanosome infection rates of 1153 Similar resultswere obtained in the Kenyan Coast in a study that reportedtrypanosome infection rates of 1262 724 950 786and 797 in Ukunda Diani Muhaka Shimba hills andMwalewa respectively [32] A comparable study inMouhounRiver Burkina Faso reported overall infection rates of 10 in1423 dissected flies screened for trypanosome infections [33]Further infection rates for T congolense and T vivax wererecorded at 76 and 225 respectively Previous studieshave also showed low prevalence of trypanosome infections(about 10) in the field caught flies and associated this withinteractions of various suppressive factors in the generallysusceptible fly population [34] In addition there are a num-ber of barriers to both establishment and maturation processof trypanosomes and thus only a small proportion of theseinfections reach maturity [24] Further these trypanosomes
6 Journal of Parasitology Research
Table 3 Trypanosome infection rates in Glossina species
Glossina species Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
G pallidipes 49 2 (385) 1 (192) 3 (577) 52G longipennis 20 6 (2308) 0 (0) 6 (2308) 26Total 69 8 (1026) 1 (128) 9 (1153) 78
Table 4 Comparison of infection rates in male and female flies
Sex Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
Male 37 5 (1163) 1 (233) 6 (1395) 43Female 32 3 (857) 0 (0) 3 (857) 35Total 69 8 (1026) 1 (128) 9 (1153) 78
may lack differentiation trigger or may be inhibited by thevectorrsquos immune response [35 36]
Theoverall trypanosomes infection rates inG longipenniswere significantly higher than in G pallidipes However inCoastal Kenya higher infection rates were reported in Gpallidipes (57) than in G longipennis (02) [37] Thedifference in infection rates in the two tsetse species inMtito Andei could be due to variation in feeding preferencesenvironmental factors and host range differences [11 33]Thenumber of parasites available to infect tsetse parasite infec-tivity to tsetse and the strain or subspecies have also beenfound to affect infection rates in tsetse [38] Moreover thenutritional status of the tsetse at the time of infective bloodmeal can also affect their ability to acquire trypanosomeinfections [9] In addition low vectorial capacity has alsobeen reported and is attributed to higher levels of attacinexpression in the proventriculus and midgut [39 40] Thiskind of trait has been reported in G pallidipes and it couldexplain disparity of infection rates between G pallidipes andG longipennis [34] Within species infection rates were alsovariable and this was attributed to individual host factorsFor instance the vulnerability of flies to T brucei infectionswas shown to be due to maternally inherited features whichare associatedwith the presence of intracellular rickettsia-likeorganisms (RLOs) [41] Tsetse carrying these RLOs and othersimultaneous infections such as bacteria fungi and virus inthemidgutweremore likely to be infectedwith trypanosomesthan those without [41ndash43]
T vivax infection was more prevalent in all trappingsites than T congolense Similar results were reported in theLuangwa Valley Zambia [44] However in Cote drsquoIvoire Tcongolense was found to be more infectious (90) comparedto T simiae and T vivax in 50 of 139 microscopically positiveflies that were further analyzed using species specific primersfor T congolense subtypes T simiae and T vivax [45]The higher T vivax infections can be ascribed to shortdevelopmental lifecycle of about 10 days which is entirelycompleted within the proboscis For T congolense and Tbrucei development period has been estimated to be 14days and 30 days respectively [46] Further compared toproboscis tsetse midgut is a hostile environment which
contains midgut lectins which are able to kill trypanosomesin vivo [47] Midgut also has proteolytic digestive enzymespotent antimicrobial substances [34] prophenoloxidase cas-cade and immune molecules such as agglutinins whichhelp in expression of immune effector genes and mediatingcommunication between the gut and the fat body contentswhen microbes are encountered [48] The presence of allthese immune molecules can prevent establishment andmaturation of T congolense and T brucei as these parasitesuse midgut in part of their lifecycles hence the low infectionsrates are detected It is also known that T congolense subtypeparasites are generally low with low parasitaemia comparedto those of T vivax [38] It is therefore possible that suchscanty parasites are rarely picked up by the flies during bloodmeals which may account for the inability of the parasite toget established in the arthropod vector
This study revealed more infection rates in males thanin females In Nigeria higher infection rates in males thanfemales were also reported [49] Some studies suggest thatfemale flies should have higher infection rates than males asthey live longer thanmales and thus they have higher chancesof getting infection [50] However this relationship has notbeen established In contrast other studies explain that moremalesmay be infected than females as they are involved in sexactivities and competition than females [51] Whether moremales or females are infected in field tsetse populations thisstudy found no statististical difference with variation in sex
Apparent densities were strongly positively correlatedwith infection rates (119903 = 095) A similar observationwas reported in Mouhoun River basin Burkina Faso whereinfection rates positively correlated with apparent densities(119903 = 097) [1] This observation validates this parameter as anindicator of AAT transmission risk
5 Conclusion
This study attests that tsetse plays a major role in maintainingthe trypanosome parasite and concludes that sustainablereduction of tsetse from Mtito Andei Division can greatlyreduce the prevalence of AATThis study also showed that thehost factors are equally important in epidemiology of AAT
Journal of Parasitology Research 7
as higher trypanosome infection rates were recorded in Glongipennis than G pallidipes but the sex of the vector is lessimportant Further the study showed that apparent density ofthe vector is a good indicator of AAT transmission risk Thestudy therefore affirms the importance of designing controlstrategies targeting the biological vector (tsetse) mechanicalvectors (Tabanus and Stomoxys spp) and the trypanosomeparasites
6 Recommendations
