Embed Size (px)
Citation preview

SWANSEA UNIVERSITY
Environmental Dynamics and Climate Change
M.Sc. PROJECT
An Evaluation of the Climatic Signal within the
Earlywood Vessel Area of Quercus petraea (Matt.)
Liebl.
STUDENT NAME: Darren Davies
STUDENT NUMBER: 552192
SUPERVISOR: Dr Neil Loader
YEAR OF SUBMISSION: 2013
This work is part-funded by the European Social Fund (ESF) through the
European Union’s Convergence program administered by the Welsh
Government

I
Abstract
This paper investigated the climatic signals contained within earlywood vessels of 10
sessile oak trees (Quercus petraea (Matt.) Liebl.) from the UK, a first in the literature,
and their suitability for producing climate reconstructions. For the period of 1860 to
2012, 4 earlywood vessel area chronologies were constructed, along with a ring-width
record. The vessel chronologies were constructed through different combinations of the
largest annual earlywood vessels, allowing examination of how the climatic signals
were expressed. Identification of climatic influences was found by comparison with
monthly meteorological records. In contrast to the ring-width chronology, each vessel
chronology was statistically weaker in terms of common signal, and sensitivity.
However, earlywood vessels were less dependent on previous year growth. Earlywood
vessels were found to contain a different climatic signal to ring-widths, suggesting merit
in their use at this location. March relative humidity demonstrated the greatest influence
on earlywood vessel area, however, this signal was different between series, indicating
that different groupings of vessels may affect the expressed signal. Examination of the
suitability of the earlywood vessel chronologies to produce a March relative humidity
reconstruction demonstrated that chronologies composed of all earlywood vessels and
those with less than 5 were unsuitable. However, a chronology consisting of the average
of the 10 largest earlywood vessels was found to produce a stable reconstruction,
resulting in a March relative humidity reconstruction for the study period. Results
indicate promise in the use of earlywood vessels as a climatic proxy within the UK. As
a result of this study further research directions are suggested.
Key Words: Earlywood Vessels; Climate Proxy; Relative Humidity; United
Kingdom; Quercus petraea (Matt.) Liebl.

II
Declaration and Statements
This is being submitted in partial fulfilment of the requirements for the degree.
Sign:
Date:
This work has not previously been accepted in substance for any other degree and is not
being concurrently submitted in candidate for any degree.
Sign:
Date:
This dissertation is the result of my own independent work investigation, except where
otherwise stated. Other sources are acknowledged by footnotes giving explicit
references. A Bibliography is appended.
Signed:
Date:
I hereby give my consent for my dissertation, if relevant and accepted to be available for
photocopy, inter-library loan and for the title and summary to be made available to
outside organisations.
Signed:
Date:
Word Count: 6,681

III
Acknowledgements
The author would like to thank Dr Neil Loader for his input and advice throughout the
project, as well as staff at the National Botanic Gardens of Wales, especially Dr Angela
Singleton. In addition, the help afforded by Millie Watts, Siôn Carpenter and Ash
Woodward in collecting samples is much appreciated. Guidance provided by Prof.
Danny McCarroll aided with analysis. Thanks are also extended to Dr Neil Robertson
for his help. The author acknowledges the E-OBS dataset from the EU-FP6 project
ENSEMBLES (http://ensembles-eu.metoffice.com) and the data providers in the
ECA&D project (http://www.ecad.eu). The author is also appreciative of the funding
provided by the European Social Fund (ESF) through the European Union’s
Convergence program administered by the Welsh Government.

IV
Contents Page
Abstract I
Declaration and Statement II
Statement of Word Count II
Acknowledgements III
Table of Contents IV
List of Abbreviations V
List of Figures VI
List of Tables VIII
Section 1 – Introduction 1
Section 2 – Methodology 8
2.1 – Site location 8
2.2 – Sample Selection, Preparation and Measurement 8
2.3 – Chronology Construction and Climate-Growth Analysis 10
2.4 – Examination of Suitability for Climate Reconstruction 13
Section 3 – Results 14
3.1 – Vessel Characteristics 14
3.2 – Chronology Analysis 14
3.3 – Climate-Growth Relationships 17
3.4 – Climate Reconstruction 20
Section 4 – Discussion 21
Section 5 – Conclusion 30
References 32
Supplementary Information 39
Administrative Appendices 49
Appendix A – SOTEAS Risk Assessment 49
Appendix B – Meeting Log 53

V
List of Abbreviations
Coefficient of Efficiency CE
Estimated Sample Depth ESD
Expressed Population Signal EPS
First Order Autocorrelation AutoR
Mean Between Tree Correlation Rbt
Mean Sensitivity MS
National Botanic Gardens of Wales NBGW
Reduction of Error RE
Ring-Width RW

VI
List of Figures
Page Number
Figure 2.1 – Map of the National Botanic Gardens of Wales and
the Waun Las National Nature Reserve.
9
Figure 2.2 - Climate diagram for the National Botanic Gardens of
Wales. Data obtained from the E-OBS 0.25o gridded dataset (1961
– 1990; Haylock et al., 2008).
9
Figure 2.3 – Schematic representation of each of the earlywood
vessel chronologies outlined in Table 2.1. A) MAX, B) 5MAX, C)
10MAX and D) MEVA. Red represents the selected vessels, white
vessels not selected.
11
Figure 3.1 – A frequency histogram of earlywood vessel lumen
area. Note vessel area below 10,000 um2 was removed from
analysis.
15
Figure 3.2 – Earlywood vessel and ring-width index chronologies:
A) MAX, B) 5MAX, C) 10MAX, D) MEVA and E) RW for the
period 1860 - 2012. Grey lines represent individual series and red is
the average chronology. F) Illustrates the sample depth. Note
truncated axis.
16
Figure 3.3 – Climate-growth correlations between precipitation,
temperature, humidity (overleaf) and the constructed chronologies.
Blue = MAX; Red = 5MAX; Green = 10MAX; Purple = MEVA
and Orange = RW. Solid lines – p<0.05; Dashed lines – p<0.01.
18
Figure 3.4 – Calibration and verification results of the 10MAX
series with March relative humidity (%). Red line – Relative
Humidity; Blue line – relative humidity reconstruction.
22
Figure 3.5 – Reconstruction of past March relative humidity (%)
for the period 1860 – 2011 for the National Botanic Gardens of
Wales produced from the 10MAX series. The black line represents
the reconstruction, while the red line is the observed relative
humidity (see Jenkins et al., 2008). Grey lines represent the
uncertainty of the reconstruction utilizing Gaussian error
propagation (±95%).
24
Figure S1 – Examples of how sample preparation influenced the
accuracy of vessel measurements. Images on the left are scans of
the cores, while on the right are images of the WinCell
Classification where red represents a rejected classification and
green are areas classified as vessels. A) No enhancement of the
sample surface, B) Vessels in-filled with chalk and C) same as (B)
but core surface coloured black with permanent marker.
42
Figure S2 – Sample coverage. Note NBGW007 and 014 are 43

VII
missing years.
Figure S3 - Spatial field analysis of August temperatures for the
period of 1951-2012 against the A) MAX, B) 5MAX, C) 10MAX
and D) MEVA chronologies. Produced through KNMI Climate
Explorer (http://climexp.knmi.nl/).
44

VIII
List of Tables
Page Number
Table 2.1 – Description of the constructed earlywood vessel
chronologies.
11
Table 3.1. - Expressed population signal (EPS), Estimated sample
depth for an EPS of 0.85 (ESD), mean correlation between
standardized chronologies (Rbt), mean sensitivity (MS) and first
order autocorrelation coefficient (AutoR) for the common period
(1947 – 2012) for each Q. Petraea (Matt.) L. earlywood vessel
chronology.
15
Table 3.2 – Pearson’s correlations and in brackets r2 values
between earlywood vessel and the ring-width chronologies for the
study common period (1947 – 2012).
17
Table 3.3 – An overview of the verification statistics used to
analyse the relationship between relative humidity and each
constructed chronology (1961 – 2011). Forward model: Calibration
period 1986 – 2011, verification period 1961 – 1985. Reverse
model: Calibration period 1961 – 1985, verification period 1986 –
2011.
20
Table S1 – Tree sample meta-data.
45
Table S2 -Filters used to identify and control abnormal
classifications.
46

1
1. Introduction
Anthropogenic activities over the last two centuries have amplified average global
temperatures by an estimated 0.7 oC (Solomon et al., 2007). One of the consequences
that have been hypothesised is a rise in extreme weather events, such as; heavy
precipitation and drought (e.g. Salinger & Griffiths, 2001; Allan & Soden, 2008;
Coumou & Rahmstorf, 2012; Westra et al., 2013). Therefore, global warming will
negatively impact the human and natural environment in the future (e.g. increased
spread of disease and reduced biodiversity; see McMichael et al., 2006 and Bellard et
al., 2012). The climate models from which such conclusions have been drawn are
calibrated and validated through the use of climate observations (Sundberg et al., 2012).
The obvious source of this data is from instrumental stations; however, these records are
temporally and spatially restricted (Ruddiman, 2008. pp. 314). To overcome these
limitations climate proxies are utilized.
Found in a number of archives including; ice cores (e.g. Petit et al., 1999), lake
sediments (e.g. Fritz et al., 1991) and peat cores (e.g. Barber et al., 1994), proxies
contain an indirect record of past climate conditions. However, these proxies do not
have the advantages afforded by tree-rings (McCarroll & Loader, 2004). For example,
as annual rings can be precisely dated, long accurate chronologies can be constructed
(e.g. Friedrich et al., 2007) and through replication, measurements of confidence can be
produced (McCarroll & Loader, 2004; McCarroll et al., 2013). As a consequence, tree
based proxies have been the foundation of a number of significant climate
reconstructions (e.g. Mann et al., 1999; Esper et al., 2002; Moberg et al., 2005).
Tree-ring width measurements have traditionally been used as a proxy (Campelo
et al., 2010), while more recently earlywood and latewood width (e.g. Kalela-Brundin,
1999), stable isotope measurements (e.g. Burk & Stuiver, 1981) and latewood density

2
(e.g. Briffa et al., 1988) have also been utilized. However, one of the challenges of
producing tree-based proxy reconstructions, and with climate proxies in general, is the
production of intra-annual reconstructions (Campelo et al., 2010). Such investigations
have utilized frost- and double-ring incidences (e.g. Brunstein, 1996; Campelo et al.,
2007), however; these approaches are limited. Thus, a frontier in contemporary
dendroclimatology is the identification of high resolution proxies, with anatomical tree-
ring features being cited as a promising research avenue (García-González & Fonti,
2006).
Wood cell features (i.e. diameter and lumen area) have long been recognised to be
modified between locations and along climatic gradients (Fonti & García-González,
2004), and through producing time-series of these characteristics it is hypothesised that
environmental information can be obtained (Schweingruber, 2001; Fonti et al., 2009).
The environmental signal within wood cell features is thought to be related to water
quantities within the tree. As water levels rise, turgor pressure increases, which will
influence the cell size (Ray et al., 1972; Boyer, 1985; Eilmann et al., 2006). However,
constraints (i.e. cost and time) have impeded the production of accurate measurements
of such features, and thus producing chronologies (Eckstein, 2004). Therefore, the
science was slow to develop until the invent of semi- and fully-automatic image
analysis systems (Eckstein, 2004). Consequently, since the latter half of the twentieth
century, there have been a number of investigations relating cell characteristics to
environmental signals, especially the water-conducting elements of conifers (e.g.
Panyushkina et al., 2003; DeSoto et al., 2011). However, similar research with
angiosperm species is lacking (Fonti & García-González, 2004).
Historically, the study of hardwood vessels was considered a technique to
differentiate between wood species, and it was sometime before it was documented that

3
over extended periods, vessel characteristics evolved with environmental influences
(Carlquist, 1988; Eckstein, 2004). However, early studies established that vessel
features were modified following extreme climatic years (i.e. Knigge & Schulz, 1961)
and that such qualities were suitable for the production of annual time-series (i.e.
Eckstein et al., 1977).
Since these early investigations, a variety of vessel features have been examined
within ring- and semi-ring porous species for an environmental signal. Pumijumnong &
Park (1999) for instance, investigated average vessel area, vessel diameter, and vessel
density in Tectona grandis L., while Tardif & Conciatori (2006) looked at the total
vessel area, largest earlywood vessel area and number of vessels in Quercus alba L. and
Quercus rubra L. The majority of research has however, focused on annual mean vessel
area. St. George et al., (2002) for example, found that reduced annual average vessel
area, within Quercus macrocarpa, to be associated with incidents of flooding. While
Schume et al., (2004) investigated the relationship between ground water and mean
vessel area, and found a reduction in vessel size with a loss of ground water contact. In
recent years however, research has specifically examined the environmental signal
contained within vessels of the earlywood, in the hope that information can be gained
on past spring and early summer events.
Traditionally, earlywood features have been avoided due to difficulties in
identifying an environmental relationship (Fonti & García-González, 2004). However,
earlywood vessels have demonstrated such a signal. For example, evidence supports a
distinct flooding signal within the average earlywood vessel area of different oak
species, representing spring and early summer flooding events (e.g. Astrade & Bégin,
1997; St. Georege & Nielsen, 2000, 2003). The majority of focus, however, has been on
climatic signals. García-González & Eckstein (2003) look explicitly at earlywood

