Embed Size (px)
Citation preview

Journal of Chemical Ecology, Vol. 23, No. 9. 1997
RESPONSE AND ADAPTATION TO ANALOGS OFDISPARLURE BY SPECIALIST ANTENNAL RECEPTOR
NEURONS OF GYPSY MOTH, Lymantria dispar
JOSEPH C. DICKENS,1 •* JAMES E. OLIVER,2
and VICTOR C. MASTRO3
1USDA, AKS, Vegetable Laboratory2USDA, ARS, Insect Chemical Ecology Laboratory
Beltsville, Maryland 20705
3USDA, APHIS. Otis Methods Development LaboratoryBldg. 1398. Otis ANGB. Massachusetts 02542
(Received November 18, 1996; accepted May 15, 1997)
Abstract—Reception and adaptation of antennal receptors In response to (•*•)•disparlure |(f)-D). the sex attractant pheromone of the gypsy moth. Lyman-tria dispar (Lepidoptera: Lymantriidae)). Its enamiomer, (-)-D. and analogsof disparlure in which the epoxide group Is replaced with various three-membered rings ((tKyclopropyl (2HC). (± difluorocyclopropyl (2FC),(±)-dichlorocyclopropyI (2CIC). andY±)-azindinyl (AZ). ( + )-AZ, and (-)-AZ] as well as a mixture of 1,1-epoxy analogs were Investigated using elec-troantennograms. Antennal receptors of male moths were more responsive(5 Mg stimulus load) to (-t-)-D than to (-)-D: while responses to each of theanalogs were significantly, less and did not differ from each other. Antennalreceptors of female moths did not respond significantly to any of the com-pounds at this stimulus load. Dose-response curves revealed greater sensitivityof male antennae to (+)-D than (- )-D. Among the cyclopropane analogs and1,1-epoxy mixture (1.1E), the order of sensitivity was: 2FC > 2C1C > 2HC> 1.1E. Antennae were least sensitive to the aziridinyl analogs; however,(+ )-AZ was more active than (- )-AZ, while an intermediate response waselicited by the racemate. Males exposed to ( + )-D adapted to (+)-D. whileresponses to ( + )-D were not significantly affected by exposure to (-)-D, 2PCor ( + )-AZ. After exposure to hexylacetate, males showed adaptation to (•*•)-D or (-)-D. but not to the other odorants. Preliminary field data correlatewell with knowledge obtained in our electrophysiological studies and dem-onstrate the usefulness of such studies. These results indicate that novelapproaches to the design and synthesis of potential agonists of disparlure
•To whom correspondence should be addressed.
2197
OW8-0)JI'97'0900-2I9'»IJ JO'O <C 199' Plenum Publishing Corporation

2198 DICKENS, OLIVER, AND MASTRO
receptors are necessary before substant ia l effects on the ac t iv i ty of disparlurearc realized.
Key Words—Analogs, semiochemicals, adaptation, receptor, neurophysiol-ogy, olfaction gypsy moth, Lymantria dispar.
INTRODUCTION
Since introduction of the gypsy moth, Lymantria dispar, into the United Statesin 1869 and its attainment of pest status (Plimmer et a l . , 1982), research on itand other potential lymantriid pests has intensified. Characterization of thefemale-produced sex attractant of the gypsy moth, disparlure (cis-7,8-epoxy-2-mcthyl-octadecane), by Bierl et al. (1970) provided a tool not only for use inpopulation surveys but also for disruption of orientation of males to females(Schwalbe et al., 1983). Subsequently, the ( + )-enantiomer of disparlure[(?/?,85)-cpoxy-2-methyl-octadecanc = (-t-)-disparlure = ( + )-D| was shownto be attractive, while its antipode, ( —)-D, was somewhat inhibitory (Vitc etal . , 1976).
Elucidation of the chemical structure of disparlure provided the frameworkfor synthesis and testing of analogs as possible agonists, antagonists, or syner-gists of pheromone responses. Field testing of a number of synthetic epoxidesrelated to disparlure clearly showed that movement of the epoxide or methylgroup resulted in substantial loss of attractiveness of the resulting molecule (Bierlet al., 1972; Adler et al . , 1972; Sarmiento et al . , 1972; Schneider et al., 1974).Replacement of the epoxide group by another group or groups resulted in somecompounds that were attractive to gypsy moths in the f i e l d , but none was asattractive as disparlure (Sheads et al., 1975).
We report results of electrophysiological studies of antennal receptorresponses of L. dispar to disparlure and analogs of disparlure in which theoxygen of the epoxide group of disparlure is replaced by carbon [cyclopropane(= CP) analogs] or nitrogen (aziridine (= AZ) analogs]. A mixture of 1 ,1-epoxy analogs previously shown to be active in field test (Sheads et a l . , 1975)was also tested. Adaptation experiments were done to determine effects ofselected analogs on disparlure receptors. A green leaf volatile, hexyl acetate,was also tested in the adaptation studies because it enhances behavioral responsesof moths to sex pheromones (Dickens et a l . , 1993a; Light et a l . , 1993). Thesestudies provide the bases for field studies and may facilitate design of otherpotential agonists of disparlure receptors.
METHODS AND MATERIALS
Insects. Pupae of Lymantria dispar were obtained from a laboratory colonymaintained at the USDA, ARS, Insect Biocontrol Laboratory, Beltsville Agri-

DISPARLURE ANALOGS 2199
cultural Research Center, Beltsville, Maryland. Pupae were sexed and placedinto containers in an incubator at 26°C and a photoperiod of 16L:8D untilemergence. Electrophysiological recordings were made from antennae of adultswithin one to three days after emergence.
