Embed Size (px)
Citation preview

Exp Brain Res (1981) 41:199-215 Experimental Brain Research �9 Springer-Verlag 1981
Restriction of Visual Experience to a Single Orientation Affects the Organization of Orientation Columns in Cat Visual Cortex* **
A Study with Deoxyglucose
W. Singer, B. Freeman 1, and J. Rauschecker
Max-Planck-Institute for Psychiatry, Kraepelinstr. 2, D-8000 Munich 40, Federal Repubfic of Germany
Summary and Conclusions
In six dark reared, 4-weak-old kittens visual experi- ence was restricted to contours of a single orienta- tion, horizontal or vertical, using cylindrical lenses. Subsequently, the deoxyglucose method was used to determine whether these artificial raising conditions had affected the development of orientation columns in the visual cortex. After application of the deoxy- glucose pulse one hemifield was stimulated with vertical, the other with horizontal contours. Thus, from interhemispheric comparison, changes in co- lumnar systems corresponding to experienced and inexperienced orientations could be determined. The following results were obtained: (1) Irrespective of the restrictions in visual experience, orientation col- umns develop in areas 17, 18, 19 and in the visual areas of the posterior suprasylvian sulcus. (2) Within area 17, spacing between columns encoding the same orientations is remarkably regular (1 ram), is not influenced by selective experience and shows only slight interindividual variation. (3) In non-striate areas the spacing of columns is less regular and the spatial frequency of the periodicity is lower. (4) The modifiability of this columnar pattern by selective experience is small within the granular layer of striate cortex but substantial in non-granular layers: Within layer IV columns whose preference corresponds to the experienced orientation are wider and more active than those encoding the orthogonal orienta- tion but the columnar grid remains basically unal- tered. Outside layer IV the columnar system is maintained only for columns encoding the experi-
* This work has partially been supported by a grant from the Deutsche Forsehungsgemeinschaft, SFB 50, A14
** Dedicated to Prof. D. Ploog on the occasion of his 60th anniversary
1 Research Fellow of the Alexander-yon Humboldt-Stiftung Offprint requests to: PD Dr. W. Singer (address see above)
enced orientations. The deprived columns by con- trast frequently fail to extend into non-granular layers and remain confined to the vicinity of layer IV. (5) These modifications in the columnar arrangement are more pronounced in striate cortex than in non- striate visual areas and, within the former, more conspicuous in the central than in the peripheral representation of the visual field. It is concluded that within layer IV the blue print for the system of orientation columns is determined by genetic instruc- tions: first order cells in layer IV develop orientation selectivity irrespective of experience whereby the preference for a particular orientation is predeter- mined by the position in the columnar grid. Depen- dent on experience is, however, the expansion of the columnar system from layer IV into non-granular layers. It is argued that all distortions following selective rearing can be accounted for by competitive interactions between intracortical pathways, the mechanisms being identical to those established for competitive processes in the domain of ocular domi- nance columns. It is proposed that such experience dependent modifiability of connections between first and second order cells is a necessary prerequisite for the development of orientation selectivity in cells with large and complex receptive fields.
Key words: Visual cortex - Development - Orienta- tion columns - Deoxyglucose
It is a common feature of the functional architecture of the mammalian visual cortex that neurons with similar orientation preferences are grouped together in orientation columns; these are sheet-like bands which extend orthogonally to the pial surface through all cortical layers (Hubel and Wiesel 1962, 1963a, b,
0014-4819/81/0041/0199/$ 3.40

200 W. Singer et al.: Development of Orientation Columns
1974a; Creutzfeldt et al. 1974; Albus 1975b, 1979; Hubel et al. 1978). When a large sample of cortical units is recorded along trajectories crossing a suffi- cient number of such columns all orientations are found to be represented by about the same number of neurons (Hubel and Wiesel 1962; Henry et al. 1974; Albus 1975b).
