Embed Size (px)
Citation preview

779
T. S. GUNASEKERA, N. D. PAUL* AND P. G. AYRES
Division of Biological Sciences, Institute of Environmental and Biological Sciences, Lancaster University, Lancaster LA1 4YQ , U.K.
The sensitivity to UV-B (290–320 nm) radiation of common phylloplane yeasts from two contrasting UV-B environments was
compared in the laboratory using mixtures of white light (PAR: 400–700 nm) and UV-B radiation from artificial lamp sources.
Sporidiobolus salmonicolor, Rhodotorula mucilaginosa and Cryptococcus sp., the dominant yeasts on leaves of tea (Camellia sinensis),
were isolated in Sri Lanka (SL), while Sporidiobolus sp. and Bullera alba, dominant on faba bean (Vicia faba), were isolated in the
U.K. Dose responses were determined separately for each yeast. UV-B reduced colony forming units (due to cell mortality or
inactivation) and colony size (due to reduced multiplication) of all yeasts. The LD&!
values and doses causing 50% reduction of cells
per colony were higher for SL isolates than U.K. isolates. Results indicated that each yeast is somewhat vulnerable to UV-B doses
representative of its natural habitat. The relative insensitivity of SL isolates was shown when SL and U.K. isolates were irradiated
simultaneously with the same dose of UV-B. Of the two U.K. yeasts, B. alba was significantly more sensitive than Sporidiobolus sp.
to UV-B. Except for R. mucilaginosa from SL, all yeasts demonstrated some photorepair in the presence of white light. White light
provided relatively little protection for the U.K. isolate of Sporidiobolus sp. although it allowed increased colony size.
The spectral responses of Sporidiobolus sp. (U.K.) and of B. alba (U.K.) were broadly similar. Wavelengths longer than 320 nm had
no measurable effect on colony forming units. However, colony survival was significantly reduced at 310 nm and all shorter
wavebands. No colonies were counted at 290 nm or below.
Leaves are displayed to intercept maximum incident solar
radiation. Leaves are characteristically colonized by epiphytic
communities of bacteria, yeasts and filamentous fungi (Last &
Price, 1969 ; Andrews & Kenerly, 1979 ; Campbell, 1985 ;
Thompson et al., 1993 ; Dix & Webster, 1995) so, at least on
unshaded surfaces, this particular microbial habitat is subject
to relatively high doses of solar radiation (Lindow, 1991). The
ultraviolet component of solar radiation is, in general,
damaging to living organisms, so it is often assumed that the
shorter wavelengths that reach the Earth’s surface (UV-B,
290–320 nm) have a major role in regulating both the size and
balance of the microbial populations (Ayres et al., 1996). There
is, however, little empirical evidence to support this
assumption.
UV fluxes reaching the Earth’s surface broadly increase with
decreasing latitude and increasing elevation. Superimposed on
this pattern are daily and, especially in temperate latitudes,
seasonal fluctuations. We tested the hypothesis that the leaf
surface microbes are adapted to their local environment by
examining the response to UV-B of phylloplane organisms
isolated from contrasting environments, that is from leaves, of
tea, growing at an elevation of 1400 m in the tropics (latitude
7°N) and from those, of faba bean, growing at sea level in
* Corresponding author.
temperate NW Europe (latitude 54°N). Experiments were
carried out in vitro so that doses of UV-B could be
precisely regulated. Yeasts were chosen for the study because
of their ubiquity on leaf surfaces and the relative ease with
each they can be quantified. We studied the types, three from
tea and two from faba bean, dominant after leaf washings were
plated on malt extract agar.
Since there is evidence that longer wavelengths of solar
radiation (white or visible light, 400–700 nm), even at
extremely low fluxes, may protect organisms from damage by
UV-B, the effect of white light on UV-B responses of the test
organisms was examined.
MATERIALS AND METHODS
Isolation of yeasts
Yeasts were isolated from leaves of tea (Camellia sinensis L.)
and bean (Vicia faba L.) in Sri Lanka and the U.K. respectively.
Leaf samples were excised and immediately brought to the
laboratory. Leaf discs were cut out using a 10 mm diam. cork
borer and shaken for 15 min in 20 cm$ sterile water containing
0±05% (v}v) Tween 80. The washings were serially diluted in
sterile water and 0±25 cm$ of washings were transferred to
9 cm diam. Petri dishes containing suitable culture medium.