(1) Since both T congolense and T vivax are pathogenicin cattle and other ruminants with the latter causingacute hemorrhagic syndrome [52] it would be impor-tant to conduct large scale integrated tsetse control inMtitoAndei by application of vector control strategiescoupled with curative treatment of livestock as theseanimals could act as source of reinfection to tsetse
(2) Surveillance studies to determine trypanosomeprevalence in livestock should be conducted in thisarea for effective trypanosomiasis control programs
(3) Entomological surveys should be conducted at dif-ferent seasons to understand seasonal dynamics ofthe vectors and the associated trypanosomiasis riskData on seasonal variations of potential vectors canbe integrated into epidemiologicalmodels to facilitatebetter understanding of the relative importance ofcyclical and mechanical vectors in Mtito Andei
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] N Kone E K NrsquoGoran I Sidibe A W Kombassere and JBouyer ldquoSpatio-temporal distribution of tsetse and other bitingflies in the Mouhoun River basin Burkina Fasordquo Medical andVeterinary Entomology vol 25 no 2 pp 156ndash168 2011
[2] G Cecchi M Paone R A Herrero M J B Vreysen and R CMattioli ldquoDeveloping a continental atlas of the distribution andtrypanosomal infection of tsetse flies (Glossina species)rdquo Para-sites amp Vectors vol 8 article 284 2015
[3] L J Morrison and A MacLeod ldquoAfrican trypanosomiasisrdquoParasite Immunology vol 33 no 8 pp 421ndash422 2011
[4] G Cecchi R C Mattioli J Slingenbergh and S De la RocqueldquoLand cover and tsetse fly distributions in sub-Saharan AfricardquoMedical and Veterinary Entomology vol 22 no 4 pp 364ndash3732008
[5] P Solano S J Torr and M J Lehane ldquoIs vector control neededto eliminate gambiense human African trypanosomiasisrdquoFrontiers in Cellular and Infection Microbiology vol 3 no 1article 33 2013
[6] A P M Shaw G Cecchi G RWWint R C Mattioli and T PRobinson ldquoMapping the economic benefits to livestock keepersfrom intervening against bovine trypanosomosis in EasternAfricardquo Preventive Veterinary Medicine vol 113 no 2 pp 197ndash210 2014
[7] J S Beadell C Hyseni P P Abila et al ldquoPhylogeography andpopulation structure of Glossina fuscipes fuscipes in Ugandaimplications for control of tsetserdquo PLoS Neglected TropicalDiseases vol 4 no 3 article e636 2010
[8] K Bardosh CWaiswa and S CWelburn ldquoConflict of interestuse of pyrethroids and amidines against tsetse and ticks inzoonotic sleeping sickness endemic areas of Ugandardquo Parasitesand Vectors vol 6 article 204 2013
[9] C Kubi J Van denAbbeele R DeDeken TMarcotty P Dornyand P Van Den Bossche ldquoThe effect of starvation on the sus-ceptibility of teneral and non-teneral tsetse flies to trypanosomeinfectionrdquoMedical andVeterinary Entomology vol 20 no 4 pp388ndash392 2006
[10] V Lutje J Seixas and A Kennedy ldquoChemotherapy for second-stage human African trypanosomiasisrdquo Cochrane Database ofSystematic Reviews vol 8 Article ID CD006201 2010
[11] M Bitew Y Amide T Zenebe and H Degefu ldquoTrypanosomeinfection rate inGlossina pallidipes andGlossina fuscipes fuscipesin Gojeb Valley Southwest Ethiopiardquo Global Veterinaria vol 6no 2 pp 131ndash135 2011
[12] R A Oluwafemi A A Ilemobade and E A O Laseinde ldquoTheimpact of African animal trypanosomiasis and tsetse on thelivelihood and well being of cattle and their owners in theBICOT study area ofNigeriardquo Scientific Research andEssays vol2 no 9 pp 380ndash383 2007
[13] R K Oniangrsquoo ldquoEnhancing peoplersquos nutritional status throughrevitalisation of Agriculture and related activitiesrdquo AfricanJournal of Food Agriculture Nutrition and Development vol 1no 1 pp 43ndash50 2001
[14] G L Mugunieri P Irungu and J M Omiti ldquoPerformance ofcommunity-based animal health workers in the delivery of live-stock health servicesrdquo Tropical Animal Health and Productionvol 36 no 6 pp 523ndash535 2004
[15] C A Spinage African EcologyThe Tsetse fly I Africarsquos Bane andBenefice Springer-Verlag Berlin Germany 2012
[16] P Solano J F Michel T Lefrancois et al ldquoPolymerase chainreaction as a diagnosis tool for detecting trypanosomes in nat-urally infected cattle in Burkina Fasordquo Veterinary Parasitologyvol 86 no 2 pp 95ndash103 1999
[17] T Tran G Napier T Rowan et al ldquoDevelopment and eval-uation of an ITS1 lsquoTouchdownrsquo PCR for assessment of drugefficacy against animal African trypanosomosisrdquo VeterinaryParasitology vol 202 no 3-4 pp 164ndash170 2014
[18] S M Thumbi F A McOdimba R O Mosi and J O JungrsquoaldquoComparative evaluation of three PCR base diagnostic assaysfor the detection of pathogenic trypanosomes in cattle bloodrdquoParasites and Vectors vol 1 article 46 2008
[19] A Cox A Tilley F McOdimba et al ldquoA PCR based assay fordetection and differentiation of African trypanosome species inbloodrdquoExperimental Parasitology vol 111 no 1 pp 24ndash29 2005
[20] L Brownhill E M Njuguna and M Miruka Strategy forMainstreaming Gender Analysis in the KARIMcGill UniversityProject Enhancing Ecologically Resilient Food Security ThroughInnovative Farming Systems in the Semi AridMidlands of KenyaKenya Agricultural Research Institute (KARI) McGill Univer-sity 3rd edition 2013
[21] M Thrusfield Veterinary Epidemiology Blackwell ScienceOxford UK 3rd edition 2005
[22] I Malele L Craske C Knight et al ldquoThe use of specific andgeneric primers to identify trypanosome infections of wildtsetse flies in Tanzania by PCRrdquo Infection Genetics and Evolu-tion vol 3 no 4 pp 271ndash279 2003
8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Journal of Parasitology Research 5
Table 2 Documented band sizes of trypanosome species [19]
Trypanosome species Expected band sizesin base pairs (bp)
T congolense forest 1513T congolense Kilifi 1422T congolense Savannah 1413T congolense Tsavo 954T brucei 1207ndash1224T vivax 611
15 kb1kb
500bp
minusVe M Tbb Tck Tcs 1 2 3 4 5 6
Figure 7 An image of an agarose gel showing PCR amplified DNAfragments using ITS nested PCR primers
infection in G pallidipes and G longipennis was 577 and2308 respectively (Table 3)The overall infection rate in allGlossina spp was recorded as 1153 There was a significant(119901 lt 005) difference in the trypanosomes infection rateamong Glossina species Infections with T vivax were themost prevalent (1026) in both species of tsetse comparedto T congolense with a prevalence of 128 (Table 3)
There was significant (119901 lt 005) difference in infectionrates among the trapping sites with Kamunyu recording thehighest rate of 641 followed by Kyusyani at 256 Ngiluniand Iviani recorded similar infection rates of 128 whileNthunguni had an infection rate of 0