Developmental studies of kittens whose visual experience was restricted to a narrow range of visible orientations have revealed that this normal distribu- tion of orientation preferences may become dis- torted: after selective exposure to a single orientation neurons with corresponding orientation preferences are encountered more frequently than cells prefer- ring the inexperienced orientations (Hirsch and Spinelli 1970, 1971; Blakemore and Cooper 1970; Freeman and Pettigrew 1973; Pettigrew et al. 1973; Tretter et al. 1975a; Blasdel et al. 1977; Singer 1976; Cynader and Mitchell 1977; Stryker et al. 1978; Rauschecker 1979; Rauschecker and Singer 1979, 1981). While the reproducibility of this phenomenon is now commonly accepted its neuronal basis has been and to some extent still is subject to alternative interpretations. As outlined in a review by Barlow (1975) either instructive or selective processes have been favored in most of the reports. More recently, however, it has been argued that these two interpre- tations need not be mutually exclusive (Rauschecker and Singer 1981). To further substantiate this conclu- sion and to localize more precisely the site of adaptive changes within striate cortex, we felt it necessary to analyze the topological rather than the numerical distribution of cells with a particular orientation preference.
As demonstrated by Hubel et al. (1977) in the monkey, by Skeen et al. (1978) in the tree shrew and by Albns (1979) in the cat visual cortex a comprehen- sive representation of the spatial distribution of neurons with a particular orientation preference can be obtained with the 14C-deoxyglucose method of Sokoloff et al. (1977). In the present study we have applied this technique to investigate the effect of selective experience on the spatial organization of orientation columns. While this work was in pro- gress, a short note by Flood and Coleman (1979) appeared in which they report of one kitten that had been studied with a similar experimental design.
Material and Methods
Raising Procedures
To relate the present morphological data to previous electrophy- siologicaI results, raising conditions were identical to those applied in prior physiological studies (Rauschecker and Singer 1979,
Table 1
Kitten Exposure age and weight label dosages No. (h) (weeks) (g) (~c/kg)
at experiment
H1 130 vert. 7 780 H 3 160 H2 130 horiz. 7 780 H 3 160 C1 143 vert. 6 700 C 14 120 C2 170 horiz. 9 1000 C 14 180 C3 210 vert. 7 800 C 14 200 C4 249 horiz. 8 840 C TM 200
1981). Six kittens were dark reared from the time of eye opening to 4 weeks of age. Subsequently, they were exposed selectively either to vertical or horizontal contours. Restriction of contour vision was assured by cylindrical lenses that had -25 dptr in one axis and 0 dptr refractive power in the orthogonal axis. As documented previously (Ranschecker and Singer 1981) such lenses render contours invisible whose orientation differs by more than + 10 deg from the 0 dptr axis. The lenses had a diameter of 2 cm and were positioned in front of the two eyes by means of individually moulded foam helmets: with the eyes in the null-position this arrangement allowed for binocular vision throughout the binocular visual field; only the most lateral parts of the monocular segment may have been occluded by the helmet. Special care was taken to assure that the axes of the two lenses were parallel. Since each kitten had its own helmet, any small (< 5 deg) differences between the orientation of the two lenses were constant throughout exposure. The kittens wore these helmets for an average of 12 h/ day and for the remaining time they were kept in the dark room. For exposure the kittens were kept in small groups in a large (1.5 x 1.5 x 1 m), well lit box, whose walls were painted with high contrast contours of all orientations. Apart from small counter rotations of the eyes the visible orientations were thus always in the same range relative to the retinal coordinates, irrespective of eye, head and body position. Total exposure time and the age of the kittens at the time of the experiment are summarized in Table 1.
As documented previously (Rauschecker and Singer 1981) this raising procedure leads to a massive distortion in the orienta- tion distribution of cortical units even after 100 h of selective experience. Cortical units maintain binocularity indicating that any interocular differences in the alignment of the lenses do not prevent binocular vision.
Tracer Application and Stimulation
For surgery the kittens were anesthetized with a 2 : 1 mixture of Ketanest and Rompun (0.6 ml/kg) i.m. After tracheotomy and insertion of a venous catheter the kittens were placed in a stereotactic head-holder, artificially respirated with a mixture of 70% N20/30% 02, and subsequently paralyzed with a continuous infusion of Flaxedil (20 mg/kg/h). Anesthesia was maintained by continuous i.v. application of Nembutal 2 mg/kg/h. As for elec- trophysiological recording, body temperature, ECG, EEG, and (CO2) in the expired air were continuously monitored. T o facilitate rapid access to the brain after the end of visual stimulation, a circular fissure was drilled along the horizontal circumference of the skull leaving the bone flap in its position until the end of the exposure.