4
vessels for a climatic signal and were able to produce a time-series composed of annual
average earlywood vessel area and found a strong positive spring precipitation signal,
which was mirrored by Eilmann et al., (2006). Contrasting these conclusions, a number
of investigations have found that earlywood vessel area has instead increased with
drought conditions (e.g. Knigge & Schulz, 1961; Pumijumnong & Park, 1999;
Corcuera et al., 2004; García-González & Fonti, 2007; Fonti & García-González, 2008).
On the other hand, a temperature signal has also been documented (e.g. Fonti & García-
Gonzalez, 2004). Both Matisons et al., (2012) and Matisons & Brumelis (2012) found
that winter and spring temperatures in Latvian Q. Robur L. controlled earlywood vessel
area. However, the signal was found to have altered in the recent century to one
dominated by summer precipitation, which was attributed to local climate change.
Eilmann et al., (2006) provides a number of explanations that could account for these
differences, including; different species response to water availability, different site
conditions, the internal control of earlywood production (e.g. Sass & Eckstein, 1995)
and differences in previous year climatic conditions.
Although there are currently difficulties in determining a common vessel response
to climatic conditions between locations and species, research has continued, and it
appears that potentially many of the early studies that took an overall vessel area
average have suffered from a mixing of climatic signals. Vessels are produced through
the growing season, thus are influenced by differing environmental conditions (García-
González & Fonti, 2006), in addition there is hypothesised to be a fixed period in which
vessels can be affected by environmental stimuli, namely, the time between cell
differentiation and the production of the secondary cell wall (Fonti & García-González,
2004). García-González & Fonti (2006) demonstrated that vessels produced at different
times contained contrasting climatic signals. For instance, the authors found that the

5
largest vessels were correlated with temperatures in March, while the smallest were
with June, and by computing an average of all earlywood vessels the correlations were
weakened. However, examining how climate signal strength responded to different
combinations of earlywood vessel areas, García-González & Eckstein (2003) found that
by removing a small proportion of the smallest vessels, the signal was weakened, and
there was no evidence of any improvement by retaining just the largest individuals.
Although the research considered to this point supports the use of vessel area
chronologies for climate reconstructions, some have concluded differently. Fonti et al.,
(2009) investigated the change in climate signal over different frequencies with a vessel
chronology of 446 years, the longest produced to date. By considering the climate signal
at low-, medium- and high-frequencies it was found that the signal was not consistent
throughout the vessel area chronology, where at high frequencies there was a negative
spring response, in contrast to an indirect positive response in the lower frequencies.
Such a response is problematic due to the contrasting signs of the relationship (Fonti et
al, 2009). In addition, an assumption of detrending is that there is a stable climate
response throughout the series, across different frequencies (Esper et al., 2005, 2009;
Fonti et al., 2009). However, when considering this conclusion it is also important to
acknowledge that at this site the authors also reported a similar result with a tree-ring
width chronology. Tree-ring width chronologies have been successfully utilized in
climatic reconstructions from other locations, thus it should be considered that this
result could be location based. For average earlywood vessel area to be considered a
‘useful’ proxy it has been said that the climatic signal contained should be different to
that found in easier to measure features, such as traditional ring-widths (Fonti & García-
González, 2004), a common finding in the literature (e.g. Pumijumnong & Park, 1999;
Fonti & García-González, 2004; Matisons & Brumelis, 2012). In contrast, Tardif &

6
Conciatori (2006) reported that for Q. alba L. and Quercus rubra L. that the average
earlywood vessel area produced a similar, but weaker relationship to growing season
conditions than a constructed ring-width chronology. Thus, at that study location the
production of a vessel chronology provided no benefit. In addition, the suitability of
vessels to produce chronologies should be questioned, as it is common to find that the
statistical quality of vessel chronologies is poor compared to ring-width chronologies.
For example, Fontí & García-González (2008) who compared ring-, latewood- and
earlywood-width to mean vessel area at three locations, found that the common
variability, signal and mean sensitivity was reduced in the vessel chronologies
compared to each width measurement chronology. This theme is repeated across the
literature (e.g. García-González & Eckstein, 2003; Fonti & García-González, 2004;
Tardiff & Conciatori, 2006; Fonti & García-González, 2008; Campelo et al., 2010).
However, it has been argued that the relationship between the proxy and the climate is
more important than the relationship between individual series (e.g. Fonti & García-
González, 2008). Although, this view could be seen as contradictory, as a reduced
common signal between trees will only result in a weakened and blurred signal that is
expressed when chronologies are combined.
From reviewing the literature it is clear that this particular field is still in its
infancy, and there are still many gaps. One of the main focal points that needs further
consideration is the relationship between vessel development and environmental
stimuli. In addition, there is a need to expand on research locations as current studies are
limited in specific areas, with currently none in the UK. To date, the majority of
published research has produced only short vessel chronologies which increases the
chance of spurious relationships with climatic records (e.g. 25 years; García-González
& Fonti, 2007, 22 years; Alla & Camarero, 2012). Thus, longer chronologies need to be

7
produced. Furthermore, the relationship between vessel chronologies and the climate to
date have only been examined with Pearson’s correlations and there is much to be
gained through using statistics such as the Reduction of Error (RE) and Coefficient of
Efficiency (CE; National Research Council, 2007). Such statistics are commonly used
to validate climate reconstructions, and will provide a better illustration of the suitability
of vessel chronologies for such an investigation. Thus, this project aims to:
1. Evaluate the quality of time-series built from average earlywood vessel area,
2. Evaluate, for the first time, the climatic signal contained within earlywood
vessels in the UK by producing an extended average earlywood vessel area
chronology and comparing it to climatic data,
3. Find how different combinations of earlywood vessel sizes affect the expression
of a climate signal,
4. Evaluate the suitability of vessel chronologies for climatic reconstructions in the
UK.

8
2. Methodology
2.1 – Site Location
Samples were collected from within the boundaries of the National Botanic
Gardens of Wales (NBGW) and the adjacent Waun Las National Nature Reserve
(51o50’ N, 04
o08’ W; 87 m a.s.l.; Figure 2.1). According to the National Vegetation
Classification System (Rodwell, 1991, 1992), the vegetation is dominated by semi-
improved grassland and dense scrub woodland (NBGW, 2013). Being a public
attraction there is evidence of minor management of the larger vegetation within the
NBGW limits, while it is minimal in the nature reserve. Average annual temperature is
9.8oC, while total average precipitation is 1365mm, with the majority falling over the
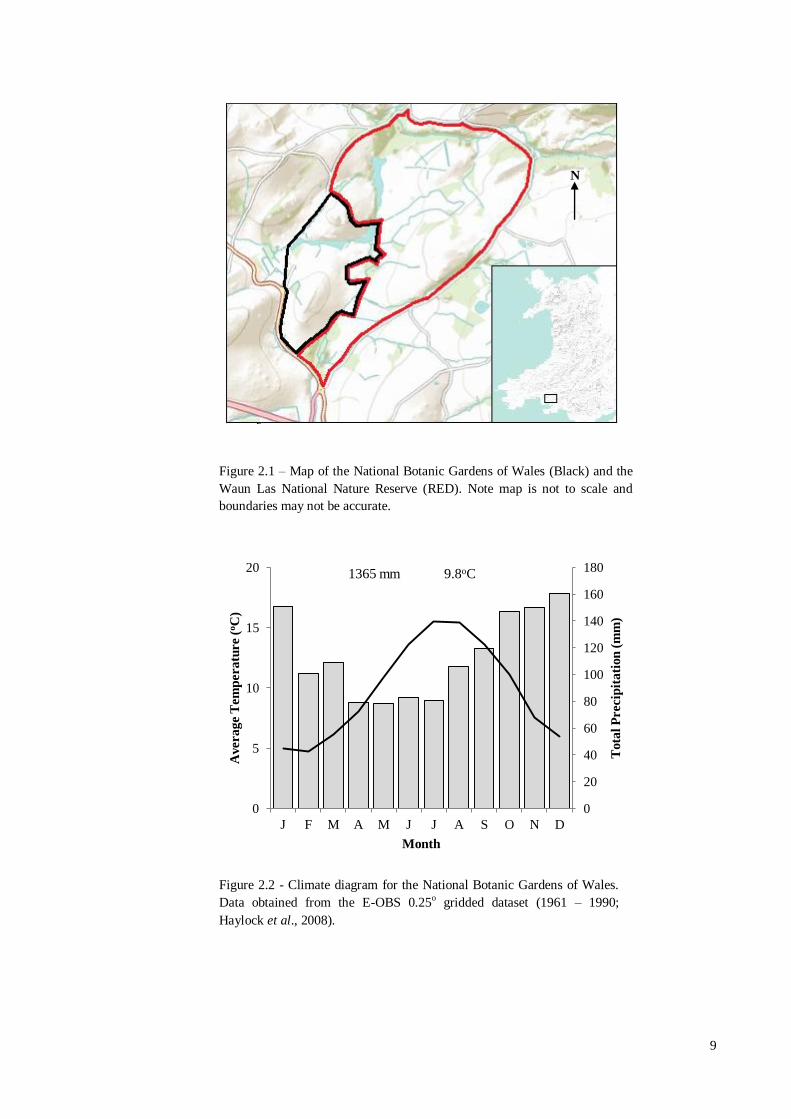
autumn and winter months (1961 – 1990; Figure 2.2) (Haylock et al., 2008).
2.2 – Sample Selection, Preparation and Measurement
Based on physical appearance (i.e. no fallen limbs, no evidence of rot) 10 sessile
oak trees (Quercus petraea (Matt.) Liebl.) were identified for investigation (Table S1).
To ensure a representative sample of earlywood vessels, dual 5-mm cores were obtained
at breast height (1.3 m) from each individual (García-González & Fonti, 2007). To
minimize the risk of reaction wood, cores were removed perpendicular to the
topographic slope (Fonti et al., 2009; Speer, 2010, p.78). Samples were left to air-dry;
before the transversal surface was progressively prepared with sandpaper (P80, P400
and P600 grades). Prior analysis, vessel lumina were cleared with compressed air, while
the wood matrix was coloured black and vessels in-filled with chalk, improving the
contrast for image analysis (e.g. Alla & Camarero, 2012) (see supplementary method).
Ring-widths were visually measured through use of a binocular microscope
(Nikon SMZ645), a Velmex positioning table and the software TSAP (Rinn, 2003),
9
0
20
40
60
80
100
120
140
160
180
0
5
10
15
20
J F M A M J J A S O N D
Tota
l P
recip
ita
tion
(m
m)
Avera
ge T
em
pera
ture (
oC
)
Month
1365 mm 9.8oC
Figure 2.1 – Map of the National Botanic Gardens of Wales (Black) and the
Waun Las National Nature Reserve (RED). Note map is not to scale and
boundaries may not be accurate.
Figure 2.2 - Climate diagram for the National Botanic Gardens of Wales.
Data obtained from the E-OBS 0.25o gridded dataset (1961 – 1990;
Haylock et al., 2008).
N

10
which also produced the master tree-ring chronology (see supplementary information).
Digital images of each core were captured using an EPSON Perfection V750 scanner at
6400 dpi resolution while, vessel analysis was conducted through the WinCELL PRO
(ver. 2013) software (Régents Instruments Inc., Québec, Canada). To ensure the
measurement of earlywood vessels a minimum size filter was applied at 10,000 μm2
(e.g. Fonti & García-González, 2008), while a suite of filters were used to avoid
inaccurate classification of ring features (Table S2).
To verify the accuracy of the chronology dating, independent ring-width
measurements were produced by the WinCELL PRO software and compared to the
original measurements. Due to inaccurate classifications by the image analysis software,
visual inspection of vessel classifications was also conducted and anomalous results
were removed (i.e. combined vessels, remaining debris).
2.3 – Chronology Construction and Climate-Growth Analysis
To examine the effect of selecting different size groups of earlywood vessels on
the climatic signal expressed, four earlywood vessel series were created (Table 2.1;
Figure 2.3), by pooling each dual core together (e.g. García-González & Eckstein,
2003), in addition to an average ring-width chronology. To compute these variables a
purposely designed computer package was created (see supplementary method).
To remove non-climatic, and preserve high frequency signals (Fonti et al., 2007),
a cubic smoothing spline of 32-year stiffness and 50% cut-off was utilized (e.g. Fonti &
García-González, 2004, 2008; Campelo et al., 2010), using the R (R Core Team, 2013)
package DetrendeR (Campelo, 2012). Detrending was applied to all series variables
individually, preventing differences arising due to utilizing contrasting methods
(García-González & Eckstein, 2003). Site chronologies were then computed by

11
Table 2.1 – Description of the constructed earlywood vessel chronologies.
Chronology Description
MAX Composed of the largest vessel from each annual ring-width.
5MAX The five largest vessels were selected from each annual
earlywood and averaged together.
10MAX The ten largest vessels were selected from each annual
earlywood and averaged together.
MEVA The average of all earlywood vessel areas from each year
Year
Figure 2.3 – Schematic representation of each of the earlywood vessel chronologies outlined
in Table 2.1. A) MAX, B) 5MAX, C) 10MAX and D) MEVA. Red represents the selected
vessels, white vessels not selected.
1921 1922 1923 1921 1922 1923
1921 1922 1923 1921 1922 1923
A B
C D

12
averaging all individual series together (e.g. Fonti & García-González, 2008). A
common period between 1947 and 1993 was produced as two trees had single core
coverage from 1993 to 2012 (Figure S2). However, the decision was made to include
these cores to increase coverage to the present, producing a common period between
1947 and 2012 (n = 65).
To review the statistical quality of the chronologies over the studies common
period, a number of statistics were selected which are common in dendroclimatology as
follows (e.g. Campelo et al., 2010); the mean between tree correlation (Rbt), Expressed
Population Signal (EPS; Wigley et al., 1984), mean sensitivity and first-order
autocorrelation. The Rbt allows for the evaluation of the series cohesion (Campelo et
al., 2010), while the EPS provides a measurement of the common variability within a
chronology (Wigley et al., 1984; Speer, 2010, p.109). It is possible to use the EPS to
determine if a chronology is being dominated by individuals, usually when the EPS is
below 0.85 (Speer, 2010, p.109). Where the EPS was found to be less than 0.85, an
estimate of the sample depth (ESD) was provided through the equation (Kiss et al.,
2011):
where EPS (x) is the desired EPS value (0.85; Speer, 2010, p.109) and Rbt is the mean
between tree correlation of the sample series. The mean sensitivity was selected to
measure the year-to-year variability in measured vessel size and ring-width (Fonti &
García-González, 2004; Speer, 2010, p.107), while the first-order autocorrelation
provides an indication of the influence of growth preceding each annual ring (Fritz,
1976, p. 259).