A correlation analysis between apparent densities andinfection rates showed a strong positive relationship with acorrelation coefficient of 095 For instance Kamunyu arearecorded the highest apparent density as well as the highestinfection rates
There was a significant difference (119901 lt 005) in T vivaxand T congolense infections amongmale and female flies Forboth T vivax and T congolense the infections were higher inmales than in females with an overall prevalence of 1395and 857 respectively (Table 4)
4 Discussion
41 Vector Distribution in Mtito Andei This study found thatthere was variation in the distribution of G pallidipes andG longipennis in the area G longipennis was the only fuscagroup of flies trapped inMtitoAndei and is known to occur inother Eastern Africa countries such as Uganda Tanzania andEthiopia [2 24]This species is found in drier areasmore thanany other tsetse of fusca group because the pupariumcontainslarge water reserves and a low permeability of its pupalmembranes which helps it to survive harsh environmentalconditions [25 26] The typical habitats for this fly are dry
thorn bush riverline thickets near acacia woodlands and theresting sites are preferably shady sites of tree trunks logsand underside of branches of multistemmed trees [24] MtitoAndei provided all these ecological conditions hence suitablefor this fly
G pallidipes is a morsitans group of flies which is widelydistributed in East Afica and is a major vector of animaltrypanosomiasis [27] It was mostly found in areas close toMtitoAndei River andAthi River as it prefers dense evergreenvegetation heavier shade and humid habitats which areclose to riverline thickets [22] It is also present in otherEastern African countries such as Uganda Ethiopia andSomalia [2 28] Previous studies have shown thatG pallidipesmight be a vector of Trypanosoma b rhodesiense causingHuman African Trypanosomiasis (HAT) in Western Kenyaand Busoga District of Uganda as this parasite has beenisolated from the vector [24 27]
Among the mechanical vectors found in Mtito AndeiTabanids were highly predominant over Stomoxines in allthe trapping sites with the latter being recorded only inKyusyani area This finding corroborates the report detailedin [1] that the species composition of mechanical flies isstrongly dependent on site and substantial differences inenvironmental and climatic conditions of the sampled areaSimilar observations were also made in other areas suchas Brazil [29] The presence of mechanical flies in the areacannot be underestimated as they have been shown tomechanically transmit T vivax and T evansi [30] and canbe responsible for seasonal epidemic patterns in low tsetsedensity areas [1 31]
42 Apparent Densities of Glossina spp This study did notshow significant difference in the apparent densities (AD)of Glossina species captured across the five trapping sitesDespite the low mean AD recorded for tsetse in this area (03flies per trap per day) it is possible that tsetse serve both asbiological transmitters and reservoirs of the parasite while themechanical vectors amplify it during high density periodsOther than amplifying the parasite the mechanical flies canintroduce it to nonendemic areas through immigration
43 Trypanosome Infection Rates This study recorded lowoverall trypanosome infection rates of 1153 Similar resultswere obtained in the Kenyan Coast in a study that reportedtrypanosome infection rates of 1262 724 950 786and 797 in Ukunda Diani Muhaka Shimba hills andMwalewa respectively [32] A comparable study inMouhounRiver Burkina Faso reported overall infection rates of 10 in1423 dissected flies screened for trypanosome infections [33]Further infection rates for T congolense and T vivax wererecorded at 76 and 225 respectively Previous studieshave also showed low prevalence of trypanosome infections(about 10) in the field caught flies and associated this withinteractions of various suppressive factors in the generallysusceptible fly population [34] In addition there are a num-ber of barriers to both establishment and maturation processof trypanosomes and thus only a small proportion of theseinfections reach maturity [24] Further these trypanosomes
6 Journal of Parasitology Research
Table 3 Trypanosome infection rates in Glossina species
Glossina species Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
G pallidipes 49 2 (385) 1 (192) 3 (577) 52G longipennis 20 6 (2308) 0 (0) 6 (2308) 26Total 69 8 (1026) 1 (128) 9 (1153) 78
Table 4 Comparison of infection rates in male and female flies
Sex Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
Male 37 5 (1163) 1 (233) 6 (1395) 43Female 32 3 (857) 0 (0) 3 (857) 35Total 69 8 (1026) 1 (128) 9 (1153) 78
may lack differentiation trigger or may be inhibited by thevectorrsquos immune response [35 36]
Theoverall trypanosomes infection rates inG longipenniswere significantly higher than in G pallidipes However inCoastal Kenya higher infection rates were reported in Gpallidipes (57) than in G longipennis (02) [37] Thedifference in infection rates in the two tsetse species inMtito Andei could be due to variation in feeding preferencesenvironmental factors and host range differences [11 33]Thenumber of parasites available to infect tsetse parasite infec-tivity to tsetse and the strain or subspecies have also beenfound to affect infection rates in tsetse [38] Moreover thenutritional status of the tsetse at the time of infective bloodmeal can also affect their ability to acquire trypanosomeinfections [9] In addition low vectorial capacity has alsobeen reported and is attributed to higher levels of attacinexpression in the proventriculus and midgut [39 40] Thiskind of trait has been reported in G pallidipes and it couldexplain disparity of infection rates between G pallidipes andG longipennis [34] Within species infection rates were alsovariable and this was attributed to individual host factorsFor instance the vulnerability of flies to T brucei infectionswas shown to be due to maternally inherited features whichare associatedwith the presence of intracellular rickettsia-likeorganisms (RLOs) [41] Tsetse carrying these RLOs and othersimultaneous infections such as bacteria fungi and virus inthemidgutweremore likely to be infectedwith trypanosomesthan those without [41ndash43]