The nictitating membranes were retracted with Neosyne- phrine and the pupils dilated with Atropin. The corneae were protected with black contact lenses containing artificial pupils of

W. Singer et al.: Development of Orientation Columns 201
2 mm diameter. The refractive power of the eyes was determined with a Rodenstock refractometer and adjusted with lenses so that the eyes were focussed on a tangent screen positioned 57 cm in front of the kitten. With the aid of a fundus camera the retinal landmarks (optic disc and area centralis) were mapped onto the tangent screen; with a hook attached to the insertion of the lateral rectus of one eye the visual axes were made convergent and adjusted until the areae centrales were superimposed on the screen. To assure non-overlapping stimulation of the right and left visual hemifields the vertical meridian was covered with a black stripe extending about 1 deg into each hemifield.
Approximately 2 h after the initiation of anesthesia visual stimulation was started. The right and left hemifields were stimulated with vertically and horizontally oriented gratings, respectively. Stimuli were provided by two identical electro- mechanical devices and consisted of square wave gratings whose spatial frequency varied continuously. These gratings were moved up and down or temperonasaUy so that at each instant the whole hemifield was covered with contours. The speed of the moving pattern was alternated every 10 min between 2 deg/s and 10 deg/s. Five minutes after the onset of stimulation tritium-labeled deoxy- glucose was administered i.v. in two kittens and C14-1abeled deoxyglucose in four kittens. The dosages are summarized in Table 1. Fifty minutes after the deoxyglucose pulse an overdose of Nembutal was given i.v., the prepared bone flap and the dura were removed and five blocks were cut containing the occipital poles, the lateral geniculate bodies and the tectum. These blocks were frozen directly onto the cryostat heads which were cooled to -80 ~ C. Subsequently, 30 Ixm thick serial horizontal sections of the visual cortices were cut at -19 ~ C. The sections from the brains containing tritium-labeled deoxyghicose were mounted on slides that were precoated with Kodak nuclear track emulsion (NTB3) whereas the sections containing C14 label were mounted on normal slides. On the precoated slides the sections were either attached by gentle pressure only or by brief melting and rapid refreezing on solid carbon dioxide plates. Initially, some of the C14 sections were stretched by pressure only, the others were attached either by melting and drying on a hot plate (90 ~ C) or by melting on the thumb and rapid refreezing on carbon dioxide. For most of the sections the latter method was applied since the liquid phase was shorter (about 6 s) than on the hot plate and tissue preservation seemed better, bubbles occurring less frequently. Attachment by pressure only was abandoned since spatial resolu- tion of colunms was little, if at all, better than with melting but tissue preservation was poor. All slides were then stored at -60 ~ C and freeze-dried for several days. The C14-1abeled sections were exposed at -60 ~ C on Agfa Mammo-ray T3 film. To facilitate comparison between the two hemispheres the sections were collated such that corresponding sections from the two hemi- spheres were exposed on the same film sheet. Optimal exposure time was assessed separately for each brain from specially pre- pared samples and ranged between 3 and 4 weeks. For later correlation between the distribution of radioactivity and cortical anatomy, all sections were NissI-stained after fixation in alcohol.
Image Processing and Quantitative Evaluation
The activity patterns on the X-ray films were processed in two ways. Firstly, corresponding sections from the two hemispheres were selected and each of the four or five serial sections contained on one slide was copied on a high resolution film (Dupont- Graphic-Arts CBS-4) with a magnification factor of 6.5. Care was taken that slides which were to be used for interhemispheric comparison had been exposed on the same X-ray film and were also copied on the same film sheet. Using a dissecting microscope the magnified negatives of these three or four serial sections were
carefully superimposed using cortical landmarks for registration and contact copies were made from these montages. This averag- ing procedure greatly increased contrast and reduced the noise introduced by freezing artifacts in individual sections. Because the activity levels in the two hemispheres turned out to be rather different, the contact prints of the montages were made at systematically varied exposure times.
