Embed Size (px)
Citation preview

Press
The discovery of resistant starch represents one of the major developments in our understanding of the importance of carbohydrates for health in the past twenty years. There has been a steady increase in knowledge of its sources, uses and physiological effects, but more information is needed on the measurement and complex physiological functions of the various types. Resistant starch is now being incorporated into commercial foods as an ingredient to increase dietary fibre intake. Both commercial and natural sources of resistant starch have been linked to an array of health benefits, especially those related to gut health.
Resistant Starch: Sources, Applications and Health Benefits covers the intrinsic and extrinsic sources of resistant starch in foods, and compares different methods of measuring resistant starch, their strengths and limitations. Applications in different food categories are addressed by recognized academic researchers and industry experts. The book includes descriptions of how resistant starch performs in bakery, dairy, snack, breakfast cereals, pasta, noodles, confectionery, meat, processed food and beverage products. It also looks at the mechanism for improving intestinal health by resistant starch in comparison to prebiotic oligosaccharides and regular dietary fibres. Other chapters cover the impact of resistant starch on blood glucose response, satiety and gut microbiota composition, as well as metabolism in animal models and individual human subjects, and the book reviews research conducted into the ways in which resistant starch can support the prevention of colon cancer. Resistant Starch: Sources, Applications and Health Benefits is unique in focusing on this versatile and important ingredient, which will be of great use to a wide range of food professionals, including food scientists, product developers and manufacturers.
About the editors
Yong-Cheng Shi is Associate Professor and Director, Carbohydrate Polymers - Technology and Product Innovation, Department of Grain Science and Industry, Kansas State University, USA.
Clodualdo C. Maningat is Vice President, Applications Technology and Technical Services, MGP Ingredients, Inc., USA; Department of Grain Science and Industry, Kansas State University, USA.
Also available from Wiley Blackwell
Oats Nutrition and Technology Edited by YiFang Chu ISBN 978-1-118-35411-7
Cereals and Pulses: Nutraceutical Properties and Health Benefits Edited by Liangli L. Yu, Rong Tsao and Fereidoon Shahidi ISBN 978-0-8138-1839-9
9 780813 809519
ISBN 978-0-8138-0951-9
Press
Resistant Starch
Shi and Maningat
Resistant Starch Sources, Applications and Health Benefits
Resistant Starch Sources, Applications and Health Benefits
Yong-Cheng Shi and Clodualdo C. Maningat
EDITORS
www.wiley.com/go/food
Shi_Resistant_9780813809519_hb.indd 1 27/08/2013 09:20

3GFFIRS 08/28/2013 13:43:24 Page 8

3GFFIRS 08/28/2013 13:43:23 Page 1
Resistant Starch

3GFFIRS 08/28/2013 13:43:23 Page 2
The IFT Press series reflects the mission of the Institute of Food Technologists — to
advance the science of food contributing to healthier people everywhere. Developed
in partnership with Wiley Blackwell, IFT Press books serve as leading-edge
handbooks for industrial application and reference and as essential texts for
academic programs. Crafted through rigorous peer review and meticulous research,
IFT Press publications represent the latest, most significant resources available to
food scientists and related agriculture professionals worldwide. Founded in 1939, the
Institute of Food Technologists is a nonprofit scientific society with 18,000 individual
members working in food science, food technology, and related professions in
industry, academia, and government. IFT serves as a conduit for multidisciplinary
science thought leadership, championing the use of sound science across the food
value chain through knowledge sharing, education, and advocacy.
IFT Press Advisory Group
Nicolas Bordenave
YiFang Chu
J. Peter Clark
Christopher J. Doona
Jung Hoon Han
Florence Feeherry
Chris Findlay
David McDade
Thomas J. Montville
Karen Nachay
Martin Okos
David S. Reid
Sam Saguy
Fereidoon Shahidi
Cindy Stewart
Herbert Stone
Kenneth R. Swartzel
Bob Swientek
Hilary Thesmar
Yael Vodovotz
Ron Wrolstad

3GFFIRS 08/28/2013 13:43:24 Page 3
Resistant StarchSources, Applications andHealth Benefits
Edited by
Yong-Cheng Shi
Department of Grain Science and Industry, Kansas State University, USA
Clodualdo C. Maningat
MGP Ingredients, Inc., USA; Department of Grain Science and Industry,
Kansas State University, USA

3GFFIRS 08/28/2013 13:43:24 Page 4
This edition first published 2013 # 2013 by John Wiley & Sons, Ltd.
Registered office: John Wiley & Sons, Ltd, The Atrium, Southern Gate, Chichester, West Sussex,PO19 8SQ, UK
Editorial offices: 9600 Garsington Road, Oxford, OX4 2DQ, UKThe Atrium, Southern Gate, Chichester, West Sussex, PO19 8SQ, UK111 River Street, Hoboken, NJ 07030-5774, USA
For details of our global editorial offices, for customer services and for information about how toapply for permission to reuse the copyright material in this book please see our website at www.wiley.com/wiley-blackwell.
The right of the author to be identified as the author of this work has been asserted in accordance withthe UK Copyright, Designs and Patents Act 1988.
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, ortransmitted, in any form or by any means, electronic, mechanical, photocopying, recording orotherwise, except as permitted by the UK Copyright, Designs and Patents Act 1988, without the priorpermission of the publisher.
Designations used by companies to distinguish their products are often claimed as trademarks. Allbrand names and product names used in this book are trade names, service marks, trademarks orregistered trademarks of their respective owners. The publisher is not associated with any product orvendor mentioned in this book.
Limit of Liability/Disclaimer of Warranty: While the publisher and author(s) have used their bestefforts in preparing this book, they make no representations or warranties with respect to the accuracyor completeness of the contents of this book and specifically disclaim any implied warranties ofmerchantability or fitness for a particular purpose. It is sold on the understanding that the publisher isnot engaged in rendering professional services and neither the publisher nor the author shall be liablefor damages arising herefrom. If professional advice or other expert assistance is required, theservices of a competent professional should be sought.
Library of Congress Cataloging-in-Publication Data
Resistant starch : sources, applications and health benefits / edited by Clodualdo C. Maningat,Yong-Cheng Shi.
pages cmIncludes bibliographical references and index.ISBN 978-0-8138-0951-9 (cloth)
1. Low-carbohydrate diet. 2. Starch–Health aspects. 3. Reducing diets. I. Maningat, Clodualdo C.,editor of compilation. II. Shi, Yong-Cheng, editor of compilation.
RM237.73R47 2013613.2’833–dc23 2013014149
A catalogue record for this book is available from the British Library.
Wiley also publishes its books in a variety of electronic formats. Some content that appears in printmay not be available in electronic books
Cover design by Andy Meaden
Set in 10.5/12.5pt Times by Thomson Digital, Noida, India.
1 2013

3GFFIRS 08/28/2013 13:43:24 Page 5
Titles in the IFT Press series
� Accelerating New Food Product Design and Development (Jacqueline H. Beckley, Elizabeth
J. Topp, M. Michele Foley, J.C. Huang, and Witoon Prinyawiwatkul)
� Advances in Dairy Ingredients (Geoffrey W. Smithers and Mary Ann Augustin)
� Bioactive Proteins and Peptides as Functional Foods and Nutraceuticals (Yoshinori Mine,
Eunice Li - Chan, and Bo Jiang)
� Biofilms in the Food Environment (Hans P. Blaschek, Hua H. Wang, and Meredith E. Agle)
� Calorimetry in Food Processing: Analysis and Design of Food Systems (G€on€ul KaletunSc)� Coffee: Emerging Health Effects and Disease Prevention (YiFang Chu)
� Food Carbohydrate Chemistry (Ronald E. Wrolstad)
� Food Ingredients for the Global Market (Yao-Wen Huang and Claire L. Kruger)
� Food Irradiation Research and Technology, Second Edition (Christoper H. Sommers and
Xuetong Fan)
� Foodborne Pathogens in the Food Processing Environment: Sources, Detection and Control
(Sadhana Ravishankar, Vijay K. Juneja, and Divya Jaroni)
� High Pressure Processing of Foods (Christopher J. Doona and Florence E. Feeherry)
� Hydrocolloids in Food Processing (Thomas R. Laaman)
� Improving Import Food Safety (Wayne C. Ellefson, Lorna Zach, and Darryl Sullivan)
� Innovative Food Processing Technologies: Advances in Multiphysics Simulation (Kai
Knoerzer, Pablo Juliano, Peter Roupas, and Cornelis Versteeg)
� Microbial Safety of Fresh Produce (Xuetong Fan, Brendan A. Niemira, Christopher J. Doona,
Florence E. Feeherry, and Robert B. Gravani)
� Microbiology and Technology of Fermented Foods (Robert W. Hutkins)
� Multiphysics Simulation of Emerging Food Processing Technologies (Kai Knoerzer, Pablo
Juliano, Peter Roupas and Cornelis Versteeg)
� Multivariate and Probabilistic Analyses of Sensory Science Problems (Jean-FranScoisMeullenet, Rui Xiong, and Christopher J. Findlay
� Nanoscience and Nanotechnology in Food Systems (Hongda Chen)
� Natural Food Flavors and Colorants (Mathew Attokaran)
� Nondestructive Testing of Food Quality (Joseph Irudayaraj and Christoph Reh)
� Nondigestible Carbohydrates and Digestive Health (Teresa M. Paeschke and William R.
Aimutis)
� Nonthermal Processing Technologies for Food (Howard Q. Zhang, Gustavo V. Barbosa-
C�anovas, V.M. Balasubramaniam, C. Patrick Dunne, Daniel F. Farkas, and James T.C. Yuan)
� Nutraceuticals, Glycemic Health and Type 2 Diabetes (Vijai K. Pasupuleti and James W.
Anderson)
� Organic Meat Production and Processing (Steven C. Ricke, Ellen J. Van Loo, Michael G.
Johnson, and Corliss A. O’Bryan)
� Packaging for Nonthermal Processing of Food (Jung H. Han)
� Practical Ethics for the Food Professional: Ethics in Research, Education and the Work-
place (J. Peter Clark and Christopher Ritson)

3GFFIRS 08/28/2013 13:43:24 Page 6
� Preharvest and Postharvest Food Safety: Contemporary Issues and Future Directions (Ross
C. Beier, Suresh D. Pillai, and Timothy D. Phillips, Editors; Richard L. Ziprin, Associate
Editor)
� Processing and Nutrition of Fats and Oils (Ernesto M. Hernandez and Afaf Kamal-Eldin)
� Processing Organic Foods for the Global Market (Gwendolyn V. Wyard, Anne Plotto,
Jessica Walden, and Kathryn Schuett)
� Regulation of Functional Foods and Nutraceuticals: A Global Perspective (Clare M. Hasler)
� Resistant Starch: Sources, Applications and Health Benefits (Yong-Cheng Shi and Clodualdo
Maningat)
� Sensory and Consumer Research in Food Product Design and Development (Howard R.
Moskowitz, Jacqueline H. Beckley, and Anna V.A. Resurreccion)
� Sustainability in the Food Industry (Cheryl J. Baldwin)
� Thermal Processing of Foods: Control and Automation (K.P. Sandeep)
� Trait - Modified Oils in Foods (Frank T. Orthoefer and Gary R. List)
� Water Activity in Foods: Fundamentals and Applications (Gustavo V. Barbosa-C�anovas,Anthony J. Fontana Jr., Shelly J. Schmidt, and Theodore P. Labuza)
� Whey Processing, Functionality and Health Benefits (Charles I. Onwulata and Peter J. Huth)

3GFFIRS 08/28/2013 13:43:24 Page 7
To my wife Lei and my son Gary – YCS
To my wife Josie, my daughter Barbara and my sister Susan – CCM

3GFFIRS 08/28/2013 13:43:24 Page 8

3GFTOC 07/20/2013 11:21:12 Page 9
Contents
Preface xvii
About the Editors xix
List of Contributors xxi
Acknowledgements xxv
1 Starch Biosynthesis in Relation to Resistant Starch 1Geetika Ahuja, Sarita Jaiswal and Ravindra N. Chibbar
1.1 Introduction 11.1.1 Starch components 1
1.1.2 Resistant starch 2
1.2 Factors Affecting Starch Digestibility 3
1.3 Starch Biosynthesis 4
1.4 Starch Biosynthesis in Relation to RS 61.4.1 ADP-glucose pyrophosphorylase (AGPase) 6
1.4.2 Starch synthases (SS) 6
1.4.3 Starch branching enzymes (SBE) 11
1.4.4 Starch debranching enzymes (DBE) 13
1.5 Concluding Remarks 13
Acknowledgements 15
References 15
2 Type 2 Resistant Starch in High-Amylose Maize Starchand its Development 23Hongxin Jiang and Jay-lin Jane
2.1 Introduction 23
2.2 RS Formation in High-Amylose Maize Starch 28
2.3 RS Formation During Kernel Development 29
2.4 Elongated Starch Granules of High-Amylose
Maize Starch 312.4.1 Structures of elongated starch granules 31
ix

3GFTOC 07/20/2013 11:21:12 Page 10
2.4.2 Formation of elongated starch granules 33
2.4.3 Location of RS in the starch granule 35
2.5 Roles of High-Amylose Modifier (HAM) Gene in
Maize ae-Mutant 36
2.6 Conclusions 37
References 38
3 RS4-Type Resistant Starch: Chemistry, Functionalityand Health Benefits 43Clodualdo C. Maningat and Paul A. Seib
3.1 Introduction 43
3.2 Historical Account of Starch Indigestibility 44
3.3 Starch Modification Yielding Increased Resistance
to Enzyme Digestibility 473.3.1 Cross-linked RS4 starches 50
3.3.2 Substituted RS4 starches 54
3.3.3 Pyrodextrinized RS4 Starches 56
3.4 Physicochemical Properties Affecting Functionality 57
3.5 Physiological Responses and Health Benefits 60
3.6 Performance in Food and Beverage Products 65
3.7 Conclusions and Future Perspectives 68
References 68
4 Novel Applications of Amylose-Lipid Complex asResistant Starch Type 5 79Jovin Hasjim, Yongfeng Ai and Jay-lin Jane
4.1 Introduction 79
4.2 Enzyme Digestibility of Amylose-Lipid Complex 804.2.1 Effects of lipid structure on the enzyme resistance
of amylose-lipid complex 81
4.2.2 Effects of the crystalline structure on the enzyme
resistance of amylose-lipid complex 82
4.2.3 Effects of amylose-lipid complex on the enzyme
resistance of granular starch 82
4.3 Production of Resistant Granular Starch Through
Starch-Lipid Complex Formation 834.3.1 Effects of fatty-acid structure on the RS content 83
4.3.2 Effects of debranching on the RS content 85
4.4 Applications of the RS Type 5 86
4.5 Health Benefits of RS Type 5 874.5.1 Glycemic and insulinemic control 87
x Contents

3GFTOC 07/20/2013 11:21:12 Page 11
4.5.2 Colon cancer prevention 89
4.6 Conclusion 91
References 92
5 Digestion Resistant Carbohydrates 95Annette Evans
5.1 Introduction 95
5.2 Starch Digestion 95
5.3 Physical Structures of Starch 975.3.1 Starch helices 98
5.3.2 Crystalline structures 99
5.3.3 Starch granule structure 99
5.4 Resistant Starch due to Physical Structure 100
5.5 Molecular Structure of Starch 102
5.6 Enzyme Resistance due to Molecular Structure 103
5.7 Conclusion 106
References 106
6 Slowly Digestible Starch and Health Benefits 111Genyi Zhang and Bruce R. Hamaker
6.1 Introduction 111
6.2 SDS and Potential Beneficial Health Effects 1126.2.1 Potential health benefit of SDS relative to RDS 113
6.3 The Process of Starch Digestion 1156.3.1 Enzyme action 115
6.4 Structural and Physiological Fundamentals of SDS 1166.4.1 Physical or food matrix structures related to SDS 117
6.4.2 Starch chemical structures leading to SDS 118
6.4.3 Other food factors that decrease digestion rate 120
6.4.4 Physiological control of food motility 121
6.5 Application-Oriented Strategies to Make SDS 1216.5.1 Starch-based ingredients 121
6.5.2 SDS generation in a food matrix 122
6.6 Considerations 123
References 123
7 Measurement of Resistant Starch and Incorporationof Resistant Starch into Dietary Fibre Measurements 131Barry V. McCleary
7.1 Introduction 131
7.2 Development of AOAC Official Method 2002.02 133
Contents xi

3GFTOC 07/20/2013 11:21:12 Page 12
7.3 Development of an Integrated Procedure for the
Measurement of Total Dietary Fibre 136
References 142
8 In Vitro Enzymatic Testing Method and DigestionMechanism of Cross-linked Wheat Starch 145Radhiah Shukri, Paul A. Seib, Clodualdo C. Maningat,
and Yong-Cheng Shi
8.1 Introduction 145
8.2 Materials and Methods 1488.2.1 Materials 148
8.2.2 General methods 148
8.2.3 Conversion of CL wheat starch to phosphodextrins
and 31PNMR spectra of the phosphodextrins 148
8.2.4 Digestibility of CL wheat starch 149
8.2.5 Thermal properties 150
8.2.6 Microscopic observation 150
8.2.7 Scanning electron microscope (SEM) 150
8.2.8 Statistical analysis 150
8.3 Results and Discussion 1518.3.1 Effects of a-amylase/amyloglucosidase digestion
on P content and chemical forms of the
phosphate esters on starch 151
8.3.2 Thermal properties 152
8.3.3 Starch granular morphology before and
after enzyme digestion 153
8.3.4 Digestibility 160
8.4 Conclusions 162
8.5 Acknowledgements 163
8.6 Abbreviations Used in This Chapter 163
References 163
9 Biscuit Baking and Extruded Snack Applicationsof Type III Resistant Starch 167Lynn Haynes, Jeanny Zimeri and Vijay Arora
9.1 Introduction 167
9.2 Thermal Characteristics of Heat-Shear Stable
Resistant Starch Type III Ingredient 168
9.3 Application to Biscuit Baking: Cookies 172
9.4 Cracker Baking 175
9.5 Extruded Cereal Application 1789.5.1 Preparation of extruded RTE cereal and analysis 179
References 189
xii Contents

3GFTOC 07/20/2013 11:21:12 Page 13
10 Role of Carbohydrates in the Prevention of Type 2 Diabetes 191Thomas M.S. Wolever
10.1 Introduction 191
10.2 Background 19110.2.1 Definition of diabetes 191
10.2.2 Types of diabetes 192
10.2.3 Complications of diabetes 192
10.2.4 Prevalence of diabetes 192
10.2.5 Risk factors for type 2 diabetes 193
10.3 Carbohydrates and Risk of Type 2 Diabetes 19310.3.1 Markers of carbohydrate quality 193
10.4 Pathogenesis of Type 2 Diabetes 195
10.5 Effect of Altering Source or Amount of Dietary
Carbohydrate on Insulin Sensitivity, Insulin Secretion
and Disposition Index 197
10.6 Mechanisms by Which Low-GI Foods Improve Beta-Cell
Function 19910.6.1 Glucose toxicity 199
10.6.2 Reduced serum free fatty acids (FFA) 200
10.6.3 Increased GLP-1 secretion 201
10.7 Conclusions 202
References 202
11 Resistant Starch on Glycemia and Satiety in Humans 207Mark D. Haub
11.1 Introduction 207
11.2 Diet and Resistant Starch 208
11.3 Resistant Starch and Insulin Sensitivity 209
11.4 Current Theoretical Mechanism 209
11.5 Satiety 211
11.6 Fermentation and Gut Microbiota 212
11.7 Effect of RS Type 212
11.8 Summary 213
References 213
12 The Acute Effects of Resistant Starch on Appetiteand Satiety 215Caroline L. Bodinham and M. Denise Robertson
12.1 Appetite Regulation 215
12.2 Measurement of Appetite in Humans 216
12.3 Proposed Mechanisms for an Effect of Resistant Starch
on Appetite 217
Contents xiii

3GFTOC 07/20/2013 11:21:13 Page 14
12.4 Rodent Data 218
12.5 Human Data 221
References 225
13 Metabolic Effects of Resistant Starch 229Martine Champ
13.1 Fermentation of RS and Its Impact on Colonic
Metabolism 230
13.2 Resistant Starch, Glycemia, Insulinaemia and Glucose
Tolerance 235
13.3 RS Consumption and Lipid Metabolism 236
13.4 RS Consumption, GIP, GLP-1 and PYY Secretion 238
13.5 RS Consumption, Satiety and Satiation and
Fat Deposition 239
13.6 Conclusion 242
References 244
14 The Microbiology of Resistant Starch Fermentation in theHuman Large Intestine: A Host of Unanswered Questions 251Harry J. Flint
14.1 Introduction 251
14.2 Identifying the Major Degraders of Resistant Starch
in the Human GI Tract 25214.2.1 The human colonic microbiota 252
14.2.2 Cultural studies 252
14.2.3 16S rRNA-based studies 253
14.3 Systems for Starch Utilization in Gut Bacteria 25414.3.1 Bacteroides spp. 255
14.3.2 Bifidobacterium spp. 255
14.3.3 Lachnospiraceae - Roseburia spp., Eubacterium
rectale and relatives 256
14.3.4 Ruminococcaceae 256
14.4 Metagenomics 256
14.5 Factors Influencing Competition for Starch as a Growth
Substrate 257
14.6 Metabolite Cross-Feeding 258
14.7 Impact of Dietary Resistant Starch upon Colonic Bacteria
and Bacterial Metabolites in Humans 259
14.8 Conclusions and Future Prospects 260
Acknowledgements 262
References 262
xiv Contents

3GFTOC 07/20/2013 11:21:13 Page 15
15 Colon Health and Resistant Starch: Human Studiesand Animal Models 267Suzanne Hendrich, Diane F. Birt, Li Li and Yinsheng Zhao
15.1 RS Classification 267
15.2 RS and Colon Health: Overview 267
15.3 RS, Gut Microbes and Microbial Fermentation 26815.3.1 RS and laxation 269
15.3.2 RS, IBS and diverticulosis 270
15.3.3 RS and IBD 270
15.3.4 RS and colon cancer risk – human studies 271
15.4 Colon Cancer Prevention – Animal Models 272
15.5 Conclusions 275
References 275
Index 279
Contents xv

3GFTOC 07/20/2013 11:21:13 Page 16

3GFPREF 07/20/2013 11:29:26 Page 17
Preface
Since the term ‘dietary fibre’ was first coined in 1953, it has undergone
several transformations with respect to its definition, composition, analytical
methodology and physiological effects. Its heterogeneous composition of
naturally-occurring non-starch polysaccharides, lignin and associated substan-
ces has grown to include other synthetic or novel fibres, comprising digestion-
resistant dextrins and resistant starches. Because of this diverse composition,
analysts are often confronted with the challenge of accurately quantifying the
level of total dietary fibre of food or beverage products. Dietary fibre is now
less frequently associated with bulk or regularity and is discussed much more
conspicuously with its role in attenuation of glycemic/insulinemic responses,
blood cholesterol lowering, satiety effects, weight management, large bowel
fermentation and changes in gut microbiota composition and metabolism in
regard to their impact on the general health and well-being of consumers.
Consumer demand for fibre-rich foods and beverages in the United States,
Europe and Asia-Pacific is rising due primarily to the preponderance of
positive epidemiological and scientific data and also an increase in consumer
awareness and support from dieticians and nutritionists. Ironically, however,
many Americans on average consume only about 50–60% of their recom-
mended daily intake of 25 g of fibre.
Resistant starch (RS), in particular, has captivated leading research scien-
tists and prominent educators, and their investigations have been featured
prominently in scientific literature on fibre. Many research activities on RS
highlighted its structure, composition, functionality, in vitro and in vivo
studies and performance in food and beverage products. RS has five types
or classes and, therefore, it provides diverse materials for research investiga-
tors. These, together with the commercial significance of RS, account for the
abundance of published articles and inventions in the scientific and patent
literature. Commercial sources of RS number around 30 – a substantial
increase since the first RS product was introduced to the market in 1993.
xvii

3GFPREF 07/20/2013 11:29:26 Page 18
The idea of writing this book was developed from the Carbohydrate
Division Symposium on resistant starch and health during the 2009 IFT
Annual Meeting in Anaheim, California. The symposium attracted speakers
who are leading researchers and scientists from the academia and the food
industry. In order to capture the important developments in RS, with
emphasis on sources, applications and health benefits, the editors embarked
on a project to write this book using the symposium papers plus the
contribution of invited scientists and academic professionals who excel
in this important area of RS.
There are 15 chapters in the book, covering various topics on RS, such as
its biosynthesis, types or classes, slowly digestible starch, methodology for
measurement and food applications, and also the physiological effects of
RS, primarily in the area of glycemic/insulinemic control, appetite/satiety,
gut microbiota metabolism and large bowel health. This book caters to a
wide audience and can be a valuable resource for students, professors,
research scientists, product developers and other food industry professio-
nals, as they investigate the ever-growing area of RS and its diverse
properties, numerous food and beverage applications, commercial signifi-
cance and physiological effects.
xviii Preface

3GFABOUT 07/20/2013 13:23:15 Page 19
About the Editors
Yong-Cheng Shi, Ph.D. is Professor and Director of the Carbohydrate
Polymers – Technology and Product Innovation group in the Department
of Grain Science and Industry at Kansas State University in Manhattan,
Kansas. He has authored or co-authored more than 40 journal articles and
book chapters and holds more than 15 patents. His research interests include:
structure and properties of starches; physical, chemical, and enzymatic
modifications of starches, biopolymers and flours; carbohydrate and health;
starch digestibility, resistant starch and dietary fibre; ingredient functionality
in cereal products; and developing technologies and products for food,
nutrition, emulsion, encapsulation, pharmaceutical and other industrial
applications.
Dr. Shi received his B.S. in Chemical Engineering from Zhejiang Univer-
sity (Hangzhou, China) and his M.S. and Ph.D. in Grain Science from Kansas
State University (Manhattan, Kansas). He is a professional member of the
American Association of Cereal Chemists International and Institute of Food
Technologists. He is an associate editor forCereal Chemistry and amember of
Advisory Board for Starch and Food Digestion journals.
Clodualdo ‘Ody’ C. Maningat, Ph.D. is Vice President of Applications
Technology and Technical Services at MGP Ingredients, Inc. in Atchison,
Kansas and Adjunct Faculty Member in the Department of Grain Science and
Industry at Kansas State University in Manhattan, Kansas. He is a member and
former chair of the Advisory Board of the Food Processing Center of the
University of Nebraska in Lincoln, Nebraska. He has authored or co-authored
more than 25 journal articles and book chapters in grain and food science
publications and holds more than 30 patents on grain-based technologies. His
research and business interests include: chemistry, modification and function-
ality of starches and proteins; analysis and function of dietary fibres; value-
addition concepts; technology of RS4-type resistant starch; physiological
xix

3GFABOUT 07/20/2013 13:23:15 Page 20
benefits of grain-derived ingredients; and research alliances with scientists and
product developers in the food industry, government and academia.
Dr. Maningat received his B.S. in Chemistry from Adamson University
(Manila, Philippines), his M.S. in Agricultural Chemistry from the University
of the Philippines at Los Banos (Laguna, Philippines) and his Ph.D. in Grain
Science from Kansas State University (Manhattan, Kansas). He is a profes-
sional member of the American Association of Cereal Chemists International,
Institute of Food Technologists, American Society of Baking and American
Chemical Society.
xx About the Editors

3GFCONT 07/20/2013 14:20:4 Page 21
List of Contributors
Geetika AhujaDepartment of Plant Sciences
College of Agriculture &
Bioresources
University of Saskatchewan
Canada
Yongfeng AiDepartment of Food Science and
Human Nutrition
Iowa State University
USA
Vijay AroraIngredient and Process Research
Mondelez International
USA
Diane F. BirtInterdepartmental Graduate Program
in Genetics
Department of Food Science and
Human Nutrition
Nutrition and Wellness Research
Center
Iowa State University
USA
Caroline L. BodinhamDepartment of Nutritional Sciences
Faculty of Health and Medical
Sciences
University of Surrey
UK
Martine ChampINRA, UMR 1280
Physiologie des Adaptations
Nutritionnelles
Universite de Nantes, CRNH,
IMAD, CHU de Nantes, Nantes
France
Ravindra N. ChibbarDepartment of Plant Sciences
College of Agriculture &
Bioresources
University of Saskatchewan
Canada
Annette EvansInnovation and Commercial
Development
Tate & Lyle
USA
xxi

3GFCONT 07/20/2013 14:20:4 Page 22
Harry J. FlintMicrobial Ecology Group
Rowett Institute of Nutrition and
Health
University of Aberdeen
Aberdeen, UK
Bruce R. HamakerWhistler Center for Carbohydrate
Research and Department of Food
Science
Purdue University
USA
Jovin HasjimQueensland Alliance for Agriculture
and Food Innovation
Centre for Nutrition and Food
Sciences
The University of Queensland
Australia
Mark D. HaubDepartment of Human Nutrition
Kansas State University
USA
Lynn HaynesIngredient and Process Research
Mondelez International
USA
Suzanne HendrichInterdepartmental Graduate Program
in Genetics
Department of Food Science and
Human Nutrition
Nutrition and Wellness Research
Center
Iowa State University
USA
Sarita JaiswalDepartment of Plant Sciences
College of Agriculture &
Bioresources
University of Saskatchewan
Canada
Jay-lin JaneDepartment of Food Science and
Human Nutrition
Iowa State University
USA
Hongxin JiangDepartment of Food Science and
Human Nutrition
Iowa State University
USA
Li LiInterdepartmental Graduate Program
in Genetics
Department of Food Science and
Human Nutrition
Nutrition and Wellness Research
Center
Iowa State University
USA
Clodualdo C. ManingatMGP Ingredients
Inc., USA; Department of Grain
Science and Industry
Kansas State University
USA
Barry V. McClearyMegazyme International
Bray Business Park
Ireland
xxii List of Contributors

3GFCONT 07/20/2013 14:20:5 Page 23
M. Denise RobertsonDepartment of Nutritional
Sciences
Faculty of Health and Medical
Sciences
University of Surrey
UK
Paul A. SeibDepartment of Grain Science and
Industry
Kansas State University
USA
Yong-Cheng ShiCarbohydrate Polymers –
Technology and Product
Innovation
Department of Grain Science and
Industry
Kansas State University
USA
Radhiah ShukriDepartment of Grain Science and
Industry
Kansas State University
USA
Thomas M.S. WoleverDepartment of Nutritional Sciences
University of Toronto
Canada; Division of Endocrinology
and Metabolism
St. Michael’s Hospital
Canada
Genyi ZhangSchool of Food Science and
Technology
Jiangnan University
China
Yinsheng ZhaoInterdepartmental Graduate Program
in Genetics
Department of Food Science and
Human Nutrition
Nutrition and Wellness Research
Center
Iowa State University
USA
Jeanny ZimeriIngredient and Process Research
Mondelez International
USA
List of Contributors xxiii

3GFCONT 07/20/2013 14:20:5 Page 24

3GFACKNOW 07/20/2013 13:31:42 Page 25
Acknowledgements
We are profoundly grateful to the chapter authors for their expertise and their
valuable contributions to make this book a reality. This is a tribute to their hard
work and the countless hours devoted in writing the chapters. A number of
scientists and academicians, to whom we extend sincere thanks, volunteered
their time to review and provide critique to the book’s contents. They are as
follows: Mike Gidley (University of Queensland), Ya-Jane Wang (University
of Arkansas), David Robbins (University of Kansas Medical Center), Jens
Walter (University of Nebraska, Lincoln), M. Denise Robertson (University of
Surrey), Paul A. Seib (Kansas State University), Steve Pickman (Consultant)
and Annette Evans (Tate & Lyle). The patience, accommodating attitude
and excellent editorial assistance of Mr. David McDade, Ms. Becky Ayre,
Mr. Sharib Asrar, Ms. Jasmine Chang and other Wiley staff are also gratefully
acknowledged.
xxv

3GFACKNOW 07/20/2013 13:31:43 Page 26

3GCH01 07/19/2013 16:58:7 Page 1
1 Starch Biosynthesis in Relationto Resistant Starch
Geetika Ahuja, Sarita Jaiswal andRavindra N. ChibbarDepartment of Plant Sciences, College of Agriculture & Bioresources,University of Saskatchewan, Canada
1.1 INTRODUCTION
1.1.1 Starch components
Starch is present in amyloplasts as semi-crystalline intracellular water-
insoluble granules, with alternating crystalline and amorphous layers. Starch
is a glucan homopolymer composed of one-quarter amylose (molecular mass
105–106Da) and three-quarters amylopectin (molecular mass 107–109Da),
along with traces of lipids (0.1–1.0%) and proteins (0.05–0.5%). Amylose is
essentially a linear glucan polymer, composed of a-1,4 linked glucose
residues with a degree of polymerization (dp) ranging between 800 (in maize
and wheat) to more than 4500 (in potato) with sparse branching
(approximately one branch per 1000 residues) (Morrison & Karkalas,
1990; Alexander, 1995). Structural and functional aspects of these glucan
polymers affect starch functionality and its end use.
Amylose chains are capable of forming single or double helices. On
the basis of orientation of its fibres in X-ray diffraction studies, amylose
can be divided into A- and B-type allomorphs (Galliard et al., 1987). In B-type
allomorph, six double helices are packed in an anti-parallel hexagonal
mode surrounding the central water channel (36 H2O per unit cell). In
A-type, the central water channel is replaced by another double helix, making
the structure more compact. In this allomorph, only eight molecules of water
per unit cell are inserted between the double helices (Galliard et al., 1987).
Amylopectin is a highly branched glucan polymer, in which a-1,4 linked
glucose residues are interspersed with a-1,6-glucosidic linkages (4–5%)
1
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH01 07/19/2013 16:58:7 Page 2
which introduce branches, and a degree of polymerization ranging from
105–107 glucose units (Myers et al., 2000). Chain lengths of 20–25 glucose
units between branch points are typical. The branches in the amylopectin
molecule are arranged in clusters (Bul�eon et al., 1998).
An amylopectin molecule typically consists of three types of chains, which
are either located within a single cluster or connect two or more clusters
(Hizukuri et al., 1986; Thompson et al., 2000). In amylopectin, only theC-chain
has a reducing end oriented towards the centre or hilum of the granule. Attached
to the C-chain with a-1,6 linkages are the B-chains. These can support other
B- or A-chains. The A-chains are the outermost chains, which do not support
any other chains. A- and B-chains form clusters and B-chains can span and
supportmultiple clusters.A-chains typically consist of 6–12 glucosemolecules,
while B-chains may contain 13–24 or up to 50 or more glucose molecules,
depending on the number of clusters they span. In the section which does not
contain a-1,6 branch points, two neighbouring glucose chains form a double
helix, and these double helices form a crystalline pattern. All of these structures
are attached by hydrogen bonds. The sections where the branch points of the
amylopectin are located are amorphous and contain amylose molecules.
1.1.2 Resistant starch
More than 50% of calorific requirement of human diet is fulfilled by starch-
based foods, and the quality and quantity of starch-based food affect overall
blood glucose and homeostasis in humans. Starch digestion in humans is
initialized by salivary a-amylases in the oral cavity, followed by pancreatic
a-amylase and the intestinal brush border glucoamylases, maltase-glucoa-
mylase, and sucrase-isomaltase (Nichols et al., 2003). Brush-border enzymes
convert the resultant products of digestive process into maltase-glucoamylase
and sucrase-isomaltase, which enter the vascular system (Lehmann & Robin,
2007). Based on its in vitro enzymatic hydrolysis, the rate of glucose release
and its absorption in the gastrointestinal tract, starch is classified as either
readily digestible starch (RDS), slowly digestible starch (SDS) or resistant
starch (RS) (Englyst et al., 1992).
According to Englyst et al. (1992), based on in vitro kinetic assay, RDS is
broken down into glucose molecules in �20 minutes, while SDS is the
fraction which gets digested in �100 minutes. Both RDS and SDS are
completely digested in the small intestine. RS is referred to that portion of
starch which is not hydrolyzed until about 120 minutes have elapsed. It passes
through the small intestine undigested, but is fermented in the large intestine
by gut microflora. Physiological benefits of RS include hypoglycaemic effects
and production of short chain fatty acids (SCFA) particularly butyrate, which
2 Resistant Starch

3GCH01 07/19/2013 16:58:7 Page 3
has been reported to lower lumen pH, making it a less conducive environment
for cancer and other diseases (Topping & Clifton, 2001; Wei et al., 2010).
A medium-to-high amount of SDS has been reported for native normal
maize starch (Axelsen et al., 1999), waxy starches (Weurding et al., 2001),
millet and sorghum (Benmoussa et al., 2006) and legumes (Hoover & Zhou,
2003). A few researchers have reported a higher rate of digestibility for cereal
starches than tuber starches such as potato (Fannon et al., 1992; Benmoussa
et al., 2006). On the basis of its botanical source, physical or chemical
processing, RS can be divided into four types. RS1 is physically inaccessible
due to its location in the food, RS2 escapes digestion because of its granular
structure, RS3 is retrograded starch and RS4 is chemically modified starch
(Brown, 2004).
1.2 FACTORS AFFECTING STARCH DIGESTIBILITY
Starch enzymatic hydrolysis and RS are influenced by several factors, both
extrinsic and intrinsic properties of starch granules. Extrinsic factors, which
include starch granule surface characteristics such as porosity of granule and
pit formation between the surface and centre of the granule (Fannon et al.,
1992), or exo-corrosion (Gallant et al., 1997), affect starch digestibility.
Intrinsic properties of starch granules, such as packing of amorphous and
crystalline regions (Gallant et al., 1992; Zhang et al., 2006), or interaction of
amylose with other components such as lipids (Crowe et al., 2000), proteins
(Escarpa et al., 1997) and/or enzyme inhibitors (Bjorck et al., 1987), also
influence starch digestibility. Reduced digestibility of tuber starch granules
has been attributed to their large and smooth surface, along with their surface
properties.
The amylose to amylopectin ratio is an important determinant of starch
digestibility. Amylose and amylopectin have different structural and physio-
logical characteristics and, hence, exhibit different reactions within the body
during digestion and subsequent release of glucose molecules for absorption.
The amylose to amylopectin ratio is a major determinant for RS2 and RS3
(Sajilata et al., 2006).
A positive correlation exists between amylose concentration and RS
formation (Ito et al., 1999). The straight chains of amylose limit the access
of small intestine b-amylases to the two terminal glucose units on the
amylose chain (besides, two terminal ends may not be accessible due to
folding of a polymer). In contrast, the highly branched structure of amylo-
pectin provides multiple terminal end glucose units that b-amylases can
access readily.
Starch Biosynthesis in Relation to Resistant Starch 3

3GCH01 07/19/2013 16:58:7 Page 4
During cooking, starch is gelatinized and amylose molecules are leached
out of the swollen starch granules as coiled polymers which, on cooling,
associate as double helices and form hexagonal networks which resist
digestion. In waxy starches, instead of this network, aggregate formation
occurs, and this is more susceptible to hydrolysis by amylases.
The intensity of starch digestion is also affected by the degree of polymeri-
zation and/or branching of glucan polymers, i.e. a reduction in the rate of
hydrolysis with increased branching due to steric hindrance (Park & Rollings,
1994). Gamma irradiation-generated rice mutants high in RS showed
increased proportion of short chains with DP� 12, decreased proportion of
intermediate chains of 13�DP� 36 and decrease in long chains with
DP� 37 (Shu et al., 2007).
Another report, by Ao et al. (2007), mentions that b-amylase and malto-
genic a-amylase mediated partial reduction of outer branch chains of amylo-
pectin reduces overall starch digestion rate, which was related to an increase in
the amount of a-1,6 linkages and decrease in a-1,4 linkages. Changes in the
amylopectin chain length distributions facilitated retrogradation to produce
B- and V- type crystalline structures, leading to more resistant starch. It is
generally believed that increased proportion of longer chains makes the starch
more resistant to digestion. A possible reason could be that longer chains form
longer and more stable helices, which are further stabilized by hydrogen
bonds distributed over the entire crystalline region and cause decreased
digestibility (Lehmann & Robin, 2007).
1.3 STARCH BIOSYNTHESIS
Plants have a unique ability to capture light energy and to fix carbon dioxide
and water to form triose sugars that act as precursor of simple and complex
carbohydrates. Photosynthesis in the plants’ chloroplast results in the produc-
tion of triose-phosphates, reducing equivalents and ATP. The triose phos-
phates are either transported by triose-phosphate transporters to the cytosol, or
are converted to phosphorylated compounds, including fructose-6-phosphate
in the plastid. During the light period, chloroplasts synthesize transitory starch
which, at night, is broken down into constituent sugars and transported to the
storage organs. In contrast, in amyloplasts, these precursors are used to
synthesize storage starch.
Analogous to chloroplasts in green tissues, storage organs contain amy-
loplasts which are albino plastids and devoid of internal membrane structure.
These specialized plastids act as processing and storage unit for starch in plant
cells. Fructose-6-phosphate in chloroplasts is used both for regeneration of
4 Resistant Starch

3GCH01 07/19/2013 16:58:7 Page 5
ribulose-1,5-bisphosphate and production of glucose-1-phosphate through
glucose-6-phosphate. Conversion of glucose-1-phosphate and ATP to
ADP-glucose by ADP-glucose pyrophosphorylase (AGPase) is the first
committed step in starch synthesis.
In addition to AGPase, other enzymes involved in the starch (especially
amylopectin) biosynthetic cascade include starch synthases (SS), starch branch-
ing enzymes (SBE) and debranching enzymes (DBE) (Smith et al., 2001; James
et al., 2003; Zeeman et al., 2010). Amylose is synthesized exclusively by
granule-bound starch synthase-I (GBSSI). The glucose moiety from ADP-
glucose is used to elongate an already existing glucan chain. Starch synthases
catalyze the formation of a-1,4 glucosidic linkage between the glucose units toform a linear chain. SS require a primer for elongation of glucose chain.
The initiation of glucan polymerization reaction is poorly understood. One
hypothesis suggests the presence of glycogenin-like self glycosylating protein
as primer for amylopectin synthesis and addition of D-glucose occurs to the
non-reducing end of a growing glucan chain (Chatterjee et al., 2005). Another
hypothesis is the de novo synthesis of glucan chains mediated by a two-site
insertion mechanism. Two glucose units from ADP-glucose complex with the
active site of starch synthase, and are subsequently added to the reducing end
of glucan chain (Mukerjea & Robyt, 2005).
Four starch synthase isoforms (SSI, SSII, SSIII, SSIV) play important role
in elongating different regions of amylopectin. Therefore, alterations in SS
activities would affect the amylopectin fine structure. Branches in amylopec-
tin and amylose are introduced by SBE, which catalyze the cleavage of an
a-1,4 linkage and join the cleaved chain to another glucan chain through a-1,6glucosidic linkage. Two classes of SBE (i.e. SBEI and SBEII) exist, which
have different substrate specificities.
Finally, debranching enzymes (isoamylase and pullulanase) act to trim the
outer branches of amylopectin molecule to form ordered branch structure and
packaging of the molecule into starch granules. Since multiple isoforms of
starch biosynthetic enzymes exist in the endosperm and have specific func-
tions, mutations in any of these genes would therefore lead to a change in
starch content, structure and functional properties.
In addition to the core enzymes, other enzymes, such as phosphorylases,
disproportionating enzymes and dikinases (glucan water dikinase, phos-
phoglucan water dikinase) also play important roles in starch metabolism.
Starch phosphorylation involves dikinases such as glucan water dikinase
(GWD, mol wt 155 kDa) and phosphoglucan water dikinase (PWD, mol wt
130 kDa), which phosphorylate the C6 and C3 positions of glucose units of
amylopectin, respectively – an important factor in starch degradation
(Fettke et al., 2009).
Starch Biosynthesis in Relation to Resistant Starch 5

3GCH01 07/19/2013 16:58:7 Page 6
1.4 STARCH BIOSYNTHESIS IN RELATION TO RS
1.4.1 ADP-glucose pyrophosphorylase (AGPase)
AGPase catalyzes the synthesis of ADP-glucose from ATP and glucose-
1-phosphate. It is the first step in starch biosynthesis, and AGPase is also a key
regulatory enzyme in the starch biosynthetic pathway. AGPase consists of two
large and two small subunits, which affect allosteric and catalytic properties of
the enzyme. Allosteric regulation of this enzyme plays a critical role in
determining the amount of starch produced (Hannah & James, 2008). AGPase
is allosterically activated by 3-phosphoglyceric acid (3-PGA) and inhibited by
inorganic phosphate (Pi) in many plant tissues (Preiss et al., 1996). Genetic
and biochemical manipulation of its sensitivity towards Pi resulted in increase
in crop productivity (starch yield) due to increased sink strength (Wang et al.,
2007; Sakulsingharoj et al., 2004; Smidansky et al., 2002). AGPase activity is
also redox regulated (Hendriks et al., 2003).
In general, the active form of AGPase is present in the plastids of mature
cereal tissues and sink tissues of non-cereal plants. Developing cereals
however, differ, with most of their AGPase activity localized mainly in the
cytosol of endosperm cells. Specific transporters/ADP-glucose transporter
channels are involved in the trafficking of the resultant ADP-glucose. In non-
cereal plants, the sucrose to starch pathway comprises plastid import of
hexose phosphates, which can be used in other biosynthetic processes in
addition to starch synthesis. In contrast, in cereals, carbon entering the plastid
as ADP-glucose is committed to starch synthesis (James et al., 2003).
MutationsinAGPaseandADP-glucosetransportershavebeenshowntoaffect
the total starch content in maize, barley, pea and potato (Hylton et al., 1992;
Shannon et al., 1998; Tjaden et al., 1998; Patron et al., 2004). The maize
Shrunken-2 and Brittle-2 mutants have lesions in the large and small subunits
of the cytosolic AGPase, respectively (Hannah & Nelson, 1976). Shrunken-2
mutant kernels are deeply dented, with floury endosperm that has 25% reduced
starch but is sweet due to high sucrose concentration (Hutchinson, 1921).
Similarly Brittle-2 mutant kernel germinates poorly, is dark and shrunken and
has25–34% lower starch thannormal (Preiss et al., 1990).Abarleymutant,Risø
16, isassociatedwithadeletioninthesmall subunitofcytosolicAGPaseresulting
in reduced starch concentration and seed weight (Johnson et al., 2003). These
changes in starch concentration have not been associated to RS (Table 1.1).
1.4.2 Starch synthases (SS)
Starch synthases catalyze the transfer of glucose unit from ADP-glucose to
non-reducing end of an already existing glucan chain, thus forming a-1,4
6 Resistant Starch


3GCH01 07/19/2013 16:58:9 Page 8
linkage. Cereal endosperms contain at least five SS classes, based on their
conserved primary amino acid sequences. SSI and SSII are present mostly in
the stroma (Fujita et al., 2006), whereas SSIII and SSIVare present both in the
stroma and starch granule (Denyer et al., 1995; Dai, 2010) and are primarily
involved in amylopectin synthesis. GBSSI is bound to starch granules and is
required for amylose synthesis. Recently, GBSSI has also been shown to
participate in the elongation of amylopectin chains, particularly for very long
branches (Yoo & Jane, 2002). The chain elongation pattern differs for each
isoform and varies with plant species (Smith et al., 1997). In addition to their
specialized functions, some SS overlap in their functional role, while others
are unique (Rold�an et al., 2007).
1.4.2.1 Granule bound starch synthase-I
GBSSI (also known as waxy protein) present in the interior of starch granule is
essential for amylose synthesis. Plants lacking GBSSI enzymatic activity
produce starch without amylose, which is also called waxy starch. In wheat,
GBSS has two isoforms, GBSSI and GBSSII (Nakamura et al., 1998; Vrinten
& Nakamura, 2000). Another isoform, GBSSIb, exclusive to the pericarp
region, has been reported in barley (James et al., 2003). This is involved in
transient starch accumulation, which enhances the sink strength of the young
caryopsis (Patron et al., 2002).
In vitro study using ADP[14C] glucose as precursor of starch biosynthesis
in isolated starch granules showed uptake of malto-oligosaccharides of DP
2–7 by GBSSI as primers for amylose synthesis (Denyer et al., 1996).
GBSSI is also reported to be involved in the elongation of long chains of
amylopectin (Yoo & Jane, 2002; Craig et al., 1998). GBSSI elongates the
glucan chains which are confined to the semi-crystalline region of the
granule and cannot form branches. Consequently, the chains remain linear
and are known as amylose, or long-branch chains of amylopectin (Jane
et al., 2010).
1.4.2.1.1 Amylose in relation to RS formation
Amylose contributes to the formation of RS2 and RS3. Deficiency of GBSS1
activity produces starchmade of only amylopectin (waxy starch). Rate of starch
digestibility is high inwaxy andpartiallywaxy starch (reducedRS) compared to
normal starch from several plants (Rooney & Plugfelder, 1986; Bertoft et al.,
2000; Li et al., 2004; Chung et al., 2006; Asare et al., 2011). In a recent study on
starch structure and in vitro enzymatic hydrolysis using barley atypical amylose
concentration starch (Table 1.1), Asare et al. (2011), using atomic force
microscopy, reported high poly-dispersity indices for normal (1.4) and
8 Resistant Starch

3GCH01 07/19/2013 16:58:9 Page 9
increased amylose starch genotype (1.25), compared to near (partially) waxy
starch genotypes (0.33). They also concluded that energy requirement for
gelatinization and hydrolysis of waxy starch is lower than for normal or
high-amylose starch.Waxy starches aremore susceptible to hydrolytic enzymes
compared to starch granules with significant amylose concentration.
Hu et al. (2004) investigated three types of rice cultivars with varying
amylose content for in vitro hydrolysis and glycemic index determination.
They concluded quicker, complete and significantly higher rates of starch
hydrolysis for waxy and low-amylose rice than for intermediate and high-
amylose rice. In a more practical approach for estimating RS contribution for
amylose, Hung et al. (2005) substituted high-amylose wheat flour for normal
wheat flour in bread-making and observed higher RS content in the substituted
bread. Physical increase in amylose content through retrogradation
and extended cooling after cooking can also lower digestibility (Blazek &
Copeland, 2010).
1.4.2.2 Starch synthase-I
In maize, SSI is responsible for extending shorter A and B1 chains up to a
critical chain length, making it unsuitable for its own catalysis (Commuri &
Keeling, 2001). In rice, retrotransposon Tos17 insertion mediated SSI-defi-
cient mutant lines showed starch phenotype with decreased amylopectin
chains of DP8–12, but increased chains of DP6–7 and 16–19. This suggests
that SSI functions in generating DP8–12 chains from shorter chains of DP6–7
emerging from the branch point of A and B1 chains (Fujita et al., 2006).
Amylose synthesis was not affected by this mutation, and its effect on starch
hydrolysis has not been reported.
1.4.2.3 Starch synthase-II
In cereal endosperm, SSII synthesizes intermediate-length branch chains of
amylopectin (see review by Jane et al., 2010). Yamamori et al. (2000)
produced triple null wheat line lacking starch granule protein-1 (SGP1),
identified as SSIIa and homologous to maize SSIIa (Li et al., 1999). Lack of
SGP1 showed amylopectin with increased short chains of DP 6–10, a decrease
in intermediate chains of DP 11–25 and a concomitant increase in apparent
amylose concentration (30.8–37.4%).
In a subsequent study (Yamamori et al., 2006), wheat lines lacking SGP1
showed an increase in resistant starch level (3.6%) compared to normal wheat
(0.02%). In a similar approach, wheat lines deficient in SSII A and B genome
polypeptides resulted in increased amylose (32%) starch, as determined by
Starch Biosynthesis in Relation to Resistant Starch 9

3GCH01 07/19/2013 16:58:9 Page 10
HP-SEC analyses (Chibbar & Chakraborty 2005; Lan et al., 2008). SSIIa
deficient maize (sugary2 mutation due to insertion in SSIIa) genotypes
showed an increase in abundance of short (DP 6–11) and medium (DP
13–25) chains. This mutation also resulted in an increase in apparent amylose
concentration from 26–40% (Zhang et al., 2004). In rice, japonica type has a
higher short to long chains ratio than indica type but, contrary to wheat and
maize, indica rice has higher amylose concentration than japonica rice
(Umemoto et al., 1999, 2002).
In barley, sex6mutation on chromosome 7H due to G!A transition results
in an early stop codon, thus inhibiting C-terminal translation of the active site
of SSIIa (Morell et al., 2003). The major effect of SSIIa inactivity is an
increase in amylose concentration (65–70%) in the mutants, which increases
RS content. In addition, a change in starch crystallinity from A-type to a
mixture of B- and V-type was also reported. V-type crystallinity indicates the
formation of amylose-lipid complexes, which inhibit starch swelling, and it
resists digestion by amylolytic enzymes (Morell et al., 2003). A barley
cultivar, Himalaya-292, which has an inactive SSIIa, produces increased
amylose starch and higher RS content. This RS-rich diet when fed to rats
changes its bowel SCFA (Bird et al., 2004).
A similar pattern of change with the SSII mutation on amylopectin fine
structure and amylose content has been reported in potato (Edwards et al.,
1999) and pea (Craig et al., 1998). SSIIa mutation in pea rug5 decreases
intermediate length amylopectin chains (B2 and B3) and produces a higher
(�35%) amylose concentration starch Table (1.1) (Craig et al., 1998).
1.4.2.4 Starch synthase-III
Amylopectin long B-chains are synthesized by SSIII. Mutation in maize SSIII
is called dull-1 (du1), which has a starch phenotype of amylopectin with
decreased proportion of long B-chains, enriched short-branch chains and
moderately increased amylose content (Wang et al., 1993). SSIII mutation
also affects SSII and SBEIIa and is capable of altering endosperm starch
structure (Gao et al., 1998). Ryoo et al. (2007) reported a mutation in rice
SSIII OsSSIIIa (floury, flo), which produced small and round starch granules
and endosperm with a loosely packed central portion, exhibiting a floury-like
phenotype. In rice flo mutant lines, amylopectin chains with DP� 30 were
reduced, suggesting that OsSSIIIa has a role in the generation of relatively
longer chains of amylopectin (i.e. B2 and B3 to B4). Concomitantly, a 2–4%
increase in the ratio of amylose to amylopectin was also observed.
In addition to its role in extending glucan chains, SSIII influences
starch structure through its association with other starch metabolizing
10 Resistant Starch

3GCH01 07/19/2013 16:58:9 Page 11
enzymes. Arabidopsis SSIII mutants AtSSIII1 and AtSSIII2 showed
increased starch concentration compared to wild type, suggesting a nega-
tive regulatory role of SSIII in biosynthesis of transient starch (Zhang
et al., 2005). However, no report is available on the effect of SSIII
mutation on starch digestibility.
1.4.2.5 Starch synthase-IV
In rice, two SSIV genes, SSIVa and SSIVb, have been shown to be expressed
during grain filling, both in the pericarp and the endosperm (Hirose & Terao,
2004). Arabidopsis SSIV mutants show a reduction in leaf starch concentra-
tion (Rold�an et al., 2007) and a striking reduction in leaf starch granules,
which suggests a role for SSIV in starch granule initiation. Recently, it has
been shown in an in vitro assay that SSIV has high SS activity when malto-
triose is used as primer (Szydlowski et al., 2009). To date, no cereal plants
deficient in SSIV activity have been characterized.
1.4.3 Starch branching enzymes (SBE)
Starch branching enzymes cleave a-1,4 linkages and transfer a free reducing
C-1 to C-6 hydroxyl group of glucose-unit in another chain, forming a new
a-1,6 branch linkage. Since branching is an essential part of amylopectin
synthesis, it will therefore be dependent on the available concentration of
needed SBE.
Based on primary amino acids sequence similarity and substrate specific-
ity, two major types of SBE (SBEI and SBEII) have been identified in cereals.
In vitro studies in maize suggest that SBEI prefers amylose as substrate and
transfers longer chains, whereas SBEII uses amylopectin as substrate and
transfers shorter chains (Guan & Preiss, 1993). In wheat, SBEII is further
divided into two�85% similar isoforms, SBEIIa and SBEIIb, with apparently
similar molecular weight (Rahman et al., 2001). In addition to this, a larger
form of SBEI, SBEIc (152 kDa) has been reported in wheat (Ba�ga et al.,
2000), which is preferentially associated with large A-type granules (Peng
et al., 2000). In dicots like pea and potato, two isoforms of SBE viz. SBEI and
SBEII (or, SBE B and SBE A) have been reported (Burton et al., 1995;
Poulsen & Kreiberg, 1993).
In maize, mutation in SBEIIb resulting in high-amylose starch is known as
amylose-extender (ae) (Stinard et al., 1993). This results in cereal starch with
high-amylose concentration (>50%) and amylopectin with more long branch-
chains and fewer short branch-chains (Jane et al., 1999). Similarly, another
report suggested a higher proportion of long chains (DP� 38) and a marked
Starch Biosynthesis in Relation to Resistant Starch 11

3GCH01 07/19/2013 16:58:9 Page 12
reduction in short chains of DP� 17 in ae rice endosperm (Nishi et al., 2001).
It also showed a significant increase in apparent amylose concentration from
25–35%.
The very long chains of ae mutant amylopectin develop B-type crystallin-
ity (Kasemsuwan et al., 1995; Hizukuri et al., 1983), which favour slow
enzymatic digestion. These results corroborated a similar study in maize (Li
et al., 2008), where aemutants showed significant increase in chain lengths of
amylopectin and higher apparent and absolute concentrations of amylose.
Further, the mutants also showed considerably higher RS content (39.4–
43.2%) compared to the parents (11.5–19.7%). A commercial product con-
taining�80% amylose, called Hi-maize, has been derived from this mutation.
Hi-maize has been added to wheat products to increase RS amount (Brown,
2004).
In a recent study, barley RNAi mediated inhibition of SBEIIa and SBEIIb
activity altered starch composition and structure (Regina et al., 2010). The
study revealed that a reduction in expression of both SBEIIaþ b to >80%
elevated the amylose content to >65% from 28% in wild type resulting in a
significant increase in RS content (Table 1.1). However, they observed minor
differences when either enzyme was down-regulated. Also, reduction in
expression of both SBEIIaþ b showed an increase in the proportion of chains
of DP<9 and DP>15 and a consequent decrease in the number of medium
chains (DP9–13).
A similar trend has previously been reported in wheat, where an increase in
amylose content (<70%) in SBEIIa mutants was observed by simultaneous
inhibition of expression of both the SBE II isoforms (Regina et al., 2006). In
addition, decrease in proportion of amylopectin chains of DP4–12 and an
increase in chains of DP> 12 was also seen. In vivo feeding studies in rats
using high-amylose wheat meal showed higher amount of RS and lower
glycemic index in comparison to wild type wheat diet (Regina et al., 2006). In
potato, inhibition of SBE A and SBE B resulted in a very high-amylose
phenotype (up to �89% by potentiometric determination), while normal high
molecular weight amylopectin was absent (Schwall et al., 2000). This type of
starch would have lower digestibility.
Yao et al. (2009) studied four corn types with different doses of amylose-
extender(ae) and floury-1(fl1) alleles in the endosperm. Amylose and RS
contents followed a similar pattern with highest values in aeaeaeae (amylose
¼ 58.3%; RS¼ 55.2%). They also observed higher proportion of longer
branch chains with DP� 25 in these mutants. Since amylose-extender
mutation reduces SBEIIb activity, it results in an increase in amylose to
amylopectin ratio, which in turn increases RS content.
12 Resistant Starch

3GCH01 07/19/2013 16:58:9 Page 13
1.4.4 Starch debranching enzymes (DBE)
Final packaging of the starch granule requires the trimming of extra branches.
Debranching enzymes have been postulated to play this important role in
amylopectin biosynthesis (Ball et al., 1996; Myers et al., 2000; Nakamura
et al., 2002). Two different mechanisms for DBE mode of action have been
proposed. The ‘preamylopectin-trimming model’ suggests that the outer
branches of preamylopectin molecules are trimmed by DBE to facilitate
chain elongation by SS (Mouille et al., 1996; Myers et al., 2000). This will
form amylopectin with an ordered branch structure and allow packaging of the
molecule in starch granules. In addition, glucan chains released by DBE’s
action on amylopectin can be elongated by GBSSI to form the amylose
fraction.
According to the ‘soluble glucan recycling model’, DBE participates in
degradation of short chain glucan molecules produced either by SS or SBE
action to prevent accumulation of highly branched soluble polymers at the
expense of amylopectin formation (Zeeman et al., 1998; Smith, 2001).
Endosperms deficient in DBE activity by lesions in DBE genes result in
the formation of phytoglycogen instead of amylopectin from soluble glucans
(Zeeman et al., 1998).
Two major DBE classes are recognized: isoamylases, which trim packed
structures (like glycogen); and pullulanases, which act on more open struc-
tures (like pullulan). Three types of isoamylases have been identified in cereal
endosperm (Kubo et al., 2005) and in potato (Hussain et al., 2003). Lack of
isoamylase-1 in rice (sugary-1, su-1), and barley (isa-1) resulted in small but
significant alteration in amylopectin chain length distribution (Kubo et al.,
2005). In mutant lines, starch granules were shrunken, irregular and com-
pound (reviewed in James et al. (2003)).
Pullulanase type DBEmutation is termed ZPU1 in maize. ZPU1 is an endo-
acting enzyme that cleaves only very short branch chains and it is activated by
redox status and inhibited by high sugar (Dinges et al., 2003). A similar report
on wheat limit-dextrinase-type-DBE activity suggests its redox regulation
(Repellin et al., 2008). Mutations in debranching enzymes, however, have not
been reported to be associated with resistant starch (Table 1.1).
1.5 CONCLUDING REMARKS
Starch biosynthesis is a complex process in which starch biosynthetic
enzymes act in a coordinated manner to produce amylopectin, which is
Starch Biosynthesis in Relation to Resistant Starch 13

3GCH01 07/19/2013 16:58:9 Page 14
architecturally conserved in starches from different botanical sources.
Genetic strategies, by identifying genotypes with lesion(s) in gene(s)
encoding starch biosynthetic enzymes, have revealed the role of each
enzyme or its isoform in the synthesis of amylose and amylopectin
constituent glucan chains and consequent alteration in starch composition
and amylopectin architecture.
It has also been found that mutations in one locus in starch biosynthetic
pathway affects one or more other starch biosynthetic enzymes. Maize ae
mutant has a lesion in SBEIIb gene, but SBEI activity is reduced or absent
and changes the properties of an isoamylase type DBE (Colleoni et al.,
2003). Conversely, genetic lesions in pullulanase (zpu-204) or isoamylase
(sul-si) type DBE reduce SBEIIa activity, although SBEIIa polypeptide is
not altered or reduced (James et al., 1995; Dinges et al., 2003). Lesions in
SSII genes which reduce SSII activity also reduce/eliminate the binding of
SSI, SBEIIa and SBEIIb within the granule matrix, although these enzymes
have not lost their affinity to amylopectin or starch (Morell et al., 2003;
Umemoto & Aoki, 2005). These observations suggest that key starch
biosynthetic enzymes form protein complexes (Tetlow et al., 2004). Using
isolated amyloplasts, starch biosynthetic enzyme complexes have been
shown in wheat and maize (Tetlow et al., 2004; Hennen-Bierwagen
et al., 2008).
In a recent proteomics study, it has been shown that phosphorylation of
GBSSI, SBEIIb and Pho 1 is needed for their incorporation in to starch
granules (Grimaud et al., 2008). The concept of starch biosynthetic enzymes
acting in a complex and its formation is dependent upon the phosphorylation
status of constituent enzymes and is an additional level of control in starch
biosynthesis.
There is significant interest in increasing amylose concentration in cereal
and tuber starches. Increased amylose concentrations have been attributed to
both SBE and SS isoforms. In addition to natural mutants in maize (ae) and
barley (sex6), amylose to amylopectin ratios in starch have been manipulated
by altering GBSSI and SBEII (waxy/amylose extender) activity in wheat
(Lafiandra et al., 2010; Sestili et al., 2010; Regina et al., 2006), in maize
(Jiang et al., 2010) and in rice (Wei et al., 2010). In wheat and barley, very
high amylose concentrations were obtained by RNAi mediated inhibition of
Sbe2a and Sbe2b genes (Regina et al., 2006, 2010). Recent advances in
understanding starch biosynthesis, combined with innovations in genomics
(Ganeshan et al., 2010), can be used to develop cereal genotypes with
increased amylose concentrations and alteration in amylopectin architecture
which can be used to produce RS.
14 Resistant Starch

3GCH01 07/19/2013 16:58:9 Page 15
ACKNOWLEDGEMENTS
Canada Research Chairs, Natural Science and Engineering Research Council,
Saskatchewan Agriculture Development Fund, Saskatchewan Pulse Growers,
and Brewing and Malting Barley Research Institute, Winnipeg are gratefully
acknowledged for supporting seed carbohydrate research in our laboratory.
REFERENCES
Alexander, R.J. (1995). Starch in plastics. Cereal Foods World 41, 426–427.Ao, Z., Simsek, S., Zhang, G., Venkatachalam, M., Reuhs, B.L., Hamaker, B. (2007). Starchwith a slow digestion property produced by altering its chain length, branch density, andcrystalline structure. Journal of Agricultural and Food Chemistry 55, 4540–4547.
Asare, E.K., Jaiswal, S., Maley, J., Ba�ga, M., Sammynaiken, R., Rossanagel, B.G.,
Chibbar, R.N. (2011). Barley grain constituents, starch composition and structure affectstarch in vitro enzymatic hydrolysis. Journal of Agricultural and Food Chemistry 59,4743–4754.
Axelsen, M., Arvidsson, L.R., Lonnroth, P., Smith, U. (1999). Breakfast glycaemicresponse in patients with type 2 diabetes: effects of bedtime dietary carbohydrates.European Journal of Clinical Nutrition 53, 706–710.
Ba�ga, M., Nair, R.B., Repellin, A., Scoles, G.J., Chibbar, R.N. (2000). Isolation of a cDNAencoding a granule-bound 152-kilodalton starch-branching enzyme in wheat. PlantPhysiology 124, 253–263.
Ball, S., Guan, H-P., James, M., Myers, A., Keeling, P., Mouille, G., Bul�eon, A., Colonna,P., Preiss, J. (1996). From glycogen to amylopectin: A model for the biogenesis of theplant starch granule. Cell 86, 349–352.
Benmoussa, M., Suhendra, B., Aboubacar, A., Hamaker, B.R. (2006). Distinctive sorghumstarch granule morphologies appear to improve raw starch digestibility. Starch - St€arke58, 92–99.
Bertoft, E., Boyer, C., Manelius, R., Avall, A-K. (2000). Observations on the alphaamylolysis pattern of some waxy maize starches from inbred line Ia453. CerealChemistry 77, 657–664.
Bird, A.R., Flory, C., Davies, D.A., Usher, S., Topping, D.L. (2004). A novel barley cultivar(Himalaya 292) with a specific gene mutation in starch synthase IIa raises large bowelstarch and short-chain fatty acids in rats. The Journal of Nutrition 134, 831–835.
Bjorck I., Nyman M., Pedersen P., Siljestrom M., Asp N.G., Eggum B.O. (1987).Formation of enzyme resistant starch during autoclaving of wheat starch: Studies invitro and in vivo. Journal of Cereal Science 6, 159–172.
Blazek, J., Copeland, L. (2010). Amylolysis of wheat starches. I. Digestion kinetics ofstarches with varying functional properties. Journal of Cereal Science 51, 265–270.
Brown, I.L. (2004). Applications and uses of resistant starch. Journal of AOAC Interna-tional 87, 727–732.
Bul�eon, A., Colonna, P., Planchot, V., Ball, S. (1998). Starch granules: structure andbiosynthesis. International Journal of Biological Macromolecules 23, 85–112.
Starch Biosynthesis in Relation to Resistant Starch 15

3GCH01 07/19/2013 16:58:9 Page 16
Burton, R.A., Bewley, D., Smith, A.M., Bhattacharyya, M.K., Tatge, H., Ring, S., Bull, V.,Hamilton, W.D.O., Martin, C. (1995). Starch branching enzymes belonging to distinctenzyme families are differentially expressed during pea embryo development. The PlantJournal 7, 3–15.
Chatterjee, M., Berbezy, P., Vyas, D., Coates, S., Barsby, T. (2005). Reduced expression ofa protein homologous to glycogenin leads to reduction of starch content in Arabidopsisleaves. Plant Science 168, 501–509.
Chibbar, R.N., Chakraborty, M. (2005). Waxy Wheat. In: Abdel-Aaal, E., Wood, P. (eds).Specialty grains for food and feed, 143–165. American Association of Cereal Chemists,Inc., St Paul, Minnesota, USA.
Chung, H-J., Lim, H.S., Lim, S-T. (2006). Effect of partial gelatinization and retrogradationon the enzymatic digestion of waxy rice starch. Journal of Cereal Science 43, 353–359.
Colleoni, C., Myers, A.M., James, M.G. (2003). One- and two- dimensional native PAGEactivity gel analyses of maize endosperm proteins reveal functional interactions betweenspecific starch metabolizing enzymes. Journal of Applied Glycoscience 50, 207–212.
Commuri, P.D., Keeling, P.L. (2001). Chain length specificities of maize starch synthaseI enzyme: studies of glucan affinity and catalytic properties. The Plant Journal 25,475–486.
Craig, J., Lloyd, J.R., Tomlinson, K., Barber, L., Edwards, A., Wang, T.L., Martin, C.,Hedley, C.L., Smith, A.M. (1998). Mutations in the gene encoding starch synthase IIprofoundly alter amylopectin structure in pea embryos. The Plant Cell 10, 413–426.
Crowe, T.C., Seligman, S.A., Copeland, L. (2000). Inhibition of enzymic digestion ofamylose by free fatty acids in vitro contributes to resistant starch formation. The Journalof Nutrition 130, 2006–2008.
Dai,Z. (2010).Activities of enzymes involved in starch synthesis inwheat (TriticumaestivumL.) grains differing in starch content. Russian Journal of Plant Physiology 57, 74–78.
Denyer, K., Hylton, C.M., Jenner, C.F., Smith, A.M. (1995). Identification of multipleisoforms of soluble and granule-bound starch synthase in developing wheat endosperm.Planta 196, 256–265.
Denyer, K., Clarke, B., Hylton, C., Tatge, H., Smith, A.M. (1996). The elongation ofamylose and amylopectin chains in isolated starch granules. The Plant Journal 10,1135–1143.
Dinges, J.R., Colleoni, C., James, M.G., Myers, A.M. (2003). Mutational analysis of thepullulanase-type debranching enzyme of maize indicates multiple functions in starchmetabolism. The Plant Cell 15, 666–680.
Edwards, A., Fulton, D.C., Hylton, C.M., Jobling, S.A., Gidley, M., R€ossner, U.,Martin, C.,Smith, A.M. (1999). A combined reduction in activity of starch synthases II and III ofpotato has novel effects on the starch of tubers. The Plant Journal 17, 251–261.
Englyst, H.N., Kingman, S.M., Cummings, J.H. (1992). Classification and measurement ofnutritionally important starch fractions.EuropeanJournal ofClinicalNutrition46, S33–S50.
Escarpa, A., Gonzalez, M.C., Morales, M.D., Saura-Calixto, F. (1997). An approach to theinfluence of nutrients and other food constituents on resistant starch formation. FoodChemistry 60, 527–532.
Fannon, J.E., Hauber, R.J., BeMiller, J.N. (1992). Surface pores of starch granules. CerealChemistry 69, 284–288.
Fettke, J., Hejazi, M., Smirnova, J., H€ochel, E., Stage, M., Steup, M. (2009). Eukaryoticstarch degradation: integration of plastidial and cytosolic pathways. Journal of Exper-imental Botany 60, 2907–2922.
16 Resistant Starch

3GCH01 07/19/2013 16:58:9 Page 17
Fujita, N., Yoshida, M., Asakura, N., Ohdan, T., Miyao, A., Hirochika, H., Nakamura, Y.(2006). Function and characterization of starch synthase I using mutants in rice. PlantPhysiology 140, 1070–1084.
Gallant, D.J., Bouchet, B., Bul�eon, A., Perez, S. (1992). Physical characteristics of starchgranules and susceptibility to enzymatic degradation. European Journal of ClinicalNutrition 46, S3–S16.
Gallant, D., Bouchet, B., Baldwin, P.M. (1997). Microscopy of starch: evidence of a newlevel of granule organization. Carbohydrate Polymers 32, 177–191.
Galliard, T. (1987). Starch: properties and potential. Starch - St€arke 39, 335–374.Ganeshan, S., Drinkwater, J.M., Repellin, A., Chibbar, R.N. (2010). Selected carbohydratemetabolism genes show coincident expression peaks in grains of in vitro- culturedimmature spikes of wheat (Triticum aestivum L.). Journal of Agricultural and FoodChemistry 58, 4193–4201.
Gao, M., Wanat, J., Stinard, P.S., James, M.G., Myers, A.M. (1998). Characterization ofdull1, a maize gene coding for a novel starch synthase. The Plant Cell 10, 399–412.
Grimaud, F., Rogniaux, H., James, M.G., Myers, A.M., Planchot, V. (2008). Proteome andphosphoproteome analysis of starch granule-associated proteins from normal maize andmutants affected in starch biosynthesis. Journal of Experimental Botany 59, 3395–3406.
Guan, H.P., Preiss, J. (1993). Differentiation of the properties of the branching isozymesfrom maize (Zea mays). Plant Physiology 102, 1269–1273.
Hannah, C.L., James, M. (2008). The complexities of starch biosynthesis in cerealendosperms. Current Opinion in Biotechnology 19, 160–165.
Hannah, L.C., Nelson, O.E. (1976). Characterization of ADP-glucose pyrophosphorylasefrom shrunken-2 and brittle-2 mutants of maize. Biochemical Genetics 14, 547–560.
Hendriks, J.H.M., Kolbe, A., Gibon, Y., Stitt, M., Geigenberger, P. (2003). ADP-glucosepyrophosphorylase is activated by posttranslational redox-modification in response tolight and to sugars in leaves of Arabidopsis and other plant species. Plant Physiology133, 838–849.
Hennen-Bierwagen, T.A., Liu, F., Marsh, R.S., Kim, S., Gan, Q., Tetlow, I.J., Emes, M.J.,James, M.G., Myers, A.M. (2008). Starch biosynthetic enzymes from developing maizeendosperm associate in multisubunit complexes. Plant Physiology 146, 1892–1908.
Hirose, T., Terao, T. (2004). A comprehensive expression analysis of the starch synthasegene family in rice (Oryza sativa L.). Planta 220, 9–16.
Hizukuri, S., Kaneko, T., Takeda, Y. (1983). Measurement of the chain length ofamylopectin and its relevance to the origin of crystalline polymorphism of starchgranules. Biochimica et Biophysica Acta 760, 188–191.
Hizukuri, S. (1986). Polymodal distribution of chain lengths of amylopectins and itssignificance. Carbohydrate Research 147, 342–347.
Hoover, R., Zhou, Y. (2003). In vitro and in vivo hydrolysis of legume starches bya-amylase and resistant starch formation in legumes – a review. Carbohydrate Polymers54, 401–417.
Hu, P., Zhao, H., Duan, Z., Linlin, Z., Wu, D. (2004). Starch digestibility and the estimatedglycemic score of different types of rice differing in amylose contents. Journal of CerealScience 40, 231–237.
Hung, P.V., Yamamori, M., Morita, N. (2005). Formation of enzyme resistant starch in breadas affected by high-amylose wheat flour substitutions. Cereal Chemistry 82, 690–694.
Hussain, H., Mant, A., Seale, R., Zeeman, S., Hinchliffe, E., Edwards, A., Hylton, C.,Bornemann, S., Smith, A.M., Martin, C., Bustos, R. (2003). Three isoforms of
Starch Biosynthesis in Relation to Resistant Starch 17

3GCH01 07/19/2013 16:58:9 Page 18
isoamylase contribute different catalytic properties for the debranching of potatoglucans. The Plant Cell 15, 133–149.
Hutchinson, C.B. (1921). Heritable characters of maize. VII Shrunken endosperm. Journalof Heredity 12, 76–83.
Hylton, C., Smith, A.M. (1992). The rb mutation of peas causes structural and regulatorychanges in ADP glucose pyrophosphorylase from developing embryos. Plant Physiol-ogy 99, 1626–1634.
Ito, T., Saito, K., Sugawara, M., Mochida, K., Nakakuki, T. (1999). Effect of raw and heat-moisture-treated high-amylose corn starches on the process of digestion in the ratdigestive tract. Journal of the Science of Food and Agriculture 79, 1203–1207.
James, M.G., Robertson, D.S., Myers, A.M. (1995). Characterization of the maize genesugary-1, a determinant of starch composition in kernels. The Plant Cell 7, 417–429.
James, M.G., Denyer, K., Myers, A.M. (2003). Starch synthesis in the cereal endosperm.Current Opinion in Plant Biology 6, 215–222.
Jane, J-L., Chen, Y.Y., Lee, L.F., McPherson, A.E., Wong, K.S., Radosavljevic, M.,Kasemsuwan, T. (1999). Effects of amylopectin branch chain length and amylosecontent on the gelatinization and pasting properties of starch. Cereal Chemistry 76,629–637.
Jane, J-L., Maningat, C.C., Wongsagonsup, R. (2010). Starch Characterization, Variety andApplication. In: Singh, B.P. (ed). Industrial Crops and Uses, 207–235.
Jiang, H., Campbell, M., Blanco, M., Jane, J-L. (2010). Characterization of maize amylose-extender (ae) mutant starches: Part II. Structures and properties of starch residuesremaining after enzymatic hydrolysis at boiling-water temperature. CarbohydratePolymers 80, 1–12.
Johnson, P.E., Patron, N.J., Bottrill, A.R., Dinges, J.R., Fahy, B.F., Parker, M.L., Waite,D.N., Denyer, K. (2003). A low starch barley mutant, Risø 16, lacking the cytosolicsmall subunit of ADP-glucose pyrophosphorylase, reveals the importance of thecytosolic isoform and the identity of the plastidial small subunit. Plant Physiology131, 684–696.
Kasemsuwan, T., Jane, J., Schnable, P., Stinard, P., Robertson, D. (1995). Characterizationof the dominant mutant amylose-extender (Ae1–5180) maize starch. Cereal Chemistry72, 457–464.
Kubo, A., Rahman, S., Utsumi, Y., Li, Z., Mukai, Y., Yamamoto, M., Ugaki, M., Harada,K., Satoh, H., Konik-Rose, C., Morell, M., Nakamura, Y. (2005). Complementation ofsugary-1 phenotype in rice endospermwith thewheat isoamylase1 gene supports a directrole for isoamylase1 amylopectin biosynthesis. Plant Physiology 137, 43–56.
Lafiandra, D., Sestili, F., D’Ovidio, R., Janni, M., Botticella, E., Ferrazzano, G., Silvestri,M., Ranieri, R., DeAmbrogio, E. (2010). Approaches for modification of starchcomposition in durum wheat. Cereal Chemistry 87, 28–34.
Lan, H., Hoover, R., Jayakody, L., Liu, Q., Donner, E., Baga, M., Asare, E.K., Hucl, P.,Chibbar, R.N. (2008). Impact of annealing on the molecular structure and physico-chemical properties of normal, waxy and high amylose bread wheat starches. FoodChemistry 111, 663–675.
Lehmann, U., Robin, F. (2007). Slowly digestible starch – its structure and healthimplications: a review. Trends in Food Science & Technology 18, 346–355.
Li, J.H., Vasanthan, T., Hoover, R., Rossanagel, B.G. (2004). Starch from hull-less barley:V. In vitro susceptibility of waxy, normal, and high amylose starches towards hydrolysisby alpha-amylases and amyloglucosidase. Food Chemistry 84, 621–632.
18 Resistant Starch

3GCH01 07/19/2013 16:58:9 Page 19
Li, L., Jiang, H., Campbell, M., Blanco, M., Jane, J-L. (2008). Characterization of maizeamylose-extender (ae) mutant starches. Part I: Relationship between resistant starchcontents and molecular structures. Carbohydrate Polymers 74, 396–404.
Li, Z., Chu, X., Mouille, G., Yan, L., Kosar-Hashemi, B., Hey, S., Napier, J., Shewry, P.,Clarke, B., Appels, R., Morell, M.K., Rahman, S. (1999). The localization andexpression of the class II starch synthases of wheat. Plant Physiology 120, 1147–1155.
Morell, M.K., Kosar-Hashemi, B., Cmiel, M., Samuel, M.S., Chandler, P., Rahman, S.,Buleon, A., Batey, I.L., Li, Z. (2003). Barley sex6 mutants lack starch synthase IIaactivity and contain a starch with novel properties. The Plant Journal 34, 173–185.
Morrison, W.R., Karkalas, J. (1990). Starch. In: Dey, P.M. (ed). Methods in PlantBiochemistry, 323–352. Academic Press, London.
Mouille, G., Maddelein, M-L., Libessart, N., Talaga, P., Decq, A., Delrue, B., Ball, S.(1996). Preamylopectin processing: A mandatory step for starch biosynthesis in plants.The Plant Cell 8, 1353–1366.
Mukerjea, R., Robyt, J.F. (2005). Starch biosynthesis: the primer nonreducing-endmechanism versus nonprimer reducing-end two-site insertion mechanism. Carbohy-drate Research 340, 245–255.
Myers, A.M., Morell, M.K., James, M.G., Ball, S.G. (2000). Recent progress towardunderstanding biosynthesis of the amylopectin crystal. Plant Physiology 122, 989–997.
Nakamura, T., Vrinten, P., Hayakawa, K., Ikeda, J. (1998). Characterization of a granule-bound starch synthase isoform found in the pericarp of wheat. Plant Physiology 118,451–459.
Nakamura, Y. (2002). Towards a better understanding of the metabolic system foramylopectin biosynthesis in plants: rice endosperm as a model tissue. Plant CellPhysiology 43, 718–725.
Nichols, B.L., Avery, S., Sen, P., Swallow, D.M., Hanh, D., Sterchi, E. (2003). The maltase-glucoamylase gene: Common ancestry to sucrase-isomaltase with complementary starchdigestion activities. Proceedings of the National Academy of Sciences of the UnitedStates of America 100, 1432–1437.
Nishi, A., Nakamura, Y., Tanaka, N., Satoh, H. (2001). Biochemical and genetic analysisof the effects of amylose-extender mutation in rice endosperm. Plant Physiology 127,459–472.
Noda, T., Kimura, T., Otani, M., Ideta, O., Shimada, T., Saito, A., Suda, I. (2002).Physicochemical properties of amylose-free starch from transgenic sweet potato.Carbohydrate Polymers 49, 253–260.
Park, J.T., Rollings, J.E. (1994). Effects of substrate branching characteristics on kinetics ofenzymatic depolymerization of mixed linear and branched polysaccharides: I. Amylose/amylopectin a- amylolysis. Biotechnology and Bioengineering 44, 792–800.
Patron, N.J., Smith, A.M., Fahy, B.F., Hylton, C.M., Naldrett, M.J., Rossanagel, B.G.,Denyer, K. (2002). The altered pattern of amylose accumulation in the endosperm of low-amylose barley cultivars is attributable to a single mutant allele of granule bound starchsynthase I with a deletion in the 50-non-coding region. Plant Physiology 130, 190–198.
Patron, N.J., Greber, B., Fahy, B.F., Laurie, D.A., Parker, M.L., Denyer, K. (2004). The lys5mutations of barley reveal the nature and importance of plastidial ADP-Glc transportersfor starch synthesis in cereal endosperm. Plant Physiology 135, 2088–2097.
Peng, M., Gao, M., Ba�ga, M., Hucl, P., Chibbar, R.N. (2000). Starch-branching enzymes
preferentially associated with A-type starch granules in wheat endosperm. PlantPhysiology 124, 265–272.
Starch Biosynthesis in Relation to Resistant Starch 19

3GCH01 07/19/2013 16:58:10 Page 20
Poulsen, P., Kreiberg, J.D. (1993). Starch branching enzyme cDNA from Solanumtuberosum. Plant Physiology 102, 1053–1054.
Preiss, J., Danner, S., Summers, P.S., Morell, M., Barton, C.R., Yang, L., Nieder, M. (1990).Molecular characterization of the brittle-2 gene effect on maize endosperm ADPglucosepyrophosphorylase subunits. Plant Physiology 92, 881–885.
Preiss, J., Sivak, M.N. (1996). Starch synthesis in sinks and sources. In: Zamaski, E.,Schaffer, A.A. (eds). Photoassimilate distribution in plants and crops – Source-sinkrelationships, 63–96.
Rahman, S., Regina,A., Li, Z.,Mukai, Y., Yamamoto,M.,Kosar-Hashemi, B., Abrahams, S.,Morell, M. (2001). Comparison of starch branching enzymes genes reveal evolutionaryrelationships among isoforms: characterization of a gene for starch branching enzymes IIafrom the wheat D genome donor Aegilops tauschii. Plant Physiology 125, 1314–1324.
Regina, A., Bird, A., Topping, D., Bowden, S., Freeman, J., Barsby, T., Kosar-Hashemi, B.,Li, Z., Rahman, S., Morell, M. (2006). High-amylose wheat generated by RNAinterference improves indices of large-bowel health in rats. Proceedings of the NationalAcademy of Sciences of the United States of America 103, 3546–3551.
Regina, A., Kosar-Hashemi, B., Ling, S., Li, Z., Rahman, S., Morell, M. (2010). Control ofstarch branching in barley defined through differential RNAi suppression of starchbranching enzyme IIa and IIb. Journal of Experimental Botany 61, 1469–1482.
Repellin, A., Ba�ga, M., Chibbar, R.N. (2008). In vitro pullulanase activity of wheat
(Triticum aestivum L.) limit-dextrinase type starch debranching enzyme is modulated byredox conditions. Journal of Cereal Science 47, 302–309.
Rold�an, I., Wattebled, F., Lucas, M., Delvall�e, D., Planchot, V., Jim�enez, S., P�erez, R., Ball,S., D’Hulst, C., M�erida, �A. (2007). The phenotype of soluble starch synthase IVdefective mutants of Arabidopsis thaliana suggests a novel function of elongationenzymes in the control of starch granule formation. The Plant Journal 49, 492–504.
Rooney, L.W., Pflugfelder, R.L. (1986). Factors affecting starch digestibility with specialemphasis on Sorghum and Corn. Journal of Animal Science 63, 1607–1623.
Ryoo, N., Yu, C., Park, C-S., Baik, M-Y., Park, I.M., Cho, M-H., Bhoo, S.H., An, G., Hahn,T-R., Jeon, J-S. (2007). Knockout of a starch synthase gene OsSSIIIa/Flo5 causes whitecore floury endosperm in rice (Oryza sativa L.). Plant Cell Reports 26, 1083–1095.
Sajilata, M.G., Singhal, R.S., Kulkarni, P.R. (2006). Resistant starch – A review. Compre-hensive Reviews in Food Science and Food Safety 5, 1–17.
Sakulsingharoj, C., Choi, S-B.,Hwang, S-K., Edwards, G.E., Bork, J.,Meyer, C.R., Preiss, J.,Okita, T.W. (2004). Engineering starch biosynthesis for increasing rice seed weight: therole of the cytoplasmic ADP-glucose pyrophosphorylase. Plant Science 167, 1323–1333.
Schwall, G.P., Safford, R., Westcott, R.J., Jeffcoat, R., Tayal, A., Shi, Y-C., Gidley, M.J.,Jobling, S.A. (2000). Production of very-high amylose potato starch by inhibition ofSBE A and B. Nature Biotechnology 18, 551–554.
Sestili, F., Botticella, E., Bedo, Z., Phillips, A., Lafiandra, D. (2010). Production of novelallelic variation for genes involved in starch biosynthesis through mutagenesis.Molecu-lar Breeding 25, 145–154.
Shannon, J.C., Pien, F-M., Cao, H., Liu, K-C. (1998). Brittle-1, an adenylate translocator,facilitates transfer of extraplastidial synthesized ADP-glucose into amyloplasts of maizeendosperms. Plant Physiology 117, 1235–1252.
Shu, X., Jia, L., Gao, J., Song, Y., Zhao, H., Nakamura, Y.,Wu, D. (2007). The influences ofchain length of amylopectin on resistant starch in rice (Oryza sativa L.). Starch - St€arke59, 504–509.
20 Resistant Starch