13
Pearson’s correlations were used to investigate climate-growth relationships with
monthly and seasonal data over the study common period (1947 – 2012) (e.g. Fonti &
García-González, 2004, 2008; Campelo et al., 2010).Precipitation and temperature data
were obtained from the E-OBS 0.25o gridded datasets (Haylock et al., 2008), while
relative humidity data was acquired from the 5km observed UK Climate Dataset
(Jenkins et al., 2008). Gridded data was selected as station coverage was poor in the
region.
2.4 – Examining Chronology Suitability to Climate Reconstructions
Where the strongest climate signal was identified, and there was a physiological
explanation for its influence, a climatic model was built through reverse linear
regression (Speer, 2010, p.178.), providing the opportunity for a number of verification
statistics to be conducted, as recommended by the National Research Council (2007).
Climate models were split into a calibration and verification period where the Reduction
of Error (RE) and Coefficient of Efficiency (CE) could be calculated. The RE statistic
compares the reconstruction to the mean of the calibration period, while the CE
compares the reconstruction to the mean of the independent verification period, and is a
harder statistic to satisfy (Kiss et al., 2011). The Pearson’s correlation coefficient (r2)
examined how much variation in the climate record could be explained by the
reconstruction. By switching the calibration and verification period it is possible to
produce a seconded set of statistics providing a comprehensive evaluation of the models
skill.

14
3. Results
3.1 – Vessel Characteristics
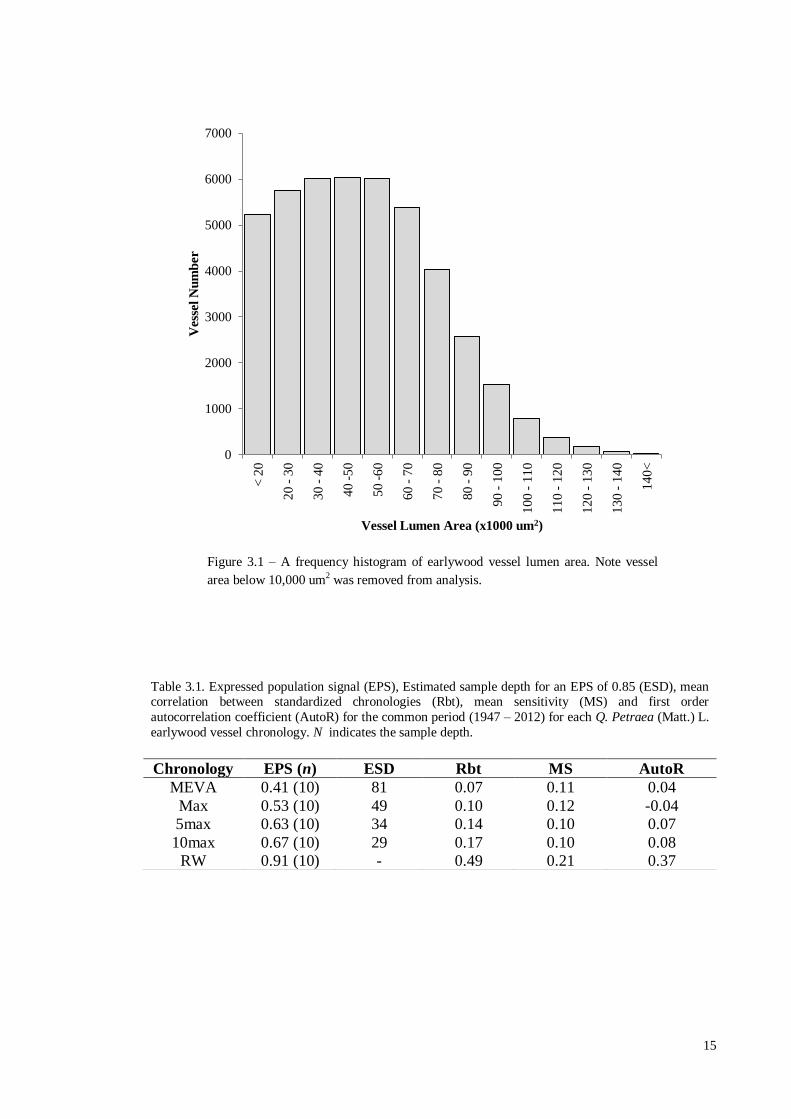
In total 43,950 earlywood vessels were measured, and it can be seen in Figure 3.1
that the frequency distribution of vessel area was skewed towards the smaller vessel
sizes. Vessel area ranged between 10,001 μm2 and 164,266 μm
2 with a mean and
median of 50,130 μm2 and
48,245 μm
2 respectively.
3.2 – Chronology Analysis
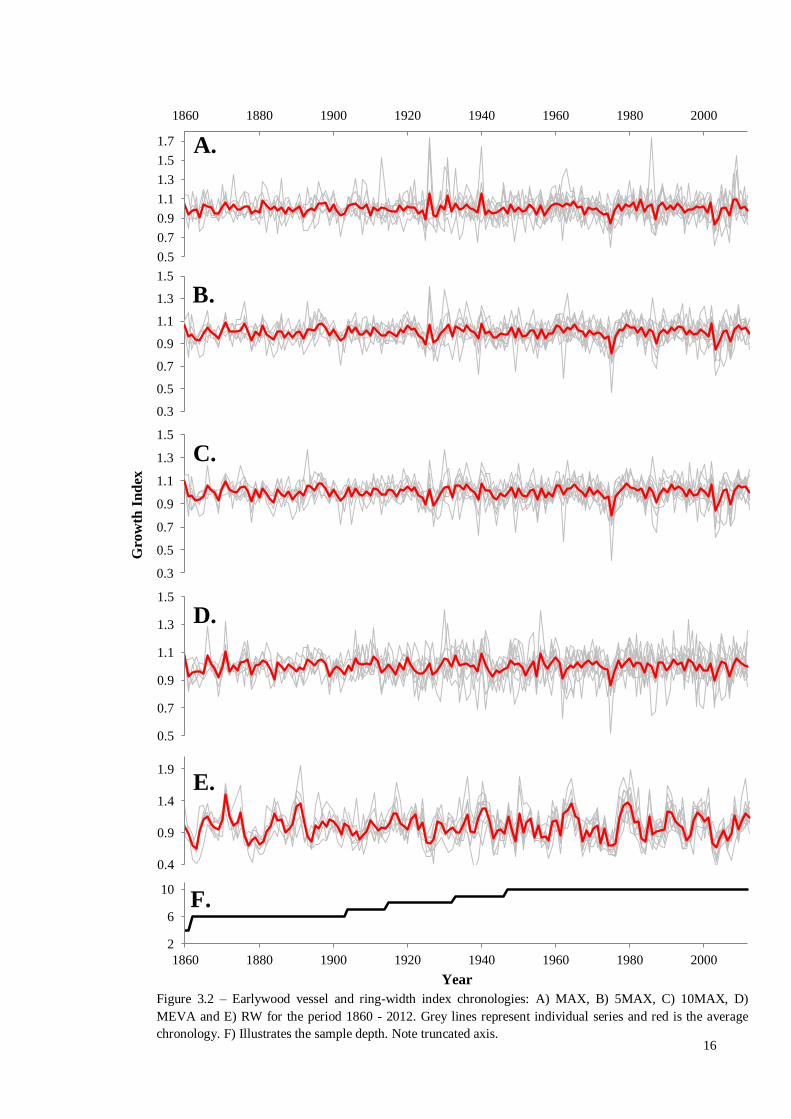
Figure 3.2 contains the five constructed time-series, while table 3.1 contains an
overview of the statistics used to assess the quality of each chronology. Ring-width
contained a greater common signal than each of the vessel chronologies. 10MAX
showed the greatest cohesion in its common signal, while in contrast MEVA
demonstrated the least. This was reflected in the ESD where 81 trees are estimated to be
required to bring the EPS to 0.85, while only 29 were required for 10MAX. Earlywood
vessels demonstrated consistently low year-to-year variability, in contrast to ring-
widths. Similarly, first order auto-regression is near absent in the vessel chronologies,
unlike ring-widths.
Pearson’s correlations revealed that RW was significantly correlated with MAX,
5MAX and 10MAX (Table 3.2). The 10MAX chronology was the most correlated
where 20% of the variation in vessel area was explained by ring width, while MEVA
demonstrated no significant correlation (Table 3.2).
Of particular interest in the vessel chronologies is the year 1975, where the
MEVA, 5MAX AND 10MAX demonstrated the smallest average vessel area (Figure
3.2 B-D). However, within the MAX (Figure 3.2 A) and RW (Figure 3.2 E)
chronologies, 1824 and 1827 were the lowest respectively.
15
0
1000
2000
3000
4000
5000
6000
7000
< 2
0
20 -
30
30 -
40
40
-5
0
50
-6
0
60
- 7
0
70
- 8
0
80
- 9
0
90
- 1
00
10
0 -
11
0
11
0 -
12
0
12
0 -
13
0
13
0 -
14
0
14
0<
Vess
el N
um
ber
Vessel Lumen Area (x1000 um2)
Table 3.1. Expressed population signal (EPS), Estimated sample depth for an EPS of 0.85 (ESD), mean correlation between standardized chronologies (Rbt), mean sensitivity (MS) and first order
autocorrelation coefficient (AutoR) for the common period (1947 – 2012) for each Q. Petraea (Matt.) L.
earlywood vessel chronology. N indicates the sample depth.
Chronology EPS (n) ESD Rbt MS AutoR
MEVA 0.41 (10) 81 0.07 0.11 0.04
Max 0.53 (10) 49 0.10 0.12 -0.04
5max 0.63 (10) 34 0.14 0.10 0.07
10max 0.67 (10) 29 0.17 0.10 0.08
RW 0.91 (10) - 0.49 0.21 0.37
Figure 3.1 – A frequency histogram of earlywood vessel lumen area. Note vessel
area below 10,000 um2 was removed from analysis.
16
0.5
0.7
0.9
1.1
1.3
1.5
1.7
1860 1880 1900 1920 1940 1960 1980 2000
A.
0.3
0.5
0.7
0.9
1.1
1.3
1.5
B.
0.3
0.5
0.7
0.9
1.1
1.3
1.5
C.
0.5
0.7
0.9
1.1
1.3
1.5
D.
0.4
0.9
1.4
1.9
E.
2
6
10
1860 1880 1900 1920 1940 1960 1980 2000
F.
Gro
wth
In
dex
Year
Figure 3.2 – Earlywood vessel and ring-width index chronologies: A) MAX, B) 5MAX, C) 10MAX, D)
MEVA and E) RW for the period 1860 - 2012. Grey lines represent individual series and red is the average
chronology. F) Illustrates the sample depth. Note truncated axis.