In the second method, the image analysis was performed directly on single autoradiographs using the computer facilities at the Technical University of Munich, which were generously made available to us by Dr. Platzer from the Dept. of Optical and Digital Image Processing. With a highly linear scanning system the density distributions on the X-ray films were coded in digital units, ranging from 1 to 1024. Spatial resolution was adjusted such that a digital value was stored for squares with a side length of 40 x 20 ~tm. All data were stored on digital tape for later quantitative processing with a PDP 11/40 to provide hardcopy reconstructions of (i) the original density distribution and (ii) the density distribution obtained with up to 19 different intensity thresholds in the range from 0 to 1023.
The average width of columns and the distance between them were measured with a microscope directly from the original X-ray patterns. For the determination of columnar spacing the distances between the central, most active zones of adjacent columns were assessed. The width of individual columns was defined as the distance between the borders within which activity was clearly identifiable as being above background. Within layer IV demarka- tion of active zones from background was sufficiently sharp to allow for a reliable determination of columnar width. This is indicated by the small standard deviations of the mean values and by the fact that different samples from the same hemisphere were never significantly different. These quantitative estimates, how- ever, had to be restricted to layer IV; firstly, because in the hemisphere stimulated with orientations corresponding to those experienced, the demarkation between adjacent columns was often not sufficiently clear in non-granular layers and, secondly, because in the other hemisphere columns frequently did not extend into these layers at all. Correlation between radioactivity distributions, cortical areas and cortical laminae was done either by mounting the film strips directly onto the corresponding Nissl sections or by the use of camera lucida drawings.
Results
C14-labeled Sections
A f t e r a p p l i c a t i o n o f t h e d e o x y g l u c o s e p u l s e t h e t w o
h a l v e s o f t h e v i s u a l f i e ld w e r e s t i m u l a t e d w i t h c o n -
t o u r s o f d i f f e r e n t o r i e n t a t i o n : h o r i z o n t a l a n d v e r t i c a l .
T h u s , c e r e b r a l g l u c o s e c o n s u m p t i o n is m e a s u r e d
w h i l e o n e h e m i s p h e r e is s t i m u l a t e d t h r o u g h b o t h
eyes w i t h o r i e n t a t i o n s c o r r e s p o n d i n g to t h o s e e x p e r i -
e n c e d d u r i n g e a r l y d e v e l o p m e n t w h i l e t h e o t h e r
h e m i s p h e r e is s t i m u l a t e d w i t h c o n t o u r s t h a t h a v e
n e v e r b e e n s e e n b e f o r e . O f t h e f o u r k i t t e n s i n v e s t i -
g a t e d w i t h C 1 4 - 1 a b e l e d d e o x y g l u c o s e t w o h a d e x p e r i -
e n c e d v e r t i c a l (C1 , C3 ) a n d t w o h o r i z o n t a l c o n t o u r s
(C2 , C4) d u r i n g t h e r a i s i n g p e r i o d . F o r e a c h o f t h e
f o u r k i t t e n s t w o f e a t u r e s w e r e o b v i o u s i n t h e ac t i v i t y
p a t t e r n s f r o m t h e o c c i p i t a l c o r t i c e s . F i r s t l y , o n e
occ ip i t a l c o r t e x a l w a y s h a d m u c h m o r e t o t a l r a d i o a c -
t iv i ty t h a n t h e o t h e r a n d s e c o n d l y , n o t w i t h s t a n d i n g

202 W. Singer et al.: Development of Orientation Columns
Fig. 1. Montages from horizontal serial sections through the medial bank in kitten C2. The montages A31 and B25 are the averages from three successive sections each (see methods) and are taken from a depth of about 4 mm where striate cortex folds to form the superior splenial sulcus. The arrows indicate these tangentially cut parts of area 17. Exposure times for the contact prints are indicated below the individual copies. Raising condition and stimulus orientation seen by the two hemispheres during the experiment are indicated by the gratings on top, A31 and B25 corresponding to the adequately and inadequately stimulated hemisphere, respectively. MB = medial bank, LS = lateral sulcus, PSS = posterior suprasylvian sulcus. Note the decreased overall activity and the better contrast of columns in area 17 of the inadequately stimulated hemisphere
this asymmetry in activity, both cortices showed clear evidence of a columnar organization. We discuss the columnar organization first.