T vivax infection was more prevalent in all trappingsites than T congolense Similar results were reported in theLuangwa Valley Zambia [44] However in Cote drsquoIvoire Tcongolense was found to be more infectious (90) comparedto T simiae and T vivax in 50 of 139 microscopically positiveflies that were further analyzed using species specific primersfor T congolense subtypes T simiae and T vivax [45]The higher T vivax infections can be ascribed to shortdevelopmental lifecycle of about 10 days which is entirelycompleted within the proboscis For T congolense and Tbrucei development period has been estimated to be 14days and 30 days respectively [46] Further compared toproboscis tsetse midgut is a hostile environment which
contains midgut lectins which are able to kill trypanosomesin vivo [47] Midgut also has proteolytic digestive enzymespotent antimicrobial substances [34] prophenoloxidase cas-cade and immune molecules such as agglutinins whichhelp in expression of immune effector genes and mediatingcommunication between the gut and the fat body contentswhen microbes are encountered [48] The presence of allthese immune molecules can prevent establishment andmaturation of T congolense and T brucei as these parasitesuse midgut in part of their lifecycles hence the low infectionsrates are detected It is also known that T congolense subtypeparasites are generally low with low parasitaemia comparedto those of T vivax [38] It is therefore possible that suchscanty parasites are rarely picked up by the flies during bloodmeals which may account for the inability of the parasite toget established in the arthropod vector
This study revealed more infection rates in males thanin females In Nigeria higher infection rates in males thanfemales were also reported [49] Some studies suggest thatfemale flies should have higher infection rates than males asthey live longer thanmales and thus they have higher chancesof getting infection [50] However this relationship has notbeen established In contrast other studies explain that moremalesmay be infected than females as they are involved in sexactivities and competition than females [51] Whether moremales or females are infected in field tsetse populations thisstudy found no statististical difference with variation in sex
Apparent densities were strongly positively correlatedwith infection rates (119903 = 095) A similar observationwas reported in Mouhoun River basin Burkina Faso whereinfection rates positively correlated with apparent densities(119903 = 097) [1] This observation validates this parameter as anindicator of AAT transmission risk
5 Conclusion
This study attests that tsetse plays a major role in maintainingthe trypanosome parasite and concludes that sustainablereduction of tsetse from Mtito Andei Division can greatlyreduce the prevalence of AATThis study also showed that thehost factors are equally important in epidemiology of AAT
Journal of Parasitology Research 7
as higher trypanosome infection rates were recorded in Glongipennis than G pallidipes but the sex of the vector is lessimportant Further the study showed that apparent density ofthe vector is a good indicator of AAT transmission risk Thestudy therefore affirms the importance of designing controlstrategies targeting the biological vector (tsetse) mechanicalvectors (Tabanus and Stomoxys spp) and the trypanosomeparasites
6 Recommendations
(1) Since both T congolense and T vivax are pathogenicin cattle and other ruminants with the latter causingacute hemorrhagic syndrome [52] it would be impor-tant to conduct large scale integrated tsetse control inMtitoAndei by application of vector control strategiescoupled with curative treatment of livestock as theseanimals could act as source of reinfection to tsetse
(2) Surveillance studies to determine trypanosomeprevalence in livestock should be conducted in thisarea for effective trypanosomiasis control programs
(3) Entomological surveys should be conducted at dif-ferent seasons to understand seasonal dynamics ofthe vectors and the associated trypanosomiasis riskData on seasonal variations of potential vectors canbe integrated into epidemiologicalmodels to facilitatebetter understanding of the relative importance ofcyclical and mechanical vectors in Mtito Andei
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] N Kone E K NrsquoGoran I Sidibe A W Kombassere and JBouyer ldquoSpatio-temporal distribution of tsetse and other bitingflies in the Mouhoun River basin Burkina Fasordquo Medical andVeterinary Entomology vol 25 no 2 pp 156ndash168 2011
[2] G Cecchi M Paone R A Herrero M J B Vreysen and R CMattioli ldquoDeveloping a continental atlas of the distribution andtrypanosomal infection of tsetse flies (Glossina species)rdquo Para-sites amp Vectors vol 8 article 284 2015
[3] L J Morrison and A MacLeod ldquoAfrican trypanosomiasisrdquoParasite Immunology vol 33 no 8 pp 421ndash422 2011
[4] G Cecchi R C Mattioli J Slingenbergh and S De la RocqueldquoLand cover and tsetse fly distributions in sub-Saharan AfricardquoMedical and Veterinary Entomology vol 22 no 4 pp 364ndash3732008
[5] P Solano S J Torr and M J Lehane ldquoIs vector control neededto eliminate gambiense human African trypanosomiasisrdquoFrontiers in Cellular and Infection Microbiology vol 3 no 1article 33 2013
[6] A P M Shaw G Cecchi G RWWint R C Mattioli and T PRobinson ldquoMapping the economic benefits to livestock keepersfrom intervening against bovine trypanosomosis in EasternAfricardquo Preventive Veterinary Medicine vol 113 no 2 pp 197ndash210 2014
[7] J S Beadell C Hyseni P P Abila et al ldquoPhylogeography andpopulation structure of Glossina fuscipes fuscipes in Ugandaimplications for control of tsetserdquo PLoS Neglected TropicalDiseases vol 4 no 3 article e636 2010
[8] K Bardosh CWaiswa and S CWelburn ldquoConflict of interestuse of pyrethroids and amidines against tsetse and ticks inzoonotic sleeping sickness endemic areas of Ugandardquo Parasitesand Vectors vol 6 article 204 2013
[9] C Kubi J Van denAbbeele R DeDeken TMarcotty P Dornyand P Van Den Bossche ldquoThe effect of starvation on the sus-ceptibility of teneral and non-teneral tsetse flies to trypanosomeinfectionrdquoMedical andVeterinary Entomology vol 20 no 4 pp388ndash392 2006
[10] V Lutje J Seixas and A Kennedy ldquoChemotherapy for second-stage human African trypanosomiasisrdquo Cochrane Database ofSystematic Reviews vol 8 Article ID CD006201 2010
[11] M Bitew Y Amide T Zenebe and H Degefu ldquoTrypanosomeinfection rate inGlossina pallidipes andGlossina fuscipes fuscipesin Gojeb Valley Southwest Ethiopiardquo Global Veterinaria vol 6no 2 pp 131ndash135 2011