17
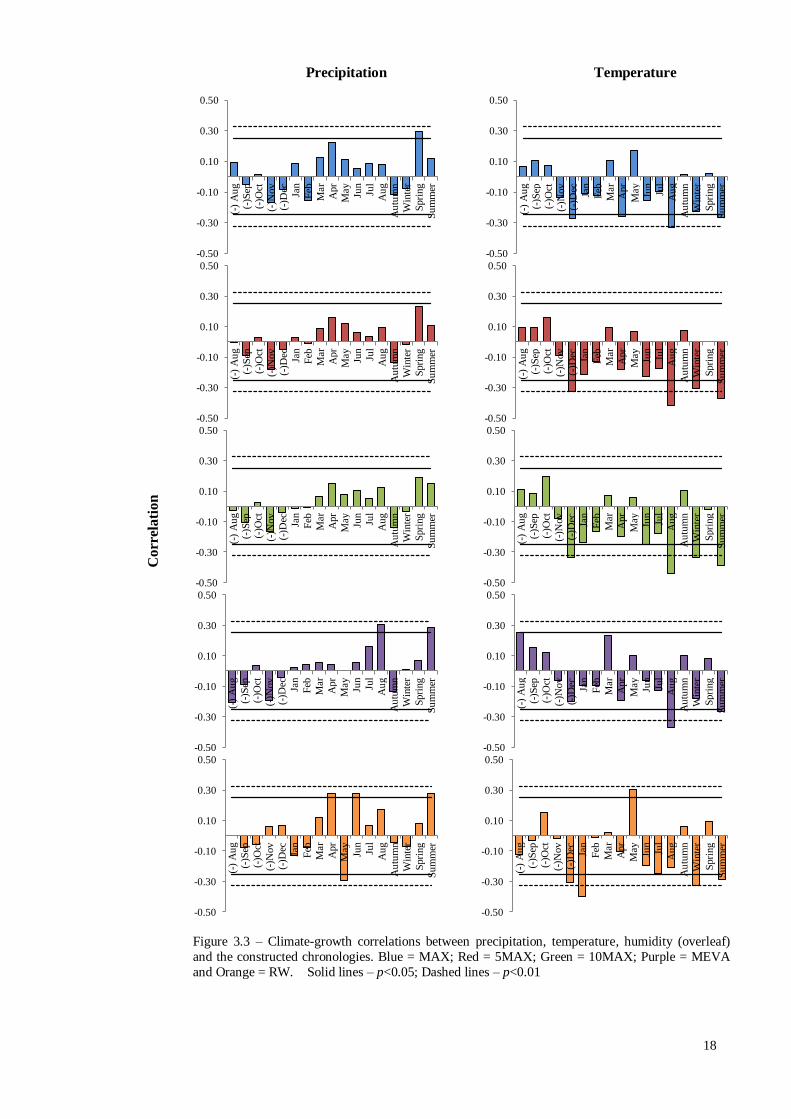
3.3 – Climate-Growth Relationships
Figure 3.3 provides an overview of the climate-growth correlations of
precipitation and temperature (1951 – 2012, n = 61), as well as humidity (1961 – 2011,
n = 50). Precipitation was weakly correlated with each vessel chronology throughout the
year, but the majority were non-significant. The highest vessel / precipitation
correlations were seen between MEVA and August as well as MAX and spring (March,
April and May), with both demonstrating a Pearson’s correlation of 0.30 (p<0.05). RW
demonstrated a weak spring precipitation signal (r = 0.08, p>0.05) due to a contrasting
relationship with April (r = 0.28, p<0.05) and May (r = -0.29, p<0.05). June saw a
return of a positive correlation (r = 0.27, p<0.05), which was reflected in the summer
(June, July & August) relationship (r = 0.28, p<0.05). Annual temperatures were on the
whole negatively correlated with earlywood vessel area. The strongest temperature
correlation was between August and 10MAX (r = -0.44, p<0.01), however the
relationship, although weaker, was evident in MAX, 5MAX and MEVA (Figure S3).
Vessel area was also significantly correlated with previous December’s temperature
which was most evident in 10MAX (r = -0.34, p<0.01). However, the relationship was
much weaker in MEVA (r = -0.20, p>0.05). RW’s strongest relationship was with
January temperatures (r = -0.40, p<0.01). In addition, there was a significant correlation
with May temperature (r = 0.30, p<0.05), while the relationship seen between vessel
area and August was weaker in the RW chronology (r = -0.21, p>0.05). The strongest
correlation with the climate data and the vessel chronologies was between March
Table 3.2 – Pearson’s correlations and in brackets r2 values between earlywood vessel and the ring-
width chronologies for the study common period (1947 – 2012).
Chronology MAX ( r2) 5MAX( r
2) 10MAX( r
2) MEVA( r
2)
RW 0.39**(0.15) 0.43**(0.18) 0.45**(0.20) 0.16(0.03) ** correlation significant at 0.01
18
Figure 3.3 – Climate-growth correlations between precipitation, temperature, humidity (overleaf)
and the constructed chronologies. Blue = MAX; Red = 5MAX; Green = 10MAX; Purple = MEVA
and Orange = RW. Solid lines – p<0.05; Dashed lines – p<0.01
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
Precipitation Temperature
Corr
elati
on
19
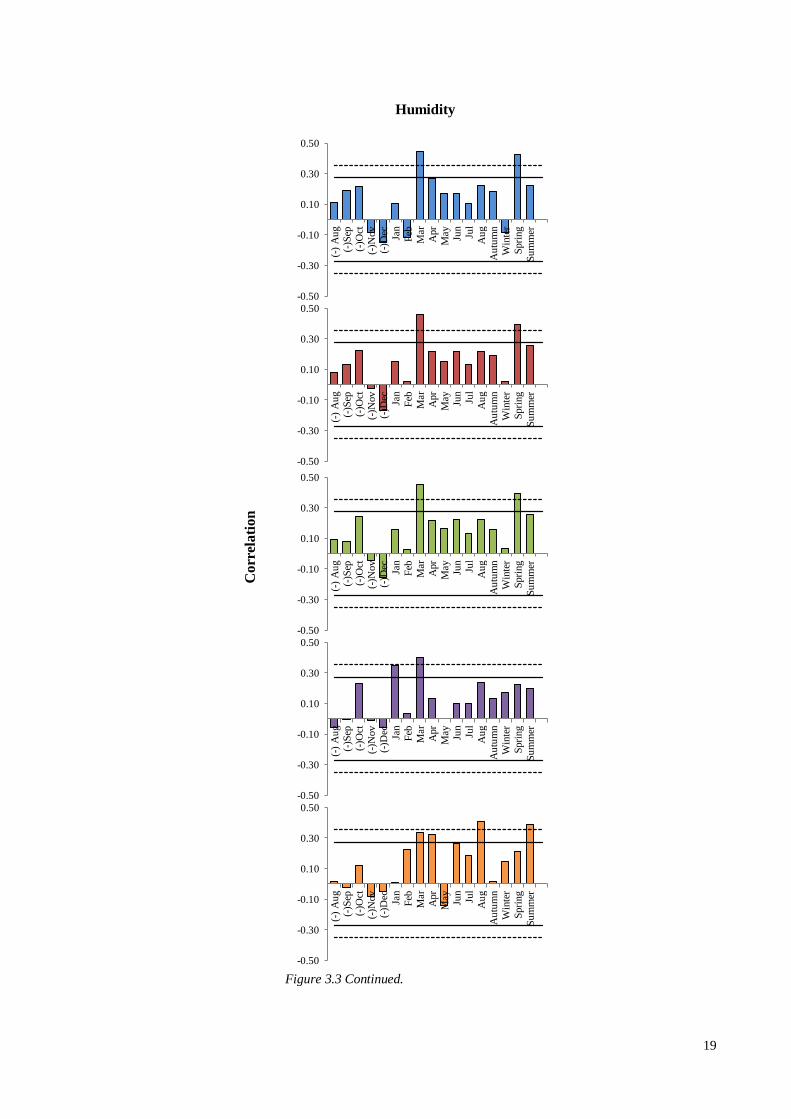
Figure 3.3 Continued.
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
-0.50
-0.30
-0.10
0.10
0.30
0.50
(-)
Au
g
(-)S
ep
(-)O
ct
(-)N
ov
(-)D
ec
Jan
Feb
Mar
Apr
May
Jun
Jul
Au
g
Au
tum
n
Win
ter
Spri
ng
Su
mm
er
Humidity
Corr
elati
on
20
relative humidity and 5MAX (r = 0.46, p<0.01), and although weaker, was present in
each vessel chronology (Figure 3.3). This trend was reflected in the significant
correlation with spring relative humidity (apart from MEVA). RW also demonstrated a
significant correlation with March relative humidity, however this was weaker than the
vessel chronologies (r = 0.34, p<0.05). RW’s strongest correlation with relative
humidity was seen in August (r = 0.40, p<0.01), which was reflected by the correlation
with average summer relative humidity (r = 0.41, p<0.01).
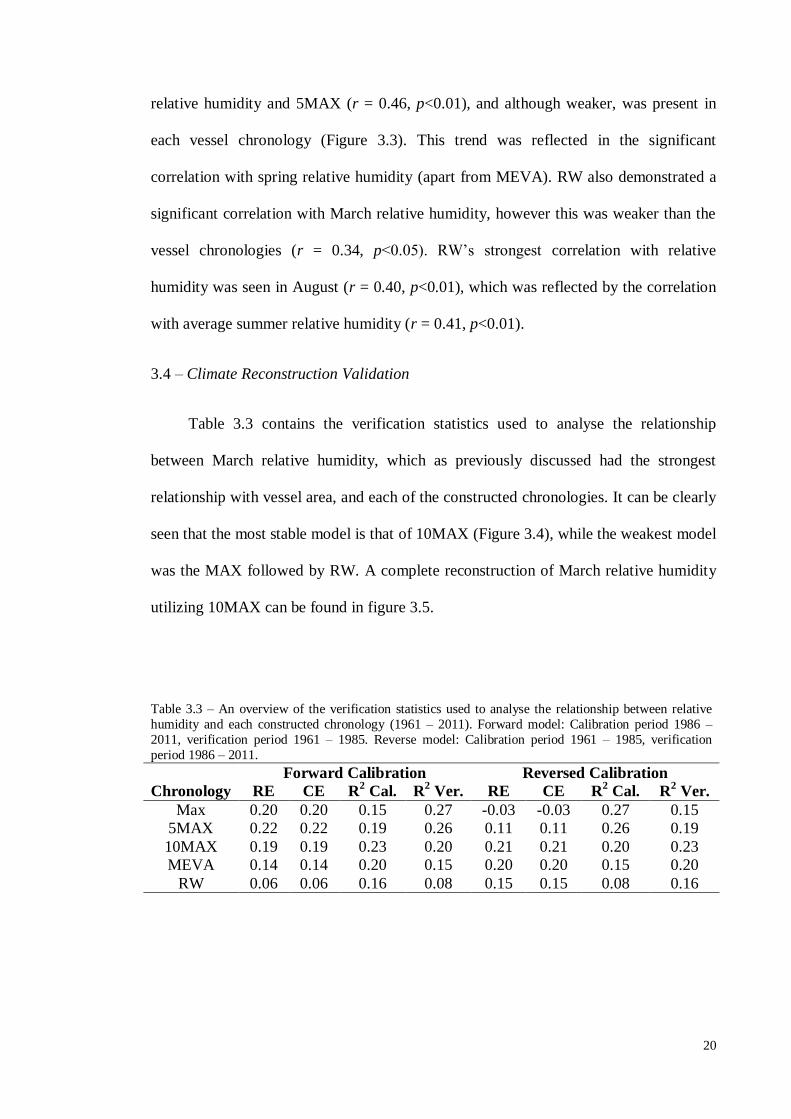
3.4 – Climate Reconstruction Validation
Table 3.3 contains the verification statistics used to analyse the relationship
between March relative humidity, which as previously discussed had the strongest
relationship with vessel area, and each of the constructed chronologies. It can be clearly
seen that the most stable model is that of 10MAX (Figure 3.4), while the weakest model
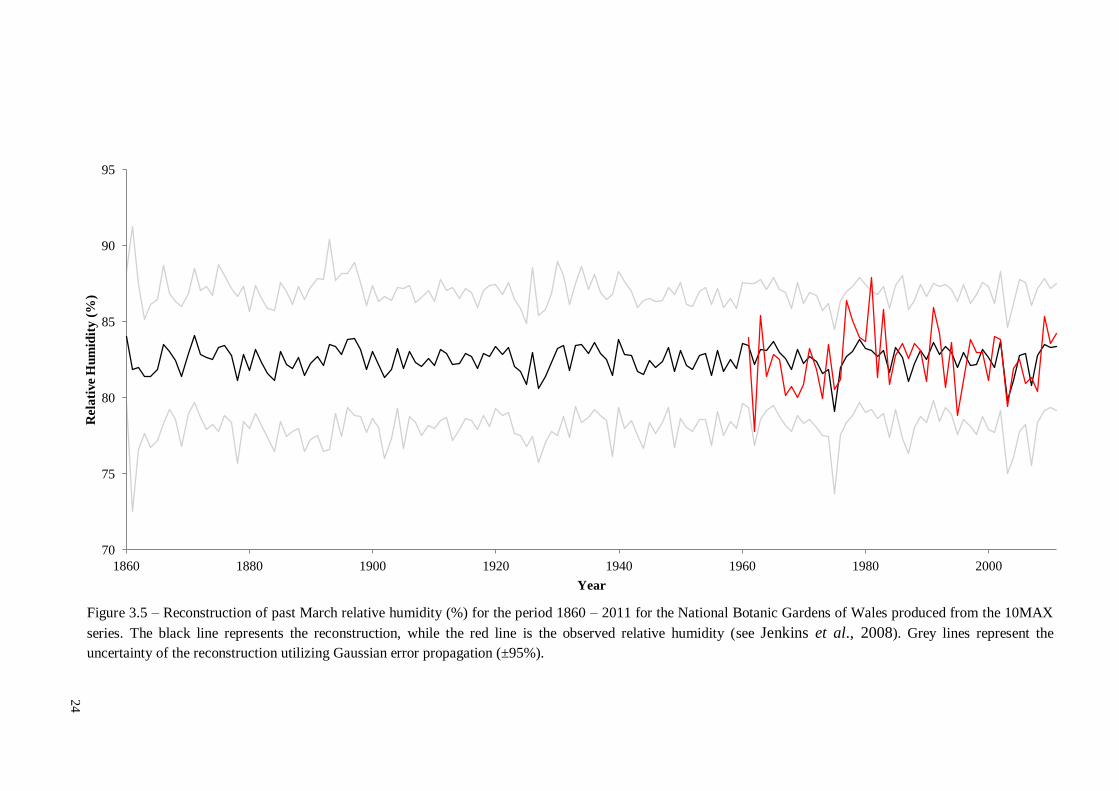
was the MAX followed by RW. A complete reconstruction of March relative humidity
utilizing 10MAX can be found in figure 3.5.
Table 3.3 – An overview of the verification statistics used to analyse the relationship between relative
humidity and each constructed chronology (1961 – 2011). Forward model: Calibration period 1986 –
2011, verification period 1961 – 1985. Reverse model: Calibration period 1961 – 1985, verification
period 1986 – 2011.
Forward Calibration Reversed Calibration
Chronology RE CE R2 Cal. R
2 Ver. RE CE R
2 Cal. R
2 Ver.
Max 0.20 0.20 0.15 0.27 -0.03 -0.03 0.27 0.15
5MAX 0.22 0.22 0.19 0.26 0.11 0.11 0.26 0.19
10MAX 0.19 0.19 0.23 0.20 0.21 0.21 0.20 0.23
MEVA 0.14 0.14 0.20 0.15 0.20 0.20 0.15 0.20
RW 0.06 0.06 0.16 0.08 0.15 0.15 0.08 0.16