3GCH01 07/19/2013 16:58:10 Page 21
Smidansky, E.D., Clancy, M., Meyer, F.D., Lanning, S.P., Blake, N.K., Talbert, L.E.,Giroux, M.J. (2002). Enhanced ADP-glucose pyrophosphorylase activity in wheatendosperm increases seed yield. Proceedings of the National Academy of Sciencesof the United States of America 99, 1724–1729.
Smith, A.M., Denyer, K., Martin, C. (1997). The synthesis of the starch granule. AnnualReview of Plant Physiology and Plant Molecular Biology 48, 67–87.
Smith, A.M. (2001). The biosynthesis of starch granules. Biomacromolecules 2, 335–341.Stinard, P.S., Robertson, D.S., Schnable, P.S. (1993). Genetic isolation, cloning, andanalysis of a mutator-induced, dominant antimorph of the maize amylose extender1locus. The Plant Cell 5, 1555–1566.
Szydlowski, N., Ragel, P., Raynaud, S., Lucas, M.M., Rold�an, I., Montero, M., Munoz, F.J.,Ovecka, M., Bahaji, A., Planchot, V., Pozueta-Romero, J., D’Hulst, C., Merida, �A.(2009). Starch granule initiation in Arabidopsis requires the presence of either class IVor class III starch synthases. The Plant Cell 21, 2443–2457.
Tetlow, I.J., Wait, R., Zhenxiao, L., Akkasaeng, R., Bowsher, C.G., Esposito, S., Kosar-Hashemi, B., Morell, M.K., Emes, M.J. (2004). Protein phosphorylation in amyloplastsregulates starch branching enzyme activity and protein-protein interactions. The PlantCell 16, 694–708.
Thompson, D.B. (2000). On the non-random nature of amylopectin branching. Carbohy-drate Polymers 43, 223–239.
Tjaden, J., M€ohlmann, T., Kampfenkel, K., Henrichs, G., Neuhaus, H.E. (1998). Alteredplastidic ATP/ADP-transporter activity influences potato (Solanum tuberosum L.) tubermorphology, yield and composition of tuber starch. The Plant Journal 16, 531–540.
Topping, D.L., Clifton, P.M. (2001). Short chain fatty acids and human colonic function:roles of resistant starch and non-starch polysaccharides. Physiological Reviews 81,1031–1064.
Umemoto, T., Aoki, N. (2005). Single-nucleotide polymorphisms in rice starch synthaseIIa that alter starch gelatinisation and starch association of the enzyme. Functional PlantBiology 32, 763–768.
Umemoto, T., Terashima, K., Nakamura, Y., Satoh, H. (1999). Differences in amylopectinstructure between two rice varieties in relation to the effects of temperature during grainfilling. Starch - St€arke 51, 58–62.
Umemoto, T., Yano, M., Satoh, H., Shomura, A., Nakamura, Y. (2002). Mapping of a generesponsible for the difference in amylopectin structure between japonica-type andindica-type rice varieties. Theoretical and Applied Genetics 104, 1–8.
Vrinten, P.L., Nakamura, T. (2000). Wheat granule-bound starch synthase I and II areencoded by separate genes that are expressed in different tissues. Plant Physiology 122,255–263.
Wang, Y.J., White, P., Pollak, L., Jane J. (1993). Characterization of starch structures of 17maize endosperm mutant genotypes with Oh43 inbred line background. Cereal Chem-istry 70, 171–179.
Wang, Z., Chen, X., Wang, J., Liu, T., Liu, Y., Zhao, L., Wang, G. (2007). Increasing maizeseed weight by enhancing the cytoplasmic ADP-glucose pyrophosphorylase activity intransgenic maize plants. Plant Cell, Tissue and Organ Culture 88, 83–92.
Wei, C., Qin, F., Zhu, L., Zhou, W., Chen, Y., Wang, Y., Gu, M., Liu, Q. (2010).Microstructure and ultrastructure of high-amylose rice resistant starch granules modifiedby antisense RNA inhibition of starch branching enzyme. Journal of Agricultural andFood Chemistry 58, 1224–1232.
Starch Biosynthesis in Relation to Resistant Starch 21

3GCH01 07/19/2013 16:58:10 Page 22
Weurding, R.E., Veldman, A., Veen, W.A.G., Aar, P.J., Verstegen, M.W.A. (2001). In vitrostarch digestion correlates well with rate and extent of starch digestion in broilerchicken. The Journal of Nutrition 131, 2336–2342.
Yamamori, M., Fujita, S., Hayakawa, K., Matsuki, J., Yasui, T. (2000). Genetic eliminationof starch granule protein, SGP-1, of wheat generates an altered starch with apparent highamylose. Theoretical and Applied Genetics 101, 21–29.
Yamamori, M., Kato, M., Yui, M., Kawasaki, M. (2006). Resistant starch and starch pastingproperties of a starch synthase IIa-deficient wheat with apparent high amylose.Australian Journal of Agricultural Research 57, 531–535.
Yao, N., Paez, A.V., White, P.J. (2009). Structure and function of starch and resistant starchfrom corn with different doses of mutant amylose-extender and floury-1 alleles. Journalof Agricultural and Food Chemistry 57, 2040–2048.
Yoo, S-H., Jane, J-L. (2002). Structural and physical characteristics of waxy and otherwheat starches. Carbohydrate Polymers 49, 297–305.
Zeeman, S.C., Umemoto, T., Lue, W-L., Au-Yeung, P., Martin, C., Smith, A.M., Chen, J.(1998). A mutant of Arabidopsis lacking a chloroplastic isoamylase accumulates bothstarch and phytoglycogen. The Plant Cell 10, 1699–1711.
Zeeman, S.C., Kossmann, J., Smith, A.M. (2010). Starch: Its metabolism, evolution andbiotechnological modifications in plants. Annual Review of Plant Biology 61, 209–234.
Zhang, G., Ao, Z., Hamaker, B.H. (2006). Slow digestion property of native cereal starches.Biomacromolecules 7, 3252–3258.
Zhang, X., Colleoni, C., Ratushna, V., Sirghle-Colleoni, M., James, M.G., Myers, A.M.(2004). Molecular characterization demonstrates that the Zea mays gene sugary2 codesfor the starch synthase isoform SSIIa. Plant Molecular Biology 54, 865–869.
Zhang, X., Myers, A.M., James, M.G. (2005). Mutations affecting starch synthase III inArabidopsis alter leaf starch structure and increase in the rate of starch synthesis. PlantPhysiology 138, 663–674.
22 Resistant Starch

CH02GR 07/30/2013 17:4:49 Page 23
2 Type 2 Resistant Starch inHigh-Amylose Maize Starchand its Development
Hongxin Jiang and Jay-lin JaneDepartment of Food Science and Human Nutrition, Iowa State University, USA
2.1 INTRODUCTION
Starch is a reserve carbohydrate widely present in seeds, roots, tubers, leaves,
stems and fruits of plants. Most starch consists of two polymers – amylopectin
and amylose – which are organized in a semi-crystalline granular structure
(Jane, 2004). Amylopectin has highly branched structures, consisting of linear
chains of a(1! 4)-linked a-D-glucopyranose units, which are connected by
approximately 5% a(1! 6) glycosidic branch linkages (French, 1984).
Amylose is an essentially linear polymer of a(1! 4)-linked a-D-glucopyr-anose units (Takeda et al., 1986, 1989, 1993).
A third component, known as intermediate component (IC), is present in
maize mutants such as high-amylose and sugary-1 maize starches. IC has
branched structures with molecular weights smaller than amylopectin but
similar to amylose (Baba & Arai, 1984; Klucinec & Thompson, 1998; Li
et al., 2008; Wang et al., 1993). The IC molecules also possess iodine-binding
capacities and b-amylolysis limits between amylopectin and amylose mol-
ecules (Kasemsuwan et al., 1995). Normal maize starch contains about 30%
amylose (Hasjim et al., 2009). High-amylose maize starches with amylose
contents of 50%, 70% and 80% have been commercialized. High-amylose
maize starch with 90% amylose has also been reported (Shi et al., 1998).
Starch is present invarieties of foods and is amajor energy source for humans.
After ingesting, starch is partially hydrolyzed by human salivary a-amylase and
is predominantly digested in the small intestine by pancreatic a-amylase and
mucosal enzymes (Sang & Seib, 2006). Starch from different botanical sources
has different digestive rate and total digestibility, which results in different
physiological effects on human health. Resistant starch (RS) is a portion of starch
23
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

CH02GR 07/30/2013 17:4:49 Page 24
in foodswhichcannot behydrolyzedbyenzymes in the small intestine andpasses
on to the large intestine as a prebiotic (Englyst & Cummings, 1985; Englyst &
Macfarlane, 1986). RS provides many health benefits to humans, as previously
reported (Behall & Howe, 1996; Behall et al., 2006a, 2006b; Dronamraju et al.,
2009;Higgins et al., 2004; Pawlak et al., 2004; Robertson et al., 2003, 2005;Van
Munster et al., 1994; Zhang et al., 2007).
Enzymatic hydrolysis of raw starch granules is a complex process. Many
characteristics of the starch determine the rate and the extent of enzymatic
hydrolysis of the granules, such as granular size, structure of the amylopectin,
polymorph, amylose content and lipid content. These characteristics are species
dependent andare also affectedbygrowth conditions andpost-harvest treatment
(Jane, 2006;Lehmann&Robin, 2007; Setiawan et al., 2010;Tester et al., 2006).
It is well known that starch granules with larger granular sizes are digested at a
slower rate than those with smaller granular sizes, because larger starch granules
have relatively smaller surface area for enzymes to attack (Tester et al., 2006).The
A-type polymorphic starch granules, such as normal andwaxymaize, rice,wheat,
and taro starch, are known to have ‘weak points’ and are more easily hydrolyzed
by enzymes than are the B- and C-type polymorphic starch granules such as
potato, high-amylosemaize, green banana and pea starch (Jane et al., 2003; Jane,
2007). This difference is attributed to the branch chain-length of amylopectin and
the packing of double helices in the starch granule (Figure 2.1).
B type A type
2.1 nm
6.9 nm
1.6 nm
7.4 nm 9.0 nm
C
A
C
A
C
A
Figure 2.1 Structure models of amylopectins for A type and B type starches.A: amorphous; C: crystalline. Reproduced from Carbohydrate Polymers, 49(3), YooS.-H. and Jane J.-l. Molecular weights and gyration radii of amylopectins determinedby high-performance size-exclusion chromatography equippedwithmulti-angle laser-light scattering and refractive index detectors, 307–314. Copyright 2002, withpermission from Elsevier.
24 Resistant Starch

CH02GR 07/30/2013 17:4:49 Page 25
The amylopectin of the A-type polymorphic starch has a larger
proportion of short A and B1 chains but smaller numbers of long B2,
B3, and B4 chains than that of the B-type polymorphic starch (Jane et al.,
1999). B2, B3, and B4 chains extend through multiple clusters of
amylopectin and their movements are constrained by the consecutive
crystalline regions in the starch granule. The short A and B1 chains,
however, extend through one cluster of amylopectin and have free ends to
move around within the cluster and to form closely packed monoclinic
crystallites. Consequently, the A-type polymorphic starch granules consist
of voids (weak points) (Figure 2.2a,c; Jane, 2006; Jane, 2007; Jane et al.,
2004; Pan & Jane, 2000), which are easily penetrated and hydrolyzed by
Figure 2.2 Internal structures of starch granules. a (maize) and b (potato): scanningelectron micrographs of inner part of granules remaining after removal of thechemical gelatinized granule surface. Reproduced from Jane, 2006; 2007. c and d:confocal laser-scanning micrographs of normal maize and potato starch granules,respectively; starch granules were stained using Rhodamine B dye. Reproduced fromJane, 2006.
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 25

CH02GR 07/30/2013 17:4:52 Page 26
enzymes to generate pin holes and channels (Fannon et al., 1992, 1993;
Jane, 2006).
The B-type polymorphic starch granules, consisting of rigidly organized
amylopectin branch chains (Figure 2.1) and without weak points or voids
(Figure 2.2b,d) (Jane, 2006; Jane, 2007; Jane et al., 2004; Jane & Shen, 1993),
are normally digested slowly by erosion of starch granules from the surface
(Jane, 2006; Robyt, 1998). After cooking, potato starch, however, is highly
digestible. The increase in amylose content in normal starch granules (Jane
et al., 2003) and in high-amylose starch granules (Evans & Thompson, 2004;
Jiang et al., 2010b; Li et al., 2008; Shi & Jeffcoat, 2001) reduce the rate of
enzymatic hydrolysis of the raw starch granules. These features can be
attributed to the fact that, in the normal starch granule, the amylose is more
concentrated at the periphery of starch granules and interacts with amylopectin
to form a hard shell (Debet & Gidley, 2007; Jane & Shen, 1993; Pan & Jane,
2000); this makes the starch granule more resistant to enzymatic hydrolysis
(Tester et al., 2006). In high-amylose maize starch, amylose forms crystalline
structure, which is highly resistant to enzymatic hydrolysis (Evans & Thomp-
son, 2004; Jiang et al., 2010b; Li et al., 2008; Shi & Jeffcoat, 2001).
In general, starch is consumed by humans after cooking. Starch gelatini-
zation during cooking is an irreversible reaction which is associated with the
dissociation of double helices and loss of birefringence viewed under a
polarized light (Jane, 2004). Gelatinization properties of starch isolated
from different botanical origins differ, and this relates to starch molecular
structures and minor components present in the starch granule (Jane, 2004;
Yoo et al., 2009). The enzymatic hydrolysis of gelatinized starch (amorphous)
is much faster than that of raw starch granules (semi-crystalline) (Lehmann &
Robin, 2007; Tester et al., 2006).
High-amylose maize (amylose-extender, aemutant) starches with amylose
contents ranging from 50–80% have been commercially available and are
widely used in food and non-food applications (Fergason et al., 1994;
Richardson et al., 2000). Maize ae single-mutant starch contains about
65% apparent amylose and about 15% RS (Li et al., 2008) – substantially
higher than normal (non-mutant) maize starch (�30% and �1.5%, respec-
tively) (Hasjim & Jane, 2009; Hasjim et al., 2009). After cross-breeding the
maize ae-mutant with a maize line carrying high-amylose modifier (HAM)
gene, the apparent amylose content of the starch (GEMS-0067) elevates to
�85% and the RS content of the starch increases to �43% (Li et al., 2008).
Maize ae-mutant starch consists of spherical and elongated starch
granules (Figure 2.3a,b), which differ from the normal maize starch in
consisting of spherical and angular granules (Figure 2.3e; Hasjim et al.,
2009; Jiang et al., 2010b; Li et al., 2007; Mercier et al., 1970; Perera et al.,
26 Resistant Starch

CH02GR 07/30/2013 17:4:52 Page 27
2001; Wolf et al., 1964). The elongated starch granule content of a double
mutant of ae and HAM genes, GEMS-0067 line, increases up to 32%
(Figure 2.3a; Jiang et al., 2010b). These elongated starch granules retain
their granular shapes after cooking at 95–100 �C with excess water and are
highly resistant to enzymatic hydrolysis (Figure 2.3c). In this chapter,
formations of RS and elongated starch granules in the maize ae-mutant
starch and how the effect of HAM gene on the RS content of the maize
ae-mutant starch will be discussed.
Figure 2.3 Scanning electron micrographs of maize starches and resistant-starch(RS) residues (Jiang et al., 2010b). (a) GEMS-0067 starch (�85% apparent amylose)showing spherical (s) and elongated (e) granules. (b) H99ae starch (�65% apparentamylose). (c) GEMS-0067 RS residue remaining after enzymatic hydrolysis at 95–100 �C (AOAC Method 991.43). (d) H99ae RS residue in a gel-like form. Arrowsindicate fragmented, hollowed and half-shell-like granules. (e) Normal maize starch.Reprinted from Jiang et al. (2010b), Copyright 2010, with permission from Elsevier.
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 27

CH02GR 07/30/2013 17:4:52 Page 28
2.2 RS FORMATION IN HIGH-AMYLOSE MAIZESTARCH
Although high-amylose maize starch has been developed for over 60 years
(BeMiller, 2009; Kramer et al., 1956; Vineyard & Bear, 1952; Vineyard et al.,
1958), interests in the RS of the high-amylose maize starch has just emerged in
the past two decades, resulting from consumer awareness of health food
benefits (Sajilata et al., 2006). The structures and properties of the high-
amylose maize starch and its RS residue have been intensively studied in order
to understand the RS formation in the starch (Evans & Thompson, 2004; Jiang
& Liu, 2002; Jiang et al., 2010b; Li et al., 2008; Shi et al., 1998; Shi &
Jeffcoat, 2001). It has been reported that molecular weight and branch chain
length distributions of RS residues remaining after pancreatic a-amylase
hydrolysis are similar to that of native starch counterparts (Evans &
Thompson, 2004; Shi & Jeffcoat, 2001).
After enzymatic hydrolysis of high-amylose maize starches at 95–100 �C(AOAC Method 991.43), the RS residues display semi-crystalline structures
with the B-type polymorph (Jiang et al., 2010b). The onset gelatinization
temperatures of the RS residues are above 100 �C, indicating the long-chain
double-helical crystallites are present in the RS residues. Gel-permeation
chromatograms show that the RS residues consist of large molecules with
degrees of polymerization (DP) 840–951 and small molecules with DP 59–
74 (Jiang et al., 2010b). This differs from molecular weight distributions of
RS residues remaining after pancreatic a-amylase hydrolysis of the native
high-amylose maize starches (Evans & Thompson, 2004; Shi & Jeffcoat,
2001).
The molecular weight distributions of the debranched RS residues remain-
ing after enzymatic hydrolysis of the high-amylose maize starches at
95–100 �C show that the RS residues consist mostly of linear molecules
and are mainly derived from amylose/IC molecules (Jiang et al., 2010b).
Thus, it is concluded that the RS residues of high-amylose maize starches
consist of mainly semi-crystalline structure of long-chain double-helices of
amylose/IC (Jiang et al., 2010b). It is likely that the amylose/IC crystallites in
native high-amylose maize starch are present in blocks, which prevent the
starch granule from swelling at 95–100 �C, maintain the semi-crystalline
structure and protect the bulk of the amylose and IC and a small portion of
amylopectin molecules from enzymatic hydrolysis (Jiang et al., 2010b).
The presence of amylose double-helical crystallites in high-amylose maize
starch is also supported by the structure and properties of the Naegeli dextrin,
which is produced by hydrolyzing the starch at 38 �C using 15.3% sulphuric
28 Resistant Starch

CH02GR 07/30/2013 17:4:52 Page 29
acid (Jiang et al., 2010e). The Naegeli dextrins of high-amylose maize
starches consist of double helices with chain lengths of DP� 12
(8.6–11.2%), DP 13–24 (38.7–54.1%), DP 25–36 (23.6–30.6%) and DP� 37
(13.1–22.1%). Because the longest detectable chains of the Naegeli dextrin
of ae waxy maize starch are DP 25 (Jane et al., 1997), the long-chain double
helices of DP� 25 present in the Naegeli dextrin are attributed to crystalline
fragments of amylose double helices (Jane & Robyt, 1984; Jiang et al.,
2010b, 2010e). The thermal properties of the Naegeli dextrins of the high-
amylose maize starches display very high peak (113.9–122.2 �C) and
conclusion gelatinization-temperatures (148.0–160.0 �C), which also sup-
port the presence of amylose double helices (Jiang et al., 2010e; Sievert &
Pomeranz, 1989, 1990).
Although high-amylose maize starches contain approximately 0.2–0.7%
lipids (Jiang et al., 2010b; Morrison, 1992, 1993, 1995; Morrison et al.,
1993), the lipids present in the starch granules also reduce the enzyme
digestibility of the starch at 95–100 �C (Jiang et al., 2010b). After the lipids
are removed from the starch granules, the RS contents of the high-amylose
maize starches reduce from 10.6–43.4% to 9.0–28.9%. The effects of lipids
on the RS content of the high-amylose maize starch are discussed in other
chapters of this book.
2.3 RS FORMATION DURING KERNEL DEVELOPMENT
Although RS and amylose/IC double-helical crystalline structures are found
in mature high-amylose maize starch granules after harvesting and drying
(Jiang et al., 2010b), it is not known whether the RS and the amylose/IC
double-helical crystalline structure are formed during kernel development
or are induced during post-harvest processing and boiling/a-amylase
hydrolysis.
Jiang et al. (2010a) studied RS formation during kernel development of
GEMS-0067 line, a maize double-mutant of ae and HAM genes. Gelatiniza-
tion thermograms of the starches harvested at 15, 20, 30, 40, 54 (mature) days
after pollination (DAP) display a major thermal transition (first peak) with the
peak temperature between 76.6–81.0 �C and an additional thermal transition
(second peak) with the peak temperature �97.1 �C (Figure 2.4). The second
peak first appears as a shoulder on 20 DAP and gradually increases into a
significant peak on 30, 40, and 54 DAP (Jiang et al., 2010a). The relative
intensity of the first peak (76.6–81.0 �C) decreases with kernel maturation and
a decrease in the amylopectin content of the starch, indicating that this peak
corresponds to the dissociation of amylopectin crystallites (Jiang et al., 2010a,
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 29

CH02GR 07/30/2013 17:4:53 Page 30
2010b; Kasemsuwan et al., 1995; Li et al., 2008). The increase in the size of
the second peak correlates with the lipid content of the starch, and this peak
disappears after removal of lipids from the starch, indicating that this
additional peak is the melting of amylose-lipid complex (Jiang et al.,
2010a; Tufvesson et al., 2003a, 2003b).
After removal of lipids, the conclusion gelatinization-temperature of the
starch does not change. The conclusion gelatinization temperature, however,
increases with kernel maturation, from 105.0 �C on 15 DAP to 117.8–122.2 �Con later dates (Jiang et al., 2010a). The percentage of the enthalpy change of
the thermal transition at temperature range above 95 �C, which corresponds tothe melting of long-chain double-helical crystallites of amylose/IC, increases
with the kernel maturation and significantly correlates with the RS content of
the starch (Jiang et al., 2010a). The RS content of the high-amylose maize
starch increases with kernel maturation and directly correlates with the
amylose/IC content of the starch (r¼ 0.99, p< 0.001) (see Table 2.1).
Thus, the authors conclude that the long-chain double-helical crystallites
of amylose/IC in the high-amylose maize starch develop with the kernel
maturation, which results in an increase in the RS content of the starch (Jiang
et al., 2010a).
140130120110100908070605040
Hea
t Flo
w E
ndo
Up
Temperature (°C)
15 DAP
20 DAP
30 DAP
40 DAP
54 DAP
Figure 2.4 Differential scanning calorimetry (DSC) thermograms of native GEMS-0067 starches harvested at different developmental stages (Jiang et al., 2010a).DAP: days after pollination. The peak area above the dashed line indicates meltingof amylose-lipid complex. Reprinted with permission from Jiang et al. (2010a).Copyright 2010 American Chemical Society.
30 Resistant Starch

CH02GR 07/30/2013 17:4:53 Page 31
2.4 ELONGATED STARCH GRANULES OFHIGH-AMYLOSE MAIZE STARCH
2.4.1 Structures of elongated starch granules
Different from normal maize starch, which contains starch granules with
spherical and angular shapes (Figure 2.3e; Hasjim et al., 2009; Perera et al.,
2001; Wongsagonsup et al., 2008), high-amylose maize starch contains
elongated granules in addition to spherical granules (Figure 2.3a,b; Boyer
et al., 1976; Jiang et al., 2010b; Mercier et al., 1970; Shi & Jeffcoat, 2001;
Sidebottom et al., 1998; Wolf et al., 1964). Starch of maize double mutant
GEMS-0067 consists of up to 32% elongated granules, which is substantially
greater than that of the maize ae single-mutant starch (�7%) and normal
maize starch (0%) (Campbell et al., 2007; Hasjim et al., 2009; Jiang et al.,
2010b; Li et al., 2007; Perera et al., 2001; Wongsagonsup et al., 2008). The
percentage of elongated starch granules increases with kernel maturation and
the increase in amylose/IC content of the starch (Jiang et al., 2010a, 2010b,
2010d; Mercier et al., 1970). Elongated starch granules are mostly found in
mature endosperm cells located in the central part of the crown region of the
high-amylose maize kernel (Boyer et al., 1976).
Starch molecules in the granule are present in semi-crystalline structures of
double helices. The birefringence pattern of a starch granule observed under a
polarized light microscope reflects the arrangement of starch molecules in the
granule (French, 1984). Normal maize starch granules display a typical
Table 2.1 Resistant starch (RS) and amylose/intermediate component (IC) contents ofGEMS-0067 starches harvested at different kernel-developmental stages (Jiang et al.,2010a). Reprinted with permission from Jiang et al. (2010a). Copyright 2010 AmericanChemical Society.
Sample RSa (%) Amylose/ICb (%)
15 DAPc 9.0�1.2 55.2�0.520 DAP 26.4�0.1 78.4�0.230 DAP 29.6�0.8 81.9�0.340 DAP 32.0�0.1 88.6�1.354 DAP 32.1�0.3 87.6�0.4Coefficient between RS and amylose/IC content 0.99d
aRS content of starch was determined using AOAC method 991.43 for total dietary fibre.bAmylose/IC content of starch was determined using Sepharose CL-2B gel-permeation chromatog-
raphy followed by the total carbohydrate (phenol-sulphuric acid) determination.cDAP: days after pollination.dp<0.001.
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 31

CH02GR 07/30/2013 17:4:53 Page 32
Maltese cross birefringence pattern (Wongsagonsup et al., 2008), reflecting
that starch molecules in the granule are aligned radially from the hilum, which
are oriented perpendicular to the granule surface.
Most spherical granules of high-amylose maize starch exhibit a normal
Maltese cross birefringence (Figure 2.5, granule a), similar to the granules of
normal maize starch. This birefringence pattern is consistent with the confocal
laser-scanning micrograph (CLSM) of the 8-aminopyrene-1,3,6-trisulfonic
acid (APTS)-derivatized spherical granule, which shows a hilum with a bright
colour at the centre of the granule (Figure 2.6a; Glaring et al., 2006; Jiang
et al., 2010c). These findings indicate that the spherical granule is developed
from one single granule nucleus.Elongated starch granules, however, display three different types of
birefringence patterns (Figure 2.5). The type 1 birefringence pattern displays
several Maltese crosses overlapping in one granule (Figure 2.5, granule b), while
the type 2 birefringence pattern shows one or moreMaltese crosses in one part of
the granule and weak or no birefringence on the rest of the granule (Figure 2.5,
granule c). The type 3 birefringence pattern exhibits a granule displayingweak or
no birefringence (Figure 2.5, granule d; Jiang et al., 2010c; Wolf et al., 1964).
The differences in birefringence patterns of elongated starch granules
indicate that arrangements of starch molecules in the granule differ between
elongated granules, and even between different parts of one elongated granule.
This feature is consistent with the CLSM images of APTS-stained elongated
granules, which showmultiple regions of different fluorescence intensity in one
elongated granule (Figure 2.6b–g) (Glaring et al., 2006; Jiang et al., 2010c).
Figure 2.5 Polarized (a) and phase contrast (b) light micrographs of high-amylosemaize (GEMS-0067) starch. Arrows indicate birefringence patterns of: (a) onegranuledisplaying one Maltese cross; (b) Maltese crosses overlapping in one granule; (c) asinglegranuleconsistingofoneormoreMaltesecrossesandweak/nobirefringenceonthe rest part of the granule; (d) granule showingweak/no birefringence. Bar¼20mm.
32 Resistant Starch

CH02GR 07/30/2013 17:4:54 Page 33
2.4.2 Formation of elongated starch granules
Starch granule formation in the amyloplast of high-amylose maize endosperm
at an early stage of kernel development (20 DAP) has been studied using
transmission electron microscopy (TEM) (Jiang et al., 2010c). Some amy-
loplasts at the subaleurone layer of high-amylose maize endosperm contain
only one spherical granule showing normal growth rings and a hilum
(Figure 2.7a), which are similar to amyloplasts in normal maize endosperm.
The majority of amyloplasts, however, contain two or more small starch
granules (Figure 2.7b).
These small granules in the amyloplast, which develop at the early stage,
fuse and grow into an elongated starch granule (Figures 2.7c–g; Jiang et al.,
2010c). This fusion is likely the result of interactions between the amylose of
adjacent granules, because high-amylose maize starch contains a large
concentration of amylose. The amylose molecules of one granule can contact
that of an adjacent granule and form anti-parallel double helices between the
Figure 2.6 Confocal laser-scanning microscope images of high-amylose maize(GEMS-0067) starch granules (Jiang et al., 2010c). Starch granules were labelled using8-aminopyrene-1,3,6-trisulfonic acid APTS). (a) Spherical granule showing brightcolour around the hilum; (b), (c), (d), (e), (f), and (g) show elongated starchgranules displaying multiple regions with intense fluorescence. Bar¼10mm. Reprintedfrom Jiang et al. (2010c), Copyright 2010, with permission from Elsevier.
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 33

CH02GR 07/30/2013 17:4:56 Page 34
two granules (Figure 2.7d; Jiang et al., 2010a, 2010b; Li et al., 2008), which
bind the two adjacent granules and prevent the amyloplast from dividing.
Eventually, the two granules grow into one elongated granule (Figure 2.7e).
A proposed mechanism of the elongated starch granule formation is shown
in Figure 2.8. The mechanism is consistent with the fact that the number of
elongated starch granules increases with the increase in the amylose content of
the starch (Jiang et al., 2010a, 2010b). In the normal maize starch, amylose
content is low (�30%) and branch chains of amylopectin are short comparing
with amylose. The short branch chains of amylopectin cannot form stable anti-
parallel double helices with branch chains of an adjacent granule in a normal
maize amyloplast during granule development. Thus, the small starch gran-
ules in the normal maize amyloplast are separated with the division of
Figure 2.7 Transmission electron micrographs of high-amylose maize (GEMS-0067) endosperm tissue harvested on 20 days after pollination (Jiang et al.,2010c).(a) Spherical starch granule with a hilumat centre of granule and growth rings.(b)Overviewofendosperm tissue. (c) Twostarchgranules initiated inoneamyloplastatearly stage of starch granule development. (d) Initial fusion of starch granules. (e) Twofused starch granules forming an elongated starch granule. (f) Two connected starchgranules and a third starch granule in the amyloplast protrusion. (g) Three smallgranules fused into one granule, with one small granule at head showing hilum andgrowth rings and the other two granules displaying no hilum or growth rings.m:membrane boundary of amyloplast. Black arrow indicates hilum. Bars¼1mm onA, C, D, F and 2mm on B, E, G. Reprinted from Jiang et al. (2010c), Copyright2010, with permission from Elsevier.
34 Resistant Starch

CH02GR 07/30/2013 17:5:0 Page 35
amyloplast (Jiang et al., 2010c). Most amyloplasts in normal maize endo-
sperm consist of a single starch granule.
2.4.3 Location of RS in the starch granule
The RS residues of GEMS-0067, consisting of about 85% amylose, are found
mostly in elongated granular structures (Figure 2.3c), whereas that of the ae
single-mutant, consisting of approximately 65% amylose, are present in a gel
form (Figure 2.3d; Jiang et al., 2010b). This is attributed to a greater content of
amylose/IC double-helical crystallites present in the granules of GEMS-0067,
particularly elongated granules (Jiang et al., 2010b).
The hollowed and half-shell-shaped granules and the gel-like starch
(Figure 2.3c,d) observed in the RS residues are the remnants of the outer
layer of spherical granules (Jiang et al., 2010b). These structures can be
attributed to the fact that the amylopectin molecules around the hilum are
loosely packed and can be promptly gelatinized and hydrolyzed by thermally
stable a-amylase at 95–100 �C. Amylose molecules, however, are more
concentrated at the periphery of the spherical granules (Jane & Shen,
1993; Jiang et al., 2010a; Jiang et al., 2010b; Li et al., 2007; Pan & Jane,
2000). The amylose molecules at the periphery of the starch granules interact
between themselves and with amylopectin molecules to form a hard shell
Figure 2.8 Proposed formation mechanism of elongated starch granule in theamyloplast (Jiang et al., 2010c). (a) Two starchgranules, eachwith hilumandgrowthrings, are initiated in amyloplast at an early stage of granule development. (b) Twogranules begin fusion through amylose interaction by forming anti-parallel doublehelices, which prevent amyloplast division. (c) Fused granules have integrated outerlayer growth rings.m:membrane boundary of amyloplast. Reprinted from Jiang et al.(2010c), Copyright 2010, with permission from Elsevier.
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 35

CH02GR 07/30/2013 17:5:1 Page 36
which is difficult to disperse and, thus, is resistant to enzymatic hydrolysis
(Debet & Gidley, 2007; Gray & BeMiller, 2004; Huber & BeMiller, 2001;
Jane & Shen, 1993; Jiang et al., 2010a, 2010b; Li et al., 2007; Pan & Jane,
2000).
2.5 ROLES OF HIGH-AMYLOSE MODIFIER (HAM)GENE INMAIZE ae-MUTANT
The role of HAM gene and its dosage effects on the physicochemical
properties of maize ae-mutant starch have been studied and reported (Jiang
et al., 2010d). Maize endosperm is a triploid tissue, resulting from the
fertilization of two maternal nuclei and one paternal nucleus (Birchler,
1993). Thus, endosperms with 0, 1, 2, and 3 doses of HAM gene are produced
by self- and inter-pollination of an ae single-mutant line and the ae and HAM
double-mutant line, GEMS-0067. The increase in HAM gene dosage in the
maize ae mutant increases the apparent amylose content of the starch (see
Table 2.2; Jiang et al., 2010d). After including three doses of HAM gene to the
maize ae-mutant, the starch has longer branch chain-lengths of IC molecules
but slightly shorter branch chain-lengths of amylopectin molecules than
maize ae single-mutant starch (Li et al., 2008). Two or three doses of
HAM gene in maize ae-mutant substantially increase the RS content of
the maize ae-mutant starch. One dose of HAM gene, however, has little effect
on the RS content.
Morphological properties of starch granules are also influenced by the
dosage of HAM gene (Jiang et al., 2010d). The greater the HAM gene dosage
in the starch, the greater the number of elongated granules is found in
maize ae-mutant starch. With the presence of three doses of HAM gene,
many maize ae-mutant starch granules display weak or no birefringence,
Table 2.2 Amylose and resistant starch (RS) contents of native maize starches isolatedfrom kernels of self- and intercrossed lines GEMS-0067 (G) and H99ae (H) (Jiang et al.,2010d). Reprinted with permission from Jiang et al. (2010a). Copyright 2010 AmericanChemical Society.
Sample HAM gene doses Amylosea (%) RSb (%)
G/G 3 69.9�0.0 35.0�0.5G/H 2 64.9�0.4 28.1�0.8H/G 1 61.1�0.1 12.9�0.2H/H 0 56.3�0.4 15.7�0.2
aAmylose content of starch was determined using iodine-colorimetric method.bRS content of starch was determined using AOAC method 991.43 for total dietary fibre.
36 Resistant Starch

CH02GR 07/30/2013 17:5:1 Page 37
indicating changes in molecular organization of the granules. The increase in
HAM gene dosage decreases the percentage crystallinity of starch, which is
consistent with the decrease in amylopectin content of the starch (Jiang et al.,
2010d). This is attributed to amylopectin having short branch chains which are
free to move and interact with one another and, thus, facilitate the formation of
double-helical crystallites.
Maize ae-mutant starches display multiple thermal-transitions during
gelatinization (Jiang et al., 2010a, 2010b, 2010d). The temperature of the
thermal transition range increases with the increase in the HAM gene dosage.
The first thermal transition peak, with the peak temperature at �82.9 �C,corresponds to the melting of amylopectin crystallites (Jiang et al., 2008;
Kasemsuwan et al., 1995; Li et al., 2008), which decreases with an increase in
HAM gene dosage (Jiang et al., 2010d). The second thermal transition peak,
with the peak temperature at �99.1 �C, corresponds to the dissociation of
amylose-lipid complexes (Jiang et al., 2010a; Li et al., 2008; Tufvesson et al.,
2003a, 2003b), which increases with an increase in HAM gene dosage (Jiang
et al., 2010d). The conclusion gelatinization-temperature of the starch is
above 100 �C and increases with the increase in HAM gene-dosage (Jiang
et al., 2010d), indicating the formation of amylose/IC crystallites (Jiang et al.,
2010a).
These findings indicate that the formation of amylose/IC crystallites and
amylose-lipid complex increasewith the increase in HAMgene dosage, which
is consistent with the increase in the RS content of the starch (Jiang et al.,
2010d).
2.6 CONCLUSIONS
Long-chain double-helical crystallites of amylose/IC in high-amylose maize
starch are developed during kernel development and are concentrated in the
elongated granules and at the outer layer of the spherical granules. The
amylose/IC crystallites, having onset gelatinization temperature above
100 �C, restrict starch granules from swelling and dispersion at 95–
100 �C and protect the starch molecules from enzymatic hydrolysis at
95–100 �C. Lipids present in the granule also protect starch granules
from enzymatic hydrolysis at 95–100 �C. The formation of elongated starch
granules in high-amylose maize starch results from amylose interactions
between adjacent small granules in the amyloplast during granule develop-
ment. The inclusion of HAM gene(s) to the maize ae-mutant increases the
crystallites of amylose/IC in the starch, which results in the increase in RS
content of the starch.
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 37

CH02GR 07/30/2013 17:5:1 Page 38
REFERENCES
Baba, T., Arai, Y. (1984). Structural features of amylomaize starch. 3. Structural charac-terization of amylopectin and intermediate material in amylomaize starch granules.Agricultural and Biological Chemistry 48(7), 1763–1775.
Behall, K.M., Howe, J.C. (1996). Resistant starch as energy. Journal of the AmericanCollege of Nutrition 15(3), 248–254.
Behall, K.M., Scholfield, D.J., Hallfrisch, J.G. (2006a). Barley [beta]-glucan reducesplasma glucose and insulin responses compared with resistant starch in men. NutritionResearch 26(12), 644–650.
Behall, K.M., Scholfield, D.J., Hallfrisch, J.G., Liljeberg-Elmstaahl, H.G.M. (2006b).Consumption of both resistant starch and beta-glucan improves postprandial plasmaglucose and insulin in women. Diabetes Care 29(5), 976–981.
BeMiller, J.N. (2009). One hundred years of commercial food carbohydrates in the UnitedStates. Journal of Agricultural and Food Chemistry 57(18), 8125–8129.
Birchler, J.A. (1993). Dosage analysis of maize endosperm development. Annual Review ofGenetics 27, 181–204.
Boyer, C.D., Daniels, R.R., Shannon, J.C. (1976). Abnormal starch granule formation inZea-Mays-L endosperms possessing amylose-extender mutant. Crop Science 16(2),298–301.
Campbell, M.R., Jane, J., Pollak, L., Blanco, M., O’Brien, A. (2007). Registration of maizegermplasm line GEMS-0067. Journal of Plant Registrations 1, 60–61.
Debet, M.R., Gidley, M.J. (2007). Why do gelatinized starch granules not dissolvecompletely? roles for amylose, protein, and lipid in granule ‘ghost’ integrity. Journalof Agricultural and Food Chemistry 55(12), 4752–4760.
Dronamraju, S.S., Coxhead, J.M., Kelly, S.B., Burn, J., Mathers, J.C. (2009). Cell kineticsand gene expression changes in colorectal cancer patients given resistant starch: arandomised controlled trial. Gut 58(3), 413–420.
Englyst, H.N., Cummings, J.H. (1985). Digestion of the polysaccharides of some cerealfood in the human small intestine. American Journal of Clinical Nutrition 42(5), 778–787.
Englyst, H.N., Macfarlane, G.T. (1986). Breakdown of resistant and readily digestiblestarch by human gut bacteria. Journal of the Science of Food and Agriculture 37(7),699–706.
Evans, A., Thompson, D.B. (2004). Resistance to a-amylase digestion in four native high-amylose maize starches. Cereal Chemistry 81(1), 31–37.
Fannon, J.E., Hauber, R.J., BeMiller, J.N. (1992). Surface pores of starch granules. CerealChemistry 69(3), 284–288.
Fannon, J.E., Shull, J.M., BeMiller, J.N. (1993). Interior channels of starch granules.Cereal Chemistry 70(5), 611–613.
Fergason, V., Jeffcoat, R., Fannon, J.E., Capitani, T.A. (1994). Low amylopectin starch. USpatent 5300145.
French, D. (1984). Organization of starch granules. In: Whistler, R.L., BeMiller, J.N.,Paschall, E.F. (eds). Starch: Chemistry and Technology, 183–247. New York: AcademicPress.
Glaring, M.A., Koch, C.B., Blennow, A. (2006). Genotype- specific spatial distribution ofstarch molecules in the starch granule: A combined CLSM and SEM approach.Biomacromolecules 7(8), 2310–2320.
38 Resistant Starch

CH02GR 07/30/2013 17:5:1 Page 39
Gray, J.A., BeMiller, J.N. (2004). Development and utilization of reflectance confocal laserscanning microscopy to locate reaction sites in modified starch granules. CerealChemistry 81(2), 278–286.
Hasjim, J., Jane, J. (2009). Production of resistant starch by extrusion cooking of acid-modified normal-maize starch. Journal of Food Science 74(7), C556–C562.
Hasjim, J., Srichuwong, S., Scott, M.P., Jane, J. (2009). Kernel composition, starchstructure, and enzyme digestibility of opaque-2 maize and quality protein maize.Journal of Agricultural and Food Chemistry 57(5), 2049–2055.
Higgins, J.A., Higbee, D.R., Donahoo, W.T., Brown, I.L., Bell, M.L., Bessesen, D.H.(2004). Resistant starch consumption promotes lipid oxidation. Nutrition Metabolism1, 8.
Huber, K.C., BeMiller, J.N. (2001). Location of sites of reaction within starch granules.Cereal Chemistry 78(2), 173–180.
Jane, J. (2004). Starch: structure and properties. In: Tomasik, P. (ed). Chemical andFunctional Properties of Food Saccharides, 81–101. New York: CRC Press.
Jane, J. (2006). Current understanding on starch granule structures. Journal of AppliedGlycoscience 53(3), 205–213.
Jane, J. (2007). Structure of starch granules. The Japanese Society of Applied Glycoscience54, 31–36.
Jane, J., Robyt, J.F. (1984). Structure studies of amylose-V complexes and retrogradedamylose by action of alpha amylases, and a new method for preparing amylodextrins.Carbohydrate Research 132(1), 105–118.
Jane, J., Shen, J.J. (1993). Internal structure of the potato starch granule revealed bychemical gelatinization. Carbohydrate Research 247, 279–290.
Jane, J., Wong, K.-S., McPherson, A.E. (1997). Branch-structure difference in starches ofA- and B-type x-ray patterns revealed by their Naegeli dextrins. Carbohydrate Research300(3), 219–227.
Jane, J., Chen, Y.Y., Lee, L.F., McPherson, A.E., Wong, K.S., Radosavljevic, M.,Kasemsuwan, T. (1999). Effects of amylopectin branch chain length and amylosecontent on the gelatinization and pasting properties of starch. Cereal Chemistry76(5), 629–637.
Jane, J., Ao, Z., Duvick, S.A., Wiklund, M., Yoo, S.-H., Wong, K.-S., Gardner, C. (2003).Structures of amylopectin and starch granules: How are they synthesized? Journal ofApplied Glycoscience 50(2), 167–172.
Jane, J., Atichokudomchai, N., Suh, D. (2004). Internal structures of starch granulesrevealed by confocal laser-light scanning microscopy. In: Tomasik, P., Yuryev, V.P.,Bertoft, E. (eds). Starch: Progress in Structural Studies, Modifications and Applica-tions, 147–156. Cracow: Polish Society of Food Technologists.
Jiang, G., Liu, Q. (2002). Characterization of residues from partially hydrolyzed potato andhigh amylose corn starches by pancreatic a-amylase. Starch - Staerke 54(11), 527–533.
Jiang, H., Campbell, M., Jane, J. (2008). Characterization of enzyme-resistant maize ae-mutant starches during kernel development. Annual Meeting of American Association ofCereal Chemists, Honolulu, Hawaii, USA.
Jiang, H., Blanco, M., Campbell, M., Jane, J. (2010a). Resistant-starch formation in high-amylose maize during kernel development. Journal of Agricultural and Food Chemistry58(13), 8043–8047.
Jiang, H., Campbell, M., Blanco, M., Jane, J. (2010b). Characterization of maize amylose-extender (ae) mutant starches. Part II: Structures and properties of starch residues
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 39

CH02GR 07/30/2013 17:5:1 Page 40
remaining after enzymatic hydrolysis at boiling-water temperature. CarbohydratePolymers 80(1), 1–12.
Jiang, H., Horner, H.T., Pepper, T.M., Blanco, M., Campbell, M., Jane, J. (2010c).Formation of elongated starch granules in high-amylose maize. Carbohydrate Polymers80(2), 534–539.
Jiang, H., Jane, J., Acevedo, D., Green, A., Shinn, G., Schrenker, D., Srichuwong, S.,Campbell, M., Wu, Y. (2010d). Variations in starch physicochemical properties from ageneration-means analysis study using amylomaize V and VII parents. Journal ofAgricultural and Food Chemistry 58, 5633–5639.
Jiang, H., Srichuwong, S., Campbell, M., Jane, J. (2010e). Characterization of maizeamylose-extender (ae) mutant starches. Part III: Structures and properties of the Naegelidextrins. Carbohydrate Polymers 81, 885–891.
Kasemsuwan, T., Jane, J., Schnable, P., Stinard, P., Robertson, D. (1995). Characterizationof the dominant mutant amylose-extender (Ae1–5180) maize starch. Cereal Chemistry72(5), 457–464.
Klucinec, J.D., Thompson, D.B. (1998). Fractionation of high-amylose maize starches bydifferential alcohol precipitation and chromatography of the fractions.Cereal Chemistry75(6), 887–896.
Kramer, H.H., Whistler, R.L., Anderson, E.G. (1956). A new gene interaction in theendosperm of maize. Agronomy Journal 48, 170–172.
Lehmann, U., Robin, F. (2007). Slowly digestible starch – its structure and healthimplications: a review. Trends in Food Science Technology 18(7), 346–355.
Li, L., Blanco, M., Jane, J.-l. (2007). Physicochemical properties of endosperm andpericarp starches during maize development. Carbohydrate Polymers 67(4), 630–639.
Li, L., Jiang, H., Campbell, M., Blanco, M., Jane, J. (2008). Characterization of maizeamylose-extender (ae) mutant starches: Part I. Relationship between resistant starchcontents and molecular structures. Carbohydrate Polymers 74(3), 396–404.
Mercier, C., Charbonniere, R., Gallant, D., Guilbot, A. (1970). Development of somecharacteristics of starches extracted from normal corn and amylomaize grains duringtheir formation. Staerke 22(1), 9–16.
Morrison, W.R. (1992). Analysis of cereal starches.Modern Methods of Plant Analysis 14(Seed Anal.), 199–215.
Morrison, W.R. (1993). Barley lipids. Barley 199–246.Morrison, W.R. (1995). Starch lipids and how they relate to starch granule structure and
functionality. Cereal Foods World 40(6), 437–446.Morrison, W.R., Law, R.V., Snape, C.E. (1993). Evidence for inclusion complexes of lipids
with V-amylose in maize, rice and oat starches. Journal of Cereal Science 18(2), 107–109.
Pan, D.D., Jane, J. (2000). Internal structure of normal maize starch granules revealed bychemical surface gelatinization. Biomacromolecules 1(1), 126–132.
Pawlak, D.B., Kushner, J.A., Ludwig, D.S. (2004). Effects of dietary glycaemic index onadiposity, glucose homoeostasis, and plasma lipids in animals. Lancet 364(9436), 778–785.
Perera, C., Lu, Z., Sell, J., Jane, J. (2001). Comparison of physicochemical properties andstructures of sugary-2 cornstarch with normal and waxy cultivars. Cereal Chemistry 78(3), 249–256.
Richardson, P.H., Jeffcoat, R., Shi, Y.-C. (2000). High-amylose starches: From bio-synthesis to their use as food ingredients. MRS Bulletin 25(12), 20–24.
40 Resistant Starch

CH02GR 07/30/2013 17:5:1 Page 41
Robertson, M.D., Currie, J.M., Morgan, L.M., Jewell, D.P., Frayn, K.N. (2003). Prior short-term consumption of resistant starch enhances postprandial insulin sensitivity in healthysubjects. Diabetologia 46(5), 659–665.
Robertson, M.D., Bickerton, A.S., Dennis, A.L., Vidal, H., Frayn, K.N. (2005). Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adiposetissue metabolism. American Journal of Clinical Nutrition 82(3), 559–567.
Robyt, J.F. (1998). Essentials of Carbohydrate Chemistry. New York: Springer-Verlag.Sajilata, M.G., Singhal, R.S., Kulkarni, P.R. (2006). Resistant starch – A review. Compre-hensive Reviews in Food Science and Food Safety 5(1), 1–17.
Sang, Y., Seib, P.A. (2006). Resistant starches from amylose mutants of corn by simulta-neous heat-moisture treatment and phosphorylation. Carbohydrate Polymers 63(2),167–175.
Setiawan, S., Widjaya, H., Rhakpongpairoj, V., Jane, J. (2010). Effects of drying conditionsof corn kernels and storage at an elevated humidity on starch structures and properties.Journal of Agricultural and Food Chemistry 58(23), 12260–12267.
Shi, Y.-C., Jeffcoat, R. (2001). Structural features of resistant starch. In:McCleary, B, Prosky,L. (eds). Advanced Dietary Fibre Technology, 430–439. Oxford, UK: Wiley-Blackwell.
Shi, Y.-C., Capitani, T., Trzasko, P., Jeffcoat, R. (1998). Molecular structure of a low-amylopectin starch and other high-amylose maize starches. Journal of Cereal Science 27(3), 289–299.
Sidebottom, C., Kirkland, M., Strongitharm, B., Jeffcoat, R. (1998). Characterization of thedifference of starch branching enzyme activities in normal and low-amylopectin maizeduring kernel development. Journal of Cereal Science 27(3), 279–287.
Sievert, D., Pomeranz, Y. (1989). Enzyme-resistant starch. I. Characterization and evalua-tion by enzymic, thermoanalytical, and microscopic methods. Cereal Chemistry 66(4),342–347.
Sievert, D., Pomeranz, Y. (1990). Enzyme-resistant starch. II. Differential scanningcalorimetry studies on heat-treated starches and enzyme-resistant starch residues.CerealChemistry 67(3), 217–221.
Takeda, Y., Hizukuri, S., Juliano, B.O. (1986). Purification and structure of amylose fromrice starch. Carbohydrate Research 148(2), 299–308.
Takeda, C., Takeda, Y., Hizukuri, S. (1989). Structure of Amylomaize Amylose. CerealChemistry 66(1), 22–25.
Takeda, Y., Tomooka, S., Hizukuri, S. (1993). Structures of Branched and Linear-Molecules of Rice Amylose. Carbohydrate Research 246, 267–272.
Tester, R.F., Qi, X., Karkalas, J. (2006). Hydrolysis of native starches with amylases.Animal Feed Science and Technology 130(1–2), 39–54.
Tufvesson, F., Wahlgren, M., Eliasson, A.-C. (2003a). Formation of amylose-lipid com-plexes and effects of temperature treatment. Part 1. Monoglycerides. Starch - Staerke 55(2), 61–71.
Tufvesson, F., Wahlgren, M., Eliasson, A.-C. (2003b). Formation of amylose-lipidcomplexes and effects of temperature treatment. Part 2. Fatty acids. Starch - Staerke55(3–4), 138–149.
Van Munster, I.P., Tangerman, A., Nagengast, F.M. (1994). Effect of resistant starch oncolonic fermentation, bile acid metabolism, and mucosal proliferation. DigestiveDiseases and Sciences 39(4), 834–842.
Vineyard, M.L., Bear, R.P. (1952). Amylose content. Maize Genetics Cooperation News-letter 26, 5.
Type 2 Resistant Starch in High-Amylose Maize Starch and its Development 41

CH02GR 07/30/2013 17:5:1 Page 42
Vineyard, M.L., Bear, R.P., MacMasters, M.M., Deatherage, W.L. (1958). Development of‘amylomaize’ – corn hybrids with high amylose starch. II. Results of breeding efforts.Agronomy Journal 58, 598–602.
Wang, Y.J., White, P., Pollak, L., Jane, J. (1993). Amylopectin and intermediate materialsin starches from mutant genotypes of the Oh43 inbred line. Cereal Chemistry 70(5),521–525.
Wolf, M.J., Seckinger, H.L., Dimler, R.J. (1964). Microscopic characteristics of high-amylose corn starches. Staerke 16(12), 377–382.
Wongsagonsup, R., Varavinit, S., BeMiller, J.N. (2008). Increasing slowly digestible starchcontent of normal and waxy maize starches and properties of starch products. CerealChemistry 85(6), 738–745.
Yoo, S.-H., Perera, C., Shen, J., Ye, L., Suh, D.-S., Jane, J.-L. (2009). Molecular Structureof Selected Tuber and Root Starches and Effect of Amylopectin Structure on TheirPhysical Properties. Journal of Agricultural and Food Chemistry 57(4), 1556–1564.
Zhang, W., Wang, H., Zhang, Y., Yang, Y. (2007). Effects of resistant starch on insulinresistance of type 2 mellitus patients. Chinese Journal of Preventive Medicine 41(2),101–104.
42 Resistant Starch

3GCH03 07/19/2013 17:21:12 Page 43
3 RS4-Type Resistant Starch: Chemistry,Functionality and Health Benefits
Clodualdo C. Maningat1 and Paul A. Seib2
1MGP Ingredients, Inc., USA; Department of Grain Science and Industry,Kansas State University, USA
2Department of Grain Science and Industry, Kansas State University, USA
3.1 INTRODUCTION
The statistics on obesity are alarming; more than one-third of adult Americans
and almost 17% of youth were classed as obese in 2009–2010 (Ogden et al.,
2012). One television news report disturbingly labelled overweight youth as
‘coronary time bombs’, as they are likely to develop heart disease when they
grow into adulthood. As a potential consequence, billions of dollars will be
spent in weight-related bills. The high price tag is simply because obesity is
not a singular medical condition. Being overweight or obese is also associated
with Type 2 diabetes, cardiovascular disease, high cholesterol, hyperlipidae-
mia, hypertension, stroke, cancer and other diseases. About one-third of
Americans have hypertension – a major risk factor for heart disease and stroke
if left untreated.
Resistant starch (RS), as a component of dietary fibre, is not only beneficial
but can be considered essential to the general health and well-being of
consumers worldwide. It demonstrates numerous positive physiological
effects and, therefore, plays a critical role as a food ingredient in addressing
the widespread metabolic syndrome that afflicts the general population. This
chapter specifically highlights the properties of RS belonging to the RS4
classification and underscores its effectiveness in alleviating diet and lifestyle-
related diseases.
43
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH03 07/19/2013 17:21:12 Page 44
3.2 HISTORICAL ACCOUNT OF STARCHINDIGESTIBILITY
From the early 1900s into the 1930s, scientists explored the in vitro and in vivo
amylase digestibility of raw and gelatinized starches from different botanical
sources (Fofanow, 1911; Thorpe, 1913; Daniels & Strickler, 1917; Langwor-
thy & Deuel, 1920, 1922; Katz, 1934) (Table 3.1). Those researchers
succeeded in developing two of the concepts that we know today about
RS: that several raw starches, especially raw potato starch, are resistant to
digestion; and that retrograded starch, which was then termed as
‘amylocoagulose’ (Katz, 1934), resisted digestion.
The linkage between dietary fibre and starch as a dietary source of fibrewas
established in 1982, when British researchers discovered the presence of
starch ‘resistant’ to digestion in the residue of cooked and cooled potatoes
recovered after amylase hydrolysis during the determination of non-starch
polysaccharides (Englyst et al., 1982). A decade after Englyst’s work, a
formal definition of resistant starch as ‘the sum of starch and products of
starch degradation not absorbed in the small intestine of healthy individuals’
was established (Asp, 1992). Then, in 2001, a revised definition of dietary
fibre, which included RS under analogous carbohydrates, was introduced by
the American Association of Cereal Chemists (2001).
Subsequently, other organizations and countries followed suit (Institute of
Medicine, 2001; European Food Safety Authority, 2007) and, finally in 2009,
the Codex Alimentarius Commission adopted a new dietary fibre definition
that included RS under carbohydrate polymers obtained from food raw
material by physical, enzymatic or chemical means (Codex Alimentarius
Commission, 2009). In 2012, Health Canada, after several years of delibera-
tion, issued a new dietary fibre definition that is consistent with the definition
adopted by Codex Alimentarius Commission, and includes RS under novel
fibres (Health Canada, 2012).
RS4-type resistant starch belongs to one of the five types or classes of RS.
Englyst & Macfarlane (1986), citing the work of Englyst (1985), reported on
the classification of starch for nutritional purposes into readily digestible
starch and resistant starch, which was further classified into RS1, RS2, RS3a,
and RS3b. The preceding two types of RS3 were differentiated by their
composition as being mainly staled amylopectin and mainly retrograded
amylose, respectively.
Englyst & Cummings (1987) proposed a classification of starch based on
its digestibility that can distinguish readily digestible starch in freshly cooked
foods, partially resistant starch in raw potato and banana starch, and resistant
44 Resistant Starch

3GCH03 07/19/2013 17:21:13 Page 45
Table 3.1 Chronology of events leading to resistant starch, its classification to RS1-RS5and its inclusion in the definition of dietary fibre.
Year Events References
1911 Raw potato starch was less digested inhuman subjects, but raw wheat, oat andrice starches were practically assimilated.
Fofanow, 1911
1917 Taka-diastase indigestible starch was foundin bakery products.
Daniels and Strickler,1917
1920–1922 Raw corn, wheat, cassava, rice and taro rootstarches were completely digested in vivo,but raw potato, arrowroot and cannastarches were less digestible.
Langworthy and Deuel,1920, 1922
1934 ‘Amylocoagulose’, a retrograded material inannealed starch gels, was not digested bymalt extract.
Katz, 1934
1953 The word ‘dietary fibre’ was first used todescribe plant cell walls in the diet.
Hipsley, 1953
1961–1962 Raw high-amylose corn starch was resistantto in vitro and in vivo digestion.
Ackerson, 1961; Leach& Schoch, 1961;Borchers, 1962;Sandstedt et al., 1962
1969–1971 Chemically modified starch was resistant todigestion by pancreatin.
Janzen, 1969; Leegwater& Luten, 1971
1971–1973 Prevalence of ‘Western’ diseases indeveloped societies, but not in Africa, wasattributed to inadequate dietary fibre(non-starch polysaccharides) intake.
Burkitt, 1971, 1973a,b;Trowell, 1972
1977 Raw banana starch resisted in vitro digestionby a-amylase.
Fuwa, 1977
1981 Protocol for measuring glycemic index wasestablished.
Jenkins et al., 1981
1982 The term ‘resistant starch’ was coined forretrograded starch found in cooked andcooled potatoes.
Englyst et al., 1982
1984 Retrograded amylose was resistant todigestion by a-amylases and yieldedresistant fragments with degree ofpolymerization of 43–50.
Jane and Robyt, 1984
1986 Starch was classified for nutritional purposesinto readily digestible starch and resistantstarch, which was further classified intoRS1, RS2, RS3a and RS3b.
Englyst & Macfarlane,1986
1992 Official definition for ‘resistant starch’ wasestablished as ‘the sum of starch andproducts of starch degradation that arenot absorbed in the small intestines ofhealthy individuals’.
Asp, 1992
(continued)
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 45

3GCH03 07/19/2013 17:21:13 Page 46
Table 3.1 (Continued)
Year Events References
1992 The in vitro Englyst method to measurerapidly digestible starch, slowly digestiblestarch and resistant starch (by difference)was established.
Englyst et al., 1992
1992 RS1, RS2 and RS3 classification of resistantstarch was introduced.
Englyst et al., 1992
1993 Hi-MaizeTM, the first commercially availableresistant starch ingredient from high-amylose maize starch, was released.
Brown et al., 1995
1993 Chemically modified starch was alluded toas another form of resistant starch.
Eerlingen et al., 1993
1995 Chemically modified starch wasindependently reported by twolaboratories to represent RS4classification.
Eerlingen and Delcour,1995; Brownet al., 1995
1995–1996 The Englyst method for in vitro assay ofresistant starch was validated in vivo.
Silvester et al., 1995;Englyst et al., 1996
2000–2001 Low prevalence of ‘Western’ diseases amongAfricans was associated with adequateconsumption of resistant starch, ratherthan the non-starch polysaccharidecomponents of dietary fibre.
Ahmed et al., 2000;Topping & Clifton,2001
2001 American Association of Cereal Chemistsissued a revised definition of dietary fibreto include resistant starch underanalogous carbohydrates; Institute ofMedicine of the National Academy ofSciences issued a new definition for dietaryfibre, functional fibre and total fibre, withresistant starch falling under theclassification of functional fibre.
American Associationof Cereal Chemists,2001; Institute ofMedicine, 2001
2002 Direct method for quantifying resistant starchwas developed for RS1, RS2 and RS3.
McCleary et al., 2002
2006 RS5 classification was proposed to representamylose-lipid complexes organized in theV-type polymorph.
Brown et al., 2006
2009 Codex Alimentarius Commission issued anew definition for dietary fibre to includeresistant starch under carbohydratepolymers obtained from food raw materialby physical, enzymatic or chemical means.
Codex AlimentariusCommission, 2009
2010–2011 Methods to determine soluble, insoluble, andtotal dietary fibre (Codex definition) weredeveloped.
McCleary et al.,2010, 2011
2012 Health Canada issued a new definition fordietary fibre to include resistant starchunder novel fibres.
Health Canada, 2012
46 Resistant Starch

3GCH03 07/19/2013 17:21:13 Page 47
starch formed as a result of food processing. RS was classified into three types
(RS1, RS2 and RS3) as discussed by Englyst et al. (1992):
� RS1 pertains to physically-entrapped starch present in whole or coarsely-
ground grains and pulses where the starch granules are encapsulated within
a cell wall so that amylase enzymes are prevented or delayed from having
access to the glycosidic bonds.� RS2 is raw starch with inherent enzyme resistance by virtue of its crystal-
line structure.� Recrystallized or retrograded amylose represents the RS3 type.
Eerlingen et al. (1993) alluded to the presence of another type of RS in
food, which they described as chemically modified starch to represent starch
that is rendered indigestible to amylase enzymes by virtue of their chemical
modification. Chemically modified starches can inhibit or reduce starch
digestibility by their functional groups blocking access of amylase enzymes,
or by the presence of atypical linkages such as (1! 2)- and (1! 3)-glycosidic
bonds that are not recognized by amylase enzymes.
A fourth type of RS (i.e. RS4) was introduced independently by Eerlingen
& Delcour (1995) and Brown et al. (1995) into the existing three types or
classes of RS. Brown et al. (2006) then added a fifth type of RS (i.e. RS5) to
represent amylose-inclusion complexes with lipids that exist as the V-type
crystals. It is worth noting that guest molecules such as emulsifiers and
surfactants can also form helical complexes with amylose and, to a lesser
extent, amylopectin (Krog, 1971; Evans, 1986; Eliasson, 1998; Faergemand&
Krog, 2006).
3.3 STARCHMODIFICATION YIELDING INCREASEDRESISTANCE TO ENZYME DIGESTIBILITY
Chemically modified food starches are traditionally used as ingredients in the
food industry to enhance the processing performance, physical attributes,
sensory properties and storage stability of consumer packaged food products.
Production of modified starches for food use includes acid treatment, oxida-
tion, esterification and etherification, or combinations of modification (Huber
& BeMiller, 2010; Mason, 2009).
Starch granules are penetrable by low molecular weight, water-soluble
solutes (or chemical reactants), but molecules larger than 1000 Daltons are
effectively excluded (Brown & French, 1977). The pathway of entry is
probably through the pores and channels located on the granule surface
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 47

3GCH03 07/19/2013 17:21:13 Page 48
(Huber & BeMiller, 2000; Kim & Huber, 2008; Han et al., 2006). These
granular features and other factors influence the chemical reactions occurring
in the granule.
Huber &BeMiller (2001, 2010) summarized the intrinsic factors (botanical
source, granule morphology, molecular structure, composition, starch prop-
erties) and extrinsic factors (reaction medium, reaction temperature, amount
of chemicals, type of chemicals, addition of salts, presence of catalysts) that
influence the chemical modification of starch. Biliaderis (1982) stated that
factors influencing the rate and selectivity of starch modification reactions at
the macromolecular level consisted of the reactivity of the starch hydroxyl
groups, specific area and organization of the starch granule, diffusion rate and
reactivity of the co-reactant, and steric factors due to bulkiness of the
substituent groups.
Other researchers discovered that cereal and legume starches are less
susceptible to structural modification than root and tuber starches (Singh
et al., 1993). Corn and amaranth starches displayed differences in optimum
reaction conditions during the preparation of sodium carboxymethyl deriv-
atives (Bhattacharya et al., 1995), while maize starches with a differing
amylose/amylopectin ratio exhibited variations in the distribution of hydrox-
ypropyl groups on the glucan chains (Azemi & Wooton, 1995).
Even though a combination of factors appears to dictate the rate and
selectivity of the modification reaction, Biliaderis (1982) and Hood&Mercier
(1978) suggested that the nature of a modifying agent is more important in
determining the substituent distribution in starch chains than the physico-
chemical characteristics of the granule. When the degree of substitution (DS)
increased to a relatively high value (e.g. DS¼ 0.85–2.89), a disorganization of
the granule crystal structure occurred (Chi et al., 2008). Small-angle X-ray
scattering measurements of acylated starches revealed that a longer substitu-
ent chain like butyrate is accommodated easily within the crystalline lamellar
structure, compared to acetate and propionate chains, leaving the nano-
structure practically unchanged (Lopez-Rubio et al., 2009).
At the molecular level, substituent groups are distributed unevenly (both
between starch molecules and over the length of molecules), indicating that
starch granules react heterogeneously (Huber & BeMiller, 2001, 2010). The
DS of the amylose fraction was much higher than that of the amylopectin
fraction, suggesting a difference in reactivity of amorphous and crystalline
regions (Huang et al., 2007).
Hoover & Sosulski (1986) hypothesized that derivatization sites for
phosphorus oxychloride (POCl3) occurred in the amorphous regions of the
granule, because no difference was observed between the x-ray diffractions of
the cross-linked and unmodified starches. Because POCl3 (or its first
48 Resistant Starch

3GCH03 07/19/2013 17:21:13 Page 49
hydrolysis product, HOPOCl2) is highly reactive, it reacts with the first starch
molecules it encounters near the surface of the granules.
The location of modifying groups was proposed earlier by Biliaderis
(1982) when he introduced two possible structures for chemically modified
amylopectin. Fast-reacting reagents such as acetic anhydride or POCl3 tended
to react at or near the surface of a granule, where the tips (or non-reducing
ends) of the amylopectin cluster chains are located. Slow-reacting reagents
like propylene oxide tended to penetrate deeper to react not only near the tips
of amylopectin cluster chains, but also in the amylopectin region, where there
is dense branching. The location of phosphate cross-links may be assumed to
be uniformly distributed near the non-reducing ends and the branch points in
amylopectin, as noted by Manelius et al. (2000) for the reaction of a cationic
epoxide with potato starch. Both locations comprise the amorphous region of
the starch granule.
According to Huber & BeMiller (2000, 2001), POCl3 reaction with potato
and sorghum starches occurred at the granule surfaces, channels and cavities,
and likely within the granule matrix, as revealed by scanning electron
microscopy with compositional backscattered electron imaging. However,
POCl3 reacted with potato starch largely on the surface of the granule, because
potato starch granules lack pores and channels. It was observed that the
penetration of POCl3 through the perimeter surface of the granules inward was
somewhat impeded by a highly associated ‘outer shell’.
A number of researchers employing in vitro and in vivo techniques reported
that chemical modification of starches restricted to various degrees the
hydrolytic action of amylase enzymes. For example, modified raw or gelati-
nized starch with varying levels of cross-linking and/or substitution had
differential reduction of susceptibility to amylase action (Bjorck et al.,
1989; Hoover & Sosulski, 1986; Wolf et al., 1999; Wooton & Chaudhry,
1979; Conway & Hood, 1976; Janzen, 1969; Ackar et al., 2010; Klaushofer
et al., 1978; Xie & Liu, 2004; Xie et al., 2006; Wepner et al., 1999; Kim &
Huber, 2008; Leegwater & Luten, 1971; Azemi & Wooton, 1995; Hood &
Arneson, 1976; Hahn & Hood, 1980; Annison et al., 1995, 2003; Bajka et al.,
2007; Wolf et al., 2001; Wang et al., 2001; Woo & Seib, 2002).
On the basis of the above-mentioned studies, the mechanism of amylase
resistance of RS4 resistant starch can be summed up as follows. The
substituent groups along the a-1,4 D-glucan chains hinder enzymatic attack
and also make neighbouring glycosidic bonds resistant to degradation. The
presence of cross-linked polymer chains inhibits granular swelling and also
provides steric hindrance to the approach of the active site of amylase
enzymes. Moreover, cross-linking of starch may restrict the movement of
a-amylase through the granule surface pores and channels. Unusual
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 49

3GCH03 07/19/2013 17:21:13 Page 50
glycosidic bonds, such as a-1,2, a-1,3 or b-1,6 bonds, which form during
pyrolysis reaction on starch, are not substrates for amylases.
3.3.1 Cross-linked RS4 starches
Depending on the level of modifying chemicals, cross-linking of starch
granules from different botanical sources had the general effect of maintaining
granular integrity, restricting swelling, resisting mechanical shear and reduc-
ing paste viscosity (Huber & BeMiller, 2010; Mason, 2009). The degree of
cross-linking of starches used as thickeners in food products is too low to elicit
significant resistance to a-amylase hydrolysis (Wurzburg & Vogel, 1984;
Ostergard et al., 1988; Woo & Seib, 2002). Numerous cross-linking agents
have been reacted with starch, such as POCl3, sodium trimetaphosphate
(STMP), epichlorohydrin, adipic anhydride, citric acid and glutaric acid.
These inorganic and organic reagents are said to have bi- or poly-functional
reactive groups. That is, they react with two or more hydroxyl groups on the
starch molecular chains.
Early descriptions in the patent literature of cross-linking starch were those
of Felton & Schopmeyer (1943), utilizing 0.005–0.25% POCl3 at pH 8–12, and
Van Patten & Powell (1969), who improved the thickening ability of starch by
adding trisodium phosphate prior to the cross-linking reaction with POCl3.
POCl3-treated starch has a monoester : diester ratio of 1 : 3, based on the
thallium : phosphorus ratio assayed by inductively coupled plasma-atomic
emission spectroscopy (Koch et al., 1982). Chemical modifications such as
phosphate cross-linking inhibit alpha-amylase hydrolysis, as reported by
different researchers (Bjorck et al., 1989; Hoover & Sosulski, 1986; Wolf
et al., 1999;Wooton&Chaudhry, 1979; Conway&Hood, 1976; Janzen, 1969).
The presence of cross-linked residues across the glucan chains provides
steric hindrance to the action of amylase enzymes. Analysis of cross-linked
granular corn starch by a combination of gel permeation chromatography,
enzymatic hydrolysis and 31P-NMR confirmed that amylose was cross-linked
with amylopectin, no cross-linkages among amylose molecules existed, and
that amylose molecules were molecularly interspersed inside the granule
(Jane et al., 1992; Kasemsuwan & Jane, 1994).
Low levels of cross-linking of potato starch by 0.05–0.10% POCl3 had no
effect on pancreatin digestion of the gelatinized starch (Janzen, 1969), but
considerable inhibition of starch hydrolysis by pancreatin was displayed at
higher levels of POCl3 treatment (0.5–1.5%). Digestibility of raw or gelati-
nized starch to pancreatic a-amylase was very slightly reduced (1–3%) by low
level cross-linking (�0.0825% POCl3) of lentil, faba bean and field pea
starches (Hoover & Sosulski, 1986). Using epichlorohydrin as a cross-linking
50 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 51
agent at 0.1%, 0.3% and 0.5% levels, starch from two Croatian wheat varieties
(Ackar et al., 2010) yielded low levels of resistant starch (0.31–2.25%)
compared to the respective native wheat starches (0.48–0.58%). By compari-
son, 0.06–0.12% STMP treatment of waxy maize starch slightly reduced the
four-hour percent hydrolysis (13.0–13.7%) of the gelatinized starch by hog
pancreatic a-amylase compared to the gelatinized unmodified waxy maize
starch (15.8%) (Hahn & Hood, 1980).
Wheat starch cross-linked with 0.1% POCl3 or 0.3% epichlorohydrin did
not generate significant RS when assayed for total dietary fibre by AOAC
Method 991.43 (Woo & Seib, 2002). However, reaction with 1–2% POCl3 or
1–2% epichlorohydrin yielded 52.7–85.6% total dietary fibre and 57.4–75.8%
total dietary fibre, respectively. The cross-linking conditions were 33% starch
solids, 15% Na2SO4, pH 11.5, reaction temperature of 25 �C, and one hour
reaction time. The use of 12% of a 99 : 1 blend of STMP and sodium
tripolyphosphate (STPP) as modifying agents at approximately the same
reaction conditions as above, except for 10% Na2SO4, reaction temperature of
45 �C, and three hours reaction time, resulted in a phosphorylated cross-linked
RS4 product with low swelling power (�3), 0.32% phosphorus and 75.7%
total dietary fibre. At a 10% level of 99: 1 ratio of STMP/STPP, the level of
phosphorus is 0.32% and total dietary fibre is 75.6%, but other ratios (25 : 75,
50 : 50, or 75 : 25) of STMP/STPP generated lower phosphorus incorporation
(0.13–0.29%) and diminished total dietary fibre (21.6–63.7%).
General cross-linking conditions of 0–20% Na2SO4, 10–19% STMP/STPP
(99 : 1 ratio), pH 11.5–12.3, reaction temperature of 25–70 �C and reaction
time of 0.5–12 hours were utilized for preparation of different phosphorylated
cross-linked RS4 wheat starches with 0.32–0.35% phosphorus and
75.7–88.1% total dietary fibre. Phosphorylated cross-linked RS4 starches
from normal wheat, waxy wheat, normal corn, waxy corn and potato showed
0.32%, 0.32%, 0.34%, 0.33% and 0.32% phosphorus and 76%, 80%, 35%,
58% and 73% total dietary fibre, respectively.
Four phosphorylated cross-linked RS4 wheat starches with phosphorus
contents of 0.13–0.38% and total dietary fibre levels of 14.0–93.4% (Woo &
Seib, 2002) were analyzed for the amount of RS by the Englyst method
(Englyst et al., 1992). The resulting RS levels by this method, before and after
gelatinization of the samples, correlated directly with total dietary fibre using
AOAC Method 991.43. Actual RS values ranged from 35.5–66.1% for the
ungelatinized samples and 2.6–15.2% for the gelatinized samples. Corn starch
cross-linked with 4%, 8%, and 12% of a 99 : 1 mixture of STMP/STPP
introduced 0.14–0.37% phosphorus to the granules, which was directly
proportional (R2¼ 0.9971) to the amount of RS (24.5–81.6%), as measured
by AOAC Method 991.43 (Chung et al., 2004).
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 51

3GCH03 07/19/2013 17:21:14 Page 52
Woo et al. (2006) reported on a pregelatinized, phosphorylated, cross-
linked RS4 starches from wheat, potato and tapioca that exhibited resistance
to a-amylase digestion, possessed fat replacement properties and demon-
strated stability to freeze-thaw processing. This particular pregelatinized RS4
starch was formulated in a number of reduced-fat foods, namely bakery
products, instant mashed potato, sausages, salad dressings, desserts, ice
cream, yogurt and cr�eme filling.
Although Kerr & Cleveland (1957) pioneered the cross-linking of starch at
alkaline pH using STMP, its combination with small levels of STPP during the
reaction formed the basis for patenting a phosphorylated cross-linked food-
grade starch resistant to a-amylase and the method of preparing the same
product (Seib & Woo, 1999). Analysis of phosphodextrins from a-amylase
and glucoamylase digestion of phosphorylated cross-linked RS4 wheat starch
by proton-decoupled 31P NMR (Sang et al., 2007) showed four types of
phosphate esters: cyclic-monostarch monophosphate (cyclic-MSMP), mono-
starch monophosphate (MSMP), monostarch diphosphate (MSDP), and dis-
tarch monophosphate (DSMP), with signals centred at d 15.9 ppm, d 3–5 ppm,
d�5 and�10 ppm, and d�1 to1 ppm, respectively. Phosphorylation of wheat
starch with STMP/STPP produced �37% MSMP and 63% DSMP, whereas
the use of POCl3 changed the proportion to �20% MSMP and 80% DSMP
(Sang et al., 2007).
Starch hydroxyls have ionization constants between pKa 12.5–13.0 at
45 �C (Lammers et al., 1993). Less than 1% of hydroxyl groups is ionized at
pH 10.5, and about 5–10% are ionized at pH 11.5. Depending on the pH used
during the reaction with STMP, the ratio of DSMP/MSMP can vary from
41 : 22 for pH 10.5 to approximately 65 : 35 for higher pH (11.5–12.5). The
lower-reaction pH of 10.5 also produced 18% MSDP and 19% cyclic-MSMP.
The proposed reaction mechanism between starch and STMP at alkaline
pH involves the initial attack by a starch alcoholate ion, resulting in ring-
opening of STMP and formation of a monostarch triphosphate intermediate.
This is followed by the reaction of another starch alcoholate ion with the
intermediate, forming DSMP and pyrophosphate. The formation of MSDP
occurs by losing the g-phosphoryl group on the monostarch triphosphate by
the peeling mechanism. Cyclic-MSMP and MSDP are reactive at high
alkalinity (pH 11.5 for 3 hours), which causes the disappearance of cyclic-
MSMP and the reduction of MSDP from 17% to 11%. The DSMP content of
phosphorylated cross-linked RS4 wheat starch correlated positively (r¼ 0.96;
P¼ 0.02) with RS content (as determined by the method of Englyst et al.
(1992)) and with total dietary fibre content (r¼ 0.90; P¼ 0.05) as assayed by
AOAC Method 991.43.
52 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 53
Sang et al. (2010) reported a phosphorylated cross-linked RS4wheat starch
(0.37% phosphorus, 88.8% total dietary fibre) exposed to NaOH at pH 9–11
and 40 �C for four hours that did not significantly change the phosphorus
content (0.35–0.37%), even with the elevated pH treatment. A much higher
alkalinity of pH 12 at the above time-temperature combination reduced the
phosphorus content to 0.29%, which indicates that about 22% of covalently-
bound phosphate was removed from the starch. In addition, cyclic-MSMP and
MSDP disappeared, as confirmed by the absence of 31P NMR signals, which
suggests the instability of those two types of phosphate esters at pH 12.
However, the cross-linked phosphate ester, DSMP, increased to �18% after
exposure at pH 12 and 40 �C for four hours, indicating that new DSMP was
formed from the reaction of MSDP or cyclic-MSMP with hydroxyl groups on
another starch chain. Total dietary fibre content was little changed (88.8% vs.
86.5%) upon exposure to pH 12, even though the total phosphorus decreased
by 22%. The 18% increase of cross-linked phosphate esters (DSMP) com-
pensated for the 22% loss of total phosphorus. The integrated NMR intensities
of DSMP andMSMP indicated that they contained 0.17% phosphorus (or 46%
of total phosphorus) and 0.077% phosphorus (or 21% of total phosphorus),
respectively.
Starch citrate is the nomenclature used in the literature for the cross-linked
product derived from high-temperature treatment of a low-moisture mixture
of starch and citric acid, a six-carbon tricarboxylic acid. The reaction
mechanism involves dehydration of citric acid to anhydride form, and this
functional group consequently forms cross-linking bonds with starch mole-
cules. Klaushofer et al. (1978) developed a cross-linked starch using citric
acid as the primary modifying agent. The process involved drying to 5–20%
moisture an aqueous mixture of starch and citric acid (5–40%, starch basis)
and heating for 1–5 hours at 110–140 �C to yield starch citrate. This product
exhibited decreased in vitro amylase digestibility with an increasing degree of
esterification. In addition, the esterified starch showed improved stability to
shear and freeze-thaw processing conditions.
Xie & Liu (2004) reported that for a corn starch/citric acid weight ratio of
5 : 2, pH 3.5 and a reaction temperature range of 120–150 �C for 3–9 hours, the
DS varied from 0.09–0.12 and the level of RS ranged from 41.1–78.8%. An
improvement of RS content to 92.9% was achieved when a reaction tempera-
ture of 150 �C was employed. On the other hand, Wepner et al. (1999)
obtained 45.9–57.5% RS from starch citrates prepared from potato, pea, corn,
and wheat starches using the following reaction conditions: starch/citric acid
weight ratio of 5 : 2, pH 3.5 and five hours at 140 �C. The DS expressed as
percent esterified citric acid ranged from 12.2–14.4%.
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 53

3GCH03 07/19/2013 17:21:14 Page 54
Citric acid-treated starch containing slowly digestible and resistant starch
fractions was described by Shin et al. (2007). The preparation procedure
consisted of a two-step process of autoclaving (121 �C for 30 minutes) an
aqueous mixture of starch-citric acid, followed by heat treatment as high as
130 �C for up to 24 hours. The resulting product has a dextrose equivalent
(DE) of 10.2, DS of 0.027, cold water solubility of 55.2%, slowly digestible
and resistant starch fractions of 54.1%, and low blood glucose response
in mice.
Other researchers reacted glutaric acid, a five-carbon dicarboxylic acid,
with adlay starch in preparing cross-linked RS4 resistant starch and charac-
terizing its properties (Kim et al., 2008). Starch glutarate with high RS content
of 65–66% was prepared from adlay starch using the following reaction
conditions: 30–40% glutaric acid (starch basis), 115–130 �C, and 6–7.5 hoursof reaction time. The modified product displayed a carbonyl absorption band
at 1730 cm�1 by FT-IR and 173 ppm and 32 ppm CP-MAS 13C NMR peaks
assigned to carbonyl and methylene groups, respectively, suggesting cross-
linking between starch chains by esterification. It was observed that formation
of b-1,6-glycosidic bonds may have also occurred during the preparation of
starch glutarate.
3.3.2 Substituted RS4 starches
Food-grade substituted starches used for technological effects are normally
treated with appropriate modifying agents (for example, acetic anhydride or
propylene oxide) to yield a DS or a molar substitution (MS) of �0.1 (Mason,
2009). These esterified or etherified starches have the general properties of
increased swelling, elevated viscosity and enhanced clarity, and their pastes
have improved stability to room temperature, as well as refrigerated or freezer
storage.
Pancreatin digestibility of gelatinized hydroxypropylated potato starch
decreased with increasing DS (Leegwater & Luten, 1971). A logarithmic plot
of DS (0.02–0.45) and reducing power (2.6–61.0mg glucose per mmol chain
unit) demonstrated that the digestibility decreased exponentially with increas-
ing DS. In a related observation, Klaushofer et al. (1978) reported declining in
vitro amylase digestibility with an increasing degree of esterification with
citrate. Furthermore, an increasing degree of hydroxypropyl substitution
among cereal starches resulted in a diminished in vitro enzymatic hydrolysis
(Azemi & Wooton, 1984).
Hahn & Hood (1980) showed that gelatinized hydroxypropylated waxy
maize starch (MS¼ 0.095–0.131) was digested to 11.4% after four hours of
hydrolysis with hog pancreatic a-amylase, compared to 15.8% hydrolysis of
54 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 55
the gelatinized unmodified waxy maize starch. In another study, the digest-
ibility of gelatinized wheat starch by porcine pancreatic a-amylase was
reduced by 13.3–23.4% by substitution with hydroxypropyl (DS¼ 0.06) or
acetyl groups (DS¼ 0.07), while cross-linking with phosphate (0.05% POCl3)
had a minor lowering effect (2.5%) on its digestibility (Wooton & Chaudhry,
1979). Etherification with hydroxypropyl groups lowers the digestibility of
wheat starch to a larger extent (23.7–25.2%) than esterification with acetyl
groups (10.8–13.3%), most likely due to the larger influence of the more bulky
hydroxypropyl group on enzyme digestion.
The relative degree of hydrolysis to hog pancreatic a-amylase
(0.0175–0.30% enzyme concentration based on starch) was 75.3–80.1%
for gelatinized tapioca hydroxypropyl distarch phosphate (1.6% hydrox-
ypropyl) and 50.6–58.2% for gelatinized hydroxypropylated tapioca starch
(4.2% hydroxypropyl), compared to 100% for gelatinized unmodified
tapioca starch (Conway & Hood, 1976). These digestibility values are
comparable to other studies conducted on hydroxypropylated starches by
Leegwater & Luten (1971) and by Hood & Arneson (1976).
Highly hydroxypropylated (DS¼ 0.12), lightly cross-linked (0.000085%
POCl3) waxy starch showed decreased digestibility (34.4%) to a-amylase/
glucoamylase compared to unmodified waxy starch (98.0%) (Wolf et al.,
1999). Moderately hydroxypropylated (DS¼ 0.07), moderately cross-linked
(0.00037% POCl3) dullwaxy starch displayed decreased digestibility (68.7%)
compared to unmodified dull waxy starch (90.6%).
Due to their beneficial health effects, acylated starches that possess much
higher DS compared to traditional food-grade modified starches were sug-
gested for possible use in food products (Annison et al., 2003; Bajka et al.,
2010). Acylated starches as a form of substituted RS4 starch was borne out of
a strategy to deliver short chain fatty acids directly to the large bowel (Annison
et al., 1995). The 2-, 3- or 4-carbon short-chain fatty acids represented by
acetic acid, propionic acid or butyric acid, respectively, can be attached to
starch by an esterification process. Annison et al. (2003) reported on the
preparation of these esters with a DS of approximately 0.20 by reacting 600 g
starch with 115ml, 180ml or 230ml of acetic anhydride, propionic anhydride
or butyric anhydride, respectively.
Butyrate esters of corn starch tended to increase starch resistance to
enzymic hydrolysis in vitro and to intestinal amylolysis in vivo (Annison
et al., 1995, 2003). Butyrate is released in the large bowel by the action of
microbial esterases and lipases, leaving the starch backbone available for
fermentation (Bajka et al., 2006, 2007). A butyrylated starch product with a
DS of approximately 0.3 was exceedingly resistant to amylolysis, but was still
capable of liberating butyrate when exposed to faecal microflora (Bajka et al.,
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 55