Plates were incubated for 72 h at 22 °C in darkness and yeasts
Mycol. Res. 101 (7) : 779–785 (1997) Printed in the United Kingdom
Responses of phylloplane yeasts to UV-B (290–320 nm)radiation: interspecific differences in sensitivity

UV-B and phylloplane yeasts 780
were isolated. Three dominant yeasts from leaves of tea and
two from faba bean were isolated and were maintained on 5%
(w}v) malt extract agar (MEA). Subcultures were identified by
the International Mycological Institute, Bakeham Lane, Egham,
Surrey, U.K., as, respectively Sporidiobolus salmonicolor Fell &
Tallman, Rhodotorula mucilaginosa (A. Jo$ rg.) F. C. Harrison and
Cryptococcus sp., from tea and Sporidiobolus sp. and Bullera alba
(W. F. Hanna) Derx, from bean.
Dose–response curves
Subcultures less than 48 h old, in an exponential growth
phase, were used. Irradiation was carried out in a dark room,
where temperature was maintained at 20³2°. Suspensions of
yeast cells of 1500–2000 cm−$ were made up in sterile
distilled water, and 0±2 cm$ was spread over the surface of
MEA Petri dishes, using a sterile glass spreader, before being
irradiated.
Broad band UV-B (Phillips TL UV-B 40W}12) and daylight
fluorescent tubes (Osram L58W}23) were used to provide
UV-B and white light respectively. UV-B sources were filtered
with 0±1 mm thick cellulose diacetate film (‘Clarifoil ’, supplied
by Courtaulds Ltd, Derby, U.K.) to remove wavelengths
below approximately 292 nm. UV irradiance was measured
using a double monochromator scanning spectroradiometer
(Macam SP991-PC, Livingstone, U.K.) and irradiances were
weighted according to the DNA action spectrum of Setlow
(1974) normalized to 1 at 300 nm. The spectroradiometer was
calibrated for wavelength using the spectral lines from a
mercury arc lamp (LOT Oriel, Leatherhead, Surrey, U.K.) and
for spectral irradiance against tungsten and deuterium sources
(Macam SR903, Livingstone, U.K.) referable to National
Physics Laboratory Standards. To provide a range of fluxes,
the distance between the Petri dishes and uv tubes was varied
by hanging the tubes at an angle approximately 30° to the hori-
zontal surface on which plates were arranged. Irradiation was
carried out against both dark and white light (400–700 nm¯250 µmol m−# s−") backgrounds in order to investigate the
possibility of photoreactivation.
After exposure to uv radiation, plates were further
incubated for up to 66 h in the dark at 20° and visible
macrocolonies were counted. To determine the number of
cells per colony, 5–10 colonies were carefully picked off into
20 cm$ water containing 0±05% (v}v) Tween 80 and shaken
for 15 min before cells were counted using a haemocytometer.
Dose–response studies were carried out separately for each
organism.
Relative sensitivity to UV-B radiation
To compare the yeasts directly all five isolates (two U.K. and
three SL) were simultaneously exposed to the same dose of
UV-B (4±98 kJ m−# UV-BDNA
). A suspension of 1500–2000
cells cm−$ was prepared as above and 0±2 cm$ was spread
onto five replicate Petri dishes (9 cm) containing 5% (w}v)
MEA. Irradiations were carried out under a white light
(400–700 nm¯ 250 µmol m−# s−") background for 6 h. Cells
were subsequently incubated for up to 66 h in the dark at 20°and visible macrocolonies were counted. Controls were
covered with a polyester filter supplied by Lee Filters,
Andover, Hants, U.K. (spectrally equivalent to Mylar, which
removes wavelengths shorter than approximately 320 nm, i.e.
removes most of the UV-B) so that they were exposed only
to white light and whatever UV-A radiation (" 320 nm) was
emitted from the UV-B sources.
Effect of white light on colony forming ability of yeast
isolates
In the experiments above, white light was provided in order
to test the possible occurrence of photoreactivation, but white
light alone might influence yeast cell growth. To test this,
each of the five yeasts was dispersed onto 9 cm Petri dishes
containing 5% (w}v) MEA and then exposed to 6 h white
light (approximately 250 µmol m−# s−"). Samples were incu-
bated for a further 60 h in the dark at 20° and numbers of
visible macrocolonies were counted. Results were compared
with those for unexposed (dark) controls. The experiment was
repeated three times and each had five replicates.
Effectiveness of different wavebands within the UV
spectrum
A concentration of approximately 500 cells cm−$ was prepared
with sterile distilled water, and 100 µl of the suspension was
spread onto 4±5 cm Petri dishes containing 5% (w}v) MEA,
with a glass spreader. Cells were then exposed to different
wavebands of uv radiation within the range of 270–380 nm.