21
4. Discussion
For a successful and accurate climate reconstruction to be produced from tree
based proxies, it is imperative that there is a strong common signal between individuals
(Fonti & García-González, 2008). Ring-width series clearly demonstrated shared
variability, while vessel chronologies were weaker. In fact, this is recurrent within the
literature (e.g. García-González & Eckstein, 2003; Fonti & García-González, 2004;
Tardiff & Conciatori, 2006; Fonti & García-González, 2008; Campelo et al., 2010).
Explanations are speculative, however, it has been suggested that earlywood vessels are
more strongly influenced by internal biological controls than external factors
(Woodcock, 1989; Fonti & García-González, 2004; Eilmanm et al., 2006). In fact, this
is reflected by the difference between the RW and vessel chronologies mean sensitivity,
where the vessel chronologies could be considered as complacent (Speer, 2010, p.107.).
The fact that there appears to be an internal control of earlywood vessel area is not
surprising. It has been found that within Q. petraea (Matt.) L., frost induced embolism
each winter, reduces conductivity within the earlywood vessels that were formed that
spring (Hacke & Sauter, 1996; Fonti et al., 2009). Earlywood vessels are important in
ring-porous oaks, as they have been found to provide up to 95% of annual hydraulic
conductivity (Cocuera et al., 2006; Alla & Camarero, 2012). Thus each spring, prior
bud-burst and leaf development, there must be an emphasis within ring-porous species
to produce large earlywood vessels to ensure sufficient hydraulic conductivity (Breda &
Granier, 1996; Fonti et al., 2009; Alla & Camarero, 2012). However, it is likely that
there is also a biological constraint on the maximum size of vessel area, as larger vessels
are thought to be more prone to embolism (Ewers, 1985; Kongoh et al., 2006).
Consequently, it is hypothesised that the reduced common signal and mean sensitivity
compared to RW, is due to a biologically controlled constraint on vessel size.
22
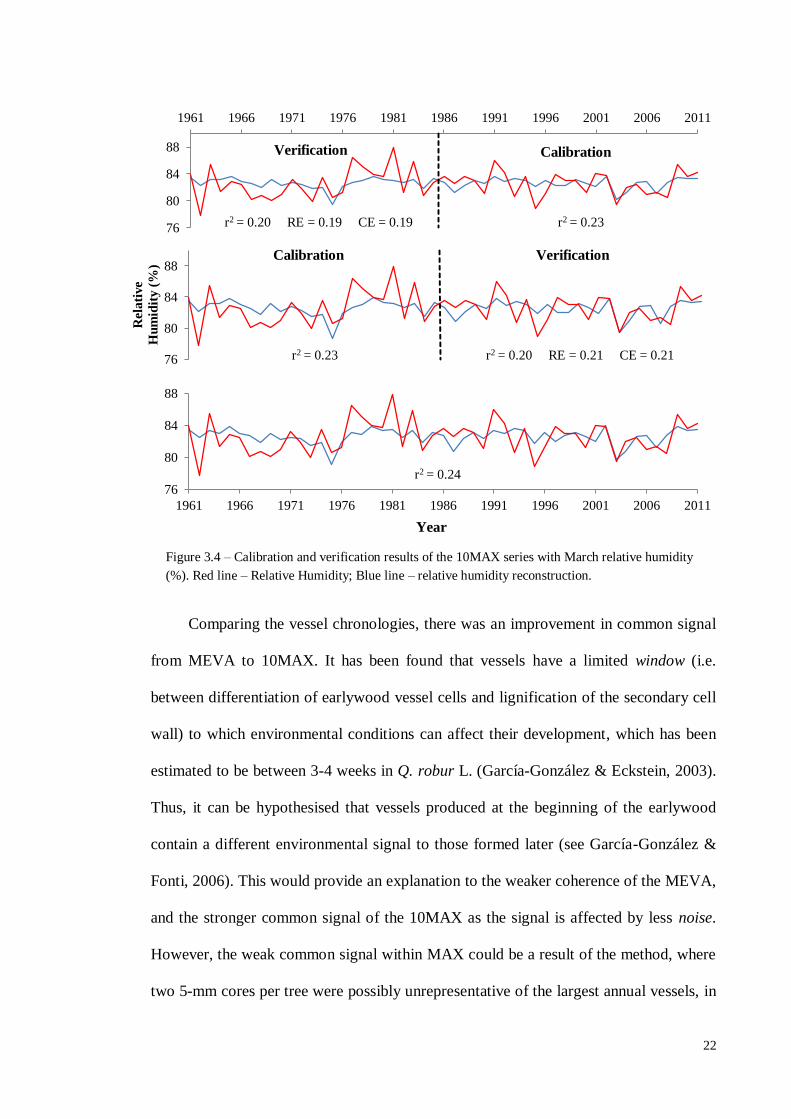
Comparing the vessel chronologies, there was an improvement in common signal
from MEVA to 10MAX. It has been found that vessels have a limited window (i.e.
between differentiation of earlywood vessel cells and lignification of the secondary cell
wall) to which environmental conditions can affect their development, which has been
estimated to be between 3-4 weeks in Q. robur L. (García-González & Eckstein, 2003).
Thus, it can be hypothesised that vessels produced at the beginning of the earlywood
contain a different environmental signal to those formed later (see García-González &
Fonti, 2006). This would provide an explanation to the weaker coherence of the MEVA,
and the stronger common signal of the 10MAX as the signal is affected by less noise.
However, the weak common signal within MAX could be a result of the method, where
two 5-mm cores per tree were possibly unrepresentative of the largest annual vessels, in
76
80
84
88
1961 1966 1971 1976 1981 1986 1991 1996 2001 2006 2011
Verification Calibration
r2 = 0.20 RE = 0.19 CE = 0.19 r2 = 0.23
76
80
84
88
1961 1966 1971 1976 1981 1986 1991 1996 2001 2006 2011
r2 = 0.24
76
80
84
88
Rela
tive
Hu
mid
ity (
%)
r2 = 0.20 RE = 0.21 CE = 0.21 r2 = 0.23
Verification Calibration
Figure 3.4 – Calibration and verification results of the 10MAX series with March relative humidity
(%). Red line – Relative Humidity; Blue line – relative humidity reconstruction.
Year

23
contrast to García-González & Fonti (2007). In fact, some investigations have taken
three cores per tree (e.g. García-González & Eckstein, 2003), although this hasn’t
necessarily been shown to be beneficial. It may be possible to improve the common
signal by increasing the sample size. Although this study predicted a large range of trees
that would be required to get an EPS of 0.85, the estimates are feasible as many dendro-
based reconstructions include hundreds of specimens (e.g. Grudd et al., 2002; Grudd,
2008; Cooper et al., 2013; Wilson et al., 2013).
For earlywood vessel area to be viewed as a useful climatic proxy, it must be
demonstrated that the signal it contains is unique, or better in respect to other ‘easier’ to
obtain tree proxies (Fonti & García-González, 2004). Although significantly correlated,
RW could only account for a small amount of variation within the earlywood vessel
chronologies. In addition, climate-growth models demonstrated differing responses.
Fonti & García-González (2008) suggest that the differences in response can be
attributed to how radial growth and vessels are affected by environmental stimuli. They
argue that radial increment is controlled by the sum of environmental and biological
processes over a year, while vessels are influenced by fewer controls, over a shorter
period of time. Thus, it can be said that at this location vessels and RW have responded
differently to climatic influences, which has been repeatedly demonstrated at different
locations in the literature (e.g. Pumijumnong & Park, 1999; Fonti & García-González,
2004; Matisons & Brumelis, 2012). Consequently, there appears to be merit in
researching the climatic content of earlywood vessel area at this location.
Due to the short period in which individual vessels can be influenced by
environmental factors (see above), it is hypothesised that they contain a high resolution
climatic signal (Eckstein, 2004). This was reflected by the vessels being most strongly
correlated with March humidity. The response to humidity, although a first in the
24
70
75
80
85
90
95
1860 1880 1900 1920 1940 1960 1980 2000
Rela
tive H
um
idit
y (
%)
Year
Figure 3.5 – Reconstruction of past March relative humidity (%) for the period 1860 – 2011 for the National Botanic Gardens of Wales produced from the 10MAX
series. The black line represents the reconstruction, while the red line is the observed relative humidity (see Jenkins et al., 2008). Grey lines represent the
uncertainty of the reconstruction utilizing Gaussian error propagation (±95%).
24

25
literature, is not surprising as vessel area is thought to be linked to moisture availability
(e.g. Knigge & Schulz, 1961; Pumijumnong & Park, 1999; García-González &
Eckstein, 2003; Corcuera et al., 2004; Fonti & García-González, 2004; Eilmann et al.,
2006). In fact, vessel area has been observed to be greater in humid than drier climates
(Kondoh et al., 2006). Thus, it is hypothesised that as relative humidity increases, so
does the moisture availability within the soil, due to precipitation and reduced
evaporation. With an increase in soil moisture content, the tree root system can take
advantage. As water quantities increase within the trees hydrosystem, turgor pressure,
which plays a large role in determining cell size, will increase (Ray et al., 1972; Boyer,
1985; Eilmann et al., 2006). Ultimately, this explains the relationship observed. The fact
that this signal was strongest in relation to March relative humidity can be explained
physiologically. Within ring-porous oak, the largest vessels are produced at the
beginning of the growing season, prior bud burst, ensuring adequate hydraulic
conductivity. Phenological observations have found that within South Wales the first-
flush begins early in the growing season, at the end of March and beginning of April
(Forestry Commission, 2001). Thus, the largest vessels would be expected to respond to
environmental stimuli during March. However, it could then be argued that the MAX
chronology should have contained the strongest response, but it is possible that an
under-representation of the largest vessels influenced the result, which is suggested by
the weak statistical quality of the chronology. In fact, it has previously been suggested
that selecting the largest vessels would not provide an optimal signal (García-González
& Fonti, 2006). To confirm these observations, more local phenological investigations
in conjunction with the extraction of micro-cores to investigate vessel development are
required (e.g. Fonti et al., 2007).

26
The highly significant correlation between August temperatures and the vessel
chronologies is harder to explain, as earlywood production is likely to have ceased prior
this point (Speer, 2010. P.43.). However, earlywood and latewood production is
dependent on environmental conditions and it is possible that earlywood production
could extend late into the summer months (Speer, 2010. P.43.). In fact, this explanation
could have some potential as the inclusion of the smaller vessels, which can be assumed
to be produced later in the year (García-González & Fonti, 2006), improved the signal
strength. In addition, it has been documented that cooler conditions slow cell formation
resulting in a longer window in which the climate can affect vessel characteristics (i.e.
Fonti & García-González, 2004) although, such an influence in the summer months is
questionable. Assuming, earlywood production had ceased prior this point other
explanations would be required. For instance, the August temperature signal could have
arisen due to its influence on earlier formed vessels, which would explain why the
MAX and 5MAX chronologies also demonstrated a significant correlation. However,
observations imply that cell wall lignification, after 3-4 weeks, inhibits alteration of
vessel characteristics (Eckstein, 2004). Although evidence suggests that August
temperature is influencing vessel area, it would be wrong to conclude this without
further research, as correlation does not mean causation devoid of a clear physiological
mechanism.
Results demonstrated that vessel chronologies were less dependent on previous
year conditions compared to RW. This has been reported within the literature (e.g.
García-González & Eckstein, 2003). In fact, it appears to be one of the advantages of
vessel chronologies over ring-width chronologies. However, significant correlations
with previous winter temperatures were found. Within oak it has been repeatedly
observed that ring-width is negatively related to winter temperatures (Pilcher, 1995) and

27
it can be expected that vessels are influenced by the same factors. It has been suggested
that this signal is a result of winter conditions affecting the storage and synthesis of
carbohydrates, which in turn influences the coming spring growth (Aloni, 1991; Alla &
Camarero, 2012). However, in contrast to Fonti & García-González (2004) no influence
was found with previous summer conditions.
Examination of the RE, CE and correlation statistics demonstrated that the climate
models varied in their reconstruction skill. The 5MAX chronology was found to have
the strongest correlation with the humidity record however, considering the RE, CE and
r2
statistics, it was found that the climate model was unsuitable. On the other hand, the
10MAX chronology was found to be more stable in its predictive ability. A possible
explanation is that chronologies constructed from the largest or five largest vessels
could be subject to greater errors in measurement, due to the use of only a select few
vessels. Again, it is also possible that relying on a small selection of vessels from just
two 5-mm cores may result in large vessels being missed, leading to an under-estimate
of annual vessel size. Thus, the 10MAX may overcome these limitations by including
more vessels (without mixing climatic signals), thus a greater chance of gaining an
adequate vessel sample, representing different vessel sizes. Although, the validation
statistics suggest the 10MAX chronology is suitable for a climatic reconstruction, it is
important to remember that the signal strength between series is not satisfactory, thus
the reconstruction could be miss-leading. The reconstruction predicted that March 1975
was the least humid of the study period, in contrast to the observed humidity data.
Although this raises question marks as to the suitability of the reconstruction, it is
interesting to note that between 1975 and 1976 Western Europe saw some of the worst
drought conditions on record (Fleig et al., 2011). Although, the drought is considered to
have begun late in the spring (Ratcliffe, 1978), there had been a period from 1970 to