Odorant stimuli. Odorant stimuli used in experimental studies are repre-sented in Figure 1. The enantiomers of disparlure [( + )-D, (7R,8S)-epoxy-2-methyl-octadecane 1, and (-)-D, (7S,8R)-epoxy-2-methyI-octadecane 2| weregreater than 98% optical purity (Oliver and Waters, 1995). Other compoundsstudied included racemic cyclopropane (2HC), dichlorocyclopropane (2C1C),and difluorocyclopropane (2FC) analogs 5, 4, and 3; the enantiomeric aziridineanalogs [( + )-AZ and (-)-AZJ 6 and 7 as well as the racemic mixture (AZ);and the mixture of 1,1-distributed epoxides 8 and 9 (1, 1E). Hexyl acetate (99%)used as the standard was obtained from Aldrich Chemical Co., Milwaukee,Wisconsin.
FIG. 1. Structures of compounds used in electrophysiological and behavioral experi-ments: 1; (+)-disparlure [(7R,8S)-epoxy-2-methyl-octadecane], the principal sex attrac-tant of the gypsy moth, 2; (—)-disparlure, an inhibitor of behavioral responses to ( + )-disparlure, 3; difluorocyclopropyl; 4, dichlorocyclopropyl; and 5, cyclopropyl, are dis-parlure analogs; 6, (+)-aziridinyl and 7, (-)-aziridinyl analogs; 8 + 9 mixture of 1,1-epoxide analogs. Stimulatory effectiveness of the compounds is (1 > 2 » 3 > 4 >5, 8 + 9 > 6 > 7).

220(1 DICKENS, OLIVER, AND MASTRO
Chemical Syntheses. Gas chromatography (GC) was performed on Shi-madzu instruments equipped with flame ionization detectors and either a 60-mSPB 608 or a 30-m DB-1 capillary column. Compositions of mixtures are asreported by Shimadzu CR501 Chromatopac recording integrators and are uncor-rected for response or other factors. Mass spectra (MS) were recorded with aFinnigan Incos 50 GC-MS with a source block temperature of 150°C.
( + )-l-Dccyl-2-(2-methylhexyl)-cyclopropane 5 was prepared from2-methyl-7-octadccene 90.4% (Z) and 9.6% (E) and diiodomethane/tetra-ethylaluminum according to the procedure of Maruoka et al. (1985) and waspurified by flash chromatography on silver nitrate-impregnated silica gel withhexane. The product after chromatography contained (by GC) 88.6% of 5, 9.7%of the trans isomer, and 1.7% of (E)-2-methyl-7-octadcccnc. The mass spec-trum of 5 showed ions (abundance) at m/z, (%): 280, M+ (1 ) , 252 (1 ) , 125(6), 111 (19), 98 (10), 97 (39), 85 (16), 84 (24), 83 (53), 82 (15), 81 (10), 71(29), 70 (42), 69 (80), 68 (15), 67 (20), 57 (83), 56 (95), 55 (75), 54 (16), 43(100), 42 (14), and 41 (76).
For ( + )-2-decyl-3-(2-methylhexyl)-l ,1-dichlorocyclopropane 4, a solutionof 2-methyl-7-octadecene [ca. 0.55 g, 90.4% (Z) and 9.6% (E)| in chloroformwas stirred and cooled to — 30°C, and a two-phase mixture prepared by dis-solving potassium hydroxide (3 g) in 40% aqueous tetrabutylammonium hydrox-ide (10 ml) was added dropwise from an addition funnel while a second 5-mlportion of chloroform was added dropwise from a second addition funnel(extremely vigorous initial reaction!). The thick mixture was allowed to slowlywarm to room temperature, stirred for 1.5 days at room temperature, and thenwas thoroughly partitioned between 2 N hydrochloric acid and hexane. Thehexane solution was washed well with water then with aqueous sodium carbon-ate, dried with magnesium sulfate, and concentrated. The residue (729 mg) wasflash chromatographed on silica gel with cyclohexane (essentially no retention)to give 687 mg (ca. 95%) of 4 as a clear oil that consisted (by GC) of 90.8%cis-4, 7.7% trans-4, 0.4% unreacted (Z)-olefin, and 1.2% unreacted (£)-olefin.The mass spectrum of 4 showed ions (abundance) at m/z 123 (10), 109 (10),97 (11), 95 (13), 83 (19), 82 (13), 81 (19), 79 (12), 71 (12), 70 (21), 69 (51),57 (49), 56 (54), 55 (39), 43 (100), 42 (11) , and 41 (58).
For (±) 2-decyl-3-(2-methylhexyl)-l,l-difluorocyclopropane 3, the generalprocedure of Crabbe et al. (1973) was modified slightly in that sodium chlo-rodifluoroaeetate was added in small portions as a solid from a solids additionfunnel to a solution of 2-methyl-7-octadecene in refluxing diglyme. The reactionwas monitored by removing aliquots for GC analysis and was continued unti lessentially all of the olefin had been consumed. Because of the relative unreac-tivity of 2-methyl-7-octadecene, rather large excesses of sodium chlorodifluo-roacetate had to be employed. The olefin employed in this reaction had beenpurified by repeated flash chromatography on silver nitrate-impregnated silica

DISPARLURE ANALOGS 2201
gel (hexanes followed by 2.5% ethyl acetate in hexanes), and the product wasflash chromatographed on silica gel with petroleum ether to give 3 as a 99:1cis-trans mixture. The mass spectrum of 3 showed ions (%) at m/z 149 (2),135 (3), 121 (5), 109 (7), 97 (12), 96 (15), 95 (15), 85 (13), 84 (9), 83 (21),82 (21), 81 (16), 77 (10), 71 (11), 70 (23), 69 (42), 68 (10), 67 (13), 57 (54),56 (49), 55 (36), 43 (100), 42 (13), and 41 (59).
The mixture of 1,2-epoxy-2-(5-methylhexyl)tridecane and 1,2-epoxy-2-(6-methylheptyl)dodecane, 8 and 9, the 1,1-disubstituted epoxides, was preparedas described by Sheads et al. (1975) and used as an unresolved mixture (twogeometric isomers, four diastereomers).