To provide defined ranges of narrow band uv, two 75 W
xenon arc lamps, each in a ‘Q arc ’ housing with four outlet
ports, were used. Each of the four output beams was
collimated, and visible and infrared radiation was attenuated
using dicroic beam turners. Narrow band interference filters
(10 nm width at half peak height) were placed in between the
specimen and the beam to obtain the required waveband. The
distance of the colonies from the light source was adjusted
until all treatments received 1±1 Wm−#. All components were
supplied by LOT Oriel (Leatherhead, Surrey, U.K.).
Statistical analysis
Dose–response curves. Analysis of variance (ANOVA) and
non-linear regressions for mortality and cells per colony
(colony size) were made using SPSS (SPSS Inc., Michigan Av.,
Chicago, U.S.A.) statistical software. Two regression models
were used ; Y¯ 1}(1}ub!(b
"
t)) and Y¯ b!b
"tb
#t# for
mortality and cells per colony respectively, where b!, b
", b
#¯
constants ; u¯ upperbound value for the logistic model ; t¯a time value or the value of an independent variable : e.g. UV-
B dose. LD&!
values and the dose causing 50% reduction of
cells per colony were calculated from fitted curves.
Response of different yeast isolates to UV-B radiation.
Analysis of variance and Tukey–HSD multiple range tests
were used to detect significant differences among treatments
using SPSS statistical software. Percentages were arcsin
transformed before applying statistical tests.

T. S. Gunasekera, N. D. Paul and P. G. Ayres 781
Effect of white light (PAR) on colony forming ability of
yeast isolates. Student’s t test and analysis of variance
(ANOVA-general factorial) were used to compare the means
of dark and light grown colonies of each isolate.
RESULTS
Dose–response curves
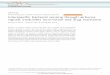
The effect of increasing UV-B dose on cell mortality and cells
per colony of S. salmonicolor (SL) is shown in Fig. 1. Dose
responses for the other organisms are not presented in such
detail, since their form was similar in each case. However, their
LD&!
values and the dose causing a 50% reduction of cells per
colony (colony size) were calculated from the fitted curves,
and these are summarized in Table 1. The values estimated for
regression models for dose–response curves, R# (coefficient of
determination), and P (significance) values are also summarized
in Table 1.
In general, increasing UV-B dose had a highly significant
(P! 0±001) effect, increasing mortality and decreasing cells
per colony of all yeasts, except B. alba which showed a less
significant (P! 0±01) effect on cells per colony under a light
background.
There was variation between yeasts of similar origin but
LD&!
values were higher for SL isolates than U.K. isolates,
irrespective of background irradiation. Similarly, higher UV-B
doses were required to reduce cells per colony (colony size) by
50% for SL isolates than for U.K. isolates, both under light and
dark backgrounds (Table 1). For S. salmonicolor (SL), Crypto-
100
9080
7060
5040
30
20
10
00 2 4 6 8 10 12 14
R2 = 0·7756
0·00E + 000 2 4 6 8 10 12 14
R2 = 0·8638
1·00E + 06
2·00E + 06
3·00E + 06
4·00E + 06
5·00E + 06
6·00E + 06
7·00E + 06
Cel
ls p
er c
olon
y
Mor
tali
ty (
%)
100
90
80
70
60
50
40
30
20
10
00 2 4 6 8 10 12 14
R2 = 0·67480
0·00E + 000 2 4 6 8 10 12
R2 = 0·88117
1·00E + 06
2·00E + 06
3·00E + 06
4·00E + 06
5·00E + 06
–10
(b)
(d )
(a)
(c)
Does (kJ m–2 d–1)
Fig. 1. Response of Sporidiobolus salmonicolor (SL) to UV-BDNA
dose : (a) colony number (per cent mortality) in the dark, (b) colony size
(cells per colony) in the dark, (c) colony number in white (PAR: 400–700 nm) light and (d ) colony size (cells per colony) in white light.
R#, coefficient of determination for the fitted curve. Each point represents data for a single Petri dish or, in the case of colony size, the
mean of 5–10 colonies per plate.
coccus sp. (SL) and B. alba (U.K.), the inhibitory effect of UV-
B on survival and colony size was reduced by concomitant
irradiation with white light, i.e. there was evidence of
photoreactivation. In the U.K. isolate of Sporidiobolus photo-
reactivation was less apparent, but colony size was increased
under a light background. In contrast, in R. mucilaginosa the
reduction in both survival and colony size by UV-B was
greater under a light background than under a dark background
(Table 1).