28
1974 of dry winters which lead to depleted ground water levels in the spring of 1975
(Ratcliffe, 1978). As vessel size is part dependent on moisture availability during spring
(Ray et al., 1972; Boyer, 1985; Eilmann et al., 2006) it is suggest here, that the reduced
vessel sizes of 1975 seen in the reconstruction, and in a number of the raw chronologies,
is a consequence of these dry conditions. This result conforms to ideas that vessels
provide a potential proxy for extreme climatic years (e.g. Knigge & Schulz, 1961).
When considering the previous discussion a number of limitations should be
considered. The humidity data set that was utilized was relatively short, thus increasing
the risk of spurious results. In addition, there is no detailed phenological data available
concerning vessel formation and leaf flushing at the research location, which would aid
in data interpretation.
Following the previous discussion it is possible to evaluate the use of vessel area
chronologies as a climate proxy within the UK. In line with other European studies,
there does appear to be a climatic signal contained within vessel area increments which
was distinct, and in some cases, stronger than ring-width measurements at the research
location in question. In addition, there does seem to be potential in the use of vessel area
chronologies in producing very high resolution reconstructions, which will benefit in
our understanding of the climate system. The prospect of vessels recording a humidity
signal is advantageous in examining climate change, past and present, as humidity plays
a major role in the energy balance of the climate system (Shu et al., 2005). However, it
is clear that the quality of the constructed chronologies is something that needs to be
questioned, although this maybe improved through larger sample sizes. Finally, it has
been demonstrated that care and knowledge must be used when utilizing vessels in a
reconstruction, as the strongest correlation may not be the best, and the use of statistics
such as the RE and CE is strongly recommended.

29
Although the results of this investigation have demonstrated the potential use of
vessels as a proxy, the field is still relatively new and further work is required. Much
has now been published on the climatic influences on vessel area, however, there are
conflicting results. Thus, further investigation on the climatic influence, site conditions
and differences between species on vessel area is required. Great benefit will also be
obtained if such investigations occur in new regions as spatially, present studies are
limited (e.g. Latvia, Matisons et al., 2013; Switzerland, Fonti & Garcia-González,
2004). In addition, it can be argued that the science needs to advance, and work should
now begin to focus on producing actual climatic reconstructions utilizing statistics such
as the RE and CE. Ultimately, this will provide a greater understanding of the suitability
of vessel area as a proxy. As many of the longest tree-ring chronologies utilize
preserved wood samples, research is required to examine the suitability of vessels from
such media, and if results are positive, there is potential that long vessel chronologies
could be built from existing samples (e.g. Friedrich et al., 2007). Finally, there is the
future possibility that if techniques are developed which allow the accurate
identification of when specific vessels were formed, that high resolution reconstructions
of the growing season maybe produced from the same core samples, providing an
invaluable source of past climatic information.

30
5. Conclusion
High resolution proxies are highly sort after within paleo-climatology, and
earlywood vessel area chronologies are thought to be a promising avenue. This thesis
aimed to investigate the applicability of earlywood vessel area as a proxy, and was a
first in the United Kingdom. From this investigation a number of conclusions can be
drawn:
1) It appears that time-series of vessel areas have a reduced statistical quality
compared to ring-width chronologies, in respect to their common signal and
variability. In addition, vessels demonstrated a reduced year to year sensitivity to
environmental stimuli. However, vessel series had the advantage of being less
dependent on previous year growth than RW.
2) Annually, earlywood vessels at this location were mainly influenced by March
relative humidity. In addition, August temperatures appeared to be negatively
related to the vessel series, although there is no clear mechanism to explain this
relationship. Vessel chronologies displayed different responses to the climate
compared to a ring-width series, suggesting there is merit in the use of vessel
area chronologies at this location.
3) A chronology constructed from the average area of all earlywood vessels
demonstrated a weaker correlation to the climate, compared to chronologies
composed of selected vessel sizes, demonstrating that different groups of vessels
may contain different environmental signals. The average of the 5 largest vessels
were most related to March relative humidity, compared to the other series,
while a series composed of the largest ten vessels was more related to August
temperatures.

31
4) Although this study produced a relative humidity reconstruction, caution is
needed in interpreting the results due to weak verification statistics. However,
there is an indication of vessels containing a record of extreme events. A larger
sample depth would probably have improved the expressed climate signal. Thus,
it would appear to be potential in using vessels as a proxy within the UK.
From this study a number areas for further research have been identified. Most
importantly is the need to conduct similar research with different species, in new
regions and climates, to understand differences in vessel responses, while, further
insight into vessel development in relation to phenological observations would also be
beneficial. In addition, emphasis should be placed on examining new methods to
identify the signals contained within specific vessel groups, which could lead to the
future possibility of an unprecedented high resolution climate proxy.

32
References
Alla, A. Q., & Camarero, J. J. (2012) “Contrasting responses of radial growth and wood
anatomy to climate in a Mediterranean ring-porous oak: implications for its
future persistence or why the variance matters more than the mean” European
Journal of Forest Research. 131. 1537 – 1550.
Allan, R. P., & Soden, B. J. (2008) “Atmospheric warming and the amplification of
precipitation” Science. 321. Pp. 1481 – 1484.
Aloni, R. (1991) Wood formation in deciduous hardwood trees. In: A. S. Raghavendra,
ed. (1991) Physiology of Trees. New York: Wiley. Pp. 75 – 197.
Astrade, L., & Bégin, Y. (1997) “Tree-ring response of Populus tremula L. and Quercus
robur L. to recent spring flodds of the Saone River, France” Ecoscience. 4. 232
– 239.
Barber, K. E., Chambers, F. M., Maddy, D., Stoneman, R., & Brew, J. S. (1994) “A
sensitive high-resolution record of late Holocene climate change from a raised
bog in northern England” The Holocene. 4. 198 – 205.
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W., & Courchamp, F. (2012)
“Impacts of climate change on the future of biodiversity” Ecology letters. 15.
365 – 377.
Boyer, J. S. (1985) “Water transport” Annual Review of Plant Physiology and Plant
Molecular Biology. 36. 473 – 516.
Breda, N., & Granier, A. (1996) “Intra- and interannual variations of transpiration, leaf
area index and radial growth of a sessile oak stand (Quercus petraea)” Annals
of Forest Science. 53. 521 – 536.
Briffa, K. R., Jones, P. D., & Schweingruber, F. H. (1988) “Summer temperature
patterns over Europe: A reconstruction from 1750 A.D. based on maximum
latewood density indices of conifers” Quaternary Research. 30. 36 – 52.
Brunstein, C. F. (1996) “Climatic significance of the bristlecone pine latewood frost-
ring record at Almagre Mountain, Colorado, USA” Arctic and Alpine
Research. 28. 65 – 76.
Burk, R. L., & Stuiver, M. (1981) “Oxygen isotope ratios in trees reflect mean annual
temperature and humidity” Science. 211. 1417 – 1419.
Campelo, F. (2012) detrendeR: Start the detrendeR Graphical User Interface (GUI). R
Package version 1.0.4. Available at: < http://CRAN.R-
project.org/package=detrendeR > [Accessed 1/9/13].
Campelo, F., Gutiérrez, E., Ribas, M., Nabais, C., & Freitas, H. (2007) “Relationships
between climate and double rings in Quercus ilex from NE Spain” Canadian
Journal of Forest Research. 37. 1915 – 1923.

33
Campelo, F., Nabais, C., Gutiérrez, E., Freitas, H., & García-González, I. (2010)
“Vessel features of Quercus iles L. Growing under Mediterranean climate have
a better climatic signal than tree-ring width” Trees. 24. 463 – 470.
Carlquist, S. (1988) Comparative wood anatomy, Systematic, ecological, and
evolutionary aspects of dicotyledon wood. Berlin: Springer.
Cooper, R. J., Melvin, T. M., Tyers, I., Wilson, R. J., & Briffa, K. R. (2013) “A tre-ring
reconstruction of East Anglian (UK) Hydroclimate variability over the last
millennium” Climate Dynamics. 40. 1019 – 1039.
Corcuera, L., Camarero, J. J., & Gil-Pelegrin, E. (2004) “Effects of a severe drought on
growth and wood anatomical properties of Quercus faginea” IAWA Journal.
25. 185 – 204.
Corcuera, L., Camarero, J. J., Sisó, S., & Gil-Pelegrin, E. (2006) “Radial-growth and
wood-anatomical changes in over aged Quercus pyrenaica coppice stands:
functional responses in a new Mediterranean landscape” Trees. 20. 91 – 98.
Coumou, D., & Rahmstorf, S. (2012) “A decade of weather extremes” Nature Climate
Change. 2, 7. 491 – 496.
DeSoto, L., Cruz, M. D. L., & Fonti, P. (2011) “Intra-annual patterns of tracheid size in
the Mediterranean tree Juniperus thurifera as an indicator of seasonal water
stress” Canadian Journal of Forest Research. 41. 1280 – 1294.
Eckstein, D. (2004) “Change in past environments – secrets of the tree hydrosystem”
New Phytologist. 163. 1 – 4.
Eckstein, D., Frisse, E., & Quiehl, F. (1977) “Holzanatomische Untersuchungen zum
Nachweis anthropogener Einflusse auf die Umweltbedingungen einer
Rotbuche” Angew. Bot. 51. 47-56.
Eilmann B., Weber, P., Rigling, A., & Eckstein, D. (2006) “Growth reactions of Pinus
sylvestris L. and Quercus pubescens Willd. to drought years at a xeric site in
Valais, Switzerland” Dendrochronologia. 23. 121 – 132.
Esper, J., Cook, E. R., & Schweingruber, F. H. (2002) “Low-frequency signals in long
tree-ring chronologies for reconstructing past temperature variability” Sciecne.
295. 2250 – 2253.
Esper, J., Krusic, P. J., Peters, K., & Frank, D. C. (2009) “Exploration of long-term
growth changes using the tree-ring program “Spotty”” Dendrochronologia. 27.
75 – 82.
Esper, J., Wilson, R. J. S., Frank, D. C., Moberg, A., Wanner, H., & Luterbacher, J.
(2005) “Climate: past ranges and future changes” Quaternary Science Reviews.
24. 2164 – 2166.

34
Ewers, F. W. (1985) “Xylem structure and water conduction in conifer trees, dicot trees,
and lianas” Int. Assoc. Wood Anat. Bull. 6. 309 – 317.
Fleig, A. K., Tallaksen, L. M., Hisdal, H., & Hannah, D. M. (2011) “Regional
hydrological drought in north-western Europe: linking a new regional drought
index with weather types” Hydrological Processes. 25. 1163 – 1179.
Fonti, P., & García-González, I. (2004) “Suitability of chestnut earlywood vessel
chronologies for ecological studies” New Phytologist. 163. 77 – 86.
Fonti, P., & García-González, I. (2008) “Earlywood vessel size of oak as a potential
proxy for spring precipitation in mesic sites” Journal of Biogeography. 35.
2249 – 2257.
Fonti, P., Solomonoff, N., & García-González, I. (2007) “Earlywood vessels of
Castanea sativa recored temperature before their formation” New Phytologist.
173. 562 – 570.
Fonti, P., Treydte, K., Osenstetter, S., Frank, D., & Esper, J. (2009) “Frequency-
dependent signals in multi-centennial oak vessel data” Palaeogeography,
Palaeoclimatology, Palaeoecology. 275. 92 – 99.
Forestry Commission (2001) Tree Phenology. [Online] Available at: <
http://www.forestry.gov.uk/images/cchg_oak_budburst_map.gif/$file/cchg_oa
k_budburst_map.gif >. [Accessed: 08/10/2013].
Friedrich, M., Remmele, S., Kromer, B., Hofmann, J., Spurk, M., Kaiser, K. F., Orcel,
C., & Kuppers, M. (2007) “The 12,460-year Hohenheim oak and pine tree-ring
chronology from Central Europe; a unique record for radiocarbon calibration
and paleoenvironment reconstructions” Radiocarbon. 46. 1111 -1122.
Fritts, H. C. (1976) Tree rings and climate. London: Academic Press.
Fritz, S. C., Juggins, S., Battarbee, R. W., & Engstrom, D. R. (1991) “Reconstruction of
past changes in salinity and climate using a diatom-based transfer function”
Nature. 352. 706 – 708.
García-González, I. G., & Eckstein, D. (2003) “Climatic signal of earlywood vessels of
oak on a maritime site” Tree Physiology. 23. 497 – 504.
García-González, I., & Fonti, P. (2006) “Selecting earlywood vessels to maximize their
environmental signal” Tree Physiology. 26. 1289 – 1296.
García-González, I., & Fonti, P. (2007) “Ensuring a representative sample of earlywood
vessels for dendroecological studies: an example from two ring-porous
species” Trees. 22. 237 – 244.
Grudd, H. (2008) “Tornetrask tree-ring width and density AD 500-2004: a test of
climatic sensitivity and a new 1500-year reconstruction of north Fennoscandian
summers” Climate Dynamics. 31. 843 – 857.