Aziridines 6 and 7 (ca. 98% enantiomeric purity) were synthesized from 2and 1, respectively, as described previously (Oliver and Waters, 1995). Althoughno direct determinations of their enantiomeric purities were made, the conver-sions involve inversion of configuration at each carbon of the three-memberedring, and the enantiomeric composition is believed to reflect that of the startingdisparlure samples [had inversion not been realized at one carbon, a trans-aziridine, not the enantiomerically opposite cis-aziridine, would have beenobtained (Oliver and Waters, 1995)]. The (7R,8S) aziridine 6 was preparedfrom (7S,8R)-2 synthesized here, which had been determined to contain about2.2% of 1 (Oliver et al., 1996), and the (7S,8R) isomer 7 was prepared from asample of 1 that contained about 1.5% of 2. It should be noted that when 6 and7 are referred to as ( + )-AZ and ( —)-AZ, respectively, the ( + ) and ( —) areused to facilitate comparison of absolute configurations to those of ( + )- and( —)-disparlure and do not necessarily indicate direction of rotation of polarizedlight, which has not been determined.
The cyclopropane analogs 3, 4, and 5 (all racemic) were prepared byaddition of the appropriate carbene to (Z)-2-methyl-7-octadecene as describedabove. The stereochemistry of the products closely reflects that of the olefinemployed, and since 4 and 5 were made from olefin containing about 10% ofthe E isomer, the predominantly cis-cyclopropanes contained about 10% of theirtrans-isomers. The olefin used for the preparation of 3 was more rigorouslypurified, and the resulting difluorocyclopropane was approximately 99% trans.
Electrophysiological Techniques. Electroantennograms (EAGs) were usedto compare the efficacy of the homologous analogs on antennal pheromonereceptors in male moths (Kaissling, 1995) and to elucidate potential presenceof receptors in females. EAGs were recorded from adult males and femalesusing a modification of an earlier technique (Schneider, 1957; Dickens et al.,1993b). An excised antenna was fixed between two glass capillary electrodesfilled with 0.1 M NaCl. Ag-AgCI wires in the glass capillaries connected thepreparation to the recording instruments: a Grass P-16 preamplifier, a Tektronix5111A analog storage oscilloscope, and a Alpken 310 stripchart recorder.
Experimental Protocol. Three experiments were performed. The objective

2202 DICKENS, OLIVER, AND MASTRO
of the first experiment was to determine the relative responsiveness of both maleand female antennal preparations to an elevated st imulus load (5 /xg) of allexperimental compounds. Thus, EAGs to 5-/zg stimulus loads were recordedfor both sexes. This experiment was replicated six times for both sexes.
In a second experiment, the responsiveness of male antennal receptors toeach odorant was determined by recording EAGs to serially increasing stimulusloads of the experimental odorants. Six replicates for both sexes were recordedfor this experiment.
The third experiment determined the effects of exposure of male moths forfive hours to volatiles emanating from 50 ptg of ( + )-D, ( — )-D, 2FC, and ( + )-AZ analogs of disparlure (5 /xl of 10 /*g//jl experimental odorant in hexanesolvent) on EAGs to ( + )-D and the plant odor, hexyl acetate [stimulus load =0.05 /ig (-t-)-disparlure or 100 /xg hexyl acetate; these stimulus loads werechosen as they elicit similar EAG depolarizations in male moths prior to adap-tation (ca. 0.5 mV)|. The adapting odorants were selected based on their highactivity relative to the other analogs in the first two experimental scries. Acontrol consisted of exposure of the moth to an equivalent amount (5 p.1) of thehexane solvent alone. During the 5-hr exposure, individual moths were placedin disposable plastic cups (4 cm diameter x 4 cm height) with paper lids andheld in the dark at ambient temperature (ca. 20°C). Ten replicates were recordedfor each adapting odorant in this experiment. Prior to exposure to the adaptingodorant, one antenna was amputated from a male moth and EAGs were recordedto ( + )-D and hexyl acetate. Following the 5-hr exposure, the remaining antennawas amputated and EAGs recorded wi th in less than 3 min in response to thesame odorants.
Odorants used to elicit EAGs were serially diluted in hexane and weredelivered as 5-pd aliquots of an appropriate serial dilution placed on a piece ofWhatman No. 1 filter paper ( 8 x 1 8 mm). The filter paper was inserted into aglass tube (80 mm long X 5 mm ID) and oriented toward the preparation withthe outlet 1 cm from the antenna. The hexane solvent was allowed to evaporatefor 15 sec prior to use of the odor cartridge. Each stimulation lasted 1 sec.Airflow was 1 liter/min as measured by a flowmeter. The time between stimuliwas 2-3 min. Five microliters of the hexane solvent served as a control afterevaporating for 15 sec from the filter paper. In order that responses from differentpreparations might be compared, hexyl acetate (100 /ig st imulus load) was usedas a standard. Hexyl acetate was selected from several green leaf volatiles(Visser et al . , 1979) since responses to it at this stimulus load were reliable.Stimulation with the standard preceded or followed the response to each exper-imental stimulus. Responses are represented as percent responses of the meanof the two nearest responses to the standard.