[12] R A Oluwafemi A A Ilemobade and E A O Laseinde ldquoTheimpact of African animal trypanosomiasis and tsetse on thelivelihood and well being of cattle and their owners in theBICOT study area ofNigeriardquo Scientific Research andEssays vol2 no 9 pp 380ndash383 2007
[13] R K Oniangrsquoo ldquoEnhancing peoplersquos nutritional status throughrevitalisation of Agriculture and related activitiesrdquo AfricanJournal of Food Agriculture Nutrition and Development vol 1no 1 pp 43ndash50 2001
[14] G L Mugunieri P Irungu and J M Omiti ldquoPerformance ofcommunity-based animal health workers in the delivery of live-stock health servicesrdquo Tropical Animal Health and Productionvol 36 no 6 pp 523ndash535 2004
[15] C A Spinage African EcologyThe Tsetse fly I Africarsquos Bane andBenefice Springer-Verlag Berlin Germany 2012
[16] P Solano J F Michel T Lefrancois et al ldquoPolymerase chainreaction as a diagnosis tool for detecting trypanosomes in nat-urally infected cattle in Burkina Fasordquo Veterinary Parasitologyvol 86 no 2 pp 95ndash103 1999
[17] T Tran G Napier T Rowan et al ldquoDevelopment and eval-uation of an ITS1 lsquoTouchdownrsquo PCR for assessment of drugefficacy against animal African trypanosomosisrdquo VeterinaryParasitology vol 202 no 3-4 pp 164ndash170 2014
[18] S M Thumbi F A McOdimba R O Mosi and J O JungrsquoaldquoComparative evaluation of three PCR base diagnostic assaysfor the detection of pathogenic trypanosomes in cattle bloodrdquoParasites and Vectors vol 1 article 46 2008
[19] A Cox A Tilley F McOdimba et al ldquoA PCR based assay fordetection and differentiation of African trypanosome species inbloodrdquoExperimental Parasitology vol 111 no 1 pp 24ndash29 2005
[20] L Brownhill E M Njuguna and M Miruka Strategy forMainstreaming Gender Analysis in the KARIMcGill UniversityProject Enhancing Ecologically Resilient Food Security ThroughInnovative Farming Systems in the Semi AridMidlands of KenyaKenya Agricultural Research Institute (KARI) McGill Univer-sity 3rd edition 2013
[21] M Thrusfield Veterinary Epidemiology Blackwell ScienceOxford UK 3rd edition 2005
[22] I Malele L Craske C Knight et al ldquoThe use of specific andgeneric primers to identify trypanosome infections of wildtsetse flies in Tanzania by PCRrdquo Infection Genetics and Evolu-tion vol 3 no 4 pp 271ndash279 2003
8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 Journal of Parasitology Research
Table 3 Trypanosome infection rates in Glossina species
Glossina species Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
G pallidipes 49 2 (385) 1 (192) 3 (577) 52G longipennis 20 6 (2308) 0 (0) 6 (2308) 26Total 69 8 (1026) 1 (128) 9 (1153) 78
Table 4 Comparison of infection rates in male and female flies
Sex Infection rates Overall infection rates () TotalUninfected T vivax T congolense forest
Male 37 5 (1163) 1 (233) 6 (1395) 43Female 32 3 (857) 0 (0) 3 (857) 35Total 69 8 (1026) 1 (128) 9 (1153) 78
may lack differentiation trigger or may be inhibited by thevectorrsquos immune response [35 36]
Theoverall trypanosomes infection rates inG longipenniswere significantly higher than in G pallidipes However inCoastal Kenya higher infection rates were reported in Gpallidipes (57) than in G longipennis (02) [37] Thedifference in infection rates in the two tsetse species inMtito Andei could be due to variation in feeding preferencesenvironmental factors and host range differences [11 33]Thenumber of parasites available to infect tsetse parasite infec-tivity to tsetse and the strain or subspecies have also beenfound to affect infection rates in tsetse [38] Moreover thenutritional status of the tsetse at the time of infective bloodmeal can also affect their ability to acquire trypanosomeinfections [9] In addition low vectorial capacity has alsobeen reported and is attributed to higher levels of attacinexpression in the proventriculus and midgut [39 40] Thiskind of trait has been reported in G pallidipes and it couldexplain disparity of infection rates between G pallidipes andG longipennis [34] Within species infection rates were alsovariable and this was attributed to individual host factorsFor instance the vulnerability of flies to T brucei infectionswas shown to be due to maternally inherited features whichare associatedwith the presence of intracellular rickettsia-likeorganisms (RLOs) [41] Tsetse carrying these RLOs and othersimultaneous infections such as bacteria fungi and virus inthemidgutweremore likely to be infectedwith trypanosomesthan those without [41ndash43]
T vivax infection was more prevalent in all trappingsites than T congolense Similar results were reported in theLuangwa Valley Zambia [44] However in Cote drsquoIvoire Tcongolense was found to be more infectious (90) comparedto T simiae and T vivax in 50 of 139 microscopically positiveflies that were further analyzed using species specific primersfor T congolense subtypes T simiae and T vivax [45]The higher T vivax infections can be ascribed to shortdevelopmental lifecycle of about 10 days which is entirelycompleted within the proboscis For T congolense and Tbrucei development period has been estimated to be 14days and 30 days respectively [46] Further compared toproboscis tsetse midgut is a hostile environment which
contains midgut lectins which are able to kill trypanosomesin vivo [47] Midgut also has proteolytic digestive enzymespotent antimicrobial substances [34] prophenoloxidase cas-cade and immune molecules such as agglutinins whichhelp in expression of immune effector genes and mediatingcommunication between the gut and the fat body contentswhen microbes are encountered [48] The presence of allthese immune molecules can prevent establishment andmaturation of T congolense and T brucei as these parasitesuse midgut in part of their lifecycles hence the low infectionsrates are detected It is also known that T congolense subtypeparasites are generally low with low parasitaemia comparedto those of T vivax [38] It is therefore possible that suchscanty parasites are rarely picked up by the flies during bloodmeals which may account for the inability of the parasite toget established in the arthropod vector
This study revealed more infection rates in males thanin females In Nigeria higher infection rates in males thanfemales were also reported [49] Some studies suggest thatfemale flies should have higher infection rates than males asthey live longer thanmales and thus they have higher chancesof getting infection [50] However this relationship has notbeen established In contrast other studies explain that moremalesmay be infected than females as they are involved in sexactivities and competition than females [51] Whether moremales or females are infected in field tsetse populations thisstudy found no statististical difference with variation in sex