Columnar Organization Common to Both Hemispheres
The basic patterns of the columnar organization of radioactivity were strikingly similar in all four kit-
tens. In accordance with the results obtained by Albus (1979) in normally raised cats, patchy activity distributions were commonly observed not only in striate cortex but also in areas 18, 19, and in the visual areas of the posterior suprasylvian sulcus (Figs. 1, 3-7). Within striate cortex the spacing of clusters with increased activity was found to be surprisingly regular with a peak-to-peak periodicity of about 1 mm (see below). Within area 18 and even more so
Fig. 2. Montages from tangential sections through supragranular layers of areas 17 and 18 of kitten C3. For each of the four prints three successive serial sections were superimposed. Sections A2/B2 and A3/B3 are from a depth of about 500 vm and 800 vm, respectively. Raising and experimental stimulus conditions as well as exposure times of individual contact prints are indicated as in Fig. 1. Here, sections A correspond to the inadequately and sections B to the adequately stimulated hemisphere. The Nissl-stained sections immediately precede those used for montages A2/B2. Note the increase of activity at the transition zone from area 17 to area 18 (arrows in A2/A3). For further description see text

W. Singer et al.: Development of Orientation Columns 203
204 W. Singer et al.: Development of Orientation Columns
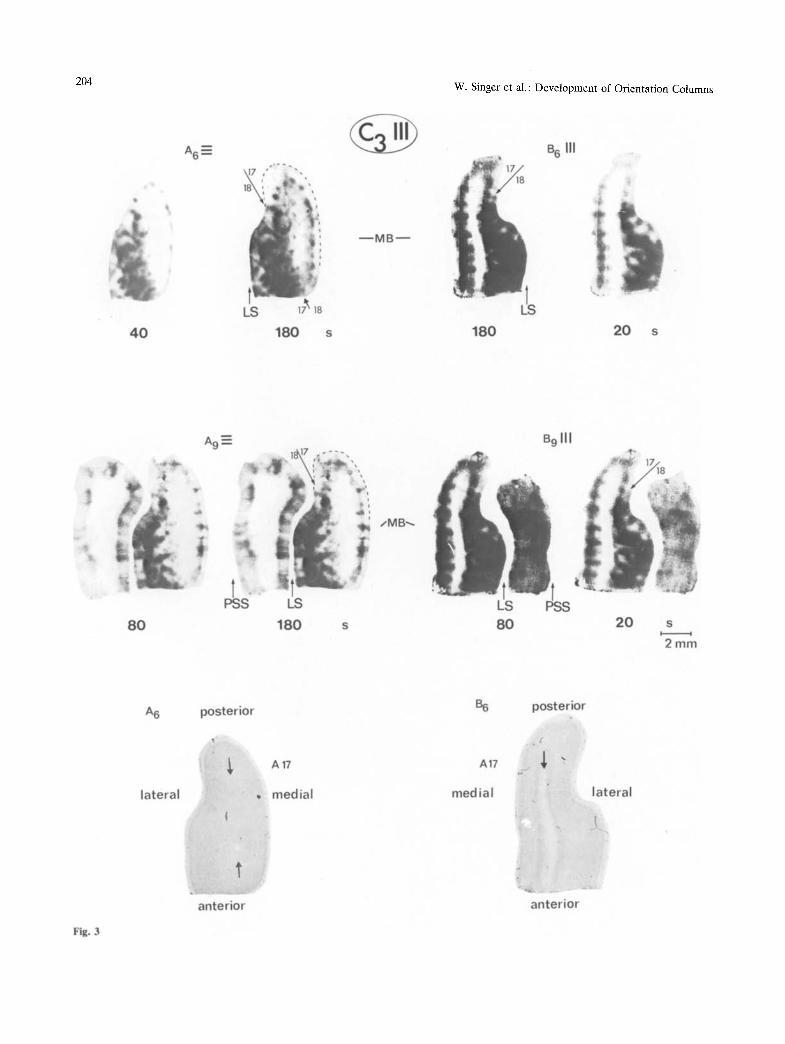
Fig. 3

W. Singer et al.: Development of Orientation Columns
in the other visual areas, this columnar arrangement becomes less regular and the width of both active and inactive areas increases markedly. The columnar pattern in non-striate visual areas is distinguishable particularly well in the hemispheres which were stimulated with the orientation that was orthogonal to the experienced one. Examples for the columnar arrangement in area 18 are shown in Fig. 2 (A2, A3) and in Fig. 3 (A6), which represent sections that run tangential to the cortical lamination. Columns in area 19 and in the visual areas of the posterior suprasyl- vian sulcuscan be seenbest in Fig. 3 (A9) and in Fig. 4 (Al l , A20) showing sections from deeper parts of the cortical areas bordering the lateral and the posterior suprasylvian sulcus.