Relative sensitivity to UV-B radiation
Sri Lankan (SL) isolates were significantly (P! 0±05) less
susceptible to damage than U.K. isolates when all five were
exposed simultaneously to UV-B (4±98 kJ m−# UV-BDNA
)
radiation (Fig. 2). Susceptibility did not differ significantly
(P! 0±05) between the three SL yeasts but the two U.K.
yeasts showed different sensitivity ; Sporidiobolus sp. was
significantly (P! 0±05) less susceptible than B. alba (Fig. 2),
which did not survive at this dose.
Effect of white light (PAR) on number of colonies of
yeast isolates
White light alone had no significant (P! 0±05) effect on the
number of colonies of any of the five yeast isolates. However,
S. salmonicolor from SL showed some slight sensitivity to
white light (Fig. 3).

UV-B and phylloplane yeasts 782
Table 1. Dose of UV-BDNA
required to cause 50% cell mortality (LD&!) and 50% reduction in cells per colony (ED
&!) of five phylloplane yeasts cultured
with white light or dark background
Source Phylloplane yeast Fitted dose–response curve
r# and P
value
LD&!}ED
&!†
(kJ m−# d−" UV-BDNA
)
(a) Mortality
White light background
SL S. salmonicolor 1}((1}100)(1±15¬0±58D)) 0±67*** 8±64³0±42SL Cryptococcus sp. 1}((1}100)(4±78¬0±45D)) 0±80*** 7±92³0±72SL R. mucilaginosa 1}((1}100)(0±29¬0±70D)) 0±55*** 7±25³0±63U.K. Sporidiobolus sp. 1}((1}100)(2±87¬0±23D)) 0±55*** 3±94³0±42U.K. B. alba 1}((1}100)(52±4¬0±02D)) 0±85** 2±23³0±18
Dark background
SL S. salmonicolor 1}((1}100)(0±83¬0±54D)) 0±67*** 7±29³0±39SL Cryptococcus sp. 1}((1}100)(0±66¬0±51D)) 0±75*** 6±24³0±47SL R. mucilaginosa 1}((1}100)(0±09¬0±69D)) 0±48*** 9±49³0±88U.K. Sporidiobolus sp. 1}((1}100)(0±85¬0±30D)) 0±55*** 4±06³0±43U.K. B. alba 1}((1}100)(8±27¬0±22D)) 0±86*** 1±78³0±13
(b) Colony size (cells per colony)
White light background
SL S. salmonicolor (6±74¬10')®(5±64¬10&¬D)(3701¬D#) 0±86*** 6±24³0±35SL Cryptococcus sp. (1±43¬10()®(1±78¬10'¬D)(54636¬D#) 0±87*** 4±68³0±41SL R. mucilaginosa (1±01¬10()®(1±42¬10'¬D)(52038¬D#) 0±82*** 4±30³0±37U.K. Sporidiobolus sp. (5±70¬10')®(2±20¬10'¬D)(204354¬D#) 0±80*** 1±50³0±12U.K. B. alba (1±17¬10')®(5±37¬10&¬D)(57736¬D#) 0±53*** 1±27³0±09
Dark background
SL S. salmonicolor (3±27¬10')®(5±52¬10&¬D)(23634¬D#) 0±88*** 3±49³0±37SL Cryptococcus sp. (1±25¬10()®(1±78¬10'¬D)(23460¬D#) 0±87*** 4±12³0±39SL R. mucilaginosa (9±53¬10')®(9±60¬10&¬D)(62733¬D#) 0±82*** 5±79³0±40U.K. Sporidiobolus sp. (3±40¬10')®(1±61¬10'¬D)(185119¬D#) 0±83*** 1±22³0±15U.K. B. alba (3±52¬10&)®(2±66¬10&¬D)(47141¬D)# 0±83*** 0±77³0±09
LD&!
and ED&!
values and their standard errors were calculated from dose–response curves (see Fig. 1 for example). The equations of the fitted dose–response
curves, their associated coefficients of determination r# and statistical significance are also given for each yeast. D¯UV-B dose, ** and ***, significant at
P! 0±01 an P! 0±001 respectively.
† LD&!, mortality ; ED
&!, colony size (cells per colony).
0
5
10
15
20
25
30
35
40
45
50
a
a
a
b
SLPY SLRY SLWY BPY BWY
c
Yeast isolate
Sur
viva
l (ar
csin
tran
sfor
med
) (%
)
Fig. 2. Effects of UV-B radiation (4±98 kJ m−# UV-BDNA
) on survival
of colonies of five yeasts. Irradiated colonies are compared with dark
controls. SLPY: Sporidiobolus salmonicolor (SL) ; SLRY: Rhodotorula
mucilaginosa (SL) ; SLWY: Cryptococcus sp. (SL) ; BPY: Sporidiobolus sp.