Statistical Analyses. Electroantennograms were evaluated by analysis of
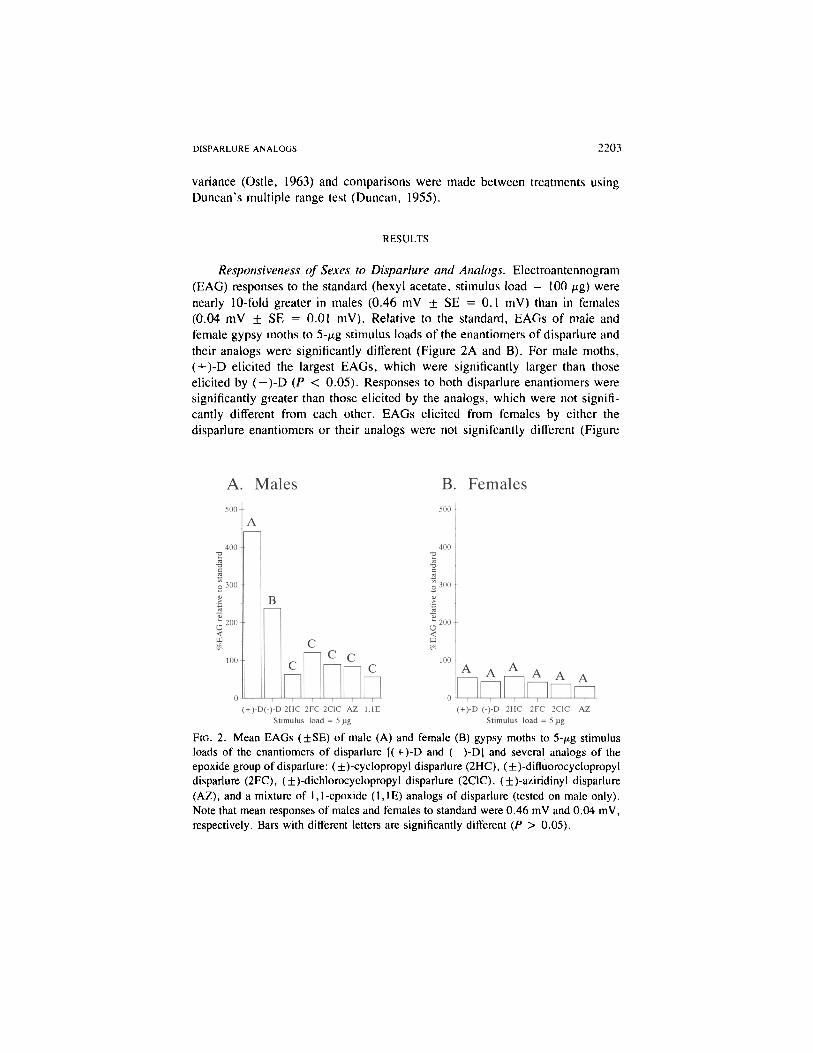
DISPARLURE ANALOGS 2203
variance (Ostle, 1963) and comparisons were made between treatments usingDuncan's multiple range test (Duncan, 1955).
RESULTS
Responsiveness of Sexes to Disparlure and Analogs. Electroantennogram(EAG) responses to the standard (hexyl acetate, stimulus load = 100 jtg) werenearly 10-fold greater in males (0.46 mV ± SE = 0.1 mV) than in females(0.04 mV ± SE = 0.01 mV). Relative to the standard, EAGs of male andfemale gypsy moths to 5-^g stimulus loads of the enantiomers of disparlure andtheir analogs were significantly different (Figure 2A and B). For male moths,( + )-D elicited the largest EAGs, which were significantly larger than thoseelicited by ( —)-D (P < 0.05). Responses to both disparlure enantiomers weresignificantly greater than those elicited by the analogs, which were not signifi-cantly different from each other. EAGs elicited from females by either thedisparlure enantiomers or their analogs were not signifcantly different (Figure
FIG. 2. Mean EAGs (±SE) of male (A) and female (B) gypsy moths to 5-/ig stimulusloads of the enantiomers of disparlure [(+)-D and (-)-D] and several analogs of theepoxide group of disparlure: (±)-cyclopropyl disparlure (2HC), (±)-difluorocyclopropyldisparlure (2FC), (±)-dichlorocyclopropyl disparlure (2C1C), (±)-aziridinyl disparlure(AZ), and a mixture of 1,1-epoxide (1,1E) analogs of disparlure (tested on male only).Note that mean responses of males and females to standard were 0.46 mV and 0.04 mV,respectively. Bars with different letters are significantly different (P > 0.05).

2204 DICKENS, OLIVIER, AND MASTRO
2B). The millivolt responses of females to each compound were significantlyless than responses elicited from male antennae (P < 0.05).
Sensitivity of Antennal Receptors of Males to Disparlure and Analogs.While the shapes of the dose-response curves for the enantiomers of disparlurewere similar, male antennae were at least 100 times more sensitive to ( + )-Dthan to (-)-D [threshold for (-t-)-D = 0.0005 ^g, (-)-D = 0.005 /ig| (Figure3A). Dose-response curves constructed from EAGs of males to cyclopropyl(2HC), difluorocyclopropyl (2FC), and dichlorocyclopropyl (2C1C) analogs wereparallel to those for the disparlure enantiomers but shifted closer to the abscissa(Figure 3B). The dose-response curve obtained for the 1,1-epoxide mixture(1 ,1E) (Figure 3D) was similar to those obtained for the cyclopropyl analogs.Threshold responses were elicited by the 0.5-/xg stimulus load for 2FC and2C1C, but only by 5 /xg for 2HC and 1,1E. Relative stimulus loads necessaryto elicit equivalent responses for ( + )-D and its analogs were estimated as:( + )-D = 1, 2FC = 100, 2C1C = 500, and 2HC, 1,1E = 1000. Dose-responsecurves for the aziridine enantiomers were steeper than those for the other com-pounds tested. EAGs elicited by ( + )-AZ [conformation corresponding to ( + )-D|were greater than those elicited by ( — )-AZ, while intermediate responses weregiven to racemic AZ (Figure 3C). Significant EAGs were recorded for theaziridine analogs only at the 5-fig stimulus load, which was at least 10-foldhigher in threshold than all other analogs.