3GCH03 07/19/2013 17:21:14 Page 56
2007). Less butyrate is released when the DS is greater than 0.3, which may be
explained by decreased solubility or increased steric obstruction to the action
of microbial esterases.
Waxy maize starch esterified with 3% octenyl succinic anhydride showed
28.3% slowly digestible starch, which rose to 42.8% after the succinylated
starch was heat-moisture treated at 120 �C for four hours and 10% moisture
(He et al., 2008). Cooked 1-octenyl succinylated corn starch (DS¼ 0.07;
DE� 3) decreased the extent of 15-hour in vitro hydrolysis (70%) by
a-amylase/amyloglucosidase (Wolf et al., 2001) compared to cooked,
unmodified corn starch (99.9%). Cooking of butyrylated high-amylose
corn starch increased in vitro a-amylase/amyloglucosidase hydrolysis from
6% to 43% (Bajka et al., 2006).
3.3.3 Pyrodextrinized RS4 starches
Pyrodextrins are conventionally produced by treatment of granular native
starch at low moisture contents and elevated temperatures with varying
amounts of mineral acids and, sometimes, carboxylic acids (Wurzburg,
1986; Tomasik et al., 1989; Wang et al., 2001; Laurentin et al., 2003; Huber
& BeMiller, 2010). These conditions destroy the original starch backbone
structure and promote hydrolysis, transglucosidation and repolymerization.
As a result, pyrodextrins have low molecular weights, possess atypical
glycosidic linkages (i.e. (1! 2)-, (1! 3)-glycosidic bonds) and contain
new branched structures.
Using dextrinization conditions of pH 2.5–2.6, a temperature of 170 �C,and a three-hour reaction time, Wang et al. (2001) demonstrated, by reducing
sugar analysis, that hydrolysis occurred during the first hour, followed by
transglucosidation and repolymerization in the remaining two hours. This
result agreed with the low content of enzyme resistant dextrins during the first
hour of pyrodextrinization, followed by a rapid increase of enzyme-resistant
dextrins in the next two hours.
Pyrodextrinization, followed by amylase treatment, was the basis for a
number of patents to produce soluble dietary fibres whose nomenclature may
fall under different names, such as indigestible dextrins, resistant dextrins,
resistant maltodextrins or digestion-resistant maltodextrins (Ohkuma et al.,
1994, 1995, 1997). A product of this type dissolves into a clear and stable
aqueous solution with low viscosity. It has a DE of 10–12, an average
molecular weight of 2000Da, and its structure is composed of (1! 2)-,
(1! 3)-, (1! 4)-, and (1! 6)-glycosidic linkages and levoglucosan
(Ohkuma & Wakabayashi, 2001; Okuma & Kishimoto, 2004). It is approxi-
mately 90% indigestible, with a low caloric count (0.5 kcal/g). Resistant
56 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 57
maltodextrins that are available commercially vary in total dietary fibre
content, ranging from 52–90%, and they are being offered either as a
hydrogenated version, in agglomerated form, in admixture with honey solids
and stevia glycosides, or in liquid corn syrup version (75% solids).
Another manufacturing process for resistant dextrins involves controlled
dextrinization, wherein the starch undergoes a degree of hydrolysis followed
by repolymerization (Lefranc-Millot et al., 2010). This process converts
starch into dietary fibre through the formation of non-digestible glycosidic
bonds that deter cleavage by amylase enzymes. When analyzed by AOAC
Method 2001.03, the resistant dextrin obtained in this process contains 85%
total dietary fibre, which consists of 50% insoluble fibre in ethanol and 35%
resistant oligosaccharides. In another study, lightly, moderately and highly
converted dextrins from common corn starch displayed digestibilities of
90.1%, 84.2% and 63.8%, respectively, when treated with a mixture of
a-amylase and glucoamylase, whereas raw common corn starch has 97.9%
digestibility (Wolf et al., 1999).
Formation of enzyme-resistant fractions during pyrodextrinization was
shown to be affected by the botanical source of starch (Laurentin et al., 2003).
These researchers also revealed that granule morphology of native starch was
similar to that of pyrodextrinized starch, and that the latter exhibited no
endotherm by differential scanning calorimetry (DSC). Extensive
depolymerization of pyrodextrinized starches occurred as evident in the
gel filtration profile, and the amount of starch hydrolysable by the combined
action of Termamyl a-amylase and amyloglucosidase was significantly
decreased.
3.4 PHYSICOCHEMICAL PROPERTIES AFFECTINGFUNCTIONALITY
The chemical name of a phosphorylated cross-linked RS4 starch produced by
treatment with a mixture of STMP and STPP at a 99 : 1 ratio (Seib & Woo,
1999) is phosphated distarch phosphate (E-Number 1413 and CAS No.
977043-58-5). The use of these two phosphate-modifying agents is regulated
to yield a food-grade product with nomore than 0.4% phosphorus, as specified
in the section on Food-StarchModified of the US Code of Federal Regulations
Title 21 Part 172.892 (Code of Federal Regulations, 2010). Commercial-scale
production yielded a product that delivered a minimum of 85% (dry basis)
total dietary fibre with an average of �94% (dry basis) total dietary fibre by
AOAC Method 991.43 (Woo et al., 2009). The fibre exists primarily as
insoluble fibre. A phosphorylated cross-linked RS4 wheat starch was
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 57

3GCH03 07/19/2013 17:21:14 Page 58
described in a US patent publication to contain 93.4% total dietary fibre and
0.38% phosphorus (Seib & Woo, 1999).
Phosphorylated cross-linked RS4 starches from wheat, corn, potato and
rice are practically insoluble in dimethyl sulphoxide or 1M KOH (Woo &
Seib, 2002; Shin & Seib, 2004). Scanning electron microscopy showed the
same shape and smooth surface as their parent starches (Seib & Woo, 1999).
Pasting curves at 8% starch solids did not rise above the baseline when heated
from 30 �C to 95 �C. Wide-angle x-ray diffraction showed phosphorylated
cross-linked RS4 starches from cereal, and potato sources gave A- and B-type
polymorphic crystal patterns, respectively.
When analyzed by DSC, the transition temperatures of phosphorylated
cross-linked RS4 potato starch were slightly affected. In contrast, the other
cross-linked RS4 starches from normal wheat, waxy wheat, normal corn
and waxy corn gave elevated To, Tp, and Tc by 4.3–8.4 �C, 5.4–8.3 �C, and3.6–10 �C, respectively (Woo & Seib, 2002). Enthalpy of gelatinization
tended to be lower for all the RS4 starches, except for RS4 potato starch.
It was surmised that annealing of starch occurred during exposure to the cross-
linking reaction conditions, and that cross-linking could inhibit cooperative
melting of crystals in starch granules (Woo & Seib, 2002; Jacobs et al., 1995).
No significant differences in DSC transition temperatures and enthalpies of
gelatinization were found after a phosphorylated cross-linked RS4 wheat
starch was held at pH 9.0–12.0 for four hours at 40 �C (Sang et al., 2010). This
alkaline pH treatment did not alter its typical A-type x-ray diffraction pattern
or its degree of crystallinity.
The phosphorus content of phosphorylated cross-linked RS4 corn starch
modified with 4–12% of a 99 : 1 blend of STMP/STPP was inversely
proportional (R2¼�0.9537) to the swelling volume of the cross-linked
product (Chung et al., 2004). Cross-linking has the added effect of increasing
the glass transition temperature of corn starch in excess water by nearly 1 �C,which was explained as a reduction in chain mobility, such that a higher
temperature (Tg) is required to induce movement. On the other hand, the glass
transition temperature of the cross-linked starches measured at 15% moisture
decreased by �1 �C, which was attributed to internal plasticization of starch
by the ionic phosphate groups. Furthermore, the retrogradation enthalpy after
one week of storage of the gelatinized starches (67% moisture) at 4 �C was
decreased by cross-linking.
Cross-linking of banana starch with 11.9% STMP and 0.1% STPP raised
the DSC peak temperature of gelatinization to 86.6 �C, compared to 79.2 �Cfor the parent native banana starch, while the enthalpy of gelatinization
decreased to 9.36 J/g from 25.1 J/g for the parent native banana starch
(Aparicio-Saguilan et al., 2008). Autoclaving at 121 �C for one hour, followed
58 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 59
by storing at 4 �C for 24 hours, decreased the RS content (total dietary fibre
assay) of phosphorylated cross-linked banana starch from 94.68% to 85.35%.
As a result of autoclaving and cooling treatments, the peak temperature and
enthalpy of gelatinization decreased to 68.5 �C and 1.5 J/g, respectively.
When a phosphorylated cross-linked RS4 wheat starch was heated in a
cooking viscometer (8% solids) from 30 �C to 95 �C, no pasting curve (i.e. flatline) was generated, which is indicative of the highly restricted swelling of the
cross-linked starch granules (Woo & Seib, 2002; Xie & Liu, 2004). The
granules remained intact after heating, and it was observed that disintegrated
or convoluted granules were not evident, and neither were granule ghosts.
DSC analysis of phosphorylated cross-linked RS4 wheat starch demonstrated
elevated initial, peak, and conclusion temperatures (4.3–10.5 �C higher), but a
slight reduction in enthalpy of gelatinization (0.9 J/g lower) compared to the
parent native wheat starch.
When heated from 35 �C to 60 �C in excess water, the granules of a
commercial phosphorylated cross-linked RS4 wheat starch maintained their
structures essentially unchanged (Ratnayake & Jackson, 2008). Above 65 �Cand up to 85 �C, the granules became increasingly swollen. Coinciding with
this observation, the DSC enthalpies of gelatinization did not change during
the 35–60 �C treatment, but gradually disappeared between 60 �C and 85 �C.X-ray crystallinity gradually decreased within the same temperature range of
60–85 �C, but the A-type polymorph was preserved up to 70 �C. Consistentwith previous studies (Seib & Woo, 1999; Woo & Seib, 2002), Ratnayake &
Jackson (2008) found that phosphorylated cross-linked RS4 wheat starch did
not completely dissolve in 90% dimethyl sulphoxide.
A phosphorylated cross-linked RS4 wheat starch with 72.9% total dietary
fibre exhibited a low swelling power of 2.8 g/g and low solubility in water of
0.5% at 95 �C (Woo & Seib, 2002). Compared to other cereal starches, it
displayed similar water vapour sorption and desorption isotherms at 25 �C and
at water activities below 0.8 (Shin et al., 2003).
Lefranc-Millot et al. (2010) described a resistant pyrodextrin that has a
clean, neutral taste, possesses no sweetness and is compatible with sugar-free
claims. The commercial product dissolves rapidly in aqueous systems and has
limited impact on viscosity. It is stable to high temperature, variable pH and
high shear processing. The resistant dextrin is officially recognized as a
soluble dietary fibre, with suggested ingredient labelling of ‘dextrins’ in food
product packages.
Consistent with heavily cross-linked starches, corn starch citrates resisted
granular swelling as exhibited by the flat RVA pasting curve (i.e. 0 RVU) when
heated from 50 �C to 95 �C at 8% starch solids (Xie & Liu, 2004). Heating
starch citrates in water (7% solids) at 100 �C for 30 minutes resulted in
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 59

3GCH03 07/19/2013 17:21:14 Page 60
9.8–16.8% loss in RS content, indicating intermediate thermal stability of this
product. The DSC enthalpy of gelatinization tended to decrease in starch
citrates (0.4–3.0 J/g), compared to the native corn starch (6.7–15.6 J/g).
Analysis by DSC demonstrated that adlay starch glutarates with RS
content of 30–66% exhibited lower To (46.0–49.9 �C), Tp (60.5–65.6 �C),Tc (73.1–73.9
�C), andDH (5.7–9.5 J/g) compared to control starch (To, 60�C;
Tp, 71.3�C; Tc, 82.6
�C; D H, 13.3 J/g), presumably because of the disruption
of molecular (double helical) and crystalline orders during the preparation of
the glutarates (Kim et al., 2008). Hot water solubility at 80 �C of the glutarate
derivatives was drastically reduced to 0.71–1.52%, compared to 81.2% for the
control starch. When subjected to a-amylase and amyloglucosidase treat-
ments, gelatinized starch glutarate lost its granular shape and its granules
cohered together. Its good stability to heat treatment in excess water was
demonstrated when the RS content before heating of starch glutarate was
shown to be similar to that after heating.
3.5 PHYSIOLOGICAL RESPONSES AND HEALTHBENEFITS
Dietary fibre sources, including RS, promote a number of physiological
benefits in humans, which include, but are not limited to, the following:
decreased intestinal transit time; enhanced satiety; reduced postprandial blood
glucose and/or insulin levels; diminished blood total and/or low-density
lipoprotein cholesterol concentrations; and fermentability by colonic micro-
biota to give short-chain fatty acids (SCFA) (Baghurst et al., 1996; Topping &
Clifton, 2001; Nugent, 2005; Topping, 2007;Witwer, 2008; Buttriss & Stokes,
2008; Bird et al., 2009).
Different types of RS can elicit significantly different glycemic responses,
as demonstrated by a comparison between RS2 high-amylose corn starch
and phosphorylated cross-linked RS4 wheat starch in human subjects
(Haub et al., 2010). The glucose response and the incremental area under
the curve of phosphorylated cross-linked RS4 wheat starch were signifi-
cantly decreased, compared with RS2 high-amylose corn starch and dex-
trose (control).
Aqueous dispersions of two RS4 potato starches (38 grams in 296ml of
water), when consumed as a drink (Haub et al., 2012), elicited a significantly
decreased glycemic response compared to dextrose (control). There were no
blood glucose response differences and no satiety effects when the same doses
of RS4 potato starches were added to dextrose and compared with dextrose
alone. Rice kernels coated with RS4 wheat starch using agar and locust bean
60 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 61
gum had lower in vitro starch digestibility, a decreased glucose response in
rats and a slower rate of blood glucose decrease, compared to uncoated rice
and rice mixed with RS4 wheat starch (Choi et al., 2010).
Using the glycemic index protocol, nutritional bars formulated with 34
weight percent of phosphorylated cross-linked RS4 wheat starch displayed
attenuated postprandial blood glucose and insulin levels, when compared to a
control glucose drink and to another nutritional bar in which the above RS4
wheat starch was replaced by an equivalent amount of puffed wheat
(Al-Tamimi et al., 2010). Incremental areas under the glucose curves were
140, 84 and 28mmol/l� 2 hour for the glucose control drink, puffed wheat bar
and cross-linked RS4 bar, respectively. The corresponding incremental areas
under the insulin curves were 17 575, 8758, and 3659 pM� 2 hour, respec-
tively. The results are noteworthy, as the bar formulas contain high levels of
glycemic carbohydrates, in the form of 11% brown sugar and 20% corn syrup.
Other researchers compared the properties in vivo of phosphorylated cross-
linked RS4 wheat starch and RS2 high-amylose corn starch in order to gauge
their impact on gastrointestinal microbiota composition and metabolism in
human volunteers (Martinez et al., 2010). During the three-week feeding
period, both types of resistant starch were well tolerated by the subjects when
fed daily with 100 grams of snack crackers (containing 33 grams of dietary
fibre contributed by the resistant starch ingredient). Pyrosequencing of faecal
samples demonstrated that, at the phyla level, RS4 crackers significantly
decreased Firmicutes and increased Bacteroidetes and Actinobacteria. These
changes were associated with a decrease in the family Ruminococcaceae and
increases in the genera Parabacteroidetes and Bifidobacterium.
Denaturing gradient gel electrophoresis revealed that RS4 crackers induced
a swift and reversible increase in band intensity of Bifidobacterium adoles-
centis, whereas RS2 crackers caused a gradual rise. Quantitative enumeration
of bifidobacteria by qRT-PCR confirmed the significant increase in cell
numbers during consumption of RS4 and RS2 crackers, compared to the
control crackers (no resistant starch). The total cell numbers of bifidobacteria
increased more than three-fold with RS4 crackers, while RS2 crackers
doubled the cell numbers.
In vitro digestion of phosphorylated cross-linked RS4 wheat starch and
phosphorylated cross-linked RS4 potato starch by successive treatments with
pepsin and pancreatin-bile yielded 82% and 74% of indigestible residues,
respectively (Thompson et al., 2011). Incubation of these indigestible residues
with fresh human faecal inoculum produced gases that increased linearly over
the 24-hour fermentation period, with the two RS4 starches exhibiting similar
gas production rates as well as similar rates of production of total short-chain
fatty acids. Acetic acid was the major SCFA produced, along with relatively
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 61

3GCH03 07/19/2013 17:21:14 Page 62
higher amounts of butyric acid than propionic acid. The fractional molar ratios
of the above acids (acetic : propionic : butyric) are 0.586 : 0.186 : 0.228 and
0.577 : 0.200 : 0.223 for RS4 wheat starch and RS4 potato starch, respectively.
Caloric contribution was calculated to be one-third lower for the two RS4
starches, compared to unmodified (non-resistant) starch.
In an animal feeding trial, hamsters on a phosphorylated cross-linked RS4
wheat starch diet consumed less feed and gained less weight than those on a
cellulose diet (Seib &Woo, 1999). Total serum cholesterol in the animals was
not significantly different between the two diets, but the concentration of high-
density lipoprotein cholesterol was significantly higher in the RS4 diet.
Consequently, the calculated levels of low- and very low-density lipoprotein
cholesterols were less in the serum of animals consuming the RS4 diet than the
cellulose diet. Analysis of fermentation products from the caecum of hamsters
showed elevated levels of SCFA (in particular butyric acid) in the RS4 diet,
compared to the cellulose diet.
In mice fed for six weeks with a high-fat diet, supplemental addition at a
15% level of phosphorylated cross-linked RS4 corn starch and phosphorylated
cross-linked RS4 high-amylose corn starch lowered body weight gain as well
as total lipid, triglyceride and total cholesterol in the serum and liver,
compared to the corresponding unmodified starches (Lee et al., 2012).
Rats fed for four weeks with a diet containing 5% phosphorylated cross-
linked RS4 corn starch (92.6% total dietary fibre) exhibited lower serum levels
of total cholesterol (12.6% decrease) and low density lipoprotein plus very
low density lipoprotein cholesterol (24.3% decrease) than rats fed with a diet
containing 5% cellulose (Song et al., 2010). In addition, this RS4 corn starch
can potentially ameliorate allergic inflammation in the mesenteric lymph
nodes of rats, because of its impact on elevating serum immunoglobulin A
levels, as well as the CD4þ T cell population and the ratio of CD4þ/CD8þ T
cells.
Annison et al. (2003) summarized the strategies to elevate SCFA in the
large bowel as follows:
a) Increase the consumption of foods high in RS content.
b) Consume SCFA directly in foods or beverages.
c) Consume foods that contain starches esterified with specific SCFA.
The rat feeding study by these researchers confirmed that starches esterified
with specific SCFA through reaction with acetic, propionic or butyric anhy-
dride resisted digestion in the small intestines and delivered those SCFAs into
the large bowel, where they were released by the action of the microflora. The
selective increase of individual SCFAwas observed even only after three days,
62 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 63
which indicates that the rat microflora can readily adapt. Esterified starches
with a DS of around 0.20–0.25 have the potential to improve the nutritional
properties of foods through delivery of SCFA directly to the large bowel.
Ingestion by ileostomists of starches esterified with acetate, propionate or
butyrate with DS of 0.23–0.25 resulted in the recovery of 73–76% of the
esterified acid at the terminal ileum, which suggests that SCFA delivery to the
large bowel could occur in humans with an intact gastrointestinal tract (Clarke
et al., 2007). Free faecal butyrate concentrations were increased in human
subjects after consumption of butyrylated high-amylose corn starch, and about
27% of total esterified butyrate was absorbed in the small intestine, 15.8%was
recovered in the faeces, and about 57.2% was released in the large bowel
(Clarke et al., 2011a). The population of Parabacteroides distasonis was also
increased by the above-mentioned butyrylated starch derivative.
Reduction in DNA single-strand breaks was twice that in rats fed butyry-
lated high-amylose corn starch compared to unesterified high-amylose corn
starch, which was interpreted as the greater ability of the butyrylated starch to
deliver butyrate to the large bowel, thereby raising the concentration of this
particular SCFA (Bajka et al., 2008). High-protein diets increased colonocyte
DNA single-strand breaks in rats, but this effect was opposed by inclusion in
the diet of butyrylated high-amylose corn starch (Conlon et al., 2012).
Butyrylated high-amylose corn starch protects colonocytes in rats from
genetic damage by high dietary protein (Bajka et al., 2008), which may be
explained by increased caecal butyrate pools.
Bajka et al. (2006) studied the effects of feeding high-amylose corn starch
and butyrylated high-amylose corn starch in rats and the effect of cooking on
resistance to in vitro and in vivo amylolysis. Cooking of butyrylated high-
amylose corn starch increased in vitro a-amylase/amyloglucosidase hydroly-
sis from 6% to 43%. Cooked butyrylated high-amylose corn starch delivered
significantly greater amounts of esterified butyrate in the rats’ large bowel,
compared to raw or cooked high-amylose corn starch. The caecum is the main
site of bacterial fermentation of carbohydrates in rats. By comparison,
ingestion of butyrylated low-amylose corn starch by rats resulted in lower
digesta pH in the large bowel and elevated butyrate concentrations in the
caecal, proximal and distal colon, as well as in the portal vein (Bajka et al.,
2010).
Butyrylated high-amylose corn starch (DS¼ 0.25) affected the faecal
microbiota composition of rats treated with azoxymethane, a carcinogenic
agent (Abell et al., 2011). It increased propionate and butyrate concentrations
in distal colonic digesta and was associated with the appearance of Para-
bacteroidetes distasonis, Lactobacillus gasseri and Phascolarctobacterium
faecium. Following acute exposure to azoxymethane, rats fed with butyrylated
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 63

3GCH03 07/19/2013 17:21:14 Page 64
high-amylose corn starch had higher caecal tissue and digesta weights and
lower pH in the large bowel, compared with rats fed with unmodified low-
amylose and high-amylose corn starches (Clarke et al., 2011b). Morphologi-
cal assessment of the rats’ distal colonic epithelium showed that butyrylation
has a positive impact, as indicated by an elevated apoptotic index. Butyrate
concentrations in hepatic portal plasma, as well as in caecal and proximal and
distal colonic digesta, were higher in azoxymethane-treated rats fed with
butyrylated high-amylose corn starch, compared to non-acylated low-amylose
and high-amylose corn starches (Clarke et al., 2008). This butyrate effect is
associated with reduced tumour incidence, number and size in rats.
Net incremental area under the blood glucose curve (Wolf et al., 2001) was
lower (P< 0.05) after fasting healthy adult subjects consumed 25 g of
1-octenyl succinylated starch (107mmol.min/l) than that for 25 g of glucose
(127mmol.min/l). A blunted glycemic response was displayed by 1-octenyl
succinylated starch, as indicated by the calculated relative glycemic response
of 93.8% compared to that of glucose.
Digestibility in the small intestine of a resistant pyrodextrin (Lefranc-
Millot et al., 2010) is in the range of 8.7–19%, which results in a low glycemic
response (�25) and low insulinemic response (�13). The low insulinemic
response is reported to induce a high satiety feeling and to reduce postprandial
hypoglycaemia. When the resistant pyrodextrin was formulated in pasta,
beverages and biscuits, consumption of these food products displayed low
glycemic responses (Lefranc-Millot et al., 2006).
In a cross-over feeding study, 20 volunteers consuming 300 g rice with
5.7 g of resistant maltodextrins displayed significantly lower postprandial
blood insulin and glucose levels, compared to rice without resistant malto-
dextrins. When 13 volunteers ate a fast-food restaurant meal with 10 g of
resistant maltodextrins, the insulin level at the 30-minute postprandial period
was significantly reduced in comparison to the same meal without resistant
maltodextrins (Gordon, 2007).
Other researchers confirmed in vivo the blunting effect of resistant malto-
dextrins on postprandial blood glucose and insulin levels, coupled with a
decrease in intestinal glucagon (Okuma & Kishimoto, 2004; Wakabayashi
et al., 1992; Tokunaga & Matsuoka, 1999). Co-ingestion of resistant malto-
dextrins with monosaccharides or disaccharides can moderate increases in
postprandial blood glucose levels, resulting in reduced insulin requirements
and preventing fat accumulation (Ohkuma & Wakabayashi, 2001). Among
Type 2 diabetics, 10–20 g of resistant maltodextrins fed at every meal for three
months significantly reduced fasting blood glucose levels, improved glucose
tolerance and decreased HbA1C (Nomura et al., 1992; Fujiwara &Matsuoka,
1995).
64 Resistant Starch

3GCH03 07/19/2013 17:21:14 Page 65
Body fat was significantly reduced when 12 overweight males with
hyperlipidaemia were fed 30 g of resistant maltodextrins each day for three
months (Gordon, 2007). Long-term feeding of diets with added resistant
maltodextrins to patients with hyperlipidaemia had the general effect of
decreased low-density lipoprotein cholesterol and triglycerides, but increased
high-density lipoprotein cholesterol (Okuma & Kishimoto, 2004).
Supplementation of resistant maltodextrins in the diet at the rate of
7.5–15 g/day for three weeks was well tolerated by human subjects and
tended to increase the faecal Bifidobacterium population (Fastinger et al.,
2008). There was also a shift in the molar proportions of SCFA towards
butyrate. In an earlier study, resistant maltodextrins increased faecal bulk and
decreased symptoms of constipation (Satouchi et al., 1993).
Flickinger et al. (1998) determined that a dextrinized corn starch with
mixed glycosidic linkages was hardly digested in the small intestine of ileally
cannulated dogs. This dextrin has an estimated caloric value of 2.2 kJ/g.
Another mixed-linkage resistant dextrin (non-viscous soluble dietary fibre)
described by Lefranc-Millot et al. (2010), and made either from wheat or corn
starch, has a caloric value of 2 kcal/g (Vermorel et al., 2004), which is in
agreement with the above reported caloric content.
3.6 PERFORMANCE IN FOOD AND BEVERAGEPRODUCTS
Product developers and designers have an arsenal of conventional dietary
fibres and resistant starch ingredients to boost the fibre content of food and
beverage products for nutrient and calorie labelling claims (Baghurst et al.,
1996; Erickson, 2005; Topping, 2007; Witwer, 2008; Bird et al., 2009).
Resistant starches from different sources and belonging to RS1–RS5 types are
traditionally white in colour and possess fine particle size and neutral flavour.
However, they have distinct differences in water-holding capacity that con-
note formulation changes because of the impact on water absorption and
processing parameters of bakery foods (Woo et al., 2009).
A number of bakery, pasta, noodle, dairy, snack and confectionery products
were formulated with a phosphorylated cross-linked RS4 wheat starch to
enhance fibre content, lower net available carbohydrates and reduce caloric
counts (Maningat et al., 2005, 2008). A white pan bread (Yeo & Seib, 2009)
containing one part of phosphorylated cross-linked RS4 wheat starch blended
with nine parts of wheat flour had a slightly lower specific volume (5.96 cc/g)
and slightly firmer texture (427.8 g), compared to a native wheat starch
formula (6.07 g/cc; 475.2 g) and a control wheat flour formula (6.28 g/cc;
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 65

3GCH03 07/19/2013 17:21:14 Page 66
406.9 g). In the same study, sugar-snap cookies were used to compare the
performance of phosphorylated cross-linked RS4 wheat starch with RS2
potato starch. Both starches produced cookies with top grain and spread
factors comparable to the control flour. However, cookies made with RS4
wheat starch exhibited similar snapping force, but RS2 potato starch demon-
strated significantly lower snapping force when compared to the control
cookie.
In a related study, Erickson (2005) formulated an oatmeal chip cookie by
total replacement of cookie flour with RS4 resistant starch. The usage level of
12.78% RS4 resistant starch in the cookie formula delivered a ‘good source of
fibre’ claim (3 g of dietary fibre per 30 g serving size).
High-protein, high-fibre white or whole wheat bread doughs formulated
with 11.6% of phosphorylated cross-linked RS4 wheat starch (based on total
formula weight) had 4–14% higher water absorption, 3–5 minute shorter
mixing time, 17–23 minute shorter proof time, and a baking time about four
minutes longer than the respective control doughs (Maningat et al., 2005).
The high-protein, high-fibre breads displayed greater loaf volume (260–325
cc higher) and higher moisture (3.2–4.6% higher), protein (6.2–9.0%
higher), and dietary fibre (12.4–15.7% higher), along with a �27% average
reduction in calories (by calculation), compared to the respective control
doughs.
Flour tortillas formulated with 5%, 10%, and 15% of phosphorylated cross-
linked RS4 wheat starch generated doughs that were characteristically soft,
extensible and easy to spread, with minimal shrink-back after pressing
(Alviola et al., 2010). The diameters of tortillas containing 10% and 15%
RS4wheat starch were significantly larger (P< 0.05), but the thickness tended
to decrease, compared to the control tortillas. The calculated specific volume
tended to increase as the dosage of RS increased. Flour tortillas with 15% RS
wheat starch had 14.3% dietary fibre and produced significantly higher
(P< 0.05) overall acceptability scores than the control tortillas (dietary
fibre¼ 2.8%).
A pregelatinized version of phosphorylated cross-linked RS4 resistant
wheat starch showed a significant (P< 0.001) and linear (R2¼ 0.97) trend of
increasing flour tortilla weight with an increased level of RS substitution
(Alviola et al., 2010). This increased water requirement results in competition
with gluten for water yielding doughs that were less extensible (resisted
spreading). The resulting tortillas were less puffed and denser than the control
tortillas, but had significantly increased tortilla weight by up to 6.2% and
dietary fibre by 13.6% at a 15% level of substitution.
The addition of RS2 and RS4 high-amylose corn starches in pasta at
2.5–10.0% levels significantly decreased water absorption and swelling index,
66 Resistant Starch

3GCH03 07/19/2013 17:21:15 Page 67
compared to control pasta (Bustos et al., 2011). As the level of RS4 starch
increases, the hardness of pasta tends to increase, but springiness, cohesive-
ness and chewiness tend to decrease.
Corn starch cross-linked with 4–8% of a 99 : 1 mixture of STMP and STPP
produced fried batters with improved crispiness and hardness and reduced oil
uptake, as indicated by the higher TA.XT2 peak number and peak force, along
with lower oil content compared to unmodified corn starch (Han et al., 2007).
The phosphorylated cross-linked RS4 corn starch may have formed a rigid
matrix in the fried batter that increased batter hardness and inhibited oil
absorption during frying.
To measure the textural contribution and test the integrity or survivability
of phosphorylated cross-linked RS4 wheat starch under adverse processing
conditions, it was subjected to a Wenger TX-57 twin-screw extrusion process
during the preparation of a ring-shaped breakfast cereal containing 0%, 5%,
10%, 15% and 20% of the RS4 wheat starch (Miller et al., 2011). Analysis of
total dietary fibre (AOAC Method 991.43) of the extruded breakfast cereals
showed that RS4 wheat starch retained about 78–89% of its fibre content after
extrusion, with an average fibre retention of around 83%. Product density
increased as the level of RS4 wheat starch increased, but internal cell wall
thickness and size or porosity were not affected. Higher addition of levels of
15% and 20% RS4 wheat starch decreased cereal ring diameter, but increased
initial (dry) cereal crispness and extended bowl life.
While not considered a food-grade starch in the USA and other countries,
starch citrate was evaluated for its performance in pasta and bakery products
by Wepner et al. (1999). Starch citrates from different botanical sources
(potato, pea, wheat, and corn), with 12.2–14.4% esterified citric acid and with
RS content ranging from 45.9–57.5%, were formulated in toast bread by
replacing 10% of the flour in the formula. Toast breads enriched with starch
citrate appeared more yellow in colour, exhibited coarser crumb structure and
possessed slightly lower loaf volume than the control bread. Approximately
60–85% retention of RS content was observed during the preparation of toast
bread.
In another type of baked product, wafers prepared by replacing 7.5–15.0%
of flour with starch citrate displayed a lower percent retention (around 50%) of
RS content compared to toast bread. The sensory attributes of the starch
citrate-fortified wafers were described as harder in texture, lighter in colour,
friable and more brittle, compared to the control wafer.
The addition of 7.5–15.0% starch citrate in a pasta formula caused a
reduction in quality, as demonstrated by reduced firmness of the cooked pasta
and increased cooking loss (Wepner et al., 1999). Twin-screw extrusion of
starch citrates at low shear force resulted in retention of 52–85% of the
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 67

3GCH03 07/19/2013 17:21:15 Page 68
theoretical RS content. By contrast, extrusion at high shear force showed
lower retention (29–65%) of the theoretical RS content. The extrudates in both
treatments had a comparable expansion index, water absorption index and
water solubility index, but extrusion at a low shear force caused a higher
breaking force (13.66–33.28N) compared to extrusion at a high shear force
(7.79–23.40N).
3.7 CONCLUSIONS AND FUTURE PERSPECTIVES
Physical activity, portion control and wholesome foods define a person’s
healthy lifestyle. While the first two items are behavioural in nature, the third
can be designed in foods. With health and wellness soundly resonating with
consumers, food developers are constantly challenged to design consumer
food products that not only have superior taste and texture, but are also
nutritious.
Resistant starch ingredients, in general, provide an easy solution to assist
food scientists in tailoring foods with remarkable end-use qualities and that
address health concerns. Since Hi-MaizeTM was released in 1993 as the first
resistant starch ingredient, the growth of RS has been phenomenal, with at
least 30 RS ingredients now sold and marketed worldwide. As discussed in
this chapter, a range of RS4-type resistant starches, in addition to the other
categories of RS, are available in the tool box of food designers. Prevailing
evidence confirms a host of physiological benefits and food product func-
tionalities of RS4-type resistant starches. There are opportunities for research
scientists and food technologists to identify additional useful attributes of
commercially-available resistant starch ingredients other than for caloric
reduction or as a fibre source.
REFERENCES
Abell, G.C.J., Christophersen, C.T., McOrist, A.L., Clarke, J.M. (2011). Dietary resistantstarch and butyrylated starches have different effects on the fecal bacterial flora ofazoxymethane-treated rats. British Journal of Nutrition 105 (10), 1480–1485.
Ackar, D., Babic, J., Subaric, D., Kopjar, M., Milicevic, B. (2010). Isolation of starch fromtwo wheat varieties and their modification with epichlorohydrin. Carbohydrate Poly-mers 81, 76–82.
Ackerson, C.W. (1961). High-amylose corn in a diet for day-old chicks. Feed Age 11,25–27.
Ahmed, R., Segal, I., Hassan, H. (2000). Fermentation of dietary starch in humans.American Journal of Gastroenterology 95, 1017–1020.
68 Resistant Starch

3GCH03 07/19/2013 17:21:15 Page 69
Al-Tamimi, E. Seib, P.A., Snyder, B.S., Haub, M.D. (2010). Consumption of cross-linkedresistant starch (RS4XL) on glucose and insulin responses in humans. Journal ofNutrition and Metabolism: Article ID 651063, 6p.
Alviola, J.N., Jondiko, T., Awika, J.M. (2010). Effect of cross-linked resistant starch onwheat tortilla quality. Cereal Chemistry 87, 221–225.
American Association of Cereal Chemists . (2001). The definition of dietary fiber. CerealFoods World 46, 112–126.
Annison, G., Topping, D.L., Illman, R.J. (1995). Fatty acid delivery system comprising ahydrolysable bond. Commonwealth Scientific and Industrial Research Organization.Patent No. WO 9513801. http://ep.espacenet.com.
Annison, G., Illman, R.J., Topping, D.L. (2003). Acetylated, propionylated or butyrylatedstarches raise large bowel short-chain fatty acids preferentially when fed to rats. Journalof Nutrition 133, 3523–3528.
Aparicio-Saguilan, A., Gutierez-Meraz, F., Garcia-Suarez, F.J., Tovar, J., Bello-Perez, L.A.(2008). Physicochemical and functional properties of cross-linked banana resistantstarch. Effect of pressure cooking. Starch - Starke 60, 286–291.
Asp, N.G. (1992). Resistant starch. Proceedings from the Second Plenary Meeting ofEURESTA: European FLAIR Concerted Action No. 11 on Physiological Implications ofthe Consumption of Resistant Starch in Man. Preface. European Journal of ClinicalNutritionSuppl. V 2, S1.
Azemi, B.M.N.M.,Wooton, M. (1984). In vitro digestibility of hydroxypropyl maize, waxymaize and high-amylose maize starches. Starch - Starke 34, 273–275.
Azemi, B.M.N.M., Wooton, M. (1995). Distribution of partial digestion products ofhydroxypropyl derivatives of maize (NM), waxy maize (WM), and high amylose(HA) maize starches. Starch - Starke 47, 465–469.
Baghurst, P.A., Baghurst, K.I., and Record, S.J. (1996). Dietary fiber, non-starch poly-saccharides and resistant starch – A review. Food Australia 48, S3–S35.
Bajka, B.H., Topping, D.L., Cobiac, L., Clarke, J.M. (2006). Butyrylated starch is lesssusceptible to enzymic hydrolysis and increases large bowel butyrate more than highamylose maize starch in the rat. British Journal of Nutrition 96, 276–282.
Bajka, B.H., Topping, D.L., Cobiac, L., Clarke, J.M. (2007). Butyrylated resistant starchesand the influence of cooking on amylolysis in vitro and short chain fatty acids in vivo. In:Salovaara, H., Gates, F., Tenkanen, M. (eds).Dietary Fiber: Components and Functions,171–185. Wageningen Academic Publishers: Wageningen, The Netherlands.
Bajka, B.H., Clarke, J.M., Cobiac, L., Topping, D.L. (2008). Butyrylated starch protectscolonocyte DNA against dietary protein-induced damage in rats. Carcinogenesis 29,2169–2174.
Bajka, B.H., Clarke, J.M., Topping, D.L., Cobiac, L., Abeywardana, M.Y., Mahinda, Y.,Patten, G.S. (2010). Butyrylated starch increase large bowel butyrate and lowers colonicsmooth muscle contractility in rats. Nutrition Research 104, 427–434.
Bhattacharya, D., Singhal, R.S., Kulkarni, P.R. (1995). A comparative account of condi-tions for synthesis of sodium carboxymethyl starch from corn and amaranth starch.Carbohydrate Polymers 27, 247–253.
Biliaderis, C.G. (1982). Physical characteristics, enzymatic digestibility, and structure ofchemically modified smooth pea and waxy maize starches. Journal of Agricultural andFood Chemistry 30, 925–930.
Bird, A.R., Lopez-Rubio, A., Shrestha, A.K., Gidley, M. (2009). Resistant starch in vitroand in vivo: Factors determining yield, structure, and physiological relevance. In:
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 69

3GCH03 07/19/2013 17:21:15 Page 70
Kasapis, S., Norton, I.T., Ubbink, J.B. (eds). Modern Biopolymer Science: Bridging theDivide Between Fundamental Treatise and Industrial Application, 449–510. Acad.Press: San Diego, CA.
Bjorck, I., Gunnarsson, A., Ostergard, K. (1989). A study of native and chemicallymodified potato starch. Part II. Digestibility in the rat intestinal tract. Starch - Starke4, 128–134.
Borchers, R. (1962). A note on the digestibility of the starch of high-amylose corn by rats.Cereal Chemistry 39, 145–146.
Brown, S.A., French, D. (1977). Specific adsorption of starch oligosaccharides in the gelphase of starch granules. Carbohydrate Research 59, 203–212.
Brown, I.L., McNaught, K.J., Maloney, E. (1995). Hi-MaizeTM: New directions in starchtechnology and nutrition. Food Australia 47, 272–275.
Brown, I.L., Yotsuzuka, M., Birkett, A., Henriksson, A. (2006). Prebiotics, synbiotics andresistant starch. Journal of Japanese Association for Dietary Fiber Research 10, 1–9.
Burkitt, D.P. (1971). Epidemiology of cancer of the colon and rectum. Cancer 62,1713–1724.
Burkitt, D.P. (1973a). Epidemiology of large bowel disease: The role of fiber. Proceedingsof the Nutrition Society 32, 145–149.
Burkitt, D.P. (1973b). Some disease characteristics of western civilization. British MedicalJournal 2, 274–276.
Bustos, M.C., Perez, G.T., Leon, A.E. (2011). Effect of four types of dietary fiber on thetechnological quality of pasta. Food Science and Technology International 17, 213–221.
Buttriss, J.L., Stokes, C.S. (2008). Dietary fiber and health: An overview.Nutrition Bulletin33, 186–200.
Chi, H., Xu, K., Wu, X., Chen, Q., Xue, D., Song, C., Zhang, W., Wang, P. (2008). Effect ofacetylation on properties of corn starch. Food Chemistry 106, 923–928.
Choi, H.J., Lee, C.J., Cho, E.J., Choi, S.J., Moon, T.W. (2010). Preparation, digestibility,and glucose response in mice of rice coated with resistant starch type 4 usinglocust bean gum and agar. International Journal of Food Science & Technology45, 2612–2621.
Chung, H.J., Woo, K.S., Lim, S.-T. (2004). Glass transition and enthalpy relaxation ofcross-linked corn starches. Carbohydrate Polymers 55, 9–15.
Clarke, J.M., Bird, A.R., Topping, D.L., Cobiac, L. (2007). Excretion of starch andesterified short-chain fatty acids by ileostomy subjects after the ingestion of acylatedstarches. American Journal of Clinical Nutrition 86, 1146–1151.
Clarke, J.M., Topping, D.L., Bird, A.R., Young, P.Y., Cobiac, L. (2008). Effects of high-amylose maize starch and butyrylated high-amylose maize starch on azoxymethane-induced intestinal cancer in rats. Carcinogenesis 29, 2190–2194.
Clarke, J.M., Topping, D.L., Christophersen, C.T., Bird, A.R., Lange, K., Saunders, I.,Cobiac, L. (2011a). Butyrate esterified to starch is released in the human gastrointestinaltract. American Journal of Clinical Nutrition 94, 1276–1283.
Clarke, J.M., Young, G.P., Topping, D.L., Bird, A.R., Cobiac, L., Scherer, B.L., Winkler, J.G., Lockett, T.J. (2011b). Butyrate delivered by butyrylated starch increases distalcolonic epithelial apoptosis in carcinogen-treated rats. Carcinogenesis doi, 10.1093/carcin/bgr254, pp. 1–6.
Code of Federal Regulations . (2010). Food-starch modified. In: Food Additives Permittedin Foods for Human Consumption, Title 21, Chapter 1, Part 172, Section 172.892. U.S.Government Printing Office, Washington.
70 Resistant Starch

3GCH03 07/19/2013 17:21:15 Page 71
Codex Alimentarius Commission . (2009). Joint FAO/WHO Food Standards Programme,Thirty second session, ALINORM 09/32/26, Rome, Italy. June 29–July 4.
Conlon, M.A., Kerr, C.A., McSweeney, C.S., Dunne, R.A., Shaw, J.M., Kang, S., Bird, A.R., Morell, M.K., Lockett, T.J., Molloy, P.L., Regina, A., Toden, S., Clarke, J.M.,Topping, D.L. (2012). Resistant starches protect against colonic DNA damage andalter microbiota and gene expression in rats fed a western diet. Journal of Nutrition 142,832–840.
Conway, R.L., Hood, L.F. (1976). Pancreatic a-amylase hydrolysis product of modifiedand unmodified tapioca starches. Starch - Starke 28, 341–343.
Daniels, A.L., Strickler, L. (1917). A comparison of the digestibility of starch in typicalbatter and dough mixtures. Journal of Home Economics 9, 109–114.
Eerlingen, R.C., Delcour, J.A. (1995). Formation, analysis, structure, and properties of typeIII enzyme resistant starch. Journal of Cereal Science 22, 129–138.
Eerlingen, R.C., Crombez, M., Delcour, J.A. (1993). Enzyme resistant starch. I. Quantita-tive and qualitative influence of incubation time and temperature of autoclaved starch onresistant starch formation. Cereal Chemistry 70, 339–344.
Eliasson, A.-C. (1998). Lipid-carbohydrate interactions. In: Hamer, R.J., Hoseney, R.C.(eds). Interactions: The Keys to Cereal Quality, 47–79. American Association of CerealChemists, St. Paul, MN.
Englyst, H.N. (1985). Dietary polysaccharide breakdown in the gut of man. PhD thesis,University of Cambridge, Cambridge, UK.
Englyst, H.N., Macfarlane, G.T. (1986). Breakdown of resistant starch and readilydigestible starch by human gut bacteria. Journal of the Science of Food and Agriculture37, 699–706.
Englyst, H.N., Cummings, J.H. (1987). Resistant starch, a ‘new’ food component: Aclassification of starch for nutritional purposes. In: Morin, I.D. (ed). Cereals in aEuropean Context, 221–233. Ellis Horwood: Chichester.
Englyst, H.N., Wiggins, H.S., Cummings, J.H. (1982). Determination of the non-starchpolysaccharides in plant foods by gas-liquid chromatography of constituent sugars asalditol acetates. Analyst 107, 307–318.
Englyst, H.N., Kingman, S.M., Cummings, J.H. (1992). Classification and measurementof nutritionally important starch fractions. European Journal of Clinical Nutrition 46,S33–S50.
Englyst, H.N., Kingman, S.M., Hudson, G.J., Cummings, J.H. (1996). Measurement ofresistant starch in vivo and in vitro. British Journal of Nutrition 75, 749–755.
Evans, I.D. (1986). An investigation of starch/surfactant interactions using viscometry anddifferential scanning calorimetry. Starch - Starke 38, 227–235.
Erickson, W. (2005). The baker’s toolbox for developing health-promoting products.Cereal Foods World 50, 6–8.
European Food Safety Authority (2007). Statement of the scientific panel on dieteticproducts, nutrition and allergies on a request from the commission related to dietaryfiber (Request No. EFSA-Q-2007-121). 17th Plenary Meeting Corresponding to Item10.1 of the Agenda, July 6.
Faergemand, M., Krog, N. (2006). Interactions of emulsifiers with other components infoods. In: Gaonkar, A.G., McPherson, A. (eds). Ingredients Interactions: Effects onFood Quality, Second Edition, 389–421, CRC Press: Boca Raton, FL.
Fastinger, N.D., Karr-Lilienthal, L.K., Spears, J.K., Swanson, K.S., Zinn, K.E., Nava, G.M., Ohkuma, K., Kanahori, S., Gordon, D.T., Fahey, Jr., G.C. (2008). A novel resistant
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 71

3GCH03 07/19/2013 17:21:15 Page 72
maltodextrin alters gastrointestinal tolerance factors, fecal characteristics, and fecalmicrobiota in healthy adult humans. Journal of the American College of Nutrition 27,356–366.
Felton, G.E., Schopmeyer, H.H. (1943). Thick-bodied starch and method of making. U.S.Patent 2,328,537.
Flickinger, E.A., Wolf, B.W., Garleb, K.A., Fahey Jr., G.C. (1998). In vitro and in vivodigestibility of selected oligosaccharides. The FASEB Journal 12, A210.
Fofanow, L. (1911). Digestion and absorption of several varieties of raw starch in the normaland diseased gastrointestinal tract. Zeitschrift f€ur Klinische Medizin 71, 257–270.
Fujiwara,K.,Matsuoka,A.(1995). Improvementofglucose tolerancebylow-viscosity,water-soluble dietary fiber, indigestible dextrin. Japanese Journal of Nutrition 53, 361–368.
Fuwa, H. (1977). Journal of the Japanese Society of Starch Science 24, 128–140.Gordon, D.T. (2007). The effect of resistant maltodextrin on blood glucose, insulin and
triacylglyceride levels, and fat accumulation after meal feeding in humans. In:Salovaara, H.Gates, F., Tenkanen, M. (eds). Dietary Fiber: Components and Functions,305–322. Wageningen Academic Publishers: The Netherlands.
Hahn, D.E., Hood, L.F. (1980). Physical characteristics of a-amylase hydrolyzates fromunmodified and chemically-modified waxy maize starches. Journal of Food Science 45,518–522.
Han, J.-A., Gray, J.A., Huber, K.C., BeMiller, J.N. (2006). Derivatization of starch granulesas influenced by the presence of channels and reaction conditions. In: Fishman, M.L., Qi,P.X., Wicker, L. (eds). Advances in Biopolymers, Molecules, Clusters, Networks andInteractions, 165–184. Am. Chem. Soc: Washington, DC.
Han, J.-A., Lee, M.-J., Lim, S.-T. (2007). Utilization of oxidized and cross-linked cornstarches in wheat flour batter. Cereal Chemistry 84, 582–586.
Haub, M.D., Hubach, K.L., Al-Tamimi, E.K., Ornelas, S., Seib, P.A. (2010). Different typesof resistant starch elicit different glucose responses in humans. Journal of Nutrition andMetabolism 2010: Article ID 230501.
Haub, M., Louk, J.A., Lopez, T.C. (2012). Novel resistant potato starches on glycemia andsatiety in humans. Journal of Nutrition and Metabolism doi: 10.1155/2012/478043,Article ID 478043, 4 pp.
He, J., Liu, J., Zhang, G. (2008). Slowly digestible waxy maize starch prepared by octenylsuccinic anhydride esterification and heat-moisture treatment: Glycemic response andmechanism. Biomacromolecules 9, 175–184.
Health Canada (2012). Policy for labeling and advertising of dietary fiber-containing foodproducts. Bureau of Nutritional Sciences, Food Directorate, Health Products and FoodBranch, 18 pp.
Hipsley, E.H. (1953). Dietary ‘fiber’ and pregnancy toxaemia. British Medical Journal2, 420.
Hood, L.F., Arneson, V.G. (1976). In vitro digestibility of hydroxypropyl distarchphosphate and unmodified tapioca starch. Cereal Chemistry 53, 282–290.
Hood, L.F., Mercier, C. (1978). Molecular structure of unmodified and chemicallymodified manioc starches. Carbohydrate Research 247, 279–290.
Hoover, R., Sosulski, F. (1986). Effect of cross-linking on functional properties of legumestarches. Starch - Starke 38, 149–155.
Huang, J.R., Schols, H.A., Jin, Z.Y., Sulmann, E., Voragen, A.G.J. (2007). Pastingproperties and (chemical) fine structure of acetylated yellow pea starch is affectedby acetylation reagent type and granule size. Carbohydrate Polymers 68, 397–406.
72 Resistant Starch

3GCH03 07/19/2013 17:21:15 Page 73
Huber, K.C., BeMiller, J.N. (2000). Channels of maize and sorghum starch granules.Carbohydrate Polymers 41, 269–276.
Huber, K.C., BeMiller, J.N. (2001). Location of sites of reactions within starch granules.Cereal Chemistry 78, 173–180.
Huber, K.C., BeMiller, J.N. (2010). Modified starch chemistry and properties. In: Bertolini,A.C. (ed). Starches: Characterization, Properties and Applications, 145–203. CRCPress, Boca Raton, FL.
Institute of Medicine . (2001). Dietary reference intakes: Proposed definition of dietaryfiber. National Academy Press, Washington, DC.
Jacobs, H., Eerlingen, R.C., Clauwaert, W., Delcour, J.A. (1995). Influence of annealing onthe pasting properties of starches from various botanical sources. Cereal Chemistry 72,480–487.
Jane, J., Xu, A., Radosavljevic, M., Seib, P.A. (1992). Location of amylose in normal starchgranules. I. Susceptibility of amylose and amylopectin to cross-linking reagents. CerealChemistry 69, 405–409.
Jane, J.-L., Robyt, J.F. (1984). Structure studies of amylose-V complexes and retrogradedamylose by action of alpha amylases, and a new method for preparing amylodextrins.Carbohydrate Research 132, 105–118.
Janzen, G.J. (1969). Digestibility of starches and phosphatised starches by means ofpancreatin. Starch - Starke 21, 231–237.
Jenkins,D.J.,Wolever,T.M.,Taylor,R.H.,Barker,H.,Fielden,H.,Baldwin, J.M.,Bowling,A.C.,Newman,H.C.,Jenkins,A.L.,Goff,D.V. (1981).Glycemicindexoffoods:Aphysiologi-cal basis for carbohydrate exchange.American Journal of Clinical Nutrition 34, 362–366.
Kasemsuwan, T., Jane. J. (1994). Location of amylose in normal starch granules. II.Location of phosphodiester cross-linking revealed by Phosphorus-31 nuclear magneticresonance. Cereal Chemistry 71, 282–287.
Katz, J.R. (1934). Abhandlungen zur physikalischen Chemie der Starke und der Brotber-eitung. XX. Uber den Zusammenhang der Anderung der Starke beim Altbackenwerdendes Brotes und beim Retrogradieren von Starkekleister. Zeitschrift f€ur PhysikalischeChemie Abt. A. 169, 321.
Kerr, R.W., Cleveland, F.C. (1957). Process for the preparation of distarch phosphate andthe resulting product. U.S. Patent 2,801,242.
Kim, H.-S., Huber, K.C. (2008). Channels within soft wheat starch A- and B-type granules.Journal of Cereal Science 48, 159–172.
Kim, M.J., Choi, S.J., Shin, S.I., Sohn, M.R., Lee, C.J., Kim, Y., Cho, W.I., Moon, T.W.(2008). Resistant glutarate starch from adlay: Preparation and properties. CarbohydratePolymers 74, 787–796.
Klaushofer, H., Berghofer, E., Steyrer, W. (1978). Starch citrates – Production andtechnical application properties. Starch - Starke 30, 47–51.
Koch, V.H., Brommer, H.D., Koppens, J. (1982). Analytical investigations on phosphatecross-linked starches. Starch - Starke 34, 16–21.
Krog, N. (1971). Amylose complexing effect of food grade emulsifiers. Starch - Starke 23,206.
Lammers, G., Stamhuis, E.J., Beenackers, A.A.C.M. (1993). Kinetics of the hydroxypro-pylation of potato starch in aqueous solution. Industrial & Engineering ChemistryResearch 32, 835–842.
Langworthy, C.F., Deuel, H.J., Jr., (1920). Digestibility of raw corn, potato and wheatstarches. Journal of Biological Chemistry 42, 27–40.
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 73

3GCH03 07/19/2013 17:21:15 Page 74
Langworthy,C.F.,Deuel,H.J., Jr., (1922).Digestibility of raw rice, arrowroot, canna, cassava,taro, tree-fern and potato starches. Journal of Biological Chemistry 52, 251–261.
Laurentin, A., Cardenas, M., Ruales, J., Perez, E., Tovar, J. (2003). Preparation ofindigestible pyrodextrins from different starch sources. Journal of Agricultural andFood Chemistry 51, 5510–5515.
Leach, H.W., Schoch, T.J. (1961). Structure of the starch granule II. Action of variousamylases on granular starches. Cereal Chemistry 38, 34–46.
Lee, K.Y., Yoo, S.-H., Lee, H.G. (2012). The effect of chemically-modified resistant starch,RS type-4, on body weight and blood lipid profiles of high fat diet-induced obese mice.Starch - Starke 64, 78–85.
Leegwater, D.C., Luten, J.B. (1971). A study on the in vitro digestibility of hydroxypropylstarches by pancreatin. Starch - Starke 23, 430–432.
Lefranc-Millot, C., Wils, D., Henry, J., Lightower, H., Saniez-Degrave, M.H. (2006).Nutriose, a resistant dextrin and Maltisorb, a sugar alcohol, two key ingredients forhealthy diets and obesity management. Obesity Reviews 7, 269.
Lefranc-Millot, C., Wils, D., Deremaux, L., Macioce, V., Saniez-Degrave, M.-H. (2010).Nutriose, more than just a soluble fiber. In: van der Kamp, J.W., Jones, J., McCleary, B.,Topping, D. (eds). Dietary Fiber: New Frontiers for Food and Health, 295–308.Wageningen Academic Publishers, Wageningen, The Netherlands.
Lopez-Rubio, A., Clarke, J.M., Scherer, B., Topping, D.L., Gilbert, E.P. (2009). Structuralmodifications of granular starch upon acylation with short-chain fatty acids. FoodHydrocolloids 23, 1940–1946.
Manelius, R., Buleon, A., Nurmi, K., Bertoft, E. (2000). The substitution pattern incationized and oxidized potato starch granules. Carbohydrate Research 329, 621–633.
Maningat, C., Bassi, S., Woo, K., Dohl, C., Gaul, J., Stempien, G., Moore, T. (2005).Formulation of high-protein, high-fiber (low-carbohydrate), reduced calorie breads. AIBResearch Department Technical Bulletin 27 (4), 1–16.
Maningat, C.C., Bassi, S.D., Ranjan, S., Gaul, J.A., Stempien, G.J., Dohl, C.T., Woo, K.(2008). High-protein, reduced-carbohydrate bakery and other food products. Austra-lian Patent No. 2004253169.
Martinez, I., Kim, J., Duffy, P.R., Schlegel, V.L., Walter, J. (2010). Resistant starches types2 and 4 have differential effects on the composition of the fecal microbiota in humansubjects. PLoS ONE 5 (11): e15046. doi: 10.1371/journal.pone.0015046.
Mason, W.R. (2009). Starch use in foods. in: BeMiller, J., Whistler, R. (eds). Starch:Chemistry and Technology, Third Edition, 747–795. Academic Press, New York.
McCleary, B.V., McNally, M., Rossiter, P. (2002). Measurement of resistant starch byenzymic digestion in starch samples and selected plant materials: Collaborative study.Journal of AOAC International 85, 1103–1111.
McCleary, B.V., DeVries, J.W., Rader, J.I., Cohen, G., Prosky, L., Mugford, D.C., Champ,M., Okuma, K. (2010). Determination of total dietary fiber (Codex definition) byenzymatic-gravimetric method and liquid chromatography: Collaborative study. Journalof AOAC International 93, 221–233.
McCleary, B.V., DeVries, J.W., Rader, J.I., Cohen, G., Prosky, L., Mugford, D.C., Champ,M., Okuma, K. (2011). Collaborative study report: Determination of insoluble, soluble,and total dietary fiber (Codex definition) by an enzymatic-gravimetric method and liquidchromatography. Cereal Foods World 56, 238–247.
Miller, R.A., Jeong, J., Maningat, C.C. (2011). Effect of RS4 resistant starch on extrudedReady-to-Eat (RTE) breakfast cereal quality. Cereal Chemistry 88, 584–588.
74 Resistant Starch

3GCH03 07/19/2013 17:21:15 Page 75
Nomura, M., Nakajima, Y., Abe, H. (1992). Effect of long-term administration ofindigestible dextrin as soluble dietary fiber on lipid and glucose metabolism. Journalof Japanese Society of Nutrition and Food Science 35, 21–25.
Nugent, A.P. (2005). Health properties of resistant starch. Nutrition Bulletin 30, 27–54.Ogden, C.L., Carroll, M.D., Kit, B.K., Flegal, K.M. (2012). Prevalence of obesity in theUnited States, 2009–2010. NCHS Data Brief, No. 82. National Center for HealthStatistics, Hyattsville, MD.
Ohkuma, K., Wakabayashi, S. (2001). Fibersol-2: A soluble, non-digestible, starch-deriveddietary fiber. In: McCleary, B.V., Prosky, L. (eds). Advanced Dietary Fiber Technology,509–523. Blackwell Science Ltd: Malden, MA.
Ohkuma, K., Hanno, Y., Inada, K., Matsuda, I., Katta, Y. (1994). Indigestible dextrin. U.S.Patent 5,364,652.
Ohkuma, K., Matsuda, I., Hanno, Y. (1995).Method for preparing low calorie dextrin. U.S.Patent 5,430,141.
Ohkuma, K., Hanno, Y., Inaba, K., Matsuda, I., Katsuda, Y. (1997). Process for preparingdextrin containing food fiber. U.S. Patent 5,620,873.
Okuma, K., Kishimoto, Y. (2004). Effect of resistant maltodextrin on metabolism ofglucose and lipids. In: Van Der Kamp, J.W., Asp, N.-G., Jones, J.M., Schaafsma, G.(eds). Dietary Fiber: Bioactive Carbohydrates for Food and Feed, 219–230. Wage-ningen Academic Publishers: The Netherlands.
Ostergard, K., Bjorck, I., Gunnarsson, A. (1988). A study of native and chemically modifiedpotato starch. I. Analysis and enzymatic availability in vitro. Starch - Starke 40, 58–66.
Ratnayake, W.S., Jackson, D.S. (2008). Thermal behavior of resistant starches RS2, RS3,and RS4. Journal of Food Science 73, C356–C366.
Sandstedt, R.M., Strahan, D., Ueda, S., Abbott, R.C. (1962). The digestibility of high-amylose corn starches. Cereal Chemistry 39, 123–131.
Sang, Y., Prakash, O., Seib, P.A. (2007). Characterization of phosphorylated cross-linkedresistant starch by 31P nuclear magnetic resonance (31P NMR) spectroscopy. Carbohy-drate Polymers 67, 201–212.
Sang, Y., Seib, P.A., Herrera, A.I., Prakash, O., Shi, Y.-C. (2010). Effects of alkalinetreatment on the structure of phosphorylated wheat starch and its digestibility. FoodChemistry 118, 323–327.
Satouchi, M., Wakabayashi, S., Ohkuma, K., Fujiwara, K., Matsuoka, A. (1993). Effects ofindigestible dextrin on bowel movements. Japanese Journal of Nutrition 51, 31–37.
Seib, P.A., Woo, K. (1999). Food-grade starch resistant to alpha-amylase and method ofpreparing the same. U.S. Patent 5,855,946.
Shin, M., Seib, P.A. (2004). Morphological change and RS level of RS mixture withcross-linked RS4 granule during in vitro digestion. Food Science and Biotechnology 13,544–548.
Shin, M., Woo, K., Seib, P. (2003). Hot water solubility and water sorption of resistantstarch at 25 �C. Cereal Chemistry 80, 564–566.
Shin, S.I., Lee, C.J., Kim, D.-I., Lee, H.A., Cheong, J.-J., Chung, K.M., Baik, M.-Y., Park,C.S., Kim, C.H., Moon, T.W. (2007). Formation, characterization, and glucose responsein mice to rice starch with low digestibility produced by citric acid treatment. Journal ofCereal Science 45, 24–33.
Silvester, K., Englyst, H.N., Cummings, J.H. (1995). Ileal recovery of starch from wholediets containing resistant starch measured in vitro and fermentation of ileal effluent.American Journal of Clinical Nutrition 62, 403–411.
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 75

3GCH03 07/19/2013 17:21:15 Page 76
Singh, V., Ali, S.Z., Divakar, S. (1993). 13C CP/MASNMR spectroscopy of native and acidmodified starches. Starch - Starke 45, 59–62.
Song, Y., Bae, C., Woo, D., Kim, Y., Park, T. (2010). Immunomodulatory activity of type-4resistant starch in the mesenteric lymph nodes of rats. Journal of Medicinal Food 13,205–210.
Thompson, L.U., Maningat, C.C., Woo, K., Seib, P.A. (2011). In vitro digestion of RS4-type resistant wheat and potato starches, and fermentation of indigestible fractions.Cereal Chemistry 88, 72–79.
Thorpe, E. (1913). A dictionary of applied chemistry. London, New York, Bombay, andCalcutta, v, 179.
Tokunaga, M., Matsuoka, A. (1999). Effects of a food for specified health use (FOSHU)which contains indigestible dextrin as an effective ingredient on glucose and lipidmetabolism. Journal of the Japan Diabetes Society 42, 61–65.
Tomasik, P., Palasinski, M., Wiejak, S. (1989). The thermal decomposition of carbohy-drates. Part II. The decomposition of starch. Advances in Carbohydrate Chemistry andBiochemistry 47, 270–335.
Topping, D. (2007). Cereal complex carbohydrates and their contribution to human health.Journal of Cereal Science 46, 220–229.
Topping, D.L., Clifton, P.M. (2001). Short-chain fatty acids and human colonic function:Roles of resistant starch and nonstarch polysaccharides. Physiological Reviews 81,1031–1064.
Trowell, H. (1972). Crude fiber, dietary fiber and atherosclerosis. Atherosclerosis 16,138–140.
Van Patten, E.M., Powell, E.L. (1969). Process for the manufacture of cross-linkedgranular starch products. U.S. Patent 3,422,089.
Vermorel, M., Condray, Y.C., Wils, D., Sinaud, S., Tressol, J.C., Montaurier, C., Vernet, J.,Brandolini, M., Bouteloup-Demange, C., Rayssiguier, Y. (2004). Energy value of a low-digestible carbohydrate, Nutriose FB, and its impact on magnesium, calcium and zincapparent absorption and retention in healthy young men. European Journal of Nutrition43, 344–352.
Wakabayashi, S., Ueda, Y., Matsuoka, A. (1992). Effects of indigestible dextrin on sugartolerance. II. Effect of continuous administration in rats fed on a high sucrose diet.Journal of the Japan Diabetes Society 35, 873–880.
Wang, Y.-J., Kozlowski, R., Delgado, G.A. (2001). Enzyme resistant dextrins from highamylose corn mutant starches. Starch - Starke 53, 21–26.
Wepner, B., Berghofer, E., Miesenberger, E., Tiefenbacher, K., Ng, P.N.K. (1999). Citratestarch – application as resistant starch in different food systems. Starch - Starke 51,354–361.
Witwer, R.S. (2008). Natural resistant starch in glycemic management: From physiologicalmechanism to consumer communication. In: Pasupuleti V.K., Anderson, J.W. (eds).Nutraceuticals, Glycemic Health and Type 2 Diabetes, 401–438. Wiley-Blackwell:Ames, IA.
Wolf, B.W., Bauer, L.L., Fahey, G.C., Jr., (1999). Effects of chemical modification on invitro rate and extent of food starch digestion: An attempt to discover slowly digestedstarch. Journal of Agricultural and Food Chemistry 47, 4178–4183.
Wolf, B.W., Wolever, T.M.S., Bolognesi, C., Zinker, B.A., Garleb, K.A., Firkins, J.L.(2001). Glycemic response to a food starch esterified by 1-octenyl succinic anhydride inhumans. Journal of Agricultural and Food Chemistry 49, 2674–2678.
76 Resistant Starch

3GCH03 07/19/2013 17:21:15 Page 77
Woo, K.S., Seib, P.A. (2002). Cross-linked resistant starch: Preparation and properties.Cereal Chemistry 79, 819–825.
Woo, K., Bassi, S.D., Maningat, C.C., Zhao, L., Trompeter, E.T., KelleyJr., G.A., Ranjan,S., Gaul, J., Dohl, C.T., DeMerittJr., G.K., Stempien, G.J., Krehbiel, K.D. (2006).Pregelatinized chemically modified resistant starch products and uses thereof. PatentApplication Publication No. US 2006/0188631.
Woo, K.S., Maningat, C.C., Seib, P.A. (2009). Increasing dietary fiber in foods: The casefor phosphorylated cross-linked resistant starch, a highly concentrated form of dietaryfiber. Cereal Foods World 54 (5), 217–223.
Wooton, M., Chaudhry, M.A. (1979). Enzymic digestibility of modified starches. Starch -Starke 31, 224–228.
Wurzburg, O.B. (1986). Converted starches. In: Wurzburg, O.B. (ed). Modified Starches:Properties and Uses, 17–40, CRC Press: Boca Raton, FL.
Wurzburg, O.B., Vogel, W.F. (1984). Modified starch – Safety and regulatory aspects. In:Phillips, G.O., Wedlock, D.J., Williams, P.A. (eds). Gums and Stabilizers for the FoodIndustry, 406–415. Pergamon Press: Oxford, UK.
Xie, X., Liu, Q. (2004). Development and physiochemical characterization of new resistantcitrate starch from different corn starches. Starch - Starke 56, 364–370.
Xie, X., Liu, Q., Cui, S.W. (2006). Studies on the granular structure of resistant starch (type4) from normal, high amylose and waxy corn starch citrates. Food Research Interna-tional 39, 332–341.
Yeo, L.L., Seib, P.A. (2009). White pan bread and sugar snap cookies containing wheatstarch phosphate, a cross-linked resistant starch. Cereal Chemistry 86, 210–220.
RS4-Type Resistant Starch: Chemistry, Functionality and Health Benefits 77

3GCH03 07/19/2013 17:21:15 Page 78

CH04GR 07/29/2013 17:14:45 Page 79
4 Novel Applications of Amylose-LipidComplex as Resistant Starch Type 5
Jovin Hasjim1, Yongfeng Ai2 and Jay-lin Jane2
1Queensland Alliance for Agriculture and Food Innovation, Centre forNutrition and Food Sciences, The University of Queensland, Australia
2Department of Food Science and Human Nutrition, Iowa State University,USA
4.1 INTRODUCTION
Amylose is known to form single helical complexes with many other sub-
stances, including iodine, monoglycerides, lysophospholipids, fatty acids and
alcohols. Amylose-lipid complex is commonly found in native starch granules
and processed starch (Becker et al., 2001; Morrison et al., 1993, 1984). The
hydrocarbon chain of the lipid interacts with the hydrophobic moiety of the
amylose chain and fills the central cavity of the amylose single helix (Jane &
Robyt, 1984; Jane et al., 1985; Morrison et al., 1993). The number of glucose
units per helical turn ranges from six to eight, depending on the size of the cross-
section of the complexing agent (Jane & Robyt, 1984; Shogren et al., 2006).
Amylopectin can also form a single helical complex with lipids. Cold-
water soluble starch prepared by heating normal maize starch in aqueous
alcohol solution shows the V-type X-ray diffraction pattern associated with
starch-lipid complex (Jane et al., 1986a, 1986b). The degree of crystallinity of
the cold-water soluble maize starch is higher than its amylose content, which
implies that amylopectin is involved in the complex formation with alcohols.
The presence of amylopectin-lipid complex also restricts the reassociation of
amylopectin branches into double helices, as observed in the decrease of the
degree of starch retrogradation during storage (Eliasson & Ljunger, 1988;
Huang & White, 1993).
Amylose-lipid complex is resistant to amylolytic enzyme hydrolysis
(Holm et al., 1983; Jane & Robyt, 1984; Kitahara et al., 1996; Seneviratne
& Biliaderis, 1991). It has been proposed as a new source of resistant starch
79
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

CH04GR 07/29/2013 17:14:45 Page 80
(RS, type 5) in addition to the four earlier proposed sources (i.e. physically
inaccessible starch (RS type 1), native B- and some C-type polymorphic starch
granules (RS type 2), retrograded amylose (RS type 3) and chemicallymodified
starch (RS type 4)) (Eerlingen & Delcour, 1995; Englyst et al., 1992).
Amylose-lipid complex has been shown to reduce postprandial glycemic
and insulinemic responses (Hasjim et al., 2010; Holm et al., 1983; Takase
et al., 1994). Removal of lipids from native high-amylose maize starch (RS
type 2) also results in a decrease in the enzyme resistance of the starch (Jiang
et al., 2010). The advantages of amylose-lipid complex as RS comparing with
other types of RS are that:
1. amylose-lipid complex is more heat stable than most native B- and C-type
polymorphic starch granules (RS type 2) such as potato and banana starches;
2. the production of amylose-lipid complex requires less extensive physical
and/or chemical processing than that of retrograded amylose (RS type 3)
and chemically modified starch (RS type 4);
3. amylose-lipid complex restores its complex structure spontaneously
during cooling after being heated above its dissociation temperature
(Biliaderis & Galloway, 1989; Hasjim et al., 2010).
4.2 ENZYME DIGESTIBILITY OF AMYLOSE-LIPIDCOMPLEX
Amylose-lipid complex is more resistant to amylolytic enzyme hydrolysis
than amorphous amylose and most A-type polymorphic starches. The enzyme
resistance of amylose-lipid complex depends on the molecular structures of
the complexing lipid and the amylose (Gelders et al., 2005; Kitahara et al.,
1996) and on the crystalline structure of the amylose-lipid complex
(Seneviratne & Biliaderis, 1991). In starch granules, amylose-lipid complex
has been shown to retard granule swelling during heating in excess water,
which reduces the enzyme accessibility to hydrolyze the starch granules
(Cui & Oates, 1999; Tester & Morrison, 1990).
In a recent study, cooking of tapioca, normal maize and high-amylose
maize starches in the presence of corn oil, soy lecithin, palmitic acid (PA),
stearic acid (SA), oleic acid (OA) or linoleic acid (LA) was shown to reduce
the susceptibility of the starch to enzyme hydrolysis (Ai et al., 2013). The
same phenomenon, however, was not observed with waxy maize starch. The
results indicate that the amylose-lipid complex formed during the cooking of
non-waxy starch and lipid was responsible for the decrease in the enzyme
digestibility of starch. The presence of amylose-lipid complex was later
80 Resistant Starch

CH04GR 07/29/2013 17:14:45 Page 81
confirmed from the endothermic peak of the dissociation of amylose-lipid
complex by differential scanning calorimetry (DSC) for starch samples
cooked with soy lecithin, PA, SA, OA, and LA. The amylose-lipid complex
in the cooked starch-corn oil mixture was confirmed by 13C nuclear magnetic
resonance (NMR) spectroscopy.
4.2.1 Effects of lipid structure on the enzyme resistanceof amylose-lipid complex
The properties of amylose-lipid complex, including the dissociation temper-
ature and the susceptibility to enzyme hydrolysis, are affected by the
molecular structure of the complexing lipid. In general, the dissociation
temperature of amylose-lipid complex increases with the hydrocarbon-chain
length of the fatty acid (FA) (Biliaderis & Galloway, 1989; Raphaelides &
Karkalas, 1988; Tufvesson et al., 2003), which reflects the stability of the
amylose-lipid complex, i.e. the hydrophobic interaction between amylose and
the hydrophobic chain of the FA. Short-chain and some medium-chain FAs
(up to 10 carbon units) are not as effective complexing agents as long-chain
FAs (12 carbon units and above). This is due to the greater water solubility of
the short- and medium-chain FAs, allowing the FAs to participate in the
aqueous solution instead of complexing with amylose (Tufvesson et al., 2003;
Yotsawimonwat et al., 2008).
The dissociation temperature of amylose-lipid complex also decreases with
the increase in the number of cis double bonds in the FA chain of complexing
lipid (Karkalas et al., 1995; Raphaelides & Karkalas, 1988; Tufvesson et al.,
2003). The kink structure of cis double bond disrupts the alignment of the FA
in the cavity of amylose single helix. The dissociation temperature of amylose
complexed with an unsaturated FA (e.g. OA and LA) is lower than that of
amylose complexed with a saturated FA of the same hydrocarbon chain-length
(e.g. SA; Tufvesson et al., 2003). Consequently, a saturated long-chain FA
produces a more enzyme-resistant amylose complex than a shorter or an
unsaturated FA (Kitahara et al., 1996).
The low water solubility and the high melting temperature of long-chain
FAs, however, reduce the dispersion of these FAs in water and reduces
their availability in the aqueous solution to form complex with amylose
(Yotsawimonwat et al., 2008). The effectiveness of the complex formation
between amylose and long-chain FA can be improved by increasing the
solubility of the long-chain FA at the pH above the pKa of the FA. Long-
chain FAs are more soluble in aqueous solution when they are in salt form,
carrying a negative charge (Tufvesson et al., 2003; Yotsawimonwat et al.,
2008).
Novel Applications of Amylose-Lipid Complex as Resistant Starch Type 5 81

CH04GR 07/29/2013 17:14:45 Page 82
4.2.2 Effects of crystalline structure on the enzymeresistance of amylose-lipid complex
On the basis of crystallinity, there are two forms of amylose-lipid complex.
The amorphous (Form I) complex is produced by mixing amylose and lipid at
a lower temperature (25–60 �C), whereas the crystalline (Form II) complex
is produced at a higher temperature (90–100 �C) (Gelders et al., 2005;
Seneviratne & Biliaderis, 1991; Tufvesson et al., 2003). Consequently, the
amorphous complex has a lower dissociation temperature (<100 �C) than
does the crystalline counterpart (>100 �C). Amorphous amylose-lipid com-
plex can be converted to crystalline complex through isothermal annealing at a
temperature above the dissociation temperature of the amorphous complex
and below that of the crystalline complex.
The enzyme resistance of amylose-lipid complex can be attributed to
the collapsed helical conformation of the starch-lipid complex, preventing
the dispersion of amylose molecules and interfering with the binding of
amylose molecules by enzyme for hydrolysis (Jane & Robyt, 1984).
In addition, the crystalline structure of amylose helical complex protects
the bulk of the amylose in the crystallites and enhances the enzyme resistance
of the amylose-lipid complex. Hence, the crystalline amylose-lipid complex
(Form II) is more resistant to amylolytic enzyme hydrolysis than the amor-
phous complex counterpart (Form I) (Gelders et al., 2005; Seneviratne &
Biliaderis, 1991).
4.2.3 Effects of amylose-lipid complex on the enzymeresistance of granular starch
Amylose-lipid complex is commonly found in native cereal starch granules
(Morrison et al., 1993, 1984). The native lipids in cereal starch granules,
including maize, rice, barley, oat, millet, and sorghum, are mostly free FAs
and lysophospholipids, and the content of lipids in cereal starch granules is
positively correlated with the amylose content of the starch (Morrison et al.,
1984). These lipids are found on the surface as well as in the inner part of
starch granules (Morrison, 1981). Amylose-lipid complex of granular starch
usually shows a dissociation temperature ranging between 70–108 �C, asanalyzed using DSC in the presence of excess water (Kugimiya et al., 1980; Li
et al., 2008).
The physicochemical properties of native starch granules are greatly
affected by the presence of amylose-lipid complex, including the decreases
in granule swelling, molecular leaching and enzyme digestibility. The forma-
tion of amylose-lipid complex with a helical conformation enhances the
82 Resistant Starch

CH04GR 07/29/2013 17:14:45 Page 83
entanglements between amylose and amylopectin molecules, restricting
amylose leaching and retarding the swelling of starch granules during heating
in excess water (Becker et al., 2001; Cui & Oates, 1999; Tester & Morrison,
1990). Consequently, it reduces the accessibility of enzyme to hydrolyze the
starch (Cui & Oates, 1999). Furthermore, a decrease in RS content was
observed when native high-amylose maize starch was defatted using methanol
solution (Jiang et al., 2010).
4.3 PRODUCTION OF RESISTANT GRANULAR STARCHTHROUGH STARCH-LIPID COMPLEX FORMATION
A technology of producing RS by increasing the amount of amylose-lipid
complex (RS type 5) in granular starch has been recently developed. The
process includes a treatment of swollen starch granules with a debranching
enzyme (isoamylase or pullulanase) to remove the branch linkages of
amylopectin. The resulting free long branch-chains of amylopectin can
function in a similar way to amylose molecules; thus, they can effectively
complex with FA. High-amylose maize starch VII (HA7) was chosen for this
technology because it consists of large proportions of amylose and long
branch-chains of amylopectin (Jane et al., 1999; Li et al., 2008).
A suspension of HA7 is pre-heated at an elevated temperature and is then
incubated with a debranching enzyme – either isoamylase (ISO) or pullula-
nase (PUL). The ISO- or PUL-treated-HA7 suspension is then mixed with FA
at a temperature above the melting temperature of the FAwith stirring for 30
minutes to allow the amylose-lipid complex formation. The complex is
recovered by centrifugation, and the excess FA is removed by washing
with 50% ethanol. FAs are effective complexing agents for making RS
and are economically favourable, because FAs are often the by-products of
edible oil refining process. The resulting HA7-FA complex products have RS
contents up to 75%, measured using the AOAC Method 991.43 for total
dietary fibres, in which the each complex product was hydrolyzed for
30 minutes with a thermally stable a-amylase in a boiling water bath
(Horwithz, 2003; see Table 4.1). The effects of debranching and complex
formation with FA on the RS contents are significant at p< 0.01.
4.3.1 Effects of fatty-acid structure on the RS content
The increase in the RS content of HA7 after complex formation with FA
(Table 4.1) is attributed to the increased amount of amylose-lipid complex (RS
type 5). PA and SA are preferred for the production of RS type 5 because of
Novel Applications of Amylose-Lipid Complex as Resistant Starch Type 5 83

CH04GR 07/29/2013 17:14:45 Page 84
their long hydrocarbon chains and straight molecular structures, which
form stable inclusion-complexes with amylose (Tufvesson et al., 2003;
Yotsawimonwat et al., 2008).
When short-chain FAs, such as sodium propionate and butyric acid (BA),
are used to form helical complexes with HA7, the products display lower RS
contents than those made with PA and SA (Table 4.1). The differences in the
RS contents are attributed to the greater water solubility of short-chain FAs,
preventing the formation of stable helical complexes with amylose. As a
result, the order of the RS contents of the complex products is: SA complex>PA complex > myristic acid (or MA) complex > BA complex.
Unsaturated FAwith cis double bond(s) (e.g. OA, which has a kink in the
hydrophobic chain), is less effective for amylose-lipid complex formation
(Karkalas & Raphaelides, 1986; Tufvesson et al., 2003) and produces a
complex product with a lower RS content (63.2%) than the complex product
of saturated FA with the same carbon number (SA, 71.6%) (Table 4.1).
The RS contents of pre-heated HA7 control and (PUL-treated HA7)-FA
complex products are substantially reduced after they are defatted using 85%
methanol solution in a Soxhlet extractor for 24 hours (Table 4.2). These results
support the fact that amylose-lipid complex contributes to the enzyme
Table 4.1 Resistant starch contents of high-amylose maize starch VII after pre-heating at80 �C, treatment with debranching enzyme and complexing with fatty acid at 80 �C.1
Treatments2 Resistant starch content (%)3
Pre-heated HA7 control 36.7�0.2HA7-SA complex 59.8�2.8HA7-PA complex 58.3�1.7ISO-treated HA7 57.8�0.1PUL-treated HA7 46.0�1.6(ISO-treated HA7)-SA complex 74.8�1.5(ISO-treated HA7)-PA complex 74.3�2.4(PUL-treated HA7)-SA complex 71.6�0.3(PUL-treated HA7)-PA complex 69.9�2.2(PUL-treated HA7)-MA complex 62.7�3.0(PUL-treated HA7)-BA complex 44.8�0.8(PUL-treated HA7)-NaPr complex 48.1�2.9(PUL-treated HA7)-OA complex 63.2�0.6
1Values are means� standard deviations.2HA7¼ high-amylose maize starch VII, SA¼ stearic acid, PA¼palmitic acid, ISO¼ isoamylase, PUL¼pullulanase, MA¼myristic acid, BA¼butyric acid, NaPr¼ sodium propionate, and OA¼oleic acid.3Resistant starch content was analyzed following the AOAC Method 991.43 for total dietary fibres
using thermally stable a-amylase in a boiling water bath.
84 Resistant Starch

CH04GR 07/29/2013 17:14:45 Page 85
resistance of the control HA7 starch and the (PUL-treated HA7)-FA complex
products. The RS contents of the defatted (PUL-treated HA7)-FA complex
products are, however, higher than those of the pre-heated HA7 control, with
andwithout defatting, which suggests that the FA in the (PUL-treatedHA7)-FA
complex products cannot be completely removed by the defatting process.
These results further support the complex formation between the PUL-
treated HA7 starch and FA because the FA in starch-lipid complex is less
extractable than uncomplexed FA (Karkalas & Raphaelides, 1986). In addi-
tion, the defatted (PUL-treated HA7)-SA complex product shows a substan-
tially higher RS content than the defatted (PUL-treated HA7)-PA complex
product, which could be attributed to the lower extractability of the longer-
chain FA (SA) in the starch-lipid complex than the shorter one (PA).
4.3.2 Effects of debranching on the RS content
The RS contents of the HA7 starch and the HA7-FA complex products
increase after the HA7 starch is debranched with ISO or PUL (Table 4.1).
Debranching enzyme (ISO or PUL) hydrolyzes the a-(1! 6) glycosidic
linkages and releases linear chains from amylopectin and also from amylose.
The free linear chains have greater mobility to associate between themselves
to form double helices, resulting in the increase of RS content as shown in
the ISO-treated and PUL-treated HA7 starch products (without complexing
with FA).
A combination of the debranching reaction and the complex formation
with FA further increases the RS content of HA7 starch; examples are
Table 4.2 Resistant starch contents of high-amylose maize starch VII-fatty acid complexproducts after defatting.1
Resistant starch content (%)3
Treatments2 Before defatting After defatting Difference
Pre-heated HA7 control 36.7�0.2 30.2�1.3 6.5(PUL-treated HA7)-PA complex 69.9�2.2 38.2�0.7 31.7(PUL-treated HA7)-SA complex 71.6�0.3 50.9�0.4 20.7
1Defatting was carried out using 85%methanol solution in a Soxhlet extractor for 24 hours. Values are
means� standard deviations.2HA7¼ high-amylose maize starch VII, PUL¼pullulanase, PA¼palmitic acid, and SA¼ stearic acid.
HA7 starch was pre-heated at 80 �C before treatments with PUL and fatty acids.3Resistant starch content was analyzed following the AOAC Method 991.43 for total dietary fibres,
using thermally stable a-amylase in a boiling water bath.
Novel Applications of Amylose-Lipid Complex as Resistant Starch Type 5 85