(U.K.) ; BWY: Bullera alba (U.K.). Different letters indicate significant
(P! 0±05) difference. Bars indicate standard error of mean of five
replicates. The experiment was repeated with similar results.
0
0·2
0·4
0·6
0·8
1
1·2
SLRY SLWY BPY BWY
Yeast isolate
cfu
(lig
ht/d
ark)
SLPY
Fig. 3. Effects of white light on five yeasts. Ratios of colony forming
units (cfu) in light and dark are shown. SLRY: Rhodotorula mucilaginosa
(SL) ; SLPY: Sporidiobolus salmonicolor (SL) ; SLWY: Cryptococcus sp.
(SL) ; BPY: Sporidiobolus sp. (U.K.) ; BWY: Bullera alba (U.K.). Bars
indicate standard error of three repeated experiments, each had five
replicates.
The effectiveness of different bands within the UV
spectrum
The spectral responses of Sporidiobolus sp. (U.K.) (Fig. 4a) and
of B. alba (U.K.) (Fig. 4b) were broadly similar. Wavelengths

T. S. Gunasekera, N. D. Paul and P. G. Ayres 783
120
100
80
60
40
20
0270 280 289 300 310 320 330 340
Wavelength (nm)
Sur
viva
l (%
)
(b)
120
100
80
60
40
200
270 280 289 300 310 320 330 340
(a)
Fig. 4. Effects of different wavebands of UV radiation on the survival
of colonies of (a) Sporidiobolus sp. and (b) Bullera alba. Irradiated
colonies are compared with dark controls. Bands transmitted by the
filters were 10 nm wide at half peak height. Four measurements were
made with each filter and are plotted at the band centres. The
histogram shows the mean values for each waveband.
longer than 320 nm had no significant effect on the number of
colonies. However, colony survival was significantly reduced
at 310 nm and all shorter wavebands. No colonies were
counted at 290 nm or below.
DISCUSSION
Both colony number and size were examined here because, as
elsewhere, the highest doses of UV-B killed cells, probably
through direct damage to DNA (Zo$ lzer & Kiefer, 1983).
Biphasic survival curves similar to those reported here have
been found for the yeast community of the apple phylloplane
and interpreted as evidence for ‘multiple-hit ’ inactivation
kinetics (Pennycook & Newhook, 1982). UV-B at lower doses
may kill only part of any population and, also, may reduce
colony size by affecting rates of cell division. Thus, James &
Nasim (1987) found uv caused an immediate post-irradiation
delay in the division of Saccharomyces cells, followed by a
further delay in the production of second-generation buds but,
thereafter, a stimulation in the rate of cell division which
persisted through several generations. Liefer et al. (1986)
found both budding and division were equally delayed in
Sacchomyces cerevisiae Meyen ex E. C. Hansen by radiation at
313 nm while radiation at 303 nm affected budding signi-
ficantly less than division. Since yeast cells growing in the
exponential phase were used in the present study, emergence
of buds and DNA synthesis may have been affected by
exposure to UV-B.
When filtered with cellulose diacetate, the spectrum of UV-
B lamps (Philips TL UV-B 40W}12) does not include the
wavelengths shorter than 292 nm. Since polyester does not
sharply separate longer wavelengths of UV-B (! 320 nm)
and shorter wavelengths of UV-A (" 320 nm) a little radiation
at wavelength" 320 nm was presented. However, the
spectral responses of two yeasts, Sporidiobolus sp. (U.K.) and B.
alba (U.K.), were examined in detail (Fig. 4) and both showed
that UV-A caused little damage. Even within the UV-B range,
shorter wavelengths were much more effective than longer
wavelengths approaching 320 nm. Our studies were not
designed to establish action spectra and we do not exclude the
possibility that the higher irradiance of UV-A present in
natural sunlight may have some deleterious effect on
phylloplane yeasts. However, our data clearly indicate that it
is the UV-B component of solar radiation that is primarily
effective in regulating populations of phylloplane yeasts.
Confirmation comes from field studies which consistently
show that polyester screens, which remove UV-B from
sunlight, significantly increase populations of phylloplane
yeasts (Ayres et al., 1996 ; Gunasekera, 1996).