Adaptation Studies. The adaptation studies revealed differing effects onEAGs to the test stimuli, ( + )-D, and hexyl acetate resulting from adaptation ofmales to ( + )-D and the analogs (Figure 4). Stimulus loads for ( + )-D and hexylacetate (0.05-/ig and 100-/ng stimulus load, respectively) were chosen as theyelicit similar EAG depolarizations in male moths prior to adaptation (ca. 0.5mV). Previous adapting exposure to ( + )-D significantly diminished EAGs to( + )-D by nearly 0.3 mV (60%) (P < 0.05) (Figure 4A). Although exposureto (-)-D reduced EAGs to ( + )-D, exposure to (-)-D, (-)-2FC, or ( + )-AZdid not significantly affect the magnitude of the EAG depolarization elicited by( + )-D. With the exception of ( + )-D, the effects of these latter odorants did notdiffer from the control.
Surprisingly, exposure to both ( + )-D and (-)-D significantly reduced EAGsto hexyl acetate (Figure 4B) (P < 0.05). Neither 2FC nor ( + )-AZ reducedpostexposure EAGs to hexyl acetate. Although postexposure EAGs to hexylacetate following these latter odorants may have been enhanced slightly, theywere not significantly different from the control.
DISCUSSION
Responsiveness and Sensitivity to Disparlure and Analogs. Dose-responsecurves for the enantiomers of disparlure observed in our study differ slightly
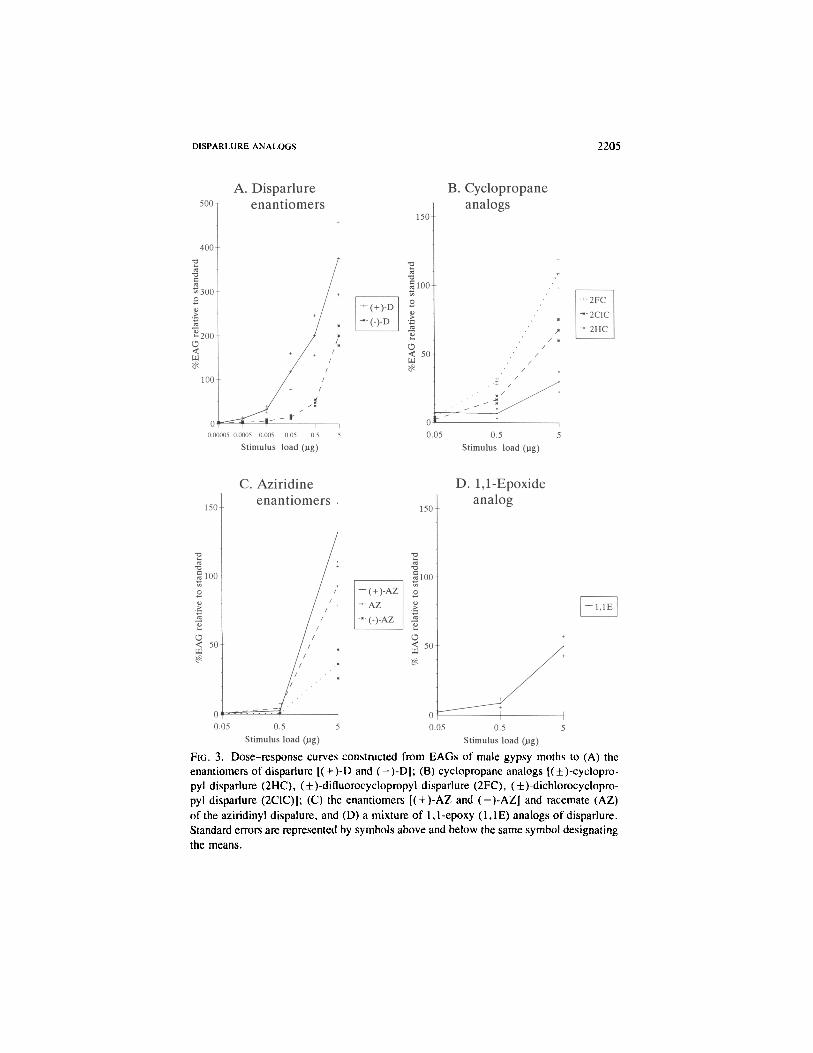
DISPARLURE ANALOGS 2205
FIG. 3. Dose-response curves constructed from EAGs of male gypsy moths to (A) theenantiomers of disparlure [( + )-D and ( — )-D]; (B) cyclopropane analogs [(±)-cyclopro-pyl disparlure (2HC), (±)-difluorocyclopropyl disparlure (2FC), (±)-dichlorocyclopro-pyl disparlure (2CIC)]; (C) the enantiomers [( + )-AZ and (-)-AZJ and racemate (AZ)of the aziridinyl dispalure, and (D) a mixture of 1,1-epoxy (1,1E) analogs of disparlure.Standard errors are represented by symbols above and below the same symbol designatingthe means.