CH04GR 07/29/2013 17:14:45 Page 86
(ISO-treated HA7)-FA and (PUL-treated HA7)-FA complex products
(Table 4.1). These results show a synergistic effect of the two treatments,
in which the linear chains produced from the debranching reaction can
form complex more efficiently with FA than can the highly branched
amylopectin molecules.
The complex formation between FAs and relatively short linear dextrins
produced from the debranching of amylopectin in waxy rice starch has been
reported (Yotsawimonwat et al., 2008). Because most high-amylose maize
starches, including HA7 starch, have amylopectin with long branch-chains
(Jane et al., 1999; Li et al., 2008), some linear long chains released from the
debranching of HA7 amylopectin are expected to function in a similar fashion
to amylose. This is supported by the ability of the starch from double-mutant
ae-waxy maize (which consists of only amylopectin, with similar structure to
the HA7 amylopectin) to form complex with iodine and give a blue colour
equivalent to 34.5% apparent amylose content (Jane et al., 1999).
The highly branched structure of amylopectin molecules creates steric
hindrance, which prevents the formation of highly ordered amylopectin-lipid
complex. Thus, the long-branch chains of amylopectin, after being released
from the parent molecules through the debranching reaction, can form helical-
complex with lipids more effectively.
4.4 APPLICATIONS OF THE RS TYPE 5
The (ISO-treated HA7)-FA and (PUL-treated HA7)-FA complex products
contain RS up to 75% after enzyme digestion at 100 �C in excess water
following the AOAC Method 991.43 (Table 4.1). The results indicate that RS
type 5 in the (ISO-treated HA7)-FA and (PUL-treated HA7)-FA complex
products are thermally stable and, thus, are suitable for food processing that
requires heating.
In addition, the heat stability of an (ISO-treated HA7)-PA complex product
has been tested in bread making. In a white bread recipe, 72% (dry basis, db)
of wheat flour was replaced with (ISO-treated HA7)-PA complex product
(60%, db) and wheat gluten (12%, db) (Hasjim et al., 2010). The analytical
result of the RS content of the bread containing RS type 5, determined using
the AOAC Method 991.43, was 34.4% (db), which was similar to the value
calculated (33.6%, db) from the RS contents of the wheat flour, (ISO-treated
HA7)-PA complex product and wheat gluten used to make the bread (Table
4.3). Although bread made with such a high content of RS type 5 was not
intended for a desirable texture, the result proves that RS type 5 is heat stable
and that the enzyme resistance remains in the bread after baking.
86 Resistant Starch

CH04GR 07/29/2013 17:14:46 Page 87
4.5 HEALTH BENEFITS OF RS TYPE 5
The high contents of RS type 5 in the (ISO-treated HA7)-FA and (PUL-treated
HA7)-FA complex products from in vitro studies (Tables 4.1 and 4.2) suggest
that they have potential health benefits in controlling postprandial glycemic
and insulinemic responses and preventing colon cancer (Higgins, 2004;
Nugent, 2005). Human and animal studies have been conducted to investigate
the health benefits of the RS type 5 (Hasjim et al., 2010; Zhao et al., 2011).
4.5.1 Glycemic and insulinemic control
Ahuman feeding study was conducted to understand the effects of RS type 5 on
postprandial glycemic and insulinemic responses (Hasjim et al., 2010). The two
types of breads used in the study were control white bread and RS-type-5 bread
containing 60%, db, (ISO-treated HA7)-PA complex product (Table 4.3). The
results showed that the total areas under the curves of the postprandial glycemic
and insulinemic responses were reduced to 55% and 43%, respectively, when
the human subjects ingested the bread containing RS type 5, compared with the
control white bread (as 100%) (Figure 4.1). Similarly, reduced postprandial
insulinemic response was also observed from the rats fed with starch-mono-
stearoylglycerol complex, which was accompanied by declined lipogenesis in
the adipose tissue and liver (Takase et al., 1994).
The results from the in vivo studies indicate that RS type 5 can be used to
prevent the occurrence of hyperglycaemia and hyperinsulinaemia, which also
Table 4.3 Resistant starch content of breadmade from RS type 5. Reprinted fromHasjimet al., 2010.
Resistant starch (%)
Samples Analytical1 Calculated2
Wheat flour 2.1 –(ISO-treated HA7)-PA complex 52.7 –Wheat gluten 11.0 –Control white bread3 3.3 2.0RS type 5 bread4 34.4 33.6
1Resistant starch analyzed using the AOAC Method 991.43.2Resistant starch calculated on the basis of resistant starch contents of wheat flour, (ISO-treated HA7)-
PA complex product and wheat gluten.3Control white bread contained 96% wheat flour.4Enzyme-resistant bread contained 24% wheat flour, 60% (ISO-treated HA7)-PA complex product and
12% wheat gluten.
Novel Applications of Amylose-Lipid Complex as Resistant Starch Type 5 87

CH04GR 07/29/2013 17:14:46 Page 88
prevents the hyperglycaemia-induced hypoglycaemia and the constant feeling
of hunger associated with it (Cryer, 1999; Weinger et al., 1995). Furthermore,
the results showed that the insulinemic responsewas reduced to 2mU/L (close
to the baseline) 120 minutes after ingesting the bread containing RS type 5,
versus about 12mU/L after ingesting the white bread (Figure 4.1; Hasjim
et al., 2010). This suggests that RS type 5 may prevent the development of
insulin resistance, an abnormality caused by persistent hyperinsulinaemia
from frequent consumption of high glycemic-response foods (Byrnes et al.,
(a) Glycemic response
(b) Insulinemic response
RS5 breadWhite bread
RS5 bread
White bread
Time (min)
Incr
ease
in g
luco
se (
mg/
dL)
Incr
ease
in in
sulin
(m
U/L
)
40
35
30
25
20
15
10
5
0
30
25
20
15
10
5
0
–15 0 15 30 45 60 75 90 105 120
Time (min)–15 0 15 30 45 60 75 90 105 120
45
Figure 4.1 (a) Average postprandial glycemic and (b) insulinemic responses of 20male human subjects after ingesting control white bread and bread containing 60%(ISO-treated HA7)-PA complex product (RS type 5). Reprinted from Hasjim et al.,2010.
88 Resistant Starch

CH04GR 07/29/2013 17:14:46 Page 89
1995; Higgins, 2004). Thus, RS type 5 has a potential for interventions of
metabolic syndrome including type 2 diabetes, obesity, hypertension, lipid
abnormalities and heart disease, which have been associated with repeated
hyperinsulinaemia and insulin resistance (Higgins, 2004; Ludwig, 2002;
Takase et al., 1994).
4.5.2 Colon cancer prevention
Potential gut health benefits of RS type 5 were studied using an animal model,
and the (PUL-treated HA7)-SA complex product was used as the source of RS
type 5 (Zhao et al., 2011). Normal maize starch (NMS) and HA7 were used as
negative and positive controls, respectively. The starches were heated in
excess water (1 g starch in 3 g water) on a stove until they were boiling or
formed consistent starch paste. After cooking, the starches, at 55% (db) of the
total weight, were mixed with other ingredients to prepare NMS, HA7 and RS-
type-5 diets following the recipe of AIN-93 Rodent Diets (Reeves, 1997)
without the addition of cellulose.
Three groups of five-week-old male Fischer-344 rats were injected with
two doses (20mg/kg body weight) of azoxymethane (AOM) in oneweek apart
and fed with NMS diet until three days after the second injection, followed by
a treatment diet (NMS, HA7, or RS-type-5 diet). After eight weeks of feeding
on the treatment diet, the rats were sacrificed, and the occurrence of mucin-
depleted foci (MDF) and aberrant crypt foci (ACF) in the colon as well as the
caecum digesta weight and pH were analyzed. MDF and ACF are commonly
considered as the precursor lesions of chemically induced colon cancer
(Arakaki et al., 2006; McLellan et al., 1991).
It was found that the average number of MDF in the colons of the rats fed
with the RS-type-5 diet was significantly lower than those in the colons of the
rats fed with the NMS and HA7 diets (Table 4.4). The average number of ACF
was also reduced by the RS-type-5 diet, although the number was not
significantly different from those of the other diets. A significantly larger
amount of caecal digesta and a lower caecum pH were observed in the group
fed with RS type 5, compared with those in the other two groups (Table 4.4).
Lowering the pH of digesta may create an environment that suppresses the
growth of pathogenic organisms in the colon (Kleessen et al., 1997; Silvi
et al., 1999). No significant differences were found in the average daily food
intake and body weight gain among the rats fed with different diets in this
study.
In another study using the same animal model and diets, it was found that a
significantly larger amount of faeces was discharged by the rats fed with RS
type 5 than by those fed with the other two diets (Table 4.5). In addition, the
Novel Applications of Amylose-Lipid Complex as Resistant Starch Type 5 89

CH04GR 07/29/2013 17:14:46 Page 90
faeces from the rats fed with RS type 5 contained significantly greater
amounts of starch and lipid than those from the other two groups fed with
NMS and HA7. Polarized-light micrographs of the faeces collected from the
rats fed with RS type 5 showed starch granules with birefringence (Figure
4.2). The results of gas chromatography showed that SA was the major
component of the lipids extracted from the faeces of the rats fed with RS type
5, indicating that a proportion of the (PUL-treated HA7)-SA complex product
was not digested in the small intestine and was not fermented in the colon
(Table 4.5). Therefore, SA was not absorbed in the digestive tract and was
discharged as starch-lipid complex. The results also suggest that 15% (db) of
Table 4.4 Mucin-depleted foci, aberrant crypt foci and caecum digesta weight and pHfrom rats injected with azoxymethane as colon cancer inducer (Zhao et al., 2011).1
Adapted from Zhao et al., 2011. Copyright 2011 American Chemical Society.
Diet2
Mucin-depleted foci(MDF)
Aberrantcrypt foci(ACF)
Caecaldigestaweight (g)
CaecumdigestapH
Cooked normalmaize starch
3.5�1.8 47.5�13.4 3.29 7.5�0.1
Cooked HA7 1.8�1.4� 39.6�18.0 13.59� 6.6�0.6�
Cooked (PUL-treatedHA7)-SA complex
0.3�0.5� 30.9�25.0 18.67� 5.7�0.2�
1Values are means� standard deviations.2Each diet contained 55% (db) starch.�Significant at p<0.05 compared with cooked normal maize starch diet.
Table 4.5 Daily weight, total starch content, and lipid content of faeces collected fromrats fed with different diets.1
Faeces weight (g/day) Starch contentof faeces (%)3
Lipid contentof faeces (%)4
Diet2 As is Dry
Cooked normalmaize starch
0.23�0.07 a 0.20�0.06 a 0.2�0.0 a 16.8�0.4 ab
Cooked HA7 0.55�0.31 a 0.45�0.25 a 16.8�1.0 b 11.7�0.3 aCooked(PUL-treated
HA7)-SAcomplex2.04�0.76 b 1.65�0.62 b 44.0�0.0 c 22.0�2.2 b
1Values are means� standard deviations. Different letters on the same column represent significant
difference at p<0.05.2Each diet contained 55% (db) starch.3Starch contentwasanalyzedusingMegazymeTotal StarchAssayKit following theAACCMethod76-13.4Lipid content analyzed using the AOAC Method 996.06.
90 Resistant Starch

CH04GR 07/29/2013 17:14:46 Page 91
the consumed RS type 5 was not utilized by the rats and the microbes residing
in the digestive tract of the rats. The inferior digestibility of the RS type 5,
however, did not affect the body weight gain of the rats.The results from these two in vivo rat studies suggest two possible
mechanisms of RS type 5 in reducing the amount of MDF in the colon.
One is the greater amount of the faeces discharged from the colon. The
larger, bulky amount of faeces can purge carcinogen (e.g. the injected
AOM) out from the colon. The other is the FA present in the starch-lipid
complex, which may have the affinity to absorb carcinogen and to remove
it from the colon.
4.6 CONCLUSION
Amylose-lipid complex has been proposed as RS type 5, because of its
resistance to enzyme hydrolysis. The presence of amylose-lipid complex in
starch granules also increases their enzyme resistance by restricting the
granule swelling during cooking. The enzyme resistance of amylose-lipid
complex depends on the molecular structure of the lipid and the crystalline
structure of the single helices. FA with a longer hydrocarbon chain length
produces amylose-lipid complex with greater enzyme resistance, whereas FA
with a greater degree of unsaturation produces amylose-lipid complex with
Figure 4.2 Polarized-light micrograph of starch isolated from faeces of rats fed with(PUL-treated HA7)-SA complex product.
Novel Applications of Amylose-Lipid Complex as Resistant Starch Type 5 91

CH04GR 07/29/2013 17:14:47 Page 92
lower enzyme resistance. Furthermore, a crystalline form of amylose-lipid
complex has greater enzyme resistance than its amorphous counterpart.
A technology has been recently developed to produce RS using amylose-
lipid complex by treating granular HA7 starch with debranching enzyme,
followed by complex formation with FA. The RS contents of the complex
products were significantly higher (up to 75%). Feeding human subjects with
this RS type 5 reduced postprandial glycemic and insulinemic responses
compared with the control diet of white bread, which suggests that RS type 5
has potential to intervene in metabolic syndromes including type-2 diabetes,
obesity, hypertension, lipid abnormalities and heart disease, as well as other
complications associated with repeated hyperinsulinaemia and insulin resist-
ance. RS type 5 also shows the ability to reduce colon cancer development, as
observed by lowering the amount of MDF in the colons of AOM-treated rats.
REFERENCES
Ai, Y., Hasjim, J., Jane, J. (2013). Effects of lipids on enzymatic hydrolysis and physicalproperties of starch. Carbohydrate Polymers 92(1), 120–127.
Arakaki, J., Suzui, M., Morioka, T., Kinjo, T., Kaneshiro, T., Inamine, M., Sunagawa, N.,Nishimaki, T., Yoshimi, N. (2006). Antioxidative and modifying effects of a tropicalplant Azadirachta indica (neem) on azoxymethane-induced preneoplastic lesions in therat colon. Asian Pacific Journal of Cancer Prevention 7(3), 467–471.
Becker, A., Hill, S.E., Mitchell, J.R. (2001). Relevance of Amylose-Lipid Complexes to theBehaviour of Thermally Processed Starches. Starch - St€arke 53(3–4), 121–130.
Biliaderis, C.G., Galloway, G. (1989). Crystallization behavior of amylose-V complexes:Structure-property relationships. Carbohydrate Research 189, 31–48.
Byrnes, S.E., Brand Miller, J.C., Denyer, G.S. (1995). Amylopectin starch promotes thedevelopment of insulin resistance in rats. Journal of Nutrition 125(6), 1430–1437.
Cryer, P.E. (1999). Symptoms of hypoglycemia, thresholds for their occurrence, andhypoglycemia unawareness. Endocrinology & Metabolism Clinics of North America28(3), 495–500.
Cui, R., Oates, C.G. (1999). The effect of amylose-lipid complex formation on enzymesusceptibility of sago starch. Food Chemistry 65(4), 417–425.
Eerlingen, R.C., Delcour, J.A. (1995). Formation, analysis, structure and properties of typeIII enzyme resistant starch. Journal of Cereal Science 22(2), 129–138.
Eliasson, A.C., Ljunger, G. (1988). Interactions between amylopectin and lipid additivesduring retrogradation in a model system. Journal of the Science of Food and Agriculture44(4), 353–361.
Englyst, H.N., Kingman, S.M., Cummings, J.H. (1992). Classification and measurementof nutritionally important starch fractions. European Journal of Clinical Nutrition46 Suppl 2, S33–S50.
Gelders, G.G., Duyck, J.P., Goesaert, H., Delcour, J.A. (2005). Enzyme and acid resistanceof amylose-lipid complexes differing in amylose chain length, lipid and complexationtemperature. Carbohydrate Polymers 60(3), 379–389.
92 Resistant Starch

CH04GR 07/29/2013 17:14:47 Page 93
Hasjim, J., Lee, S.-O., Hendrich, S., Setiawan, S., Ai, Y., Jane, J. (2010). Characterizationof a novel resistant-starch and its effects on postprandial plasma-glucose and insulinresponses. Cereal Chemistry 87(4), 257–262.
Higgins, J.A. (2004). Resistant starch: Metabolic effects and potential health benefits.Journal of AOAC International 87(3), 761–768.
Holm, J., Bj€orck, I., Ostrowska, S., Eliasson, A.C., Asp, N.G., Larsson, K., Lundquist, I.(1983). Digestibility of amylose-lipid complexes in-vitro and in-vivo. Starch - St€arke35(9), 294–297.
Horwithz, W. (ed). (2003). Official Methods of Analysis of AOAC International. Gaithers-burg, Maryland: AOAC International.
Huang, J.J., White, P.J. (1993). Waxy corn starch: Monoglyceride interaction in a modelsystem. Cereal Chemistry 70(1), 42–47.
Jane, J., Robyt, J.F. (1984). Structure studies of amylose-V complexes and retrogradedamylose by action of alpha amylases, and a new method for preparing amylodextrins.Carbohydrate Research 132(1), 105–118.
Jane, J.L., Robyt, J.F., Huang, D.H. (1985). 13C-N.M. R. study of the conformation ofhelical complexes of amylodextrin and of amylose in solution. Carbohydrate Research140(1), 21–35.
Jane, J.-L., Craig, S.A.S., Seib, P.A., Carl Hoseney, R. (1986a). A granular coldwater-solublestarch gives a V-type X-ray diffraction pattern. Carbohydrate Research 150(1), C5–C6.
Jane, J., Craig, S.A.S., Seib, P.A., Hoseney, R.C. (1986b). Characterization of granular coldwater-soluble starch. Starch - St€arke 38(8), 258–263.
Jane, J., Chen, Y.Y., Lee, L.F., McPherson, A.E., Wong, K.S., Radosavljevic, M.,Kasemsuwan, T. (1999). Effects of amylopectin branch chain length and amylosecontent on the gelatinization and pasting properties of starch. Cereal Chemistry76(5), 629–637.
Jiang, H., Campbell, M., Blanco, M., Jane, J. (2010). Characterization of maize amylose-extender (ae) mutant starches: Part II. Structures and properties of starch residuesremaining after enzymatic hydrolysis at boiling-water temperature. CarbohydratePolymers 80(1), 1–12.
Karkalas, J., Raphaelides, S. (1986). Quantitative aspects of amylose-lipid interactions.Carbohydrate Research 157, 215–234.
Karkalas, J., Ma, S., Morrison, W.R., Pethrick, R.A. (1995). Some factors determining thethermal properties of amylose inclusion complexes with fatty acids. CarbohydrateResearch 268(2), 233–247.
Kitahara,K., Suganuma, T., Nagahama, T. (1996). Susceptibility of amylose-lipid complexesto hydrolysis by glucoamylase from Rhizopus niveus. Cereal Chemistry 73(4), 428–432.
Kleessen, B., Stoof, G., Proll, J., Schmiedl, D., Noack, J., Blaut, M. (1997). Feedingresistant starch affects fecal and cecal microflora and short-chain fatty acids in rats.Journal of Animal Science 75(9), 2453–2462.
Kugimiya, M., Donovan, J.W., Wong, R.Y. (1980). Phase transitions of amylose-lipidcomplexes in starches: A calorimetric study. Starch - St€arke 32(8), 265–270.
Li, L., Jiang, H., Campbell, M., Blanco, M., Jane, J. (2008). Characterization of maizeamylose-extender (ae) mutant starches. Part I: Relationship between resistant starchcontents and molecular structures. Carbohydrate Polymers 74(3), 396–404.
Ludwig, D.S. (2002). The glycemic index: Physiological mechanisms relating to obesity,diabetes, and cardiovascular disease. Journal of the American Medical Association 287,2414.
Novel Applications of Amylose-Lipid Complex as Resistant Starch Type 5 93

CH04GR 07/29/2013 17:14:48 Page 94
McLellan, E.A., Medline, A., Bird, R.P. (1991). Sequential analyses of the growth andmorphological characteristics of aberrant crypt foci: Putative preneoplastic lesions.Cancer Research 51(19), 5270–5274.
Morrison, W.R. (1981). Starch lipids: A reappraisal. Starch - St€arke 33(12), 408–410.Morrison, W.R., Milligan, T.P., Azudin, M.N. (1984). A relationship between the amylose
and lipid contents of starches from diploid cereals. Journal of Cereal Science 2(4),257–271.
Morrison, W.R., Law, R.V., Snape, C.E. (1993). Evidence for inclusion complexes of lipidswith V-amylose in maize, rice and oat starches. Journal of Cereal Science 18(2),107–109.
Nugent, A.P. (2005). Health properties of resistant starch. Nutrition Bulletin 30(1), 27–54.Raphaelides, S., Karkalas, J. (1988). Thermal dissociation of amylose-fatty acid com-
plexes. Carbohydrate Research 172(1), 65–82.Reeves, P.G. (1997). Components of the AIN-93 diets as improvements in the AIN-76A
diet. Journal of Nutrition 127(5), 838S–841S.Seneviratne, H.D., Biliaderis, C.G. (1991). Action of a-amylases on amylose-lipid
complex superstructures. Journal of Cereal Science 13(2), 129–143.Shogren, R.L., Fanta, G.F., Felker, F.C. (2006). X-ray diffraction study of crystal
transformations in spherulitic amylose/lipid complexes from jet-cooked starch. Carbo-hydrate Polymers 64(3), 444–451.
Silvi, S., Rumney, C.J., Cresci, A., Rowland, I.R. (1999). Resistant starch modifies gutmicroflora and microbial metabolism in human flora-associated rats inoculated withfaeces from Italian and UK donors. Journal of Applied Microbiology 86(3), 521–530.
Takase, S., Goda, T., Watanabe, M. (1994). Monostearoylglycerol-starch complex: Itsdigestibility and effects on glycemic and lipogenic responses. Journal of NutritionalScience and Vitaminology 40(1), 23–36.
Tester, R.F., Morrison, W.R. (1990). Swelling and gelatinization of cereal starches.I. Effects of amylopectin, amylose, and lipids. Cereal Chemistry 67(6), 551–557.
Tufvesson, F., Wahlgren, M., Eliasson, A.-C. (2003). Formation of amylose-lipid com-plexes and effects of temperature treatment. Part 2. Fatty acids. Starch - St€arke 55(3–4),138–149.
Weinger, K., Jacobson, A.M., Draelos, M.T., Finkelstein, D.M., Simonson, D.C. (1995).Blood glucose estimation and symptoms during hyperglycemia and hypoglycemiain patients with insulin-dependent diabetes mellitus. The American Journal of Medicine98(1), 22–31.
Yotsawimonwat, S., Sriroth, K., Kaewvichit, S., Piyachomkwan, K., Jane, J.-L., Sirithu-nyalug, J. (2008). Effect of pH on complex formation between debranched waxyrice starch and fatty acids. International Journal of Biological Macromolecules43(2), 94–99.
Zhao, Y., Hasjim, J., Li, L., Jane, J.-L., Hendrich, S., Birt, D.F. (2011). Inhibition ofazoxymethane-induced preneoplastic lesions in the rat colon by a cooked stearic acidcomplexed high-amylose cornstarch. Journal of Agricultural and Food Chemistry59(17), 9700–9708.
94 Resistant Starch

CH05GR 07/29/2013 16:2:52 Page 95
5 Digestion Resistant Carbohydrates
Annette EvansInnovation and Commercial Development, Tate & Lyle, USA
5.1 INTRODUCTION
The rates of obesity, diabetes, cardiovascular and other diseases have
increased dramatically over the last century, which has led to increased
consumer awareness of the importance of nutrition in the development and
progression of these diseases. As a result of this awareness, and increased
scientific research in this area, demand for ‘healthier options’ is high.
Consumers understand the value and potential health benefits of dietary fiber,
so one top demand is for fiber-containing foods that taste good. For food
ingredient companies, this presents both an opportunity and a challenge to
supply the market with new and nutritionally improved carbohydrate ingre-
dients, including starch and fiber.
To develop fiber-containing ingredients, it is important to understand the
enzymes involved in food digestion, as well as the properties of a food that
make it either highly digestible or highly enzyme-resistant and, therefore, high
in dietary fiber. In the case of starch, the structures responsible for enzyme
resistance can be either physical or molecular. This chapter examines the
physical and molecular structures of starch that influence enzyme digestibil-
ity, and describes examples of commercially available resistant starch ingre-
dients and their source of enzyme resistance related to starch structure.
5.2 STARCH DIGESTION
Starch is the major source of carbohydrates in the human diet. Starch can be
obtained from fruit, vegetables, roots and grains. Starch and starch derivatives
are a nutritive, abundant and economical food source. Today, starch is
consumed unprocessed in the form of raw fruits and vegetables, or in the
95
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

CH05GR 07/29/2013 16:2:52 Page 96
form of more shelf-stable processed foods (Moore et al., 1984). The process-
ing of starch evolved in the last 80 years from a simple isolation process to the
production of diversified lines of sweeteners and starch products (O’Dell,
1979). Food starches contribute to the characteristic viscosity, texture, mouth-
feel and consistency of many food products.
Starch is a glucose polymer composed of highly branched amylopectin
(a-1,4 and a-1,6-linkages) and essentially linear amylose molecules (mainly
a-1,4-linkages) (Figure 5.1).
Starch digestion in the human body is mainly done by a-amylases. Salivary
a-amylase is the first amylase that comes in contact with the food in the
mouth. It catalyzes the hydrolysis of amylose to maltose, maltotriose and
maltotetraose, and the hydrolysis of amylopectin to the same products plus
two a-limit dextrins (Robyt, 1984). The salivary a-amylase quickly passes
from the mouth to the stomach, together with the food starch. The pH in the
stomach is about 2, which inactivates the salivary a-amylase. After some
resident time in the stomach, the partially hydrolyzed starch passes into the
small intestine, where it is neutralized. The main hydrolysis of the starch is
then accomplished by pancreatic a-amylase, secreted from the pancreatic
duct, via a multiple attack mechanism (Mazur & Nakatani, 1993).
Two other enzymes are necessary to convert the hydrolysis products of the
a-amylases into glucose, which can be actively transported across the small
intestine membrane. These two enzymes are the brush border glucogenic
enzymes maltase-glucoamylase and sucrase-isomaltase (Swallow et al., 2001;
Nichols et al., 2003). Maltase-glucoamylase converts maltose, maltotriose,
and maltotetraose into D-glucose by successive action from the non-reducing
O
OH
OH
HHO
HO
H
H H
H
O
O
OH
HHO
H
H H
H
O
O
OH
OHH
OHH
H H
H
O
HOO
OH
OHH
HOH
H H
H
OH
α-1,6-linkage
α-1,4-linkage
Figure 5.1 Structures found in starch molecules.
96 Resistant Starch

CH05GR 07/29/2013 16:2:52 Page 97
end (Robyt, 1984). Sucrase-isomaltase hydrolyses the a-1,6-linkages. Theresulting D-glucose can then actively be transported across the luminal
membrane of the small intestine and thence into the blood via the sodium-
glucose cotransporter (SGLT-1), which is located at the luminal surface of
enterocytes (Pencek et al., 2002).
Starch digestion is influenced by factors that affect enzyme activity and the
susceptibility of the starch substrate to the hydrolytic enzymes (Tester et al.,
2004). Enzyme activity may depend on quantity of the enzymes present,
enzyme isoform and enzyme inhibitors either present in the consumed food or
produced during enzyme hydrolysis of the food. While brush border enzymes
are capable of cleaving both a-1,4 and a-1,6 glycosidic bonds as well as otherglycosidic bonds (e.g. a 1,2), the main hydrolysis of starch is performed by the
a-amylases. Starch digestion by a-amylases requires a series of steps where
the enzymes first need to diffuse to the starch matrix into the food, then bind
to the substrate, then finally cleave the a-1,4-glycosidic linkages (Brayer
et al., 2000).
5.3 PHYSICAL STRUCTURES OF STARCH
To understand factors that may influence the susceptibility of a particular
starch substrate to a-amylase hydrolysis, four aspects of hydrolysis should be
considered: the diffusion of the enzyme towards the substrate; the porosity of
the starch substrate; the adsorption of the enzyme on the substrate; and the
catalytic event (Colonna et al., 1992). Porosity has to be considered at two
levels of scale: at the macro (1–100mm) and at the micro molecular level
(<10 nm) (Colonna et al., 1992). The physical form of the starch also
influences enzyme binding. Physical entrapment of starch in the food matrix,
starch granular structure, and also smaller scale structures such as crystallinity
and helicity, may influence the binding of enzymes to the starch substrate.
For native starch granules, the limiting factor for the hydrolysis has been
shown to be the penetration of the enzyme into the granules by successive
formation of pits and larger pores (Gallant et al., 1973).
Before a substrate can be hydrolyzed, adsorption of the enzyme onto the
substrate is crucial. Any factors that influence the binding of the a-amylases to
the starch substrate can slow down or limit starch digestion. The hydrolysis
products maltose and maltotriose have been shown to act as substrate
inhibitors for a-amylase (Seigner et al., 1985).
For an enzyme to be able to hydrolyze a substrate, proper enzyme-substrate
recognition is necessary. Substrate binding by means of the enzyme sub-sites
has to be energetically favoured. For the catalytic action of the enzyme to
Digestion Resistant Carbohydrates 97

CH05GR 07/29/2013 16:2:52 Page 98
occur, the substrate to be hydrolyzed has to be oriented properly inside the
active side of the enzyme. Only the part of the starch substrate that can fit into
the active site cavity of the a-amylase can be hydrolyzed. Structures such as
double helices are too big to fit into the active site cavity and can only be
hydrolyzed in the unwound state.
5.3.1 Starch helices
Amylose is known to form single helices in solution. The addition of
complexing ligands such as iodine, fatty acids, alcohols, emulsifiers or
flavour compounds to aqueous starch solutions induces the formation of
single helices (Heinemann et al., 2003). V-amylose is the generic term for
amylose obtained by complexation, and the compounds used to obtain the
single helices may or may not be present in the final V-type starch (Buleon
et al., 1998). The amylose single helix has been described as a left-handed
helix with six glucose residues per turn, with a pitch height of about 0.7 nm
(Buleon et al., 1998). The outer surface of the single helix is thought to be
mostly hydrophilic, while the centre channel of the helix is hydrophobic.
Single helices are stabilized by 2-O . . . O-6 hydrogen bonds between
spatially close glucose residues of the next spiral turn (intrahelical hydrogen
bonds). The structure of the single helices is further cooperatively strength-
ened by 2-O . . . O-3 hydrogen bonds between adjacent glucose units
(Immel & Lichtenthaler, 2000).
The formation of singles helices from branched material has been little
studied. Formation of single helices from amylopectin with complexing
agents has been shown (Heinemann et al., 2003). Considering the structure
of amylopectin molecules, it would be expected that external chains that are in
close proximity to each other may easily form double helices.
The external chains of amylopectin and amylose chains are known to be
able to form double helices within starch granules and in non-granular form
(Wu & Sarko, 1978a, 1978b, 1978c). Evidence and models for both right-
handed (Mueller et al., 1995) and left-handed double helices (Imberty et al.,
1988) have been provided. The left-handed form is the energetically favoured
form and is commonly accepted as the form of starch double helices (Imberty
et al., 1988; Buleon et al., 1998). The starch double helix has a pitch of
2.13 nm with six glucose units per turn (Imberty et al., 1988, Buleon et al.,
1998). Starch double helices are stabilized by hydrogen bonds between
2-O . . . O-6 of glucose units on different strands of the double helix
(interhelical hydrogen bonds). The outer surface of the double helix has an
irregular distribution of hydrophobic and hydrophilic areas (Immel and
Lichtenthaler, 2000).
98 Resistant Starch

CH05GR 07/29/2013 16:2:52 Page 99
5.3.2 Crystalline structures
Single helices formed by non-granular starch can arrange themselves to form
crystalline entities. If the crystalline arrays are large enough to diffract x-rays,
an x-ray diffraction pattern can be observed. The characteristic structure
of crystalline V-amylose has been extensively studied (Rappenecker &
Zugenmaier, 2001; Godet et al., 1995). The double helices formed by external
chains of amylopectin or amylose molecules can also form crystalline entities.
Double helical starch can form different crystalline polymorphs. Native
starch granules can be found with A-type crystallinity (cereal starches) and
B-type crystallinity (tubers, high-amylose maize starches) (Banks & Green-
wood, 1975; Zobel, 1988). The double helical structures within the two
polymorphic forms are essentially identical (Imberty et al., 1991), but the
packing of the helices within the crystalline structure is different. In general,
A-type crystals are less hydrated and more densely packed than B-type
crystals (Imberty et al., 1987; Imberty & Perez, 1988).
Once starch has lost its granular structure, it can be crystallized into one of
the two crystalline polymorphs, depending on the starch structure and
crystallization conditions. The A-type polymorph is the more thermo-
dynamically stable form and is preferably formed by shorter chains, higher
temperature and lower starch concentrations. The B-type polymorph is
formed by longer starch chains, lower temperatures and higher starch con-
centrations (Gidley & Bulpin, 1987). B-type crystallites are commonly
believed to be more enzyme resistant, while A-type polymorphs are more
heat stable compared to B-type formed from chains with similar molecular
weight (Whittam et al., 1990; Le Bail et al., 1995).
5.3.3 Starch granule structure
Starch granules are the naturally occurring storage form of starch. Different
levels of organization exist in native starch granules. At the lowest such level,
the starch granules consist of amorphous and semi-crystalline shells that are
often called growth rings. At a higher level of structure are the so-called
‘blocklets’ (Gallant et al., 1997). One blocklet is thought to contain several
amorphous and crystalline lamellae that are, in turn, created by the chain
organization of amylopectin molecules. The side chains of amylopectin
can form double helices and, due to the close proximity of the chains to
each other, the double helices can easily form crystalline arrays. These
crystalline arrays, and the non-crystalline zones between them that have
been shown to contain the amylopectin branch points (Jenkins et al., 1993),
are organized into lamellae.
Digestion Resistant Carbohydrates 99

CH05GR 07/29/2013 16:2:52 Page 100
Heating of starch granules can lead to partial or complete melting of the
crystallites and helices present in the granules. Depending on the heat
treatment conditions used, the heat treatment can be classified as gelatiniza-
tion, annealing or heat-moisture treatment. Gelatinization of starch is defined
as the complete melting of granular structures in the presence of excess water.
Annealing and heat-moisture treatment are both heat treatments that only
partially melt structures in the starch granules by using temperature/moisture
combinations below the melting point of the treated starch.
Annealing is a term used in polymer chemistry to describe a treatment that
uses heat and moisture to increase the order of polymer chains. In the case of
starch, the term ‘annealing’ is used to describe a heat treatment that employs
temperatures below the melting point of the starch at moisture contents above
40%. Heat-moisture treatment of starch is described as a heat treatment with
temperatures below the melting point of starch, at moisture levels below 35%
(Stute, 1992; Jacobs & Delcour, 1998). These treatments are generally used to
‘perfect’ crystalline order in starch granules.
5.4 RESISTANT STARCH DUE TO PHYSICAL STRUCTURE
Resistant starch has been defined as ‘the starch and products of starch
digestion that are not absorbed in the small intestine of healthy individuals’
(Asp, 1992). Based on the source of the enzyme resistance, resistant starch has
been classified into five different types (Table 5.1). In type 2, type 3 and type 5
resistant starches, the enzyme resistance is due to the physical structure of the
starch molecules.
The physical structures responsible for formation of type 2 resistant starch
are part of the natural organization of starch granules. Examples of starches
high in type 2 resistant starch are green banana, potato and high-amylose
Table 5.1 Classification of resistant starch (RS).
RS type Source of enzyme resistance Examples
1 Whole grains Seeds, grains2 Raw starch granule structure Green banana, high-amylose,
potato3 Structures formed on re-association or
retrogradation of starch after heattreatment
Cooked pasta, cooked potato
4 Chemical modifications of the starch Modified starch5 Starch lipid complex Amylose containing starch
100 Resistant Starch

CH05GR 07/29/2013 16:2:52 Page 101
maize. Several commercial type 2 resistant starch ingredients are available
These are usually high-amylose starches that might undergo heat-moisture
treatment to enhance their inherent RS content (US Patent 5593503).
Type 3 resistant starch is formed by reassociation of starch chains after heat
treatment. Different factors, such as amylose/amylopectin ratio, chain length,
lipid content and processing conditions, influence the amount and quality of
the type 3 RS (Eerlingen & Delcour, 1995). Annealing, heat moisture
treatment and gelatinization of starch all melt crystalline structures present
in native starch granules either fully or partially. Upon cooling, linear amylose
molecules, as well as linear regions of amylopectin molecules, organize into a
mix of amorphous areas as well as helices and crystallites, with varying
degrees of enzyme resistance and, therefore, varying levels of resistant starch.
In the case of gelatinization, this reassociation of starch chains upon cooling is
called retrogradation.
The resulting resistant starch often exhibits B-type crystallinity, but A-type
crystalline RS can also be obtained if the retrogradation or crystallization
occurs at high temperatures or involved starch molecules with short chain
length. Formation of crystalline structures that can form RS 3 take place above
the glass transition temperature (and below the melting temperature), and any
components present that influence the glass transition temperature can,
therefore, be expected to influence the formation (yield and quality) of the
formed type 3 RS.
Amylose content has been positively correlated with RS yield (Sievert &
Pomeranz, 1990) and the formation of type 3 RS is strongly related to the
crystallization process of amylose (Eerlingen & Delcour, 1995). The yield of
RS formed is dependent on the water content and temperature used. At high
starch concentrations, the starch chains interact more easily, leading to
increased crystal and RS formation. Water does act as a plasticizer in the
system, and a minimum water content is necessary to achieve the chain
mobility needed to form crystalline structure resistant to enzyme digestion.
The presence of lipids has been shown to decrease the formation of type 3 RS,
due to formation of amylose lipid complexes (type 5 RS) (Czuchajowaska
et al., 1991).
Annealing and heat-moisture treatment can be used for the manufacture
of resistant starch, since the more ‘perfect’ structures formed often lead to
an increase in enzyme resistance of the starch (Jacobs & Delcour, 1998;
Chiu et al., 1999; Brumovsky & Thompson, 2002).
Some commercial examples of type 3 resistant starch are Hi-maize 330
(National Starch and Chem. Co) and Promitor Resistant Starch 60 (Tate &
Lyle). Both of these ingredients are manufactured from high-amylose maize
starch. In both cases, the starch is first gelatinized and then cooled to
Digestion Resistant Carbohydrates 101

CH05GR 07/29/2013 16:2:52 Page 102
retrograde the starch chains. Heat-moisture treatments are then applied to the
retrograded starch to increase the molecular order and, therefore, the enzyme
resistance of the starch (US5281276; US7189288). Type 3 resistant starches
have also been produced by a process in which the starch is first treated with a
debranching enzyme to increase the amount of linear chains, followed by
controlled reassociation or crystallization of the linear chains to form enzyme
resistant starch. One commercially available example of type 3 resistant starch
produced in this fashion is ActistarTM RM (Cargill Inc.), which is produced
by debranching and crystallization of tapioca maltodextrin (Patent EP
0846704 B1).
Tate & Lyle recently described a resistant starch process in which a starch is
first treated with a glucanotransferase enzyme to elongate the external chains
of amylopectin, followed by debranching and then crystallization of the linear
chains (US Patent Application 2007/0059432 A1).
The enzyme resistance in Type 5 resistant starch is due to the molecular
structure of amylose lipid complexes that can either be present in the native
starch or formed by controlled reactions using non-granular starch and
lipids to form the resistant amylose lipid complex (Brown et al., 2006;
Hasjim et al., 2010).
5.5 MOLECULAR STRUCTURE OF STARCH
Starch is comprised of two polymers. Amylose is an essentially linear
polymer, in which glucose monomers are linked by a-1,4-glycosidic bonds.Amylopectin is a highly branched polymer that, in addition to the linear
a-1,4-linkages, also contains a-1,6-linked glucose monomers that create
branch points.
Certain processing treatments alter the molecular structure of starch and
may create additional linkage types. The amount and distribution of a-1,6-branch points can be altered by enzymatic treatment of starch. Debranching
enzymes such as isoamylase or pullulunase can be used to decrease the branch
points, and branching enzymes can be used to increase the amount of a-1,6-linkages in starch. Transferase enzymes can alter the distribution of branch
points in starch.
In addition to altering the amount and distribution of linkages present in
native starch, processing treatments can be used to create new types of
linkages not found in native starch molecules. When heat is applied to starch
at low moisture and low pH conditions, dextrinization of the starch occurs
(Wurzburg, 1986). During the initial stages of dextrinization, acid-catalyzed
hydrolysis occurs. This is followed by a recombination of the fragments to
102 Resistant Starch

CH05GR 07/29/2013 16:2:52 Page 103
form branched structures. The dextrinization process converts a portion of the
normal a-1,4-, a-1,6-linkages to random a-1,4-, a-1,6-, a-1,2-, a-1,3 and
even b-type linkages (Wurzburg, 1986). Figure 5.2 shows a representative
molecular structure of a dextrin.
Chemical modification of starch is a process widely used in the food
industry to alter the texture and processing stability of starch. Awide range of
properties can be achieved, depending on the botanical origin of the starch,
amylose and amylopectin structure, and the amount and type of modification
agent. Starch modification can be achieved through derivatization such as
etherification, esterification and cross-linking of starch (Figure 5.3). Chemical
modification involves the introduction of functional groups into the starch
molecules, resulting in significantly different properties. Starches with altered
gelatinization, pasting and retrogradation behaviour can be produced through
chemical modification (Singh et al., 2007).
5.6 ENZYME RESISTANCE DUE TOMOLECULARSTRUCTURE
The linear regions of the starch molecules are easily digested by human
pancreatic a-amylase. The small sugar and oligosaccharide digestion
products of a-amylase can then be digested further by the intestinal brush
border enzymes.
Branch points can create a steric hindrance for enzyme digestion, irre-
spective of the type of glycosidic linkage that creates the branching of the
starch chain. For a-amylase hydrolysis to occur, a linear portion of the starch
chain has to fit in the enzyme’s active site. This linear portion has to be long
enough to create energetically favourable binding. Glucose units close to the
branch points have less favourable binding energy with the sub-sites of the
enzyme than glucose units further down the chain; sub-site binding therefore
affects the efficiency with which the enzyme hydrolyses a linkage close to a
branch point. Linkages directly adjacent may not be hydrolyzed at all, leading
to the formation of limit dextrins (Shannon et al., 1984; Colonna et al., 1992).
Chemical modification of starch can be viewed as a modification that
creates chain irregularities or branches in the starch chains. Cross-linking of
starch covalently links two starch chains together, in effect creating a branch
point on both chains. Chemical substitution introduces a bulky side group to
the starch chains. The introduction of these chemical groups may create a
steric hindrance to one or more human digestive enzymes.
Several type 4 resistant starch ingredients are available commercially, with
total dietary fiber values of up to 90%. These type 4 products, from different
Digestion Resistant Carbohydrates 103
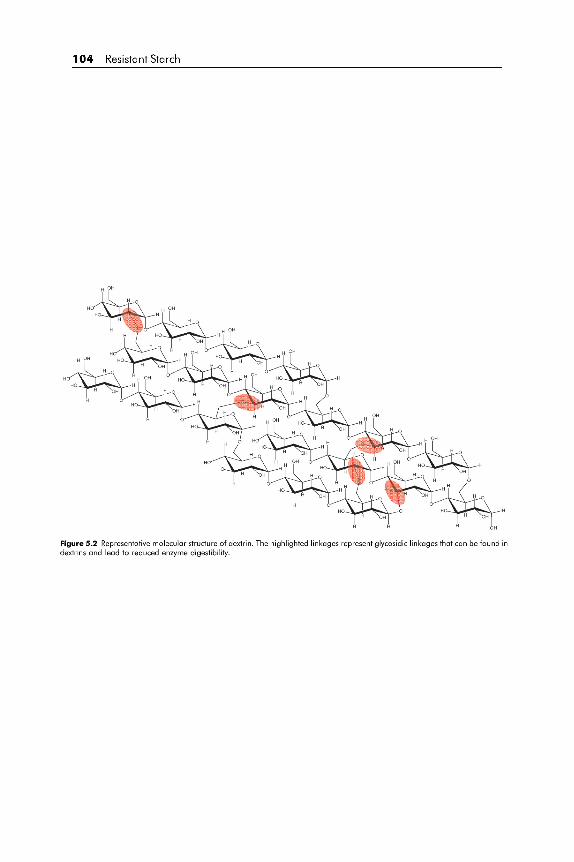

CH05GR 07/29/2013 16:2:53 Page 105
botanical origins, contain high amounts of resistant starch due to a high level
of chemical cross-linking (US 2006/0188631 A1). Substituted starches are not
currently produced as resistant starch ingredients.
Dextrinization of starch leads to the formation of potentially indigestible
linkages. The glycosidic linkages in dextrins are usually predominately a-1,4-, but a-1,6-, a-1,2-, a-1,3 and, possibly, even b-type linkages can be found(Wurzburg, 1986). The a-1,4- and a-1,6-bonds are digestible by human
pancreatic a-amylase and brushborder enzymes, respectively. However,
while some of the a-1,2-bonds might also be digestible by brushborder
enzymes, other bonds formed during dextrinization are not digestible by
human enzymes.
Commercially, the technique of dextrinization is used by several compa-
nies to create dietary fiber ingredients that (depending on their resulting
molecular size) can be classified as resistant starch or resistant maltodextrins.
The Nutriose1 line of dietary fiber ingredients is marketed by Roquette
(Roquette Fr�eres). Nutriose1 soluble fibres are food dextrins made from
wheat or maize starch (US5620871). Fibersol 2 is produced and marketed by
a joint venture between ADM (Archer Daniels Midland Company) and
Matsutani (Matsutani LLC). It is classified as resistant maltodextrin in the
USA (US5358729) and is produced by dextrinization of starch followed by a
proprietary enzymatic molecular weight reduction of the dextrins. Tate &
Lyle’s PromitorTM Soluble Corn Fiber ingredients are produced by heat
treatment of starch hydrolysis products at low pH (US7608436). They can
be classified as corn syrup or maltodextrin, depending on the molecular
weight of the resulting product.
O
HO
OO
O
HO
OO
O
HO
O
O
O
O
OP O
–O
bridgingphosphatediester
O
HO
O
OO
HO
OOOH
bulkyhydroxy-propylether
Figure 5.3 Structures found in modified starch. Left: structure of chemical cross-link(cross-linking with POCl3). Right: structure of chemical substitution (substitution withpropylene oxide).
Digestion Resistant Carbohydrates 105

CH05GR 07/29/2013 16:2:53 Page 106
5.7 CONCLUSION
Carbohydrates play an important role in the human diet for a number of
reasons. The World Health Organization recommends that at least 55% of the
energy of a diet be derived from carbohydrates. Starch is the major source of
carbohydrates in the human diet. Many carbohydrate-containing foods also
contain important micronutrients and phytochemicals.
Dietary fiber is a class of carbohydrates that are important in overall health.
Today, food ingredient companies have the knowledge and expertise to
maximize the health-giving potential of starch ingredients by modulating
not only the texture and sensory properties of commercial starches, but also
the digestibility and physiological effect of a starch ingredient.
REFERENCES
Asp, N.-G. (1992). Resistant Starch. Proceedings from the second plenary meeting ofEURESTA: European FLAIR concerned action no. 11 on physiological implications ofthe consumption of resistant starch in man. European Journal of Clinical Nutrition 46,S1.
Banks, W., Greenwood, C.T. (1975). Starch and its components. John Wiley & Sons,New York, NY.
Brayer, G.D., Sidhu, G., Maurus, R., Rydberg, E.H., Brown, C., Wang, Y.L., Nguyen, N.T.,Overall, C.H., Withers, S.G. (2000). Subsite mapping of the human pancreatic alpha-amylase active site through structural, kinetic, and mutagenesis techniques. Bio-chemistry 39, 4778–4791.
Brown, I.L., Yotsuzuka, M., Birkett, A., Henriksson, A. (2006). Prebiotics, synbiotics andresistant starch. Journal of Japanese Association for Dietary Fiber Research 10, 1–9.
Brumovsky, J.O., Thompson, D.B. (2002). Production of boiling-stable type II resistantstarch by partial acid hydrolysis and hydrothermal treatments of high-amylose maizestarch. Cereal Chemistry 78, 680–689.
Buleon, A.P., Colonna, P., Planchot, V., Ball, S. (1998). Starch granules: structure andbiosynthesis. International Journal of Biological Macromolecules 23, 85–112.
Chiu, C.-W., Shi, Y.-C., Sedam, M. (1999). Process for producing amylase resistantgranular starch. US Patent 5,902,410.
Colonna, P., Leloup, V., Buleon, A. (1992). Limiting factors of starch hydrolysis. EuropeanJournal of Clinical Nutrition 46, S17–S32.
Czuchajowska, Z., Sievert, D., Pomeranz, Y. Enzyme-resistant starch. IV. Effects ofcomplexing lipids. Cereal Chemistry. 68, 537–542.
Eerlingen, R.C., Delcour, J.A. (1995). Formation, analysis, structure and properties of TypeIII enzyme resistant starch. Journal of Cereal Science 22, 129–138.
Gallant, D.J., Derrien, A., Aumaitre, A., Guilbot, A. (1973). Degradation in vitro del’amidon par le suc pancreatique. Starch 25, 56–64.
Gallant, D.J., Bouchet, B., Baldwin, P.M. (1997). Microscopy of starch: evidence of a newlevel of granule organization. Carbohydrate Polymers 32, 177–191.
106 Resistant Starch

CH05GR 07/29/2013 16:2:53 Page 107
Gidley, M.J., Bulpin, P.V. (1987). Crystallization of malto-oligosaccharides as models ofthe crystalline form of starch: minimum chain-length requirement for the formation ofdouble helices. Carbohydrate Research 161, 291–300.
Godet, M.C., Bizot, H., Buleon, A. (1995). Crystallization of amylose-fatty acid complexesprepared with different amylose chain lengths. Carbohydrate Polymers 27, 47–52.
Hasjim, J., Lee, S.-O., Hendrich, S., Setiawan, S., Ai, Y., Jane, J. (2010). Characterizationof a novel resistant starch and its effects on postprandial plasma glucose and insulinresponses. Cereal Chemistry 87, 257–262.
Heinemann, C., Escher, F., Conde-Petit, B. (2003). Structural features of starch-lactoneinclusion complexes in aqueous potato starch dispersions: the role of amylose andamylopectin. Carbohydrate Polymers 51, 159–168.
Imberty A., Chanzy, H., Perez, S., Buleon, A., Tran, V. (1987). New three-dimensionalstructure for A-type starch. Macromolecules 20, 2634–2636.
Imberty A., Chanzy, H., Perez, S. (1988). The double-helical nature of the crystallinestructure for A-starch. Journal of Molecular Biology 201, 365–378.
Imberty A., Perez, S. (1988). A revisit to the three-dimensional structure of B-type starch.Biopolymers 27, 1205–1221.
Imberty, A., Buleon, A., Tran, V., Perez, S. (1991). Recent advances in knowledge of starchstructure. Starch 43, 375–384.
Immel, S., Lichtenthaler, F.W. (2000). The hydrophobic topographies of amylose and itsblue iodine complex. Starch 1, 1–8.
Jacobs, H, Delcour, J.A. (1998). Hydrothermal modifications of granular starch withretention of granular structure: a review. Journal of Agricultural and Food Chemistry46, 2895–2905.
Jenkins, P.J., Cameron, R.E., Donald, A.M. (1993). A universal feature in the structure ofstarch granules from different botanical sources. Starch 45, 417–420.
Le Bail, P., Bizot, H., Pontoire, B., Buleon, A. (1995). Polymorphic transitions ofamylose-ethanol crystalline complexes induced by moisture exchanges. Starch 47,229–232.
Mazur, A.K., Nakatani, H. (1993). Multiple attack mechanism in the porcine pancreaticalpha-amylase hydrolysis of amylose and amylopectin. Archives of Biochemistry andBiophysics 306, 29–38.
Moore, C.O., Tuschhoff, J.V., Hastings, C.W., Schanefelt, R.V. (1984). Applications ofstarches in foods. In: Whistler, R.L., BeMiller, J.N., Paschall, E.F. (eds). Starch:Chemistry and Technology, 575–590. New York, Academic Press, Inc.
Mueller, J.J., Gernat, C., Schulz, W., Mueller, E.-C. (1995). Computer simulations of x-rayscattering curves: gelation and crystallization process in amylose solutions. Biopolymers35, 271–288.
Nichols, B. L, Avery, S.E., Sen, P., Swallow, D.M., Hanh, D., Sterchi, E.E. (2003). Themaltase-glucoamylase gene: common ancestry to sucrase-isomaltase with complemen-tary starch digestion activities. Proceeding of the National Academy of Sciences in theUnited States of America 100, 1432–1437.
O’Dell, J. (1979). Starch. In: Blanshard, J.M.V, Mitchell, J.R. (eds). Polysaccharides inFoods, 171. London, Butterworths.
Pencek, R.R., Koyama, Y., Lacey, D.B., James, F.D., Fueger, P.T., Jabbour, K., Wiliam, P.E., Wasserman, D.H. (2002). Transporter-mediated absorption is the primary route ofentry and is required for passive absorption of intestinal glucose into the blood ofconscious dogs. Journal of Nutrition 132, 1929–1934.
Digestion Resistant Carbohydrates 107

CH05GR 07/29/2013 16:2:53 Page 108
Rappenecker, G., Zugenmaier, P. (2001). Conformation and packing analysis ofpolysaccharides and derivatives. 8. Detailed refinement of the crystal structure ofVh-amylose. Carbohydrate Research 89, 11–19.
Robyt, J.F. (1984). Enzymes in the hydrolysis and synthesis of starch. In: Whistler, R.L.,BeMiller, J.N., Paschall, E.F. (eds). Starch: Chemistry and Technology, 87–118.New York, Academic Press, Inc.
Seigner C., Provanov, E., Marchismouren, G. (1985). On porcine pancreatic alpha-amylase action – kinetic evidence for the binding of 2 maltooligosaccharidemolecules (maltose, maltotriose and orthonitrophenylmaltoside) by inhibition studies– correlation with the 5-subsite energy profile. European Journal of Biochemistry 148,161–168.
Shannon, J.C., Garwood, D.L. (1984). Genetics and physiology of starch development. In:Whistler, R.L., BeMiller, J.N., Paschall, E.F. (eds). Starch: Chemistry and Technology,26–79. New York, Academic Press, Inc.
Sievert, C., Pomeranz, Y. (1990). Enzyme-resistant starch. II. Differential scanningcalorimetry study on heat-treated starches and enzyme-resistant starch residues. CerealChemistry. 68, 86–91.
Singh, J., Kaur, L., McCarthy, O.J. (2007). Factors influencing the physio-chemical,morphological, thermal and rheological properties of some chemically modified starchesfor food applications – a review. Food Hydrocolloids 21, 1–22.
Stute, R. (1992). Hydrothermal modification of starches: the difference between annealingand heat-moisture treatment. Starch 44, 205–214.
Swallow, D.M., Poulter, M., Hollox, E.J. (2001). Intolerance to lactose and other dietarysugars. Drug Metabolism and Disposition 29, 513–516.
Tester, R.F., Karkalas, J., Qi, X. (2004). Starch – composition, fine structure andarchitecture. Journal of Cereal Science 39, 151–165.
US2006/0188631 A1. (2005). Pregelatinized chemically modified resistant starch prod-ucts and uses thereof. Woo, K., Bassi, S.D., Maningat, C.C., Zhao, L., Trompeter, E.,Kelley, G.A., Ranjan, S., Gaul, J., Dohl, C.T., DeMeritt, G.K., Stempien, G.J., Krehbiel,K.D.
US5593503 Process for producing amylase resistant granular starch. Shi, Y.-C., Trzasko,P.T.
US5281276 Process for making amylase resistant starch from high amylose starch. Chiu,C.-W., Henley, M., Altieri, P.
US7189288 Enzyme-resistant starch and method for its production. Stanley, K.D.,Stanley, E.D., Richmond, P.A., Yackel, W.C., Harris, D.W., Eilers, T.A., Marion, E.A.
US7674897 Production of Crystalline Short Chain Amylose. Norman, B., Pedersen, S.,Stanley, K.D., Stanley, E.D., Richmond, P.A.
Whittam, M.A., Noel, T.R., Ring, S.G. (1990). Melting behaviour of A- andB-type crystalline starch. International Journal of Biological Macromolecules 12,359–362.
Wu, H.C.H., Sarko, A. (1978a). Packing analysis of carbohydrates and polysaccharides. 8.Double helical molecular-structure of crystalline B-amylose. Carbohydrate Research61, 7–25.
Wu, H.C.H., Sarko, A. (1978b). Packing analysis of carbohydrates and polysaccharides. 9.Double helical molecular-structure of crystalline A-amylose. Carbohydrate Research61, 27–40.
108 Resistant Starch

CH05GR 07/29/2013 16:2:53 Page 109
Wu, H.C.H., Sarko, A. (1978c). Crystal-structures of A-polymorphs, B-polymorphs andC-polymorphs of amylose and starch. Carbohydrate Research 61, 73–78.
Wurzburg, O.B. (1986). Modified starches: Properties and uses, 33–34. CRC Press.Zobel, H.F. (1988). Molecules to granules: A comprehensive starch review. Starch 40,44–50.
Digestion Resistant Carbohydrates 109

CH05GR 07/29/2013 16:2:53 Page 110

3GCH06 07/20/2013 8:2:37 Page 111
6 Slowly Digestible Starch and HealthBenefits
Genyi Zhang1and Bruce R. Hamaker2
1 School of Food Science and Technology, Jiangnan University, China2Whistler Center for Carbohydrate Research and Department of FoodScience, Purdue University, USA
6.1 INTRODUCTION
Starch is the major glycaemic carbohydrate of foods, and its nutritional
property is related to its rate and extent of digestion and glucose absorption
in the small intestine. A classification of starch into rapidly digestible starch
(RDS), slowly digestible starch (SDS) and resistant starch (RS) has been used
to specify the nutritional quality of starch. SDS is the most elusive fraction of
the three, perhaps due to its somewhat transient nature, and it is certainly less
studied than RS. Yet, the physiological consequences of SDS may be just as
profound as those of RS, in terms of regulating glucose delivery to the body
and its associated metabolic effects, triggering gut hormones to affect
gastrointestinal tract motility and, perhaps, even reducing appetite and
increasing satiety. This chapter covers the current understanding of SDS,
including its concept and potential health benefits, the starch digestion
process, structure and mechanism of SDS and approaches to making SDS.
On one level, available carbohydrates are an important part of the diet and
should provide 45–65% of total caloric intake as recommended in the Dietary
Guidelines for Americans (U.S. Department of Agriculture, 2010). However,
quantity alone is not an accurate assessment of nutritive quality and rate – the
extent of digestion and absorption of glycaemic carbohydrates is an important
parameter (Jenkins et al., 1981; Englyst et al., 2003).
Over thirty years ago, Jenkins et al. (1981) proposed the concept of
glycaemic index (GI), which is represented by area under the in vivo
glycaemic response curve (AUC) of a tested food compared to a reference
glucose or white bread containing the same amount (50 g) of available
111
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH06 07/20/2013 8:2:37 Page 112
carbohydrate. Glycaemic response is a broader concept that includes the
nature and shape of the postprandial blood glucose profile. The digestion
property of glycaemic carbohydrates is the main determinant of the GI value
of different foods, although factors that affect glucose transport through the
gut enterocytes, as well as other human factors, also impact rate of rise of
blood glucose levels (Wolever et al., 2006).
Brand et al. (1991) categorized foods based on their GI values as high GI
(GI> 70), low GI (GI< 55) and intermediate GI foods. Since the advent of the
GI concept, there have been extensive clinical and nutritional investigations
on dietary carbohydrates, particularly regarding potential health benefits of
low-GI foods (Wolever, 2003), and an extensive list of different foods have
been tested and compiled by Foster-Powell et al. (2002).
There exists a controversy that has developed within both scientific and
industrial groups regarding the use of the GI concept in the context of a tool to
affect public health. This is in large part due to a body of studies with varying
results on the relationship between low-GI food and health (Howlett &
Ashwell, 2008), even though this includes carefully conducted studies with
mechanistic underpinnings. Slowly digestible glycaemic carbohydrates is one
way to achieve low glycaemic response in foods, but it is not the only way. A
better understanding of the structural and mechanistic basis of low glycaemic
response foods will provide important information to this field of study and to
the development of low glycaemic response products.
6.2 SDS AND POTENTIAL BENEFICIALHEALTH EFFECTS
Slowly digestible starch offers the possibility of moderated glucose delivery to
the body, which can be beneficial to glucose homeostasis and related
processes, as well as distal small intestinal glucose release that may affect
motility of food through the gastrointestinal tract and, perhaps, fullness and
satiety. Due to its importance as the central metabolic fuel, levels of glucose
are tightly controlled in the body through a series of mechanisms. Although
glucose may be supplied internally through glycogenolysis and gluco-
neogenesis, dietary derived glucose is critical for growth and development
and for energy-consuming organs such as the brain and central nervous system
(Peters et al., 2004).
Glucose is also a signal molecule in energy metabolism that stimulates
insulin secretion, governs glucose utilization and inhibits endogenous
glucose production. Starch, generally being the most abundant glycaemic
112 Resistant Starch

3GCH06 07/20/2013 8:2:37 Page 113
carbohydrate consumed, is themajor source of dietary glucose. Importantly, the
manner of digestion of starch and subsequent release of glucose in the digestive
system is the starting-point of the glucose homeostasis regulatory cascade, and
its timing and location of release controls glucose delivery of foods.
6.2.1 Potential health benefit of SDS relative to RDS
The nutritional property of starch has been ascribed to relative amounts of RDS,
SDSandRS.RDS results in a largefluctuation inbloodglucose that can generate
stress on the regulatory system of glucose homeostasis (Ludwig, 2002), and this
may even be associated with cell, tissue and organ damages under long-term
stress (Brownlee, 2001). In commonly consumed starchy foods that are proc-
essed, RDS is usually dominant. High RDS correlates with high value of GI and
glycaemic load (GL, GI� amount). Moreover, the high postprandial glycaemic
response elicited by RDS coincides with high levels of plasma insulin, the
principal hormone to regulate postprandial blood glucose levels.
RS, that starch which escapes digestion by host enzymes, is ascribed to a
number of positive physiological effects and benefit to the colon microbiota
(Annison & Topping, 1994; Ferguson et al., 2000; Topping et al., 2008). Most
of the health benefits ascribed to SDS are deduced from low-GI foods which
may have a similar glycaemic response as SDS. However, not all low-GI foods
have a slow digesting component. The property of SDS has not been studied
clinically to a significant degree.
When considering possible physiological effect/differences between
chronic consumption of foods high in RDS versus SDS, one might first
look at the level of the gut itself. RDS releases glucose quickly after leaving
the pyloric valve of the stomach, and is absorbed in the duodenum or proximal
jejunum. The exact fate of SDS is somewhat less clear. While most SDS-
containing foods still have a rapidly digesting component, certainly some
portion of the starch or its intermediate degradation products is digested and
releases glucose into the distal jejunum and ileum.
Such differences in starch digestion rate have, in recent years, been
documented to cause changes at the level of the gut enterocytes in terms
of gene expression that lead to changes in levels of the mucosal a-glucosi-dases (Mochizuki et al., 2010a, 2010b), glucose transporters (Shimada et al.,
2009) and gut hormones (Wachters-Hagedoorn et al., 2006; Shimada et al.,
2008), all of which could affect human physiology and health.
Seal et al. (2003) showed that,when fed to human subjects, normal uncooked
corn starch,which is a good source of SDS, resulted in a slowandprolonged (�5
hours) postprandial release of glucose, as well as low insulin levels throughout
Slowly Digestible Starch and Health Benefits 113

3GCH06 07/20/2013 8:2:37 Page 114
the postprandial phase (Figure 6.1). Unlike RDS, there was a lack of a
hypoglycaemic episode with SDS ingestion and there was a shift in peak blood
glucose to a longer time. SDS consumption also extended exogenous glucose
oxidation and resulted in a lower level of plasma-free fatty acids.
In the study byWachters-Hagedoorn et al. (2006), ingestion of normal corn
starch resulted in elevated and prolonged increase in the incretin hormones
glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic poly-
peptide (GIP) – the former up to 300 minutes postprandial. Apart from its
insulin stimulation role, GLP-1 has been implicated in decreasing gastric
emptying and improvement in insulin sensitivity and glycaemic response in
diabetics. This suggests that slowly digesting carbohydrates (SDC) in general
may be beneficial on energy homeostasis and regulation. SDC may have
additional benefits in areas of cognitive/mental performance (Benton et al.,
2003; Lang et al., 2003) and satiety (Sparti et al., 2000; Araya and Alvi~na,2004). The review of Lehmann & Robin (2007) discuss these other areas of
possible benefit.
Thus, SDS is associated with positive health outcomes that may include
moderated postprandial glycaemia, circulating free fatty acids and oxidative
stress (Jenkins et al., 2002). Harbis et al. (2004) showed that slowly available
glucose, the in vivo counterpart to SDS, when provided to obese and insulin-
resistant individuals, resulted in some improved metabolic profiles, including
lower postprandial insulinaemia and circulating triacylglycerols. Moreover,
high levels of SDS-containing legumes are considered beneficial to diabetic
control (McCrory et al., 2010), and it has been shown that a bedtime meal
containing SDS (corn starch) improved glucose tolerance for the second
Figure 6.1 Depiction of slowly digesting side-by-side mechanism and resultinginside-out digestion pattern (Zhang et al., 2006a). Reprinted with permissionfrom Zhang et al., 2006a. Copyright 2006 American Chemical Society.
114 Resistant Starch

3GCH06 07/20/2013 8:2:38 Page 115
morning meal (Axelsen et al., 1999). SDS-containing foods may be a dietary
intervention to curb or delay the prevalence of metabolic syndrome and
related diseases such as cardiovascular disease and diabetes (Cook et al.,
2003; Hu et al., 2004; Giugliano et al., 2006).
6.3 THE PROCESS OF STARCH DIGESTION
Starch is digested in the mammalian gastrointestinal tract first by salivary and
pancreatic a-amylases which produce small linear and branched a-limit
dextrins, which are further digested to glucose by the a-glucosidases,maltase-glucoamylase (MGAM) and sucrase-isomaltase (SI) at the mucosal
brush border of the small intestines (Swallow et al., 2001; Nichols et al., 2003).
The resulting glucose is then absorbed by the sodium-glucose active transporter
(SGLT-1) found in the apicalmembrane of the enterocytes (Pencek et al., 2002).
This is a very efficient system of reducing starch to glucose, though the rate can
be influenced by a number of intrinsic factors related to the starchy food (food
matrix, starch molecular form (dispersed, retrograded, crystalline), natural
enzyme inhibitors) (Tester et al., 2004).Gelatinized starches that are completely
dispersed are particularly susceptible to enzyme action.
A significant, though often overlooked, factor in determining the rate of
starch digestion and glucose absorption is the transit time of food through the
upper gastrointestinal tract. This is related principally to gastric emptying
time, which is controlled by gut hormones as triggered by macronutrients or
short chain fatty acids generated by fermentation of dietary fibre, including
resistant starch, as well as food form (solids vs. liquids, viscosity). Viscosity
itself can delay starch digestion by limiting access of enzymes to substrate. It
is possible that these factors can be used to generate a slow starch digestion
effect.
6.3.1 Enzyme action
Salivary and pancreatic a-amylases (E.C. 3.2.1.1) are a-1,4 endo-
glucosidases. It is generally considered that starch is only slightly digested
by salivary amylase, with the bulk of hydrolysis occurring by the action of
pancreatic a-amylase. There are five sub-sites for starch binding for human
a-amylase (Brayer et al., 2000), and cleavage of the a-1,4 glycosidic linkagesuses a multiple attack mechanism (Mazur & Nakatani, 1993) to produce
a-limit dextrins. On the other hand, the brush border a-glucosidases hydro-lyze both a-1,4 and a-1,6 glycosidic bonds and produce glucose. Regarding
kinetics of bond cleavage, a-1,6 glycosidic bonds hydrolyze more slowly
Slowly Digestible Starch and Health Benefits 115

3GCH06 07/20/2013 8:2:38 Page 116
compared to a-1,4 bonds (Kerr et al., 1951), and this approach has been usedto slow starch digestion rate (Ao et al., 2007).
While pancreatic a-amylase plays a pivotal role in starch digestion
(Englyst et al., 1992), the mucosal a-glucosidases are a recognized control
point for glucose generation and, consequently, are of equal importance in
strategies to slow glucose delivery from starch. Even the a-amylase degrada-
tion products of maltotriose, maltotetraose, and maltopentose have been
shown to have some inhibitory action on the most active of the four
a-glucosidases, the glucoamylase (Quezada-Calvillo et al., 2007, 2008).
Mucosal MGAM (E.C. 3.2.1.20 and 3.2.1.3, encoded by the gene MGAM,
located on chromosome 7q34) and SI (E.C. 3.2.1.48 and 3.2.1.10, encoded by
the gene SI, located on chromosome 3q26) (Nichols et al., 1998, 2003) are
complexes of two enzymes each that belong to the glucohydrolyase Family 31.
All four enzyme subunits have a-1,4 exo-glucosidic activity from the non-
reducing ends of maltooligosaccharides. Though commonly termed maltase,
glucoamylase, isomaltase and sucrase, these enzymes all have maltase activity
and other activities overlap into different subunits. This lack of specificity of
terminology has led to the use of terms associated with their location relative
to the anchor point on the brush border enterocytes. Thus, N-terminal MGAM
is maltase, C-terminal MGAM is glucoamylase, N-terminal SI is isomaltase
and C-terminal SI is sucrase. The main debranching enzyme is Nt-SI although
Nt-MGAM also has some isomaltase activity (Sim et al., 2010). Ct-MGAM
has higher hydrolytic property for a-glucans with longer chains (Sim et al.,
2008), and can even digest native starch amylopectin (Lin et al., 2012).
6.4 STRUCTURAL AND PHYSIOLOGICALFUNDAMENTALS OF SDS
SDS is not a defined entity of starch, as is RS with its subcategories of RS 1-4
types, and it is more of a concept than a specific material; it is an in vitro
measurement of starch digestion between 20 and 120 minutes when using the
enzymes and conditions as stated by Englyst et al. (1992). It corresponds to
the in vivo concept of slowly available glucose (SAG), although SAG can
come also from other slowly digestible glycaemic carbohydrates. Accord-
ingly, knowledge of starch structure and the various ways that SDS can be
generated is necessary to develop strategies for SDS product development. In
our view, there are four basic mechanisms for generating SDS:
1. physical or food matrix structures that decrease enzyme accessibility;
2. chemical structures that limit rate of hydrolysis;
116 Resistant Starch

3GCH06 07/20/2013 8:2:38 Page 117
3. other food factors that decrease digestion rate (e.g. viscosity, inhibitors);
4. physiological control of food motility (i.e. gastric emptying).
6.4.1 Physical or food matrix structures related to SDS
Processed foods, such as pasta, often have structures related to food matrices
that lead to SDS. These are generally where starch in the food is less
accessible due to other surrounding food components, such as the dense
protein matrix of pasta (Fardet et al., 1998) or hard-to-digest protein matrix in
sorghum porridges (Zhang & Hamaker, 1998).
It is raw starch itself, with its natural semi-crystalline granular structure
comprised of concentric layers of amorphous and crystalline regions, that is
perhaps the best example of SDS physical structure. Amylopectin fine
structure differences form the basis of the semi-crystalline A, B, and C
x-ray diffraction patterns (Thompson, 2000). Due to a combination of
crystallite packing and accessibility of the granule to enzymes afforded by
surface pores leading to interior channels, raw cereal starches naturally have a
high amount of SDS material (Seal et al., 2003).
G�erard et al. (2001), Jane et al. (1997), and Planchot et al. (1997) have
shown that B-type tuber starches are resistant to enzyme digestion, while
A-type cereal starches are mostly slowly digestible. While amorphous
regions of the starch granule have been thought to be more easily digested
than crystalline regions (Gallant et al., 1992), Zhang et al. (2006a, 2006b)
revealed that degree of crystallinity did not change after amylolysis,
indicating that amorphous and crystalline regions are digested simulta-
neously. The proposed side-by-side digestion mechanism starts from the
channels (see Figure 6.1) and proceeds to the observed inside-out digestion
pattern. Apparently, this is caused by tight linkages between amorphous and
crystalline layers. The layer-by-layer crystalline (A-type) and amorphous
regions are the structural basis for the slow digestion property of native
cereal starches, providing a strategy for developing starch-based structures
that are slowly digestible.
As mentioned, supramolecular level food matrices, such as exist in cooked
pasta with protein networks, may cause slow starch digestion due to limited
accessibility of starch-degrading enzymes to starch (Fardet et al., 1998). To
better control starch digestion rates for experimental purposes, we made
starch-entrapped microspheres that consist of starch in a quick-setting gel
(sodium alginate in calcium chloride) (Venkatachalam et al., 2009; Figure
6.2). The porous polysaccharide structure permits entrance of amylase to
gelatinized starch and the rate of digestion is controlled by pore size. This
provides a strategy to achieve SDS.
Slowly Digestible Starch and Health Benefits 117
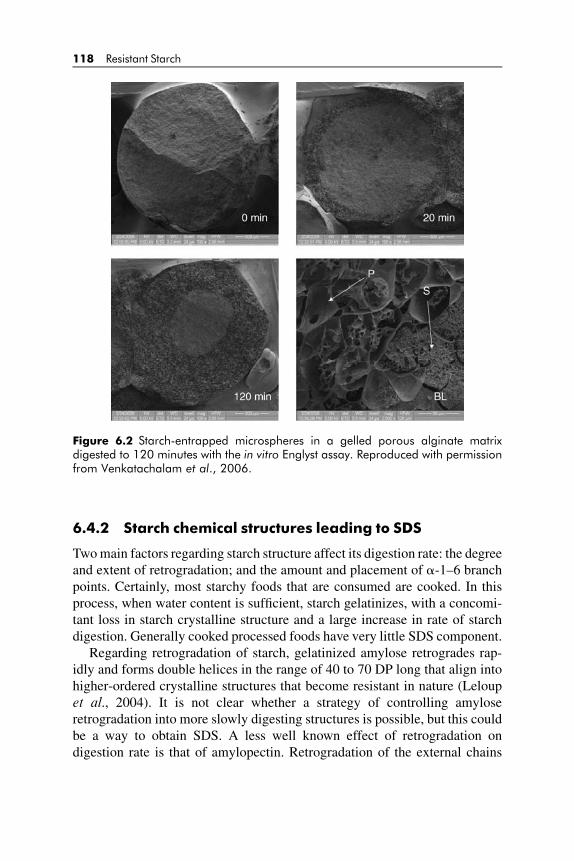
3GCH06 07/20/2013 8:2:38 Page 118
6.4.2 Starch chemical structures leading to SDS
Twomain factors regarding starch structure affect its digestion rate: the degree
and extent of retrogradation; and the amount and placement of a-1–6 branch
points. Certainly, most starchy foods that are consumed are cooked. In this
process, when water content is sufficient, starch gelatinizes, with a concomi-
tant loss in starch crystalline structure and a large increase in rate of starch
digestion. Generally cooked processed foods have very little SDS component.
Regarding retrogradation of starch, gelatinized amylose retrogrades rap-
idly and forms double helices in the range of 40 to 70 DP long that align into
higher-ordered crystalline structures that become resistant in nature (Leloup
et al., 2004). It is not clear whether a strategy of controlling amylose
retrogradation into more slowly digesting structures is possible, but this could
be a way to obtain SDS. A less well known effect of retrogradation on
digestion rate is that of amylopectin. Retrogradation of the external chains
Figure 6.2 Starch-entrapped microspheres in a gelled porous alginate matrixdigested to 120 minutes with the in vitro Englyst assay. Reproduced with permissionfrom Venkatachalam et al., 2006.
118 Resistant Starch

3GCH06 07/20/2013 8:2:38 Page 119
slows rate of digestion (Zhang et al., 2008b). Along this line, retrogradation of
partially debranched amylopectin after isoamylase treatment has been used to
make SDS (Shi et al., 2003; Shin et al., 2004).
In our own investigations (Zhang et al., 2008a, 2008b), we showed that
amylopectin fine structure differences can drive the SDS amount. Using a
range of maize starch mutants, two paths towards increase were found,
represented by a parabolic relationship between SDS content and the weight
ratio of amylopectin short chains (DP< 13) to long chains (DP� 13)
(Figure 6.3). Amylopectin with either a high amount of short chains (higher
branch points and shorter external chains), or a higher proportion of long
internal chains, had a higher content of SDS (Figure 6.3).
Two mechanisms were found to drive slow digestion of gelatinized
starches: inherently high branch density somewhat slows digestion due to
the slower kinetics associated with digestion of a-1,6 linkages, while high
long chains drives faster retrogradation, as has been noted by others (Klucinec
& Thompson, 1999; Matalanis et al., 2009). In the latter retrograded type, the
SDS property appeared to be transient in nature, as increased storage time
converted some SDS to RS (Zhang et al., 2008b). Regarding the effect of
branched density, another a-glucan structure with high a-1,6 linkages, thoughnot in a branched structure like amylopectin, is pullulan, and digestion was
likewise affected (Wolf et al., 2003). Thus, amylopectin seems associated
with the SDS property.
R2 = 0.7147
Group I Group II
SF/LF weight ratio
SD
S (
%)
25
20
15
10
5
01 0.8 0.6 0.4 0.2 0
Figure 6.3 A parabolic relationship between SDS percentage and the weight ratio ofshort-chain (SF) to long-chain fraction (LF) (Zhang et al., 2008a). Reprinted withpermission from Zhang et al., 2008a. Copyright 2008 American Chemical Society.
Slowly Digestible Starch and Health Benefits 119

3GCH06 07/20/2013 8:2:38 Page 120
6.4.3 Other food factors that decrease digestion rate
Much focus on decreasing starch digestion rate has been on food viscosity, as
well as a-amylase and a-glucosidase natural inhibitors, and other factors suchas organic acids. These have been associated mostly with changes in gly-
caemic response or GI.
Water-soluble polysaccharides that impart significant viscosity effect have
been shown to result in slower digestion and absorption of glycaemic
carbohydrates, and also reduced postprandial glycaemic and insulinaemic
responses (Jenkins et al., 1976, 1977). Common fibres in this category include
b-glucans, psyllium and guar gum, and these have been shown to moderate
glycaemic response even when incorporated into food products (Wood et al.,
1990; Yokayama et al., 1997; Jenkins et al., 2002). At a mechanistic level,
viscosity appears to cause a slower gastric emptying as well as lower
accessibility of enzymes to substrates (Lecl�ere et al., 1994; Slavin, 2005),
accompanied by a reduced diffusion rate of digested carbohydrates to the
mucosal surface (Briani et al., 1987).
Natural a-amylase and a-glucosidase inhibitors can moderate starch
digestion to blunt both glycaemic and insulinaemic responses. Strong inhibi-
tion can result in an increase in resistant starch, though partial inhibition can
produce a slowly digestible effect. Acarbose, a modified tetrasaccharide
formed by the genus Actinoplanes, is one of the best studied of such inhibitors,
with most specific inhibition of the C-terminal a-glucosidases (Hiele et al.,
1992; Eskandari et al., 2011). Due to its blunting of postprandial glycaemic
response, it has been used in treatment of diabetes (Chiasson et al., 1994;
Conniff et al., 1994). A well-studied a-amylase inhibitor comes from Great
Northern beans (phaseolamin) and has been shown to promote glucose
homeostasis (Boivin et al., 1988).
There is a significant literature on natural inhibitors of both a-amylase and
a-glucosidases. A number of phenolic compounds, such as those found in green
tea extract (Konishi et al., 2006), inhibit enzyme activity and lower glycaemic
response. As a strategy to slow glycaemic carbohydrate digestion and/or absorp-
tion, use of natural inhibitors in foods necessitates more study to determine
physiologically relevant levels and other potential unintended consequences.
Lower glycaemic and insulinaemic responses have been noted with
consumption of organic acids, such as found in sourdough fermentation
(Liljeberg and Bj€orck, 1996, 1998; Liljeberg et al., 1995). For example,
sodium propionate added to bread decreased postprandial blood glucose and
insulin responses and also increased satiety compared to control breads. This
was attributed to delayed gastric emptying. Vinegar added to a starchy meal
had a similar effect (Liljeberg and Bj€orck, 1998).
120 Resistant Starch

3GCH06 07/20/2013 8:2:38 Page 121
6.4.4 Physiological control of foodmotility
Slow digestion of starch can perhaps be best achieved by control of transit
time of food through the upper gastrointestinal tract, and this is mainly
through control of the rate of gastric emptying. Both the ileal break and the
colonic break have the effect of slowing gastric emptying. The former relates
to sensing of macronutrients (fat, protein, carbohydrate) in the ileum, through
triggering of gut hormones and the nervous system that causes longer
retention of food in the stomach (Van Citters & Lin, 2006). The colonic
break relates mainly to dietary fibre, including RS, fermentation that also
triggers the gut hormone peptide YYand, perhaps, GLP-1, which slows gastric
emptying (Massimino et al., 1998; Cuche et al., 2000).
Starch that is portioned out of the stomach to the small intestine over an
extended period of time is, in essence, a ‘slowly digestible starch’. Dietary fibre
fermentation also can cause the ‘second meal effect’, as noted by Jenkins et al.
(1982), Wolever et al. (1988) and Brighenti et al. (2006), which moderates
glycaemic and insulinaemic responses of the subsequent meal. Resistant starch
has specifically been shown to have this effect (Bj€orck et al., 2000).
6.5 APPLICATION-ORIENTED STRATEGIESTOMAKE SDS
Currently, there are very few approaches to make commercially viable SDS.
Practically, at this time strategies can be reduced to two areas: starch
molecular structures that reduce digestion rate; and food matrices with
slow starch digestion. A central question that remains, however, is what
degree of slow digestion effect, and what amount consumed, is necessary to
obtain a desired physiological, health-related outcome. This is one of the main
unanswered questions that impedes SDS development of commercial
products.
6.5.1 Starch-based ingredients
As mentioned above, starch amylopectin structures with high branching
and short external chains, or with a high proportion of long chains, have
higher SDS contents. Additionally, enzymatic, physical, and chemical
modifications of starch have been attempted, with the aim of decreasing
digestion rate.
Modification of starch molecular structures through genetic manipulation
of synthesizing enzymes, or through enzyme modification by processing,
Slowly Digestible Starch and Health Benefits 121

3GCH06 07/20/2013 8:2:38 Page 122
is an approach with good potential for the creation of SDS products. Main
strategies would be to produce highly branched structures with short external
chains or structures that retrograde to slowly digestible crystallites. b-amylase
has been used to shorten non-reducing external chains of amylopectin to
increase branch density and branching enzymes to add branches (Hamaker &
Han, 2006; Ao et al., 2007). The patent of Backer & Saniez (2005) showed a
method to produce a highly branched soluble a-glucan using an enzymatic
approach. Shi et al. (2003), in their patent, showed a method of making
retrograded debranched linear chains that are slowly digestible.
Physical modification of starch to slow starch digestion rate has been
achieved by heat-moisture treatment, temperature cycling and storage condi-
tions. Hydrothermal treatment of sweet potato starch resulted in A-type
crystalline structure that coincided with a large increase in SDS over native
granules (Shin et al., 2004). Using waxy potato starch, Lee et al. (2011)
increased SDS content to 42% by heat-moisture treatment and gave a
favourable glycaemic response in mice. Temperature-cycling retrogradation
of waxy rice starch also resulted in a high SDS value of 52% (Zhang et al.,
2011).
Regarding chemical modification, digestion properties of starch can be
changed through the addition of functional groups to the linear chains of
starch. Many modification methods make some degree of RS, such as
oxidation, acetylation, cross-linking and etherification (Wolf et al., 1999).
Octenyl succinic anhydride (OSA) esterification, in particular, appears to
increase SDS content (Han & BeMiller, 2007), and Wolf et al. (2001) found
modified in vivo glycaemic response of OSA-modified starch. OSA-modified
starch molecules slow enzyme digestion through an uncompetitive inhibitor
mechanism (He et al., 2008).
6.5.2 SDS generation in a food matrix
Another approach to making SDS products is through use of food processing
technologies to create matrices that slow access to starch and to limit the
extent of starch gelatinization, which also slows digestion. Dense food
matrices, such as dense pastas, are normally associated with SDS, though
the interactions of specific food components with starch can also slow starch
digestion. The most recognized interaction is the inclusion complex between
amylose and lipid.
Lower digestibility has been noted in foods that have amylose-
monoglyceride complexes (Holm et al., 1983; Seneviratne & Biliaderis,
1991; Murray et al., 1998). Saturated monoglycerides with long aliphatic
tails cause slower digestion (Eliasson & Krog, 1985). Patil et al. (1998)
122 Resistant Starch

3GCH06 07/20/2013 8:2:39 Page 123
showed that amylose-lipid complexes mixed in the diet of dogs somewhat
lowered glycaemic and insulinaemic responses. Starch conjugated through the
Maillard reaction with protein or amino acids has also been shown to reduce
digestibility (Yang et al., 1998). Legume or pulses, particularly when cooked
as a whole grain, have a comparably high content of SDS that is at least
partly due to physical entrapment in cell walls (Englyst et al., 1996; McCrory
et al., 2010).
6.6 CONSIDERATIONS
There is growing evidence that the rate and location of starch digestion in the
upper gastrointestinal tract has physiological and metabolic consequences that
relate to health. This applies not only for starch, which is the major glycaemic
carbohydrate in most diets, but also for other slowly digestible carbohydrates.
While SDS is distinctly different from RS in that it is digestible and generates
glucose directly for absorption in the small intestine, starch digestion rate is
impacted by RS and its effect on gastric emptying. Additionally, glucose
release in the ileum of the small intestine also likely slows gastric emptying.
When considering the wide range of time of postprandial digestion of starches
in foods, perhaps control of gastric emptying has the largest single effect on
rate of starch digestion, and SDS may itself lengthen emptying time through
feedback mechanisms.
However, even considering that some methods exist to make SDS ingredi-
ents or foods, there is a lack of knowledge as to the amount and manner of
delivery that is needed to produce a desired effect. Further understanding of
consequences of SDS that might include energy balance, metabolic diseases,
cognitive and physical performance and satiety, are needed. New technologies
also must be identified to make robust and more defined SDS materials for use
in the food industry.
REFERENCES
Annison, G., Topping D.L. (1994). Nutritional role of resistant starch: chemical structurevs. physiological function. Annual Review of Nutrition 14, 297–320.
Ao, Z., Simsek, S., Zhang, G., Venkatachalam, M., Reuhs, B., Hamaker. B.R. (2007).Starch with slow digestion property produced by altering branch density, chain lengthand crystalline structure. Journal of Agricultural and Food Chemistry 55, 4540–4547.
Araya, H., Alvi~na, M. (2004). Rapid carbohydrate digestion rate produced lesser short-term satiety in obese preschool children. European Journal of Clinical Nutrition 58,637–642.
Slowly Digestible Starch and Health Benefits 123