Apparent densities were strongly positively correlatedwith infection rates (119903 = 095) A similar observationwas reported in Mouhoun River basin Burkina Faso whereinfection rates positively correlated with apparent densities(119903 = 097) [1] This observation validates this parameter as anindicator of AAT transmission risk
5 Conclusion
This study attests that tsetse plays a major role in maintainingthe trypanosome parasite and concludes that sustainablereduction of tsetse from Mtito Andei Division can greatlyreduce the prevalence of AATThis study also showed that thehost factors are equally important in epidemiology of AAT
Journal of Parasitology Research 7
as higher trypanosome infection rates were recorded in Glongipennis than G pallidipes but the sex of the vector is lessimportant Further the study showed that apparent density ofthe vector is a good indicator of AAT transmission risk Thestudy therefore affirms the importance of designing controlstrategies targeting the biological vector (tsetse) mechanicalvectors (Tabanus and Stomoxys spp) and the trypanosomeparasites
6 Recommendations
(1) Since both T congolense and T vivax are pathogenicin cattle and other ruminants with the latter causingacute hemorrhagic syndrome [52] it would be impor-tant to conduct large scale integrated tsetse control inMtitoAndei by application of vector control strategiescoupled with curative treatment of livestock as theseanimals could act as source of reinfection to tsetse
(2) Surveillance studies to determine trypanosomeprevalence in livestock should be conducted in thisarea for effective trypanosomiasis control programs
(3) Entomological surveys should be conducted at dif-ferent seasons to understand seasonal dynamics ofthe vectors and the associated trypanosomiasis riskData on seasonal variations of potential vectors canbe integrated into epidemiologicalmodels to facilitatebetter understanding of the relative importance ofcyclical and mechanical vectors in Mtito Andei
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] N Kone E K NrsquoGoran I Sidibe A W Kombassere and JBouyer ldquoSpatio-temporal distribution of tsetse and other bitingflies in the Mouhoun River basin Burkina Fasordquo Medical andVeterinary Entomology vol 25 no 2 pp 156ndash168 2011
[2] G Cecchi M Paone R A Herrero M J B Vreysen and R CMattioli ldquoDeveloping a continental atlas of the distribution andtrypanosomal infection of tsetse flies (Glossina species)rdquo Para-sites amp Vectors vol 8 article 284 2015
[3] L J Morrison and A MacLeod ldquoAfrican trypanosomiasisrdquoParasite Immunology vol 33 no 8 pp 421ndash422 2011
[4] G Cecchi R C Mattioli J Slingenbergh and S De la RocqueldquoLand cover and tsetse fly distributions in sub-Saharan AfricardquoMedical and Veterinary Entomology vol 22 no 4 pp 364ndash3732008
[5] P Solano S J Torr and M J Lehane ldquoIs vector control neededto eliminate gambiense human African trypanosomiasisrdquoFrontiers in Cellular and Infection Microbiology vol 3 no 1article 33 2013
[6] A P M Shaw G Cecchi G RWWint R C Mattioli and T PRobinson ldquoMapping the economic benefits to livestock keepersfrom intervening against bovine trypanosomosis in EasternAfricardquo Preventive Veterinary Medicine vol 113 no 2 pp 197ndash210 2014
[7] J S Beadell C Hyseni P P Abila et al ldquoPhylogeography andpopulation structure of Glossina fuscipes fuscipes in Ugandaimplications for control of tsetserdquo PLoS Neglected TropicalDiseases vol 4 no 3 article e636 2010
[8] K Bardosh CWaiswa and S CWelburn ldquoConflict of interestuse of pyrethroids and amidines against tsetse and ticks inzoonotic sleeping sickness endemic areas of Ugandardquo Parasitesand Vectors vol 6 article 204 2013
[9] C Kubi J Van denAbbeele R DeDeken TMarcotty P Dornyand P Van Den Bossche ldquoThe effect of starvation on the sus-ceptibility of teneral and non-teneral tsetse flies to trypanosomeinfectionrdquoMedical andVeterinary Entomology vol 20 no 4 pp388ndash392 2006
[10] V Lutje J Seixas and A Kennedy ldquoChemotherapy for second-stage human African trypanosomiasisrdquo Cochrane Database ofSystematic Reviews vol 8 Article ID CD006201 2010
[11] M Bitew Y Amide T Zenebe and H Degefu ldquoTrypanosomeinfection rate inGlossina pallidipes andGlossina fuscipes fuscipesin Gojeb Valley Southwest Ethiopiardquo Global Veterinaria vol 6no 2 pp 131ndash135 2011
[12] R A Oluwafemi A A Ilemobade and E A O Laseinde ldquoTheimpact of African animal trypanosomiasis and tsetse on thelivelihood and well being of cattle and their owners in theBICOT study area ofNigeriardquo Scientific Research andEssays vol2 no 9 pp 380ndash383 2007
[13] R K Oniangrsquoo ldquoEnhancing peoplersquos nutritional status throughrevitalisation of Agriculture and related activitiesrdquo AfricanJournal of Food Agriculture Nutrition and Development vol 1no 1 pp 43ndash50 2001
[14] G L Mugunieri P Irungu and J M Omiti ldquoPerformance ofcommunity-based animal health workers in the delivery of live-stock health servicesrdquo Tropical Animal Health and Productionvol 36 no 6 pp 523ndash535 2004
[15] C A Spinage African EcologyThe Tsetse fly I Africarsquos Bane andBenefice Springer-Verlag Berlin Germany 2012
[16] P Solano J F Michel T Lefrancois et al ldquoPolymerase chainreaction as a diagnosis tool for detecting trypanosomes in nat-urally infected cattle in Burkina Fasordquo Veterinary Parasitologyvol 86 no 2 pp 95ndash103 1999
[17] T Tran G Napier T Rowan et al ldquoDevelopment and eval-uation of an ITS1 lsquoTouchdownrsquo PCR for assessment of drugefficacy against animal African trypanosomosisrdquo VeterinaryParasitology vol 202 no 3-4 pp 164ndash170 2014
[18] S M Thumbi F A McOdimba R O Mosi and J O JungrsquoaldquoComparative evaluation of three PCR base diagnostic assaysfor the detection of pathogenic trypanosomes in cattle bloodrdquoParasites and Vectors vol 1 article 46 2008
[19] A Cox A Tilley F McOdimba et al ldquoA PCR based assay fordetection and differentiation of African trypanosome species inbloodrdquoExperimental Parasitology vol 111 no 1 pp 24ndash29 2005