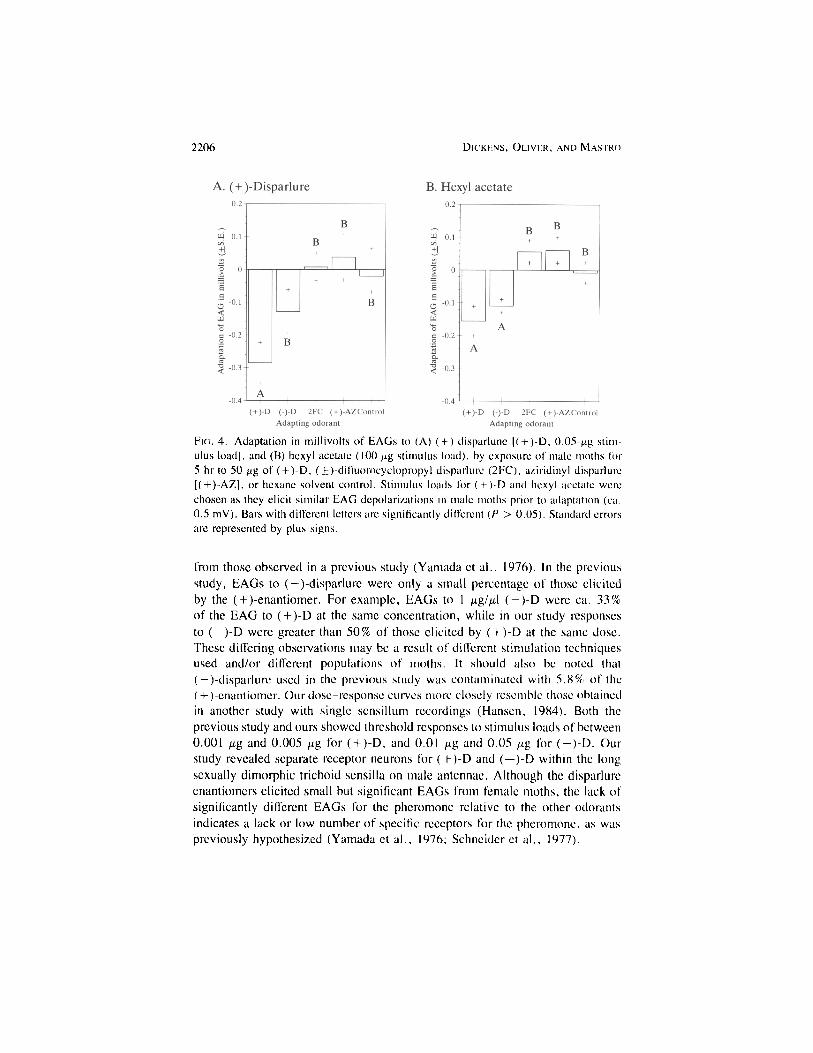
2206 DICKENS, OLIVIER, AND MASTRO
FIG. 4. Adaptation in millivolts of EAGs to (A) ( + )-disparlune |( + )-D, 0.05-/ig stim-ulus load], and (B) hexyl acetate (100 ̂ g stimulus load), by exposure of male moths for5 hr to 50 pg of ( + )-D, (±)-difluorocyclopropyl disparlure (2FC), aziridinyl disparlure[( + )-AZ], or hexane solvent control. Stimulus loads for ( + )-D and hexyl acetate werechosen as they elicit similar EAG depolarizations in male moths prior to adaptation (ca.0.5 mV). Bars with different letters are significantly different (P > 0.05). Standard errorsarc represented by plus signs.
from those observed in a previous study (Yamada et al., 1976). In the previousstudy, EAGs to (-)-disparlure were only a small percentage of those elicitedby the ( + )-enantiomer. For example, EAGs to 1 /ag//il (-)-D were ca. 33%of the EAG to ( + )-D at the same concentration, while in our study responsesto (-)-D were greater than 50% of those elicited by ( + )-D at the same dose.These differing observations may be a result of different stimulation techniquesused and/or different populations of moths. It should also be noted that(—)-disparlure used in the previous study was contaminated with 5.8% of the( + )-enantiomer. Our dose-response curves more closely resemble those obtainedin another study with single sensillum recordings (Hansen, 1984). Both theprevious study and ours showed threshold responses to st imulus loads of between0.001 /xg and 0.005 /*g for ( + )-D, and 0.01 /xg and 0.05 ^g lor (-)-D. Ourstudy revealed separate receptor neurons for (-l-)-D and (—)-D wi th in the longsexually dimorphic trichoid sensilla on male antennae. Although the disparlureenantiomers elicited small but significant EAGs from female moths, the lack ofsignificantly different EAGs for the pheromone relative to the other odorantsindicates a lack or low number of specific receptors for the pheromone, as waspreviously hypothesized (Yamada et al., 1976; Schneider et al., 1977).

DISPARLURE ANALOGS 2207
Similarity in shapes of dose-response curves elicited by the disparlure enan-tiomers and the cyclopropane (C) analogs may be indicative of their interactionwith the same receptors (Kaissling, 1971). Greater activity for dihalogenC ana-logs relative to the cyclopropyl analog (2HC) may be explained by inductiveeffects of the halogen atoms on neighboring carbons that mimic the electron-withdrawing effects of oxygen in the epoxide pheromone. The fact that thedifluoroC (2FC) analog was more active than the dichloroC (2C1C) analog maybe accounted for by the smaller size of the fluorine atom compared to chlorine,thus providing less steric hindrance for 2FC. The greater effectiveness of the2FC analog relative to the 2HC analog is explained principally by inductiveeffects since the atomic radii of fluorine and hydrogen are similar.
Greater BAG activity of ( + )-AZ relative to ( —)-AZ correlates with therelationship of these analogs with ( + )-D and (-)-D. Nitrogen (nitrile) analogsof aldehyde pheromone components of the boll weevil and the southwesterncornborer moth were active in electrophysiological studies and slightly active infield tests with the boll weevil (Ujvary and Dickens, 1991).
Both racemic AZ and 1,1E of disparlure were previously synthesized andtested in the field (Sheads et al., 1975). While racemic AZ was somewhatattractive, 1,1E was nearly as attractive as racemic disparlure, although insectcaptures in unbaited traps were not reported. When combined with disparlure,no effect on the attractiveness of disparlure was noted for either racemic AZ or1,1E. The authors stated that "it would be interesting to determine whetherelectroantennogram responses of the compounds would parallel captures ofinsects in the field". Our current results show antennal receptors of male gypsymoths to be 1000-fold less sensitive to 1,1E than ( + )-D and perhaps even lesssensitive to racemic AZ. Thus, our results do not correlate with the observationsmade in the earlier study.