35
Grudd, H., Briffa, K. R., Karlen, W., Bartholin, T. S., Jones, P. D., & Kromer, B. (2002)
“A 7400-year tree-ring chronology in northern Swedish Lapland: natural
climatic variability expressed on annual to millennial timescales” The
Holocene. 12. 657 – 665.
Hacke, U., & Sauter, J. J. (1996) “Xylem dysfunction during winter and recovery of
hydraulic conductivity in diffuse-porous and ring-porous trees” Oecologia.
105. 435 – 439.
Haylock, M. R., Hofstra, N., Klein Tank, A. M.G., Klok, E. J., Jones, P. D., & New, M.
(2008) “A European daily high-resolution gridded dataset of surface
temperature and precipitation” Journal of Geophysical Research
(Atmospheres) 113. D20119. DOI: 10.1029/2008JD10201.
Jenkins, G. J., Perry, M. C., & Prior, M. J. (2008) The climate of the United Kingdom
and recent trends. [pdf] Exeter: Met Office Hadley Center. Available at: <
http://www.ukcip.org.uk/wordpress/wp-content/PDFs/UKCP09_Trends.pdf >
[Accessed 25/09/13].
Kalela-Brundin, M. (1999) “Climatic information from tree-rings of Pinus sylvestris L.
and a reconstruction of summer temperature back to AD 1500 in
Femundsmarka, eastern Norway, using partial least squares regression (PLS)
analysis” The Holocene. 9. 59 – 77.
Kiss, A., Wilson, R., & Bariska, I. (2011) “An experimental 392-year documentary-
based multi-proxy (vine and grain) reconstruction of May-July temperatures
for Koszeg, West-Hungary” Int. J. Biometeorol. 55. 595 – 611.
Knigge, W., & Schulz, H. (1961) “Einfluss der Jahreswitterung 1959 auf die
Zellartenverteilung, Faserlange und Gefassweite verschiedener Holzarten”
Holz als Roh- und Werkstoff. 19. 293 – 303.
Kondoh, S., Yahata, H., Nakashizyka, T., & Kongoh, M. (2006) “Interspecific variation
in vessel size, growth and drought tolerance of broad-leaved trees in semi-arid
regions of Kenya” Tree Physiology. 26. 899 – 904.
Mann, M. E., Bradley, R. S., & Hughes, M. K. (1999) “Northern hemisphere
temperatures during the past millennium: inferences, uncertainties and
limitations” Geophysical Research Letters. 26. 759 – 762.
Matisons, R., & Brumelis, G. (2012) “Influence of climate on tree-ring and earlywood
vessel formation in Quercus robur in Latvia” Trees. 26. 1251 – 1266.
Matisons, R., Elferts, D., & Brumelis, G. (2012) “Changes in climatic signals of English
oak tree-ring width and cross-section area of earlywood vessels in Latvia
during the period 1900-2009” Forest Ecology and Management. 279. 34 – 44.
Matisons, R., Elferts, D., & Brumelis, G. (2013) “Pointer years in tree-ring width and
earlywood-vessel area time series of Quercus robur – relation with climate
factors near its northern distribution limit” Dendrochronologia. 31. 129 – 139.

36
McCarroll, D., & Loader, N. J. (2004) “Stable isotopes in tree rings” Quaternary
Science Reviews. 23. 771 – 801.
McCarroll, D., Loader, N. J., Jalkanen, R., Gagen, M. H., Grudd, H., Gunnarson, B. E.,
Kirchhefer, A. J., Friedrich, M., Linderholm, H. W., Lindholm, H. W.,
Boettger, T., Los, S., Remmele, S., Kononov, Y. M., Yamazaki, Y. H., Young,
G. H. F., & Zorita, E. (2013) “A 1200-year multiproxy record of tree growth
and summer temperature at the northern pine forest limit of Europe” The
Holocene. DOI: 10.1177/0959683612467483.
McMichael, A. J., Woodruff, R. E., & Hales, S. (2006) “Climate change and human
health: present and future risks” The Lancet. 367. 859 – 869.
Moberg, A., Sonechkin, D. M., Holmgren, K., Datsenko, N. M., & Karlén, W. (2005)
“Highly variable northern hemisphere temperatures reconstructed from low-
and high- resolution proxy data” Nature. 433. 613 – 617.
National Botanic Gardens of Wales (2013) Waun Las Grasslands. [online] Available at:
< http://www.gardenofwales.org.uk/science/waun-las-grasslands > [Accessed
03/10/2013.
National Research Council (2007) Surface Temperature Reconstructions for the Last
2,000 Years. National Academies Press: Washington, DC.
Panyushkina, I. P., Hughes, M. K., Vaganov, E. A., & Munro, M. A. R. (2003)
“Summer temperature in northeastern Siberia since 1642 reconstrcuted from
tracheid dimensions and cell numbers of Larix cajanderi”. Canadian Journal
of Forest Research. 33. 1905 – 1914.
Petit, J. R., Jouzel, J., Raynaud, D., Barkov, I., Barnola, J. M., Basile, I., Bender, M.,
Chappellaz, J., Davis, M., Delaygue, G., Delmotte, M., Kotylakov, V. M.,
Legrand, M., Lipenkov, V. Y., Lorius, C., Pepin, L., Ritz, C., Saltzman, E., &
Stievenard, M. (1999) “Climate and atmospheric history of the past 420,000
years from the Vostok ice core, Antarctica” Nature. 399. 429-436.
Pilcher, J. R. (1995) Biological consideration in the interpretation of stable isotope
ratios in oak tree-rings. In B. Frenzel (ed) Problems of stable isotopes in tree-
rings, lake sediments and peat-bogs as climatic evidence for the Holocene.
157-161.
Pumijumnong, N., & Park, W. K. (1999) “Vessel chronologies from teak in northern
Thailand and their climatic signals” IAWA Journal. 20. 285 – 294.
R Core Team (2013). R: A language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria. Available at <
http://www.R-project.org/ > [Accessed 1/09/13].
Ratcliffe, R. A. S., Miles, M. K., Rydz, B., & Shaw, E. M. (1978) “Meteorological
aspects of the 1975 – 76 drought [and discussion]” Proceedings of the Royal
Society A. 363. 3 – 20.

37
Ray, P. M., Green, P. B., & Cleland, R. (1972) “Role of turgor in plant-cell growth”
Nature. 239. 163 – 164.
Rinn, F. (2003) TSAP-Win-time series analysis and presentation dendrochronology and
related applications. Heidelberg: Frank Rinn.
Rodwell, J. S. (1991) British Plant Communities. Volume 1. Woodlands and scrub.
London: Cambridge University Press.
Rodwell, J. S. (1992) British Plant Communities. Volume 3. Grassland and montane
communities. London: Cambridge University Press.
Ruddiman, W. F. (2008) Earth’s Climate, Past and Future. New York: W. H. Freeman.
Salinger, M. J., & Griffiths, G. M. (2001) “Trends in New Zealand daily temperature
and rainfall extremes” International Journal of Climatology. 21. 1437 – 1452.
Sass, U., & Eckstein, D. (1995) “The variability of vessel size in beech (Fagus sylvatica
L.) and its ecophysiological interpretation” Trees. 9. 247 – 252.
Schume, H., Grabner, M., & Eckmullner, O. (2004) “The influence of an altered
groundwater regime on vessel properties of hybrid poplar” Trees. 18. 184 –
194.
Schweingruber, F.H. (2001) Dendroökologische Holzanatomie. Bern: Paul Haupt.
Shu, Y., Feng, X., Gazis, C., Anderson, D., Faiia, A. M., Tang, K., & Ettl, G. J. (2005)
“Relative humidity recorded in tree rings: A study along a precipitation
gradient in the Olympic Mountains, Washington USA” Geochimica et
Cosmochimica Acta. 69. 791 – 799.
Solomon, S., D. Qin, M. Manning, R.B. Alley, T. Berntsen, N.L. Bindoff, Z. Chen, A.
Chidthaisong, J.M. Gregory, G.C. Hegerl, M. Heimann, B. Hewitson, B.J.
Hoskins, F. Joos, J. Jouzel, V. Kattsov, U. Lohmann, T. Matsuno, M. Molina,
N. Nicholls, J. Overpeck, G. Raga, V. Ramaswamy, J. Ren, M. Rusticucci, R.
Somerville, T.F. Stocker, P. Whetton, R.A. Wood and D. Wratt, 2007:
Technical Summary. In: Climate Change 2007: The Physical Science Basis.
Contribution of Working Group I to the Fourth Assessment Report of the
Intergovernmental Panel on Climate Change [Solomon, S., D. Qin, M.
Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller
(eds.)]. Cambridge University Press, Cambridge, United Kingdom and New
York, NY, USA.
Speer, J. H. (2010) Fundamentals of tree-ring research. Tucson: The University of
Arizona Press.
St. George, S. & Nielsen, E. (2003) “Palaeoflood records for the Red River, Manitoba,
Canada, derived from anatomical tree-ring signitures” The Holocene. 13. 547 –
555.

38
St. George, S., & Nielsen, E. (2000) “Signatures of high-magnitude 19th century floods
in Quercus macrocarpa tree rings along the Red River, Manitoba, Canada”
Geology. 28. 899 – 902.
St. George, S., Nielsen, E., Conciatori, F., & Tardif, J. (2002) “Trends in Quercis
macrocarpa vessel areas and their implications for tree-ring paleoflood studies”
Tree-Ring Research. 58. 3 – 10.
Sundberg, R., Mober, A., & Hind, A. (2012) “Statistical framework for the evaluation
of climate model simulations by use of climate proxy data from the last
millennium – Part 1: Theory” Climate of the Past. 8. 1339 – 1353.
Tardif, J. C., & Conciatori, F. (2006) Influence of climate on tree rings and vessel
features in red oak and white oak growing near their northern distribution limit,
southwestern Quebec, Canada. Canadian Journal for Forest Research. 36.
2317 – 2330.
Westra, S., Alexander, L. V., & Zwiers, F. W. (2013) “Global increasing trends in annual
maximum daily precipitation” Journal of Climate. 26. 3904 – 3918.
Wigley, T. M. L., Briffa, K. R., & Jones, P. D. (1984) “On the average value of
correlated time series, with applications in Dendroclimatology and
Hydrometeorology” Journal of Climate and Applied Meteorology. 23. 201 –
213.
Wilson, R., Miles, D., Loader, N. J., Melvin, T., Cunningham, L., Cooper, R., & Briffa,
K. (2013) “A millennial long March-July precipitation reconstruction from
southern-central England” Climate Dynamics. 40. 997 – 1017.
Woodcock, D. W. (1989) “Climate sensitivity of wood-anatomical features in a ring-
porous oak (Quercus macrocarpa)” Canadian Journal for Forest Research. 19.
639 – 644.

39
Supplementary Materials for
An Evaluation of the Climatic Signal within the
Earlywood Vessel Area of Quercus petraea (Matt.) Liebl.
Darren Davies (552192)
This Section includes:
Supplementary Materials and Methods
Figs. S1 to S3
Tables S1 to S2
Captions for databases S1 to S3
Other Supplementary Materials for this manuscript includes the following:
CD Content Database S1 to S5:
1. Site cross-dating statistics
2. Comparison between constructed master chronology and that of Pilcher et al.,
(1984)
3. Master Chronology
4. Raw Vessel Chronologies
5. Detrended Vessel Chronologies

40
Materials and Methods
Chronology Dating
Provided in Database S1 is the cross-dating statistics used to analyse the relationship
between the individual sample cores. Where the t-value Ballie-Pilcher (TVBP), produced
by TSAP (Rinn, 2003), was found to be greater than 3.5, a significant relationship was
assumed (Baillie & Pilcher, 1973). Where significant relationships were identified, visual
inspection of ring-width graphs were used to identify any possibility of missed rings from
the measurement stage. Once these criteria were satisfied, cores were added to the master
chronology which was produced by TSAP (Rinn, 2003). To confirm the accuracy of the
master chronology, it was cross-dated with the master chronology of Pilcher et al., (1984)
where the TVBP value was 7.9 (Database S2). Consequently, the master chronology
produced was thought to be accurate (Database S3). The constructed chronology was found
to cover the period of 1773 – 2012, however, due to a reduced coverage and time
constraints only the period of 1860 – 2012 was considered for further analysis.
Sample Preparation
Initial analysis of core samples was conducted without alteration of the cores sanded
surface. However, it was found that WinCELL PRO (ver. 2013) software (Régents
Instruments Inc., Québec, Canada) was unable to accurately identify and measure vessel
lumina correctly in their natural form (Figure S1A). Initially, vessels were cleared with
compressed air and in-filled with white chalk by hand, to see if there was an improvement,
however, results were not satisfactory (Figure S1B). Therefore, following a popular
approach in the literature (e.g. Fonti et al., 2007; Fonti & García-González, 2008; Alla &
Camarero, 2012) the wood matrix was coloured black with permanent marker, in addition