[20] L Brownhill E M Njuguna and M Miruka Strategy forMainstreaming Gender Analysis in the KARIMcGill UniversityProject Enhancing Ecologically Resilient Food Security ThroughInnovative Farming Systems in the Semi AridMidlands of KenyaKenya Agricultural Research Institute (KARI) McGill Univer-sity 3rd edition 2013
[21] M Thrusfield Veterinary Epidemiology Blackwell ScienceOxford UK 3rd edition 2005
[22] I Malele L Craske C Knight et al ldquoThe use of specific andgeneric primers to identify trypanosome infections of wildtsetse flies in Tanzania by PCRrdquo Infection Genetics and Evolu-tion vol 3 no 4 pp 271ndash279 2003
8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Journal of Parasitology Research 7
as higher trypanosome infection rates were recorded in Glongipennis than G pallidipes but the sex of the vector is lessimportant Further the study showed that apparent density ofthe vector is a good indicator of AAT transmission risk Thestudy therefore affirms the importance of designing controlstrategies targeting the biological vector (tsetse) mechanicalvectors (Tabanus and Stomoxys spp) and the trypanosomeparasites
6 Recommendations
(1) Since both T congolense and T vivax are pathogenicin cattle and other ruminants with the latter causingacute hemorrhagic syndrome [52] it would be impor-tant to conduct large scale integrated tsetse control inMtitoAndei by application of vector control strategiescoupled with curative treatment of livestock as theseanimals could act as source of reinfection to tsetse
(2) Surveillance studies to determine trypanosomeprevalence in livestock should be conducted in thisarea for effective trypanosomiasis control programs
(3) Entomological surveys should be conducted at dif-ferent seasons to understand seasonal dynamics ofthe vectors and the associated trypanosomiasis riskData on seasonal variations of potential vectors canbe integrated into epidemiologicalmodels to facilitatebetter understanding of the relative importance ofcyclical and mechanical vectors in Mtito Andei
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] N Kone E K NrsquoGoran I Sidibe A W Kombassere and JBouyer ldquoSpatio-temporal distribution of tsetse and other bitingflies in the Mouhoun River basin Burkina Fasordquo Medical andVeterinary Entomology vol 25 no 2 pp 156ndash168 2011
[2] G Cecchi M Paone R A Herrero M J B Vreysen and R CMattioli ldquoDeveloping a continental atlas of the distribution andtrypanosomal infection of tsetse flies (Glossina species)rdquo Para-sites amp Vectors vol 8 article 284 2015
[3] L J Morrison and A MacLeod ldquoAfrican trypanosomiasisrdquoParasite Immunology vol 33 no 8 pp 421ndash422 2011
[4] G Cecchi R C Mattioli J Slingenbergh and S De la RocqueldquoLand cover and tsetse fly distributions in sub-Saharan AfricardquoMedical and Veterinary Entomology vol 22 no 4 pp 364ndash3732008
[5] P Solano S J Torr and M J Lehane ldquoIs vector control neededto eliminate gambiense human African trypanosomiasisrdquoFrontiers in Cellular and Infection Microbiology vol 3 no 1article 33 2013
[6] A P M Shaw G Cecchi G RWWint R C Mattioli and T PRobinson ldquoMapping the economic benefits to livestock keepersfrom intervening against bovine trypanosomosis in EasternAfricardquo Preventive Veterinary Medicine vol 113 no 2 pp 197ndash210 2014
[7] J S Beadell C Hyseni P P Abila et al ldquoPhylogeography andpopulation structure of Glossina fuscipes fuscipes in Ugandaimplications for control of tsetserdquo PLoS Neglected TropicalDiseases vol 4 no 3 article e636 2010
[8] K Bardosh CWaiswa and S CWelburn ldquoConflict of interestuse of pyrethroids and amidines against tsetse and ticks inzoonotic sleeping sickness endemic areas of Ugandardquo Parasitesand Vectors vol 6 article 204 2013
[9] C Kubi J Van denAbbeele R DeDeken TMarcotty P Dornyand P Van Den Bossche ldquoThe effect of starvation on the sus-ceptibility of teneral and non-teneral tsetse flies to trypanosomeinfectionrdquoMedical andVeterinary Entomology vol 20 no 4 pp388ndash392 2006
[10] V Lutje J Seixas and A Kennedy ldquoChemotherapy for second-stage human African trypanosomiasisrdquo Cochrane Database ofSystematic Reviews vol 8 Article ID CD006201 2010
[11] M Bitew Y Amide T Zenebe and H Degefu ldquoTrypanosomeinfection rate inGlossina pallidipes andGlossina fuscipes fuscipesin Gojeb Valley Southwest Ethiopiardquo Global Veterinaria vol 6no 2 pp 131ndash135 2011
[12] R A Oluwafemi A A Ilemobade and E A O Laseinde ldquoTheimpact of African animal trypanosomiasis and tsetse on thelivelihood and well being of cattle and their owners in theBICOT study area ofNigeriardquo Scientific Research andEssays vol2 no 9 pp 380ndash383 2007
[13] R K Oniangrsquoo ldquoEnhancing peoplersquos nutritional status throughrevitalisation of Agriculture and related activitiesrdquo AfricanJournal of Food Agriculture Nutrition and Development vol 1no 1 pp 43ndash50 2001
[14] G L Mugunieri P Irungu and J M Omiti ldquoPerformance ofcommunity-based animal health workers in the delivery of live-stock health servicesrdquo Tropical Animal Health and Productionvol 36 no 6 pp 523ndash535 2004
[15] C A Spinage African EcologyThe Tsetse fly I Africarsquos Bane andBenefice Springer-Verlag Berlin Germany 2012
[16] P Solano J F Michel T Lefrancois et al ldquoPolymerase chainreaction as a diagnosis tool for detecting trypanosomes in nat-urally infected cattle in Burkina Fasordquo Veterinary Parasitologyvol 86 no 2 pp 95ndash103 1999
[17] T Tran G Napier T Rowan et al ldquoDevelopment and eval-uation of an ITS1 lsquoTouchdownrsquo PCR for assessment of drugefficacy against animal African trypanosomosisrdquo VeterinaryParasitology vol 202 no 3-4 pp 164ndash170 2014
[18] S M Thumbi F A McOdimba R O Mosi and J O JungrsquoaldquoComparative evaluation of three PCR base diagnostic assaysfor the detection of pathogenic trypanosomes in cattle bloodrdquoParasites and Vectors vol 1 article 46 2008
[19] A Cox A Tilley F McOdimba et al ldquoA PCR based assay fordetection and differentiation of African trypanosome species inbloodrdquoExperimental Parasitology vol 111 no 1 pp 24ndash29 2005
[20] L Brownhill E M Njuguna and M Miruka Strategy forMainstreaming Gender Analysis in the KARIMcGill UniversityProject Enhancing Ecologically Resilient Food Security ThroughInnovative Farming Systems in the Semi AridMidlands of KenyaKenya Agricultural Research Institute (KARI) McGill Univer-sity 3rd edition 2013
[21] M Thrusfield Veterinary Epidemiology Blackwell ScienceOxford UK 3rd edition 2005
[22] I Malele L Craske C Knight et al ldquoThe use of specific andgeneric primers to identify trypanosome infections of wildtsetse flies in Tanzania by PCRrdquo Infection Genetics and Evolu-tion vol 3 no 4 pp 271ndash279 2003