The low sensitivity of antennal receptors for the cyclopropyl, aziridinyl,and 1,1-epoxy analogs of disparlure may be indicative of the stringent require-ments for interaction of related compounds with disparlure receptors. This lowsensitivity of disparlure-sensitive neurons was also apparent in single sensillumrecordings in which only a few spikes were elicited by 2FC even at the 5-p.gstimulus load (Dickens, unpublished). We realize that comparisons made betweenEAGs elicited by the compounds at different stimlus loads are imprecise due topossible differences in the number of molecules evaporating from the filter paperas a result of differing volatilities of the compounds and inherent molecularproperties causing differential binding to the filter paper. It was suggested in arecent study with another difluorinated pheromone analog that the retarded evap-oration rate from filter paper observed for the analog was caused by interactionof flurorine with the hydroxyl groups of cellulose (Klun et al., 1996). Of thecompounds considered here, 2C1C (4), (-t-)-AZ (6), and (-)-AZ (7) are theleast volatile, but we lack a formula for estimating the amounts of material

2208 DICKENS, OLIVIER, AND MASTRO
actually volatilizing from the paper. For purposes of comparison, the order ofelution from a nonpolar GC column is 3 > 5 > l = 2 > 6 = 7>8 + 9>4. The order of activity observed for the compounds in BAG studies was 1 >2 » 3 > 4 > 5 > 8 + 9 » 6 > 7 . Thus differences in volatility alonedo not explain substantial differences in EAGs observed in our study.
Adaptation Studies. Interaction of the disparlure enantiomers and their ana-logs with receptor processes for (+ )-D were further revealed by the adaptationexperiments. Adaptation of responses to ( + )-D by prolonged exposure to it wasexpected, and provided a test of the experimental procedure. The fact thatexposure to (-)-D did not cause adaptation to ( + )-D was not totally unexpectedsince specialized neurons housed in the same trichoid sensilla respond selectivelyto either ( —)-D or ( + )-D (Hansen, 1984). Thus, processes for the transport andinteraction of the two molecules with their receptors may not significantly over-lap, even though under certain conditions both molecules may bind to the samepheromone binding protein in the sensillar lymph (Vogt et al., 1989). The lackof an effect of 2FC and (-l-)-AZ analogs on postexposurc responses to (-t-)-Dfurther reflect their ineffectiveness in interactions with receptor processes for thepheromone.
Adaptation to hexyl acetate by (4-)-D and ( —)-D was unexpected. Receptorneurons for plant volatiles are housed in different sensilla than those housingneurons responsive to the pheromone (Kaissling, 1971). Processes for transportand interaction of such molecules with responsive dendrites may differ, as, forexample, the localization of pheromone binding proteins and general odorantbinding proteins (GOBPs) in different sensilla (Laue et al . , 1994). However,sex pheromones in saturniid moths may bind general odorant binding proteinsin females (Vogt and Riddiford, 1981; Vogt, 1987; Vogt et al., 1991). Thismay indicate a degree of flexibility in this and possibly other perireceptor events.In any case, the physical or biochemical processes responsible for our resultsremain unclear and will require further experimentation.
The relative lack of activity of the three-membered ring analogs of dispar-lure in our electrophysiological studies was recently verified in field tests inwhich 2FC was unattractive alone and had no significant effect on the numberof male moths responding to ( + )-D or (±)-D (Mastro et al., unpublished). Thelack of activity of 2FC of the other compounds tested in our electrophysiologicalstudies reflect the specificity of receptors for ( + )-D and (-)-D, as previouslyreported for a number of analogs used in other studies (Bierl et al., 1972; Adleret al., 1972; Sanniento et al., 1972; Schneider et al., 1974). These resultsindicate that novel approaches to the design and synthesis of potential agonistsor antagonists of disparlure receptors are necessary before substantial effects onthe activity of disparlure are realized.
Acknowledgments—The authors wish to thank Dr. D. Rowe, USDA, ARS, Mississippi State,Mississippi for assistance in statistical analyses; Dr. B. A. Leonhardt. USDA, ARS, Plant Sciences

DISPARLURE ANALOGS 2209
Institute, Beltsville, Maryland, for encouragement and support; and Dr. John Davis and Ms. SiniPanicker tor technical assistance. Critical reviews of the manuscript were provided by ProfessorF. E. Hanson, Department of Biological Sciences. University of Maryland-B.-C., Baltimore, Mary-land, and Dr. D. M. Light, USDA, ARS, Western Regional Research Center, Albany, California.
REFERENCES
ADLER, V., BEROZA, M., BIeRL, B. A., and SARMIeNTO, R. 1972. Electroantennograms and fieldattraction of the Gypsy moth sex attractant disparlure and related compounds. J. Econ. Entomol.65:679-681.
BIERL, B. A., BeROzA, M., and COLLIER, C. W. 1970. Potent sex attractant of the Gypsy moth:its isolation, identification and synthesis. Science 170:87-89.
BIERL, B. A., BEROzA, M., and COLLIER, C. W. 1972. Isolation, identification and synthesis of theGypsy moth sex attractant. J. Econ. Entomol. 65:659-664.
CRABBE, P., CERVANTES, A., CRUZ, A., GALEAZZI, E., IRIARTE, J., and VELARDE, E. 1973. Chem-istry of difluorocyclopropyl acetates. Application of difluorocarbene chemistry to homologationreactions. J. Am. Chem. Soc. 95:6655-6665.
DICKENS, J. C., VISSER, J. H., and VAN DER PERS, J. N. C. I993a. Detection and deactivation ofpheromone and plant odor components by the beet armyworm, Spodopteru exigua (Hubner)(Lepidoptera: Noctuidae). J. Insect Physiol. 39:503-516.
DICKENS, J. C., SMITH, J. W., and LIGHT, D. M. 1993b. Green leaf volatiles enhance sex attractantpheromone of the tobacco budworm, Heliothis virescens (Lep.: Noctuidae). Chemoecology4:175-177.