41
to completing the previous steps. To prohibit chalk falling out of the vessels a coating of
hairspray was applied. Altogether, these steps greatly improved the identification and
measurement of the vessel areas (Figure S1C).
Vessel Sorting Package
As there was no readymade software for sorting the measured vessels by size, a package
was created in Microsoft Excel (ver. 2007) using Visual Basic for Applications code. The
Macro used within the package works in a logical fashion by selecting the area of each
vessel, from each year, and sorting them in ascending order. Following this the code
separates, from the annual vessel area size list, a selection of the largest, five largest, ten
largest and all the vessel areas and where appropriate produces an average. This is repeated
for each year of the series producing the chronologies described in table 2.1.
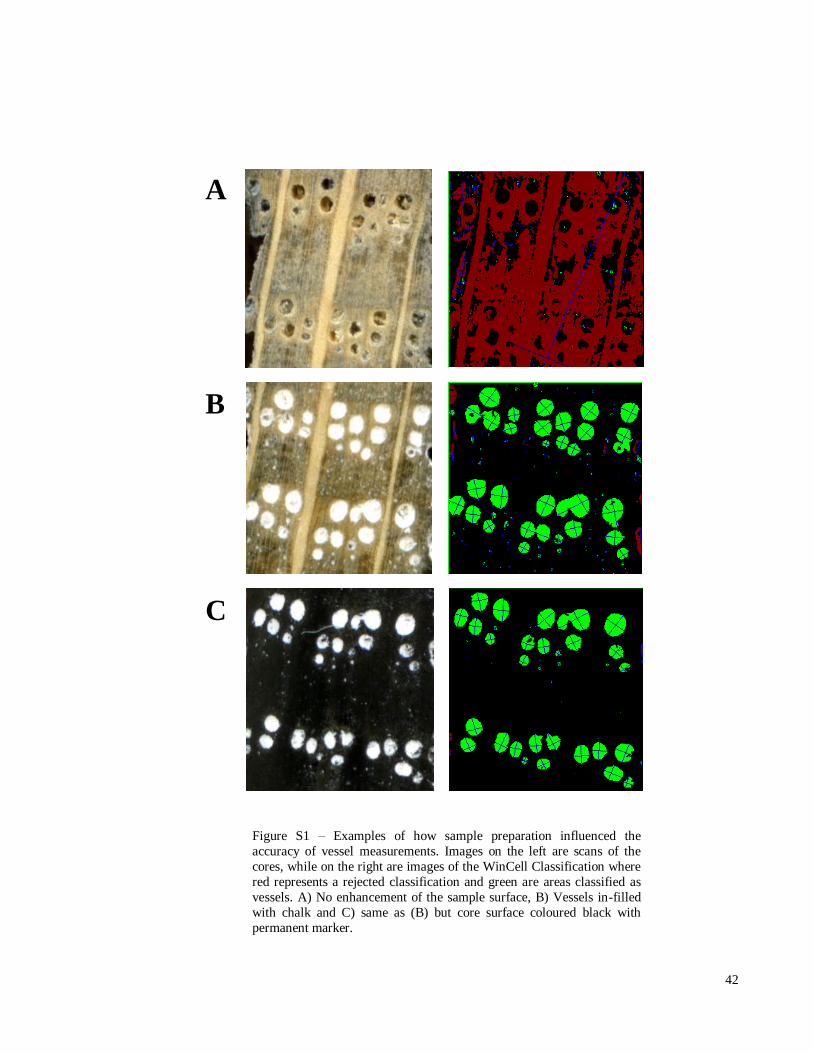
42
Figure S1 – Examples of how sample preparation influenced the
accuracy of vessel measurements. Images on the left are scans of the
cores, while on the right are images of the WinCell Classification where
red represents a rejected classification and green are areas classified as
vessels. A) No enhancement of the sample surface, B) Vessels in-filled
with chalk and C) same as (B) but core surface coloured black with
permanent marker.
A
B
C

43
Figure S2 – Sample coverage. Note NBGW007 and 014 are missing years.
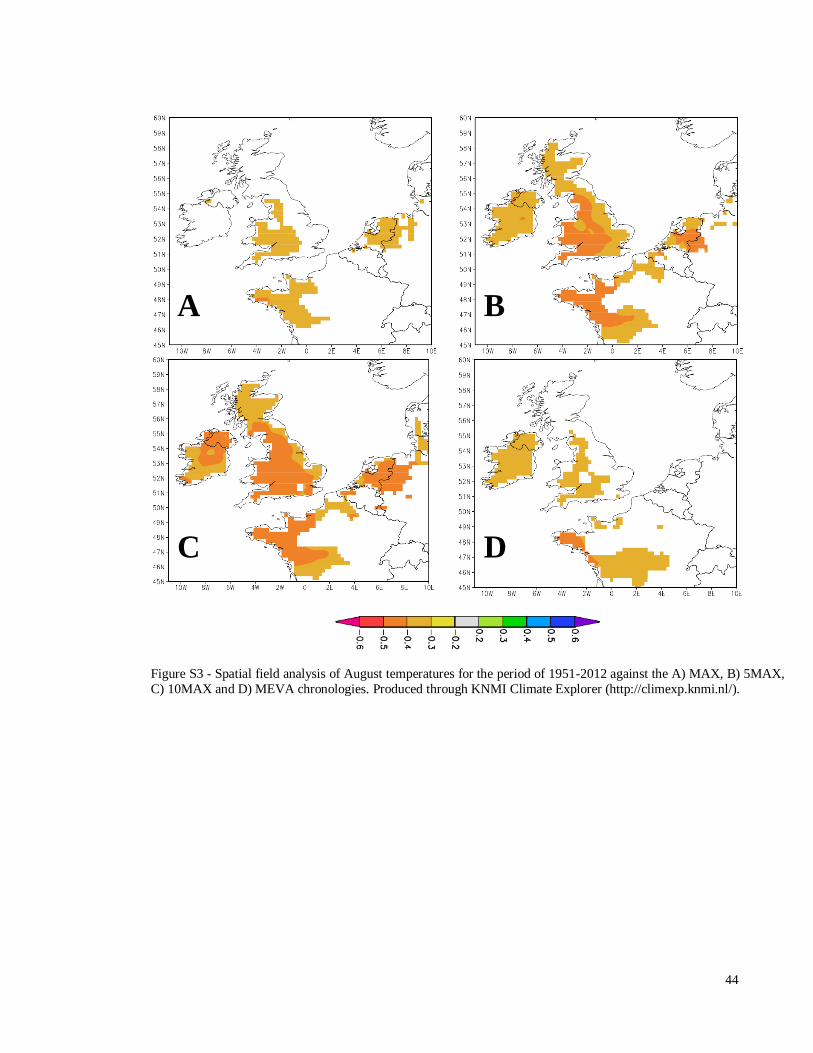
44
Figure S3 - Spatial field analysis of August temperatures for the period of 1951-2012 against the A) MAX, B) 5MAX,
C) 10MAX and D) MEVA chronologies. Produced through KNMI Climate Explorer (http://climexp.knmi.nl/).
A B
C D
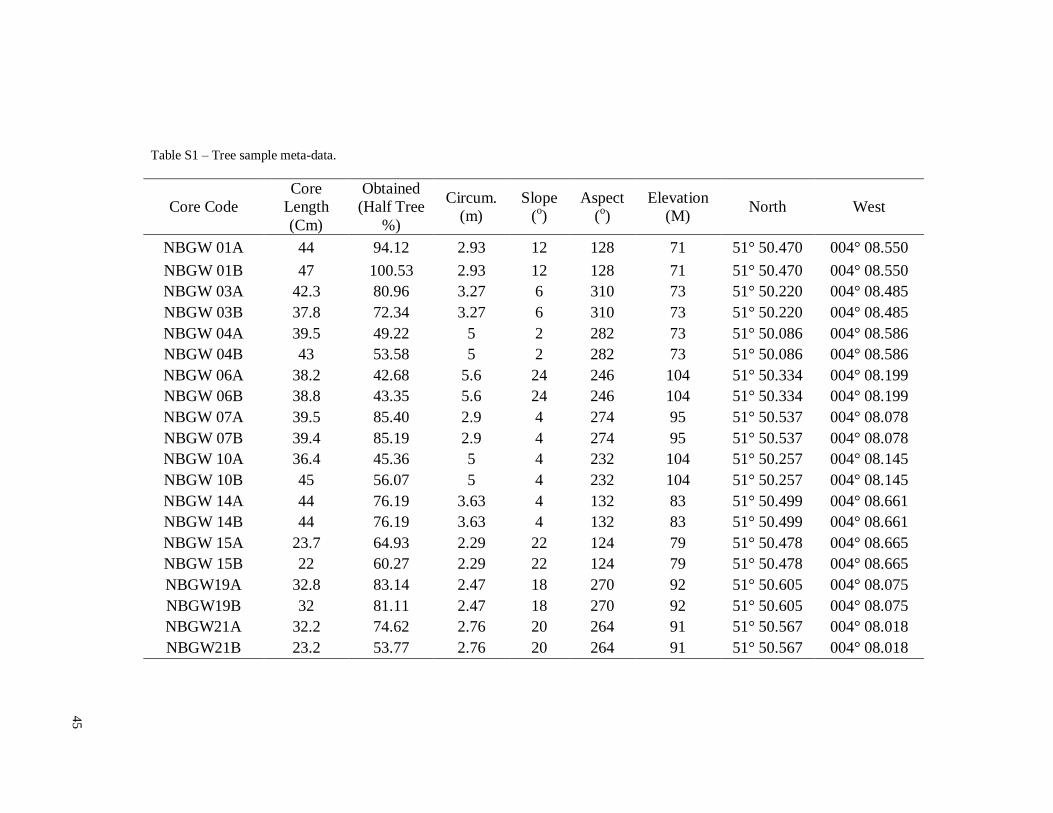
45
Table S1 – Tree sample meta-data.
Core Code
Core
Length
(Cm)
Obtained
(Half Tree
%)
Circum.
(m)
Slope
(o)
Aspect
(o)
Elevation
(M) North West
NBGW 01A 44 94.12 2.93 12 128 71 51° 50.470 004° 08.550
NBGW 01B 47 100.53 2.93 12 128 71 51° 50.470 004° 08.550
NBGW 03A 42.3 80.96 3.27 6 310 73 51° 50.220 004° 08.485
NBGW 03B 37.8 72.34 3.27 6 310 73 51° 50.220 004° 08.485
NBGW 04A 39.5 49.22 5 2 282 73 51° 50.086 004° 08.586
NBGW 04B 43 53.58 5 2 282 73 51° 50.086 004° 08.586
NBGW 06A 38.2 42.68 5.6 24 246 104 51° 50.334 004° 08.199
NBGW 06B 38.8 43.35 5.6 24 246 104 51° 50.334 004° 08.199
NBGW 07A 39.5 85.40 2.9 4 274 95 51° 50.537 004° 08.078
NBGW 07B 39.4 85.19 2.9 4 274 95 51° 50.537 004° 08.078
NBGW 10A 36.4 45.36 5 4 232 104 51° 50.257 004° 08.145
NBGW 10B 45 56.07 5 4 232 104 51° 50.257 004° 08.145
NBGW 14A 44 76.19 3.63 4 132 83 51° 50.499 004° 08.661
NBGW 14B 44 76.19 3.63 4 132 83 51° 50.499 004° 08.661
NBGW 15A 23.7 64.93 2.29 22 124 79 51° 50.478 004° 08.665
NBGW 15B 22 60.27 2.29 22 124 79 51° 50.478 004° 08.665
NBGW19A 32.8 83.14 2.47 18 270 92 51° 50.605 004° 08.075
NBGW19B 32 81.11 2.47 18 270 92 51° 50.605 004° 08.075
NBGW21A 32.2 74.62 2.76 20 264 91 51° 50.567 004° 08.018
NBGW21B 23.2 53.77 2.76 20 264 91 51° 50.567 004° 08.018
45

46
Table S2 -Filters used to identify and control abnormal
classifications
Filter Criteria
Area < 10,000 μm2
Area > 150,000 μm2
Length < 150 μm
Width < 50 μm
Width > 500 μm
Length / Width ratio < 0.05
Length / Width ratio > 1.9

47
Additional Data (separate files on CD)
Contained on the attached CD is a copy of the cross-dating statistics for the individual tree
cores (Database S1) and between the constructed master chronology and the master
chronology of Pilcher et al., (1984) (Database S2). Database S3 contains the master
chronology. Also contained is the raw (Database S4) and detrended vessel data (Database
S5).

48
Supplementary References
Alla, A. Q., & Camarero, J. J. (2012) “Contrasting responses of radial growth and wood
anatomy to climate in a Mediterranean ring-porous oak: implications for its future
persistence or why the variance matters more than the mean” European Journal of
Forest Research. 131. 1537 – 1550.
Baillie, M. G. L., & Pilcher, J. R. (1973) “A simple cross-dating program for tree-ring
research” Tree-Ring bulletin. 33. 7 – 14.
Fonti, P., & García-González, I. (2008) “Earlywood vessel size of oak as a potential proxy
for spring precipitation in mesic sites” Journal of Biogeography. 35. 2249 – 2257.
Fonti, P., Solomonoff, N., & García-González, I. (2007) “Earlywood vessels of Castanea
sativa recored temperature before their formation” New Phytologist. 173. 562 –
570.
Pilcher, J. R., Baillie, M. G. L., Schmidt, B., & Becker, B. (1984) “A 7,272 year tree-ring
chronology for western Europe” Nature. 312. 150 – 152.
Rinn, F. (2003) TSAP-Win-time series analysis and presentation dendrochronology and
related applications. Heidelberg: Frank Rinn.
49
Appendix 1 - SOTEAS Risk Assessment

50

51

52
53
Appendix 2 – Meeting Logs
An Evaluation of the Climatic Signal within the Earlywood Vessel Area of Quercus petraea
Dr Neil Loader
54
55





