3GCH06 07/20/2013 8:2:39 Page 124
Axelsen, M., Arvidsson, L.R., Lonnroth, P., Smith, U. (1999). Breakfast glycaemicresponse in patients with type 2 diabetes: effects of bedtime dietary carbohydrates.European Journal of Clinical Nutrition 53, 706–710.
Backer, D., Saniez, M.H. (2005). Soluble highly branched glucose polymers and theirmethod of production. US Patent 2005/0142167A1.
Benton, D., Ruffin, M., Lassel, T., Nabb, S., Messaoudi, M., Vinoy, S., Desor, D., Lang, V.(2003). The delivery rate of dietary carbohydrates affects cognitive performance in bothrats and humans. Psychopharmacology 166, 86–90.
Bj€orck, I., Liljeberg, H., Ostman, E. (2000). Low glycaemic-index foods. British Journal ofNutrition 83, S149–55.
Boivin, M., Flourie, B. Rizza, R.A., Go, V.L., DiMagno, E.P. (1988). Gastrointestinaland metabolic effects of amylase inhibition in diabetics. Gastroenterology 94,387–394.
Brand, J.C., Colagiuri, S., Crossman, S., Allen, A., Roberts, D.C., Truswell, A.S. (1991).Low-glycemic index foods improve long-term glycemic control in NIDDM. DiabetesCare 14, 95–101.
Brayer, G.D., Sidhu, G., Maurus, R., Rydberg, E.H., Braun, C., Wang, Y., Nguyen, N.,Overall, C.M., Withers, S.G. (2000). Subsite mapping of the human pancreatica-amylase active site through structural, kinetic, and mutagenesis techniques.Biochemistry 39, 4778–4791.
Briani, G., Bruttomesso, D., Bilardo, G., Giorato, C., Duner, E., Iori, E., Sgnaolin,E., Pedrini, P., Tiengo, A. (1987). Guar-enriched pasta and guar gum in the dietarytreatment of type II diabetes. Phytotherapy Research 1, 177–179.
Brighenti, F., Benini, L., Rio, D.D., Casiraghi, C., Pellagrini, N., Scazzina, F., Jenkins, D.J.,Vantini, I. (2006). Colonic fermentation of indigestible carbohydrate contributes to thesecond-meal effect. American Journal of Clinical Nutrition 83, 817–822.
Brownlee, M. (2001). Biochemistry and molecular cell biology of diabetic complications.Nature 414, 813–820.
Chiasson, J.L., Josse, R.G., Hunt, J.A., Palmason, C., Rodger, N.W., Ross, S.A., Ryan, E.A., Tan, M.H., Wolever, T.M. (1994). The efficacy of acarbose in the treatment ofpatients with non-insulin-dependent diabetes mellitus. A multicenter controlled clinicaltrial. Annals of Internal Medicine 121, 928–935.
Conniff, R.F., Shapiro, J.A., Seaton, T.B. (1994). Long-term efficacy and safety of acarbosein the treatment of obese subjects with non-insulin-dependent diabetes mellitus.Archives of Internal Medicine 154, 2442–2448.
Cook, S., Weitzman, M., Auinger, P., Nguyen, M., Dietz, W.H. (2003). Prevalence of ametabolic syndrome phenotype in adolescents. Archives of Pediatrics & AdolescentMedicine 157, 821–827.
Cuche, G., Cuber, J.C., Malbert, C.H. (2000). Ileal short-chain fatty acids inhibit gastricmotility by a humoral pathway. American Journal of Physiology. Gastrointestinal andLiver Physiology 279, G925–930.
Eliasson, A.C., Krog, N. (1985). Physical properties of amylose monoglyceride complexes.Journal of Cereal Science 3, 239–248.
Englyst, H.N., Kingman, S.M., Cummings, J.H. (1992). Classification and measurementof nutritionally important starch fractions. European Journal of Clinical Nutrition 46,S33–50.
Englyst, H.N., Kingman, S.M., Hudson, G., Cummings, J.H. (1996). Measurement ofresistant starch in vitro and in vivo. British Journal of Nutrition 75, 749–755.
124 Resistant Starch

3GCH06 07/20/2013 8:2:39 Page 125
Englyst, K.N., Vinoy, S., Englyst, H.N., Lang, V. (2003). Glycaemic index of cerealproducts explained by their rapidly and slowly available glucose. British Journal ofNutrition 89, 329–339.
Eskandari, R.K., Jones, K.R., Reddy, K., Jayakanthan, M., Chaudet, D.R. Rose, B.M. Pinto.(2011). Probing the intestinal a-glucosidase enzyme specificities of starch-digestingmaltase-glucoamylase and sucrase-isomaltase: synthesis and inhibitory properties of 30-and 50- maltose-extended de-o-sulfonated ponkoranol. Chemistry – A European Journal17, 14817–14825.
Fardet, A., Hoebler, C., Baldwin, P.M., Bouchet, B., Gallant, D.J., Barry, J.L. (1998).Involvement of the protein network in the in vitro degradation of starch from spaghettiand lasagne: a microscopic and enzymatic study. Journal of Cereal Science 27,133–145.
Ferguson, L.R., Tasman-Jones, C., Englyst, H., Harris, P.J. (2000). Comparative effects ofthree resistant starch preparations on transit time and short-chain fatty acid production inrats. Nutrition and Cancer 36, 230–237.
Foster-Powell, K., Holt, S.H., Brand-Miller, J.C. (2002). International table ofglycemic index and glycemic load values. American Journal of Clinical Nutrition76, 5–56.
Gallant, D.J., Bouchet, B., Bul�eon, A., Perez, S. (1992). Physical characteristics of starchgranules and susceptibility to enzymatic degradation. European Journal of ClinicalNutrition 46, S3–16.
G�erard, C., Colonna, P., Bul�eon, A., Planchot, V. (2001). Amylolysis of maize mutantstarches. Journal of the Science of Food and Agriculture 81, 1281–1287.
Giugliano, D., Ceriello, A., Esposito, K. (2006). The effects of diet on inflammation.Journal of the American College of Cardiology 48, 677–685.
Hamaker, B.R., Han, X. (2006). Slowly digestible starch. US Patent 20060257977.Han, J., BeMiller, J.N. (2007). Preparation and physical characteristics of slowly digestingmodified food starches. Carbohydrate Polymers 67, 366–374.
Harbis, A., Perdreau, S., Vincent-Baudry, S., Charbonnier, M., Bernard, M.C., Raccah, D.,Senft, M., Lorec, A.M., Defoort, C., Portugal, H., Vinoy, S., Lang, V., Lairon, D. (2004).Glycemic and insulinemic meal responses modulate postprandial hepatic and intestinallipoprotein accumulation in obese, insulin resistant subjects. American Journal ofClinical Nutrition 80, 896–902.
He, J., Liu, J., Zhang, G. (2008). Slowly digestible waxy maize starch prepared by octenylsuccinic anhydride esterification and heat-moisture treatment: glycemic response andmechanism. Biomacromolecules 9, 175–184.
Hiele, M., Ghoos, Y., Rutgeerts, P., Vantrappen, G. (1992). Effects of acarbose on starchhydrolysis. Study in healthy subjects, ileostomy patients, and in vitro.Digestive Diseasesand Sciences 37, 1057–1064.
Holm, J, Bj€orck, I., Ostrowska, S., Eliasson, A.C., Asp, N.G., Larsson, K., Lundquist, I.(1983). Digestibility of amylose-lipid complexes in vitro and in vivo. Starch - St€arke 35,294–297.
Howlett, J., Ashwell, M. (2008). Glycemic response and health: summary of a workshop.American Journal of Clinical Nutrition 87, 212S–6S.
Hu, G., Qiao, Q., Tuomilehto, J., Balkau, B., Borch-Johnsen, K., Pyorala, K. (2004).Prevalence of the metabolic syndrome and its relation to all-cause and cardiovascularmortality in nondiabetic European men and women. Archives of Internal Medicine164, 1066–1076.
Slowly Digestible Starch and Health Benefits 125

3GCH06 07/20/2013 8:2:39 Page 126
Jane, J.L., Wong, K.S., McPherson, A.E. (1997). Branch structure difference in starches ofA- and B-type x-ray patterns revealed by their Naegeli dextrin. Carbohydrate Research300, 219–227.
Jenkins, A.L., Jenkins, D.J.A., Zdravkovic, U., Wursch, O., Vuksan, V. (2002). Depressionof the glycemic index by high levels of b-glucan fiber in two functional foods tested intype 2 diabetes. European Journal of Clinical Nutrition 56, 622–628.
Jenkins, D.J., Wolever T.M., Taylor, R.H. (1982). Slow release dietary carbohydrateimproves second meal tolerance. American Journal of Clinical Nutrition 35, 1339–46.
Jenkins, D.J.A., Leeds A.R., Gassul, M.A., Wolever, T.M.S., Goff, D.V., Alberti,K.G.M.M., Hockaday, T.D.R. (1976). Unabsorbable carbohydrates and diabetes.Decreased post-prandial hyperglycemia. Lancet 2, 172–174.
Jenkins, D.J.A., Leeds R., Gassul, M.A., Cochet, R., Alberti, K.G.M.M. (1977). Decreasein post-prandial insulin and glucose concentrations by guar and pectin. Annals ofInternal Medicine 86, 20–23.
Jenkins, D.J.A., Kendall, C.W.C., Augustin, L.S.A., Franseschi, S., Hamaidi, M., Marchie,A., Wolever T.M.S., Taylor, R.H. (1981). Glycemic index of foods: a physiological basisfor carbohydrate exchange. American Journal of Clinical Nutrition 34, 362–366.
Jenkins, D.J.A., Kendall, C.W.C., Augustin, L.S.A., Franseschi, S., Hamaidi, M., Marchie,A., Axelsen, M. (2002). Glycemic index: overview of implications in health and disease.American Journal of Clinical Nutrition 76, 266S-273S.
Kerr, R.W., Cleveland, F.C., Katzbeck, W.J. (1951). The action of amylo-glucosidase onamylose and amylopectin. Journal of the American Chemical Society 73, 3916–3921.
Klucinec, J.D., Thompson, D.B. (1999). Amylose and amylopectin interact in retrograda-tion of dispersed high-amylose starches. Cereal Chemistry 76, 282–291.
Konishi, Y., Aitani, M., Yamaguchi, H., Arimura, T., Okada, T., Murai, H. (2006).Methodof treating diabetes and obesity using carbohydrate absorption inhibitors. US Patent20060024396.
Lang, V., Degouy, M., Champenois, Y. (2003). Use of a cereal product for improvingcognitive performance and mental well-being in a person, particularly in a child and anadolescent. US Patent 20030161861.
Lecl�ere, C.J., Champ, M., Boillot, J., Guille, G., Lecannu, G., Molis, C., Bornet, F.,Krempf, M., Delort-Laval, J., Galmiche, J.P. (1994). Role of viscous guar gums inlowering the glycemic response after a solid meal. American Journal of ClinicalNutrition 59, 914–921.
Lee, C.J., Kim, Y., Choi, S.J., Moon, T.W. (2011). Slowly digestible starch from heat-moisture treated waxy potato starch: Preparation, structural characteristics, and glucoseresponse in mice. Food Chemistry online, 10.1016/j.foodchem.2011.09.098.
Lehmann, U., Robin, F. (2007). Slowly digestible starch – its structure and healthimplications: A review. Trends in Food Science & Technology 18, 346–355.
Leloup, V.M., Colonna, P., Ring, S.G. (2004). a-amylase adsorption on starch crystallites.Biotech. Bioengineering 38, 127–134.
Liljeberg, H.G., Bj€orck, I. M. (1996). Delayed gastric emptying rate as a potentialmechanism for lowered glycaemia after eating sourdough bread: studies in humansand rats using test products with added organic acids or an organic salt. AmericanJournal of Clinical Nutrition 64, 886–93.
Liljeberg, H., Bj€orck, I. (1998). Delayed gastric emptying rate may explain improvedglycaemia in healthy subjects to a starchy meal with added vinegar. European Journal ofClinical Nutrition 52, 368–371.
126 Resistant Starch

3GCH06 07/20/2013 8:2:39 Page 127
Liljeberg, H.G., Lonner, C.H., Bj€orck, I.M. (1995). Sourdough fermentation or addition oforganic acids or corresponding salts to bread improves nutritional properties of starch inhealthy humans. Journal of Nutrition 125, 1503–1511.
Lin, A.H.M., Nichols, B.L., Ao, Z., Quezada-Calvillo, R., Avery, S.E., Sim, L., Rose, D.R.,Naim, H.Y., Hamaker, B.R. (2012). Unexpected high digestion rate of cooked starch bythe ct-maltase-glucoamylase small intestinal mucosal a-glucosidase subunit. PLoS One7(5), e35473.
Ludwig, D.S. (2002). The glycemic index physiological mechanism relating to obesity,diabetes, and cardiovascular disease. Journal of the American Medical Association 287,2414–2423.
Massimino, S.P., McBurney, M.I., Field, C.J., Thompson, A.B.R., Keelan, M., Hayek,M.G., Sunvoid, G.D. (1998). Fermentable dietary fiber increases GLP-1 secretion andimproves glucose homeostasis despite increased intestinal glucose transport capacity inhealthy dogs. Journal of Nutrition 128, 1786–1793.
Matalanis, A.M., Campanella, O.H. and Hamaker, B.R. (2009). Storage retrogradationbehavior of sorghum, maize and rice starch pastes related to amylopectin fine structure.Journal of Cereal Science 50, 74–81.
Mazur, A.K., Nakatani, H. (1993). Multiple attack mechanism in the porcine pancreaticalpha-amylase hydrolysis of amylose and amylopectin. Archives of Biochemistry andBiophysics 306, 29–38.
McCrory, M.A., Hamaker, B.R., Lovejoy, J.C., Eichelsdoerfer, P.E. (2010). Pulse con-sumption, satiety and weight management. Advances in Nutrition 1, 17–30.
Mochizuki, K., Hanai, E., Suruga, K., Kuranuki, S., Goda, T. (2010a). Changes ina-glucosidase activities along the jejunal-ileal axis of normal rats by the a-glucosidaseinhibitor miglitol. Metabolism 59, 1442–1447.
Mochizuki, K., Sato, Y., Takase, S., Goda, T. (2010b). Changes in mucosal a-glucosidaseactivities along the jejuna-ileal axis by Hm-HACS diet intake are associated withdecreased lipogenic enzyme activity in epididymal adipose tissue. Journal of Agricul-tural and Food Chemistry 58, 6923–6927.
Murray, S., Patil, A., Fahey, G., Merchen, N., Wolf, B., Lai, S.C., Gareb, K. (1998).Apparent digestibility of a debranched amylopectin-lipid complex and resistant starchincorporated into enteral formulas fed to ileal-cannulated dogs. Journal of Nutrition 128,2032–2035.
Nichols, B.L., Eldering, J., Avery, S., Hahn, D., Quaroni, A., Sterchi, E. (1998). Humansmall intestinal maltase-glucoamylase cDNA cloning. Homology to sucrase-isomaltase.Journal of Biological Chemistry 273, 3076–3081.
Nichols, B.L., Avery, S.E., Sen, P., Swallow, D.M., Hahn, D., Sterchi, E.E. (2003). Themaltase-glucoamylase gene: common ancestry to sucrase-isomaltase with complemen-tary starch digestion activities. Proceedings of The National Academy Of Sciences OfThe United States Of America 100, 1432–1437.
Patil, A.R., Murray, S.M., Fahey, G.C., Merchen, N.R., Wolf, B.W., Lai, C., Garleb, K.A.(1998). Apparent digestibility and glycemic responses to a debranched amylopectin-lipid complex and resistant starch incorporated into enteral formulas fed to dogscannulated in the ileum. FASEB Journal 12, A210 (abstract).
Pencek, R.R., Koyama, Y., Lacy, D.B., James, F.D., Fueger, P.T., Jabbour, K., William, P.E.,Wasserman, D.H. (2002). Transporter-mediated absorption is the primary route of entryand is required for passive absorption of intestinal glucose into the blood of consciousdogs. Journal of Nutrition 132, 1929–1934.
Slowly Digestible Starch and Health Benefits 127

3GCH06 07/20/2013 8:2:39 Page 128
Peters, A., Schweiger, U., Pellerin, L., Hubold, C., Oltmanns, K.M., Conrad, M., Schultes,B., Born, J., Fehm, H.L. (2004). The selfish brain: competition for energy resources.Neuroscience & Biobehavioral Reviews 28, 143–180.
Planchot, V., Colonna, P., Bul�eon, A. (1997). Enzymatic hydrolysis of a-glucan crystal-lites. Carbohydrate Research 298, 319–326.
Quezada-Calvillo, R., Robayo-Torres, C.C., Ao, Z., Hamaker, B.R., Quaroni, A., Brayer,G.D., Sterchi, E.E., Baker, S.S., Nichols, B.L. (2007). Luminal substrate ‘brake’ onmucosal maltase-glucoamylase activity regulates total rate of starch digestion toglucose. Journal of Pediatric Gastroenterology and Nutrition 45, 32–43.
Quezada-Calvillo, R., Sim, L., Ao, Z., Hamaker, B.R., Quaroni, A., Brayer, G.D., Sterchi,E.E., Robayo-Torres, C.C., Rose, D.R., Nichols, B.L. (2008). Localization of luminalstarch substrate ‘brake’ on maltase-glucoamylase activity within the glucoamylasesubunit. Journal of Nutrition 138, 685–692.
Seal, C.J., Daly, M.E., Thomas, L.C., Bal, W., Birkett, A.M., Jeffcoat, R., Mathers, J.C.(2003). Postprandial carbohydrate metabolism in healthy subjects and those with type 2diabetes fed starches with slow and rapid hydrolysis rates determined in vitro. BritishJournal of Nutrition 90, 853–864.
Seneviratne, H.D., Biliaderis C.G. (1991). Action of a-amylases on amylose-lipid complexsuperstructures. Journal of Cereal Science 13, 129–143.
Shi, Y.C., Cui, X., Birkett, A.M, Thatcher, M.G. (2003). Slowly digestible starch products.US Patents 20030219520, 20030215562.
Shimada, M., Mochizuki, K., Goda, T. (2008). Dietary resistant starch reduces levels ofglucose-dependent insulinotropic polypeptide mRNA along the jejunum-ileum inboth normal and type 2 diabetic rats. Bioscience, Biotechnology & Biochemistry 72,2206–2209.
Shimada, M., Mochizuki, K., Goda, T. (2009). Feeding rats dietary resistant starch shiftsthe peak of SGLT1 gene expression and histone H3 acetylation on the gene fromthe upper jejunum toward the ileum. Journal of Agricultural and Food Chemistry 57,8049–8055.
Shin, S.I., Choi, H.J., Chung, K.M., Hamaker, B.R., Park, K.H., Moon, T.W. (2004). Slowlydigestible starch from debranched waxy sorghum starch: preparation and properties.Cereal Chemistry 81, 404–408.
Sim, L., Quezada-Calvillo, R., Sterchi, E.E., Nichols, B.L., Rose, D.R. (2008). Humanintestinal maltase–glucoamylase: crystal structure of the n-terminal catalytic subunit andbasis of inhibition and substrate specificity. Journal of Molecular Biology 375, 782–792.
Sim, L., Willemsma, C., Sim, L., Willemsma, C., Mohan, S., Naim, H.Y., Pinto, B.M.,Rose, D.R. (2010). Structural basis for substrate selectivity in human maltase-glucoamylase and sucrase-isomaltase N-terminal domains. Journal of BiologicalChemistry 285, 17763–17770.
Slavin, J.L. (2005). Dietary fiber and body weight. Nutrition 21, 411–418.Sparti, A., Milon, H., Vetta, V.D., Schneiter, P., Toppy, L., Jequier, E., Schuz, Y. (2000).
Effect of diets high or low in unavailable and slowly digestible carbohydrates on thepattern of 14-h substrate oxidation and feeling of hunger in humans. American Journal ofClinical Nutrition 72, 1461–1468.
Swallow, D.M., Poulter, M., Hollox, E.J. (2001). Intolerance to lactose and other dietarysugars. Drug Metabolism and Disposition 29, 513–516.
Tester, R.F., Karkalas, J., Qi, X. (2004). Starch structure and digestibility enzyme-substraterelationship. World’s Poultry Science Journal 60, 186–195.
128 Resistant Starch

3GCH06 07/20/2013 8:2:39 Page 129
Thompson, D.B. (2000). On the non-random nature of amylopectin branching. Carbohy-drate Polymers 43, 223–239.
Topping, D.L., Bajka, B.H., Bird, A.R., Clarke, J.M., Cobiac, L., Conlon, M.A.,Morrell, M.K., Toden, S. (2008). Resistant starches as a vehicle for deliveringhealth benefits to the human large bowel. Microbial Ecology in Health and Disease20, 103–108.
U.S. Department of Agriculture and U.S. Department of Health and Human Services(2010). Dietary Guidelines for Americans, 2010, 7th edition. US Government PrintingOffice, Washington, DC.
Van Citters, G.W., Lin, H.C. (2006). Ileal brake: Neuropeptidergic control of intestinaltransit. Current Gastroenterology Reports 5, 367–373.
Venkatachalam, M., Kuschnick, M.R., Zhang, G., Hamaker, B.R. (2009). Starch-entrappedbiopolymer microspheres as a novel approach to vary blood glucose profiles. Journal ofthe American College of Nutrition 28, 583–590.
Wachters-Hagedoorn, R.E., Priebe, M.G., Heimweg, J.A.J., Heiner, A.M., Englyst, K.N.,Holst, J.J., Stellaard, F., Vonk, R.J. (2006). The rate of intestinal glucose absorption iscorrelated with plasma glucose-dependent insulinotropic polypeptide concentrations inhealthy men. Journal of Nutrition 136, 1511–1516.
Wolever, T.M. (2003). Carbohydrate and the regulation of blood glucose and metabolism.Nutrition Reviews 61, S40–S48.
Wolever, T.M., Jenkins, D.J., Ocana, A.M., Rao, V.A., Collier, G.R. (1988). Second mealeffect: low-glycemic-index foods eaten at dinner improve subsequent breakfast glyce-mic response. American Journal of Clinical Nutrition 48, 1041–1047.
Wolever, T.M., Yang, M., Zeng, X.Y., Atkinson, F., Brand-Miller, J.C. (2006). Foodglycemic index, as given in glycemic index tables, is a significant determinant ofglycemic responses elicited by composite breakfast meals. American Journal of ClinicalNutrition 83, 1306–1312.
Wolf, B.W., Bauer, L.L., Fahey, G.C. (1999). Effects of chemical modification on in vitrorate and extent of food starch digestion: An attempt to discover a slowly digested starch.Journal of Agricultural and Food Chemistry 47, 4178–4183.
Wolf, B.W., Wolever, T.M.S., Bolognesi, C., Zinker, B.A., Garleb, K.A., Firkins, J.L.(2001). Glycemic response to a food starch esterified by 1-octenyl succinic anhydride inhumans. Journal of Agricultural and Food Chemistry 49, 2674–2678.
Wolf, B.W., Garleb, K.A., Choe, Y.S., Humphrey, P.M., Maki, K.C. (2003). Pullulan is aslowly digestible carbohydrate in humans. Journal of Nutrition 133, 1051–1055.
Wood, P.J., Braaten J.T., Fraser W.S., Riedel, D., Poste, L.M. (1990). Comparisons ofviscous properties of oat and guar gum and the effects of these and oat bran on glycemicindex. Journal of Agricultural and Food Chemistry 38, 753–757.
Yang, W., Hattori, M., Kawaguchi, T., Takahashi, K. (1998). Properties of starchesconjugated with lysine and poly(lysine) by theMaillard reaction. Journal of Agriculturaland Food Chemistry 46, 442–445.
Yokoyama, W.H., Hudson, C.A., Knuckles, B.E., Chiu, M.M., Sayre, R.N., Turnlund, J.R.,Schneeman, B.O. (1997). Effect of b-glucan in durum wheat pasta on human glycemicresponse. Cereal Chemistry 74, 293–296.
Zhang, G., Hamaker, B.R. (1998). Low a-amylase starch digestibility of cooked sorghumflours and the effect of protein. Cereal Chemistry 75, 710–713.
Zhang, G., Ao, Z., Hamaker, B.R. (2006a). Slow digestion property of native cerealstarches. Biomacromolecules 7, 3252–3258.
Slowly Digestible Starch and Health Benefits 129

3GCH06 07/20/2013 8:2:39 Page 130
Zhang, G., Venkatachalam, M., Hamaker, B.R. (2006b). Structural basis for the slowdigestion property of native cereal starch. Biomacromolecules 7, 3259–3266.
Zhang, G., Ao, Z., Hamaker, B.R. (2008a). Nutritional property of endosperm starchesfrom maize mutants: A parabolic relationship between slowly digestible starch andamylopectin fine structure. Journal of Agricultural and Food Chemistry 56, 4686–4694.
Zhang, G., Maghaydah, S., Hamaker, B.R. (2008b). Slowly digestible state of starch:mechanism of slow digestion property of gelatinized maize starch. Journal of Agricul-tural and Food Chemistry 56, 4695–4702.
Zhang, L., Hu, X., Xu, X., Jin, Z., Tian, Y. (2011). Slowly digestible starch preparedfrom rice starches by temperature-cycled retrogradation. Carbohydrate Polymers 84,970–974.
130 Resistant Starch

3GCH07 07/20/2013 8:10:27 Page 131
7 Measurement of Resistant Starch andIncorporation of Resistant Starch intoDietary Fibre Measurements
Barry V. McClearyMegazyme International, Bray Business Park, Ireland
7.1 INTRODUCTION
Resistant starch (RS) is that portion of the starch that is not broken down and
absorbed in the small intestine of humans. It enters the large intestine, where it
is partially or wholly fermented. The presence of a starch fraction resistant to
enzymic hydrolysis was first recognized by Englyst et al. (1982) during their
research on the measurement of non-starch polysaccharides.
Several in vivo approaches have been adopted for the measurement of
resistant starch, including: the hydrogen breath test; direct collection of ileal
effluent from patients (ileostomy patients) who have had the colon removed;
and direct collection of the ileal effluent from healthy subjects using a long
triple lumen tube (Champ et al., 2001). Of these, the ileostomy model is
considered to be the best, but not necessarily perfect.
It is generally accepted that any in vitro method used to measure resistant
starch must give values in line with those obtained with ileostomy patients.
Berry (1986) modified the in vitro method of Englyst et al. (1982) to mimic
physiological conditions more closely. Incubations were performed at 37 �C.Pancreatic a-amylase and pullulanase was again employed, but the initial
heating step at 100 �C was omitted. Using this method, the measured resistant
starch contents of samples were much higher than those previously obtained.
This was subsequently confirmed by Englyst & Cummings (1985, 1986, 1987)
through studies with healthy ileostomy subjects.
131
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH07 07/20/2013 8:10:28 Page 132
These authors (Englyst et al. 1992) also divided RS into three classes,
namely:
� RS1: physically trapped starch as found in coarsely ground or chewed
cereals, legumes, and grains.� RS2: resistant starch granules or non-gelatinized starch granules which are
highly resistant to digestion by a-amylase until gelatinized, e.g. uncooked
potato, green banana and high-amylose starch).� RS3: retrograded starch polymers (mainly amylose), which are produced
when starch is cooled after gelatinization or during heat-moisture treatment
on annealing of starch granules (during which the starch is not gelatinized).
A fourth type of resistant starch (RS4; chemically modified starch) was
introduced by Brown et al. (1998). This starch, unlike RS1, RS2 and RS3,
contains additional chemical groups. Englyst et al. (1992) also reported on a
method for the measurement of readily digested starch (RDS), slowly digested
starch (SDS) and resistant starch (RS). In this method, resistant starch is
calculated by subtracting the sum of RDS plus SDS from total starch.
Although the method can yield useful information, it is very laborious and
gives poor reproducibility without extensive training of the analyst (Champ
et al., 2001). Accuracy is severely hampered by the fact that, with samples
containing high levels of starch with low resistant starch content, one large
analytical value is subtracted from another large value. In fact, the errors in the
measurement may be as large as the resistant starch value, e.g. materials with
approximately 70% starch and 2% resistant starch.
By the early 1990s, the physiological significance of RS was fully realized.
Several new or modified methods for its measurement were developed during
the European Research Program, EURESTA (Englyst et al., 1992; Champ,
1992). The Champ (1992) method, was based on modifications to the method
of Berry (1986), and gave a direct measurement of RS. Basically, sample size
was increased from 10mg to 100mg, the sample was digested with pancreatic
a-amylase only, and incubations were performed at pH 6.9 (pH 5.2 was used
by Englyst et al. (1982, 1992) and Berry (1986)). RS determinations were
performed directly on the pellet.
Muir & O’Dea (1992) developed a procedure for RS in which samples
were chewed, treated with pepsin and then with a mixture of pancreatic
a-amylase and amyloglucosidase (AMG) in a shaking water bath at pH 5.0,
37 �C for 15 hours. The residual pellet (containing RS) was recovered
by centrifugation and washed with acetate buffer by centrifugation, and the
RS was digested by a combination of heat, DMSO and thermostable
a-amylase treatments.
132 Resistant Starch

3GCH07 07/20/2013 8:10:28 Page 133
Other methods for RS determination were developed by Faisant et al.
(1995), Goni et al. (1996), Akerberg et al. (1998) and Champ et al. (1999).
These modifications included changes in enzyme concentrations employed,
types of enzymes used (all used pancreatic a-amylase, but pullulanase was
removed, and in some cases replaced by AMG), sample pre-treatment
(chewing), pH of incubation and the addition (or not) of ethanol after the
a-amylase incubation step (Champ et al., 1999).
7.2 DEVELOPMENT OF AOAC OFFICIAL METHOD2002.02
While significant steps were made in the development of in vitro methods for
the measurement of resistant starch during the 1990s, none of these methods
were successfully subjected to interlaboratory evaluation. This prompted
McCleary & Monaghan (2002) to look at each of these methods in detail,
to evaluate all of the parameters involved and to identify sources of variability.
The ultimate aim was to develop a procedure that gave values in line with
those obtained with ileostomy patients, but also a method that could survive
the rigors of interlaboratory evaluation. Parameters investigated included:
a) incubation conditions (shaking/stirring, pH, temperature, time);
b) level of pancreatic a-amylase employed;
c) level of AMG employed;
d) the importance of protease pre-treatment;
e) procedures for recovery of resistant starch;
f) method for the dissolution of RS; and
g) glucose determination procedure.
Incubations were performed at physiological temperature (37 �C) in both ashaking water bath and in an arrangement in which the contents of the tubes
were continually stirred at different speeds. Incubations were allowed to
proceed for up to 24 hours, and the RS values obtained for a set of samples
were compared to values obtained from ileostomy studies. Typical results
obtained for regular maize starch (RMS) and high-amylose maize starch
(HAMS) are shown in Figure 7.1.
These incubations were performed in the presence of optimal levels of
AMG and at a pH of 6.0. This pH was chosen as a compromise to allow for the
different pH optima of a-amylase (pH 6.9) and AMG (pH 4.5) and the
determined stability of the two enzymes on extended incubation at different
pH values. At pH 6.0, pancreatic a-amylase has approximately 80% of the
Measurement of Resistant Starch and Incorporation of Resistant Starch 133

3GCH07 07/20/2013 8:10:28 Page 134
activity at the optimal pH of 6.9, and AMG has 20% of the activity at the
optimal pH of 4.5. The level of each enzyme is adjusted to optimize the assay.
AMG is very stable at both pH 6.0 and 6.9, losing less than 5% of initial
activity on incubation at 37 �C for 16 hours. Pancreatic a-amylase is less
stable, however; even at pH 6.0, approximately 30% of activity remains on
incubation of the enzyme under assay conditions of 16 hours at 37 �C(compared to 50% at pH 6.9) (Figure 7.2).
There was considerable flexibility in the concentration of pancreatic
a-amylase used. RS values obtained for RMS and HAMS varied little,
with enzyme concentrations ranging from 15–60 units/ml of incubation
mixture (see Figure 7.1).
AMG in the incubation mixture had a considerable effect on the deter-
mined RS values (McCleary &Monaghan, 2002); this is considered to be due
to the known inhibitory effect of maltose on pancreatic a-amylase. The AMG
removed the maltose by hydrolyzing it to glucose, which has no inhibitory
effect. The effect of protease on determined RS values was studied by
including a pre-treatment with pepsin at pH 2. Results obtained indicated
that the protease pre-treatment had no significant effect on determined RS
values (McCleary & Monaghan, 2002). This may be due, in part, to the
presence of an active protease in the pancreatic a-amylase preparation used.
Figure 7.1 The effect of the concentration of pancreatic a-amylase and incubationtime on the determined RS value of regular maize starch (RMS) and high-amylosemaize starch (HAMS) at pH 6.0 and 37 �C for up to 25 hours. HAMS with pancreatica-amylase concentrations of (black triangle) 15, (black circle) 30 and (black square)60 U/mL; RMS with pancreatic a-amylase concentrations of (open triangle) 15,(open circle) 30 and (open square) 60 U/mL.
134 Resistant Starch

3GCH07 07/20/2013 8:10:28 Page 135
Resistant starch is crystalline and is difficult to dissolve. Solvents used to
dissolve this material include dimethyl sulphoxide (DMSO) and 2–4M
potassium hydroxide. For all samples studied, the RS was completely dis-
solved by stirring the RS containing pellet in 2MKOH in an ice/water bath for
20 minutes (conditions used by several authors). On neutralization, it was
essential to hydrolyze this starch rapidly to avoid re-crystallization (which
would again render the starch resistant to hydrolysis by AMG. To achieve this
and to simplify neutralization, a concentrated sodium acetate buffer (1.2M,
pH 3.8) was added, followed immediately by AMG (320 units/test; one unit of
AMG activity is the amount of enzyme required to release one micromole of
glucose from soluble starch per minute at pH 4.5 and 40 �C).This method for the measurement of RS was subjected to evaluation under
the auspices of AOAC International and AACC International, to determine the
interlaboratory performance statistics. The materials used in the study repre-
sented food materials containing RS (cooked kidney beans, green banana and
cornflakes) and a range of commercial starches, most of which naturally
contain (or were processed to contain) elevated RS levels. Thirty-seven
laboratories tested eight pairs of blind duplicate starch or plant material
samples, with RS values between 0.6 (regular maize starch) and 64% (fresh
weight basis). For samples excluding regular maize starch, RSDr values
Figure 7.2 Stability of pancreatic a-amylase and AMG on incubation at pH 6.0 and6.9 and 37 �C for up to 25 hours. AMG at pH 6.0 (closed triangle) and 6.9 (opentriangle); pancreatic a-amylase at pH 6.0 (open circle) and 6.9 (closed circle).
Measurement of Resistant Starch and Incorporation of Resistant Starch 135

3GCH07 07/20/2013 8:10:28 Page 136
ranged from 1.97–4.2% and RSDR values ranged from 4.58–10.9%. The range
of applicability of the test is 2–64% RS.
The method was not suitable for samples with less than 1%RS (e.g. regular
maize starch; 0.6% RS). For such samples, RSDr and RSDR values are
unacceptably high. On the basis of this evaluation, the method was accepted
asAOACOfficialMethod 2002.02 andAACCRecommendedmethod 32–40.01
(McCleary et al., 2002).
7.3 DEVELOPMENT OF AN INTEGRATED PROCEDUREFOR THE MEASUREMENT OF TOTAL DIETARY FIBRE
Hipsley (1953) coined the term dietary fibre to cover the non-digestible
constituents of plants that make up the plant cell wall (known to include
cellulose, hemicellulose and lignin) with the aim of defining some property of
the constituent of the food that could be related to physiological behaviour in
the human small intestine. This definition was broadened by Trowell et al.
(1976) to become primarily a physiological definition, based on edibility and
resistance to digestion in the human small intestine. Thus, the definition
included indigestible polysaccharides such as gums, modified celluloses,
mucilages and pectin, and non-digestible oligosaccharides (NDO).
Methods which were developed to meet this analytical requirement
focused on the use of enzymes to remove starch and protein. The enzymes
employed require a defined level of activity and must be devoid of contami-
nating enzymes active on dietary fibre components such as pectin, b-glucan,arabinoxylan and other hemicelluloses. Following extensive international
collaboration, the method that evolved was AOAC Official Method 985.29,
‘Total dietary fibre in foods; enzymatic-gravimetric method’ (Prosky et al.,
1985, 1994) This method was subsequently extended to allowmeasurement of
total dietary fibre (TDF), soluble dietary fibre (SDF) and insoluble dietary
fibre (IDF) in foods (AOAC Official Method 991.43) (Lee et al., 1992) Other
modifications to these methods for fibre analysis have also been approved by
AOAC International (Theander & Aman, 1982).
In concurrent research in the UK, methods were developed for the measure-
ment of non-starch polysaccharides (NSP)(Englyst et al., 1982; Englyst &
Cummings, 1984, 1985; Englyst & Hudson, 1987), based on the original work
of Southgate (1969) and Southgate et al. (1978). These NSP procedures
measure only NSP; RS and NDO are excluded. Starch in the sample is
completely dissolved in hot dimethyl sulphoxide (DMSO), diluted in buffer
and depolymerized with thermostable a-amylase, followed by a mixture of
136 Resistant Starch

3GCH07 07/20/2013 8:10:28 Page 137
pancreatin and pullulanase. The NSP recovered is acid hydrolyzed to
monosaccharides, which are measured by high-performance liquid chroma-
tography (HPLC), gas-liquid chromatography (after derivatization) (Englyst
& Cummings, 1984) or colorimetrically (Englyst & Hudson, 1987). These
methods have not been successfully subjected to international interlabor-
atory evaluation.
A survey of scientists initiated in 1993 (Lee & Prosky, 1995) showed that
65% of the respondents favoured the inclusion of NDO and 80% favoured
inclusion of RS in the definition of dietary fibre. This led to the development of
methods for measurement of RS (AOAC Method 2002.02) and for a number
of NDO, including fructo-oligosaccharides (AOAC Methods 997.08 and
999.03), polydextrose (AOAC Method 2000.11), resistant maltodextrins
(AOAC Method 2001.03), and galacto-oligosaccharides (AOAC Method
2001.02).
In 1998, the American Association of Cereal Chemists began a critical
review of the current state of dietary fibre science, including consideration
of the state of the dietary fibre definition. Over the course of the following
year, the committee held three workshops and provided an international
website, available to all Web users worldwide, to receive comments. All
interested parties were provided with additional opportunity for comment.
After due deliberation, an updated definition of dietary fibre was delivered to
the AACC Board of Directors for adoption in early 2000 and published
(Anon, 2001) namely:
‘Dietary fibre is the edible parts of plants or analogous carbohydrates that are
resistant to digestion and absorption in the human small intestine with
complete or partial fermentation in the large intestine. Dietary fibre includes
polysaccharides, oligosaccharides, lignin, and associated plant substances.
Dietary fibres promote beneficial physiological effects including laxation, and/
or blood cholesterol attenuation, and/or blood glucose attenuation.’
Several definitions of dietary fibre have appeared over the past ten years.
The Food Nutrition Board of the Institute of Medicine of the National
Academies (USA) (2002) defined dietary fibre as follows: ‘Dietary fibre
consists of nondigestible carbohydrates and lignin that are intrinsic and
intact in plants. Added fibre consists of isolated, nondigestible carbohydrates
that have beneficial physiological effects in humans. Total fibre is the sum of
dietary fibre and added fibre.’
At the 30th session of the CODEX Committee on Nutrition and Foods for
Special Dietary Uses (CCNFSDU; 2008), the Committee agreed on the
following definition for dietary fibre:
Measurement of Resistant Starch and Incorporation of Resistant Starch 137

3GCH07 07/20/2013 8:10:28 Page 138
‘Dietary fibre is carbohydrate polymers1 with ten or more monomeric units,2
which are not hydrolyzed by the endogenous enzymes in the small intestine of
humans and belong to the following categories:
� Edible carbohydrate polymers naturally occurring in the food as consumed.� Carbohydrate polymers which have been obtained from food rawmaterial by
physical, enzymatic or chemical means and which have been shown to have a
physiological effect of benefit to health as demonstrated by generally
accepted scientific evidence to competent authorities,� Synthetic carbohydrate polymers which have been shown to have a physio-
logical effect of benefit to health as demonstrated by generally accepted
scientific evidence to competent authorities.
The fact that a single method to measure all dietary fibre components is
needed has been known for some time. While it is possible to measure many
individual fibre components with specific and non-specific methods, total
dietary fibre cannot simply be calculated by adding the values for these
specific components to the determined value of high molecular weight dietary
fibre, as measured with AOACOfficial Methods 985.29 or 991.43. Since these
latter methods also measure some of the RS and various NDO in food
materials, summation leads to ‘double counting’ of this material (Figure 7.3;
McCleary et al., 2009).
An integrated method for the measurement of total dietary fibre was
published in 2007 (McCleary, 2007). This method allows the accurate
measurement of total high molecular weight dietary fibre (HMWDF), which
includes IDF (including RS) and higher molecular weight soluble dietary fibre
which precipitates in the presence of 76% aqueous ethanol (SDFP), as well as
lower molecular weight soluble dietary fibre which remains soluble in the
presence of 76% aqueous ethanol (SDFS). Details of this procedure are
outlined in Figure 7.4.
1When derived from a plant origin, dietary fibre may include fractions of lignin and/or other
compounds when associated with polysaccharides in the plant cell walls and if these compounds
are quantified by the AOAC gravimetric analytical method for dietary fibre analysis: Fractions
of lignin and the other compounds (proteic fractions, phenolic compounds, waxes, saponins,
phytates, cutin, phytosterols, etc.) intimately ‘associated’ with plant polysaccharides in the
AOAC 991.43 method. These substances are included in the definition of fibre insofar as they are
actually associated with the poly- or oligo-saccharidic fraction of fibre. However when
extracted or even re-introduced in to a food containing non digestible polysaccharides,
then they cannot be defined as dietary fibre. When combined with polysaccharides, these
associated substances may provide additional beneficial effects.2Decision on whether to include carbohydrates of 3 to 9 monomeric units should be left up to
national authorities.’
138 Resistant Starch

3GCH07 07/20/2013 8:10:28 Page 139
The use of pancreatic a-amylase more closely simulates digestion in the
human digestive tract and yields RS values in line with those obtained with
AOACOfficial Method 2002.02, and with results from ileostomy patients. For
most food and ingredient samples analyzed, the RS values obtained with
AOAC Method 2009.01 were higher than those obtained with AOAC Method
985.29. The notable exception is for phosphate cross-linked starch (RS4),
where values obtained with Method 2009.01 are much lower than those
obtained with Method 985.29. The physiological significance of these results
will be discussed separately (McCleary et al., 2013).
This method was successfully subjected to interlaboratory evaluation
(McCleary et al., 2009) and accepted as AOAC Method 2009.01. In this
study, total HMWDF and SDFS were measured.
In an AOACI/AACCI interlaboratory study recently completed, the method
was evaluated for the measurement of IDF, SDFP and SDFS. IDF and SDFP
are measured using the standard gravimetric procedures, with allowance for
ash and non-digested protein. SDFS is analyzed by HPLC using D-sorbitol as
internal standard and a Waters Corporation Sugar Pak1 chromatographic
column. To obtain resistant starch values in line with those obtained in vivo
with ileostomy patients, incubations are performed either in a shaking water
Figure 7.3 Schematic representation of dietary fibre components measured, and notmeasured, by AOAC Official Methods 985.29 and 991.43. Also depicted are theproblems of partial measurement of RS, Polydextrosej and resistant maltodextrins bycurrent AOAC total dietary fibre methods. Most of the SDFS (galactooligosacchar-ides, fructooligosaccharides, etc) are not measured. The currently describedintegrated total dietary fibre procedure measures all components shown, with nodouble counting.
Measurement of Resistant Starch and Incorporation of Resistant Starch 139

3GCH07 07/20/2013 8:10:29 Page 140
bath in orbital motion, or with a VarioMag1 magnetic stirrer at 37 �C for
16 hours. The pH is adjusted, and a-amylase and AMG are inactivated and
protein denatured by heating the sample to about 100 �C. Incubation with
protease is followed by pH adjustment and filtration to separate IDF and
soluble fibres. SDFP is precipitated with ethanol and recovered and dried.
After weighing, IDF and SDFP residues are analyzed for residual protein
and ash. The SDFS fraction (in the ethanolic filtrate) is concentrated by
rotary evaporation, re-dissolved and adjusted to pH 4.2–4.7 and incubated
with AMG to remove completely any traces of higher molecular weight
Add 40 ml of 50 mM Na maleate buffer, pH 6.0 (+ CaCl2) Containing pancreatic α-amylase + amyloglucosidase
Sample (1.00 g) in sealed 250 ml Duran bottle (in duplicate)
Incubate in shaking water bath at 37°C for 16 hours
Add 3.0 ml 0.75 M Trizma base to adjust pH to ≈ 8.2
Incubate at > 90°C for 20 min. Cool to ≈ 60°C
Add 0.1 ml protease
Incubate at 60°C for 30 min. Cool to room temperature
Add 4.0 ml of 2 M acetic acid (to adjust pH to ≈4.5) + 1 ml internal standard
Remove 1 ml for available CHO determination
Add 4 volumes of ethanol, stir, store at room temp for 1 hour, then filter
HMWDF determination LMWSDF determination
Figure 7.4 Schematic representation of the integrated TDF assay procedure, alsoshowingwhere samples canbe removed for determination of available carbohydrates.
140 Resistant Starch

3GCH07 07/20/2013 8:10:29 Page 141
maltodextrins. The solution is then either desalted by passage through a
column of mixed cation and anion ion exchange resins (Figure 7.5) or,
alternatively, is desalted on line using a de-ashing pre-column.
Quantitation is greatly simplified by including an internal standard. The
preferred internal standard is D-sorbitol and this is added to the sample just
prior to adding ethanol to precipitate the SDFP. A number of compounds were
evaluated as potential internal standards, including 1,5-pentanediol, dieth-
ylene glycol and triethylene glycol. Of these, diethylene glycol appeared best.
However, on closer study, some of this is lost when the SDFS fraction is rotary
evaporated, presumably by adsorbing to the glass rotary evaporator flask. This
was not observed with D-sorbitol. When D-sorbitol is rotary evaporated with a
range of sugars and NDO, the ratio of the components remains the same.
Results of this interlaboratory study have been published (McCleary et al.,
2012).
A major advantage of the described method for the measurement of total
dietary fibre is that it allows the separate measurement of IDF, SDFP and
SDFS. There is some international debate as to whether NDO (SDFS) should
be included in the dietary fibre measurements. Until there is agreement, this
Figure 7.5 Arrangement for the deionization of samples using amixed bed resin in aBio-Rad, Econo-Pacj disposable chromatographic column (cat. No. 732–1010) withan Alltech One-Way Stopcock (cat. No. 211524). Also shown is a Gilson Minipulsj
Evolution pump and collection bottles.
Measurement of Resistant Starch and Incorporation of Resistant Starch 141

3GCH07 07/20/2013 8:10:29 Page 142
oligosaccharide material can be measured and simply reported as NDO. With
minor modification, the method can also be adapted to measure digestible
carbohydrates (fructose, glucose, sucrose, maltodextrins and non-resistant
starch, and the glucose component of lactose; McCleary, 2007).
REFERENCES
Akerberg, K.E., Liljberg, G.M. Granfeldt, Y.E., Drews, A.W., Bjorck M.E. (1998). An in-vitro method based on chewing, to predict resistant starch content in foods allowsparallel determination of potentially available starch and dietary fiber. Journal ofNutrition 128, 651–660.
Anon (2001) The definition of dietary fiber. Cereal Foods World 46, 112–126.Berry, C.S. (1986). Resistant starch: formation and measurement of starch that survives
exhaustive digestion with amylolytic enzymes during the determination of dietary fibre.Journal of Cereal Science 4, 301–314.
Brown, I.L., Wang, X., Topping, D.L., Playne, M.J., Conway, P.L. (1998). High amylosemaize starch as a versatile prebiotic for use with probiotic bacteria. Food Australia 50,603–622.
Champ, M. (1992). Determination of resistant starch in foods and food products:interlaboratory study. European Journal of Clinical Nutrition 46(Suppl. 2), S51–S62.
Champ, M., Martin, L., Noah, L., M. Gratas. (1999). Analytical methods for resistantstarch. In: Cho, S.S., Prosky, L., Dreher, M. (eds). Complex Carbohydrates in Foods,169–187. Marcel Dekker, Inc., New York, USA.
Champ, M., Kozlowski, F., Lecannu, G. (2001). In-vivo and In-vitro methods for resistantstarch measurement. In: McCleary, B.V., Prosky, L. (eds). Advanced Dietary FibreTechnology106–119. Blackwell Science Ltd., Oxford, UK.
Codex Committee on Nutrition and Foods for Special Dietary Uses 28th Session. ChiangMai, Thailand, 30th October–3rd November 2006.
Englyst, H.N., Cummings, J.H. (1984). Simplified method for the measurement of non-starch polysaccharides by gas-liquid chromatography of constituent sugars as alditolacetates. Analyst 109, 937–942.
Englyst, H.N., Cummings, J.H. (1985). Digestion of the polysaccharides of some cerealfoods in the human small intestine. American Journal of Clinical Nutrition 42,778–787.
Englyst, H.N., Cummings, J.H. (1986). Digestion of the carbohydrates of banana (Musaparadisiaca sapientum) in the human small intestine. American Journal of ClinicalNutrition 44, 42–50.
Englyst, H.N., Cummings, J.H. (1987). Digestion of polysaccharides of potato in the smallintestine of man. American Journal of Clinical Nutrition 45, 423–431.
Englyst, H.N., Hudson, G.J. (1987). Colorimetric method for the routine measurement ofdietary fibre as non-starch polysaccharides. A comparison with gas-liquid chromatog-raphy. Food Chemistry 24, 63–67.
Englyst, H.N., Wiggins, H.L., Cummings, J.H. (1982). Determination of the non-starchpolysaccharides in plant foods by gas-liquid chromatography of constituent sugars asalditol acetates. Analyst 107, 307–318.
142 Resistant Starch

3GCH07 07/20/2013 8:10:29 Page 143
Englyst, H.N., Kingman, S.M., Cummings, J.H. (1992). Classification and measurementof nutritionally important starch fractions. European Journal of Clinical Nutrition 46,S33–S50.
Faisant, F., Planchot, V., Kozlowski, F., Pacouret, P., Colonna, P., Champ, M. (1995).Resistant starch determination adapted to products containing high levels of resistantstarch. Sciences des Aliments 15, 83–89.
Goni, I., Manas, E., Garcia-Diz, F. Saura-Calixto, F. (1996). Analysis of resistant starch: amethod for food and food products. Food Chemistry 56, 445–449.
Hipsley, E.H. (1953). Dietary fibre and pregnancy toxaemia. British Medical Journal 2,420–422.
Horwitz, W. (ed). (2002). Official Methods of analysis of AOAC International, 17thEdition. The Association, Gaithersberg, USA.
Institute of Medicine (2002). In:Dietary reference intakes: Energy, Carbohydrates, Fiber,Fat, Fatty Acids, Cholesterol, Protein and Amino Acids. National Academies Press,Washington, DC.
Lee, S.C., Prosky, L. (1995). International survey on dietary fiber definition, analysis andreference materials. Journal of the Association of Official Analytical Chemists 78, 22–36.
Lee, S.C., Prosky, L., DeVries, J.W. (1992). Determination of total, soluble, and insolubledietary fiber in foods – Enzymatic-gravimetric method,MES-TRIS buffer: Collaborativestudy. Journal of the Association of Official Analytical Chemists 75, 395–416.
McCleary, B.V. (2007). An integrated procedure for the measurement of total dietary fibre(including resistant starch), non-digestible oligosaccharides and available carbohy-drates. Analytical Bioanalytical Chemistry 389, 291–308.
McCleary, B.V., Sloane, N., Draga, A., Lazewska, I. (2013) Cereal Chemistry, in press.McCleary, B.V., Monaghan, D.A. (2002). Measurement of resistant starch. Journal of theAssociation of Official Analytical Chemists 85, 665–675.
McCleary, B.V., Rossiter, P. (2004). Measurement of novel dietary fibres. Journal of theAssociation of Official Analytical Chemists 87, 707–717.
McCleary, B.V., McNally, M., Rossiter, P. (2002). Measurement of resistant starch byenzymic digestion in starch samples and selected plant materials: Collaborative Study.Journal of the Association of Official Analytical Chemists 85, 1103–1111.
McCleary, B.V., Mills, C., Draga, A. (2009). Development and evaluation of an integratedmethod for the measurement of total dietary fibre. Quality Assurance and Safety ofCrops and Foods 1, 213–224.
McCleary, B.V., DeVries, J.W., Rader, J.I., Cohen, G., Prosky, L., Mugford, D.A., Champ,M., Okuma, K. (2010). Determination of total dietary fiber (CODEX definition) byenzymatic-gravimetric method and liquid chromatography: collaborative study. Journalof the Association of Official Analytical Chemists 93, 221–33.
McCleary, B.V., DeVries, J.W., Rader, J.I., Cohen, G., Prosky, L, Mugford, D.C., Okuma,K. (2012). Determination of insoluble, soluble, and total dietary fiber (CODEX defini-tion) by enzymatic-gravimetric method and liquid chromatography: collaborative study.Journal of AOAC International 95, 824–844.
Muir, J.A., O’Dea, K. (1992). Measurement of resistant starch: Factors affecting theamount of starch escaping digestion in vitro. American Journal of Clinical Nutrition56, 123–127.
Prosky, L., Asp, N-G., Furda I, DeVries, J.W., Schweizer, T.F., Harland, B.F. (1985).Determination of total dietary fiber in food and food products: collaborative study.Journal of the Association of Official Analytical Chemists 68, 677–679.
Measurement of Resistant Starch and Incorporation of Resistant Starch 143

3GCH07 07/20/2013 8:10:30 Page 144
Prosky, L., Asp, N-G., Schweizer, T.F. DeVries, J.W., Furda, Lee, S.C. (1994). Determi-nation of soluble dietary fiber in foods and food products: Collaborative study. Journal ofthe Association of Official Analytical Chemists 77, 690–694.
Southgate, D.A.T. (1969). Determination of carbohydrates in foods. II. Unavailablecarbohydrates. Journal of the Science of Food and Agriculture 20, 326–330.
Southgate, D.A.T., Hudson, G.J., Englyst, H.N. (1978). The analysis of dietary fibre, thechoices for the analyst. Journal of the Science of Food and Agriculture 29, 979–988.
Theander, O., Aman, P. (1982). Studies on dietary fibre: A method for the analysis andchemical characterization of total dietary fibres. Swedish Journal of AgriculturalResearch 9 (3), 97–106.
Trowell, H.C., Southgate, D.A.T., Wolever, T.M.S., Leeds A.R., Gassull, M.A., Jenkins,D.J.A. (1976). Dietary fiber redefined. Lancet 1, 967.
144 Resistant Starch

CH08GR 07/29/2013 16:22:47 Page 145
8 In Vitro Enzymatic Testing Methodand Digestion Mechanism ofCross-linked Wheat Starch
Radhiah Shukri,1 Paul A. Seib,1 Clodualdo C.Maningat,1,2 and Yong-Cheng Shi1
1Department of Grain Science and Industry, Kansas State University, USA2MGP Ingredients, Inc., USA
8.1 INTRODUCTION
Early studies have shown that a fraction of some starches consumed by
humans escape the small intestine (Anderson et al., 1981; Stephen et al.,
1981; Englyst & Cummings, 1985; Asp et al., 1987). The total amount of
starch and products of starch degradation that resist digestion in the small
intestine of healthy people is termed resistant starch (RS) (Asp, 1992). RS is
further categorized into five classes: physically inaccessible starch known as
RS1; granular starch known as RS2; cooked and retrograded starches known
as RS3; chemically modified starches known as RS4; and amylose-lipid
complex (Brown et al., 2006; Hasjim et al. 2010). The degree of starch
digestibility is affected by the structure of starch granules, the physical
characteristics of food, the amylose and amylopectin ratio, retrogradation
of amylose and the presence of other nutrients and anti-nutrients (Sharma
et al., 2008; Bird et al., 2009).
Chemical modification of starch has been proved to affect the extent and
rate of digestibility in the small intestine (Wolf et al., 1999), based on starch
source, the type and degree of modification, extent of granule gelatinization
and the source of enzyme used (Filer, 1971). Cross-linked (CL) starch is
one of the most highly produced and utilized chemically modified starches in
the food industry (Wurzburg, 1986). The highest cross-linking levels that are
acceptable as food starch with 0.4% phosphorus (P) add-ons are achievable by
145
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

CH08GR 07/29/2013 16:22:47 Page 146
using 12% sodium trimetaphosphate (STMP) and sodium tripolyphosphate
(STPP) as phosphorylating agents (Woo & Seib, 2002).
Several in vitro methods have been suggested to quantify RS in food,
all of which involve enzymatic digestions on starch, removal of the
digested starch and quantitation of remaining RS. Quantitation of RS can
be carried out using direct methods or indirect methods. While direct
methods determine RS in the remaining starch after removing the
digested portion (Berry, 1986; Champ, 1992; Faisant et al., 1993;
McCleary et al., 2002), the indirect methods measure RS by subtracting
the digested starch from the total starch (Englyst, et al., 1992; Tovar
et al., 1990). Determination of RS is achieved by solubilizing RS and
quantifying with colorimetric methods (Berry, 1986; McCleary et al.,
2002), or by isolating RS gravimetrically after ethanol precipitation
(Lee et al., 1992; McCleary et al., 2010).
Most methods use a-amylase (Englyst et al., 1992) or a cocktail of
a-amylase and amyloglucosidase (McCleary et al., 2002, 2010) to hydrolyze
the RDS and SDS fractions. Enzyme cocktails are used to avoid possible
inhibition of a-amylase by maltose and maltotriose (Sharma et al., 2008). The
extrinsic factor (i.e., extent of chewing, rate of orocecal transit), incubation
temperature, incubation time and enzyme source may affect the in vitro yield
of these assays (Englyst et al., 1992).
Table 8.1 shows a comparison of in vivo and in vitro RS content using
Englyst, AOAC Method 991.43, AOAC Method 2002.02 and AOAC
Method 2009.01 in several samples. The RS content of each sample either
differed slightly or significantly when compared to the in vitro methods
used. Several samples determined by the Englyst method (raw potato
starch, cornflakes and raw green banana) and AOAC Method 2002.02
(raw potato starch and raw green banana) showed results consistent with the
in vivo data. Although AOAC Method 991.43 was reported to have
consistency with ileostomy patients fed with bean and potato flakes
(Schweizer et al., 1990), the RS reported in Table 8.1 revealed that
none of the samples except cornflakes had similar in vivo compatibility
with the RS determined by AOAC Method 991.43. The significantly lower
RS (determined by AOAC Method 991.43) of raw potato starch, high-
amylose corn starch and raw green banana was likely due to a higher
incubation temperature (100 �C).The most current in vitro method, namely AOAC Method 2009.01, claims
to effectively determine non-starch polysaccharides, RS and non-digestible
oligosaccharides content in samples (McCleary et al., 2010). The latter
method applies key features of AOAC Method 985.29, AOAC Method
991.43, AOAC Method 2001.03 and AOAC Method 2002.02 (McCleary
146 Resistant Starch

CH08GR 07/29/2013 16:22:48 Page 147
et al., 2010). The starch digestion conditions in AOAC Method 2009.01 were
similar to those used in AOACMethod 2002.02, which involve the incubation
of a sample with a pancreatic a-amylase and amyloglucosidase cocktail for 16
hours at 37 �C. Although the digestive enzyme concentration in AOAC
Method 2009.01 was significantly higher, results produced by AOACMethod
2009.01 and AOAC Method 2002.02 were claimed to be similar for most
samples (McCleary, 2007).
The RS content by AOAC Method 2009.01 in the samples listed in Table
8.1 showed no consistency with in vivo data. Based on a comparison of in vitro
methods (Englyst Method, AOAC Method 991.43, AOAC Method 2009.01)
on CLwheat starch, AOACMethod 2009.01 provided a significantly lower RS
content (Table 8.1). AOAC Method 2002.02 was not included in the latter
comparison, because CL wheat starch is not able to solubilize in 2M
potassium hydroxide for determination of glucose content using the colori-
metric procedure.
The findings of RS content in CL wheat starch raise questions as to the
efficiency of AOAC Method 2009.01 to quantify RS and the compatibility
with in vivo results. As opposed to the Englyst Method, AOAC Method
2009.01 differs in enzyme concentration, buffers, measurement method,
incubation time and enzyme sources, but both methods employ the same
Table 8.1 Comparison of resistant starch (RS: % total starch) content in raw potatostarch, high-amylose corn starch, corn flakes and raw green banana determined byin vitro methods (Englyst Method, AOAC Method 991.43, AOAC Method 2002.02 andAOAC Method 2009.01) and in vivo method (ileostomy model).
In vitro RS (%)
Source of starch EnglystAOAC991.43
AOAC2002.02
AOAC2009.01
In vivoRS (%)
Raw potato starch 66.5a 0.9b 64.9b 56.8b 67.9c
High-amylose corn starch 71.4a 25.6b 50.0b 49.3b 43.7c
Corn flakes 3.9a 3.3e 2.2b 2.4b 3.1–5.0d
Raw green banana 54.2b 7.5b 51b 38b 55.3c
Cross-linked wheat starch� 81.7 82.3 –�� 23.9 –
aEnglyst et al. (1992).bMcCleary (2007).cLangkilde & Andersson (1995).dMuir and O’Dea, (1992).ePendlington (1999).� RS content was determined by in vitro methods in Kansas State University lab.�� Unable to determine RS in cross-linked wheat starch due to insolubility of cross-linked starch in 2M
potassium hydroxide.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 147

CH08GR 07/29/2013 16:22:48 Page 148
incubation temperature (37 �C). The digestion kinetics, specifically the
mechanism of enzyme action and incubation time in AOAC Method
2009.01, was our major interest. We predicted that 16 hours of incubation
time, with a relatively high amount and concentration of digestive enzyme
(40ml a-amylase (50U/ml) and amyloglucosidase (3.4U/ml)), would be too
harsh of a condition for CLwheat starch. Hence, the objectives of this research
were to study the in vitro digestion behaviour of CL wheat starch at various
time intervals, the progressive changes of CL wheat starch granules during the
in vitro digestion and the mechanism of the digestive enzymes during the
digestion period in AOAC Method 2009.01.
8.2 MATERIALS ANDMETHODS
8.2.1 Materials
CL wheat starch (Fibersym1 RW) and native wheat starch (MidsolTM 50)
were obtained from MGP Ingredients, Inc1 (Atchison, Kansas). The inte-
grated total dietary fibre assay kit (catalogue no. INTDF 06/12) was purchased
from Megazyme International Ireland Ltd. (Wicklow, Ireland). All chemicals
were reagent grade.
8.2.2 General methods
P content was assayed using the procedure of Smith & Caruso (1964.)
Moisture content was obtained according to AACC Method 44-15 (AACC
2000).
8.2.3 Conversion of CL wheat starch to phosphodextrinsand 31PNMR spectra of the phosphodextrins
The treatment was based on the method described by Sang et al. (2010).
Starch (1.0 g db) was weighed into a 50ml centrifuge tube. The starch was
slurried with 30ml 2.0mM calcium chloride at pH 8.2, and heat-stable
a-amylase (100ml) was then added. The mixture was heated in a boiling
water bath for 30 minutes and 100ml of heat stable a-amylase was added into
the mixture again. After cooling to room temperature, the pH of the mixture
was adjusted to 4.5, and amyloglucosidase (200ml) was incorporated for one
hour to incubate at 60 �C. The mixture was adjusted to pH 7.0, centrifuged
(1500� g, 10min), and the supernatant was freeze-dried.
148 Resistant Starch

CH08GR 07/29/2013 16:22:48 Page 149
The freeze-dried starch digest (1.0 g, db) was dissolved in deuterium
oxide (1.0ml) containing 20mM EDTA and 0.002% sodium azide. The pH
of the solution was adjusted to 8.0 by adding 0.1M sodium hydroxide. The
proton-decoupled 31P NMR spectra were obtained using a 11.75 Tesla
Varian NMR System (Varian Inc., Palo Alto, CA). Using a method by Sang
et al. (2007), the 1H NMR was operated at 499.84MHz and 31P at
202.34MHz. The 31P NMR data collection was carried out at 25 �C using
a delay of six seconds between pulses, with a pulse width of 15.0ms and a
sweep width at 12,730Hz. The samples were run for eight hours for 31P
spectra detection. The obtained spectra were processed and analyzed using
Varian software VNMRJ Version 2.2C. Chemical shifts were reported in d(ppm) from the reference signal of an 85% phosphoric acid external
standard.
8.2.4 Digestibility of CL wheat starch
Digested starch of native and CL wheat starches were determined at 0, 1, 2, 4,
6, 8, 16 and 24 hours, using the a-amylase and amyloglucosidase incubation
procedure from AOAC Method 2009.01. To collect undigested starch for
characterization, CL wheat starch was weighed to 1.0 g (db), wet with ethanol
(1.0ml), and a pancreatic a-amylase/amyloglucosidase mixture (40ml) was
added to a 250ml glass bottle. The pancreatic a-amylase/amyloglucosidase
mixture contained 50 units (U)/ml of a-amylase and 3.4U/ml amylogluco-
sidase. 1 U of a-amylase at pH 5.8 and 37 �C was defined as the amount of
enzyme required to release one micromole of D-glucose per minute from
soluble starch. 1 U of amyloglucosidase at pH 4.5 and 40 �Cwas defined as the
amount of enzyme required to release one micromole of D-glucose per minute
from soluble starch.
The bottle was capped and placed in a water bath at 37 �C with a
continuous (170 rpm) stirring via a stir bar. Fourteen bottles were incu-
bated for each sample, and two bottles were taken out after 1, 2, 4, 6, 8, 16
and 24 hours. After cooling to 25 �C, the pH of the mixture was decreased
to 2.5 to inactivate the enzyme activity with the addition 1.0M hydro-
chloric acid. The mixture was held at pH 2.5 for one hour before
increasing the pH to 6.0 via the addition of 1.0M sodium hydroxide.
Subsequently, the mixture was centrifuged (1500� g, 10min), supernatant
decanted, pellet washed (two times) with water and centrifuged (1500� g,
10min), and the pellet was then oven-dried at 37 �C for 5 hours. The dried
pellet (undigested portion of the CL wheat starch) was gently ground using
a pestle and mortar and was stored at room temperature in an airtight
container.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 149

CH08GR 07/29/2013 16:22:48 Page 150
8.2.5 Thermal properties
The gelatinization temperatures and the enthalpy of digested CL wheat starch
were measured by differential scanning calorimetry (DSC) (TA Instruments
Q100, TA Instruments, New Castle, DE). The total solids content of samples
were 33.3% (w/w, dry basis). After hydration for one hour at 25 �C, 30mg of
well-stirred sample suspensions were weighed into 40ml aluminium pans and
hermetically sealed immediately to prevent moisture loss. Scans were per-
formed from 10–130 �C at a constant heating rate of 10 �C/min. A sealed
empty pan was used as a reference and the DSC was calibrated using indium.
The gelatinization enthalpy (DH) and transition temperatures, namely the
onset temperature (To), peak temperature (Tp) and conclusion temperature
(Tc), were determined on the basis of the first-run DSC heating curves.
The DH was evaluated.
8.2.6 Microscopic observation
CLwheat starch digested after 1, 2, 4, 6, 8, 16 and 24 hours of incubation were
placed on microscope slides and observed under an Olympus BX-51 micro-
scope (Olympus, Tokyo, Japan) with a 40� objective. CL wheat starch before
digestion was also observed under a microscope for comparison with the
assayed CL wheat starch.
8.2.7 Scanning electron microscope (SEM)
The samples were sprinkled lightly onto a carbon double-sided adhesive
tape on metal specimen stubs, which were then coated with gold-palladium
(60 : 40 ratio) under vacuum with a Desk II Sputter/Etch Unit (Denton
Vacuum, LLC, Moorestown, NJ). Micrographs of the samples were obtained
at 1000� and 5000� magnifications by SEM (S-3500N, Hitachi Science
Systems, Ltd, Japan) at an accelerating potential of 20 kV using an X-ray
Detector-Link Pentafet 7021 (Oxford Instruments Microanalysis Limited,
Bucks, England).
8.2.8 Statistical analysis
All data were statistically analyzed by analysis of variance (ANOVA) using
Statistical Analysis Software (SAS) (version 9.2, SAS Institute, Cary, NC),
and the values are expressed as means � standard deviations from two
replicates, unless stated otherwise.
150 Resistant Starch

CH08GR 07/29/2013 16:22:49 Page 151
8.3 RESULTS AND DISCUSSION
8.3.1 Effects of a-amylase/amyloglucosidase digestion onP content and chemical forms of the phosphateesters on starch
The P content of CL wheat starch after different incubation times with
a-amylase/amyloglucosidase is reported in Table 8.2. The initial P content
of CL wheat starch (at 0 hours) was 0.36%. The P content of CL wheat starch
remained constant for the first two hours but started to increase at four hours.
The P content progressively increased with incubation time, until it reached
0.56% at 24 hours, indicating that the molecules with higher P density were
more resistant to digestion. The regions with no or low bound P were
selectively removed, thus elevating the concentration of P in CL wheat starch
residue.
The 31P NMR spectra of phosphate derivatives of CL wheat starch and
their digestive residues after various a-amylase/amyloglucosidase incuba-
tion periods are also tabulated in Table 8.2. The effect of digestive enzymes
Table 8.2 Phosphorus content and levels of different phosphate esters of cross-linkedwheat starch and their digestive residues after various a-amylase/amyloglucosidaseincubation periods.
Phosphorus content (% dry basis)in the form of:
MSMP2
Digestiontime (h)
Totalphosphorus(%)�
Cyclic-MSMP1
C3 C6 DSMP3
0 0.36�0.01d 0.000 0.078 0.054 0.2281 0.36�0.02d 0.000 0.078 0.052 0.2302 0.36�0.00d 0.000 0.080 0.052 0.2294 0.38�0.02cd 0.004 0.090 0.057 0.2296 0.40�0.01c 0.004 0.089 0.061 0.2458 0.41�0.00c 0.004 0.099 0.062 0.24616 0.47�0.01b 0.004 0.104 0.069 0.29224 0.56�0.00a 0.005 0.136 0.080 0.338
�Values within columns bearing the same small letter superscript/s do not significantly differ
at (p<0.05).1Cyclic-MSMP, cyclic-monostarch phosphate.2MSMP, monostarch monophosphate positioned at C-3 and C-6.3DSMP, distarch monophosphate.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 151

CH08GR 07/29/2013 16:22:49 Page 152
on theCLwheat starch started after four hours of incubation, causing an increase
in the cyclic-monostarch monophosphate (cyclic-MSMP) and monostarch
monophosphate (MSMP). The distarch monophosphate (DSMP) of CL
wheat starch started to increase drastically after six hours of incubation. The
remaining periods of incubation showed a significant increase in MSMP
and DSMP, whereas cyclic-MSMP remained constant until the digestion
was terminated.
The results suggest that cyclic-MSMP and other phosphate esters
increased due to hydrolysis of starch molecules that may not be linked
to P molecules, leaving the remaining starch granules that were cross-
linked to P molecules. 31P NMR spectra of CL wheat starch after 16 hours
of digestion, as shown in Figure 8.1b , has signals of cyclic-MSMP, MSMP
positioned at C3 and C6, inorganic phosphate (Pi) and DSMP at d15.4 ppm, d 1–3 ppm, d 3.5 ppm and d 0 to –1 ppm, respectively. The
cyclic-MSMP had stronger peak detention as compared to the spectra
before digestion (Figure 8.1a), which was consistent with the quantified
cyclic-MSMP (Table 8.2).
8.3.2 Thermal properties
The gelatinization transition temperatures, consisting of To, Tp, and Tc as
well as DH of CL wheat starches at different times of a-amylase/amylo-
glucosidase incubation, are reported in Table 8.3. Gelatinization of all
starches took place between 72 �C (for To) to 94 �C (for Tc), and an
endothermic peak was observed in the DSC curves of all samples. The
data was slightly higher than that reported by Woo & Seib (2002), presum-
ably due to higher P content (0.36%) in the commercial CL wheat starch in
our study as opposed to the laboratory-prepared CL wheat starch (0.32%).
There was a slight increase in the Tp and Tc for digested CL wheat starch
after 16 and 24 hours incubation. The DH for CL wheat starch at all
incubation times was comparable to the DH before digestion, which was
inconsistent with potato starch (Jiang & Liu, 2002) and maize starch (Shresta
et al., 2012). An amylose-lipid complex peak was absent for CL wheat starch
at all incubation times. Similar values of the thermal parameters imply that
the remaining CL wheat starch residual after the a-amylase/amyloglucosi-
dase digestion was likely to be intact, with little or no effect on starch granule
crystallinity. Native wheat starch (Colonna et al., 1988), native barley starch
(Lauro et al., 1999) and native maize starch (Brewer et al., 2012) had been
reported to have decreased crystallinity after enzymatic digestion. The
current result showed that cross-linking of wheat starch aided in the retention
of starch crystallinity up to 24 hours amylolysis.
152 Resistant Starch

CH08GR 07/29/2013 16:22:49 Page 153
8.3.3 Starch granular morphology before and afterenzyme digestion
Figures 8.2 and 8.3 represent light microscopic and SEM micrographs
respectively of CL wheat starch after various a-amylase/amyloglucosidase
incubation periods. The SEM results were correlated with the microscopic
C6
Pi
MSDP
(a) C3
15 10 5 0 –5 –10 –15 –20
Cylic-MSMP
(b)
Chemical shift (ppm)
Figure 8.1 31P nuclear magnetic resonance spectra of dextrins prepared from cross-linked wheat starch before (a) and after 16 hours of a-amylase/amyloglucosidasedigestion (b). Cyclic-MSMP: cyclic-monostarch monophosphate; C-3: monostarchmonophosphate positioned at C3; C-6: monostarch monophosphate positioned atC6; MSDP: monostarch diphosphate; Pi: inorganic phosphate.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 153

CH08GR 07/29/2013 16:22:50 Page 154
evidence. However, the SEM micrographs provide a better observation of the
morphological changes of CLwheat starch granules after an incubation period
ranging from 0 to 24 hours.At the start (0 hours), the CL wheat starch granules had a smooth surface,
with round, shallow indentations of flower petal-like patterns which are
restricted to only some of the granules. In the first hour of digestion, mild
erosion on the surface of most starch granules was observed, indicating the
susceptibility of CL starch granules early during the incubation.
The progression of starch granule corrosion continued to be observed after
incubation from 1–4 hours. Scattered erosion on the surface of starch granules
increased and deepened with time. Most starch granules still, however,
remained intact, with the retention of the Maltese Cross pattern. After
incubation for six hours and eight hours, surface erosion of CL wheat starch
granules intensified, causing severe damage and the disappearance of granular
identity of some starch granules. The severity of damage on CL wheat starch
granules was more prominent after incubation for 16 hours and 24 hours, when
smaller fragments of broken starch granules were evident. However, some
starch granules remained intact and retained crystallinity, as depicted by the
Maltese Cross. This observation indicated that hydrolysis of CL wheat starch
granules by an a-amylase/amyloglucosidase cocktail was progressing con-
tinuously and did not stop until the 24 hours incubation was terminated.
CL wheat starch showed unequal granule degradation by digestive
enzymes at all incubation times. From 1–24 hours incubation, some of the
starch granules remained intact, while others had mild to extensive
Table 8.3 Thermal properties of cross-linked wheat starch and their digestive residuesafter various a-amylase/amyloglucosidase incubation periods.
Temperature (�C)Digestiontime (h) To Tp Tc DH (J/g)
0 72.0�0.2a 76.2�0.2c 89.8�0.7bc 10.7�0.3bc
1 72.2�0.1a 76.1�0.1c 90.4�0.2c 10.8�0.2bc
2 72.2�0.1a 76.1�0.1c 90.7�0.3c 11.8�0.1b
4 73.2�0.4a 77.3�0.5b 89.3�0.6bc 11.5�0.1b
6 72.3�0.3a 76.6�0.3c 91.5�0.5b 12.2�0.1a
8 72.3�0.1a 77.6�0.6b 91.7�0.5b 12.2�0.3a
16 73.2�0.1a 78.3�0.3ab 93.6�0.4a 11.3�0.4b
24 73.7�0.1a 78.9�0.0ab 93.8�0.3a 11.6�0.4b
Values within columns bearing the same small letter superscript(s) do not significantly differ at
(p>0.05).
154 Resistant Starch

CH08GR 07/29/2013 16:22:50 Page 155
Figure 8.2 Microscopic photos (40� objective) of cross-linked wheat starch atdifferent incubation times of a-amylase/amyloglucosidase digestion.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 155

CH08GR 07/29/2013 16:22:53 Page 156
Figure 8.2 (Continued)
156 Resistant Starch

CH08GR 07/29/2013 16:23:2 Page 157
Figure 8.3 Scanning electron micrographs (1000� and 3000� magnifications) ofcross-linked wheat starch at different times of a-amylase/amyloglucosidasedigestion.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 157

CH08GR 07/29/2013 16:23:8 Page 158
Figure 8.3 (Continued)
158 Resistant Starch

CH08GR 07/29/2013 16:23:11 Page 159
degradation. The same phenomenon was observed on the native wheat starch
granules (Figure 8.4 ), although the starch damage was more prominent at one
and two hours of digestion. This could be attributed to heterogeneous action of
the digestive enzymes on the wheat starch granules. Starch granules are also
Figure 8.4 Scanning electron micrographs (1000� and 3000� magnifications) ofnative wheat starch at one hour and two hours of a-amylase/amyloglucosidasedigestion.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 159

CH08GR 07/29/2013 16:23:13 Page 160
described as not equally susceptible to enzymatic hydrolysis, greatly depend-
ing on the adsorption manner of amylases on the granule and the method of
isolation of the starch (Colonna et al., 1988; Oates, 1997).
Colonna et al. (1988) had a similar observation of unequal amylase
digestion on native wheat starch, in which some granules had minimal pitting
and a fewwere severely cratered. However, all large granules were completely
degraded.
The mechanism of enzyme attack on starch granules is due to exo-
corrosion (Shresta et al., 2012), endo-corrosion or a combination of both
(Manelius et al., 1997; Apinan et al., 2007). The microscopic and SEM results
suggest that the starch hydrolysis by amylase progressed, starting from the
surface and proceeding inside of native wheat and CL wheat starch granules
by exo-corrosion. In addition, the starch granules were most probably digested
by the digestive enzymes using the side-by-side mechanism, as observed by
Zhang et al. (2006) and Shresta et al. (2012), in which the enzyme digested on
the amylose and amylopectin as well as the amorphous and crystalline regions
(Zhang et al., 2006).
Unlike digested maize starches, which are reported to have holes or tunnels
deepening into the interior of granules, or ‘Swiss cheese’ shell appearance
(Robyt, 2009; Lauren et al., 2012), CL wheat starch granules (Figure 8.3) and
native wheat starch (Figure 8.4) had a roughened superficial surface and
exposure of layered internal structures that intensified with increased incuba-
tion time, causing the formation of cavities on some starch granules. In
addition, CL wheat starch was theoretically obstructed by phosphate groups
within the granule surface pores and channels, inhibiting the diffusion of
amylase molecules (Thompson et al., 2011).
8.3.4 Digestibility
The extent of digestibility of CL wheat starch at different incubation times (up
to 24 hours) was determined using the AOAC Method 2009.01. Wheat starch
was also assayed using the same method as control, since wheat starch is
easily hydrolyzed by digestive enzymes (Bj€orck et al., 1986). The amount of
digested starch after each incubation period is shown in Figure 8.5. Both
starches showed rapid digestion at one hour, and the rate of digestion slowed
until the end of the incubation period. The latter observation is in agreement
with that of Bertoft & Manelius (1992), which showed two stages of starch
digestion, the first being the initial rapid hydrolysis, followed by a slower and
more constant rate of hydrolysis.
For wheat starch, the amount of starch digested was close to 70% at one
hour of incubation and 99% at six hours, indicating a high susceptibility of
160 Resistant Starch

CH08GR 07/29/2013 16:23:13 Page 161
wheat starch granules to hydrolysis. The SEM results (Figure 8.4) showed the
severity of starch hydrolysis at the early stage of digestion (one hour and two
hours). Our result is different from that reported by Colonna et al. (1988), who
observed 74% and 91% starch hydrolysis after incubation for ten hours and 21
hours, respectively. The difference could be due to enzyme source. Colonna
et al. (1988) used Bacillus subtilus a-amylase (17.5 Phadebas Unit/mgprotein), whereas the current study used a combination of pancreatic a-amy-
lase (50U/ml) and amyloglucosidase (3.4U/ml). Pancreatic a-amylase had
been suggested to be the most effective enzyme to digest native starch,
followed by barley, bacterial and fungal amylases (Kimura & Robyt, 1995).
When subjected to a cocktail of a-amylase and amyloglucosidase at 37 �Cfor 24 hours, CL wheat starch was digested at a slower rate than native wheat
starch and, by 24 hours, 84% of the CL starch was digested. The cross-linking
treatment decreased the susceptibility of starch granules to digestive enzymes
due to the stabilizing effect of granules with a high degree of cross-linking
(Seib & Woo, 1999; Woo et al., 2009), therefore causing a slower rate of
digestion. The amount of digested CL wheat starch was negatively correlated
with P content, but had little effect on starch crystallinity.
Digestion time (hours)
Native wheat starch CL wheat starch
Dig
este
d st
arch
(%
)
0 5 10 15 20 25
100
90
80
70
60
50
40
30
20
10
0
Figure 8.5 Digested starch of cross-linked (CL) wheat starch and native wheat starchat different incubation times of a-amylase/amyloglucosidase digestion.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 161

CH08GR 07/29/2013 16:23:13 Page 162
O’Brien&Wang (2009) proposed that amylopectin couldbemore reactive in
the cross-linking process, due to higher retained phosphate salts in the branched
structure of the polymer. Since amylopectin is comprisedof amorphous (tightly-
spaced branches) and crystalline regions (parallel glucans) (Oostergetel & van
Bruggen, 1989), phosphate groups may stabilize both amorphous and crystal-
line regions by promoting stronger interactions between the two regions.
Therefore, the undigested portion of the residual starch had increased stability
and was less susceptible to digestive enzymes, as depicted by the intact,
yet highly roughened starch granule surfaces observed in SEM micrographs
(Figure 8.3). Increased P content with increased incubation time showed that
phosphate groups were retained in the undigested residuals, reflecting the
incapability of digestive enzyme to cleave bonds close to a glucose linked
to a phosphate group.
Previous RS quantitation of CLwheat starch, using the EnglystMethod and
AOAC Method 991.43, showed 83% and 76%, respectively (Woo & Seib,
2002; Yeo & Seib, 2009; Thompson et al., 2011). While the Englyst Method
(incubation at 37 �C) and AOAC Method 991.43 (incubation at 100 �C) use120 minutes and 35 minutes of incubation time, respectively, AOAC Method
2009.01 incubates samples for 16 hours at 37 �C, yielding 25% RS content for
CL wheat starch.
The significantly lower RS content of CL wheat starch, as determined by
AOAC Method 2009.01, compared to the values obtained by the Englyst
Method and AOAC Method 991.43, raised questions about the validity of
AOAC Method 2009.01 to measure accurately RS content that is consistent
with in vivo human conditions. At 24 hours, RS content of CL wheat starch
further decreased to 16%, indicating that the digestive action of the
a-amylase and amyloglucosidase cocktail on CL wheat starch was still
progressing.
In humans, the transit time of food in the intestine takes 3–4 hours (Perera
et al., 2010). Long-term hydrolysis may lead to an increase in substrate
surface availability caused by amylase attack (Colonna et al., 1988). At 16
hours incubation time in AOAC Method 2009.01, a much lower yield of RS
content resulted, as indicated in the current study, and may not reflect in vivo
response, which is important in the food industry.
8.4 CONCLUSIONS
CL wheat starch was assayed using the digestive enzymes and incubation
conditions of AOAC Method 2009.01, and samples were collected at
predetermined times. The digestion of CL wheat starch continued to
162 Resistant Starch