DUNCAN, D. B. 1955. Multiple range and multiple F tests. Biometrics 11:1-42.HANSEN, K. 1984. Discrimination and production of disparlure enantiomers by the gypsy moth and
nun moth. Physiol. Entumol. 9:9-18.KAISSLING, K.-A. 1971. Insect olfaction, pp. 351-431, in L. Beidler (ed.). Handbook of Sensory
Physiology. Vol. IV. Chemical Senses. I. Springer Verlag, New York.KAISSLING, K.-A. 1995. Single unit and electroantennogram recordings in insect olfactory organs,
pp. 361-377, in A. I. Spielman and J. G. Brand (eds.) Experimental Cell Biology of Tasteand Olfaction. CRC Press, Boca Raton, Florida.
KLUN, J. A., OLIVER, J. E., K H R I M I A N , A. P., DICKENS, J. C., and POTTS, W. J. E. 1997. Behavioraland electrophysiological activity of the racemate and enantiomers of a monofluorinated analogof European com borer sex pheromone. J. Entomol. Sci. 32:37-49.
LAUE, M., STEINBRECHT, R. A., and ZIEGELBERGER, G. 1994. Immunocytochemical localization ofgeneral odorant-binding protein in olfactory sensilla of the silkmoth Antheraea polyphemus.Naturwissenschaften 81:178-180.
LIGHT, D. M., FLATH, R. A., BUTTERY, R. G., ZALOM, F. G., RICE. R. E., DICKENS, J. C., andJANG, E. B. 1993. Host plant green leaf volatiles synergize the synthetic sex pheromones ofthe com earworm and codling moth (Lepidoptera). Chemoecology 4:145-152.
MARUOKA, K., FUKUTANI , Y., and YAMAMOTO, H. 1985. Trialkylaluminum-alkylidene iodide. Apowerful cyclopropanation agent with unique selectively. J. Org. Chem. 50:4412-4414.
OLIVER, J. E., and WATERS, R. M. 1995. Determining the enantiomeric purity of disparlure. J.Chem. Ecol. 21:199-211.
OLIVER, J. E., WATERS, R. M., and HARRISON, D. J. 1996. Semiochemicals viaepoxide inversion.J. Chem. Ecol 22:287-294.
OSTLE, B. 1963. Statistics in Research, Iowa State University Press, Ames, Iowa, xv + 585 pp.PLIMMER, J. R., LEONHARDT, B. A., WEBB, R. E., and SCHWALBE, C. P. 1982. Management of
the gypsy moth with its sex attractant pheromone. Am. Chem. Soc. Symp. Ser. 190:231-242.

2210 DICKENS, OLIVER, AND MASTRO
SARMIENTO, R., BEROZA, M., B I E R L , B. A., and TARDIF , J. G. R. 1972. Ac t iv i ty of compoundsrelated to disparlure, the sex attractant of the gypsy moth. J. Econ. Entomol. 65:665-667
SCHNEIDER, D. 1957. Elektrophysiologische Untersuchungen von Chemo- und Mechanorezeptorender Antenne des Seidenspinners Bombyx mori. Z. Vergl. Physiol. 40:8-41.
SCHNEIDER, D., LANGE, R., SCHWARZ, F., BEROZA, M., and B I E R I , B. A. 1974. Attraction of malegypsy and nun moths to disparlure and some of its chemical analogues, (Oecologia 14:10-36.
SCHNEIDER, D., K A F K A , W. A., BEROZA, M., and B I E R I , B. A. 1977. Odor receptor responses ofmale gypsy and nun moths (Lepidoptera, Lymantriidae) to disparlure and its analogs. J. Comp.Physiol. 113:1-15.
SCHWALBE, C. P., PASZEK, E. C., BIERL-LEONHARDT, B. A., and PLIMMER, J. R. 1983. Disruptionof gypsy moth (Lepidoptera: Lymantriidae) mating w i t h disparlure. J. Econ. Entomol.76:841-844.
SHEADS, R. E,., BEROZA, M., and PASZER, E. C. 1975. Chemicals related to the gypsy moth sexpheromone as attractants and as synergists or inhibi tors of the pheromone. J. Agric. FoodChem. 23:60-64.
U J V A R Y , I., and D I C K E N S , J. C. 1991. Novel nitrogen-containing funct ional group mimies of alde-hyde insect pheromones, pp. 45-50, in I. Hrdy (ed.) . Insect Chemical Ecology. SPB AcademicPubl., The Hague, The Netherlands.
VISSER, J. H., VAN STRATEN, S., and MAARSE., H. 1979. Isolation and ident i f ica t ion of v o l a t i l e s inthe foliage of potato, Solanum tuberosum, a host plant of the Colorado beetle, Leptinotarsadecemlineata. J. Chem. Ecol. 5:13-25.
VITE, J. P., KLIMETZEK, D., LOSKANT, G.. HEDDEN, R., and MORI, K. 1976. Chiralily of insectpheromones: Response interruption by inactive antipodes Naturwissenschaften 63:5X2 5X3.
VOGT, R. G. 1987. The molecular basis of pheromone reception: its influence on behavior, pp.385-431, in G. D. Prestwich and G. J. Blomquist (eds.). Pheromone Biochemistry. AcademicPress, New York.
VOGT, R. G. and RIDDIFORD, L. M. 1981. Pheromone binding and inactivation by moth antennae.Nature 293:161-163.
VOGT, R. G., KOHNE, A. C., D U B N A U , J. T., and PRESTWICH, G. D. 1989. Expression of pheromonebinding proteins during antennal development in the gypsy moth Lymantria dispar. J. Neurosci.9:3332-3346.
VOGT, R. G., PRESTWICH, G. D., and L E R N E R , M. R. 1991. Odorant-binding-protein subfamil iesassociate with distinct classes of olfactory receptor neurons in insects. J. Neurobiol. 22:74-84.
YAMADA, Y., SAITO, T., KATAGIRI, K., IWAKI. S., and MARUMO, S. 1976. Electroantennogramand behavioural responses of the Gypsy moth to enantiomers of disparlure and its trans analogs.J. Insect Physiol. 22:755-761.