Apart from these common features of the colum- nar organization within the visual cortices there were, however, numerous and marked differences between the two hemispheres, indicating that selec- tive visual experience had led to changes in the organization of orientation columns.
Interhemispheric Differences
Activity Levels. In all kittens, interhemispheric com- parison of the average activity level of the occipital cortex showed that it was consistently higher in that hemisphere which was stimulated with the same orientation as the orientation to which it had been exposed during the raising period. Henceforth, this kind of stimulation will be referred to as "adequate" stimulation for that hemisphere and conversely, the stimulation of a hemisphere with contours to which the hemisphere had never been exposed, will be described as "inadequate". As exemplified in Figs. 1-7, interhemispheric differences in overall activity are clearly visible both in the photographic montages and in the individual computer processed sections. It can further b e seen that these interhemispheric activity differences are more pronounced in area 17 than in the other visual areas. This is mainly because the activity in the inadequately stimulated area 17 is particularly low when compared to other visual
205
areas. On the adequately stimulated side, activity levels are approximately the same over the various visual areas while on the inadequately stimulated side they are much lower over area 17 than over non- striate areas (Fig. 4 (Al l ; A20) and Fig. 5 (B10, B20)). As can be seen particularly well in Fig~ 2 (A2) but also in must of the other figures, the 17/18 border on the inadequately stimulated side, therefore, appears as a demarkation line between zones of particularly low and medium activity. From the computer processed sections these interhemispheric differences in overall activity can be assessed in a quantitative way. As exemplified in Figs. 6 and 7 for kittens C3 and C4, the activity in most parts of the inadequately stimulated area 17 is below 250 relative units, the scale ranging from 1 to 1023 U. Only in very circumscribed patches, peak activities between 550 and 700 U are reached. In corresponding sections of the contralateral, adequately stimulated striate cortex, the activity over most of the area is in the range from 550 to 700 U and there exist rather large patches where the activity exceeds the level of 700 units. In non-striate visual areas of the inadequately stimulated hemisphere activity is commonly exceed- ing 250 U, reaching peak values between 400 and 550 U. Over non-striate areas of the adequately stimulated hemisphere the activity is in the same range as over striate cortex of the same side (from 550 to more than 700 U)o
Related to this difference in overall activity is the finding that in all four kittens the columnar structure is visible much more clearly in the inadequately stimulated hemisphere than in the adequately stimu- lated one. This trend is apparent both for the central (Fig. 2, Fig. 3 (A6/B6), and Fig. 6 (A2/B2)) and the peripheral (Figs. 1, 4, 5, 7) representations of the visual field. Quantitative evaluation of computer processed sections suggests as the main reason for the blurring of the columnar structure in the adequately stimulated hemisphere the increased activity in the troughs between the dusters of high activity. As indicated by test slides exposed for shorter periods of time, this reduction in contrast could not be compen- sated for by shorter exposure.