Both dose–response studies and simultaneous irradiation
(using the same dose) showed the isolates from Sri Lanka were
less sensitive to UV-B than were the U.K. isolates. Five is a
small sample on which to base a firm conclusion other than
that there are large and significant differences between species
in their sensitivity to UV-B, and the extent of intra-specific
variability remains to be investigated, but the results do
suggest that the incidence of UV-B-tolerant organisms may
increase with decreasing latitude. Among higher plants similar
differences are well known. Sullivan & Teramura (1988)
showed that species native to higher elevations, where UV-B
doses are relatively high, were less susceptible to UV-B than
those from lower elevations. Similarly, plants native to high
latitudes are often more sensitive to UV-B injury than those
originating from lower latitudes (Bachelet et al., 1991). Isolates
of the same filamentous fungus, Septoria tritici Roberge, from
geographical regions with contrasting UV-B climates had
different tolerances of UV-B (Rasanayagam et al., 1995). When
the extension of germ-tubes was compared, isolates from
Tunisia, where doses of UV-B are relatively high, were more
tolerant of UV-B than isolates from the U.K. where doses are
relatively low, although a later survey (Paul et al., 1996)
showed significant variation in UV-B response between
isolates from a single site in the U.K.
The maximum mid-day irradiance at Lancaster under clear
sky conditions predicted by the model of Bjo$ rn & Murphy
(1985) is 0±08 W m−# s−", and the maximum integrated dose
is 1±7 kJ m−# d−" UV-BDNA
. The corresponding maximum
value predicted for Sri Lanka is 3±8 kJ m−# d−" UV-BDNA
during March. Most importantly, and supporting the
hypothesis that this work tested, survival and}or colony size
of all isolates are predicted to be affected in vivo, to greater or
lesser extent, by doses of UV-B occurring at their own site of
origin (Table 2). The two U.K. isolates would be differently
affected, so B. alba and Sporidiobolus sp. are predicted to show
11±7 and 3±8% mortality, respectively. Similarly, the three SL
isolates would also show contrasting responses ; Cryptococcus
sp. would show no mortality, while S. salmonicolor and R.
mucilaginosa would show 6±4 and 20±2% mortality, re-
spectively. Colony size would be reduced for both U.K. and
SL isolates at ‘ local ’ doses, but the effect would be greater
with the U.K. isolates than with the SL isolates (Table 2).
None of the yeasts showed total insensitivity to its ‘ local ’
UV-B environment. This suggests there is intra-specific
variation in response and that insensitivity to (or tolerance of)
UV-B probably has a cost, or disadvantage. Thus, natural

UV-B and phylloplane yeasts 784
Table 2. Estimated mortality and reduction of cells per colony (colony
size) for yeasts under UV-B doses representative of their natural habitat
Source Yeast
Dose
(kJ m−# d−"
UV-BDNA
)
Mortality
(%)
Reduction
of cells
colony−"
(%)
SL S. salmonicolor 3±78 6±35 30±80SL Cryptococcus sp. 3±78 0±11 41±65SL R. mucilaginosa 3±78 20±18 45±48U.K. Sporidiobolus sp. 1±7 3±81 54±96U.K. B. alba 1±7 11±72 63±54
Values derived from the dose–response curves (Fig. 1). UV-B doses of
1±7 kJ m−# d−" UV-BDNA
(equivalent to mid-summer Lancaster level under
clear sky) and 3±8 kJ m−# d−" UV-BDNA
(maximum theoretical value for Sri
Lanka) were used for U.K. and Sri Lankan isolates respectively.
populations may be comprised of cells that collectively have
a range of tolerances. The UV-B environment is not uniform;
the more sensitive cells may occupy sites that provide some
protection against radiation, such as depressions between
cells, substomatal cavities, or the base of trichomes.
White light provided relatively little protection for the U.K.
isolate of Sporidiobolus sp. but, excepting R. mucilaginosa, all
yeasts demonstrated decreases in UV-B damage in the
presence of white light, i.e. evidence of photoreactivation or
other light mediated repair mechanisms (PR). The capacity for
PR is not a universal characteristic among yeasts, thus S.
cerevisiae is photoreactivable while Schizosaccharomyces pombe
Lindner is not (Harm, 1980), and may be related to habit, e.g.
leaf or animal surface, as well as taxonomy (the yeast growth
form occurs in taxonomically diverse groups). The absence of
PR in R. mucilaginosa does not prove an absence of that
capacity. Though PR is usually induced by wavelengths
ranging from 310 to 490 nm, some wavelengths may be
particularly effective. For example, PR occurs between 320
and 440 nm with a peak around 375 nm in S. cerevisiae (Jagger,
1985). The quantity and spectral balance of white light
provided here may not have been enough to induce PR in
R. mucilaginosa. The inhibitory effect of white light, in the
presence of UV-B, on R. mucilaginosa is problematical.