CH08GR 07/29/2013 16:23:13 Page 163
increase up to 24 hours. The RS value obtained at 16 hours was lower than
the RS content measured by the Englyst Method and AOACMethod 991.43.
Microscopic and SEM results of the indigestible residues collected at the
predetermined times showed progressive degradation of CL wheat starch
granules. However, some starch granules remained intact, which explained
the comparable DSC data for CL wheat starch at all incubation times. The
mechanism of enzyme attack on the CL wheat starch was exo-corrosive, as
identified by the surface erosions and the non-existence of pinholes on the
granules.
Future studies will include assaying CL starches from different botanical
sources in order to understand further the mechanism of enzyme attack as
affected by cross-linking.
8.5 ACKNOWLEDGEMENTS
We thankMGP Ingredients, Inc. for donating the starch samples and Dr. Susan
Sun for the use of DSC. This is Contribution no. 13–233-J from the Kansas
Agricultural Experiment Station.
8.6 ABBREVIATIONS USED IN THIS CHAPTER
DH, gelatinization enthalpy; ANOVA, analysis of variance; CL, cross-linked;DSC, differential scanning calorimeter; P, phosphorus; RDS, rapidly digest-
ible starch; RS, resistant starch; SDS, slowly digestible starch; SEM, scanning
electron microscope; STMP, sodium trimetaphosphate; STPP, sodium tripo-
lyphosphate; Tc, conclusion temperature; To, onset temperature; Tp, peak
temperature.
REFERENCES
American Association of Cereal Chemists (2000). Approved Methods of the AACC, 17thed. Method 32–40. The Association: St. Paul, MN.
Anderson, I.H., Levine, A.S., Levitt, M.D. (1981). Incomplete absorption of the carbohy-drate in all purpose wheat flour. The New England Journal of Medicine 304, 891–892.
AOAC Official Methods of Analysis (2011). Method 2002.02 and Method 2009.01, 18thed. Rev 4. Horwitz, W., Latimer, G. (eds). AOAC International: Gaithersburg, Maryland.
Apinan, S., Yujiro, I., Hidefumi, Y., Takeshi, F., Myllarinen, P., Forssell, P., and Poutanen,K. (2007). Visual observation of hydrolyzed potato starch granules by a-amylase withconfocal laser scanning microscopy. Starch - Starke 59, 543–548.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 163

CH08GR 07/29/2013 16:23:13 Page 164
Asp, N.G. (1992). Preface: resistant starch. Proceedings of the 2nd plenary meeting ofEURESTA: European Flair Concerted Action No. 11 on Physiological Implications ofthe Consumption of Resistant Starch.Manual of European Journal of Clinical Nutrition46, S1.
Asp, N.G., Bjorck, I., Holm, J., Nyman, M., Siljestrom, M. (1987). Enzyme resistant starchfractions and dietary fibre. Scandinavian Journal of Gastroenterology 129, 29–32.
Berry, C.S. (1986). Resistant starch: formation and measurement of starch that survivesexhaustive digestion with amylolytic enzymes during the determination of dietary fiber.Journal of Cereal Science 4, 301–314.
Bertoft, E., Manelius, R. (1992). Amethod for the study of the enzymic hydrolysis of starchgranules. Carbohydrate Research 227, 269–283.
Bird, A.R., Lopez-Rubio, A., Shresta, A.K., Gidley, M.J. (2009). Resistant starch in vitroand in vivo: Factors determining yield, structure and physiological relevance. In:Kasapis, S., Norton, I.T., Ubin, J.B. (eds). Modern Biopolymer Science, 449–510.San Diego, CA, Academic Press.
Bj€orck, I., Nyman, M., Pedersen, B., Siljestr€om, M., Asp, N.G., Eggum, B. O. (1986). Onthe digestibility of starch in wheat bread – studies in-vitro and in-vivo. Journal of CerealScience, 4, 1–11.
Brewer, L. R., Cai, L., Shi, Y. C. (2012). Mechanism and enzymatic contribution to in vitrotest method of digestion for maize starches differing in amylose content. Journal ofAgricultural and Food Chemistry 60, 4379–4387.
Brown, I.L., Yotsuzuka, M., Birkett, A., Henriksson, A. (2006). Prebiotics, synbiotics andresistant starch. Journal of Japanese Association for Dietary Fiber Research 10, 1–9.
Champ, M. (1992). Determination of resistant starch in foods and food products:interlaboratory study. European Journal of Clinical Nutrition 46, S51–S61.
Colonna, P., Buleon, A., Lemarie, F. (1988). Action of Bacillus subtilis amylase on nativewheat starch. Biotechnology and Bioengineering 31, 895–904.
Englyst, H.N., Cummings, J.H. (1985). Digestion of the polysaccharides of some cerealfoods in the human small intestine. The American Journal of Clinical Nutrition 42,778–787.
Englyst, H.N., Kingman, S.M., Cummings, J.H. (1992). Classification and measurementof nutritionally important starch fractions. European Journal of Clinical Nutrition 46,S33–S50.
Faisant N., Champ M., Colonna P., Buleon A., Molis C., Langkilde A.M., Schweizer T.,Flourie B., Galmiche J.P. (1993). Structural features of resistant starch at end of thehuman small intestine. European Journal of Clinical Nutrition 47, 285–296.
Filer, L.J. (1971). Modified food starches for use in infant foods. Nutrition Reviews 29,55–59.
Hasjim, J., Lee, S.O., Hendrich, S., Setiawan, S., Ai, Y., Jane, J.L. (2010). Characterizationof a novel resistant starch and its effects on postprandial plasma-glucose and insulin-responses. Cereal Chemistry 87, 257–262.
Jiang, G., Liu, Q. (2002). Characterization of residues from partially hydrolyzed potato andhigh amylose corn starches by pancreatic a-amylase. Starch 54, 527–533.
Kimura, A., Robyt, J.F. (1995). Reaction of enzymes with starch granules: Kinetics andproducts of the reaction with glucoamylase. Carbohydrate Research 277, 87–107.
Langkilde, A.M., Andersson, H. (1995). In vivo quantitation of resistant starch inEURESTA reference materials using the ileostomy model. In: Asp, N.G., van Amels-voot, J.M.M., Hantvast, J.G.A.J. (eds). Proceedings of the concluding plenary meeting of
164 Resistant Starch

CH08GR 07/29/2013 16:23:13 Page 165
EURESTA31–32. European Flair Concerted Action on Resistant Starch, No. 11 (COST911).
Lauro, M., Forssell, P.M., Suortti, M.T., Hulleman, S.H.D., Poutanen, K.S. (1999).a-amylolysis of large barley starch granules. Cereal Chemistry 76, 925–930.
Lee, S., Prosky, L., DeVries, J. (1992). Determination of total, soluble, and insolubledietary fiber in foods: Enzymatic-gravimetric method, MES-TRIS buffer: collaborativestudy. Journal of AOAC International 75, 395–416.
Manelius, R., Qin, Z., Avall, A.K., Andtfolk, H., Bertoft, E. (1997). The mode of action ongranular wheat starch by bacterial a-amylase. Starch - Starke 49, 142–147.
McCleary, B.V. (2007). An integrated procedure for the measurement of total dietary fibre(including resistant starch), non-digestible oligosaccharides and available carbohy-drates. Analytical Bioanalytical Chemistry 389, 291–308.
McCleary, B.V., McNally, M., Rossiter, P. (2002). Measurement of resistant starch byenzymatic digestion in starch and selected plant materials: collaborative study. Journalof AOAC International 85, 1103–1111.
McCleary, B.V., DeVries, J.W., Rader, J. I., Cohen, G., Prosky, L., Mugford, D.C., Champ,M., Okuma, K. (2010). Determination of total dietary fiber (CODEX definition) byenzymatic-gravimetric method and liquid chromatography: collaborative study. Journalof AOAC International 93, 221–233.
Muir, J.G., O’Dea, K. (1992). Measurement of resistant starch: factors affecting the amountof starch escaping in vitro. American Journal of Clinical Nutrition 56, 123–127.
O’Brien, S., Wang, Y.J. (2009). Effects of shear and pH on starch phosphates prepared byreactive extrusion as a sustained release agent. Carbohydrate Polymers 77, 464–471.
Oates, C.G. (1997). Towards an understanding of starch granule structure and hydrolysis.Trends in Food Science and Technology 8, 375–382.
Oostergetel, G.T., Van Bruggen, E.F.J. (1989). On the origin of a low angle spacing instarch. Starch 41, 331–335.
Pendlington, A.W. (1999). Progress in the certification of five new food reference materialsby AOAC, Englyst and Uppsala methods of dietary fiber analysis. In: Cho, S.S., Prosky,L., Dreher, M. (eds). Complex Carbohydrate in Foods, 251–266. Marcel Dekker,Inc., NY.
Perera, A., Meda, V. Tyler, R.T. (2010). Resistant Starch: A Review of Analytical Protocolsfor Determining Resistant Starch and of Factors Affecting the Resistant Starch Contentof Foods. Food Research International 43, 1959–1974.
Robyt, J.F. (2009). Enzymes and their action on starch. In: BeMiller, J.N., Whistler, R.(eds). Starch: Chemistry and Technology, 3rd edition, 237–292. Academic Press, USA.
Sang, Y., Prakash, O., Seib, P.A. (2007). Characterization of phosphorylated cross-linkedresistant starch by 31P nuclear magnetic resonance (31P NMR) spectroscopy. Carbohy-drate Polymers 67, 201–212.
Sang, Y., Seib, P.A., Herrera, A.I., Prakash, O., Shi, Y.C. (2010). Effects of alkalinetreatment on the structure of phosphorylated wheat starch and its digestibility. FoodChemistry 118, 323–327.
Schweizer, T.F., Andersson, H., Langkilde, A.M., Reimann, S., Torsdottir, I. (1990).Nutrients excreted in ileostomy effluents after consumption of mixed diets with beansor potatoes. II. Starch, dietary fibre and sugars. European Journal of Clinical Nutrition44, 567–575.
Seib, P.A., Woo, K. (1999). Food-grade starch resistant to a-amylase and method ofpreparing the same. US Patent 5,855,946.
In Vitro Enzymatic Testing Method and Digestion Mechanism of Cross-linked 165

CH08GR 07/29/2013 16:23:14 Page 166
Sharma, A., Yadav, B.S., Ritika, B.Y. (2008). Resistant starch: physiological roles and foodapplications. Food Reviews International 24, 193–234.
Shresta, A.K., Blazek, J., Flanagan B.M., Dhital, S., Larroque, O., Morell, M.K., Gilbert, E.P., Gidley, M.J. (2012). Molecular, mesoscopic and microscopic structure evolutionduring amylase digestion of maize starch granules. Carbohydrate Polymers 90, 23–33.
Smith, R.J., Caruso, J. (1964). Determination of phosphorus. Methods in CarbohydrateChemistry 4, 42–46.
Stephen, A.M., Haddad, A.C., Phillips, S.F. (1981). Dietary and endogenous sources ofcarbohydrate substrate in the human colon. Gut 22, A893.
Thompson, L.U., Maningat, C.C., Woo, K., Seib, P.A. (2011). In Vitro Digestion of RS4-Type Resistant Wheat and Potato Starches, and Fermentation of Indigestible Fractions.Cereal Chemistry 88, 72–79.
Tovar, J., Bj€orck, I., Asp, N.G. (1990). Analytical and nutritional implications of limitedenzymic availability of starch in cooked red kidney beans. Journal of Agricultural andFood Chemistry 38, 488–493.
Wolf, B.W., Bauer, L.L., Fahey, G.C. (1999). Effects of chemical modification on in vitrorate and extent of food starch digestion: An attempt to discover a slowly digested starch.Journal of Agricultural and Food Chemistry 47, 4178–4183.
Woo, K.S. Seib, P.A. (2002). Cross-linked resistant starch: Preparation and properties.Cereal Chemistry 79, 819–825.
Woo, K.S., Maningat, C.C., Seib, P.A. (2009). Increasing dietary fiber in foods: The casefor phosphorylated cross-linked resistant starch, a highly concentrated form of dietaryfiber. Cereal Foods World 54, 217–223.
Wurzburg, O.B. (1986). Cross-linked starches. In: Wurzburg, O.B. (ed).Modified starches:Properties and uses, 41–53. CRC Press, Florida, USA.
Yeo, L.L., Seib, P.A. (2009). White pan bread and sugar-snap cookies containing wheatstarch phosphate, a cross-linked resistant starch. Cereal Chemistry 86, 210–220.
Zhang, G., Ao, Z., Hamaker, B.R. (2006). Slow digestion property of native cereal starches.Biomacromolecules 7, 3253–3258.
166 Resistant Starch

CH09GR 07/29/2013 16:34:3 Page 167
9 Biscuit Baking and Extruded SnackApplications of Type III ResistantStarch
Lynn Haynes, Jeanny Zimeri and Vijay AroraIngredient and Process Research, Mondelez International, USA
9.1 INTRODUCTION
Enzyme-resistant starch may be defined as the sum of starch and products of
starch degradation not absorbed in the small intestine of healthy individuals
(Eerlingen, 1994). It may be classified into four types, designated I to IV.
Physically inaccessible starch, locked in the plant cell, is classified as
type I. This type can be found in partially milled grains and seeds and legumes.
Native granular starch found in uncooked ready-to-eat starch-containing
foods, such as in bananas, is classified as type II resistant starch. Enzyme
susceptibility of type II resistant starch is reduced by the high density and
partial crystallinity of the granular starch.
The amount of type I and type II resistant starch is generally less than about
12% by weight, after grain milling and cooking, based on the amount of starch
contained in the starch source and in the formula. Type I and type II resistant
starches have low melting points and do not survive a baking process where
temperatures are substantially above 100�C and there is sufficient moisture in
the formula to gelatinize the starch (e.g. >20%). Type I and type II resistant
starch do not survive high shear processing such as extrusion, which would
disrupt granular integrity.
Starch may undergo a treatment process to obtain an indigestible starch
fraction. Depending upon the type of treatment, a type III resistant starch or a
type IV resistant starch may be produced. An indigestible starch fraction
which forms after certain heat-moisture treatments of the starch may be
present in, for example, cooked, cooled potatoes; this is type III enzyme-
resistant starch.
167
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

CH09GR 07/29/2013 16:34:3 Page 168
In type IV resistant starch, the enzyme resistance is introduced by
chemically modifying or thermally modifying the starch. For example,
glycosidic bonds other than a-(1–4) and a-(1–6) bonds may be formed by
heat treatments. Other glycosidic bonds may reduce the availability of starch
for amylolytic enzymes. In addition, the digestibility of starch may be reduced
by chemical cross-linking (Woo & Seib, 2002). Production and use of such
ingredients may be subject to legal limitations imposed by the US Food and
Drugs Administration.
Details of a patented process and flour replacer performance in a cookie
formula have been published (Haynes et al., 2002), utilizing a process
comprised of gelatinization, nucleation/propagation and heat treatment stages
for an amylose extender corn starch (aeWx VII). The characteristics of a
thermal-stable, shear-stable enzyme-resistant starch type III RS (X150) and
its use in biscuit baking and extrusion applications, without the formation
of glycosidic bonds other than a-(1–4) and a-(1–6) glycosidic bonds,
are described.
9.2 THERMAL CHARACTERISTICS OF HEAT-SHEARSTABLE RESISTANT STARCH TYPE III INGREDIENT
The starch-based ingredient is comprised of about 60% by weight enzyme-
resistant starch type III, which has a melting point of about 150�C andwhich is
formed by gelatinization, followed by at least one cycle of crystal nucleation
and crystal growth or propagation. In the critical cooling step, the gelatinized
starch is cooled to a crystal nucleating temperature of >60�C, above the
melting point of amylopectin starch such that the nucleation of amylolipid
crystalline complex is not favoured. A crystal propagating temperature of
about 130�C is used to maximize the formation of the heat-stable RS III,
which has a thermal melt temperature of 150�C. A debranching enzyme, such
as pullulanase may be used to increase the yield of the high-melting enzyme-
resistant starch type III.
Shown in Figures 9.1 and 9.2 are the MDSC curves for enzyme-resistant
starch type III ingredient and isolate. In the MDSC technique, the material
being analyzed is heated at a steady rate with a programmed saw-tooth pattern
of heating and cooling imposed upon the steady rate. The amplitude of the
fluctuation in temperature allows a more precise analysis of the equilibrium
melting point because it separates overlapping thermal events such as
irreversible decomposition. In Figure 9.1, a single crystalline melt is observed
at about 150�C for resistant starch type III bulk ingredient. Enzyme-resistant
168 Resistant Starch

CH09GR 07/29/2013 16:34:3 Page 169
Figure 9.1 Modulated differential scanning calorimetry (MDSC) curve for anenzyme-resistant starch type III ingredient (X150) or bulking agent, obtained froma single nucleation temperature of about 70�C and a propagation temperature ofabout 130�C.
Figure 9.2 MDSC curve for the isolated, heat-treated enzyme-resistant starch type IIIbulking agent.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 169

CH09GR 07/29/2013 16:34:4 Page 170
starch can be isolated from the bulk ingredient by a method adopted and
modified from the Prosky method for Total Dietary Fibre in baked goods set
forth in AOAC Method 991.43.Shown in Figure 9.2 is the MDSC thermal profile of enzyme-resistant type
III starch isolated from the bulk ingredient.
The second stage nucleation/propagation temperature cycling is followed
by a third stage involving heat treatment of the enzyme-resistant starch type III
product. The heat treatment is conducted at a temperature of about 130�C for
about one hour at a moisture content of 18%. Figure 9.3 shows a MDSC curve
for the isolated, heat-treated enzyme-resistant starch type III bulking agent.
The heat treatment increases the total dietary fibre content and improves
baking functionality. Corresponding MDSC curves for type II resistant starch
isolate (Figure 9.4) and type II resistant starch bulk ingredient after heat
treatment (Figure 9.5) show no high temperature melting.An enzyme-resistant starch type III bulking agent produced from gelatini-
zation and recrystallization at high temperatures, followed by heat treatment,
has about 60% total dietary fibre and is resistant to enzymes such as
a-amylase, b-amylase, amyloglucosidase and pancreatin, and it provides a
reduced calorie or low calorie, highly functional ingredient for replacing flour
in baking or extruded applications. As a result of the very highmelting point of
Figure 9.3 MDSC curve for an isolated enzyme-resistant starch type III ingredient(X150) obtained from the bulk.
170 Resistant Starch

CH09GR 07/29/2013 16:34:4 Page 171
Figure 9.4 MDSC curve for isolated, heat-treated enzyme-resistant granular starchtype II obtained by heat-treating granular type II (Novelose 240) bulking agent at130�C for one hour, followed by isolation of the RS from the heat-treated bulkingagent. (Novelose 240).
Figure 9.5 MDSC curve for heat-treated enzyme-resistant granular starch type II,obtained by heat treating an enzyme-resistant granular starch type II (Novelose 240)bulking agent.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 171

CH09GR 07/29/2013 16:34:5 Page 172
150�C, the enzyme-resistant starch ingredient is extremely heat-stable. The
resistant starch content of the type III resistant starch bulking ingredient
survives the high shear conditions of extrusion, enabling its use as a low
calorie, high fibre flour replacer in baking and extrusion applications.
9.3 APPLICATION TO BISCUIT BAKING: COOKIES
Cookies are baked products which generally contain three major ingredients,
i.e. flour, sugar and fat. Cookie quality attributes are their size and tender bite
(Miller & Hoseney, 1997). Also, cookies baked from soft wheat flour display
not only a large spread but also a uniform surface cracking pattern (Miller
et al., 1997). In cookie production, mixing disperses ingredients evenly and
promotes water absorption, rather than developing a true dough structure
(Huebner et al., 1999). Due to the high levels of fat and sugar, the development
of the gluten network is limited (Slade & Levine, 1994). Sugar is pre-
dissolved in water at a concentration in the formula water which exceeds
35% weight per weight, usually in the presence of formula fat. The dough
moisture is typically about 17–18%. The total solvent, sugar plus formula
water is about 64% (wt per cwt flour).
The cookie baking method used to evaluate resistant starch ingredient
performance is a standard wire-cut cookie baking method (AACC 10–53)
designed at the Nabisco Biscuit Company for the evaluation of ingredient
functionality and predictive correlation between sensory and mechanical
texture analysis (mechanical texture by Instron three-point bend or puncture
test). The AACC 10–53 was adopted as the approved method of the American
Association of Cereal Chemists after collaborative testing in 1992. Shown in
Table 9.1 is the formula used in the AACC 10–53 wire-cut cookie baking
method.
Cookie geometry and moisture bake-out is critical in the industrial setting,
which demands high through-put and automated packing of the product.
Correct cookie baking results in proper colour, taste and texture which, in turn,
influences consumer acceptance of the finished product. Achieving the right
geometry, colour and moisture balance is highly dependent upon ingredients
used in the formula.
In order to obtain a significant reduction in calories or an increase in total
dietary fibre per serving, usually 25% to 50% of the flour must be replaced. At
this level, solvent binding properties of flour replacers can disturb the colour
and moisture balance in the baked product. The water holding capacity of
conventional un-gelatinized wheat flour may be about 0.6 grams of water per
gram of dry flour. The bulking agent solvent holding properties ideally would
172 Resistant Starch

CH09GR 07/29/2013 16:34:6 Page 173
approximate that of the flour that is being replaced. Typically, gelatinized
starch used as a baking ingredient has an extremely high water holding
capacity of about 10 g of water per gram of starch (Pomeranz, 1964). The
resistant starch type III ingredient bulking agent, ground to a particle size
distribution the same as that of conventional wheat flour, has a water holding
capacity of less than 3 g of water per gram of dry matter. The lower water
holding capacity generally results in lower viscosity dough and has a
beneficial effect upon spread and baking characteristics. The water holding
is substantially unaltered by baking.
Baking performance of the resistant starch ingredients is tested by replac-
ing 50% by weight of the wheat flour in the standard dough formulation with
enzyme-resistant starch ingredient to obtain a blend. For each of the resistant
starch compositions, the amount of wheat flour replacement, the ingredient
type, the enzyme-resistant starch content or yield, the AOAC total dietary fibre
content, the cookie width and the MDSC enthalpy are set forth in Table 9.2.
Also in Table 9.2 are the L�a�b� colour measurements.
As shown in Table 9.2 and in Figure 9.6, the heat-treated RS Type III
(X150) ingredient exhibits a superior baking characteristic of greater cookie
spread. Along with cookie geometry, the product colour and moisture are
closest to control in the cookie made with RS type III and are likely to deliver
optimum commercial production efficiencies.
The enzyme resistance and low caloric value of the very high melting
enzyme-resistant type III starch ingredient component is substantially
unaltered by baking. The pure (or 100% by weight) enzyme-resistant starch
Table 9.1 AACC 10–53 wire-cut cookie formula.
Ingredient Weight (g)Percwt
Ingredientmoist
Ingredientdry weight(g)
Low trans shortening 90 40.00 0.20% 89.82Sucrose, fine gran. 94.5 42.00 0.50% 94.03Salt 2.81 1.25 0.50% 2.8Non-fat dry milk 2.25 1.00 4.00% 2.16High fructose corn syrup 3.38 1.50 29.00% 2.4Ammonium bicarbonate 1.13 0.50 99.50% 0.01Sodium bicarbonate 2.25 1.00 50.00% 1.13Flour 225 100.00 13.00% 195.75water 49.5 22.00 100.00% 0Total weight 470.82 Dry solids 388.08Total moist 17.57%
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 173


CH09GR 07/29/2013 16:34:7 Page 175
type III has a caloric value of essentially zero, or less than about 0.5 calories/
gram even after baking. The caloric value for starch which is not resistant
starch is about 4 calories per gram. Thus, a resistant starch type III ingredient
or bulking agent with at least a 30% yield of RS type III will exhibit a caloric
value of less than about 2.8 calories/gram (0.70� 4 cal/gþ 0.30� 0
cal/g¼ 2.8 calories/gram). The resistant starch type III ingredient or bulking
agent exhibits excellent baking characteristics in terms of oven spread, edge
contour, oil release, surface cracking, odour, colour or browning, mouth-feel
and texture. It may be used alone or in combination with other fibres or whole
grain flour to produce a healthy, wholesome product.
9.4 CRACKER BAKING
Chemically leavened cracker dough formulas differ from cookie formulas in
two important aspects. First, the level of sugar in a cracker formula, measured
as wt sugar/wt sugarþ formula water, is lower at about 28%. Second, the total
solvent (sugarþwater) per cwt of flour is lower at about 48% (wt per cwt
flour), most of which is water. The cracker formula moisture is about 35% of
formula, significantly higher than cookie formulas. The cracker formula
conditions of low sugar and higher formula moisture allow for the develop-
ment of wheat flour gluten, such that a cohesive, elastic dough can be formed
Figure 9.6 AACC 10–53 wire cut cookies made with 50% replacement of flour withresistant starch ingredient.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 175

CH09GR 07/29/2013 16:34:8 Page 176
into a thin sheet from which cracker dough pieces are cut. Ingredients with
high water absorption are undesirable, because a tough, dry, less elastic dough
results. To compensate, more formula water is added, which can contribute to
longer bake time, requiring greater amounts of heat energy to remove the
excess moisture.
Type II granular resistant starch, Type III resistant starch (X150), made
from preferred process as described (gelatinization, nucleation/propagation
and heat treatment stages), and Type IV chemically modified resistant starch
(Fibersym1 70 or its old name Fiberstar 70) are evaluated in a cracker
formula. Table 9.3 shows the formula amounts for sugar, which calculates to
24% (weight sugar divided by weight of sugar plus formula water), total
solvent (sugarþ syrupþwater) per cwt of flour of about 48% and the total
formula moisture of about 35%. The cracker dough is sheeted and dough
pieces cut and weighed and baked.
As a result of the lower solvent holding capacity of the high melting point,
thermally stable type III resistant starch (X150), formula water requirements
are lower and superior baking characteristics are achieved. Although there is
sufficient moisture to gelatinize starch, due to the high melting point of Type
III RS made by the preferred process there is no loss of enzyme resistance
upon baking. Differences in dough sheet toughness and extensibility, com-
pared to control flour, were noted for all dough made with resistant starch.
Reducing the flour content of the formula reduces the amount of flour gluten
protein available for dough development. Figure 9.7 shows the sheeting
property of dough when flour is replaced at the 50% level.
Replacement of flour with resistant starch makes the dough less tough/
cohesive and less extensible. Using stronger flour with the resistant starch
does help to improve dough toughness, although there is still a notable
Table 9.3 Model formula for chemically leavened cracker baking test.
Ingredients Total weight Per cwt
Stage 1 Pre-dissolved sucrose 6.12 11.08Stage 1 Corn starch 3.1 5.61Stage 1 Salt 0.61 1.10Stage 1 Mono calcium phosphate 0.31 0.56Stage 1 Sodium bicarbonate 0.16 0.29Stage 2 Low trans oil 2.75 4.98Stage 2 Tap water (110�F) 18.87 34.17Stage 2 High fructose corn syrup 1.84 3.33Stage 3 Flour 55.23 100.00
Total weight 88.99 161.13Total %moist 0.3532
176 Resistant Starch

CH09GR 07/29/2013 16:34:9 Page 177
decrease from control. Addition of vital wheat gluten shows increase in
toughness and extensibility of the dough sheet, as vital wheat gluten level
increases, toughness and extensibility also increase.
The finished product is baked for the same length of time and the moisture
measured by percentage weight loss. Resistant starch type II (Novelose 260)
requires more water to form dough, so current bake time results in out-of-
specification cracker moisture. Tougher dough tends to result in crackers with
higher moisture. Type III resistant starch (X150) at a 50% level demonstrates a
mechanical break force closest to control, as shown in Figure 9.8.
Figure 9.7 Force required to stretch dough and distance stretched before breakingare shown for resistant starch ingredient used to replace flour before and after doughresting. Note: additional water (19.5g) is added to the Novelose sample becausedough did not develop with formula amount.
Figure 9.8 Finished product modulus and stress force for cracker made withresistant starch replacing 50% of regular flour. Final moisture contents of finishedproducts are also shown.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 177

CH09GR 07/29/2013 16:34:11 Page 178
Retention of enzyme resistance during baking was confirmed by AOAC
991.43 fibre analysis of the finished cracker product. The results of fibre
analysis are shown in Table 9.4.
9.5 EXTRUDED CEREAL APPLICATION
Ready-to-eat (RTE) cereals are typically constituted by a flour blend (wheat
flour, ground cereal fines, barley malted flour), which constitutes approxi-
mately 96% by weight of the solids, 2% granulated sugar, 1.25% fine granular
salt and 0.5% of other minor ingredients. Enzyme-resistant starch type III
(X150) has been used to replace up to 50% of the flour blend in RTE cereals.
In the processing of RTE cereals, the solid ingredients are extruded using
a twin-screw extruder at a moisture content of approximately 9% moisture
(w.b.) under medium-shear screw profile, configured as shown in Figure 9.9.
The extrudate expands at the exposure to atmospheric pressure just outside of
the die, referred to as ‘flashing off’ of moisture.
The very-high-melting, high-shear-surviving enzyme-resistant starch is
substantially unaltered by extrusion, i.e. it remains substantially enzyme
resistant and exhibits a reduced calorie value of less than about
1.6–2.0 kcal/gram (60–50% by weight RS type III, having a melting point
or endothermic peak temperature of at least 140�C, as determined by MDSC),
as determined by fibre analysis. Enthalpy values for the high-melting enzyme-
resistant starch in the cereal are typically in the order of 6 Joules/g (based on
grams of resistant starch in the flour blend), at a temperature of from 130�C to
about 165�C. This is highly advantageous for producing reduced-calorie
extruded cereals because, if the crystal structure that provides enzyme-
resistance is destroyed or melts during extrusion, and if the crystal recrys-
tallizes into a lower-melting form which is not enzyme-resistant, then calorie
reduction will not be achieved in the extruded product. For example, when
Table 9.4 Results of AOAC method 991.43 fibre analysis in finished crackers.
ProductFlourreplacer
Productflour
Flourreplacement
% Dietaryfibre(theoretical)
% Dietaryfibre(actualanalysis)
% Fibreretentionafterprocessing
Cracker,
unoiled
Control 75.8% 0% 2.2 2.4 109%
X150 75.8% 50% 19 17.3 91%
Novelose
260
75.8% 50% 22.7 19.9 88%
178 Resistant Starch

CH09GR 07/29/2013 16:34:11 Page 179
using a type II or type IV resistant starch, the calorie reduction would be less,
since these types of resistant starch do not survive high temperature or high
shear processing, such as extrusion.
The RS type III ingredient gives the extruded RTE cereal excellent
extrusion characteristics in terms of density, moisture content, texture (brit-
tleness) and cell structure.
9.5.1 Preparation of extruded RTE cereal and analysis
The extrusion functionality of an enzyme-resistant starch type III prepared in
accordance to the above procedure was compared to the extrusion function-
ality of a commercially available, enzyme-resistant starch type II ingredient
(Hi-MaizeTM 1043, produced by National Starch and Chemical Co.) and
a commercially available, enzyme-resistant starch type IV ingredient
(Fibersym1 70, produced by MGP Ingredients) using conventional, non-
heat-treated wheat flour as Control.
Hi-MaizeTM 1043 has the following characteristics, as claimed by National
Starch and Chemical Co.:
� Moisture: 13% maximum;� Total dietary fibre (AOAC method 991.43): 60% minimum (dry basis);� Calories: approx. 1.6 kcal/g.
Figure 9.9 Medium-shear screw profile used in extruder.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 179

CH09GR 07/29/2013 16:34:11 Page 180
Fibersym1 70 (also known by its old name as Fiberstar 70) has the
following characteristics, as claimed by MGP:
� Moisture: 10.6% typical;� Total dietary fibre (AOAC method 991.43): 70% minimum (dry basis);� Calories: approx. 3.6 kcal/g (not corrected for insoluble fiber)
Extrusion functionality was evaluated bymeasurement of density, moisture
content, texture (brittleness) and cell structure. Resistant starch ingredients
that resulted in cereal properties as close to the properties achieved with the
wheat flour control were considered to have the best extrusion functionality.
Total dietary fibre was used to determine thermal and shear stability of the
resistant starch ingredients.
Control was produced following a standard formula for extruded, expanded
RTE cereals. In test samples, 50% of the total flour weight in the formula
(including wheat flour, ground cereal fines and barley malted flour) was
replaced by a resistant starch, namely RS III (X150) (variable 1), Hi-Maize
1043 (variable 2), or Fibersym 70 (variable 3), with the objective of testing
their functionality as flour replacers.
9.5.1.1 Formulas
The formulas are shown in Table 9.5.
9.5.1.2 Extrusion
The operating conditions used in the extruder to produce the cereal samples
are shown in Table 9.6.
9.5.1.3 Results
As shown in Table 9.7, the bulk density of the three test samples was lower
than that of the control, in the following descending order: Fibersym
70>RSIII>Hi-Maize 1043. Bulk density is an important parameter when
dealing with packing of a standard weight of cereals in a standard volume.
Also as shown in Table 9.7, resistant starch raw materials RSIII, Hi-Maize
1043 and Fibersym 70 had MDSC enthalpies of 8.39 J/g, 0 J/g and 0 J/g,
respectively, at temperatures above 140�C. Hi-Maize 1043 presented an
endotherm at 101.6�C and an enthalpy of 4.4 J/g, while Fibersym 70 presented
an endotherm at 74.9�C and an enthalpy of 11.71 J/g, representing low-
melting resistant starches.
180 Resistant Starch



CH09GR 07/29/2013 16:34:16 Page 183
The control did not present a MDSC enthalpy above 140�C, indicating thatno enzyme-resistant starch was present and that none was created during
processing. Cereal samples containing RSIII presented a MDSC enthalpy of
9.05 J/g (based on g of RSIII in the flour blend), comparable to that of the
RSIII raw material, indicating that the RSIII ingredient was stable during
extrusion. In addition, it presented a high melting point of 151.2�C, corre-sponding to enzyme-resistant starch ingredients. Since it remained unchanged
during extrusion, RSIII was very identifiable in the final product.
Cereal samples containing Hi-Maize 1043 presented a MDSC enthalpy of
4.35 J/g (based on g of Hi-Maize 1043 in the flour blend) at 144.7�C, value,which was equivalent to the enthalpy exhibited by the Hi-Maize 1043
ingredient, but presented a shift in temperature of about 45�C with respect
to the original melting temperature of 101.6�C in the Hi-Maize 1043
ingredient. Thus, generation of crystalline resistant starch, as expected
from high temperature/moisture processes, occurred during extrusion and/
or DSC analysis. A smaller enthalpy than that for RSIII represented a sample
with less degree of crystallinity and, thus, less content of enzyme-resistant
starch. In addition, the generated enzyme-resistant starch had a lower melting
point than that of RSIII.
Cereal samples containing Fibersym 70 presented a MDSC enthalpy of
1.2 J/g (based on g of Fibersym in the flour blend) at 122�C, a value that was
Table 9.7 Final product property measurements.
Bulkdensity(g/cm3)
Moisture(%)
MDSC enthalpy@ >140�C(J/g)�
RS meltingpeaktemperature(�C) L�, a�, b�
RSIII, raw material – – 8.39 151.2 –
Hi-MaizeTM 1043, raw
material
– – 0 101.6 –
Fibersym�R 70, rawmaterial
– – 0 74.9 –
Control cereal 0.452 9.73 0 – 70.61, 3.81,
25.33RSIII cereal, 50% flour
replacement
0.309 9.99 9.05 151.2 68.49, 4.62,
25.59
Hi-MaizeTM 1043
cereal, 50% flour
replacement
0.195 10.02 4.35 144.7 75.29, 2.47,
22.26
Fibersym�R 70 cereal,
50% flour
replacement
0.365 7.81 0 122 73.39, 3.21,
26.74
� Joules per g of flour, g of RSIII in flour blend, g of Hi-MaizeTM in flour blend, or g of Fibersym�R inflour blend, respectively.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 183

CH09GR 07/29/2013 16:34:16 Page 184
very different in both magnitude and temperature from the original Fibersym
70 raw material. Since Fibersym 70 is a cross-linked starch, as opposed to
granular and retrograded starches in the case of Hi-Maize 1043 and X-150,
enthalpy might not correlate well with the amount of enzyme-resistant starch.
A better measurement would correspond to amount of total dietary fibre, as
will be discussed later.
As shown in Table 9.8, the theoretical total dietary fibre content in the
resistant starches corresponded to: RSIII¼ 50%; Hi-Maize 1043¼ 62%;
Fibersym 70¼ 70% (although it assayed at 80% using the AOAC 991.43
method), based on fibre content of ingredient declared in specifications.
Although cereal samples containing Hi-Maize 1043 had a higher theoreti-
cal dietary fibre content than cereal samples containing RSIII, calculated
based on fibre content of the ingredient declared in the specifications, the
measured AOAC total dietary fibre for cereal samples containing RSIII was
higher than that for cereal samples containing Hi-Maize 1043. The calculated
% fibre retention (i.e. fibre that survived processing plus that being generated
during processing) corresponded then to 96% for RSIII cereals and only 67%
for Hi-Maize 1043 cereals.
Table 9.8 Fibre content of extruded, RTE cereals.
% totaldietaryfibre,theoreticala
% AOACtotal dietaryfibreb
% fibre inRS ingredientafterprocessingc
% fibreretentionin RSingredientd
RSIII ingredient 50 – – –Hi-MaizeTM ingredient 62 – – –Fibersym�R 70
ingredient70, min. 80 – –
Control cereal 5.58 7.7 – –RSIII cereal, 50% flour
replacement23.3 22.4 48 96
Hi-MaizeTM 1043cereal, 50% flourreplacement
28.9 19.4 42 67
Fibersym�R 70 cereal,50% flourreplacement
37.3 12.60 27 34
aCalculated based on fibre content of ingredient declared in specifications.bAs measured by AOAC 991.43.cCalculated based on measured AOAC TDF in final product.dCalculated based on % total dietary fibre in RS ingredient, declared in specifications, vs. % fibre in RS
ingredient after processing.
184 Resistant Starch

CH09GR 07/29/2013 16:34:16 Page 185
Although cereal samples containing Fibersym 70 had the highest theoreti-
cal dietary fibre content than the rest of the samples, calculated based on
measured fibre content of the ingredient (80%), the measured AOAC total
dietary fibre for cereal samples containing Fibersym 70 was the lowest of all
the samples. The calculated percentage fibre retention corresponded to only
34%, which suggests that Fibersym 70 did not completely survive high
temperature and high shear conditions present during extrusion.
Cereal brittleness is presented in Figure 9.10. A high peak force and low
peak distancewould correspond to a brittle sample, while a low peak force and
high peak distancewould correspond to a less brittle sample. The x- and y-axis
error bars represent �1 standard deviation, while the diamonds represent
averages. Overlapping error bars represent samples that were not significantly
different from one another. In this case, control, X-150 and Fibersym
70 samples did not present a significantly different peak force or a signifi-
cantly different peak distance, indicating that they possessed a similar
brittleness. Hi-Maize 1043 samples showed a larger peak distance than the
rest of the samples, indicating a lower brittleness. Hi-Maize 1043 samples did
not present a significantly different peak force than RSIII or Fibersym
70samples, but theywere significantly softer (lowerpeak force) than the control.
As shown in Figure 9.11, air cells were more numerous and larger in the
control than in the rest of the samples, a characteristic which can be correlated
to bulk density. Cell structure is the result of expansion during extrusion,
governed by moisture content, moisture flash-off and flour blend (matrix)
physicochemical properties.
As shown in Figure 9.12, cereal samples containing RSIII showed superi-
ority in bowl life when compared to the other two resistant starch-containing
Figure 9.10 Extruded cereal brittleness.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 185

CH09GR 07/29/2013 16:34:16 Page 186
samples (i.e. RSIII samples force vs. distance (deformation) behaviour after
soaking in 8�C water for 30 minutes was closer to the control than that of
Fibersym 70 and Hi-Maize 1043). For example, if a distance of 10mm was
chosen on the force vs. distance plot, it can be seen that the control was harder
(higher force) than the rest of the samples, and that RSIII was harder (higher
force) than Hi-Maize 1043 and Fibersym 70. The same behaviour was
Figure 9.11 Cell structure of extruded cereals.
0 5 10 15 20 25 30 35
300000
250000
200000
150000
100000
50000
0
–50000
Force (g)
Distance (mm)
Control RS III
Hi-Maize™
Fibersym® 70
Figure 9.12 Extruded cereal bowl life.
186 Resistant Starch
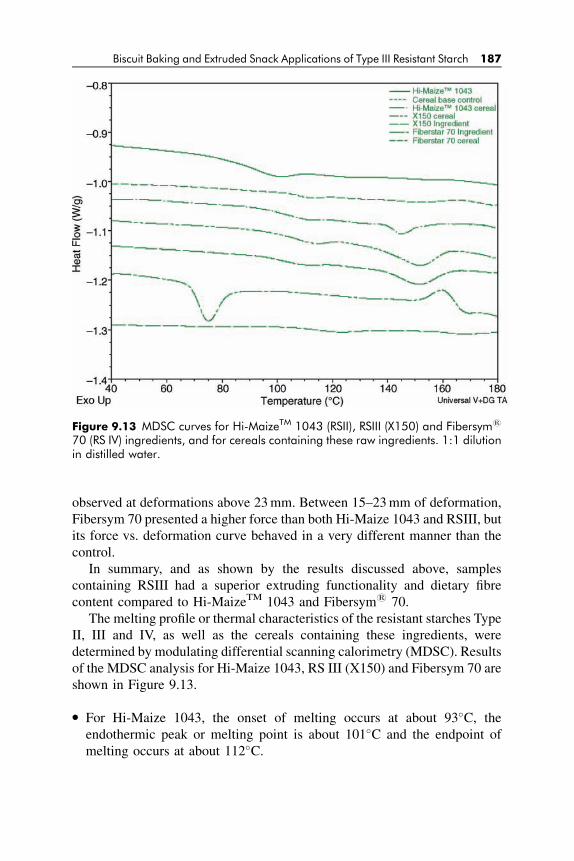
CH09GR 07/29/2013 16:34:17 Page 187
observed at deformations above 23mm. Between 15–23mm of deformation,
Fibersym 70 presented a higher force than both Hi-Maize 1043 and RSIII, but
its force vs. deformation curve behaved in a very different manner than the
control.
In summary, and as shown by the results discussed above, samples
containing RSIII had a superior extruding functionality and dietary fibre
content compared to Hi-MaizeTM 1043 and Fibersym1 70.
The melting profile or thermal characteristics of the resistant starches Type
II, III and IV, as well as the cereals containing these ingredients, were
determined by modulating differential scanning calorimetry (MDSC). Results
of the MDSC analysis for Hi-Maize 1043, RS III (X150) and Fibersym 70 are
shown in Figure 9.13.
� For Hi-Maize 1043, the onset of melting occurs at about 93�C, the
endothermic peak or melting point is about 101�C and the endpoint of
melting occurs at about 112�C.
Figure 9.13 MDSC curves for Hi-MaizeTM 1043 (RSII), RSIII (X150) and Fibersym1
70 (RS IV) ingredients, and for cereals containing these raw ingredients. 1:1 dilutionin distilled water.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 187

CH09GR 07/29/2013 16:34:19 Page 188
� For RSIII, the onset of melting occurs at about 135�C, the endothermic peak
or melting point is about 151.2�C and the endpoint of melting occurs at
about 165�C.� For Fibersym 70, the onset of melting occurs at about 68�C, the endother-
mic peak or melting point is about 75�C and the endpoint of melting occurs
at about 95�C.
The software calculates the enthalpy of the endothermic peak in J/g.
Results of the MDSC analysis for cereals containing resistant starch
ingredients are also shown in Figure 9.13.
� For Hi-Maize-containing cereal, the onset of melting occurs at about
135�C, the endothermic peak or melting point is about 145�C and the
endpoint of melting occurs at about 155�C.� For RSIII-containing cereal, the onset of melting occurs at about 135�C, the
endothermic peak or melting point is about 152�C and the endpoint of
melting occurs at about 165�C.� For Fibersym 70-containing cereal, the onset of melting occurs at about
107�C, the endothermic peak or melting point is about 122�C and the
endpoint of melting occurs at about 133�C.
Figure 9.14 MDSC curve for Novelose 330 (RSIII). 1:1 dilution in distilled water.
188 Resistant Starch

CH09GR 07/29/2013 16:34:20 Page 189
MDSC results for Novelose 330 (National Starch & Chemical Co.,
Bridgewater, NJ), another commercially available type III resistant starch,
are shown in Figure 9.14. The onset of melting occurs at about 104� C, theendothermic peak or melting point is about 118.2 and the endpoint of melting
occurs at about 135� C. Thus, even though this is a type III resistant starch, itmelts at a lower temperature than RSIII, indicating that it would not survive
high-temperature extrusion conditions such as those used in breakfast cereal
manufacture, having very little functionality as a fibre.
REFERENCE S
Anderson, S., Haynes, L., Zhou, N., Slade, L., Levine, H., Kweon, M., Locke, J., Zimeri, J.(2005). Reduced-Calorie Flour Containing Type 3 Resistant Starch Used In ModelCracker System. AACCI Annual Meeting, September 11, 2005, Orlando, FL.
Eerlingen, R. (1994). Formation, Structure and Properties of Enzyme Resistant Starch.Doctoral dissertation. Katholieke Universiteit Leuven. Available at: http://www.biw.kuleuven.be/lmt/labolmc/DOCS/Eerlingen.htm
Haynes, L, Gimmler, N., Locke, J.P.III, Kweon, M-R, Slade, L., Levine, H. Production ofan enzyme-resistant starch for a reduced calorie flour replacer. United States Patent,US6352733, March 5, 2002.
Huebner, F.R., Bietz, J.A., Nelsen, T., Bains, G.S., Finney, P.L. (1999). Soft Wheat Qualityas Related to Protein Composition. Cereal Chemistry 76(5), 650–655.
Kweon, M., Haynes, L., Locke, J., Slade, L., Levine, H. (2004). Baking functionality of areduced-calorie flour replacer containing type-3 resistant starch. AACC/TIA JointMeeting, Sep 19 – 22, 2004, San Diego, CA.
Miller, R.A., Hoseney, R.C. (1997). Factors in Hard Wheat Flour Responsible for ReducedCookie Spread. Cereal Chemistry 74(3), 330–336.
Miller, R.A., Hoseney, R.C., Morris, C.F. (1997). Effect of Formula Water Content on theSpread of Sugar-Snap Cookies. Cereal Chemistry 74(5), 669–671.
Pomeranz, Y. (1964). Wheat: Chemistry and Technology, Vol. II, 333. Washington StateUniversity: Pullman, WA.
Slade, L., Levine, H. (1994). Structure-function relationships of cookie and crackeringredients. In: Faridi, H. (ed). The Science of Cookie and Cracker Production,23–141. Chapman & Hall: New York.
Woo, K.S., Seib, P.A. (2002). Cross-Linked Resistant Starch: Preparation and Properties.Cereal Chemistry 79(6), 819–825.
Zimeri, J.E., Haynes, L., Olson, A., Arora, V.K., Slade, L., Levine, H., Kweon, M.Production of low calorie, extruded, expanded foods having a high fiber content.United States Patent, US7648723, January 19, 2010.
Biscuit Baking and Extruded Snack Applications of Type III Resistant Starch 189

CH09GR 07/29/2013 16:34:20 Page 190

3GCH10 07/20/2013 8:53:8 Page 191
10 Role of Carbohydrates in thePrevention of Type 2 Diabetes
Thomas M.S. WoleverDepartment of Nutritional Sciences, University of Toronto, Canada; Divisionof Endocrinology and Metabolism, St. Michael’s Hospital, Canada
10.1 INTRODUCTION
The main objectives of this chapter are to review basic facts about diabetes
mellitus (definition, types, prevalence and risk factors) and to understand how
dietary carbohydrates affect the risk of developing type 2 diabetes and some of
the mechanisms by which dietary carbohydrates may be able to prevent type
2 diabetes, namely reduced postprandial glucose responses and increased
colonic fermentation.
10.2 BACKGROUND
10.2.1 Definition of diabetes
Diabetes is a condition in which the body either cannot produce insulin, or
cannot use the insulin it produces, resulting in a rise in the blood glucose
concentration. Therefore, the diagnosis of diabetes is made based on the
finding of high blood glucose. For men and non-pregnant women, diabetes is
present if any one of the following three conditions is met (Canadian Diabetes
Association Clinical Practice Guidelines Expert Committee, 2008):
� fasting plasma glucose equal to or greater than 7.0mmol/l (126mg/dl);� random (i.e. at any time of the day) plasma glucose equal to or greater than
11.1mmol/l (200mg/dl); or� a plasma glucose equal to or greater than 11.1mmol/l (200mg/dl) two hours
after consuming 75 g glucose (oral glucose tolerance test).
191
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH10 07/20/2013 8:53:8 Page 192
10.2.2 Types of diabetes
There are three main types of diabetes: type 1, type 2 and gestational diabetes:
� Type 1 diabetes, formerly known as insulin-dependent diabetes or juvenile
onset diabetes, is an autoimmune disease in which the insulin-producing
cells of the pancreas are destroyed, so that the body no longer produces any
– or only a very small amount of –insulin. Type 1 diabetes usually develops
in children or adolescents, affects about 5–10% of people with diabetes and
is treated with lifelong injections of insulin.� Type 2 diabetes, formerly known as noninsulin-dependent diabetes or
maturity onset diabetes, occurs when the pancreas cannot produce enough
insulin to meet the body’s needs and/or the body is unable to respond
normally to the actions of insulin (insulin resistance). Type 2 diabetes
usually develops in middle-aged or elderly adults (although it can occur in
children), affects about 90–95% of people with diabetes and is treated by
diet and exercise and, if necessary, oral medications and/or insulin.� Gestational diabetes is high blood glucose that develops during pregnancy.
It affects about 2–4% of pregnant women and is treated by diet and, if
necessary, insulin. Blood glucose levels usually return to normal after
delivery.
10.2.3 Complications of diabetes
Diabetes has serious complications, including heart disease, kidney disease,
eye disease, impotence and nerve damage, which are caused by high blood
glucose. Treatment of diabetes to reduce blood glucose levels is known to
prevent, delay the onset of or reduce the severity of these complications.
10.2.4 Prevalence of diabetes
Diabetes is a major public health issue because of the increased morbidity and
mortality associated with it, the high cost of treatment, and the fact that its
prevalence is rising rapidly in all parts of the world. In 1998, it was estimated
that there were 135 million people in the world with diabetes and that this
number would more than double to 300 million by 2025 (King et al., 1998).
However, the prevalence of diabetes is increasing faster than previously
expected; it is now estimated that there are 285 million people worldwide
who are affected by diabetes, and that this number is increasing by seven
million per year to reach 390 million by 2025 (Canadian Diabetes Associa-
tion, 2010).
192 Resistant Starch

3GCH10 07/20/2013 8:53:8 Page 193
10.2.5 Risk factors for type 2 diabetes
Since most people with diabetes have type 2 diabetes and since diet plays a
larger role in the prevention of type 2 than type 1 diabetes, the rest of this
chapter is concerned with type 2 diabetes. Consideration of dietary strategies
to prevent type 2 diabetes is particularly important for people at high risk of
developing the condition. Major factors associated with increased risk for type
2 diabetes include (Canadian Diabetes Association Clinical Practice Guide-
lines Expert Committee, 2008):
� being overweight or obese (body mass index over 25 kg/m2);� age over 40 years; family history of diabetes in a first degree relative (parent
or sibling);� being a member of a high risk population (Aboriginal, African, Hispanic,
Asian or South Asian);� having previously had gestational diabetes or high blood glucose (impaired
fasting glucose which is fasting glucose between 5.6–6.9mmol/l or
100–125mg/dl or impaired glucose tolerance, which is blood glucose
two hours after oral glucose tolerance test between 7.8–11.0mmol/l or
140–199mg/dl);� having high serum triglycerides or low serum high-density lipoprotein
(HDL) cholesterol;� the presence or history of peripheral-, coronary- or cerebro-vascular
disease;� the presence or history of schizophrenia.
10.3 CARBOHYDRATES AND RISK OF TYPE 2 DIABETES
With respect to the prevention of type 2 diabetes, there is strong evidence that
the quality of dietary carbohydrate is more important than the quantity. There
is little evidence that the amount of carbohydrate consumed influences risk for
type 2 diabetes. In six out of seven recent prospective studies in which it was
examined, the amount of carbohydrate consumed had no significant effect on
type 2 diabetes (see Table 10.1).
10.3.1 Markers of carbohydrate quality
The quality of dietary carbohydrate has traditionally been assessed by
considering simple versus complex carbohydrates. However, there is no
evidence that a high intake of simple carbohydrates causes diabetes (Janket
Role of Carbohydrates in the Prevention of Type 2 Diabetes 193

3GCH10 07/20/2013 8:53:9 Page 194
et al., 2003; Hodge et al., 2004), although consumption of sugar-sweetened
beverages may increase diabetes risk (Schulze et al., 2004a).
Measures of carbohydrate quality which do influence risk for type
2 diabetes include cereal fibre and glycemic index (Table 10.1). Cereal fibre
is considered here to be a qualitative indicator of carbohydrate intake,
because it represents a small proportion of total carbohydrate (usually less
than 10%) and is a marker of the types of carbohydrates consumed. In six out
of seven studies in which it was examined, a high intake of cereal fibre was
associated with a significant reduction in the relative risk of developing type 2
diabetes; the mean reduction in risk was about 30%.
The other marker of carbohydrate quality is glycemic index (GI). GI is
defined as 100� F/R, where F is the incremental area under the glycemic
response curve (AUC) elicited by a portion of food containing 50 g available
carbohydrate, and R is the mean AUC elicited by 50 g available carbohydrate
from the reference food or glucose tested 2–3 times by the same subject. The
GI is the mean of these values for a group of subjects (typically n¼ 10)
(Brouns et al., 2003; Wolever, 2006). In seven out of eight studies in which it
was examined, a high GI diet significantly increased risk for type 2 diabetes,
with the mean increase in risk being about 30%.
A marker of both carbohydrate quality and quantity is glycemic load,
defined as GI� g, where GI is the food glycemic index and g is the amount of
available carbohydrate in the food. Glycemic load is considered by many to be
Table 10.1 Relative risk of type 2 diabetes for 5th vs. 1st quintiles of intake of totalcarbohydrate, cereal fibre, glycemic index and glycemic load.
Totalcarbohydrate
Cerealfibre
Glycemicindex
Glycemicload
Salmeron et al.,1997a
ns 0.70 1.37 ns
Salmeron et al.,1997b
ns 0.72 1.37 1.47
Meyer et al., 2000 ns 0.64 ns nsSchulze et al.,
2004bns 0.63 1.59 ns
Hodge et al., 2004 ns ns 1.23 nsZhang et al., 2006 ns 0.77 1.30 1.61Villegas et al., 2007 1.28 – 1.21 1.34Krishnan et al.,
2007– 0.82 1.23 1.22
Values are relative risks which are significantly different from 1 for fully adjusted models; ns¼ non-
significant effect; ‘–’ is shown for studies where no result is given.
194 Resistant Starch

3GCH10 07/20/2013 8:53:9 Page 195
a more complete indicator of the physiological impact of dietary carbohy-
drate. However, it is not a consistent predictor of diabetes risk, with a
significant effect only found in four out of the eight studies in which it
was examined (Table 10.1).
10.4 PATHOGENESIS OF TYPE 2 DIABETES
Prospective studies suggest that the quality of dietary carbohydrate (high
cereal fibre intake and/or low GI) is more important than the quantity
consumed for preventing type 2 diabetes. To determine if there are plausible
mechanisms by which high cereal fibre and low GI diets reduce the risk for
type 2 diabetes, it is necessary to understand the pathophysiological mecha-
nisms which cause type 2 diabetes.
Many factors influence blood glucose, but perhaps the most important is
insulin. Insulin controls the blood glucose concentration via a classical
feedback loop. A rise in blood glucose stimulates the secretion of insulin
from the beta-cells of the pancreas, and the resulting rise in blood insulin
stimulates muscle and adipose tissue to increase glucose uptake, hence
causing blood glucose to fall. The ability of a rise in plasma glucose to
stimulate insulin secretion is termed insulin secretion, and the ability of
insulin to stimulate glucose uptake from the blood is termed insulin sensitivity.
In normal subjects, there is a hyperbolic relationship between insulin secretion
and insulin sensitivity (Clausen et al., 1996). Thus, people with high insulin
sensitivity (or low insulin resistance) have low insulin secretion in response to
a rise in blood glucose, while those with low insulin sensitivity (or high insulin
resistance) have high insulin secretion. The product of insulin secretion and
insulin sensitivity is termed disposition index, which is a marker of beta-cell
function.
It is commonly thought that diabetes develops because of decreased insulin
sensitivity (or increased insulin resistance). However, the relationship
between insulin secretion and insulin sensitivity suggests that blood glucose
remains normal as long as changes in insulin sensitivity are compensated for
by changes in insulin secretion. Type 2 diabetes develops when the disposition
index falls – that is, when insulin secretion cannot be increased enough to
compensate for reductions in insulin sensitivity (Kahn, 2003).
Most studies investigating the role of diet on the pathogenesis of type
2 diabetes only measure insulin sensitivity, but this does not necessarily
indicate risk for type 2 diabetes, because the deleterious effects of reduced
insulin sensitivity or the beneficial effects of increased insulin sensitivity on
Role of Carbohydrates in the Prevention of Type 2 Diabetes 195

3GCH10 07/20/2013 8:53:9 Page 196
blood glucose could be compensated for by compensatory changes in insulin
secretion. Indeed, studies in which both insulin secretion and insulin sensi-
tivity have been measured have shown that factors which increase insulin
sensitivity, such as exercise (Kahn et al., 1990), are accompanied, at least in
the short term, by a compensatory reduction in insulin secretion. Conversely,
factors which cause a reduction in insulin sensitivity, such as pregnancy
(Catalano et al., 1993) or use of nicotinic acid (Kahn et al., 1989) are
associated with compensatory increases in insulin secretion, such that the
disposition index did not change.
The dynamic relationship between insulin secretion and insulin sensitivity
makes teleological sense, because it allows blood glucose control to be
maintained despite changes in insulin sensitivity that normally occur through-
out life due to physiological factors such as puberty, pregnancy and changes in
body composition that occur during aging, and also by environmental factors
such as short-term changes in activity and the quality and quantity of food
intake.
The relative importance of changes in insulin secretion and changes in
insulin sensitivity in causing the development of type 2 diabetes has been
shown in two studies in which both were measured longitudinally in the same
subjects (Weyer et al., 1999; Festa et al., 2006). These studies show that a
reduction in insulin sensitivity is not necessarily associated with the develop-
ment of diabetes but, rather, it is the change in insulin secretion that
determines whether an individual develops diabetes or not. In the larger
study of this kind, oral glucose tolerance was assessed, and insulin secretion
and insulin sensitivity were measured, using a frequently sampled intravenous
glucose tolerance test in nearly 800 subjects at baseline and after a mean
period of 5.2 years follow-up.
Figures 10.1a and 10.1c show the results for subjects who had normal
glucose tolerance at baseline divided into three groups: those who remained
normal at follow-up; those who developed impaired glucose tolerance
(IGT); and those who developed diabetes. All three of these groups of
subjects gained weight and became more insulin resistant (less insulin
sensitive) with time. What distinguished them was that subjects who
remained normal increased insulin secretion more than those who developed
impaired glucose tolerance, while those who developed diabetes had no
change in insulin secretion. Similar results are seen for subjects who had
impaired glucose tolerance at baseline (Figures 10.1b and 10.1d). Remark-
ably, some subjects with impaired glucose tolerance at baseline became
normal at follow-up, despite the fact that they became more insulin resistant;
the increase in insulin secretion was large enough to overcome the reduction
in insulin sensitivity.
196 Resistant Starch

3GCH10 07/20/2013 8:53:9 Page 197
10.5 EFFECT OF ALTERING SOURCE OR AMOUNT OFDIETARY CARBOHYDRATE ON INSULINSENSITIVITY, INSULIN SECRETION ANDDISPOSITION INDEX
The glycemic impact of the diet can be reduced by changing the source of
dietary carbohydrate (i.e. using low-GI foods) or by reducing the amount of
(a)NormalIGTDiabetes
(c)
(b) (d)
Normal at baseline IGT at baseline
Change over 5.2 years Change over 5.2 years
Weight Si AIR Weight Si AIR
Insu
lin s
ecre
tion
3
2
1
0
–1
0 1 2 3 0 1 2 3
100
80
60
40
20
0
Insulin sensitivity Insulin sensitivity
IGT → N (n = 66)
IGT → IGT (n = 95)
IGT → D (n = 59)
N → N(n = 350)
N → IGT(n = 105)
N → D(n = 40)
N → NN → IGTN → D
IGT → NIGT → IGTIGT → D
Figure 10.1 Changes in body weight, insulin sensitivity (Si) and insulin secretion (AIR,area under the insulin response curve after intravenous glucose) over 5.2 years insubjects who had normal glucose tolerance at baseline (a and c) and subjects whohad impaired glucose tolerance at baseline (b and d), and whose glucose toleranceat follow-up had remained, or become, normal (N), impaired (IGT) or diabetic (D).Adapted from Festa et al. (2006).
Role of Carbohydrates in the Prevention of Type 2 Diabetes 197

3GCH10 07/20/2013 8:53:9 Page 198
carbohydrate consumed. Since epidemiological studies suggest that carbohy-
drate quality is associated with risk for diabetes, but that carbohydrate
quantity is not (Table 10.1), we hypothesized that, on weight-maintaining
diets, reducing diet GI without reducing carbohydrate intake would have a
more beneficial effect on insulin sensitivity and insulin secretion than a
moderate reduction in carbohydrate intake.
To test this, we studied 35 subjects with IGT, whom we identified by
screening subjects with risk factors for type 2 diabetes using a 75 g OGTT
(Wolever &Mehling, 2002). They were randomly assigned to consume one of
three diets for four months:
1. a diet containing 50–55% carbohydrate using at least one high-GI food at
every meal (high-GI, n¼ 11);
2. a diet containing 50–55% carbohydrate using at least one low-GI food at
every meal (low-GI, n¼ 13); or
3. a diet in which carbohydrate intake was reduced to 40–45% of energy and
monounsaturated fat (MUFA) intake was increased (MUFA, n¼ 11).
Subjects were provided with high- and low-GI foods and food sources of
MUFA to use in their diets. Insulin sensitivity, insulin secretion and disposi-
tion index (the product of insulin sensitivity and insulin secretion) were
measured using a frequently-sampled intravenous glucose tolerance test at
baseline and after four months on the diet. To compare the effects of the three
diets on blood glucose, insulin and free-fatty acid (FFA) responses while
consuming the study diets, subjects also underwent eight-hour metabolic
profiles. At baseline, all subjects consumed breakfast and lunch meals
reflecting the high-GI diet. At four months they consumed breakfast and
lunchmeals reflecting the diet towhich they had been randomized; subjects on
high-GI consumed the same meals as at baseline, subjects on low-GI
consumed meals of the same energy and carbohydrate content but using
low-GI instead of high GI foods, and subjects onMUFA consumedmeals with
the same energy as at baseline but containing less carbohydrate and more
MUFA.
The results showed that disposition index tended to deteriorate on the high-
GI and MUFA diets, but significantly increased on the low-GI diet. The
change in disposition index on the low-GI diet was significantly different from
the changes on both the high-GI and MUFA diets (Figure 10.2).
Similar effects have been seen in other studies. For example, 72 subjects
with metabolic syndrome were randomly assigned to receive a diet containing
oat, wheat and potatoes (foods shown to elicit high glucose and insulin
responses) or a diet containing whole rye breads and pasta (low glucose and
198 Resistant Starch

3GCH10 07/20/2013 8:53:9 Page 199
insulin responses). Beta-cell function improved significantly on the rye-pasta
diet compared to the oat-wheat-potato diet (Laaksonen et al., 2005). These
studies suggest that low-GI foods reduce diabetes risk by improving beta-cell
function.
10.6 MECHANISMS BY WHICH LOW-GI FOODSIMPROVE BETA-CELL FUNCTION
Mechanisms by which low-GI foods may increase beta-cell function include
reduced glucose toxicity, reduced serum FFA and increased levels of incretin
hormones such as GLP-1.
10.6.1 Glucose toxicity
Glucose toxicity refers to damaging effects of high blood glucose concentra-
tions per se on body tissues and regulatory processes due to a variety of
mechanisms, including increased flux through the polyol and glucosamine
pathways, increased non-enzymatic glycation products and glycosylation of
proteins, activation of diacylglycerol and protein kinase C and increased
oxidative and carbonyl stress (Brownlee, 2005).
Insulin sensitivity →
Insu
lin s
ecre
tion
→
p = 0.05
p = 0.05
Low GI
High GI
MUFA
Figure 10.2 Changes in insulin sensitivity and insulin secretion in subjects withimpaired glucose tolerance after four months on a high-carbohydrate, high-glycemicindex (GI) diet (high-GI), a high-carbohydrate, low-GI diet (low-GI) and a low-carbohydrate, high-monounsaturated fat (MUFA) diet. Adapted from Wolever andMehling (2002).
Role of Carbohydrates in the Prevention of Type 2 Diabetes 199

3GCH10 07/20/2013 8:53:9 Page 200
Glucose toxicity has been implicated in the pathogenesis of diabetes by
reducing beta-cell function (Leahy et al., 1988). However, if glucose toxicity
was the only mechanism involved, any method of reducing blood glucose
should have a beneficial effect on beta-cell function. That this is not the case is
shown by the fact that, in subjects with IGT, reducing diet GI improved beta-
cell function, but reducing the amount of carbohydrate consumed did not,
despite the fact that both diets reduced mean eight-hour blood glucose
compared to the high-GI diet and did so to an equivalent extent (Wolever
& Mehling, 2003). Similarly, in subjects with type 2 diabetes a low-GI diet
reduced serum c-reactive protein as a marker of chronic inflammation, while a
lower carbohydrate, high MUFA diet did not (Wolever et al., 2008). Both of
these studies suggest that reducing glucose toxicity is not the only mechanism
by which low-GI foods improve beta-cell function.
10.6.2 Reduced serum free fatty acids (FFA)
The beta-cell secretes insulin in response to a rise in blood glucose, and it has
been proposed that the intracellular mechanism by which glucose stimulates
insulin secretion is via intracellular long-chain fatty acids. A rise in blood
glucose increases glucose entry into the beta-cell and, hence, increases flux
through the glycolytic pathway and glucose oxidation. Increased glucose
oxidation within the beta-cell inhibits the oxidation of fatty acids, causing the
intracellular concentration of fatty acids to rise and this, in turn, stimulates
the release of insulin from the cell (Prentki & Corkey, 1996). This is confirmed
by studies showing that the addition of fatty acids to the medium of cultured
beta-cells increases insulin secretion (Zhou & Grill, 1994) and, in humans
in vivo, acutely increasing the serum FFA concentration increases insulin
secretion (Carpentier et al., 1999).
However, in the presence of chronically high concentrations of intra-
cellular fatty acids and glucose, the glucose signal is lost. Prolonged
exposure of cultured beta cells to high levels of FFA and glucose reduces
insulin secretion (Zhou & Grill, 1994). Similarly, in humans, a prolonged
increase in serum FFA inhibits insulin secretion (Carpentier et al., 1999).
For this reason, we measured serum FFA concentrations after four months
on the low-GI, high-GI and MUFA diets and showed that 0–8 hours mean
serum FFA concentration on the low-GI diet was significantly lower than on
high-GI, whereas the levels of FFA on the MUFA diet were similar to those
on high-GI (Wolever & Mehling, 2003). This suggests that it may be
necessary to reduce both glucose and FFA in order to improve beta-cell
function.
200 Resistant Starch

3GCH10 07/20/2013 8:53:9 Page 201
10.6.3 Increased GLP-1 secretion
GLP-1 (glucagon-like polypeptide-1) is an incretin, secreted from the L-cells
in the lower end of the small intestine, which has multiple biological
functions, including delaying gastric emptying, increasing satiety, increasing
insulin sensitivity and increasing insulin secretion (Drucker, 1998). The latter
is thought to be achieved not only by increasing the amount of insulin secreted
by each beta-cell, but also by increasing the number of beta-cells. GLP-1 is
secreted in response to the presence of nutrients in the distal small intestine
and also via a neuroendocrine loop, whereby GIP secretion in the upper
intestine sends a signal via the vagus nerve and the brain to the L-cells
(Brubaker & Anini, 2003).
Animal studies suggest that GLP-1 secretion is also stimulated by the
short-chain fatty acids (SCFA) produced upon fermentation of dietary fibre
and resistant starch in the large intestine (Reimer & McBurney, 1996).
This might be a mechanism by which diets high in cereal fibre reduce
diabetes risk. The cereal fibre most commonly consumed in North America
is wheat bran; however, studies lasting 4–12 weeks have shown no signifi-
cant effect of wheat fibre on blood glucose control or oral glucose tolerance
(Munoz et al., 1979; Kestin et al., 1990; Jenkins et al., 2002; Costabile
et al., 2008).
One way to reconcile discrepancy between epidemiological studies show-
ing that cereal fibre is a strong predictor of diabetes risk (Table 10.1), and
long-term clinical trials showing that wheat bran has little or no effect on
glucose metabolism, might be that the clinical trials lasted only four weeks to
three months, which is not long enough for the colonic bacteria to adapt to the
change in fibre intake. The colon is a very complex ecosystem, containing
hundreds of species of bacteria. If an input into an established ecosystem (here
the input is the amount of fibre entering the colon) is changed, the output of the
ecosystem becomes unstable and varies with time until a new equilibrium is
reached. The more complex the ecosystem, the longer it takes for a new
equilibrium to be established (Feng & Chai, 2008).
Studies in animals demonstrate this concept. For example, in a four-month
study, when the resistant starch content of the diet of rats was increased, the
SCFA content of the caecum began to increase and continued to increase over
entire period of the study (Le Blay et al., 1999). This suggests that it takes the
colonic ecosystem more than four months to equilibrate after a change in fibre
intake. Thus, we hypothesized that cereal fibre, in the form of All-Bran1
cereal would significantly increase serum SCFA concentrations and GLP-1
secretion, but that this would only be apparent after a long adaptation period of
at least six months.
Role of Carbohydrates in the Prevention of Type 2 Diabetes 201

3GCH10 07/20/2013 8:53:10 Page 202
To test this, we randomly assigned 28 hyperinsulinemic subjects to
consuming two cups of a low fibre cereal per day, or one cup of All-
Bran1 daily for one year (Freeland et al., 2010). At baseline, and after every
three months, subjects underwent a metabolic profile in which the glucose,
insulin, FFA, SCFA and GLP-1 responses elicited by the diet were measured.
The results showed significant time� treatment interactions for serum ace-
tate, butyrate and GLP-1; that is, the difference between the diets depended
upon the time. Acetate increased temporarily after nine months on the diet,
butyrate began to increase at three months and reached a plateau after nine
months, and GLP-1 tended to gradually increase with time to reach a level
significantly higher than the control after 12 months. These results supported
the hypothesis that cereal fibre may reduce diabetes risk by increasing GLP-1
secretion mediated by colonic short chain fatty acids, but that these events take
many months to occur, as the colonic ecosystem gradually adapts to the
increased fibre intake.
10.7 CONCLUSIONS
The quality of dietary carbohydrates may be an important factor in determin-
ing risk for type 2 diabetes. The results of recent studies suggest that a
deterioration in beta-cell function may be the critical pathophysiological event
which causes type 2 diabetes. Evidence has been presented here that low-GI
carbohydrates may preserve beta-cell function by reducing postprandial
glucose fluctuations and, hence, glucose toxicity, while at the same time
reducing serum FFA concentrations. Cereal fibre may preserve beta-cell
function, at least in part, by increasing colonic fermentation and up-regulating
GLP-1 secretion, but these effects may not occur until the colon has adapted to
the increased intake of fibre – a process which may take 6–9 months or more.
However, whether low-GI foods or cereal fibre will actually prevent type 2
diabetes in humans remains to be shown.
REFERENCES
Brouns, F., Bjorck, I., Frayn, K.N., Gibbs, A.L., Lang, V., Slama, G., Wolever, T.M.S.(2003). Glycaemic index methodology. Nutrition Research Reviews 18, 145–171.
Brownlee, M. (2005). The pathobiology of diabetes complications: a unifying mechanism.Diabetes 54, 1615–1625.
Brubaker, P.L., Anini, Y. (2003). Direct and indirect mechanisms regulating secretion ofglucagon-like peptide-1 and glucagon-like peptide-2. Canadian Journal of Physiologyand Pharmacology 81, 1005–1012.
202 Resistant Starch

3GCH10 07/20/2013 8:53:10 Page 203
Canadian Diabetes Association . (2010). The prevalence and costs of diabetes. URL: http://www.diabetes.ca/about-diabetes/what/prevalence/ [accessed 14 January, 2010].
Canadian Diabetes Association Clinical Practice Guidelines Expert Committee. (2008).Canadian Diabetes Association 2008 clinical practice guidelines for the prevention andmanagement of diabetes in Canada. Canadian Journal of Diabetes 32 (suppl 1),S10–S16.
Carpentier, A., Mittelman, S.D., Lamarche, B., Bergman, R.N., Giacca, A., Lewis, G.F.(1999). Acute enhancement of insulin secretion by FFA in humans is lost with prolongedFFA elevation. American Journal of Physiology 276, E1055–E1066.
Catalano, P.M., Tyzbir, E.D.,Wolfe, R.R., Calles, J., Roman, N.M., Amini, S.B., Sims, E.A.(1993). Carbohydrate metabolism during pregnancy in control subjects and women withgestational diabetes. American Journal of Physiology 264, E60–E67.
Clausen, J.O., Borch-Johnsen, K., Ibsen, H., Bergman, R.N., Hougaard, P., Winther, K.,Pedersen, O. (1996). Insulin sensitivity index, acute insulin response, and glucoseeffectiveness in a population-based sample of 380 young healthy Caucasians: analysis ofthe impact of gender, body fat, physical fitness and life-style factors. Journal of ClinicalInvestigation 98, 1195–1209.
Costabile, A., Klinder, A., Fava, F., Napolitano, A., Fogliano, V., Leonard, C., Gibson,G.R., Tuohy, K.M. (2008). Whole-grain wheat breakfast cereal has a prebiotic effect onthe human gut microbiota: a double-blind, placebo-controlled, crossover study. BritishJournal of Nutrition 99, 110–120.
Drucker, D.J. (1998). Glucagon-like peptides. Diabetes 47, 159–169.Feng, Q.Y., Chai, L.H. (2008). A new statistical dynamic analysis on vegetation patterns inland ecosystems. Physica A 387, 3583–3593.
Festa, A., Williams, K., D’Agostino, R., Jr., Wagenknecht, L.E., Haffner, S.M. (2006). Thenatural course of beta-cell function in nondiabetic and diabetic individuals: the InsulinResistance Atherosclerosis Study. Diabetes 55, 1114–1120.
Freeland, K.R., Wilson, C., Wolever, T.M.S. (2010). Adaptation of colonic fermentationand glucagon-like peptide-1 secretion with increased wheat fibre intake for 1 year inhyperinsulinaemic human subjects. British Journal of Nutrition 103, 82–90.
Hodge, A.M., English, D.R., O’Dea, K., Giles, G.G. (2004). Glycemic index and dietaryfiber and the risk of type 2 diabetes. Diabetes Care 27, 2701–2706.
Janket, S.J., Manson, J.E., Sesso, H., Buring, J.E., Liu, S. (2003). A prospective study ofsugar intake and risk of type 2 diabetes in women. Diabetes Care 26, 1008–1015.
Jenkins, D.J.A., Kendall, C.W.C., Augustin, L.S.A., Martini, M.C., Axelsen, M., Faulkner,D., Vidgen, E., Parker, T., Lau, H., Connelly, P.W., Teitel, J., Singer, W., Vandenbroucke,A.C., Leiter, L.A., Josse, R.G. (2002). Effect of wheat bran on glycaemic control and riskfactors for cardiovascular disease in type 2 diabetes. Diabetes Care 25, 1522–1528.
Kahn, S.E. (2003). The relative contributions of insulin resistance and beta-cell dysfunctionto the pathophysiology of type 2 diabetes. Diabetologia 46, 3–19.
Kahn, S.E., Beard, J.C., Schwartz, M.W., Ward, W.K., Ding, H.L., Bergman, R.N.,Taborsky, G.J.Jr., Porte, D.Jr., (1989). Increased beta-cell secretory capacity asmechanism for islet adaptation to nicotinic acid-induced insulin resistance. Diabetes38, 562–568.
Kahn, S.E., Larson, V.G., Beard, J.C., Cain, K.C., Fellingham, G.W., Schwartz, R.S., Veith,R.C., Stratton, J.R., Cerqueira, M.D., Abrass, I.B. (1990). Effect of exercise on insulinaction, glucose tolerance, and insulin secretion in aging. American Journal of Physiol-ogy 258, E937–E943.
Role of Carbohydrates in the Prevention of Type 2 Diabetes 203

3GCH10 07/20/2013 8:53:10 Page 204
Kestin, M., Moss, R., Clifton, P.M., Nestel, P.J. (1990). Comparative effects ofthree cereal brans on plasma lipids, blood pressure, and glucose metabolism inmildly hypercholesterolemic men. American Journal of Clinical Nutrition 52,661–666.
King, H., Aubert, R.E., Herman, W.H. (1998). Global burden of diabetes, 1995–2025.Prevalence, numerical estimates, and projections. Diabetes Care 21, 1414–1431.
Krishnan, S., Rosenberg, L., Singer, M., Hu, F.B., Djousse, L., Cupples, A., Palmer JR.,(2007). Glycemic index, glycemic load, and cereal fiber intake and risk of type 2 diabetesin US black women. Archives of Internal Medicine 167, 2304–2309.
Laaksonen, D.E., Toppinen, L.K., Juntunen, K.S., Autio, K., Liukkonen, K.H., Poutanen,K.S., Niskanen, L., Mykk€anen, H.M. (2005). Dietary carbohydrate modification enhan-ces insulin secretion in persons with the metabolic syndrome. American Journal ofClinical Nutrition 82, 1218–1227.
Leahy, J.L., Bonner-Weir, S., Weir, G.C. (1988). Minimal chronic hyperglycemia is acritical determinant of impaired insulin secretion after incomplete pancreatectomy.Journal of Clinical Investigation 81, 1407–1414.
Le Blay, G., Michel, C., Blotti�ere, H.M., Cherbut, C. (1999). Enhancement of butyrateproduction in the rat caecocolonic tract by long-term ingestion of resistant potato starch.British Journal of Nutrition 82, 419–426.
Meyer, K.A., Kushi, L.H., Jacobs, D.R.Jr., Slavin, J., Sellers, T.A., Folsom, A.R. (2000).Carbohydrates, dietary fiber, and incident type 2 diabetes in older women. AmericanJournal of Clinical Nutrition 71, 921–930.
Munoz, J.M., Sandstead, H.H., Jacob, R.A. (1979). Effects of dietary fiber on glucosetolerance of normal men. Diabetes 28, 496–502.
Prentki, M., Corkey, B.E. (1996). Are the b-cell signaling molecules malonyl-CoA andcytosolic long-chain acyl-CoA implicated in multiple tissue defects of obesity andNIDDM? Diabetes 45, 273–283.
Reimer, R.A., McBurney, M.I. (1996). Dietary fiber modulates intestinal proglucagonmessenger ribonucleic acid and postprandial secretion of glucagon-like peptide-1 andinsulin in rats. Endocrinology 137, 3948–3956.
Salmeron, J., Ascherio, A., Rimm, E.B., Colditz, G.A., Spiegelman, D., Jenkins, D.J.,Stampfer, M.J., Wing, A.L., Willett, W.C. (1997a). Dietary fiber, glycemic load, and riskof NIDDM in men. Diabetes Care 20, 545–550.
Salmeron, J., Manson, J.E., Stampfer, M.J., Colditz, G.A., Wing, A.L., Willett, W.C.(1997b). Dietary fiber, glycemic load, and risk of non-insulin-dependent diabetesmellitus in women. Journal of the American Medical Association 277, 472–477.
Schulze, M.B., Manson, J.E., Ludwig, D.S., Colditz, G.A., Stampfer, M.J., Willett, W.C.,Hu, F.B. (2004a). Sugar-Sweetened Beverages, Weight Gain, and Incidence of Type 2Diabetes in Young and Middle-Aged Women. Journal of the American MedicalAssociation 292, 927–934.
Schulze, M.B., Liu, S., Rimm, E.B., Manson, J.E., Willett, W.C., Hu, F.B. (2004b).Glycemic index, glycemic load, and dietary fiber intake and incidence of type 2 diabetesin younger and middle-aged women. American Journal of Clinical Nutrition 80,348–356.
Villegas, R., Liu, S., Gao, Y.-T., Yang, G., Li, H., Zheng, W., and Shu, X.O. (2007).Prospective study of dietary carbohydrates, glycemic index, glycemic load, and inci-dence of type 2 diabetes mellitus in middle-aged Chinese women. Archives of InternalMedicine 167, 2310–2316.
204 Resistant Starch

3GCH10 07/20/2013 8:53:10 Page 205
Weyer, C., Bogardus, C., Mott, D.M., Pratley, R.E. (1999). The natural history of insulinsecretory dysfunction and insulin resistance in the pathogenesis of type 2 diabetesmellitus. Journal of Clinical Investigation 104, 787–794.
Wolever, T.M.S. (2006). The Glycaemic Index: A Physiological Classification of DietaryCarbohydrate. CABI Publishing, Wallingford, UK.
Wolever, T.M.S., Mehling, C. (2002). High-carbohydrate/low-glycaemic index dietaryadvice improves glucose disposition index in subjects with impaired glucose tolerance.British Journal of Nutrition 87, 477–487.
Wolever, T.M.S., Mehling, C. (2003). Long-term effect of varying source or amount ofdietary carbohydrate on postprandial plasma glucose, insulin, triacylglycerol and freefatty acid concentrations in subjects with impaired glucose tolerance. American Journalof Clinical Nutrition 77, 612–621.
Wolever, T.M.S., Gibbs, A.L., Mehling, C., Chiasson, J.-L., Connelly, P.W., Josse, R.G.,Leiter, L.A., Maheux, P., Rabasa-Lhoret, R., Rodger, N.W., Ryan, E.A. (2008). TheCanadian trial of Carbohydrates in Diabetes (CCD), a 1-y controlled trial of low-glycemic-index dietary carbohydrate in type 2 diabetes: no effect on glycated hemo-globin but reduction in C-reactive protein. American Journal of Clinical Nutrition 87,114–125.
Zhang, C., Liu, S., Solomon, C.G., Hu, F.B. (2006). Dietary fiber intake, dietary glycemicload, and the risk for gestational diabetes mellitus. Diabetes Care 29, 2223–2230.
Zhou, Y.-P., Grill, V.E. (1994). Long-term exposure of rat pancreatic islets to fatty acidsinhibits glucose-induced insulin secretion and biosynthesis through a glucose fatty acidcycle. Journal of Clinical Investigation 93, 870–876.
Role of Carbohydrates in the Prevention of Type 2 Diabetes 205

3GCH10 07/20/2013 8:53:10 Page 206

CH11GR 07/29/2013 16:40:56 Page 207
11 Resistant Starch on Glycemiaand Satiety in Humans
Mark D. HaubDepartment of Human Nutrition, Kansas State University, USA
11.1 INTRODUCTION
There is a major public health effort to decrease the prevalence of obesity-
related health outcomes. Given the role of diet and nutrition on the aetiology
of obesity, it is imperative that dietary options should be targeted to assist with
preventing these comorbid conditions such as type 2 diabetes, dyslipidaemia
and other cardiometabolic complications.While resistant starch (RS) has been
studied by cereal chemists and food companies for several years, studies
conducted by clinical scientists have been limited, but these are increasing and
providing interesting results indicating a strong evidence that RS may provide
significant metabolic health benefits. The majority of studies indicates a
capacity to lower glycemia and improve insulin sensitivity. Other data, while
less conclusive, indicate the potential to attenuate satiety and decrease
subsequent food intake. Thus, there seems to be an inverse relationship
between glycemic response and food intake following the consumption of
RS or foods containing adequate amounts of RS. Moreover, while the data are
limited, evidence seems to indicate that different types of RS can elicit
different metabolic responses.
This review will focus on the glucoregulatory aspects of RS, with discus-
sions on outcomes from primarily human clinical trials relating to satiety, food
intake and potential to address the obesity issue present in most industrialized
societies.
207
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

CH11GR 07/29/2013 16:40:56 Page 208
11.2 DIET AND RESISTANT STARCH
Diabetes, obesity and cardiovascular disease are interrelated. Diet is one
factor that links them all together. Specifically, the current obesity epidemic
has been suggested to be a result of increased carbohydrate intake (Gaesser,
2007), even though dietary fat has frequently been touted as the primary
dietary culprit of many deleterious metabolic conditions. The fact is that we,
as a nation, actually increased carbohydrate consumption – perhaps to
address the public health message to avoid dietary fat – during the latter
years of the past century, just as the obesity epidemic began to surge
(Gaesser, 2007). Thus, the national response to decrease fat may have
inadvertently led to overconsumption of carbohydrates. From one perspec-
tive, the public health effort to decrease the percent of energy derived from
fat in our diets was moderately successful; however, total energy intake and
attaining a weight-maintaining energy balance was not affected by those
behaviours.
So, while it is laudable to try to change the behaviour of individuals (to
choose other foods), it might be more effective to address this nutritional issue
by providing bioactive compounds that ‘behave’ like traditional starch, yet
elicit a more favourable metabolic profile. In other words, let people continue
to choose the foods they prefer, but simply make those foods healthier by
incorporating bioactive ingredients and/or ingredients that affect energy
metabolism. This might seem to be an achievable goal, but some past efforts
have not proven to be acceptable to consumers.
To address this issue, RS shows promise at successfully improving
glucoregulation in a consumer-acceptable fashion, while eliciting side effects
that are similar to typical dietary fibres (e.g. bran) that have had difficulty
being accepted by a most consumers. Compared with typical monosacchar-
ides and disaccharides, RS elicits minimal glucose and insulin excursions and
can be incorporated easily into regular food items with general acceptance by
consumers, which is a barrier to many dietary fibres. That said, there is a
paucity of evidence, especially from humans, to illustrate the glucose-low-
ering mechanisms. The common theory is based on the glucose-fatty acid
cycle, whereby increasing fat oxidation allows for increased insulin sensitivity
and glucose disposal (Randle et al., 1963).
Testing this theory in human clinical trials will increase our understanding
of the mechanisms through which RS, and other modified starches with
similar properties, elicit their benefits. This will help food and pharmaceutical
scientists to develop and/or enhance products to better prevent, manage and
treat metabolic conditions. Currently, evidence indicates a glucoregulatory
benefit, but how this occurs is not well established.
208 Resistant Starch