8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 Journal of Parasitology Research
[23] M Desta S Menkir and A Kebede ldquoThe study on tsetse fly(Glossina species) and their role in trypanosome infection ratein Birbir valley Baro-Akobo river system Western EthiopiardquoJournal of Veterinary Medicine and Animal Health vol 5 no 7pp 186ndash194 2013
[24] J R Franco P P Simarro A Diarra and J G Jannin ldquoEpide-miology of human African trypanosomiasisrdquo Clinical Epidemi-ology vol 6 no 1 pp 257ndash275 2014
[25] S J Childs ldquoA model of pupal water loss in Glossinardquo Mathe-matical Biosciences vol 221 no 2 pp 77ndash90 2009
[26] S J Childs ldquoA model of teneral dehydration in Glossinardquo ActaTropica vol 131 no 1 pp 79ndash91 2014
[27] J O Ouma J S Beadell C Hyseni et al ldquoGenetic diversity andpopulation structure of Glossina pallidipes in Uganda andwestern Kenyardquo Parasites and Vectors vol 4 article 122 2011
[28] E S Krafsur ldquoTsetse flies genetics evolution and role asvectorsrdquo Infection Genetics and Evolution vol 9 no 1 pp 124ndash141 2009
[29] T Barros and L Foil ldquoSeasonal occurrence and relative abun-dance of Tabanidae (Diptera) from the Pantanal region BrazilrdquoMemoirs on Entomology International vol 14 pp 387ndash3961999
[30] M Desquesnes F Biteau-Coroller J Bouyer M L Dia and LFoil ldquoDevelopment of a mathematical model for mechanicaltransmission of trypanosomes and other pathogens of cattletransmitted by tabanidsrdquo International Journal for Parasitologyvol 39 no 3 pp 333ndash346 2009
[31] A H Rahman ldquoObservations on the trypanosomosis problemoutside the tsetse belts of SudanrdquoRevue Scientifique et Technique(International Office of Epizootics) vol 24 no 3 pp 965ndash9722005
[32] W F Snow S A Tarimo C Staak and L Butler ldquoThe feedinghabits of the tsetse Glossina pallidipes Austen on the southKenya coast in the context of its host range and trypanosomeinfection rates in other parts of East Africardquo Acta Tropica vol45 no 4 pp 339ndash349 1988
[33] J Bouyer N Kone and Z Bengaly ldquoDynamics of tsetse naturalinfection rates in the Mouhoun river Burkina Faso in relationwith environmental factorsrdquo Frontiers in Cellular and InfectionMicrobiology vol 3 article 47 Article ID Article 47 2013
[34] S Aksoy ldquoControl of tsetse flies and trypanosomes usingmolecular geneticsrdquo Veterinary Parasitology vol 115 no 2 pp125ndash145 2003
[35] C Hu and S Aksoy ldquoInnate immune responses regulate try-panosome parasite infection of the tsetse fly Glossina morsitansmorsitansrdquoMolecularMicrobiology vol 60 no 5 pp 1194ndash12042006
[36] E T Macleod A C Darby I Maudlin and S C WelburnldquoFactors affecting trypanosomematuration in tsetse fliesrdquo PLoSONE vol 2 no 2 article e239 2007
[37] Z K Njiru J N Makumi S Okoth J M Ndungu and W CGibson ldquoIdentification of trypanosomes in Glossina pallidipesand G longipennis in Kenyardquo Infection Genetics and Evolutionvol 4 no 1 pp 29ndash35 2004
[38] C Hu R V M Rio J Medlock et al ldquoInfections with immuno-genic trypanosomes reduce tsetse reproductive fitness poten-tial impact of different parasite strains on vector populationstructurerdquo PLoS Neglected Tropical Diseases vol 2 no 3 articlee192 2008
[39] ZHao I Kasumba and S Aksoy ldquoProventriculus (cardia) playsa crucial role in immunity in tsetse fly (Diptera Glossinidiae)rdquo
Insect Biochemistry and Molecular Biology vol 33 no 11 pp1155ndash1164 2003
[40] D Nayduch and S Aksoy ldquoRefractoriness in Tsetse flies(Diptera Glossinidae) may be a matter of timingrdquo Journal ofMedical Entomology vol 44 no 4 pp 660ndash665 2007
[41] F NWamwiri U Alam P CThande et al ldquoWolbachia Sodalisand trypanosome co-infections in natural populations ofGlossina austeni and Glossina pallidipesrdquo Parasites and Vectorsvol 6 article 232 2013
[42] U Alam C Hyseni R E Symula et al ldquoImplications ofmicrofauna-host interactions for trypanosome transmissiondynamics in Glossina fuscipes fuscipes in Ugandardquo Applied andEnvironmentalMicrobiology vol 78 no 13 pp 4627ndash4637 2012
[43] R E Symula U Alam C Brelsfoard et al ldquoWolbachia associ-ation with the tsetse fly Glossina fuscipes fuscipes reveals highlevels of genetic diversity and complex evolutionary dynamicsrdquoBMC Evolutionary Biology vol 13 article 31 2013
[44] M E JWoolhouse K Bealby J JMcNamara and J SilutongweldquoTrypanosome infections of the tsetse fly Glossina pallidipes inthe Luangwa Valley Zambiardquo International Journal for Para-sitology vol 24 no 7 pp 987ndash993 1994
[45] P Solano L Argiro J M Reifenberg Y Yao and G DuvalletldquoField application of the polymerase chain reaction (PCR) tothe detection and characterization of trypanosomes inGlossinalongipalpis (Diptera Glossinidae) in Cote drsquoIvoirerdquo MolecularEcology vol 4 no 6 pp 781ndash786 1995
[46] P Van Den Bossche S Chitanga J Masumu T Marcotty andV Delespaux ldquoVirulence in Trypanosoma congolense Savannahsubgroup A comparison between strains and transmissioncyclesrdquo Parasite Immunology vol 33 no 8 pp 456ndash460 2011
[47] N A Dyer C Rose N O Ejeh and A Acosta-Serrano ldquoFlyingtryps survival and maturation of trypanosomes in tsetse fliesrdquoTrends in Parasitology vol 29 no 4 pp 188ndash196 2013
[48] I Roditi andM J Lehane ldquoInteractions between trypanosomesand tsetse fliesrdquo Current Opinion in Microbiology vol 11 no 4pp 345ndash351 2008
[49] S M Samdi A O Fajinmi J O Kalejaye et al ldquoPrevalenceof trypanosomosis in cattle at slaughter in Kaduna centralAbattoirrdquoAsian Journal of Animal Sciences vol 5 no 2 pp 162ndash165 2011
[50] S Mihok E Munyoki R A Brett et al ldquoTrypanosomiasis andthe conservation of black rhinoceros (Diceros bicornis) at theNgulia Rhino sanctuary Tsavo West National park KenyardquoAfrican Journal of Ecology vol 30 no 2 pp 103ndash115 2008
[51] M Zuk and K A McKean ldquoSex differences in parasite infect-ions patterns and processesrdquo International Journal for Parasitol-ogy vol 26 no 10 pp 1009ndash1024 1996
[52] J W Magona J Walubengo and J T Odimin ldquoAcute haem-orrhagic syndrome of bovine trypanosomosis in Ugandardquo ActaTropica vol 107 no 2 pp 186ndash191 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology



















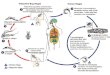







![Trypanosoma [1]](https://img.pdfslide.net/doc/110x75/58cedaba1a28abd4098b6285/trypanosoma-1.jpg)