However, somewhat similar results have been reported for
human limphoblastoid cells (Tyrell & Amaudruz, 1984), where
UV-A and visible wavelengths sensitized those cells to the
lethal action of UV-B.
Apart from the capacity for PR, tolerance of UV-B may be
related to pigmentation, which for micro-organisms living on
aerial plant surfaces is believed to play a protective function
against injurious solar radiation (Dickinson, 1986). In many
instances, it has been suggested that pigments in fungi are the
main protective mechanism against uv radiation (for example,
Maddison & Manners, 1973 ; Dickinson, 1986 ; Asthana &
Tuveson, 1992). It is not known whether UV-B promotes
carotenoid synthesis in Sporidiobolus sp., but in Rhodotorula
minuta the action spectrum for photoinduced carotenogenesis
has a prominent, broad peak centred on 280 nm (Tada et al.,
1990). Of the two U.K. isolates, B. alba (U.K.) was more
sensitive than Sporidiobolus sp. (U.K.), possibly because B. alba
lacks particular carotenoid pigments possessed by Sporidiobolus
sp. (Simpson et al., 1971). The same relative sensitivities were
noticed in the field when Sporidiobolus sp. and B. alba were
inoculated on young bean leaves and exposed to depleted or
full sunlight (Ayres et al., 1996).
The results show that spatial and temporal variations in
current UV-B doses may have a major role in regulating
populations. The ramifications of climate change, in which
UV-B doses are increased, are intriguing because these micro-
organisms typically grow in microbial communities and often
interact with each other. Thus, since B. alba is more sensitive
than Sporidiobolus sp. to UV-B, a change of UV-B climate
would give B. alba advantage over Sporidiobolus sp. and could
alter the balance between these two organisms which occupy
the same ecological niche. Similarly, since leaf surface yeasts
are known to interact with plant pathogens and affect disease
development in the field (Bashi & Fokkema, 1977 ; Fokkema et
al., 1979 ; Fokkema, 1984), increased doses of UV-B could
affect the balance between saprotrophs and parasites and
influence the incidence of disease in the field.
We thank the Tea Research Institute of Sri Lanka (TSG) and
the U.K. Department of Environment (Contract PECD
7}12}21 : NDP) for financial support.
REFERENCES
Andrews, J. H. & Kenerly, C. M. (1979). Microbial populations associated
with buds and young leaves of apple. Canadian Journal of Botany 58,
847–855.
Asthana, A. & Tuveson, R. W. (1992). Effects of UV and phototoxins on
selected fungal pathogens of citrus. International Journal of Plant Sciences
153, 442–452.
Ayres, P. G., Gunasekera, T. S., Rasanayagam, M. S. & Paul, N. D. (1996).
Effects of UV-B radiation on foliar saprophytes and pathogens. In Fungi and
Environmental Change (ed. J. C. Frankland, N. Magan & G. M. Gadd),
pp. 32–50. Cambridge University Press : Cambridge, U.K.
Bachelet, D., Barnes, P. W., Brown, D. & Brown, M. (1991). Latitudinal and
seasonal variation in calculated ultraviolet-B irradiation for rice-growing
regions of Asia. Photochemistry and Photobiology 54, 411–422.
Bashi, E. & Fokkema, N. J. (1977). Environmental factors limiting growth of
Sporobolomyces roseus, an antagonist of Cochliobolus sativus, on wheat leaves.
Transactions of the British Mycological Society 68, 7–25.
Bjo$ rn, L. O. & Murphy, T. M. (1985). Computer calculation of solar
ultraviolet radiation at ground level. Physiologie VeUgeU tale 23, 555–561.
Campbell, R. (1985). Plant Microbiology. Edward Arnold : London, U.K.
Dickinson, C. H. (1986). Adaptation of microorganisms to climatic conditions
affecting aerial plant surfaces. In Microbiology of the Phyllosphere (ed. N. J.
Fokkema & J. van den Heuvel), pp. 77–100. Cambridge University Press :
Cambridge, U.K.
Dix, J. N. & Webster, J. (1995). Fungal Ecology. Chapman & Hall : London, U.K.
Fokkema, N. J. (1984). Competition for endogenous and exogenous nutrients
between Sporobolomyces roseus and Cochliobolus sativus. Canadian Journal of
Botany 62, 2463–2468.