CH11GR 07/29/2013 16:40:56 Page 209
11.3 RESISTANT STARCH AND INSULIN SENSITIVITY
RS is a dietary carbohydrate that has been frequently studied. Most RS
research, however, has focused on fermentation characteristics, while few
studies have been designed to investigate mechanisms involved with increas-
ing insulin sensitivity. Most of the mechanistic studies have used rodents and
other small animals rather than using a human clinical trial.
Basing RSmechanisms on animal study outcomes is problematic, given the
coprophagic tendency of some species, whichmakes it difficult to interpret the
results and apply those potentially confounded results to humans. Moreover,
humans prefer eating a varied diet throughout the day and over time, while
animal studies test ingredients with the animals eating the same food at every
meal, every day. An additional research design and translational science issue
is that we do not fully understand how to relate animal outcomes to humans,
given that the dose and duration of treatment given to animals may not apply
to typical human scenarios.
There has been one human clinical trial (Robertson et al., 2005) that
specifically investigated the effects of resistant starch on insulin sensitivity
using the standard euglycaemic-hyperglycaemic clamp. They observed that
individuals consuming 30 g/day of RS over four weeks elicited significant
increase in glucose disposal and decreased insulin area under the curve during
a meal tolerance test. However, while they observed increased insulin
sensitivity, they were unable to discern clearly how RS elicited this response.
Results from other human clinical trials (Yamada et al., 2005; Robertson
et al., 2003; Haub et al., 2010; Behall et al., 2006; Al-Tamimi et al., 2010),
support this notion that RS decreases postprandial glucose and insulin
responses. While that evidence is fairly strong and consistent, the long-
term benefits of RS are still not well established. What is less clear is the
mechanism through which long-term RS ingestion exerts its benefits, and
whether RS consistently affects health and lifestyle outcomes (e.g. obesity,
satiety, and food intake) other than glucose metabolism in humans.
11.4 CURRENT THEORETICAL MECHANISM
A testable theory to explain the metabolic regulation by RS pertains to the
formation of short chain fatty acids via fermentation, which subsequently
increases fat oxidation (centrally and peripherally). While this theory has not
formally been tested relative to RS, it is supported by the observation that
increasing fat oxidation is a key factor to increase insulin sensitivity in those
with insulin resistance (Figure 11.1).
Resistant Starch on Glycemia and Satiety in Humans 209

CH11GR 07/29/2013 16:40:56 Page 210
Moreover, it was reported by Higgins et al. (2004) that the ingestion of RS
leads to acute increases in lipid oxidation, which may lead to decreased fat
accumulation and/or accretion over time. While the role of short chain fatty
acids in this process is plausible, inducing an energy deficit also increases fat
oxidation and insulin sensitivity by decreasing the inhibition on fat oxidation
elicited by insulin and other glucoregulatory hormones. Thus, since RS only
yields half the energy of regular starch (�2 kcal/g), it is plausible that a lower
energy state of the cell or tissue may increase fat oxidation.
Specifically, based on the outcomes of Robertson et al. (2003), it was
suggested that the increased insulin sensitivity arises from alterations in
substrate oxidation due to the increased availability of short chain fatty acids
derived from fermentation of RS (Figure 11.1). However, these researchers
failed to include a treatment that only increased short chain fatty acids,
thereby limiting their capacity to better understand how RS elicits the ‘second
meal effect’ or whether other factors, such as an energy deficit, were
contributing factors.
However, in a study by Johnston et al. (2010), the RS treatment (40 g/d)
over 12 weeks elicited increased insulin sensitivity without a concomitant
change in body weight or intracellular lipid content. These data indicate that
Resistant starch
Fermentation
Improved metabolic health
Energy deficit‘Weight loss’
Short chain fatty acids
blood glucoseand insulin
?
?
Figure 11.1 This figure depicts means through which RS is suggested to elicitmetabolic benefits. It has been established that RS decreases blood glucose andinsulin, which decreases risk for type 2 diabetes over time. It has, also, been shown tobe fermented by gut microbiota, thereby changing the gut ecology and producingproducts (short chain fatty acids) that have been suggested to improve metabolichealth. To date, however, it is not knownwhether some of the changes attributed to RSare due to changes in energy balance as RS may decrease energy intake acutely and/or chronically. Thus, some benefits attributed to RS may be indirect due to decreasedenergy being absorbed, and/or different ratios of macronutrients due to conversionof some RS into fatty acids by the gut microbiota.
210 Resistant Starch

CH11GR 07/29/2013 16:40:56 Page 211
RS may even improve glycemia and insulinaemia, regardless of energy intake
or lipid metabolism.
11.5 SATIETY
OneuniqueeffectofRSthatseemstobedevelopingistheimpactthatthesestarches
seemtohaveonsatiety.With lifestylemodification–specifically reducingenergy
intake – being a regular recommendation to prevent and treat various metabolic
diseases and conditions (Poirier et al., 2006; Jakicic et al., 2001), this bioactive
effect could be extremely beneficial from a public health perspective.
The means through which RS has been shown to induce satiety and lead to
decrease energy intake (may not decrease volume or weight of food con-
sumed) seems likely to involve alterations in PYY and other neuroendocrine
components (Zhou et al., 2008). Zhou et al. (2008) administered RS to mice
over 32 days and observed that active forms of PYYand GLP-1 increased. The
mice eating RS also gained less fat mass and had a lower ratio of body fat to
body weight, while eating the same weight of food.
Similarly, So et al. (2007) fed mice high and low RS diets and measured
numerous body composition and metabolic parameters after the eight-week
intervention. They observed several favourable outcomes following the high
RS treatment, compared with the low RS:
1. decreased intracellular fat deposition;
2. decreased body weight and adiposity;
3. smaller adipocytes;
4. their brain imaging results indicated greater satiety despite decreased
energy intake.
While those studies were completed in mice, the satiating effect of RS has
been reported in humans. Although hormone levels were not reported, Willis
et al. (2009) observed satiety and feelings of fullness were evident longer
following trials with RS and corn bran, compared with lower fibre bleached
oat bran, b-glucan, and polydextrose. In support with those satiety results,
outcomes from Anderson et al. (2010) indicate that foods containing 40–70%
RS seem to reduce food intake during meals later in the day. In a study out of
our lab (Haub et al., 2012), RS4 from potato starches elicited similar ratings of
satiety when ingested with an energy dense beverage (dextrose solution) or
ingested with water. Taken together, the mechanisms for the reduced intake
and ratings of satiety are not well understood, as gut hormone responses have
either not been studied or did not elicit consistent results.
Resistant Starch on Glycemia and Satiety in Humans 211

CH11GR 07/29/2013 16:40:56 Page 212
11.6 FERMENTATION ANDGUT MICROBIOTA
A new area of research has been better in understanding how gut microbiota
(especially lactobacilli and bifidobacteria), fermentation, and short chain
fatty acids impact human health (Roberfroid et al., 2010), with a few of these
aspects depicted in Figure 11.1. The food industry has been actively placing
prebiotic and probiotic foods on the market to provide consumers with foods
to try to meet this health outcome. Specific to RS, Martinez et al. (2010)
reported changes to gut microbiota following the ingestions of RS2 and RS4
by human volunteers. They observed that RS intake significantly altered gut
microbiota in as little as one to three weeks, and that RS2 and RS4 elicited
different changes in microbiota.
Moreover, short chain fatty acids (SCFA) formed from the fermentation of
dietary fibre and RS sources have been selected as potential factors for
increased satiety. However, a study using oligosaccharides (Hess et al.,
2010), reported that increased fermentation does not necessarily alter satiety
in men or women. Collectively, RS and the by-products of fermentation may
interact synergistically to elicit feelings of fullness. This is an area requiring
more research to understand better how the effects of gut changes in bacteria
and alterations in the fermentation process affect human health acutely and
over time.
11.7 EFFECT OF RS TYPE
An area of RS research that needs more investigation is comparing the forms
and types of RS currently available. Specifically, there is a paucity of data
comparing RS compounds of different types (e.g. Type 2 vs. Type 3) within
the same trial to determine similarities and differences on in vivo outcomes.
This information is imperative for consumers and others interested in select-
ing RS to incorporate into foods and diets.
To our knowledge, there is only one peer-reviewed paper to have compared
commercially available RS types from different companies on glycemia.
Haub et al. (2010) compared the glucose lowering effects of a commonly
tested type of RS (RS2) with a less studied cross-linked type (RS4). It was
observed that the RS4 elicited a greater reduction in capillary blood glucose
than RS2. This study clearly indicated that the type of RS used in clinical
studies needs to be accounted for, since different types can elicit significantly
different responses.
Together with the results fromMartinez et al. (2010), it can be erroneous to
conclude that all RS types will elicit the same physiological or psychological
212 Resistant Starch

CH11GR 07/29/2013 16:40:56 Page 213
responses. Further research investigating the differences in RS types will help
illustrate how much of each RS is necessary to elicit a desired response. This
outcome is critical when developing recipes and/or budgets for individual
consumers or companies.
11.8 SUMMARY
Over the past two decades, data from animal and human intervention studies
demonstrate that RS seems to elicit favourable changes in glucoregulatory
outcomes and factors relating to gut health. The results from animal studies
indicate a clearer effect on body composition and satiety, while the human
data are not as conclusive in that regard. More studies designed to test body
composition effects are needed to enhance our understanding of RS as a
potential ingredient to significantly affect obesity-related outcomes. Likewise,
while it is apparent that RS, as with other dietary fibres, does elicit changes in
metabolic and gut health outcomes, the appropriate dose and/or RS-contain-
ing foods that elicit desired outcomes and significant health effects are still not
well known.
REFERENCES
Al-Tamimi, E.K., Seib, P.A., Snyder, B.S., Haub, M.D. (2010). Consumption of Cross-Linked Resistant Starch (RS4(XL)) on Glucose and Insulin Responses in Humans.Journal of Nutrition and Metabolism Article ID 651063, 6p.
Anderson, G.H., Cho, C.E., Akhavan, T., Mollard, R.C., Luhovyy, B.L., Finocchiaro, E.T.(2010). Relation between estimates of cornstarch digestibility by the Englyst in vitromethod and glycemic response, subjective appetite, and short-term food intake in youngmen. American Journal of Clinical Nutrition 91, 932–939.
Behall, K.M., Scholfield, D.J., Hallfrisch, J.G., Liljeberg-Elmstahl, H.G. (2006). Con-sumption of both resistant starch and beta-glucan improves postprandial plasma glucoseand insulin in women. Diabetes Care 29, 976–981.
Gaesser, G.A. (2007). Carbohydrate quantity and quality in relation to body mass index.Journal of the American Dietetic Association 107, 1768–1780.
Haub,M.D., Hubach, K.L., Al-Tamimi, E.K., Ornelas, S., Seib, P.A. (2010). Different typesof resistant starch elicit different glucose responses in humans. Journal of Nutrition andMetabolism Article ID 230501.
Haub, M.D., Louk, J.A., Lopez, T.C. (2012). Novel resistant potato starches on glycemiaand satiety in humans. Journal of Nutrition and Metabolism Article ID 478043, 4p.
Hess, J.R., Birkett, A.M., Thomas, W., Slavin, J.L. (2010). Effects of short-chainfructooligosaccharides on satiety responses in healthy men and women. Appetite56(1), 128–34.
Resistant Starch on Glycemia and Satiety in Humans 213

CH11GR 07/29/2013 16:40:57 Page 214
Higgins, J.A., Higbee, D.R., Donahoo W.T., Brown, I.L., Bell, M.L., Bessesen, D.H.(2004). Resistant starch consumption promotes lipid oxidation. Nutrition &Metabolism1, 8.
Jakicic, J.M., Clark, K., Coleman, E., Donnelly, J.E., Foreyt, J., Melanson, E., Volek, J.,Volpe, S.L. (2001). American College of Sports Medicine position stand. Appropriateintervention strategies for weight loss and prevention of weight regain for adults.Medicine and Science in Sports and Exercise 33, 2145–2156.
Johnston, K.L., Thomas, E.L., Bell, J.D., Frost, G.S., Robertson, M.D. (2010). Resistantstarch improves insulin sensitivity in metabolic syndrome. Diabetic Medicine 27,391–397.
Martinez, I., Kim, J., Duffy, P.R., Schlegel, V.L., Walter, J. (2010). Resistant starches types2 and 4 have differential effects on the composition of the fecal microbiota in humansubjects. PLoS ONE 5(11): e15046.
Poirier, P., Giles, T.D., Bray, G.A., Hong, Y., Stern, J.S., Pi-Sunyer, F.X., Eckel, R.H.(2006). Obesity and cardiovascular disease: pathophysiology, evaluation, and effect ofweight loss: an update of the 1997 American Heart Association Scientific Statement onObesity and Heart Disease from the Obesity Committee of the Council on Nutrition,Physical Activity, and Metabolism. Circulation 113, 898–918.
Randle, P.J., Garland, P.B., Hales, C.N., Newsholme, E.A. (1963). The glucose fatty-acidcycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus.Lancet 1, 785–789.
Roberfroid, M., Gibson, G.R., Hoyles, L., McCartney, A.L., Rastall, R., Rowland, I.,Wolvers, D., Watzl, B., Szajewska, H., Stahl, B., Guarner, F., Respondek, F., Whelan, K.,Coxam, V., Davicco, M.J., L�eotoing, L., Wittrant, Y., Delzenne, N.M., Cani, P.D.,Neyrinck, A.M., Meheust, A. (2010). Prebiotic effects: metabolic and health benefits.British Journal of Nutrition 104(Suppl 2), S1–63.
Robertson, M.D., Currie, J.M., Morgan, L.M., Jewell, D.P., Frayn, K.N. (2003). Prior short-term consumption of resistant starch enhances postprandial insulin sensitivity in healthysubjects. Diabetologia 46, 659–665.
Robertson, M.D., Bickerton, A.S., Dennis, A.L., Vidal, H., Frayn, K.N. (2005). Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adiposetissue metabolism. American Journal of Clinical Nutrition 82, 559–567.
So, P.W., Yu, W.S., Kuo, Y.T., Wasserfall, C., Goldstone, A.P., Bell, J.D., Frost, G. (2007).Impact of resistant starch on body fat patterning and central appetite regulation. PLoSONE 2, e1309.
Willis, H.J., Eldridge, A.L., Beiseigel, J., Thomas, W., Slavin, J.L. (2009). Greater satietyresponse with resistant starch and corn bran in human subjects. Nutrition Research 29,100–105.
Yamada, Y., Hosoya, S., Nishimura, S., Tanaka, T., Kajimoto, Y., Nishimura, A., Kajimoto,O. (2005). Effect of bread containing resistant starch on postprandial blood glucoselevels in humans. Bioscience, Biotechnology and Biochemistry 69, 559–566.
Zhou, J., Martin, R.J., Tulley, R.T., Raggio, A.M., McCutcheon, K.L., Shen, L., Danna, S.C., Tripathy, S., Hegsted, M., Keenan, M.J. (2008). Dietary resistant starch upregulatestotal GLP-1 and PYY in a sustained day-long manner through fermentation in rodents.American Journal of Physiology – Endocrinology and Metabolism 295, E1160–1166.
214 Resistant Starch

3GCH12 07/20/2013 9:50:29 Page 215
12 The Acute Effects of Resistant Starchon Appetite and Satiety
Caroline L. Bodinham and M. Denise RobertsonDepartment of Nutritional Sciences, Faculty of Health andMedical Sciences, University of Surrey, UK
12.1 APPETITE REGULATION
Appetite is defined as the desire or physical craving to eat and is typically
separated into hunger, satiation and satiety. In a review byMattes et al. (2005),
hunger is defined as ‘sensations that promote food consumption’, satiation as
‘sensations that govern meal size and duration’ and satiety as ‘sensations
that determine the inter-meal period of fasting’. Therefore, all are important
for determining energy intake. Appetite is difficult to quantify, as it varies
greatly between individuals. It is closely regulated by physiological processes,
but it can also be influenced by many other aspects, including psychological
factors (e.g. learned habits) and emotional factors (e.g. as a response to stress),
as well as by external cues (e.g. accessibility of food, the presence or absence
of other individuals or the hedonic properties of the food itself).
The hypothalamus plays a vital role in appetite regulation and the subsequent
control of food intake; it receives peripheral signals from the digestive tract
(gut peptides) and adipose tissue (through signalling molecules, including
leptin and insulin) (Murphy & Bloom, 2004; Wynne et al., 2005). While the
hypothalamus is composed of many areas (nuclei), the arcuate nucleus (ARC)
is thought to be the most important in appetite regulation and is essential for
interpreting these peripheral signals (Wynne et al., 2005; Dhillo, 2007). The
ARC has an incomplete blood-brain barrier, so peripheral signals are able to
cross the barrier, interact with receptors and consequently cause the release of
neuropeptides that subsequently regulate ingestion (Neary et al., 2004).
There are two classes of neurons in the ARC: those that stimulate
and those that inhibit food intake. The neurons that stimulate appetite
(and, therefore, promote food intake) are neuropeptide Y (NPY) and
215
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH12 07/20/2013 9:50:29 Page 216
agouti-related peptide (AgRP) (Wynne et al., 2005). NPY causes an increase
in food intake through activation of the G-protein-coupled receptors Y1 and
Y5, while AgRP causes an increase in food intake through antagonism of
the melanocortin-4 (MC4) receptor and, therefore, the inhibition of
a-melanocyte-stimulating hormone (a-MSH) (Neary et al., 2004; Wynne
et al., 2005). The neurons that inhibit food intake are pro-opiomelanocortin
(POMC) and cocaine- and amphetamine-regulated transcript (CART). These
neurons stimulatea-MSH,which interacts with theMC4 receptor (Murphy&
Bloom, 2004; Neary et al., 2004; Wynne et al., 2005) that promotes sub-
sequent anorexigenic activity. Once the peripheral signals are interpreted in
the ARC, this can both inhibit and stimulate the different classes of neurons
whether to promote food intake or to prevent further intake.
The peripheral signals from the digestive tract are hormones that are both
anorexigenic (increase satiety) and orexigenic (promote hunger). The hor-
mone ghrelin is unique in being orexigenic, and it is often termed the
‘hormone of hunger’. Anorexigenic hormones are released postprandially
in response to ingested nutrients; they act by both direct and indirect (via the
vagus) mechanisms and promote feelings of satiety. These hormones include
glucagon-like peptide-1 (GLP-1) and peptide YY (PYY).
12.2 MEASUREMENT OF APPETITE IN HUMANS
Appetite is particularly difficult to directly measure in humans, as it is a
subjective sensation that can be influenced by many factors. Human studies,
therefore, have used different indirect measures in order to assess appetite
sensations (Mattes et al., 2005). The most frequently used indirect methods
are recording food intake, using questionnaires regarding appetite feelings and
measuring biomarkers (Mattes et al., 2005).
Records of food intake rely on participants accurately recording all they
have to eat and drink over a certain time period, as well as on accurate
interpretation by the investigators. As such, food records may not always
provide precise results. Food intake can also be assessed at weighed and
controlled ad libitum meals. While these do provide accurate measures of
food intake, they might not show habitual intakes, as they are often completed
in artificial environments.
Questionnaires relating to subjective appetite feelings are often used in
human studies. These questionnaires rely on those filling out the questions
reporting on their thoughts and feelings relating to hunger, satiation and
satiety. Typically, there are two main types of questionnaire that are used –
category scales and visual analogue scales (VAS). These are quick and easy to
216 Resistant Starch

3GCH12 07/20/2013 9:50:29 Page 217
complete and have been proven to be reproducible and valid in appetite
research.
Biomarkers of appetite are also often measured in human studies, and
include changes to gut hormone concentrations. These are often a preferred
measurement of changes to appetite, as they are less likely to be influenced by
other factors.
12.3 PROPOSED MECHANISMS FOR AN EFFECT OFRESISTANT STARCH ON APPETITE
Although the exact mechanism as to how resistant starch (RS) may affect
appetite is not known, various mechanisms have been proposed. These
mechanisms may be specific to RS or may be ubiquitous for all dietary fibres.
As RS is not digested in the small intestine but is fermented in the colon, the
most likely mechanism behind an effect on appetite is due to fermentation and
the subsequent increase in production of short-chain fatty acids (SCFA). The
SCFA may increase production of GLP-1 and PYY which, in turn, may
activate the neurons in the hypothalamus, therefore affecting behaviour and
causing changes to food intake (Figure 12.1).
The cells that secrete GLP-1 and PYY are primarily located in the ileum
and colon. GLP-1 receptors have been located in the hypothalamus (Murphy
& Bloom, 2004; Holst, 2007), therefore suggesting a role for GLP-1 in
directly affecting food intake. This has been shown in rodent models, where
central infusion of GLP-1 reduced food intake (Turton et al., 1996). GLP-1
may also indirectly affect food intake via the vagus nerve, as studies in rodents
Resistant starch
consumption
Increased SCFA
Increased GLP-1 and
PYY
Activation ofneurons in
ARC
Modifiedbehaviour/appetite
Decreasedfood intake
Figure 12.1 Possible pathway by which resistant starch consumption affects appetiteand food intake.
The Acute Effects of Resistant Starch on Appetite and Satiety 217

3GCH12 07/20/2013 9:50:29 Page 218
have shown that, following a vagotomy, the anorexic effects of GLP-1 are lost
(Dhillo, 2007). Studies where GLP-1 has been infused in humans provide
mixed results, with some studies showing an effect (Flint et al., 1998; Naslund
et al., 1999) and others none (Long et al., 1999; Naslund et al., 1998).
However, a meta-analysis (which included the above studies in addition to
others) showed a dose-dependent reduction in food intake following
GLP-17–36 (biologically active form) peripheral administration in humans
(Verdich et al., 2001).
Similarly, studies that have infused PYY in rodents and humans have found
a reduction in food intake (Batterham et al., 2002; Challis et al., 2003). In one
instance, the decrease in energy intake was 30% following peripheral infusion
of the active form (PYY3–36) in humans (Batterham et al., 2003). PYY may
also be released due to neural reflex, potentially mediated by the vagus nerve
(Fu-Cheng et al., 1997).
It has also been shown that while these hormones can reduce food intake
independently, when they are administered together, the effect on reducing
food intake is additive (Neary et al., 2005; Steinert et al., 2010).
Therefore, as these two hormones have been shown to reduce food intake,
they are likely to be the most affected by RS consumption due to fermentation
in the colon.
12.4 RODENT DATA
Results from rodent studies have shown promising effects of RS intake on
appetite and food intake, as well as on gut hormone regulation. However, these
rodent studies have not been acute (10–65 days), so therefore the translation of
the data from these experiments to acute affects in humans is not possible. As
these studies do provide some evidence for potential mechanisms, they will be
discussed briefly below and are summarised in Table 12.1.
Based on the potential pathway by which RS may affect appetite and food
intake described above (Figure 12.1), there is evidence in rodent models for
each of the steps as shown in Figure 12.2 and discussed in more detail below.
A few experiments in rodents have shown increased concentrations of
SCFA following RS consumption in studies relating to appetite work, as well
as effects of the RS consumption on changes to gene expression. Keenan et al.
(2006) proved significantly higher caecal SCFA concentrations following a
RS diet compared to a non-fermentable cellulose energy controlled diet; they
also observed increased PYY and proglucagon (the precursor molecule for
GLP-1) gene transcription in the caecum and large intestine following RS
compared to the control. Zhou et al. (2006) showed that, after a RS diet,
218 Resistant Starch


3GCH12 07/20/2013 9:50:29 Page 220
compared to control, PYY and proglucagon mRNA expression were signifi-
cantly unregulated in the caecum and colon. Further work by Zhou et al.
(2008) also showed in vitro that the fermentation of RS increased the
concentrations of SCFA, and that the SCFA (when at concentrations similar
to those observed in the caecal content in the study by Keenan et al. (2006))
directly stimulated proglucagon gene expression.
Several studies in animals have shown increased GLP-1 and PYY concen-
trations following RS consumption. The study by Keenan et al. (2006) found
greater serum PYY and GLP-1 concentrations following the RS diet, but no
effect on serum ghrelin or cholecystokinin (CCK) concentrations. Plasma
PYYand GLP-1 concentrations were also increased in the study by Shen et al.
(2009). Zhou et al. (2008) found RS resulted in significantly higher plasma
GLP-1 and PYY concentrations throughout the day. Aziz et al. (2009)
compared high-amylose with high amylopectin diets and found that the
high-amylose diets resulted in higher plasma GLP-1 and PYY concentrations.
There is also evidence from rodent studies for an effect of RS consumption
on activation of neurons in the hypothalamus. Shen et al. (2009) found that,
following a high RS diet, POMC expression in the ARC was significantly
upregulated, but there was no effect on NPY or AgRP. So et al. (2007)
conducted manganese-enhanced magnetic resonance imaging during ad
libitum food intake to monitor neuronal activity in the areas of the hypo-
thalamus involved in appetite regulation and found that there appeared to be a
satiating effect of the RS diet compared to the rapidly digestible starch.
Most of these studies, in the course of looking at possible mechanisms
behind an effect of RS on appetite, have also reported changes to food intake
and weight, with mixed findings. Aziz et al. (2009) reported that the high-
amylose diets, when consumed ad libitum, resulted in lower energy intakes,
weight gain and fat pad mass. Similarly, So et al. (2007) also reported lower
Resistant starch
consumption
Increased SCFA
Increased GLP-1 and
PYY
Activation ofneurons in
ARC
Modifiedbehaviour/appetite
Decreasedfood intake
Increased gene
expression in caecum and
colon
Lower body weight and
fat mass
Figure 12.2 Possible pathway by which resistant starch consumption affects foodintake and behaviour in rodents.
220 Resistant Starch

3GCH12 07/20/2013 9:50:29 Page 221
energy intakes in the RS group and found that, although body weight was the
same in both groups, the RS group had lower total, subcutaneous, visceral and
liver fat, and also smaller adipocytes. However, the studies by Keenan et al.
(2006), Zhou et al. (2008) and Shen et al. (2009) found similar food intakes
between groups, but lower body weights and fat mass in the RS group.
From these studies in rodents, it is clear that high RS intakes result in
increases in both GLP-1 and PYY (satiety hormones), most likely due to
increased production of SCFA from fermentation. Although, in some of these
studies, RS appears to decrease both body weight and fat mass, the effects on
actual energy and food intake are less clear.
In many of these animal experiments, the amount of RS given would be
much greater than would normally be consumed by a human. Similarly, the
length of time (normally days to weeks) that the rodents were fed the starch is
difficult to compare to the same time span in humans. The gut physiology of
rodents is also very different (greater large bowel length) from that of humans
and, therefore, fermentation effects in a rodent can not be directly translated
into similar effects to be observed in humans.
While the evidence for an effect of RS on appetite in rodent studies is
positive, these results have yet to be demonstrated fully in humans.
12.5 HUMAN DATA
Compared to the data from rodent studies, the acute effects (up to 48 hours) of
RS on appetite are less clear in humans.
There are few human studies that have directly investigated the effects of
RS on appetite and food intake, with these studies providing mixed findings
(summarised in Table 12.2). Direct comparisons are hampered by the use of
different protocols and different types of RS (the majority of these studies
used type 2 or 3). Some early studies have also compared the different effects
of amylose and amylopectin on appetite and food intake. As RS is high in
amylose, the results from these studies are included.
Based on the potential pathway by which RS may affect appetite and food
intake described above (Figure 12.1), there is only some evidence in humans for
some of the steps, as shown in Figure 12.3 and discussed in more detail below.
Very few human studies have reported the effects of RS on GLP-1 and
PYY. In the study by Raben et al. (1994) it was found that there were lower
plasma GLP-1 concentrations with RS, compared with their placebo.
Bodinham et al. (2013) also found lower GLP-1 concentrations following
a breakfast containing RS compared to a placebo. A study by Klosterbuer et al
(2012) found no effect of RS on GLP-1 concentrations. Although a study by
The Acute Effects of Resistant Starch on Appetite and Satiety 221
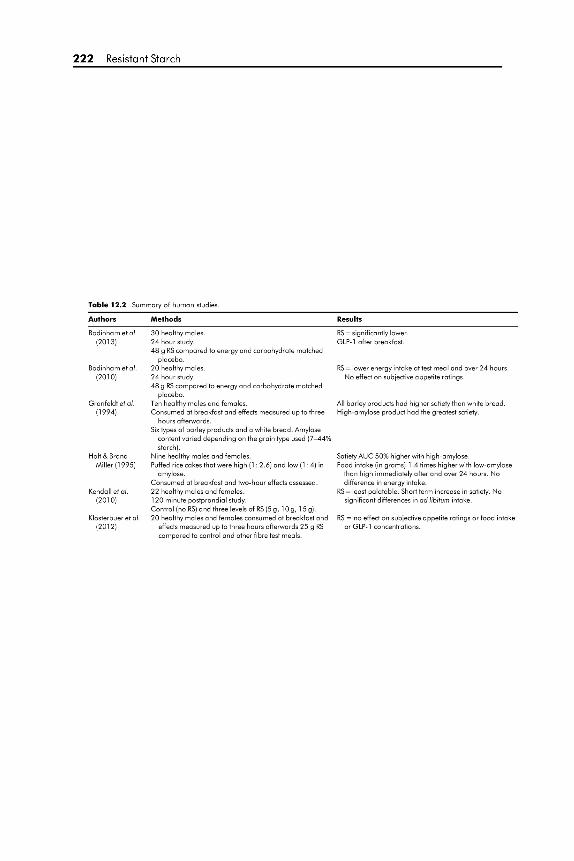


3GCH12 07/20/2013 9:50:30 Page 224
Robertson et al. (2003) was not investigating the effects of RS on appetite, one
of their outcomes was the effects of RS consumption on GLP-1 concentra-
tions, and this also showed no effect. The lack of evidence for an effect on gut
hormones in humans could be due to the neuronal effects of GLP-1 and PYY
which would not be measured in the general circulation. Alternatively, as
GLP-1 is rapidly broken down in the circulation by dipeptidyl peptidase IV
(DPP-IV) and both hormones are quickly cleared from the blood via the liver
(Holst, 2007), the hormones may not appear in peripheral blood at raised
concentrations, but there may be raised concentrations in splanchnic blood.
Several studies have measured the effects of RS on modifying appetite
using changes to subjective questionnaires. These studies have also reported
mixed findings:
� three studies have reported no differences between RS/high-amylose and a
control (Weststrate & van Amelsvoort, 1993; Bodinham, et al., 2010;
Klosterbuer et al. 2012);� one study reported lower satiety with RS compared to a control (Raben
et al., 1994);� several studies have reported greater satiety with RS (Kendall et al., 2010;
Willis et al., 2009; Nilsson et al., 2008; Liljeberg et al., 1999; Holt & Brand
Miller, 1995; Granfeldt et al., 1994; van Amelsvoort & Weststrate, 1992).
However, some of these studies also found that the RS supplement was less
palatable than the control used (Kendall et al., 2010; Willis et al., 2009; van
Amelsvoort &Weststrate, 1992), which could account for the effects observed.
Only a few studies have investigated the effects of RS on actual food intake.
Two of these studies (Kendall et al., 2010; Klosterbuer et al., 2012) found no
Resistant starch
consumption
Increased SCFA
Increased GLP-1 and
PYY
Activation ofneurons in
ARC
Modifiedbehaviour/appetite
Decreasedfood intake
Increased gene
expression
Lower body weight and
fat mass
Figure 12.3 Possible pathway by which resistant starch consumption affects foodintake and behaviour in humans. Circles show areas where there is no acute evidencein human work.
224 Resistant Starch

3GCH12 07/20/2013 9:50:30 Page 225
significant differences between treatments for energy intake at the ad libitum
meal. However, two other studies (Bodinham et al., 2010; Holt & BrandMiller,
1995) found a reduced intake with RS. The study by Bodinham et al. (2010)
found the reduced energy intake at both an ad libitum test meal (seven hours
postprandially) and over a 24-hour period. Holt & Brand Miller (1995) found
that intake (in grams) following the high-amylose rice cake was significantly
lower after two hours and over the rest of the day, compared to the low-amylose
rice cake. However, the differences in energy were not significant.
Based on the evidence from the studies using RS and differences in
amylose/amylopectin ratios, there is an acute effect of RS on appetite. In the
few studies where GLP-1 concentrations have been measured (Raben et al.,
1994, Robertson et al., 2003) in human studies, RS does not appear to have
an effect. As the data in rodents suggest that effects on satiety hormones may
be a mechanism by which RS affects appetite, satiety and food intake,
further investigations in humans would be required. Further studies with
similar study designs are also required to determine the dose that is needed
for an effect to occur, as well as other possible mechanisms behind
the effects.
REFERENCES
Aziz, A.A., Kenny, L.S., Goulet, B., Abdel-Aal, E.S. (2009). Dietary starch type affectsbody weight and glycemic control in freely fed but not energy-restricted obese rats.Journal of Nutrition 139(10), 1881–9.
Batterham, R.L., Cowley, M.A., Small, C.J., Herzog, H., Cohen, M.A., Dakin, C.L., Wren,A.M., Brynes, A.E., Low, M.J., Ghatei, M.A., Cone, R.D., Bloom, S.R. (2002). Guthormone PYY3–36 physiologically inhibits food intake. Nature 418, 650–4.
Batterham, R.L., Cohen, M.A., Ellis, S.M., Le Roux, C.W., Withers, D.J., Frost, G.S.,Ghatei, M.A., Bloom, S.R. (2003). Inhibition of food intake in obese subjects by peptideYY3–36. New England Journal of Medicine 349, 941–8.
Bodinham, C.L., Frost, G.S., Robertson, M.D. (2010). Acute ingestion of resistantstarch reduces food intake in healthy adults. British Journal of Nutrition 103,917–22.
Bodinham, C.L., Al-Mana, N.M., Smith, L., Robertson, M.D. (2013). Endogenous plasmaglucagon-like peptide-1 following acute dietary fibre consumption. British Journal ofNutrition doi:10.1017/50007114513000731.
Challis, B.G., Pinnock, S.B., Coll, A.P., Carter, R.N., Dickson SL., O’Rahilly, S. (2003).Acute effects of PYY3–36 on food intake and hypothalamic neuropeptides expression inthe mouse. Biochemical and Biophysical Research Communications 311, 915–9.
Dhillo, W.S. (2007). Appetite regulation: an overview. Thyroid 17(5), 433–45.Flint, A., Raben, A., Astrup, A., Holst. J.J. (1998). Glucagon-like peptide 1 promotessatiety and suppresses energy intake in humans. Journal of Clinical Investigation 101,515–20.
The Acute Effects of Resistant Starch on Appetite and Satiety 225

3GCH12 07/20/2013 9:50:31 Page 226
Fu-Cheng, X., Anini, Y., Chariot, J., Castex, N., Galmiche, J.P., Roze, C. (1997).Mechanisms of peptide YY release induced by an intraduodenal meal in rats: neuralregulation by proximal gut. Pfl€ugers Archiv 433, 571–9.
Granfeldt, Y., Liljeberg, H., Drews, A., Newman, R., Bjorck, I.M. (1994). Glucose andinsulin responses to barley products: influence of food structure and amylose-amylo-pectin ratio. American Journal of Clinical Nutrition 59, 1075–82.
Holst, J.J. (2007). The physiology of glucagon-like peptide 1. Physiological Reviews 87,1409–39.
Holt, S.H.A., Brand Miller, J. (1995). Increased insulin responses to ingested foods areassociated with lessened satiety. Appetite 24, 43–54.
Keenan, M.J., Zhou, J., McCutcheon, K.L., Raggio, A.M., Bateman, H.G., Todd, E., Jones,C.K., Tullet, R.T., Melton, S., Martin, R.J., Hegsted, M. (2006). Effects of resistantstarch, a non-digestible fermentable fibre, on reducing body fat. Obesity 14, 1523–34.
Kendall, C.W.C., Esfahani, A., Sanders, L.M., Potter, S.M., Vidgen, E. (2010). The effectof a pre-load meal containing resistant starch on spontaneous food intake and glucoseand insulin responses. Journal of Food Technology 8, 67–73.
Klosterbuer, S.S., Thomas , W., Slavin, J.L. (2012). Resistant starch and pullulan reducepostprandial glucose, insulin and GLP-1, but have no effect on satiety in healthy humans.Journal of Agricultural and Food Chemistry 60(48), 11928–11934.
Liljeberg, H.G.M., Akerberg, A.K.E., Bjorck, I.M. (1999). Effect of the glycemic index andcontent of indigestible carbohydrates of cereal-based breakfast meals on glucosetolerance at lunch in healthy subjects. American Journal of Clinical Nutrition 69,647–55.
Long, S.J., Sutton, J.A., Amaee, W.B., Giouvanoudi, A., Spyrou, N.M., Rogers, P.J.,Morgan, L.M. (1999). No effect of glucagon-like peptide-1 on short-term satiety andenergy intake in man. British Journal of Nutrition 81, 273–9.
Mattes, R.D., Hollis, J.H., Hayes, D., Stunkard, A.J. (2005). Appetite: measurementand manipulation misgivings. Journal of the American Dietetic Association 105,S87–97.
Murphy, K.G., Bloom, S.R. (2004). Gut hormones in the control of appetite. ExperimentalPhysiology 89, 507–16.
Naslund, E., Gutniak, M., Skogar, S., Rossner, S., Hellstrom, P.M. (1998). Glucagon-likepeptide 1 increases the period of postprandial satiety and slows gastric emptying in obesemen. American Journal of Clinical Nutrition 68, 525–30.
Naslund, E., Barkeling, B., King, N., Gutniak, M., Blundell, J.E., Holst, J.J., Rossner,S., Hellstrom, P.M. (1999). Energy intake and appetite are suppressed by glucagon-likepeptide-1 (GLP-1) in obese men. International Journal of Obesity and RelatedMetabolic Disorders 23, 304–11.
Neary, N.M., Goldstone, A.P., Bloom, S.R. (2004). Appetite regulation: from the gut to thehypothalamus. Clinical Endocrinology 60, 153–60.
Neary, N.M., Small, C.J., Druce, M.R., Park, A.J., Ellis, S.M., Semjonous, N.M., Dakin,C.L., Filipsson, K., Wang, F., Kent, A.S., Frost, G.S., Ghatei, M.A., Bloom, S.R. (2005).Peptide YY3–36 and glucagon-like Peptide-17–36 inhibit food intake additively. Endo-crinology 146, 5120–7.
Nilsson, A.C., Ostman, E.M., Holst, J.J., Bjorck, I.M. (2008). Including indigestiblecarbohydrates in the evening meal of healthy subjects improves glucose tolerance,lowers inflammatory markers, and increases satiety after a subsequent standardizedbreakfast. Journal of Nutrition 138, 732–9.
226 Resistant Starch

3GCH12 07/20/2013 9:50:31 Page 227
Raben, A., Tagliabue, A., Christensen, N.J., Madsen, J., Holst, J.J., Astrup, A. (1994).Resistant starch: the effect on postprandial glycemia, hormonal response and satiety.American Journal of Clinical Nutrition 60, 544–51.
Robertson, M.D., Currie, J.M., Morgan, L.M., Jewell, D.P., Frayn, K.N. (2003). Prior short-term consumption of resistant starch enhances postprandial insulin sensitivity in healthysubjects. Diabetologia 46, 659–65.
Shen, L., Keenan, M.J., Martin, R.J., Tulley, R.T., Raggio, A.M., McCutcheon, K.L., Zhou,J. (2009). Dietary resistant starch increases hypothalamic POMC expression in rats.Obesity 17, 40–5.
So, P.W., Yu,W.S., Kuo, Y.T.,Wasserfall, C., Goldstone, A.P., Bell, J.D., Frost, G.S. (2007).Impact of resistant starch on body fat patterning and central appetite regulation. PLoSOne 2(12), e1309.
Steinert, R.E., Poller, B., Castelli, M.C., Drewe, J., Beglinger, C. (2010). Oral administra-tion of glucagon-like peptide 1 or peptide YY 3–36 affects food intake in healthy malesubjects. American Journal of Clinical Nutrition 92, 810–7.
Turton, M.D., O’Shea, D., Gunn, I., Beak, S.A., Edwards, C.M.B., Meeran, K., Choi, S.J.,Taylor, G.M., Heath, M.M., Lambert, P.D., Wilding, J.P.H., Smith, D.M., Ghatei, M.A.,Herbert, J., Bloom, S.R. (1996). A role for glucagon-like peptide-1 in the centralregulation of feeding. Nature 379, 69–72.
van Amelsvoort, J.M.M., Westsrate, J.A. (1992). Amylose-amylopectin ratio in a mealaffects postprandial variables in male volunteers. American Journal of Clinical Nutrition55, 712–8.
Verdich, C., Flint, A., Gutzwiller, J.P., Naslund, E., Beglinger, C., Hellstrom, P.M., Long,S.J., Morgan, L.M., Holst, J.J., Astrup, A. (2001). A meta-analysis of the effect ofglucagon-like peptide-1 (7–36) amide on ad libitum energy intake in humans. Journal ofClinical Endocrinology and Metabolism 86, 4382–9.
Weststrate, J.A., van Amelsvoort, J.M.M. (1993). Effects of the amylose content ofbreakfast and lunch on postprandial variables in male volunteers. American Journalof Clinical Nutrition 58, 180–6.
Willis, H.J., Eldridge, A.L., Beiseigel, J., Thomas, W., Slavin. J.L. (2009). Greater satietyresponse with resistant starch and corn bran in human subjects. Nutrition Research 29,100–5.
Wynne, K., Stanley, S., McGowan, B., Bloom, S. (2005). Appetite control. Journal ofEndocrinology 184, 291–318.
Zhou, J., Hegsted, M., McCutcheon, K.L., Keenan, M.J., Xi, X., Raggio, A.M., Martin, R.J.(2006). Peptide YY and proglucagon mRNA expression patterns and regulation in thegut. Obesity 14, 683–9.
Zhou, J., Martin, R.J., Tulley, R.T., Raggio, A.M., McCutcheon, K.L., Shen, L., Danna,S.C., Tripathy, S., Hegsted, M., Keenan, M.J. (2008). Dietary resistant starch up-regulates total GLP-1 and PYY in a sustained day-long manner through fermentationin rodents. American Journal of Physiology. Endochrinology and Metabolism 295(5),E1160–6.
The Acute Effects of Resistant Starch on Appetite and Satiety 227

3GCH12 07/20/2013 9:50:31 Page 228

CH13GR 07/29/2013 16:49:28 Page 229
13 Metabolic Effects of Resistant Starch
Martine ChampINRA, UMR 1280, Physiologie des Adaptations Nutritionnelles, Universite deNantes, CRNH, IMAD, CHU de Nantes, Nantes, France
Resistant starch (RS) is currently defined as the sum of starch and products of
starch degradation not absorbed in the small intestine of healthy individuals
(Asp, 1992). As a consequence, RS includes not only fractions which are
resistant to endogenous enzyme digestion, but also starch and dextrins which
are potentially digestible but which, due to too short interaction with endog-
enous alpha-amylase, arrive in the colon. RS, whatever, its bioavailability, is
submitted to digestion and fermentation by colonic microbiota. Most of the RS
disappears in the colon and is metabolized into short-chain fatty acids (acetate,
propionate and butyrate) and gases (CO2, H2 and CH4).
Metabolic effects of resistant starch could a priori be classified according
to:
1. the type of metabolic effect of RS or of its metabolites; or
2. the mechanism which is involved – it can potentially be ‘direct’, due to a
matrix effect as shown for soluble (and viscous) dietary fibre, or mediated
by the short chain fatty acids produced from RS fermentation by the
colonic microbiota.
A third category is a subject of controversy. Indeed, it is linked to the effect
of the substitution of digestible starch by resistant starch. As a consequence, it
is not the metabolic effect of RS per se, but the decrease of the ‘digestible’ part
of the starch, which is mostly responsible for the metabolic effect that can be
observed.
In most investigations of metabolic studies with RS, the mechanism(s)
involved has not been elucidated fully, and/or two different mechanisms can
be simultaneously responsible for the physiological effects which have been
described. As a consequence, the first classification will be adopted all along
the review, but mechanisms involved will be discussed.
229
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

CH13GR 07/29/2013 16:49:28 Page 230
In order to conclude to a metabolic effect of RS through its intrinsic
properties, the design of the study has to be faultless, particularly with regard
to the formulation of the experimental diet. Indeed, with most RS sources
containing digestible starch, the ‘true’ RS concentration in the food/ingredient
(as eaten) has to be determined prior to the clinical trial in order to adapt the
level of incorporation of the food/ingredient to this concentration.
Unfortunately, this statement is neglected in several studies, leading to a
misinterpretation of the results. Moreover, physiological or metabolic effects
can be misattributed to RS when it is present in a diet/food/ingredient which
also has a low glycemic index.
This review will mostly refer to metabolic impact of resistant starch.
Broader overviews on the physiological effects of resistant starch have been
published earlier (Champ, 2004; Champ et al., 2003).
13.1 FERMENTATION OF RS AND ITS IMPACTON COLONIC METABOLISM
RS fermentation is characterized by a significant ratio of butyrate among
SCFAs produced by its fermentation, compared to most dietary fibres (see
Tables 13.1 and 13.2).Most of the data available on RS fermentation are issued from in vitro
fermentation of RS by animal or human faecal inocula (see Table 13.1).
Studies made with effluents from ileostomy from human patients are a priori
the most accurate (Langkilde et al., 2002; Silvester et al., 1995). However an
appropriate (validated procedure) predigestion of an RS-containing food can
be considered as a relevant method to obtain the samples which have to be
submitted to in vitro fermentation (F€assler et al., 2006). Finally, many
studies have been performed on rodents or other animal species, but their
microbiota are significantly different from human microbiota and fermen-
tation profiles cannot be extrapolated to humans even if, in some cases,
a similar feature is observed between species (Wang et al., 2004; Murray
et al., 2001).
Several data of concentration of SCFAs in human stools are available in the
literature (Hylla et al., 1998; Jenkins et al., 1998; Noakes et al., 1996;
Cummings et al., 1996; Phillips et al., 1995; van Munster et al., 1994). These
are a partial reflection of fermentation, as most SCFA are absorbed in
the colon. However, they probably reflect both the intensity and the profile
of the fermentation (Table 13.2).
230 Resistant Starch

CH13GR 07/29/2013 16:49:28 Page 231
Recent study from Verbeke et al. (2010) demonstrated higher concentra-
tion of butyrate in both plasma and urine from volunteers who ingested 13C
enriched intact barley kernels (containing RS and non starch polysaccharides)
compared to barley porridge (containing only non starch polysaccharides).
The investigation of RS fermentation in pigs by quantification of SCFA in
portal blood (Martin et al., 2000) raised the question of the significance of the
measurement of butyrate out of the site of production. Indeed, butyrate uptake
by the colonic mucosa seems to vary according to fermentation rate of RSwith
a higher uptakewhen fermentation is slow compared to fast fermentation. As a
consequence, quantification of butyrate in blood, urine or faeces only reflects
part of the production of butyrate.
Butyrate is the main fuel of the healthy colonocyte and is consumed by
these cells, explaining why only minor amounts of butyrate can be found in the
peripheral blood. Its depletion is thought to induce damage to the colonic
epithelium. As a consequence, dietary fibres inducing butyrate production,
such as RS, are described as ingredients that have the potential to prevent
colonic diseases such as colon cancer (Le Leu et al., 2009, 2007a, 2007b;
Clarke et al., 2008; Bauer-Marinovic et al., 2006; Perrin et al., 2001;
Wollowski et al., 2001; Hylla et al., 1998; Young & Gibson, 1995).
Butyratewould also be of interest in the healing of ulcerative colitis (Vernia
et al., 2003), even if most recent data only partly confirm earlier findings
(Hamer et al., 2010). Most studies, however, confirm that butyrate oxidation
by the colonic mucosa is impaired in various situations of inflammation (de
Preter et al., 2009; Jacobasch et al., 1999).
It is now admitted that RS can be a good substrate for butyrate production
in the colon (Le Leu et al., 2009, 2007a). The restoration of the integrity of
rat caeco-colonic mucosa in chemically induced colitis by RS, but not
by fructo-oligosaccharides (Moreau et al., 2003), probably reveals that
fermentation profile and kinetic of fermentation are determinant factors of
the benefits of dietary fibres on colonic diseases (in its broadest definition;
Howlett et al., 2010).
According to several authors, RS would improve colonic mucosal integrity
in healthy animals and/or reduce colonic and systemic immune reactivity for
which health benefits in inflammatory conditions are likely to be associated
(Nofrar�ıas et al., 2007). It would confirm the interest of RS in a prevention
context of several colonic diseases.
Apart from butyrate, RS fermentation induces production of acetate and
propionate. Both SCFAs are absorbed, but part is metabolized by the liver,
whereas mostly acetate can be found in the blood and used by peripheral
organs (Wong et al., 2006).
Metabolic Effects of Resistant Starch 231

3GCH13 08/31/2013 15:32:54 Page 232
232 Resistant Starch
Table 13.1 In vitro fermentation of resistant starches.
Sample Specie Total SCFA Acetate
mmol/L mmol/g mmol/L mmol/g molar
ileal subst. ileal subst. ratio
Raw banana (flour) Ileal
effluents
Human
(ileostomy)
120�2.8 5.5�0.1 62.3�3.0 2.9�0.1 55�1
vs.
Cooked banana (flour) 107�3.1 7.0�0.2 54�2.0 3.5�0.1 55�1
High RS Ileal
effluents
Human
(ileostomy)
69.6�6.9 4.9� 0.4 44.1�4.5 3.2�0.3 66�1.6
vs.
Low RS 40.9�3.9 4.8�0.3 27.9�2.5 3.3�0.2 72�1.1
mmol mmol/L molar
ratio
Raw potato starch Ileal
effluents
Pig (ileal
cannulation)
13.3 20 53.6
vs.
Control 12.3 17.5 52.3
mmol/
g OM
mmol/
g OM
Barley Native Ileal
effluents
Dog 3.61 2.66
HT extrusion (ileal
cannulation)
6.08 4.27
Corn Native 3.13 2.26
HT extrusion 3.68 3.08
Potato Native 1.64 1.46
starch LT extrusion 6.86 4.56
Rice Native 5.45 3.82
HT extrusion 7.45 5.74
Sorghum Native 3.70 2.67
HT extrusion 6.47 4.13
Wheat Native 2.85 2.17
HT extrusion 8.13 5.38
mmol molar ratio
Preparation RS3 in vitro
predig. (ba)
161 31
(retrograded long chain in vitropredig. (dy)
217 35
maltodextrin)
Ileal
effluents
Human
(ileostomy)
189 34
Preparation RS2 in vitropredig. (ba)
150 32
(native high amylose
in vitropredig. (dy)
87 23corn starch)
ba: batch; dy: dynamic.

3GCH13 08/31/2013 15:32:54 Page 233
Propionate Butyrate Reference
mmol/L mmol/g molar mmol/L mmol/g molar
ileal subst. ratio ileal subst. ratio
17.0�0.5 0.8�0.0 14.0�0.5 34.8�1.1 1.6�0.1 31.0�0.9 Langkilde
et al., 2002
18.3�0.6 1.2�0.1 18 0�4 27.0�0.7 1.7�0.1 27.0�0.9
11.8�1.3 0.8�0.1 17�0.7 11.1�1.1 0.8�0.1 17�1.1 Silvester et al.,1995
6.3�0.9 0.7�0.1 16�1.1 4.5�0.6 0.5�0.1 12�0.7
mmol mmol/L molar mmol mmol/L molar
ratio ratio
6.1 8.2 22.5 4.2 5.7 15.5 Wang et al.,2004
5.9 7.8 23.2 3.2 4.5 13.5
mmol/g OM mmol/g OM
0.59 0.36 Murray et al.,2001
0.85 0.96
0.45 0.42
0 0.60
0.11 0.07
1.42 0.88
0.92 0.71
1.53 1.18
0.67 0.36
1.61 0.73
0.52 0.16
1.50 1.25
molar ratio mmol molar ratio
48 32.6 20 F€assler et al.,
2006
49 35.5 16
57 17.0 9
43 35.8 25
51 22.5 26
Metabolic Effects of Resistant Starch 233


CH13GR 07/29/2013 16:49:36 Page 235
13.2 RESISTANT STARCH, GLYCEMIA, INSULINAEMIAAND GLUCOSE TOLERANCE
When substituting digestible starch, RS obviously reduces glycemic load of
the food/diet and, consequently, reduces insulin secretion (Raben et al., 1994).
When comparing raw potato starch (50 g containing 54% RS) to pre-
gelatinized potato starch (50 g), postprandial plasma concentrations of
glucose and insulin were lower with the RS diet, compared to the control
diet. Ranganathan et al. (1994) showed no alteration of postprandial glucose
response to a 50 g glucose load when lintnerized starch (30 g of lintner from
maize starch containing 70% amylose) was added to the experimental meal. A
lintner is a starch which has been treated with acidic solution and is poorly
fermented by the microbiota. The conclusion of the study was that the acute
effect of the ingestion of a lintnerized starch on the measured metabolic
indexes is similar to that of cellulose (insoluble and poorly fermentable fibre).
Resistant starch (RS3, PromitorTM RS, Tate & Lyle Inc, IL, USA) (from
5–25 g in a meal) has been shown to have no effect on incremental area under
the curve (iAUC) for glucose or insulin. However, there was a decrease of the
incremental blood glucose and insulin levels after a meal containing 25 g RS at
90 and 120 minutes after the meal (p¼ 0.004 and 0.001 for glucose, p¼ 0.043
and p¼ 0.042 for insulin, respectively) (Kendall et al., 2010).
The comparison of 70% amylopectin to 70% amylose corn starch (proba-
bly RS2) in normal and hyperinsulinaemic men (14-week intervention cross-
over study) revealed that the magnitude of the response in carbohydrate and
fat oxidation was blunted in hyperinsulinaemic subjects consuming excess
levels of the amylose diet (Howe et al., 1996). The hypotheses of the authors
were that this was due to:
1. an improvement in overall insulin response; or
2. a change in available substrates for oxidation resulting from microbial
fermentation.
The data from Brighenti et al. (2006) would tend to support the hypothesis
of a second-meal effect of fermentable carbohydrates such as RS. Indeed, both
a ‘slowly digestible, partly fermentable high-amylose starch (Hylon VII) plus
cellulose’ meal and an ‘amylopectin starch plus lactulose’ meal, ingested
within breakfast, improved glucose tolerance at lunch. According to these
authors, a reduction of non-esterified fatty acids (NEFA) competition for
glucose disposal might be a mechanism involved in this metabolic effect, even
if the result is not statistically significant for the RS containing meal, whereas
it is with the lactulose-containing meal (Brighenti et al., 2006).
Metabolic Effects of Resistant Starch 235

CH13GR 07/29/2013 16:49:36 Page 236
This observation has not been made by Liljeberg et al. (1999), who
described a beneficial effect of slow absorption and digestion of starch
from a breakfast meal on glucose tolerance at the second meal (lunch),
whereas the content of indigestible carbohydrates (RS and dietary fibre) had
no effect. But the same group (Nilsson et al., 2008) demonstrated that an
evening meal containing RS and dietary fibre (barley kernel bread) signifi-
cantly improved glucose tolerance at a following standardized breakfast
(28%, compared to white wheat bread), independently of the glycemic index
of the evening meal. The benefits of RSþDF were attributed to colonic
fermentation, which may be characterized by a prebiotic effect.
de Roos et al. (1995) compared daily supplement of 30 g RS2 (high-
amylose corn starch) to RS3 (retrograded high-amylose corn starch), using
glucose as a control. They determined C-peptide excretion as a measure for
the 24-hour insulin secretion. They concluded that consumption of 30 g/day
RS3 (but not RS2) during one week reduced the insulin secretion. This result
would also suggest an impact of RS fermentation rate on insulin.
Robertson et al. (2005) suggested that dietary supplementation with RS
had the potential to improve insulin sensitivity. Indeed, insulin sensitivity
assessed by euglycaemic-hyperinsulinaemic clamp and meal tolerance test
was higher after RS supplementation (30 g RS/day during four weeks) than
after placebo treatment of healthy subjects. More recently, the same group
(Johnston et al., 2010) confirmed an improvement of insulin sensitivity of
insulin resistant subjects (40 g RS/day during 12 weeks); however, this
modification was not associated with changes in whole-body composition.
These results apparently contradict those obtained from studies on rodents.
Indeed, a high-RS diet has been shown to decrease intra-hepatic fat storage
and reduce inflammatory markers (Kabir et al., 1998; So et al., 2007).
However, these studies on animal models have compared RS to digestible
starch, which is not the case with Robertson’s studies (Robertson et al., 2005).
Kwak et al. (2012) observed that a four-week dietary treatment with rice
containing resistant starch (6.51 g RS per day, compared with control com-
posed of rice with low level of RS) reduced postprandial blood glucose and
oxidative stress, measured as plasma malondialdehyde (MDA), urinary 8-epi-
PGF (2a) (whereas RH-PAT index and total nitric oxide were increased) in
patients with prediabetes or newly diagnosed type 2 diabetes.
13.3 RS CONSUMPTION AND LIPID METABOLISM
De Deckere et al. (1993) concluded, from a study on rats fed semi-purified
diets containing a low (0.8 g/MJ) or a high (9.6 g/MJ) amount of RS, that
236 Resistant Starch

CH13GR 07/29/2013 16:49:36 Page 237
dietary RS can reduce serum total cholesterol and triacylglycerols concentra-
tion and fat accretion (epididymal fat pads).
Cheng& Lai (2000) observed reduced serum and hepatic cholesterol in rats
fed a diet containing 63% rice starch (richer in RS than corn starch), compared
to animals receiving corn starch as a control. The authors attributed this effect
to the significant increase of propionate in rats fed on rice starch. Propionate
may, indeed, be involved in the control of hepatic cholesterol synthesis.
However, the experimental diets had different amount of digestible starch and
it cannot be ignored that the impact on cholesterol concentrations could be
explained by the different insulin responses to the digestible starch fractions of
the diets.
Moreover, a study on germ-free rats (Sacquet et al., 1983) also demon-
strated a reduction of plasma cholesterol with RS (RS2: high-amylose starch).
In this study, the hypothesis of an effect of SCFA cannot explain the beneficial
metabolic effect. Ten years later, the same group (Math�e et al., 1993)
described a reduction of plasma cholesterol in genetically obese (fa/fa)
and lean (fa/-) rats when fed a high-amylose corn starch diet. Among the
possible mechanisms, the authors suggested an increased biliary secretion of
bile acids linked to an accelerated catabolism of lipoproteins by the liver. A
second hypothesis, which has been mentioned earlier, is the involvement on
insulin (glucagon/insulin ratio) secretion, which would explain the reduced
plasma lipid and lipoprotein levels.
The impact of RS consumption in healthy human subjects is controver-
sial. Three of the studies (Stewart et al., 2010; Heijnen et al., 1996; Behall
& Howe, 1995) failed to demonstrate any favourable effect of RS con-
sumption on lipaemia or cholesterol concentration, whereas two earlier
studies (Behall et al., 1989; Reiser et al., 1989a, 1989b) concluded to a
significant decrease of total cholesterol and triacylglycerol when the
subjects received, respectively, 247 g and 183 g per day of high-amylose
corn starch during four and five weeks (compared to high amylopectin
starch and fructose as a control). Heijnen’s 1996 study was a randomized,
single-blind, 3� 3 Latin-square study (total of 57 subjects), in which the
subjects received 30 g of raw (RS2; Hylon VII, National Starch) or
retrograded RS (RS3; Novelose, National Starch) vs. glucose during three
weeks. In the 1995 study of Behall & Howe, the subjects received
approximately 200 g raw high-amylose (vs. low-amylose) corn starch
(RS2) per day. They observed a significant increase in total cholesterol
and a decrease in triacylglycerol after the first four weeks of supplementa-
tion, whereas Achour et al. (1997) concluded from an acute study that
retrograded high-amylose corn starch (RS3) leads to a reduction in lipolysis
in the post-absorptive period.
Metabolic Effects of Resistant Starch 237

CH13GR 07/29/2013 16:49:36 Page 238
Noakes et al. (1996) failed to observe any metabolic changes when high-
amylose starch (Hi-MaizeTM, Starch Australasia) (17 g and 25 g/day in
women and men, respectively) was introduced in the diet of hypertriglycer-
idaemic patients, whereas oat bran had a significant effect on plasma
triacylglycerol.
Results from Faisant et al. (unpublished personal data) indicated that
supplementing a normal meal with RS3 had little or no effect on postprandial
parameters of normal subjects. However, triacylglycerol metabolism was
improved, especially in subjects with higher (upper limit of the ‘normal’
range) basal triacylglycerol levels, most probably by an improvement in
clearance of postprandial chylomicron remnants.
13.4 RS CONSUMPTION, GIP, GLP-1 ANDPYY SECRETION
Glucagon-like peptide-1 (GLP-1) and peptide YY (PYY) are gut-secreted
peptides that have been proposed as potential anti-diabetes/obesity drugs.
These two hormones are also naturally secreted in response to meal ingestion,
but they degrade rapidly after endogenous secretion or exogenous injection.
Thus, pharmaceutical means to maintain substantial high plasma levels of
GLP-1 and PYY are intensely targeted.
RS has been reported by numerous authors to increase plasma total GLP-
1 and total PYY in rodents of both sexes at different ages (Keenan et al.,
2006; Zhou et al., 2006). For instance, Keenan et al. (2006) observed that
rats fed fermentable RS (RS2, Hi-MaizeTM 260, National Starch) had
(compared to methylcellulose) increased caecal weights and plasma PYY
and GLP-1 and increased gene transcription of PYYand proglucagon. They
conclude from their study that inclusion of RS in the diet may affect energy
balance through its effect as a fibre or a stimulation of PYY and GLP-1
expression. In the same year, Zhou et al. (2006), using in vivo and in vitro
approaches, concluded that the distal part of the gut has the ability to sense
nutrients such as butyrate, resulting in the up-regulation of PYY and
proglucagon gene expression.
Later on, the same group (Zhou et al., 2008) concluded from a study on
RS-fed rodents (Hi-MaizeTM 260 as RS2) that:
1. RS stimulated GLP-1 and PYY secretion in a substantial day-long manner,
independent of meal effect on changes in postprandial glycemia;
2. fermentation and the liberation of SCFAs in the lower gut are associated
with increased proglucagon and PYY gene expression;
238 Resistant Starch

CH13GR 07/29/2013 16:49:36 Page 239
3. glucose tolerance, an indicator of increased active forms of GLP-1 and
PYY, was improved in RS-fed diabetic mice.
They concluded from their study that fermentation of RS is most likely to
be the primarymechanism for increased endogenous secretions of total GLP-1
and PYY in rodents. Any factor that affects fermentation should be considered
when dietary fermentable fibre is used to stimulate GLP-1 and PYY secretion.
Tachon et al. (2013) observed that mice fed an energy-controlled diets
containing 18% or 36% type 2 RS (from high-amylose maize) were colonized
by higher levels of Bacteroidetes and Bifidobacterium, Akkermansia and
Allobaculum species in proportions that were dependent on the concentration
of the dietary fibre. The proportions of Bifidobacterium and Akkermansia
were positively correlated with mouse feeding responses, gut weight and
expression levels of proglucagon, the precursor of GLP-1.
To our knowledge, Raben et al. (1994) were the first to show that, in
humans (healthy subjects), when substituting digestible starch, RS reduces
insulin secretion but increases gastric inhibitory polypeptide (GIP), GLP-1
and epinephrine.
More recently, Nilsson et al. (2008) showed that improved tolerance was
observed after a standardized breakfast when RS and fibre (both from barley)
were ingested during the preceding evening meal. This was associated with
decreased concentration of free fatty acids and IL-6 and increased GLP-1 and
adiponectin at the time of the breakfast, thus providing evidence for a link
between the gut microbial metabolism and key factors associated with insulin
resistance.
13.5 RS CONSUMPTION, SATIETY AND SATIATIONAND FAT DEPOSITION
An acute randomised, single-blind crossover study aiming at determining the
effects of consumption of 48 g RS on appetite compared to an appropriate
control (energy and available carbohydrate-matched placebo) recently
showed a significantly lower energy intake following the RS supplement,
compared to the placebo supplement at the ad libitum test meal (Bodinham
et al., 2010). However, there was no associated effect on subjective appetite
measures.
According to de Roos et al. (1995), 30 g/day RS2 (raw high-amylose corn
starch) or RS3 (extruded and retrograded high-amylose corn starch) had little
influence on appetite and food intake. However, supplementation with RS2
caused significantly lower (p< 0.05) appetite scores than did supplementation
Metabolic Effects of Resistant Starch 239

CH13GR 07/29/2013 16:49:36 Page 240
with RS3 or glucose, though subjects paradoxically felt less full while
consuming RS2. This feeling might be related to a higher hydration capacity
of extruded retrograded starch compared to raw starch.
Kendall et al. (2010) compared the impact of lower doses of RS3 (from
5–25 g of PromitorTM RS, Tate & Lyle Inc, IL, USA) in a meal. Feelings of
fullness were greater with the 5 g dose of RS compared to control, while the
satiety quotient for overall appetite was significantly greater for 25 g RS in the
early phase after the eating period (p¼ 0.036, 0.075 and 0.97 at 45, 30 and
15 minutes after lunch).
In several studies, RS seems to have an impact on satiety which could be
mediated by acetate produced during colonic fermentation. Indeed, RS
reduced subjective satiety ratings in the intermediate absorptive period (Raben
et al., 1994; Achour et al., 1997) but led, in the post-absorptive period, to
higher subjective scores of satiety, which may reduce subsequent food
intake during the next meal (Achour et al., 1997). The lower subjective
scores for satiety and fullness after the RS (RS2; raw potato starch) than
after digestible starch (pregelatinized potato starch) were associated with
lower postprandial plasma concentration of glucose, insulin, GIP and GLP-1
(Raben et al., 1994).
Nilsson et al. (2008) observed the effect of an evening meal containing RS
(high-amylose barley kernel) or b-glucan (b-glucan-rich barley kernel) on
satiety after a subsequent standardized breakfast (ten hours after the evening
meal). Satiety was positively correlated to breath hydrogen, an indicator of
colonic fermentation.
Monsivais et al. (2010) recently showed that soluble fibre-dextrin (SFD)
enhanced the satiating power of beverages, compared to soluble corn fibre,
polydextrose and resistant starch. Indeed, only soluble fibre dextrin signifi-
cantly suppressed energy intake (p¼ 0.023). This SFD could be considered as
part of RS3. The SFD is probably highly fermentable, and its satiating effect
might also be mediated by SCFA produced within the four hours following the
fibre intake. Similar results on resistant dextrins were obtained by Nazare
et al. (2011).
Anderson et al. (2010) concluded from their recent study that estimates of
starch digestibility by the in vitro Englyst method were able to predict food
intake in young men 30 minutes and 120 minutes after consumption of a soup
containing 50 g maltodextrin, whole grain, high-amylose, regular cornstarch
or no added starch.
As shown by Tagliabue et al. (1995), RS (potato starch, in this study) seems
to have no thermogenic effect and its presence does not influence the size of
the thermic response to digestible starch. RS intake was, as expected followed
by lower oxidation and greater fat oxidation.
240 Resistant Starch

CH13GR 07/29/2013 16:49:36 Page 241
Lerer-Metzger et al. (1996) concluded that the replacement of 570 g wheat
starch/kg diet with mung-bean starch for five weeks resulted in a reduction in
plasma triacylglycerol concentration and adipocyte volume in both normal
and diabetic rats. Thus, the type of starch mixed into the diet may have
important metabolic consequences in normal and diabetic rats.
Robertson et al. (2005) showed that a supplementation with RS lead to
increased insulin sensitivity and a reduction in glycerol and free fatty acids
across subcutaneous adipose tissue. This was suggestive of a change in
adipocyte metabolism that may lead to a change in insulin sensitivity. RS
and inulin have been demonstrated to increase the release of gut hormones
with roles in appetite regulation and possibly, leptin release (Delzenne et al.,
2005; Keenan et al., 2006).
Keenan et al. (2006) and Zhou et al. (2006) have shown that dietary
resistant starch increased GLP-1 and PYY secretion. The same group (Zhou
et al., 2008) found that:
1. RS stimulated GLP-1 and PYY secretion in a substantial day-long manner,
independent of meal effect on changes in postprandial glycemia;
2. fermentation and the liberation of SCFAs in the lower gut are associated
with increased proglucagon and PYY gene expression;
3. glucose tolerance, an indicator of increased active forms of GLP-1 and
PYY was improved in RS-fed diabetic mice.
They concluded from their study that fermentation of RS is most likely the
primary mechanism for increased endogenous secretions of total GLP-1 and
PYY in rodents.
In an acute, randomizeddouble-blind, cross-over study comparing the effects
of four fibre and a low-fibre treatment on satiety, Willis et al. (2009) observed
that resistant starch (Novelose 330 and Hi-MaizeTM 260, National Starch;
RS3 and RS2, respectively) and corn bran had the most impact on satiety,
whereas polydextrose had little effect and behaved like the low-fibre treatment.
The same group (Willis et al., 2010) showed that satiety, gut hormone
(ghrelin, GLP-1, and PYY (3–36)) response, and food intake at subsequent
meals did not change in a dose-dependent manner after subjects consumed 0,
4, 8 and 12 g of mixed fibre in muffins for breakfast. However, despite lack of
differences in satiety, gut hormone levels differed among treatments. Ghrelin
was higher after the 12 g fibre dose than after the 4 g and 8 g fibre doses; GLP-
1 was higher after the 0 g fibre dose than after the 12 g and 4 g fibre doses; and
PYY(3–36) did not differ among fibre doses.
According to Shen et al. (2009), the mechanism of decreased body fat by
RS would be linked to increased neuropeptide POMC gene expression in the
Metabolic Effects of Resistant Starch 241

CH13GR 07/29/2013 16:49:36 Page 242
hypothalamus, and such an effect is independent of the involvement of
visceral afferent capsaicin-sensitive neurons.
So et al. (2007) investigated the impact of RS on body fat patterning and
central appetite regulation in mice. Mice receiving high RS (HRS; high-
amylose corn starch (60% RS)) diet during eight weeks had similar body
weights than those fed the low RS (LRS) diet, although their total body
adiposity, subcutaneous and visceral fat, intrahepatocellular lipids, plasma
leptin, plasma adiponectin and plasma insulin/glucose ratios were signifi-
cantly lower than the low RS group. Adipocytes isolated from the HRS group
were significantly smaller and had higher insulin-stimulated glucose uptake.
Manganese-enhanced magnetic resonance imaging (MEMRI) of the ventro-
medial and paraventromedial and paraventricular hypothalamic nuclei
suggested a satiating effect of the HRS diet despite a lower energy intake.
These data suggest that there may be appetite regulatory differences between
animals exposed to varying amount of RS, with the LRS animals producing
MEMRI data closer to fasted animals, and the HRS animal producingMEMRI
data closer to that of satiated animals.
So et al. (2007) suggest a link between short chain fatty acids production
from the fermentation of RS and the change in adipocyte morphology. Indeed,
the G protein receptor (GPCR43) on adipocytes may play a role in adipocyte
differentiation and proliferation; the ligands for these receptors appear to
be the short-chain fatty acids, acetate and propionate (Brown et al., 2003).
There is recent evidence that activation of GPCR43 by acetate and propionate
causes a decrease in leptin secretion and an increase in adipocyte differentia-
tion (Hong et al., 2005).
Significantly larger adipocyte size and over-insulin-stimulated glucose
oxidation in rats fed diets low in RS, compared with rats fed high RS diets,
had been described earlier by Kabir et al. (1998). So et al. (2007) also noticed
a significant difference in plasma leptin concentrations, with the low RS group
having significantly higher levels despite similar body weights. This would
confirm the observation made earlier by Skurk et al. (2007) that leptin levels
are driven by the lipid content and size of the adipocyte rather than by the
overall body weight. The concomitant increase in adiponectin and leptin in the
mice with low RS diet, compared to ‘high RS’ mice, is consistent with an
induction of leptin resistance in these mice (So et al., 2007).
13.6 CONCLUSION
Resistant starches are not homogeneous entities, as they can be totally or
partly fermented in the colon and their fermentability rate differs from one to
242 Resistant Starch

CH13GR 07/29/2013 16:49:36 Page 243
another. These various properties probably explain some of the contradictory
results described in the literature. Their fermentation produces a significant
amount of butyrate but, depending on the methodology used, butyrate or
propionate can be the second SCFA produced during fermentation of RS, after
acetate. Butyrate, being the nutrient of the colonocyte, is usually taken up
by the colonic mucosa, and only minor amounts can be found in the plasma
(or in urine).
When digestible starch is substituted in the diet by RS, glycemic load
and, consequently, insulin secretion is reduced. In the long term, RS
seems to be able to improve insulin response in hyperinsulinaemic
subjects. The impact of RS consumption at dinner on a subsequent
breakfast revealed the involvement of RS fermentation on improvement
of glucose tolerance and insulin sensitivity in healthy and insulin-
resistant subjects.
GLP-1 and PYY, two gut-secreted peptides which have been proposed as
potential anti-diabetes/obesity drugs, are also naturally secreted in response to
meal ingestion. RS has been reported to increase:
1. plasma total GLP-1 and total PYY in rodents; and
2. gene expression of PYY and proglucagon, probably through its
fermentation.
GLP-1 has been shown to increase in healthy subjects when part of the
digestible starch was substituted by RS. This impact of RS on gut peptides has
been confirmed within studies, showing an improved tolerance observed after
a standardized breakfast when RS (and fibre) was ingested during the
preceding dinner. It is associated to increased GLP-1 and adiponectin at
the time of the breakfast. These findings provide evidence for a link between
RS fermentation and the beneficial impact on insulin resistance. GLP-1
secretion would also explain the satiating effect of RS at several hours after
its ingestion.
Recent works have suggested a change in adipocyte metabolism that may
also lead to a change in insulin sensitivity. A link between short-chain fatty
acids production from the fermentation of RS and smaller adipocytes,
associated with better prognostic in metabolic syndrome, also supports the
beneficial metabolic impact of some RS. There is, however, still much work to
be performed in order to obtain new claims on several of the resistant starches.
Indeed, the effects will have to be demonstrated on each type of RS, unless a
clear correlation can be made between each fermentation pattern and corre-
sponding metabolic effects.
Metabolic Effects of Resistant Starch 243

CH13GR 07/29/2013 16:49:36 Page 244
REFERENCES
Achour, L., Flourie, B., Briet, F., Franchisseur, C., Bornet, F., Champ, M., Rambaud, J.C.,Messing, B. (1997). Metabolic effects of digestible and partially indigestible cornstarch:a study in the absorptive and postabsorptive periods in healthy humans. AmericanJournal of Clinical Nutrition 66(5), 1151–1159.
Anderson, G.H., Cho, C.E., Akhavan, T., Mollard, R.C., Luhovyy, B.L., Finocchiaro, E.T.(2010). Relation between estimates of cornstarch digestibility by the Englyst in vitromethod and glycemic response, subjective appetite, and short-term food intake in youngmen. American Journal of Clinical Nutrition 91(4), 932–9.
Asp, N.G. (1992). Resistant starch – Proceedings from the second plenary meeting ofEURESTA: European FLAIR Concerted Action No.11 on physiological implications ofthe consumption of resistant starch in man. Preface. European Journal of ClinicalNutrition 46, S1.
Bauer-Marinovic, M., Florian, S., M€uller-Schmehl, K., Glatt, H., Jacobasch, G. (2006).Dietary resistant starch type 3 prevents tumor induction by 1,2-dimethylhydrazine andalters proliferation, apoptosis and dedifferentiation in rat colon. Carcinogenesis 27(9),1849–59.
Behall, K.M., Howe, J.C. (1995). Contribution of fiber and resistant starch to metabolizableenergy. American Journal of Clinical Nutrition 62(5 Suppl), 1158S–1160S.
Behall, K.M., Scholfield, D.J., Yuhaniak, I., Canary, J. (1989). Diets containing highamylose vs. amylopectin starch: effects on metabolic variables in human subjects.American Journal of Clinical Nutrition 49(2), 337–44.
Bodinham, C.L., Frost, G.S., Robertson, M.D. (2010). Acute ingestion of resistant starchreduces food intake in healthy adults. British Journal of Nutrition 103(6), 917–22.
Brighenti, F., Benini, L., Del Rio, D., Casiraghi, C., Pellegrini, N., Scazzina, F., Jenkins,D.J., Vantini, I. (2006). Colonic fermentation of indigestible carbohydrates contributesto the second-meal effect. American Journal of Clinical Nutrition 83(4), 817–22.
Brown, A.J., Goldsworthy, S.M., Barnes, A.A., Eilert, M.M., Tcheang, L., Daniels, D.,Muir, A.I., Wigglesworth, M.J., Kinghorn, I., Fraser, N.J., Pike, N.B., Strum, J.C.,Steplewski, K.M., Murdock, P.R., Holder, J.C., Marshall, F.H., Szekeres, P.G.,Wilson, S., Ignar, D.M., Foord, S.M., Wise, A., Dowell, S.J. (2003). The Orphan Gprotein-coupled receptors GPR41 and GPR43 are activated by propionate and other shortchain carboxylic acids. Journal of Biological Chemistry 278(13), 11312–9.
Champ, M. (2004). Physiological aspects of resistant starch and in vivo measurements.Journal of AOAC International 87, 749–755.
Champ, M., Langkilde, A.M., Brouns, F., Kettlitz, B., Le Bail-Collet, Y. (2003). Advancesin dietary fibre characterisation II: consumption, chemistry, physiology and measure-ment of resistant starch: implications for health and food labelling. Nutrition ResearchReviews 16, 143–161.
Cheng, H.H., Lai, M.H. (2000). Fermentation of resistant rice starch produces propionatereducing serum and hepatic cholesterol in rats. Journal of Nutrition 130(8), 1991–5.
Clarke, J.M., Topping, D.L., Bird, A.R., Young, G.P., Cobiac, L. (2008). Effects of high-amylose maize starch and butyrylated high-amylose maize starch on azoxymethaneinduced intestinal cancer in rats. Carcinogenesis 29(11), 2190–4.
Cummings, J.H., Beatty, E.R., Kingman, S.M., Bingham, S.A., Englyst, H.N. (1996).Digestion and physiological properties of resistant starch in the human large bowel.British Journal of Nutrition 75(5), 733–47.
244 Resistant Starch

CH13GR 07/29/2013 16:49:36 Page 245
de Deckere, E.A, Kloots, W.J., van Amelsvoort, J.M. (1993). Resistant starch decreasesserum total cholesterol and triacylglycerol concentrations in rats. Journal of Nutrition123(12), 2142–51.
De Preter, V., Bulteel, V., Suenaert, P., Geboes, K.P., De Hertogh, G., Luypaerts, A.,Geboes, K., Verbeke, K., Rutgeerts, P. (2009). Pouchitis, similar to active ulcerativecolitis, is associated with impaired butyrate oxidation by intestinal mucosa. Inflamma-tory Bowel Diseases 15(3), 335–40.
de Roos, N., Heijnen, M.L., de Graaf, C., Woestenenk, G., Hobbel, E. (1995). Resistantstarch has little effect on appetite, food intake and insulin secretion of healthy youngmen. European Journal of Clinical Nutrition 49(7), 532–41.
Delzenne, N.M., Cani, P.D., Daubioul, C., Neyrinck, A.M. (2005). Impact of inulin andoligofructose on gastrointestinal peptides. British Journal of Nutrition 93(Suppl 1),S157–61.
Delzenne, N.M., Cani, P.D., Neyrinck, A.M. (2007). Modulation of glucagon-like peptide 1and energy metabolism by inulin and oligofructose: experimental data. Journal ofNutrition 137(11 Suppl), 2547S–2551S.
F€assler, C., Arrigoni, E., Venema, K., Hafner, V., Brouns, F., Amad�o, R. (2006).Digestibility of resistant starch containing preparations using two in vitro models.European Journal of Nutrition 45(8), 445–53.
Hamer, H.M., Jonkers, D.M., Vanhoutvin, S.A., Troost, F.J., Rijkers, G., de Bru€ıne, A.,Bast, A., Venema, K., Brummer, R.J. (2010). Effect of butyrate enemas on inflammationand antioxidant status in the colonic mucosa of patients with ulcerative colitis inremission. Clinical Nutrition 29(6), 738–44.
Heijnen, M.L., van Amelsvoort, J.M., Deurenberg, P., Beynen, A.C. (1996). Neither rawnor retrograded resistant starch lowers fasting serum cholesterol concentrations inhealthy normolipidemic subjects. American Journal of Clinical Nutrition 64(3),312–8.
Hong, Y.H., Nishimura, Y., Hishikawa, D., Tsuzuki, H., Miyahara, H., Gotoh, C., Choi, K.C., Feng, D.D., Chen, C., Lee, H.G., Katoh, K., Roh, S.G., Sasaki, S. (2005). Acetate andpropionate short chain fatty acids stimulate adipogenesis via GPCR43. Endocrinology146(12), 5092–9.
Howe, J.C., Rumpler, W.V., Behall, K.M. (1996). Dietary starch composition and level ofenergy intake alter nutrient oxidation in ‘carbohydrate-sensitive’ men. Journal ofNutrition 126(9), 2120–9.
Howlett, J.F., Betteridge, V.A., Champ, M., Craig, S.A., Meheust, A., Jones, J.M. (2010).The definition of dietary fiber – discussions at the Ninth Vahouny Fiber Symposium:building scientific agreement. Food & Nutrition Research 1; 54.
Hylla, S., Gostner, A., Dusel, G., Anger, H., Bartram, H.P., Christl, S.U., Kasper, H.,Scheppach, W. (1998). Effects of resistant starch on the colon in healthy volunteers:possible implications for cancer prevention. American Journal of Clinical Nutrition67(1), 136–42.
Jacobasch, G., Schmiedl, D., Kruschewski, M., Schmehl, K. (1999). Dietary resistantstarch and chronic inflammatory bowel diseases. International Journal of ColorectalDisease 14(4–5) 201–11.
Jenkins, D.J., Vuksan, V., Kendall, C.W., W€ursch, P., Jeffcoat, R., Waring, S., Mehling,C.C., Vidgen, E., Augustin, L.S., Wong, E. (1998). Physiological effects of resistantstarches on fecal bulk, short chain fatty acids, blood lipids and glycemic index. Journalof the American College of Nutrition 17(6), 609–16.
Metabolic Effects of Resistant Starch 245

CH13GR 07/29/2013 16:49:36 Page 246
Johnston, K.L., Thomas, E.L., Bell, J.D., Frost, G.S., Robertson, M.D. (2010). Resistantstarch improves insulin sensitivity in metabolic syndrome. Diabetic Medicine 27(4),391–397.
Kabir, M., Rizkalla, S.W., Champ, M., Luo, J., Boillot, J., Bruzzo, F., Slama, G. (1998).Dietary amylose-amylopectin starch content affects glucose and lipid metabolism inadipocytes of normal and diabetic rats. Journal of Nutrition 128(1), 35–43.
Keenan, M.J., Zhou, J., McCutcheon, K.L., Raggio, A.M., Bateman, H.G., Todd, E., Jones,C.K., Tulley, R.T., Melton, S., Martin, R.J., Hegsted, M. (2006). Effects of resistantstarch, a non-digestible fermentable fiber, on reducing body fat. Obesity (Silver Spring)14(9), 1523–1534.
Kendall, C.W.C., Esfahani, A., Sanders, L.M., Potter, S.M., Vidgen, E. (2010). The effectof a pre-load meal containing resistant starch on spontaneous food intake and glucoseand insulin responses. Journal of Food Technology 8(2), 67–73.
Kwak, J.H., Paik, J.K., Kim, H.I., Kim, O.Y., Shin, D.Y., Kim, H.J., Lee, J.H., Lee, J.H.(2012). Dietary treatment with rice containing resistant starch improves markers ofendothelial function with reduction of postprandial blood glucose and oxidative stress inpatients with prediabetes or newly diagnosed type 2 diabetes. Atherosclerosis (Epubahead of print: PubMed PMID: 22954674).
Langkilde, A.M., Champ, M., Andersson, H. (2002). Effects of high-resistant-starchbanana flour (RS(2)) on in vitro fermentation and the small-bowel excretion of energy,nutrients, and sterols: an ileostomy study. American Journal of Clinical Nutrition 75,104–111.
Le Leu, R.K., Brown, I.L., Hu, Y., Esterman, A., Young, G.P. (2007a). Suppression ofazoxymethane-induced colon cancer development in rats by dietary resistant starch.Cancer Biology & Therapy 6(10), 1621–1626.
Le Leu, R.K., Brown, I.L., Hu, Y., Morita, T., Esterman, A., Young, G.P. (2007b). Effect ofdietary resistant starch and protein on colonic fermentation and intestinal tumouri-genesis in rats. Carcinogenesis 28(2), 240–5.
Le Leu, R.K., Hu, Y., Brown, I.L., Young, G.P. (2009). Effect of high amylose maizestarches on colonic fermentation and apoptotic response to DNA-damage in the colon ofrats. Nutrition & Metabolism 6, 11.
Lerer-Metzger, M., Rizkalla, S.W., Luo, J., Champ, M., Kabir, M., Bruzzo, F., Bornet, F.,Slama, G. (1996). Effects of long-term low-glycaemic index starchy food on plasmaglucose and lipid concentrations and adipose tissue cellularity in normal and diabeticrats. British Journal of Nutrition 75(5), 723–32.
Liljeberg, H.G., Akerberg, A.K., Bj€orck, I.M. (1999). Effect of the glycemic index andcontent of indigestible carbohydrates of cereal-based breakfastmeals on glucose toleranceat lunch in healthy subjects. American Journal of Clinical Nutrition 69(4), 647–55.
Martin, L.J., Dumon, H.J., Lecannu, G., Champ, M.M. (2000). Potato and high-amylosemaize starches are not equivalent producers of butyrate for the colonic mucosa. BritishJournal of Nutrition 84(5), 689–96.
Math�e, D., Riottot, M., Rostaqui, N., Sacquet, E., Navarro, N., L�ecuyer, B., Lutton, C.(1993). Effect of amylomaize starch on plasma lipoproteins of lean and obese Zuckerrats. Journal of Clinical Biochemistry and Nutrition 14, 17–24.
Monsivais, P., Carter, B.E., Christiansen, M., Perrigue, M.M., Drewnowski, A. (2010).Soluble fiber dextrin enhances the satiating power of beverages. Appetite 56, 9–14.
Moreau, N.M., Martin, L.J., Toquet, C.S., Laboisse, C.L., Nguyen, P.G., Siliart, B.S.,Dumon, H.J., Champ, M.M. (2003). Restoration of the integrity of rat caeco-colonic
246 Resistant Starch