Fokkema, N. J., Houter, J. G., Kosterman, Y. J. C. & Nelis, A. L. (1979).
Manipulation of yeasts on field grown wheat leaves and their antagonistic
effect on Cochliobolus sativus and Septoria nodorum. Transactions of the British
Mycological Society 72, 19–29.
Gunasekera, T. S. (1996). Effect of UV-B (290–320 nm) radiation on
microorganisms on the leaf surface. Ph.D. Thesis, Lancaster University,
Lancaster, U.K.
Harm, W. (1980). Biological Effects of Ultraviolet Radiation. Cambridge
University Press : Cambridge, U.K.
Jagger, J. (1985). Solar UV Actions on Living Cells. Praeger Publishers : New
York, U.S.A.
James, A. P. & Nasim, A. (1987). Effects of radiation on yeasts. In The Yeasts
(ed. A. H. Rose & J. S. Harrison), pp. 73–97. Academic Press : London, U.K.

T. S. Gunasekera, N. D. Paul and P. G. Ayres 785
Kiefer, J., Schall, A. & Talibi, A. (1986). Physiological responses of yeast cells
to UV of different wavelengths. In Stratospheric Ozone Reduction, Solar
Ultraviolet Radiation and Plant Life : NATO ASI Series Vol. G8 (ed. R. C.
Worrest & M. M. Caldwell), pp. 151–159. Springer-Verlag : Berlin,
Germany.
Last, F. T. & Price, D. (1969). Yeasts associated with living plants and their
environs. In The Yeasts, vol. I (ed. A. H. Rose & J. S. Harrison), pp. 183–218.
Academic Press : London, U.K.
Lindow, S. E. (1991). Determinants of epiphytic fitness in bacteria. In Microbial
Ecology of Leaves (ed. J. H. Andrew & S. S. Hirano), pp. 295–314. Springer-
Verlag : New York, U.S.A.
Maddison, A. C. & Manners, J. G. (1973). Lethal effects of artificial ultraviolet
radiation on cereal rust uredospores. Transactions of the British Mycological
Society 60, 471–494.
Paul, N. D., Rasanayagam, M. S., Moody, S. A., Hatcher, P. E. & Ayres, P. G.
(1996). The role of interactions between trophic levels in determining the
effects of UV-B on terrestrial ecosystems. Vegetatio 128, 1–13.
Pennycook, S. R. & Newhook, F. J. (1982). Ultraviolet sterilization in
phylloplane studies. Transactions of the British Mycological Society 78,
360–361.
Rasanayagam, M. S., Paul, N. D., Royle, D. J. & Ayres, P. G. (1995). Variation
in responses of spores of Septoria tritici and S. nodorum to UV-B irradiation.
Mycological Research 99, 1371–1377.
(Accepted 8 November 1996)
Setlow, R. B. (1974). The wavelengths in sunlight effective in producing skin
cancer : a theoretical analysis. Proceedings of the National Academy of Sciences,
U.S.A. 71, 3363–3366.
Simpson, K. L., Chichester, C. O. & Phaff, H. J. (1971). Carotenoid pigments
of yeasts. In The Yeasts : Physiology and Biochemistry of Yeasts, Vol. II (ed.
A. H. Rose & J. S. Harrison), pp. 493–516. Academic Press : London, U.K.
Sullivan, J. H. & Teramura, A. H. (1988). Effects of ultraviolet-B irradiation on
seedling growth in the Pinaceae. American Journal of Botany 75, 225–230.
Tada, M., Watanabe, M. & Tada, Y. (1990). Mechanism of photoregulated
carotenogenesis in Rhodotorula minuta. VII. Action spectrum for photo-
induced carotenogenesis. Plant Cell Physiology 31, 241–246.
Thompson, I. P., Bailey, M. J., Fenlon, J. S., Fermor, T. R., Lilley, A. K., Lynch,
J. M., McCormack, P. J., McQuilken, M. P., Purdy, K. J. & Rainey, P. B.
(1993). Quantitative and qualitative seasonal changes in the microbial
community from the phyllosphere of sugarbeet (Beta vulgaris). Plant and Soil
150, 177–191.
Tyrrell, R. M. & Amaudruz, F. A. (1984). Alpha polymerase involvement in
excision repair of damage induced by solar radiation of defined wavelengths
in human fibroblasts. Photochemistry and Photobiology 40, 449–452.
Zo$ lzer, F. & Kiffer, J. (1983). Wavelength dependencies of inactivation and
mutagenesis in haploid yeast cells of different sensitivities. Photochemistry
and Photobiology 37, 39–48.