CH13GR 07/29/2013 16:49:37 Page 247
mucosa by resistant starch, but not by fructo-oligosaccharides, in dextran sulfatesodium-induced experimental colitis. British Journal of Nutrition 90(1), 75–85.
Murray, S.M., Flickinger, E.A., Patil, A.R., Merchen, N.R., Brent, J.L.Jr, Fahey, G.C.Jr.(2001). In vitro fermentation characteristics of native and processed cereal grains andpotato starch using ileal chyme from dogs. Journal of Animal Science 79(2), 435–44.
Nazare, J.A., Sauvinet, V., Normand, S., Gu�erin-Deremaux, L., Gabert, L., D�esage, M.,Wils, D, Laville, M. (2011). Impact of a resistant dextrin with a prolonged oxidationpattern on day-long ghrelin profile. Journal of the American College of Nutrition 30(1),63–72.
Nilsson, A.C., Ostman, E.M., Holst, J.J., Bj€orck, I.M. (2008). Including indigestiblecarbohydrates in the evening meal of healthy subjects improves glucose tolerance,lowers inflammatory markers, and increases satiety after a subsequent standardizedbreakfast. Journal of Nutrition 138(4), 732–9.
Noakes, M., Clifton, P.M., Nestel, P.J., Le Leu, R., McIntosh, G. (1996). Effect of high-amylose starch and oat bran on metabolic variables and bowel function in subjects withhypertriglyceridemia. American Journal of Clinical Nutrition 64(6), 944–51.
Nofrar�ıas, M., Mart�ınez-Puig, D., Pujols, J., Maj�o, N., P�erez, J.F. (2007). Long-term intakeof resistant starch improves colonic mucosal integrity and reduces gut apoptosis andblood immune cells. Nutrition (11–12) 861–70.
Perrin, P., Pierre, F., Patry, Y., Champ, M., Berreur, M., Pradal, G., Bornet, F., Meflah, K.,Menanteau, J. (2001). Only fibres promoting a stable butyrate producing colonicecosystem decrease the rate of aberrant crypt foci in rats. Gut 48(1), 53–61.
Phillips, J., Muir, J.G., Birkett, A., Lu, Z.X., Jones, G.P., O’Dea, K., Young, G.P. (1995).Effect of resistant starch on fecal bulk and fermentation-dependent events in humans.American Journal of Clinical Nutrition 62(1), 121–30.
Raben, A., Tagliabue, A., Christensen, N.J., Madsen, J., Holst, J.J., Astrup, A. (1994).Resistant starch: the effect on postprandial glycemia, hormonal response, and satiety.American Journal of Clinical Nutrition 60(4), 544–51.
Ranganathan, S., Champ, M., Pechard, C., Blanchard, P., Nguyen, M., Colonna, P.,Krempf, M. (1994). Comparative study of the acute effects of resistant starch anddietary fibers on metabolic indexes in men. American Journal of Clinical Nutrition59(4), 879–83.
Reiser, S., Powell, A.S., Scholfield, D.J., Panda, P., Ellwood, K.C., Canary, J.J. (1989a).Blood lipids, lipoproteins, apoproteins, and uric acid in men fed diets containing fructoseor high-amylose cornstarch. American Journal of Clinical Nutrition 49(5), 832–9.
Reiser, S., Powell, A.S., Scholfield, D.J., Panda, P., Fields, M., Canary, J.J. (1989b). Day-long glucose, insulin, and fructose responses of hyperinsulinemic and nonhyperinsu-linemic men adapted to diets containing either fructose or high-amylose cornstarch.American Journal of Clinical Nutrition 50(5), 1008–14.
Robertson, M.D., Bickerton, A.S., Dennis, A.L., Vidal, H., Frayn, K.N. (2005). Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adiposetissue metabolism. American Journal of Clinical Nutrition 82(3), 559–67.
Sacquet, E., Leprince, C., Riottot, M. (1983). Effect of amylomaize starch on cholesteroland bile acid metabolisms in germfree (axenic) and conventional (holoxenic) rats.Reproduction Nutrition Development 23(4), 783–92.
Shen, L., Keenan, M.J., Martin, R.J., Tulley, R.T., Raggio, A.M., McCutcheon, K.L., Zhou,J. (2009). Dietary resistant starch increases hypothalamic POMC expression in rats.Obesity (Silver Spring) 17(1), 40–5.
Metabolic Effects of Resistant Starch 247

CH13GR 07/29/2013 16:49:37 Page 248
Silvester, K.R., Englyst, H.N., Cummings, J.H. (1995). Ileal recovery of starch from wholediets containing resistant starch measured in vitro and fermentation of ileal effluent.American Journal of Clinical Nutrition Aug; 62(2), 403–11. Erratum in: AmericanJournal of Clinical Nutrition 1996 Mar; 63 (3) 407.
Skurk, T., Alberti-Huber, C., Herder, C., Hauner, H. (2007). Relationship betweenadipocyte size and adipokine expression and secretion. Journal of Clinical Endocrinol-ogy & Metabolism 92(3), 1023–33.
So, P.W., Yu, W.S., Kuo, Y.T., Wasserfall, C., Goldstone, A.P., Bell, J.D., Frost, G. (2007).Impact of resistant starch on body fat patterning and central appetite regulation. PLoSOne 2(12), e1309.
Stewart, M.L., Nikhanj, S.D., Timm, D.A., Thomas, W., Slavin, J.L. (2010). Evaluation ofthe effect of four fibers on laxation, gastrointestinal tolerance and serum markers inhealthy humans. Annals of Nutrition and Metabolism 56(2), 91–8.
Tachon, S., Zhou, J., Keenan, M., Martin, R., Marco, M.L. (2013). The intestinal micro-biota in aged mice is modulated by dietary resistant starch and correlated to improve-ments in host responses. FEMS Microbiology Ecology 83(2), 299–309.
Tagliabue, A., Raben, A., Heijnen, M.L., Deurenberg, P., Pasquali, E., Astrup, A. (1995).The effect of raw potato starch on energy expenditure and substrate oxidation. AmericanJournal of Clinical Nutrition 61(5), 1070–5.
van Munster, I.P., Tangerman, A., Nagengast, F.M. (1994). Effect of resistant starch oncolonic fermentation, bile acid metabolism, and mucosal proliferation. DigestiveDiseases and Sciences 39(4), 834–42.
Verbeke, K., Ferchaud-Roucher, V., Preston, T., Small, A.C., Henckaerts, L., Krempf, M.,Wang, H., Vonk, R.J., Priebe, M.G. (2010). Influence of the type of indigestiblecarbohydrate on plasma and urine short-chain fatty acids profiles in healthy humanvolunteers. European Journal of Clinical Nutrition 64(7), 678–84.
Vernia, P., Annese, V., Bresci, G., d’Albasio, G., D’Inc�a, R., Giaccari, S., Ingrosso, M.,Mansi, C., Riegler, G., Valpiani, D., Caprilli, R.;Gruppo Italiano per lo Studio del Colonand del Retto. (2003). Topical butyrate improves efficacy of 5-ASA in refractory distalulcerative colitis: results of a multicentre trial. European Journal of Clinical Investiga-tion 33(3), 244–8.
Wang, J.F., Zhu, Y.H., Li, D.F., Wang, Z., Jensen, B.B. (2004). In vitro fermentation ofvarious fiber and starch sources by pig fecal inocula. Journal of Animal Science 82(9),2615–22.
Weaver, G.A., Krause, J.A., Miller, T.L., Wolin, M.J. (1989). Constancy of glucose andstarch fermentations by two different human faecal microbial communities. Gut 30(1),19–25.
Weaver, G.A., Krause, J.A., Miller, T.L., Wolin, M.J. (1992). Cornstarch fermentationby the colonic microbial community yields more butyrate than does cabbage fiberfermentation; cornstarch fermentation rates correlate negatively with methanogenesis.American Journal of Clinical Nutrition 55(1), 70–7.
Willis, H.J., Eldridge, A.L., Beiseigel, J., Thomas, W., Slavin, J.L. (2009). Greater satietyresponse with resistant starch and corn bran in human subjects. Nutrition Research29(2), 100–5.
Willis, H.J., Thomas, W., Eldridge, A.L., Harkness, L., Green, H., Slavin, J.L.(2010). Increasing doses of fiber do not influence short-term satiety or foodintake and are inconsistently linked to gut hormone levels. Food & NutritionResearch 29, 54.
248 Resistant Starch

CH13GR 07/29/2013 16:49:37 Page 249
Wollowski, I., Rechkemmer, G., Pool-Zobel, B.L. (2001). Protective role of probioticsand prebiotics in colon cancer. American Journal of Clinical Nutrition 73(2 Suppl),451S–455S.
Wong, J.M., de Souza, R., Kendall, C.W., Emam, A., Jenkins, D.J. (2006). Colonic health:fermentation and short chain fatty acids. Journal of Clinical Gastroenterology 40(3),235–43.
Young, G.P., Gibson, P.R. (eds). (1995). Butyrate and the human cancer cell. Cambridge,UK: Cambridge University Press.
Zhou, J., Hegsted, M., McCutcheon, K.L., Keenan, M.J., Xi, X., Raggio, A.M., Martin, R.J.(2006). Peptide YY and proglucagon mRNA expression patterns and regulation in thegut. Obesity (Silver Spring) 14(4), 683–9.
Zhou, J., Martin, R.J., Tulley, R.T., Raggio, A.M., McCutcheon, K.L., Shen, L., Danna,S.C., Tripathy, S., Hegsted, M., Keenan, M.J. (2008). Dietary resistant starch upregulatestotal GLP-1 and PYY in a sustained day-long manner through fermentation in rodents.American Journal of Physiology. Endochrinology and Metabolism 295(5), E1160–6.
Metabolic Effects of Resistant Starch 249

CH13GR 07/29/2013 16:49:37 Page 250

3GCH14 07/29/2013 17:4:13 Page 251
14 The Microbiology of Resistant StarchFermentation in the Human LargeIntestine: A Host of UnansweredQuestions
Harry J. FlintMicrobial Ecology Group, Rowett Institute of Nutrition and Health,University of Aberdeen, Aberdeen, UK
14.1 INTRODUCTION
The human intestinal tract carries a large and diverse community of resident
microorganisms. The highest cell densities (1011 per gram) are found in the
large intestine, where flow rates are slowest and conditions in the gut lumen
become anaerobic. Some energy for microbial growth is derived from
endogenous sources through secreted proteins such as mucin and digestive
enzymes, and sloughed epithelial cells. In general, however, the main energy
sources are carbohydrates that have remained undigested in the stomach and
the small intestine (Macfarlane & Gibson, 1997).
Many plant oligosaccharides and cell wall polysaccharides are undegrad-
able, or poorly degraded, by mammalian digestive enzymes. Perhaps surpris-
ingly, however, for many diets it is a fraction of the potentially digestible
polysaccharide starch (‘resistant starch’) that escapes small intestinal diges-
tion which is estimated to provide the single largest energy source for
microbial growth in the colon (Cummings et al., 1996). There is increasing
evidence that the fermentation of resistant starch in the human large intestine
may lead to health benefits that include prevention of colorectal cancer and
improved insulin responses (Hylla et al., 1998; Bird et al., 2000, 2008;
Robertson et al., 2005).
Despite extensive early cultural studies on polysaccharide utilization by
human colonic bacteria (Salyers et al., 1977a, 1977b), our understanding of
the microbiology of resistant starch fermentation is still rather limited. One
251
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH14 07/29/2013 17:4:13 Page 252
problem is that it is not straightforward to reproduce, under in vitro conditions,
the state of the undigested starch particles that arrive in the large intestine
(Annison & Topping, 1994). Microbiological studies on pure cultures neces-
sarily tend to use autoclaved starch preparations, whose properties may differ
from those of starch in small intestinal effluent (Englyst &Macfarlane, 1986).
A second problem lies with the microbial community, which, in the colon,
is dominated by anaerobic bacteria that are not easily cultivated. Despite a
rapid increase recently in sequence-based diversity studies, relatively few
laboratories work routinely with anaerobes, making functional information
scarce. There are, however, excellent prospects now for combining molecular
and cultural approaches to obtain new insights into the impact of resistant
starches on microbial populations in the human colon.
14.2 IDENTIFYING THE MAJOR DEGRADERS OFRESISTANT STARCH IN THE HUMANGI TRACT
14.2.1 The human colonic microbiota
Direct sequence analysis of DNA and RNA from faecal and gut samples has
revealed a more complete picture of the diversity of bacteria that colonize the
human large intestine than was possible using culture-based approaches. Most
studies agree that only one-third of the bacterial diversity estimated from
16S rRNA gene sequencing is currently represented by named, cultured
species (Suau et al., 1999; Hold et al., 2002; Eckburg et al., 2005). On
the other hand, a much higher percentage of the numerically predominant
species are available as cultured isolates, so that more than half of amplified
ribosomal sequences detected in faecal samples can correspond to cultured
species (Walker et al., 2011).
The two most abundant bacterial phyla are the gram-positive Firmicutes
and gram-negative Bacteroidetes, followed by the Actinobacteria and
Proteobacteria, with small numbers of other phyla such as Verrucomicrobia.
The molecular diversity of eukaryotic gut microbes (protozoa and fungi) has
received comparatively little study to date.
14.2.2 Cultural studies
Extensive early bacterial isolation work using anaerobic techniques revealed a
large number of human gut bacteria that are capable of growth on starch.
Salyers et al. (1977a, 1977b) reported finding starch utilizers within the
Bacteroidetes, Firmicutes and Actinobacteria, with this activity being
252 Resistant Starch

3GCH14 07/29/2013 17:4:13 Page 253
particularly common among isolates of Bacteroides spp. (Table 14.1).
Subsequent selective isolations on media using starch as the sole energy
source produced a high proportion of Bifidobacterium spp. (Actinobacteria)
(Macfarlane & Englyst, 1986). The number of starch-degrading species from
these early studies among what is normally the most abundant phylum, the
Firmicutes, remained low, but it now appears that this may reflect the
relatively low coverage of this phylum previously by cultured strains (Eckburg
et al., 2005).
14.2.3 16S rRNA-based studies
While 16S rRNA-based community analysis reveals the diversity of human
colonic bacteria, it does not by itself provide any functional information. On
the other hand, when used in conjunction with selective functional screens, it
can potentially reveal uncultured starch-degrading species. Leitch et al.
(2007) used 16S rRNA sequencing to study the colonization of insoluble
resistant starch (Hylon VII) by mixed human faecal bacteria in an anaerobic
fermentor system, and 80% of starch-attached sequences recovered from four
different microbial communities corresponded to four species – Eubacterium
rectale, Ruminococcus bromii, Bifidobacterium adolescentis and Bifidobac-
terium breve (Figure 14.1).
This represents a very small subset of the diversity present in the starting
inocula, showing that the ability to bind to and colonize resistant starch is quite
limited among human colonic bacteria. The fact that the predominant attached
bacteria belong to cultured species suggests that the major colonizers may
already have been identified from the earlier isolation studies.
Another elegant functional approach is provided by stable isotope probing.
Kovatcheva-Datchary et al. (2009) incubated 13C labelled resistant starch
with mixed human faecal bacteria and recovered 13C labelled nucleic
acid, resulting from starch utilization, by density gradient centrifugation.
Table 14.1 Amylolytic activity in human gut anaerobes.
Growth with:
Strains tested amylose amylopectin
Bacteroides spp. 188 72% 73%High GþC Actinobacteria 52 31% 52%Low GþC Firmicutes 38 16% 53%
Data from Salyers et al. (1977a, 1977b).
The Microbiology of Resistant Starch Fermentation in the Human Large Intestine 253

3GCH14 07/29/2013 17:4:13 Page 254
16S rRNAanalysis again revealed that themajor utilizerswere bifidobacteria,
R. bromii andE. rectale. The last twoare low%GþCbacteria belonging to the
Firmicutes, emphasizing that members of this phylum also play an important
role in starch utilization. This is also apparent from analysis of the responses
among the faecal microbiota to added dietary starch in vivo, discussed
further below.
14.3 SYSTEMS FOR STARCH UTILIZATIONIN GUT BACTERIA
The availability of genome sequence data for an increasing number of starch-
utilizing bacteria opens up new prospects for analyzing the amylolytic gene
complement of starch-degrading species. Interpretation of these sequence
data, however, depends critically on the relatively small number of detailed
Figure 14.1 Colonization of insoluble substrates by human faecal bacteria. Mixedfaecal bacteria were introduced into a porous bag containing insoluble substrate(Hylon VII starch, wheat bran or porcine mucin) within an anaerobic continuous flowfermentor system. The experiment was repeated with inocula from four differenthealthy individuals. The predominant bacteria attached to each type of substratewere estimated by amplification and sequencing of 16S rRNA genes, as described byLeitch et al. (2007).
254 Resistant Starch

3GCH14 07/29/2013 17:4:15 Page 255
functional investigations that have been carried out in cultured gut bacteria
(Flint et al., 2008).
14.3.1 Bacteroides spp.
The starch utilization system of B. thetaiotaomicron is better understood than
that of any other human gut bacterium, thanks to the detailed studies of Salyers
et al. (reviewed Xu & Gordon, 2003; Flint et al., 2008). This gram-negative
species has a double cell membrane enclosing a periplasmic space. Soluble
starch molecules are proposed to be trapped by a complex of Sus proteins
located on the outer membrane surface. The molecules are then subject to
limited hydrolysis and translocation into the periplasmic space, where the
majority of the amylolytic activity is located. The major amylases are
described as neopullulanases, with the ability to hydrolyse a(1–4) linkagesin amylopectin and pullulan as well as in amylose.
14.3.2 Bifidobacterium spp.
Several species of bifidobacteria found in the human colon – mainly B. breve,
B adolescentis and B. pseudocatenulatum – show amylolytic activity, and
these include isolates with activity against high-amylose starches (Wang et al.,
1999; Ryan et al., 2006). A major cell surface amylase was identified in
B. breve that contains separate pullulanase and amylase catalytic domains,
together with a C terminal sortase cell wall anchoring signal (Figure 14.2).
The pullulanase domain was shown to debranch amylopectin by cleaving
Figure 14.2 Domain structures of the major cell surface amylases from two speciesof Gram-positive bacteria found in the human large intestine. GH13 – glycosidehydrolase family 13 catalytic domain; CBM – carbohydrate bindingmodule. Adaptedfrom Flint et al. (2012) Gut Microbes 3:4, 289-306. Fig. 3A.
The Microbiology of Resistant Starch Fermentation in the Human Large Intestine 255

3GCH14 07/29/2013 17:4:16 Page 256
a(1–6) linkages and acts in synergy with the a-amylase that cleaves a(1–4)linkages. Disruption of this gene in B. breve eliminated the ability to grow on
starch (Motherway et al., 2008).
14.3.3 Lachnospiraceae - Roseburia spp., Eubacteriumrectale and relatives
Bacteria related to Roseburia spp. and Eubacterium rectale represent an
important group of butyrate-producers in the human colon, accounting for
around 7–10% of bacteria in human faeces (Aminov et al., 2006). Most
isolates from this group can utilize starch as a growth substrate (Ramsay
et al., 2006). R. inulinivorans A2-194 produces a prominent amylase that is
bound to the cell surface via a sortase-mediated step. This multi-domain
enzyme includes putative starch binding modules, together with an a(1, 4)amylase (neopullulanase) catalytic domain that acts on pullulan and amylose.
The R. inulinivorans amylase, therefore, has many similarities to the enzyme
described from B. breve (Figure 14.2). These features appear consistent with
extracellular hydrolysis of starch molecules combined with the uptake and
processing of malto-oligosaccharides, as in B. breve.
14.3.4 Ruminococcaceae
This family of Firmicutes bacteria is numerically abundant in the human
colon, but is quite poorly represented by cultured strains. Ruminococcus
bromii has been regarded as a specialist starch-degrading species, but little if
any work appears to have been done on its amylase system. Recent work,
however, showed that the enzyme system of R. bromii was more effective
in degrading several particulate resistant starches than the amylases of
B. thetaiotaomicron, E. rectale or B. adolescentis (Ze et al., 2012). This
difference was most apparent when these starches were incubated without
heating, or after boiling for ten minutes, whereas autoclaving increased
degradability by the other species. Relatives of R. bromii were previously
shown to be preferentially associated with particulate material in human
faecal samples (Walker et al., 2008).
14.4 METAGENOMICS
Extraordinary recent advances in rapid sequencing capability are permitting
analysis of the gene complement of complex microbial communities (e.g.
Kurokawa et al., 2007). This allows the frequency of amylase genes, for
256 Resistant Starch

3GCH14 07/29/2013 17:4:16 Page 257
example, to be monitored in the community in response to dietary change.
This approach should be seen as complementing, rather than replacing, the
need for organism-based studies. First, the annotation of genomic data
depends crucially on functional information, which is still limited. Second,
as discussed above, the possession of an amylase gene does not define the role
of an organism in starch utilization, which is determined by the complete
enzyme complement and the organization of the relevant catalytic activities
and transport capabilities.
14.5 FACTORS INFLUENCING COMPETITION FORSTARCH AS A GROWTH SUBSTRATE
It is expected that the chemical and physical structure of resistant starch
entering the large intestine will influence the types of micro-organism capable
of exploiting it.While it seems likely that the different types of enzyme system
described above are equipped to deal with different forms of starch, there is as
yet little experimental evidence to support this notion. The failure to detect
Bacteroides among bacteria attached to resistant starch particles in vitro
(Leitch et al., 2007) perhaps suggests that Bacteroides are better able to utilize
solubilized than particulate starches, but this possibility needs more rigorous
testing. Several studies suggest that bifidobacteria are particularly active
against high-amylose starches (Wang et al., 1999) but, again, direct compari-
sons with amylolytic Firmicutes bacteria do not appear to have been made.
The gut environment can also play a role in determining the types of
bacteria that utilize a given form of starch, and therefore which bacterial
populations may be stimulated by the addition of resistant starch to the diet. A
good example is the influence of gut pH. Active microbial fermentation in the
proximal colon results in mildly acidic conditions, whereas pH is closer to
neutrality when fermentation is less active, e.g. in the distal colon.
In a fermentor system supplied continuously with soluble starch and other
polysaccharides at pH 5.5, the microbial community comprised approxi-
mately 25% of E. rectale-related butyrate-producing bacteria and 25%
Bacteroides relatives. On switching to pH 6.5, however, Bacteroides spp.
became dominant (80%) andE. rectale relatives became undetectable (Walker
et al., 2005; Duncan et al., 2009).
Since Bacteroides spp. appear to be less tolerant of growth inhibition by
weak acids at reduced pH than many gram-positive species, it appears that the
mildly acidic pH creates the opportunity for the gram-positive species to
compete successfully with Bacteroides for the polysaccharide substrates.
The consequence of these differences in pH tolerance, via their effects on
The Microbiology of Resistant Starch Fermentation in the Human Large Intestine 257

3GCH14 07/29/2013 17:4:16 Page 258
the balance of the microbial community, was a four-fold higher butyrate
concentration in the fermentor at pH 5.5, compared to that at pH 6.5 (Walker
et al., 2005).
14.6 METABOLITE CROSS-FEEDING
The anaerobic microbial communities of the rumen and large intestine are
characterized by high cell densities and by extensive interspecies cross-
feeding of fermentation products (Flint et al., 2007). Addition of resistant
starch to the diet could, therefore, lead potentially to the indirect stimulation of
many groups of bacteria and their metabolic products, in addition to the
primary amylolytic species. B. adolescentis L2-32, for example, degrades
starch to produce lactate, acetate and formate. E. hallii cannot degrade starch,
but it can convert acetate and lactate into butyrate. Co-cultures of these two
bacteria convert starch largely into butyrate, with growth of both B. adoles-
centis and E. hallii (Duncan et al., 2004; Belenguer et al., 2006). Cross-
feeding of partial degradation products from polysaccharide substrates is also
a well established phenomenon (Belenguer et al., 2006; Falony et al., 2006;
Figure 14.3).
Starch
Bifidobacterium spp.
Oligosaccharides
acetateL-lactateformate
Eubacterium hallii
12
fermentation (1 –
Roseburia hominisproducts
substrate 2 –“spillover”) butyrate
Figure 14.3 Potential contribution of metabolite cross-feeding to butyrate formationfrom resistant starches. Two distinct mechanisms are shown, one involving cross-feeding of fermentation products (lactate and acetate) and the other cross-feeding ofpartial breakdown products from starch. E. hallii is a lactate-utilizing bacterium thatproduces butyrate, while R. hominis is a non-lactate utilizing butyrate producer;neither species can grow by itself on resistant starch, but both grew when partneredwith a starch-degrading Bifidobacterium adolescentis strain. Based on Belengueret al. (2006).
258 Resistant Starch

3GCH14 07/29/2013 17:4:16 Page 259
14.7 IMPACT OF DIETARY RESISTANT STARCH UPONCOLONIC BACTERIA AND BACTERIALMETABOLITES IN HUMANS
The application of molecular approaches to bacterial enumeration in human
dietary intervention studies can reveal microbial responses to resistant starch
(RS) in vivo. Almost invariably, these responses have to be monitored in stool
samples. These can be expected to provide a good reflection of the microbiota
of the colonic lumen, but the temporal relationship between changes in the
proximal colon and changes seen in stool samples, of course, depends on
transit time.
Abell et al. (2008), using 16S rRNA based DGGE profiling, detected an
increase in relatives of R. bromii following increased RS intake. Another
recent study looked at the impact of diets high in NSP (wheat bran) or high in
type III resistant corn starch, each consumed for three weeks by 14 overweight
males, using a cross-over design (Walker et al., 2011). Bacterial groups were
monitored by qPCR, and for six subjects by 16S rRNA sequencing. This
analysis detected rapid increases in R. bromii-related bacteria and in the
Eubacterium rectale/Roseburia groups on the RS diet, although responses
varied markedly between individuals. Perhaps surprisingly, only one individ-
ual showed a strong response for bifidobacteria. A previous study failed to
detect a response in Eubacterium-related bacteria in humans consuming diets
high in RS (Schwiertz et al., 2002), but E. rectale was not monitored.
Physiological effects of RS intake are also evident from human studies, in
particular changes in faecal SCFA and improvements in insulin resistance
(Robertson et al., 2005). Evidence for protection against colorectal cancer
comes largely from animal studies and is proposed to occur mainly through
the butyrogenic effect of resistant starch (McIntyre et al., 1993). Among the
amylolytic groups known to be abundant in the human colon, only relatives of
E. rectale are butyrate-producers. As mentioned above, however, metabolite
cross-feeding might account for a butyrogenic effect of RS, even when the
dominant bacteria stimulated by starch are not butyrate-producers. Changes in
SCFA production rates have the potential to alter physiological functions
including appetite, inflammation and gut motility through signalling to gut
receptors (Brown et al., 2003).
The delivery of additional fermentable carbohydrates to the colon can have
numerous other consequences. Depression of colonic pH has been already
discussed above, and this is known to influence Ca2þ availability as well as
microbial competition. Another important effect is the increased incorpora-
tion of nitrogen into microbial cell protein, resulting from increased bacterial
The Microbiology of Resistant Starch Fermentation in the Human Large Intestine 259

3GCH14 07/29/2013 17:4:16 Page 260
growth and biomass, since this tends to decrease the concentration of
nitrogenous compounds available for catabolism to toxic or carcinogenic
products in the colon (Gill & Rowland, 2002).
14.8 CONCLUSIONS AND FUTURE PROSPECTS
Animal studies have shown that different types of RS can differ substantially
in their effects on gut metabolism and host physiology (Le Leu et al., 2009).
Such variation can be assumed also to occur in humans. At the same time,
there is evidence that RS fermentation may vary markedly between indi-
vidual humans (Walker et al., 2011). Host factors, the nature of the dietary
substrate and the composition of the gut microbiota are all likely to play a
role in determining the fermentability of resistant starch in the large
intestine (Figure 14.4).
The likely inter-dependence of these various factors is explored briefly
below.
1. RS structure will determine its rate and site of fermentation in the large
intestine. The structure of dietary starch (particle structure, association
with other polymers, crystallinity, branching, retrogradation, cross-linking)
Dietary starchDietary factors –
starch structure, particle size
cooking, storage
host amylases
digestion
(upper GI tract)
amylase inhibitors Host factors –
secretions
turnover
Resistant starch
gut environment
microbial amylases
Gut microbiota –
species
compositionfermentation
(large intestine)
Figure 14.4 Factors influencing the fate of starch in the intestine.
260 Resistant Starch

3GCH14 07/29/2013 17:4:16 Page 261
is known to determine its digestibility in the small intestine. It can be
assumed that these characteristics will also determine how rapidly resistant
starch is fermented in the colon, but less research has been done on this.
Rapid fermentation ofRSmay lead to complete fermentation in the proximal
colon, whereas slower rates will also support fermentation in more distal
regions, with the possibility of incomplete colonic fermentation overall.
2. Different types of RS may promote different groups of colonic bacteria. It
is possible that different forms of RS will prove to be accessible by
different groups of colonic microorganisms. This could result in selective
effects of starch intake upon the species composition of the colonic
microbiota, as well as differential effects on gut metabolism. For example,
it was suggested above that Bacteroides spp. may be best adapted to
utilizing soluble starch molecules. It has also been proposed that certain
types of RS might be more bifidogenic because of the high activity of this
group on high-amylose starches. If such selective effects occur, then
individuals who differ in the composition of their gut microbiota can
be expected to show different responses to RS intake as, indeed, is
suggested by the recent work of Martinez et al. (2010).
3. Host and dietary factors can influence what fraction of starch comprises
RS. Host factors (e.g. affecting secretion of digestive enzymes or transit
times) could play an important role in determining the undigested starch
fraction that arrives in the large intestine and its subsequent rate of
fermentation in the colon. This is also likely to be true for dietary factors
(e.g. the pattern of starch ingestion over time) and the presence of amylase
inhibitors of dietary origin (Wolin et al., 1999). Thus, it may be that the
types of starch that comprise RS will differ between individuals and also
within individuals over time, driven by changes in dietary intake.
4. Multiple factors (host genotype, diet, health, medication) have the poten-
tial to alter the amylolytic community within the large intestine. It seems
possible that a few species of amylolytic anaerobe play key roles in the
initial degradation of RS in the colon, especially when the RS exists in
particulate form. If so, variation in these ‘keystone’ species between
individuals could result in individual differences in fermentability of
RS (Ze et al., 2012). One possibility is that certain species might be
eliminated by antibiotic treatment during an individual’s lifetime and may
fail to re-establish, as has been proposed, for example, to explain variations
in oxalate degradation between humans (Duncan et al., 2002).
We have to conclude that some of the most fundamental questions about
the factors that influence starch fermentation in the human colon, and its
consequences for human health, remain unanswered. A combination of
The Microbiology of Resistant Starch Fermentation in the Human Large Intestine 261

3GCH14 07/29/2013 17:4:16 Page 262
cultural microbiology, microbial genomics and molecular ecology promises
progress in understanding microbial fermentation of starch by colonic micro-
organisms. This effort will clearly need to be allied, however, to an under-
standing of starch chemistry, and to physiological and nutritional studies both
in humans and in animal models.
ACKNOWLEDGEMENTS
The author would like to acknowledge support from the Scottish Government
Rural and Environment Research and Analysis Directorate.
REFERENCES
Abell, G.C.J., Cooke, C.M., Bennett, C.N., Conlon, M.A., McOrist, A.L. (2008). Phylo-types related to Ruminococcus bromii are abundant in the large bowel of humans andincrease in response to a diet high in resistant starch. FEMS Microbiology Ecology 66,505–515.
Aminov, R.I., Walker, A.W., Duncan, S.H., Harmsen, H.J.M., Welling, G.W., Flint, H.J.(2006). Molecular diversity, cultivation, and improved FISH detection of a dominantgroup of human gut bacteria related to Roseburia spp. or Eubacterium rectale. Appliedand Environmental Microbiology 72, 6371–6376.
Annison, G., Topping, D.L. (1994). Nutritional role of resistant starch: chemical structurevs. physiological function. Annual Review of Nutrition 14, 297–320.
Belenguer, A., Duncan, S.H., Calder, G., Holtrop, G., Louis, P., Lobley, G.E., Flint, H.J.(2006). Two routes of metabolic cross-feeding between bifidobacteria and butyrate-producing anaerobes from the human gut. Applied and Environmental Microbiology 72,3593–3599.
Bird, A.R., Brown, I.L., Topping, D.L. (2000). Starches, resistant starches, the gutmicroflora and human heath. Current Issues in Intestinal Microbiology 1, 25–37.
Bird, A.R., Vuaran, M.S., King, R.A., Noakes, M., Keogh, J., Morell, M.K., Topping, D.L.(2008). Wholegrain foods made from a novel high-amylose barley variety (Himalaya292) improve indices of bowel health in human subjects. British Journal of Nutrition 99,1032–1040.
Brown, A.J., Goldsworthy, S.M., Barnes, A.A., Eilert, M.M., Tcheang, L., Daniels, D.,Muir, A.I., Wigglesworth, M.J., Kinghorn, I., Fraser, N.J., Pike, N.B., Strum, J.C.,Steplewski, K.M., Murdock, P.R., Holder, J.C., Marshall, F.H., Szekeres, P.G., Wilson,S., Ignar, D.M., Foord, S.M., Wise, A., Dowell, S.J. (2003). The orphan G-coupledreceptors GPR41 and GPR43 are activated by propionate and other short chain fattyacids. Journal of Biological Chemistry 278, 1132–1139.
Cummings, J.H., Beatty, E.R., Kingman, S.M., Bungham, S.A., Englyst, H.N. (1996)Digestion and physiological properties of resistant starch in the human large bowel.British Journal of Nutrition 75, 73–747.
262 Resistant Starch

3GCH14 07/29/2013 17:4:16 Page 263
Duncan, S.H., Louis, P., Flint, H.J. (2004). Lactate-utilizing bacteria, isolated from humanfeces, that produce butyrate as a major fermentation product. Applied and Environ-mental Microbiology 70, 5810–5817.
Duncan, S.H., Louis, P., Thomson, J.M., Flint, H.J. (2009). The role of pH in determiningthe species composition of the human colonic microbiota. Environmental Microbiology11, 2112–2122.
Duncan, S.H., Richardson, A.J., Kaul, P., Holmes, R.P., Allison, M.J., Stewart, C.S. (2002).Oxalobacter formigenes and its potential role in human health. Applied and Environ-mental Microbiology 68, 3841–3847.
Eckburg, P.B., Bik, E.M., Bernstein, C.N., Purdom, E., Dethlefsen, L., Sargent, M., Gill,S.R., Nelson, K.E., Relman, D.A. (2005). Diversity of the human intestinal microbialflora. Science 308, 1635–1638.
Englyst, H.N., Macfarlane, G.T. (1986). Breakdown of resistant starch and readilydigestible starch by human gut bacteria. Journal of the Science of Food and Agriculture37, 699–706.
Falony, G., Viachou, A., Verbrugge, K., de Vuyst, L. (2006) Cross-feeding betweenBifidobacterium longum BB536 and acetate converting, butyrate-producing colonbacteria during growth on oligofructose. Applied and Environmental Microbiology72, 7835–7841.
Flint, H.J., Duncan, S.H., Scott, K.P., Louis, P. (2007). Interactions and competition withinthe microbial community of the human colon: links between diet and health. Environ-mental Microbiology 9, 1101–1111.
Flint, H.J., Bayer, E.A., Rincon, M.T., Lamed, R., White, B.A. (2008). Polysaccharideutilization by gut bacteria: potential for new insights from genomic analysis. NatureReviews Microbiology 6, 121–131.
Gill, C.I.R., Rowland, I.R. (2002). Diet and cancer: assessing the risk. British Journal ofNutrition 88, S73–S87 (Suppl 1).
Hold, G.L., Pryde, S.E., Russell, V.J., Furrie, E., Flint, H.J. (2002). Assessment ofmicrobial diversity in human colonic samples analysed by 16S rDNA sequence analysis.FEMS Microbiology Ecology 39, 33–39.
Hylla,S.,Gostner,A.,Dusel,G.,Anger,H.,Bartram,H-P.,Christl,S.U.,Kasper,H.,Scheppach,W. (1998). Effects of resistant starch on the colon in healthy volunteers: possibleimplications for cancer prevention. American Journal of Clinical Nutrition 67, 136–142.
Kovatcheva-Datchary, P., Egert, M., Maathuis, A., Rajilic-Stojanovic, M., de Graaf, A.A.,Smidt, H., de Vos, W.M., Venema, K. (2009). Linking phylogenetic identities of bacteriato starch fermentation in an in vitro model of the large intestine by RNA-based stableisotope probing. Environmental Microbiology 11, 914–926.
Kurokawa, K., Itoh, T., Kuwahara, T., Oshima, K., Toh, H., Toyoda, A., Takami, H., Morita,H., Sharma, V.K., Srivastava, T.P., Taylor, T.D., Noguchi, H., Mori, H., Ogura, Y.,Ehrlich, D.S., Itoh, K., Takagi, T., Sakaki, Y., Hayashi, T., Hattori, M. (2007).Comparative genomics revealed commonly enriched genes sets in human gut micro-biomes. DNA Research 14, 169–181.
Leitch, E.C.M., Walker, A.W., Duncan, S.H., Holtrop, G., Flint, H.J. (2007). Selectivecolonisation of insoluble substrates by human faecal bacteria. Environmental Micro-biology 9, 667–679.
Le Leu, R.K., Hu, Y., Brown, I.L., Young, G.P. (2009). Effect of high amylose maizestarches on colonic fermentation and apoptotic response to DNA-damage in the colon ofrats. Nutrition & Metabolism 6, 11–19.
The Microbiology of Resistant Starch Fermentation in the Human Large Intestine 263

3GCH14 07/29/2013 17:4:16 Page 264
Macfarlane, G.T., Englyst, H.N. (1986). Starch utilization by the human large intestinalmicroflora. Journal of Applied Bacteriology 60, 195–201.
Macfarlane, G.T., Gibson, G.R. (1997) Carbohydrate fermentation, energy transductionand gas metabolism in the human large intestine. In: Mackie, R.I., White, B.A. (eds).Gastrointestinal Microbiology Vol 1, 269–318. Chapman & Hall.
Martinez, I., Kim, J., Duffy, P.R., Schlegel, V.L., Walter, J. (2010). Resistant starchestypes 2 and 4 have differential effects on the composition of the fecal microbiota inhuman subjects. PLoS One 5,11 e15046.
McIntyre, A.P., Gibson, P., Young, G.P. (1993). Butyrate production from dietary fibre andprotection against large bowel cancer in a gut model. Gut 34, 386–391.
O’Connell Motherway, M., Fitzgerald, G.F., Neirynck, S., Ryan, S., Steidler, L., vanSinderen, D. (2008). Characterization of ApuB, an extracellular type II amylopullula-nase from Bifidobacterium breve UCC2003. Applied and Environmental Microbiology74, 6271–6279.
Ramsay, A.G., Scott, K.P., Martin, C.J., Rincon, M.T., Flint, H.J. (2006). Cell-associateda-amylases of butyrate-producing Firmicute bacteria from the human colon. Micro-biology 152, 3281–3290.
Robertson, M.D., Bickerton, A.S., Dennis, A.L., Vidal, H., Frayn, K.N. (2005). Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adiposetissue metabolism. American Journal of Clinical Nutrition 82, 559–567.
Ryan, S.M., Fitzgerald, G.F., van Sinderen, D. (2006). Screening for and identification ofstarch-, amylopectin-, and pullulan-degrading activities in Bifidobacterial strains.Applied and Environmental Microbiology 72, 5289–5296.
Salyers, A.A., Vercelotti, J.R., West, S.H.E., Wilkins, T.D. (1977a). Fermentation of mucinand plant polysaccharides by strains of Bacteroides from the human colon. Applied andEnvironmental Microbiology 33, 319–322.
Salyers, A.A., West, S.E.H., Vercellotti, J.R., Wilkins, T.D. (1977b). Fermentation ofmucins and plant polysaccharides by anaerobic bacteria from the human colon. Appliedand Environmental Microbiology 34, 529–533.
Schwiertz, A., Lehmann, U., Jacobasch, G., Blaut, M. (2002). Influence of resistant starchon the SCFA production and cell counts of butyrate-producing Eubacterium spp. in thehuman intestine. Journal of Applied Microbiology 93, 157–162.
Suau, A., Bonnet, R., Sutren, M., Godon, J.J., Gibson, G.R., Collins, M.D., Dor�e, J. (1999).Direct analysis of genes encoding 16S rRNA from complex communities reveals manynovel molecular species within the human gut. Applied and Environmental Micro-biology 24, 4799–4807.
Walker, A.W., Duncan, S.H., Leitch, E.C.M., Child, M.W., Flint, H.J. (2005). pH andpeptide supply can radically alter bacterial populations and short chain fatty acid ratioswithin human colonic microbial communities. Applied and Environmental Micro-biology 71, 3692–3700.
Walker, A.W., Duncan, S.H., Harmsen, H.J.M., Holtrop, G., Welling, G.W., Flint, H.J.(2008). The species composition of the human intestinal microbiota differs betweenparticle-associated and liquid phase communities. Environmental Microbiology 10,3275–3283.
Walker, A.W., Ince J, Duncan, S.H., Webster, L.M., Holtrop, G., Ze, X., Brown, D., Stares,M.D., Scott, P., Bergerat, A., Louis, P., McIntosh, F., Johnstone, A.M., Lobley, G.E.,Parkhill, J., Flint, H.J. (2011). Dominant and diet responsive groups of bacteria withinthe human colonic microbiota. The ISME Journal 5, 220–230.
264 Resistant Starch

3GCH14 07/29/2013 17:4:16 Page 265
Wang, X., Conway, P.L., Brown, I.L., Evans, A.J. (1999). In vitro utilization of amylopectinand high amylose maize (amylomaize) starch granules by human colonic bacteria.Applied and Environmental Microbiology 65, 4848–4854.
Wolin, M.J., Miller, T.J., Yerry, S., Zhang, Y.C., Bank, S., Weaver, G.A. (1999). Changes infermentation pattern of fecal microbial communities associated with a drug treatmentthat increases dietary starch in the human colon. Applied and Environmental Micro-biology 65, 2807–2812.
Xu, J., Gordon, J.I. (2003). Honor thy symbionts. Proceedings of The National Academy OfSciences Of The United States Of America 100, 10452–10459.
Ze, X., Duncan, S.H., Louis, P., Flint, H.J. (2012). Ruminococcus bromii is a keystonespecies for the degradation of resistant starch in the human colon. The ISME Journal 6,1535–1543.
The Microbiology of Resistant Starch Fermentation in the Human Large Intestine 265

3GCH14 07/29/2013 17:4:16 Page 266

3GCH15 07/20/2013 10:52:57 Page 267
15 Colon Health and Resistant Starch:Human Studies and Animal Models
Suzanne Hendrich, Diane F. Birt, Li Liand Yinsheng ZhaoInterdepartmental Graduate Program in Genetics, Department of FoodScience and Human Nutrition, Nutrition and Wellness Research Center,Iowa State University, USA
15.1 RS CLASSIFICATION
Resistant starches (RS) are named for their resistance to digestion in the small
intestine. This resistance allows them to pass into the large intestine and serve
as a substrate for microbial fermentation. RS have been categorized into four
types (Haralampu, 2000). Type 1 RS is physically inaccessible to digestive
enzymes such as starch from coarse grains. RS type 2 is ungelatinized starch,
including starch from potatoes or high-amylose corn starch, with long chains
that are uniformly packed (Jane, 2006). RS type 3 is starch that has been
cooked, then cooled, and is retrograded (Haralampu, 2000). It is the most
thermally stable RS and is exemplified by potato starch in cold potato salad.
RS4 is chemically modified starch that is, for example, cross-linked in a
manner to restrict swelling of the starch granules (Jane, 2006).
15.2 RS AND COLON HEALTH: OVERVIEW
Colon health involves maintenance of normal function of the colon and the
prevention of colon diseases. Healthy colon function permits regular bowel
movement on a daily basis or somewhat more frequently, consisting of
relatively soft but non-diarrhetic faeces. Colon health may be reflected in
the measurement of laxation (the frequency and total amount of faeces
excreted) and in the avoidance of constipation, which may be defined as
bowel movement frequency less than three times per week (Marks, 2009).
267
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.

3GCH15 07/20/2013 10:52:57 Page 268
There is little data to support what daily faecal weight should be considered to
be healthy, but one study of 53 Australians showed that the 16 subjects with
daily faecal mass of >150 g had higher faecal butyrate concentrations and
lower ammonia concentrations in faeces, which were thought to be protective
against colon cancer (Birkett et al., 1997).
Colon diseases include colon cancer, irritable bowel syndrome (IBS – also
referred to as spastic colon), inflammatory bowel diseases or colitis (e.g.
Crohn’s disease, ulcerative colitis), and diverticulosis/diverticulitis. The roles
of RS in preventing or protecting from each of these conditions are not yet
well understood, but seem to be most closely related to improved laxation and
increased short chain fatty acid (SCFA) production. The ability of RS to shift
gut microbial populations to increase probiotics (i.e. beneficial microbes)
and/or to decrease detrimental or pathogenic microbial species may also play
a role in disease protection. The biology and chemistry of interactions
between RS and the gut has been characterized to at least some extent in
varying models. This work provides a basis for further investigation of RS in
colon health and disease.
15.3 RS, GUT MICROBES ANDMICROBIALFERMENTATION
Several innovative model systems have enriched the understanding of how RS
affects gut microbial populations and their metabolic products. Germ-free
F344/N rats were orally dosed with pooled human faeces from three indi-
viduals from the UK or from Italy and were fed either a high sucrose diet or
15% RS (CrystaLeanTM) substituted for a portion of the sucrose for four
weeks (Silvi et al., 1999). Body weights of male rats fed RS were significantly
greater than for sucrose-fed males, whereas body weights of female rats fed
RS and bearing microbes from individuals from the UKwere significantly less
than in sucrose-fed females. Counts of Lactobacilli and Bifidobacteria were
increased, and Enterobacteria were decreased by RS feeding compared with
sucrose feeding. Staphylococci and Streptococci were increased by RS in rats
bearing microflora from humans from the UK, whereas Staphylococci were
decreased in rats fed RS and bearing Italian human microbes. Cecal b-gluco-sidase activity was increased and ammonia concentration was decreased by
RS. Cecal b-glucuronidasewas only decreased by RS in rats bearing microbes
from humans from the UK. Cecal butyrate was increased and propionate was
decreased by RS feeding (Silvi et al., 1999). Butyrate enhanced tight
junctions, improving the epithelial barrier function of Caco-2 cells, a widely
used model for human intestinal uptake and metabolism (Peng et al., 2009).
268 Resistant Starch

3GCH15 07/20/2013 10:52:57 Page 269
This barrier function is thought to be important for protection from colon
diseases.
As reviewed by Wollowski et al. (2001), the above changes in human
microflora-associated rats by RS to increase lactic acid bacteria (LAB;
Lactobacilli and Bifidobacteria) may help to detoxify some carcinogens, lower
faecal pH which is associated with colon cancer prevention, and stimulate
butyrate production, which improves protection of colonocytes from oxidative
stress, and maintains normal colon cell function. Note that LAB produce
lactate, which is then a substrate for butyrate-forming species (such as
Clostridia cluster XIVa microbes) (Collins et al., 1994), so the interactions
among gut bacteria needed for healthy colonic metabolism are complex.
Additional studies of RS feeding to female BALB/c mice showed that
increasing high-amylose corn starch intake from 0–30 (or 40% of the diet)
increased Bifidobacteria counts after 25 days (Wang et al., 2002). When
Bifidobacterium Lafti 8B was co-administered orally, the 40% high-amylose
starch diet also increased faecal butyrate concentrations by about fourfold
compared to control starch feeding (Wang et al., 2002), lending further
support to the concept that RS may benefit both gut bacterial composition
and microbial metabolism.
In vitro anaerobic incubations of resistant starches or their residues (starch
remaining after a-amylase treatment to simulate human starch digestion) from
various sources with human faeces also provide models for examining gut
microbes andmicrobialmetabolism.Pyrodextrin residues (heat-treated starches
creating RS type 4) from potato, lentil and cocoyam increased the propionate:
acetate ratio, but did not alter butyrate in human faecal incubations, compared
with residues from the non-resistant parent starches (Laurentin & Edwards,
2004).AnRS type 3 polymorphmade from thermal processing of high-amylose
cornstarch (HACS) increased butyrate production and Bifidobacteria counts in
continuous batch fermentations with human faecal inocula over 11 days,
compared with the control HACS (Lesmes et al., 2008). Thus, a human faecal
incubationmodel produces generally similar results to rodentmodels, regarding
the ability of RS to increase butyrate and Bifidobacteria, two factors thought to
benefit colon health. Proof of such benefits in human feeding studies remains to
be determined. A few relevant and recent studies regarding some aspects of
colon health are further described in the following.
15.3.1 RS and laxation
Because RS provides indigestible carbohydrates to the colon, RS should be
considered to be a subclass of dietary fibre. RS would therefore potentially
have health benefits similar to that of dietary fibres in general. Improved
Colon Health and Resistant Starch: Human Studies and Animal Models 269

3GCH15 07/20/2013 10:52:57 Page 270
laxation was observed in 14 subjects given 25 g RS (PROMITORTM) or 25 g
wheat bran fibre per day for 14 days in a randomized crossover design (Maki
et al., 2009). Both the RS and wheat bran increased faecal weight significantly
and did not differ from each other; both increased daily faecal weight above
150 g. This suggests that colon diseases associated with lesser laxation,
including diverticulosis/diverticulitis and IBS of the constipation type, may
be benefited by intake of RS. Colon cancer might also be more likely to be
prevented by RS intakes great enough to enhance laxation, based on the
findings of (Birkett et al., 1997).
15.3.2 RS, IBS and diverticulosis
No human studies have been performed on the role of RS in IBS to date, and
there are no clearly relevant animal models of this condition; neither has the
relation between RS intake and diverticulosis been studied. In an 18-month
study of nine groups of a total of 1800 Wistar rats given 0–17% wheat fibre,
incidence of diverticula was negatively associated with fibre intake and ranged
from 9% in rats fed the greatest fibre to 47% in rats fed no fibre. However, the
fibre intakes of the six treatment groups between 0–17% dietary fibre ranged
only from 0.5–4.2%, with incidence of diverticula ranging from 28–41%
(Fisher et al., 1985).
Estimated dietary fibre intakes in the US from the Continuing Survey of
Food Intakes of Individuals (1998), 10th–90th percentiles of intake ranged
from 7.1–24.8 g/d (Food&Nutrition Board, 2005) or�1.4–5.0% by weight of
diet (based on 500 g intake). This would suggest modest variance across
dietary fibre intakes in the US population in incidence of diverticula, which is
a common condition associated with aging. The significance of RS as a type of
dietary fibre in preventing diverticula in humans is probably scant.
15.3.3 RS and IBD
The ability of RS to prevent or reduce inflammatory bowel diseases (IBD –
e.g. ulcerative colitis) has been better studied and seems promising. In
Sprague-Dawley rats induced with colitis by 5% dextran sulphate sodium
in drinking water given for seven days, subsequent intake of 4% RS (type 3)
for seven days significantly improved colon histopathology scores compared
to controls, and also increased SCFA concentrations in cecal contents (Moreau
et al., 2003).
In rats with colitis induced by trinitrobenzene sulphonate (TNBS), fed
�6% RS for 14 days before and 21 days after TNBS, increased SCFA
concentrations were found in caecal and colon contents and healing was
270 Resistant Starch

3GCH15 07/20/2013 10:52:57 Page 271
accelerated compared with controls (Jacobasch et al., 1999). Pigs fed 17–24%
RS (raw potato starch, type 2 RS), compared with �5–7% RS from corn
starch, for 14 weeks, showed increased butyrate in colon contents and reduced
immune system reactivity in intestinal epithelia, including reduced numbers
of T-helper cells. This suggests that pigs fedmore RSwould be able to respond
to intestinal inflammation better (Nofrarias et al., 2007).
Twenty-two patients with quiescent ulcerative colitis (UC) consumed 60 g
oat bran (20 g dietary fibre) for 12 weeks. Faecal butyratewas increased at four
weeks, compared with baseline, but not at later time points in these subjects.
They did not show an increase in gastrointestinal symptoms during this time,
compared with ten control UC patients not ingesting oat bran, who showed
increased symptom scores according to a questionnaire (Hallert et al., 2003).
This study suggests that RS, which would share fermentation characteristics
with oat bran, might also be of some benefit in ulcerative colitis. However,
well-controlled trials of RS in colitis patients remain to be reported. Such
studies would be quite difficult to do; recruitment of sufficient subjects of
similar disease state, measuring compliance with test diets and monitoring of
treatments and symptoms over sufficient time periods would be challenging.
The needs of patients for standard treatments, depending on disease state,
might confound effects of RS.
15.3.4 RS and colon cancer risk – human studies
As with IBD, the role of RS in colon cancer risk has been studied to a very
limited extent. The following review of existing data generally does not
support the ability of increased RS intake to decrease colorectal cancer risk in
humans. This review updates similar findings (Young, 2004). Faecal samples
from 17 Native Africans (from South Africa), 17 African Americans and
18 Caucasian Americans showed that the Africans had greater faecal SCFA
concentrations than either group of Americans. This finding was attributed to
the diet of the Africans, assumed to consist largely of maize porridge, which
could contribute 30–60 g of RS per day (O’Keefe et al., 2009).
Subjects who had been treated previously for hereditary nonpolyposis
colon cancer (Lynch syndrome) were given 13 g RS/d (Novelose) for 29
months or a placebo (waxy maize starch, composed entirely of amylopectin).
The incidence of colon neoplasms after this time period did not differ between
the two treatments; 18.7% of 358 subjects given RS vs. 18.4% of 369 subjects
given the placebo (Burn et al., 2008).
Twenty-four pre-operative colorectal cancer patients given the same RS as
in the Burn et al. study showed a lesser proportion of mitotic cells in the upper
half of colon crypts than did ten similar subjects given amylopectin placebos
Colon Health and Resistant Starch: Human Studies and Animal Models 271

3GCH15 07/20/2013 10:52:57 Page 272
after ingesting the starches for four weeks (Dronamraju et al., 2009). The
reduction in this marker of premalignancy indicated that RS may protect
against colon cancer. In sporadic adenoma patients, 19 g of RS consumed
daily for two months did not alter epithelial cell proliferation in colorectal
biopsies (bromodeoxyuridine labelling index) compared with controls
ingesting placebo starch (n¼ 28 per treatment; van Gorkom et al., 2002).
After colon adenomectomy, 23 patients consumed 45 g of highly digestible
maltodextrin or digestion resistant maltodextrin (28 g/d of RS type 2) for four
weeks. Faecal bile acid concentrations decreased significantly in subjects
given RS, but SCFAs and colorectal cell proliferation did not differ between
the treatments (Grubben et al., 2001).
To date, few studies have been done on RS and colon cancer risk in humans.
Two studies suggest possible antineoplastic effects of RS, in suppressing
colon cell proliferation (Dronamraju et al., 2009) and lessening faecal bile
acids (Grubben et al., 2001) which may be colon tumour promoters (Nagen-
gast et al., 1995). Three studies (van Gorkom et al., 2002; Grubben et al.,
2001; Burn et al., 2008) did not show differences in colon cell proliferation
between RS and placebo treatments in colon cancer patients. Such patients are
at greater risk of colon cancer than the general population and are considered
good populations to attempt dietary interventions. Because cancers develop
over the lifespan and more often emerge later in life, long-term dietary
intervention trials of RS in general populations or in individuals with genetic
traits predisposing to colon cancer might be considered, if animal models are
sufficiently supportive of a protective role for RS.
15.4 COLON CANCER PREVENTION – ANIMALMODELS
There has been considerable interest in the ability of RS to reduce the yield
of colon preneoplastic lesions and colon cancers in rodent models. In most
cases, RS type 1 or 2 has been fed in diets preceding and/or following treatment
with a chemical carcinogen. In one study, Sprague-Dawley rats were fed diets
containing 0%, 10% or 20% raw high-amylose corn starch for four weeks prior
to treatment with azoxymethane (AOM), and colon cancers were assessed 25
weeks after AOM treatment. Both doses of starch inhibited colon tumour
development and increased SCFA including butyrate in the distal colon (Le Leu
et al., 2007).
In a series of studies, the ability of RS to counteract DNA damage in the
colon that was induced by high protein intakes was assessed. A high-amylose
butrylated starch was compared with high-amylose starch in preventing the
272 Resistant Starch

3GCH15 07/20/2013 10:52:57 Page 273
DNA damage caused by a high protein diet, as measured by the comet assay in
rats. Both diets were effective in protecting against DNA damage by the high
protein diet, but the diet with butrylated starch was the most effective. This
protection paralleled increasing butyrate in the large intestine (Bajka et al.,
2008). Further, feeding 20% high-amylose corn starch for four weeks with
cooked beef or chicken prevented the genetic damage caused by these meats
(Toden et al., 2007).
Although much of the research has focused on RS type 1 or 2, one study
was conducted with a hydrothermally processed type 3 resistant starch
(Novelose 330) fed for 20 weeks following treatment of Sprague-Dawley
rats with 1,2-dimethylhydrazine. The RS completely prevented the develop-
ment of tumours, compared to rats fed control starch. The RS-3 starch also
increased apoptosis and decreased proliferation in the colon (Bauer-Mari-
novic et al., 2006).
Since not all studies have observed reduced tumour yields in rodents fed
RS, a study was designed to assess the timing of feeding RS relative to AOM
treatment in Wistar rats. Raw potato starch was fed for a three-week interval,
either before or after AOM treatment, and the number of aberrant crypt foci
(ACF) was increased in the rats fed RS before AOM, in comparison with those
fed control diet, while the yield of ACF was suppressed in a dose-dependent
manner in the rats fed RS following AOM treatment (Liu & Xu, 2008). These
authors concluded that RS fed following carcinogen treatment was more
likely to inhibit the colon lesions induced by AOM than RS fed preceding
carcinogen treatment.
SCFA, including butyrate, acetate and propionate, have been implicated as
mediators of the resistant starch impact on colon carcinogenesis (Topping &
Clifton, 2001). These short-chain fatty acids are produced by microbes that
ferment the resistant starch that reaches the large intestine. The hypothesis
generally implicates production of butyrate, because of observations that
butyrate promotes differentiation of colonocytes, but data in support of this
hypothesis have been mixed. For example, studies comparing cellulose and
high-amylose resistant starch revealed the greatest increase in caecal butyrate
with the resistant starch, but the greatest inhibition of colon tumours with
cellulose (Sakamoto et al., 1996). Furthermore, Nakanishi et al. (2003) treated
rats with Clostridium butyricum, and with or without high-amylose starch,
during treatments with AOM. Treatment with C. butyricum alone increased
butyrate in the caecum, but it did not decrease aberrant crypt foci. However,
treatment with both C. butyricum and high-amylose starch decreased aberrant
crypt foci and increased acetate and propionate concentrations in the caecum.
Elevatedb-glucuronidase activitywas also observed in the caecal contents fromthe rats treated with both C. butyricum and high-amylose starch.
Colon Health and Resistant Starch: Human Studies and Animal Models 273

3GCH15 07/20/2013 10:52:57 Page 274
Studies were conducted with AOM-treated F344 rats, and the key end-
points were ACF and mucin-depleted foci (MDF). Secondary endpoints were
caecal pH, weight and SCFA concentrations (Zhao et al., 2011). ACF are
morphologically altered crypts, alone or in cluster, identified by microscopic
examination of methylene blue-stained whole mount preparations of colonic
mucosa from carcinogen-treated rodents (Bird, 1995). They are characterized
by being at least two to three times larger than normal crypts, having increased
pericryptal space and a slit-like opening, with thicker epithelium that stains
darker with methylene blue.
In recent years, a subset of AOM-induced ACF were believed to be better
predictors of cancer; these are MDF that are observed in alcian blue-stained
tissue which stains mucins. MDF are identified by the absence of the deep blue
staining indicative of mucins. These lesions highly correlated to tumour
development in AOM-treated rats (Caderni et al., 2003). More recently, these
lesions were observed in humans (Femia et al., 2008). AOM is selective for the
colon, and only a few small bowel/stomach tumours are noted. Carcinomas are
generally found approximately 30 weeks after carcinogen administration, and
tumour incidence and multiplicity can be accurately titrated (Pories et al.,
1993). AOMproduces both adenomas and carcinomas, with a predominance of
lesions induced in the distal colon, consistent with human sporadic cancers.
A high-amylose starch with exceptionally high resistance to digestion
was prepared by complexing high-amylose starch with lipids in a manner
that was expected to make it less digestible and, thus, to pass into the large
intestine and modify the gut microbial populations (Hasjim et al., 2010).
Studies evaluated the impact on pre-cancer lesions in the colon of rats by
including this resistant starch, fed in comparison with raw high-amylose
corn starch or normal corn starch, or cooked with high moisture as would be
used in a pudding, in comparison with similarly cooked high-amylose corn
starch or normal corn starch. The rats were fed a normal corn starch, a high-
amylose starch or the processed high-amylose starch after treatment with the
carcinogen AOM (Zhao et al., 2011).
This study showed that the high-amylose starch and the processed high-
amylose starch reduced pH and increased caecum weight, irrespective of
whether they were cooked by a high-moisture method. The raw high-amylose
starch reduced ACF, but ACF were not reduced by the processed starch in
comparison with raw normal starch or raw high-amylose starch (Zhao et al.,
2011). When all three starches were cooked by boiling in water, the processed
starch reduced MDF the most, high-amylose starch had an intermediate
reduction and the most MDF were found in the rats fed the normal starch.
Parallel observations were made with ACF, the more abundant precancerous
lesion induced by AOM (Zhao et al., 2011). This research supports the ability
274 Resistant Starch

3GCH15 07/20/2013 10:52:57 Page 275
of the processed high-amylose starch to prevent the development of colon
cancer in this rat model.
15.5 CONCLUSIONS
Research in humans and laboratory animals suggest potential health benefits
of dietary RS in protection against diseases of the colon, including irritable
bowel syndrome, inflammatory bowel diseases (colitis) and colon cancer. To
date, the research suggests that the type and cooking of RS, the control starch
used, the time of feeding relative to the disease process and the nature of the
disease may all impact the ability of RS to benefit colon health. Although the
research has shown considerable promise for RS, further research is needed at
all levels to provide a firm foundation for the use of RS in the maintenance of
human health.
REFERENCES
Bajka, B.H., Clarke, J.M., Cobiac, L., Topping, D.L. (2008). Butyrylated starch protectscolonocyte DNA against dietary protein-induced damage in rats. Carcinogenesis 29(11),2169–2174.
Bauer-Marinovic, M., M€uller-Schmehl. K., Glatt, H., Jacobasch, G. (2006). Dietaryresistant starch type 3 prevents tumor induction by 1,2-dimethylhydrazine and altersproliferation, apoptosis and dedifferentiation in rat colon. Carcinogenesis 27(9), 1849–1859.
Bird, R.P. (1995). Role of aberrant crypt foci in understanding the pathogenesis of coloncancer. Cancer Letters 93, 55–71.
Birkett, A.M., Jones, G.P., de Silva, A.M., Young, G.P., Muir, J.G. (1997). Dietary intakeand faecal excretion of carbohydrate by Australians: importance of achieving stoolweights greater than 150 g to improve faecal markers relevant to colon cancer risk.European Journal of Clinical Nutrition 51(9), 625–632.
Burn, J., Bishop, D.T., Mecklin, J.P., Macrae, F., M€oslein, G., Olschwang, S., Bisgaard, M.L., Ramesar, R., Eccles, D., Maher, E.R., Bertario, L., Jarvinen, H.J., Lindblom, A.,Evans, D.G., Lubinski, J., Morrison, P.J., Ho, J.W., Vasen, H.F., Side, L., Thomas, H.J.,Scott, R.J., Dunlop, M., Barker, G., Elliott, F., Jass, J.R., Fodde, R., Lynch, H.T.,Mathers, J.C. (2008). Effect of Aspirin or Resistant Starch on Colorectal Neoplasia in theLynch Syndrome. New England Journal of Medicine 359(24), 2567–2578.
Caderni, G., Femia, A.P., Giannini, A., Favuzza, A., Luceri, C., Salvadori, M., Dolara, P.(2003). Identification of Mucin-depleted Foci in the Unsectioned Colon of Azoxy-methane-treated Rats: Correlation with Carcinogenesis. Cancer Research 63, 2388–2392.
Collins, M.D., Lawson, P.A., Willems, A., Cordoba, J.J., Fernandez-Garayzabal, J., Garcia,P., Cai, J., Hippe, H., Farrow, J.A. (1994). The phylogeny Of The genus Clostridium –
Colon Health and Resistant Starch: Human Studies and Animal Models 275

3GCH15 07/20/2013 10:52:57 Page 276
Proposal of 5 New Genera and 11 New Species Combinations. International Journal ofSystematic Bacteriology 44(4), 812–826.
Dronamraju, S.S., Coxhead, J.M., Kelly, S.B., Burn, J., Mathers, J.C. (2009). Cell kineticsand gene expression changes in colorectal cancer patients given resistant starch: arandomised controlled trial. Gut 58(3), 413–420.
Femia, A.P., Giannini, A., Fazi, M., Tarquini, E., Salvadori, M., Roncucci, L., Tonelli, F.,Dolara, P., Caderni, G. (2008). Identification of Mucin Depleted Foci in the HumanColon. Cancer Prevention Research 1(7), 562–567.
Fisher, N., Berry, C.S., Fearn, T., Gregory, J.A., Hardy, J. (1985) Cereal Dietary FiberConsumption and Divertcular Disease: a Lifespan Study in Rats. American Journal ofClinical Nutrition 42(5), 788–804.
Food and Nutrition Board (2005). Dietary Reference Intakes for Energy, Carbohydrate,Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids (Macronutrients)National Academies Press, Washington, D.C., pp. 1226–43.
Grubben, M., Van den Braak, C.C.M., Essenberg, M., Olthof, M., Tangerman, A., Katan,M.B., Nagengast, F.M. (2001). Effect of resistant starch on potential biomarkers forcolonic cancer risk in patients with colonic adenomas – a controlled trial. DigestiveDiseases and Sciences 46(4), 750–756.
Hallert, C., Bjorck, I., Nyman, M., Pousette, A., Gr€ann€o, C., Svensson, H. (2003).Increasing fecal butyrate in ulcerative colitis patients by diet: Controlled pilot study.Inflammatory Bowel Diseases 9(2), 116–121.
Haralampu, S.G. (2000). Resistant starch – a review of the physical properties andbiological impact of RS3. Carbohydrate Polymers 41(3), 285–292.
Hasjim, J., Lee, S.-O., Hendrich, S., Setiawan, S., Ai, Y., Jane, J-L. (2010). Characteriza-tion of a Novel Resistant-Starch and Its Effects on Postprandial Plasma-Glucose andInsulin Responses. Cereal Chemistry 87(4), 257–262.
Jacobasch, G., Schmiedl, D., Kruschewski, M., Schmehl, K. (1999). Dietary resistantstarch and chronic inflammatory bowel diseases. International Journal of ColorectalDisease 14(4–5), 201–211.
Jane, J.L. (2006). Current understanding on starch granule structure. Journal of AppliedGlycoscience 53, 151–159.
Laurentin, A., Edwards, C.A. (2004). Differential fermentation of glucose-based carbohy-drates in vitro by human faecal bacteria – A study of pyrodextrinised starches fromdifferent sources. European Journal of Nutrition 43(3), 183–189.
Le Leu, R.K., Brown, I.L., Hu, Y., Esterman, A., Young, G.P. (2007). Suppression ofAzoxymethane-Induced Colon Cancer Development in Rats by Dietary ResistantStarch. Cancer Biology and Therapy 6(10), 1621–1626.
Lesmes, U., Beards, E.J., Gibson, G.R., Tuohy, K.M., Shimoni, E. (2008). Effects of resistantstarch type III polymorphs onhumancolonmicrobiota and short chain fatty acids in humangut models. Journal of Agricultural and Food Chemistry 56(13), 5415–5421.
Liu, R.P., Xu, G.F. (2008). Effects of resistant starch on colonic preneoplastic aberrantcrypt foci in rats. Food and Chemical Toxicology 46(8), 2672–2679.
Marks, J.W. (2009). http://www.medicinenet.com/constipation/article.htmMaki, K.C., Sanders, L.M., Reeves, M.S., Kaden, V.N., Rains, T.M., Cartwright, Y. (2009).
Beneficial effects of resistant starch on laxation in healthy adults. International Journalof Food Sciences and Nutrition 60, 296–305.
Moreau, N.M., Martin, L.J., Toquet, C.S., Laboisse, C.L., Nguyen, P.G., Siliart, B.S.,Dumon, H.J., Champ, M.M. (2003). Restoration of the integrity of rat caeco-colonic
276 Resistant Starch

3GCH15 07/20/2013 10:52:58 Page 277
mucosa by resistant starch, but not by fructo-oligosaccharides, in dextran sulfatesodium-induced experimental colitis. British Journal of Nutrition 90(1), 75–85.
Nagengast, F. M., M. Grubben, et al. (1995). Role of Bile-Acids in Colorectal Carcino-genesis. European Journal of Cancer 31A(7–8), 1067–1070.
Nakanishi, S., Kataoka, K., van Munster, I.P. (2003). Effects of high amylose maize starchand Clostridium butyricum on microbiota and formation metabolism in colonic ofazoxymethane-induced aberrant crypt foci in the rat colon. Microbiology and Immu-nology 47(12), 951–958.
Nofrarias, M., Martinez-Puig, D., Pujols, J., Maj�o, N., P�erez, J.F. (2007). Long-term intakeof resistant starch improves colonic mucosal integrity and reduces gut apoptosis andblood immune cells. Nutrition 23, 861–870.
O’Keefe, S.J.D.,Ou, J.H.,Aufreiter, S.,O’Connor,D., Sharma, S., Sepulveda, J., Fukuwatari,T., Shibata, K., Mawhinney, T. (2009). Products of the colonic microbiota mediate theeffects of diet on colon cancer risk. Journal of Nutrition 139(11), 2044–2048.
Peng,L.Y.,Li,Z.R.,Green,R.S.,Holzman,I.R.,Lin,J.(2009).ButyrateEnhancestheIntestinalBarrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated ProteinKinase in Caco-2 Cell Monolayers. Journal of Nutrition 139(9), 1619–1625.
Pories, S.E., Ramchurren, N., Summerhayes, I., Steele, G. (1993). Animal models for coloncarcinogenesis. Archives of Surgery 128, 647–653.
Sakamoto, J., Nakaji, S., Sugawara, K., Iwane, S., Munakata, A. (1996). Comparison ofresistant starch with cellulose diet on 1,2-dimethylhydrazine-induced colonic carcino-genesis in rats. Gastroenterology 110, 116–120.
Silvi, S., Rumney, C.J., Cresci, A., Rowland, I.R. (1999). Resistant starch modifies gutmicroflora and microbial metabolism in human flora-associated rats inoculated withfaeces from Italian and UK donors. Journal of Applied Microbiology 86(3), 521–530.
Toden, S., Bird, A.R., Topping, D.L., Conlon, M.A. (2007). High red meat diets inducegreater numbers of colonic DNA double-strand breaks than white meat in rats:attenuation by high-amylose maize starch. Carcinogenesis 28(11), 2355–2362.
Topping, D.L., Clifton, P.M. (2001). Short-chain fatty acids and human colonic function:Roles of resistant starch and nonstarch polysaccharides. Physiological Reviews 81(3),1031–1064.
van Gorkom, B.A.P., Karrenbeld, A., van der Sluis, T., Zwart, N., van der Meer, R., deVries, E.G.E., Kleibeuker, J.H. (2002). Calcium or resistant starch does not affectcolonic epithelial cell proliferation throughout the colon in adenoma patients: Arandomized controlled trial. Nutrition and Cancer – an International Journal 43(1),31–38.
Wang, X., Brown, I.L., Khaled, D., Mahoney, M.C., Evans, A.J., Conway, P.L. (2002).Manipulation of colonic bacteria and volatile fatty acid production by dietary highamylose maize (amylomaize) starch granules. Journal of Applied Microbiology 93(3),390–397.
Wollowski, I., Rechkemmer, G., Pool-Zobel, B.L. (2001). Protective role of probiotics andprebiotics in colon cancer. American Journal of Clinical Nutrition 73(2), 451S–455S.
Young, G.P. (2004). Resistant starch and colorectal neoplasia. Journal of AOAC Interna-tional 87(3), 775–786.
Zhao, Y., Hasjim, J., Li, L., Jane, J.-L., Hendrich, S., Birt, D.F. (2011). Inhibition ofAzoxymethane-Induced Preneoplastic Lesions in the Rat Colon by a Cooked StearicAcid Complexed High-Amylose Cornstarch. Journal of Agricultural and Food Chem-istry 59(17), 9700–9708.
Colon Health and Resistant Starch: Human Studies and Animal Models 277

3GCH15 07/20/2013 10:52:58 Page 278

BINDEX 07/22/2013 7:47:26 Page 279
Index
Note: Page numbers in italic refer to figures and tables.
acetate 231, 232, 234, 240, 242,
273
ActistarTM 102
acylated starches 55–6, 63–4
adipose fat 241–2, 243
ae starch see amylose-extender
AGPase 5, 6
All-Bran1 201
amylases
a-amylase 4, 96–7, 115–16
b-amylase 3, 4
isoamylase 83
natural inhibitors 120
resistance to 47–50, 103–5
see also enzymatic hydrolysis
amylopectin 1–2, 102
amylose/amylopectin ratios 3–4
helices 98
retrogradation and digestion
rate 118–19
types and granule structure 24–6
and waxy starch 8–9
amylopectin-lipid complex 79, 85–6
amylose
allomorphs/structure 1, 102
amylopectin/amylose ratios 3–4
helices 98
starch biosynthesis and 5, 8, 11–12,
14
see also high-amylose starch
amylose-extender starch
flour replacement product 168
role of HAM gene 36–7
structures 26–7
amylose-lipid complex (RS5) 79–80,
91–2, 122–3
advantages of 80
enzyme resistance and 80–1
complex crystallinity 82
granular starch properties 82
lipid structure 81
formation of starch-lipid
complex 83
debranching and RS content
85–6
fatty-acid choice and RS
content 83–5
health benefits of 87–91
products and applications 86–7
AOAC methodology 133–6, 137, 138,
146, 147
appetite and satiety 211, 215
appetite regulation 215–16, 239–40,
241, 242
Resistant Starch: Sources, Applications and Health Benefits, First Edition.
Edited by Yong-Cheng Shi and Clodualdo C. Maningat.
� 2013 John Wiley & Sons, Ltd. Published 2013 by John Wiley & Sons, Ltd.
279

BINDEX 07/22/2013 7:47:26 Page 280
appetite and satiety (Continued)
anorexigenic and orexigenic
hormones 216
role of hypothalamus 215–16
measuring effects of RS intake
human data 216–17, 221–5,
239–40
rodent data 218–21
proposed mechanisms for RS
effects 217–18, 229
satiety, satiation and fat
deposition 239–42
applications see food and beverage
applications
assay see measurement
bacteria see fermentation; microbiota
Bacteroidetes bacteria 252–3, 255,
257–8, 261
baking see biscuit baking
barley mutants 6, 12
batters 67
beta-cell function 199–202
and glucose toxicity 199–200
and serum free fatty acids 200
Bifidobacterium spp. 253, 255, 258,
269
biosynthesis, starch
overview of 4–6, 13–14
starch branching enzymes 11–12
starch debranching enzymes 13
starch synthases 6–11
biscuit baking 167
ae flour replacement starch
(RS3) 168–72
cookies 172–5
crackers 175–8
extruded ready-to-eat cereals
178–89
Brittle-2 mutant 6
bowel diseases see colon health
branching enzymes 11–12
breads 65, 66, 86
butyrate 230–1, 233, 234, 243, 258
and colon health studies 27, 268–9,
272
butyrylated starch 63–4
cancer, colon 89–91, 231, 268, 271–2
animal models and prevention
272–5
cereal starches
compared to tuber starches 3, 48
mutants 6, 7, 10–12
see also high-amylose starch
chemically modified (RS4) starch 46,
103, 105, 132, 145
amylase resistance 47–9, 103–4
mechanism 49–50
cross-linked starches 50–4
physicochemical properties 57–9
physiological responses to 60–4
food grade starches 57–8, 105
performance in food products
65–8
health benefits 60–5
pyrodextrinized starches 56–7
physicochemical properties 59
physiological responses to 64–5
substituted starches 54–6
physiological responses to 63–5
chloroplasts 4–5
cholesterol 237–8
citrates, starch 53–4, 59–60
colon health studies 267–8
cancer 268, 271–2
animal studies 272–5
inflammatory bowel diseases 268,
270–1
irritable bowel syndrome 268, 270
related to microbial
fermentation 268–9
colonic fermentation/microbiota see
fermentation; microbiota
constipation 267–8, 269–70
cookies 66
baking 172–5
cooking 4, 26
280 Index

BINDEX 07/22/2013 7:47:26 Page 281
high-amylose maize (RS2)
starches 27
in presence of lipids 80–1
see also biscuit baking
corn see maize
crackers see biscuit baking
cross-linked (RS4) starches 50–4, 105,
145–6
in vitro enzymatic testing see wheat
starch
physicochemical properties 57–9
physiological responses to 60–4
crystalline starch 28–9, 99
debranching enzymes 5, 13, 83
dextrins, resistant 56–7, 59, 104, 105,
240
in vitro testing of wheat starch
148–9, 151–2, 153
diabetes 43, 95, 208
carbohydrate quality/quantity and
risk 193–5
complications and prevalence
192–3
definition and types 191–2
insulin sensitivity and
secretion 195–6, 209, 210,
235–6
glycemic impact of
carbohydrate 197–8
low-GI foods and improved beta-cell
function 199–200
see also glycemic response
diet, healthy 208
dietary fat see fat
dietary fibre 43, 44, 45, 46, 57–8, 105
definitions of 13, 136, 137, 138
measurement of 131, 138–42
digestion, starch 2–3, 23–4, 96–7,
115
chronology of indigestibility
studies 44–7
digestibility of
biosynthetic mutants 7
cross-linked wheat starch 149,
160–2
factors affecting 3–4
slowly versus readily digestible
starch 113–15
see also enzymatic hydrolysis
diverticulosis 268, 270
enzymatic hydrolysis 2, 3, 24, 96–7,
115–16
amylose-lipid complex (RS5) 79,
80–1
factors affecting 97–100
high-amylose starches 27
inhibitors of 120
resistance to
and molecular structure 102–5
and physical structure 97–102
enzymatic testing see measurement
Eubacterium spp. 256
extruded cereal 178–89
fat 208
deposition 241–2, 243
lipid metabolism 236–8
fatty acids see short-chain fatty acids
fermentation, microbiotic 209–10,
212, 217–18, 220, 221, 260
metabolic studies reviewed 230–5,
239–40
colon health 268–9
microbiology of see microbiota
fiber/fibre see dietary fibre
Fibersol 105
Fibersym1 179, 183–9
Firmicutes bacteria 252–3, 254, 257
flour replacement (RS3) starch 168
food and beverage applications
amylose-lipid complex (RS5) 86–7
high-amylose (RS3) starch 168–72
cookies 172–5
crackers 175–9
extruded cereal 179–89
modified (RS4) starches 65–8
Index 281

BINDEX 07/22/2013 7:47:26 Page 282
food grade starches 57–8, 105
food viscosity 120
gelatinization 4, 26
cross-linked wheat starch 150,
152–3
high-amylose starch 29, 30, 35,
168
and slowly digestible starch 118–19
see also retrograded starch
GI concept 111–12
GLP-1 (glucagon-like peptide or
polypeptide) 201–2, 238–9,
243
role in appetite 216–25, 241
glucan polymers 1–2
and starch biosynthesis 5
glucose 2, 112–13
slowly available concept 116
toxicity/tolerance 199–200, 235–6
see also glycemic response
a-glucosidases 96–7, 115–16
natural inhibitors 120
glutarates, starch 54, 60
glycemic index (GI) 111–12, 113,
194–5, 197–9
low-GI foods and beta-cell
function 199–200
glycemic load (GL) 194–5, 235, 243
glycemic response 2–3, 60–1, 112
amylose-lipid complex (RS5) 87–9
low GI carbohydrates 197–9
natural inhibitors and food
viscosity 120
regulation by resistant starch
208–11, 212, 235–6
slowly versus readily digestible
starch 113–15
granule see starch granule
HAM gene 36–7
health benefits 2–3, 43, 60–5
amylose-lipid complex (RS5)
87–91
healthy diet 208
see also colon health
helices, starch 98
Hi-MaizeTM 179, 183–9, 238
high-amylose maize (RS2) starch
11–12, 14, 23–7, 101
amylose-extender mutant
role of HAM gene 36–7
structures 26–7
elongated starch granules 26–7
formation in the amyloplasts
33–5
structures 27, 31–3, 34
resistant starch
formation and kernel
development 29–31
location in the starch granule
35–6
semi-crystalline amylose/IC
structures 28–9
use in amylose-lipid complexes 83,
84, 85
use in (RS3) flour replacer 168
high-amylose modifier (HAM)
gene 36–7
hunger see appetite
hydroxypropylation 54–5
hypoglycemia see glycemic control
hypothalamus 215, 220, 242
insulin
sensitivity and resistance to 195–7,
209, 210, 235–6, 241
see also diabetes; glycemic
response
irritable bowel syndrome 268,
270
isoamylase 83
Lachnospiraceae bacteria 256
laxation 267–8, 269–70
lipid complexes see amylose-lipid
complex
lipid metabolism 236–8
282 Index

BINDEX 07/22/2013 7:47:26 Page 283
maize starch 12–13, 23
mutant starches 6, 7, 10, 11–12, 14,
23
structure 26–7
see also high-amylose maize (RS2)
starch
maltase-glucoamylase 96–7, 115–16
maltodextrins 64, 65, 105
measurement of resistant starch
131–3
AOAC official method 133–6,
146
enzymatic of wheat starch see wheat
starch
in vitro and in vivo compared 146–8
integrated procedure for total dietary
fibre 136–42
metabolic studies, review of
fermentation and colonic
metabolism 230–5, 239,
259–62
GIP, GLP-1 and PYY
secretion 238–9
glycemic response and glucose
tolerance 235–6
lipid metabolism 236–8
satiety and fat deposition 239–42
see also appetite and satiety
metagenomics 256–7
microbiota, gut 60, 113, 229, 239,
251–2
colon health studies 268–9
common microbes 252–3
impact of RS 259–62
microbiological studies 260–2
16S rRNA-based studies 253–4
cultural studies 252–3
metagenomics and 256–7
starch as a growth substrate 257–8
metabolite cross-feeding 258
starch utilization systems 254–5
Bacteroides spp. 255
Bifidobacterium spp. 255–6
Lachnospiraceae 256
Ruminococcaceae 256
see also fermentation
modified starch see chemically modified
molecular structure, starch 102–3, 104
mutant starches 6, 7, 10, 11–12, 14, 23
see also high-amylose maize (RS2)
starch
Nutriose1 105
obesity 43, 95, 193, 207
fat deposition studies 241–2
pasta 66–7, 67–8
peptide YY see PYY
phosphated distarch phosphate 57
phosphorylated starches 47, 48,
50–3
in vitro testing see wheat starch
physicochemical properties 57–9
physiological responses to 60–3
photosynthesis 4–5
products see food and beverage
applications
PromitorTM 105, 235, 240
propionate 231, 233, 234, 242, 243,
273
pullulanase 83
pyrodextrinized (RS4) starches 56–7,
59
physicochemical properties 59–60
physiological responses to 64, 269
PYY (peptide YY) 216, 238–9, 243
role in appetite 217–24, 241
readily digestible starch (RDS) 2–3
glycemic response compared to
SDS 113–15
resistant starch (RS), overview of
definitions 2–3, 44, 229
factors affecting enzyme
resistance 97–105
history of indigestibility studies
44–7
Index 283

BINDEX 07/22/2013 7:47:26 Page 284
resistant starch (RS), overview of
(Continued)
measurement of see measurement
physiological effects and benefits
2–3, 60–5, 207, 259–62
types and classification see RS1/2/3/
4/5
retrograded starch 3, 44, 45, 47, 80,
101, 132
Roseburia spp. 256
RS1/2/3/4/5 starches
described 3, 44, 45, 46, 47, 80,
100–2, 167–8
RS2 starch see high-amylose maize
(RS2) starch
RS3 starch 101–2, 167–72
RS4 starch see chemically modified
RS5 starch see amylose-lipid
complex
Ruminococcaceae bacteria 256
satiety see appetite and satiety
SDS see slowly digestible starch
second-meal effect 121, 210, 235
short-chain fatty acids 60, 62, 63,
209–10
and amylose-lipid complex 84
and beta-cell function 200
and colon health 268, 272, 273
fermentation in the gut 209–10,
212, 217–18, 220, 221
studies reviewed 230–5, 242
Shrunken-2 mutant 6
slowly digestible starch (SDS) 2–3,
111–13, 132
basic concept of SDS 111–13,
116–17
digestion of starch 115–16
glycemic response compared to
RDS 113–15
physiological/structural
factors 116–17
food matrix structures 117–18
food transit time 121
starch structures 118–19
viscosity and enzyme
inhibitors 120–1
strategies to make SDS 121–3
sodium trimetaphosphate 50, 51–2
starch, general overview of
classification/types 2–3
composition and structure 1–2,
23–6, 27, 96, 97–8
crystalline structures 99
granules see granule structure
molecular structure 102–3
starch helices 98
see also resistant starch
starch citrates 53–4, 59–60
products 67–8
starch digestion see digestion
starch glutarates 54, 60
starch granule structures 25, 26, 27,
99–100
cross-linked wheat starch 153–60
elongated high-amylose 27, 31–3,
34
and lipids 82
and resistant starch location 35–6
starch synthases 5, 6–8
synthase-I 8, 9
synthase-II 9–10
synthase-III 10–11
synthase-IV 11
starch types 2–3
ae corn starch III RS (X150) 168
see also RS1/2/3/4/5
structures, starch see starch; starch
granules
substituted (RS4) starches 54–6, 56–7
sucrase-isomaltase 96, 97, 115
sugary-1 mutants 13, 23
synthases see starch synthases
tortillas 66
total dietary fibre, assay of 136–42
transit time 115, 121
triacylglycerols (triglycerides) 373–4
284 Index

BINDEX 07/22/2013 7:47:26 Page 285
tuber starches 3, 48
Type 2 diabetes see diabetes
types of starch see starch types
viscosity, food 120
wafers 67
waxy starches 8–9, 54–5, 56, 80, 86
wheat starch (cross-linked), in vitro
testing of 145–8
materials and methods 148–50
results and discussion 151–63
Index 285





