Fig. 3. Montages of horizontal sections through the superficial regions of the occipital cortex of kitten C3. For each of the four prints three successive serial sections were superimposed. The sections A6/B6 are from a depth of 1.8 mm. In their central part they are tangential to the transition zone between layer VI and white matter (see arrows in corresponding Nissl stained sections below ). Sections Ag/B9 are 400 ~m further down and now the posterior pole of the suprasylvian gyrus is also contained in the sections. Raising and stimulus conditions as well as exposure times of the Contact prints are indicated as in the previous figures. Sections A are from the inadequately and sections B from the adequately stimulated hemisphere. The beaded band of activity in A17 of A6 and A9 corresponds to layer IV. The 17/18 boundaries are again indicated by arrows. Note the incomplete translaminar extent of columns in A17 of the inadequately stimulated side (A6/A9) and the trend for columns on the other side (B6/B9) to merge with each other in supragranular layers. In all sections circumscribed patches and bands of increased activity are distinguishable also over area 18. In sections A9 columns of increased activity extending throughout all laminae are apparent over the lateral bank of the lateral sulcus (LS) and over the medial bank of the posterior suprasylvian sulcus (PSS)
206 W. Singer et al.: Deve lopment of Orientat ion Columns
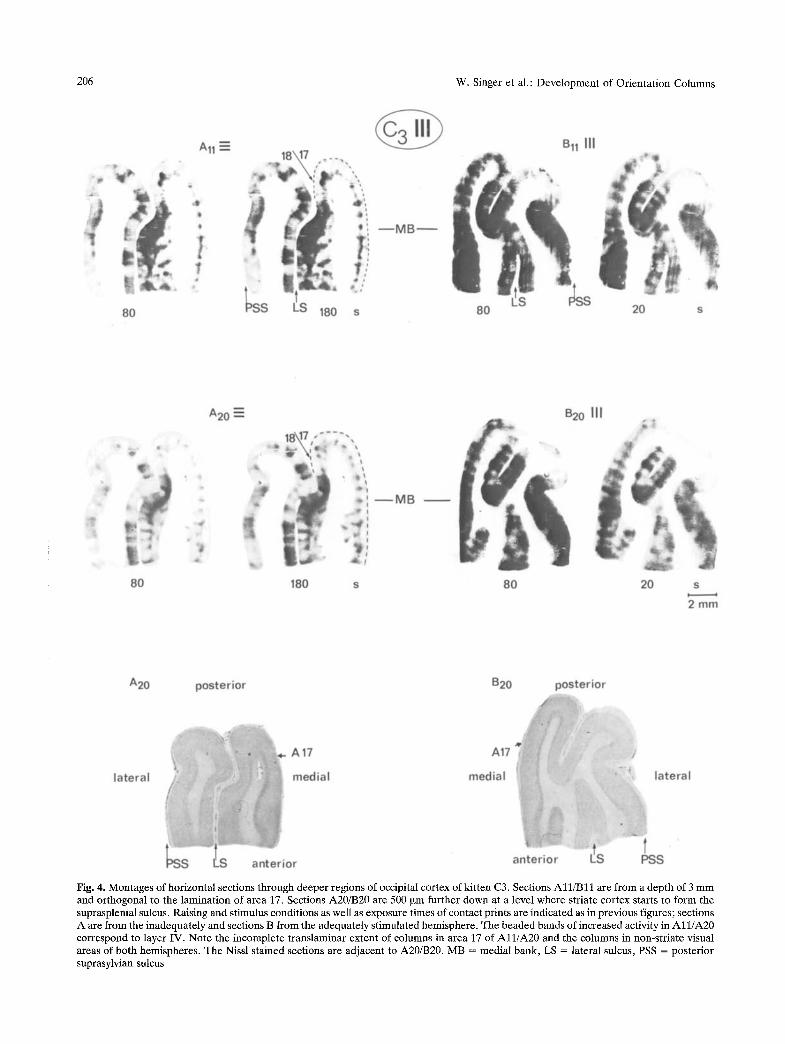
Fig. 4. Montages of horizontal sections through deeper regions of occipital cortex of kitten C3. Sections A l l / B l l are f rom a depth of 3 m m and orthogonal to the laminat ion of area 17. Sections A20/B20 are 500 ~m further down at a level where striate cortex starts to form the suprasplenial sulcus. Raising and st imulus conditions as well as exposure t imes of contact prints are indicated as in previous figures; sections A are from the inadequately and sections B from the adequately s t imulated hemisphere . The beaded bands of increased activity in A l l / A 2 0 correspond to layer IV. Note the incomplete t ranslaminar extent of columns in area 17 of A l l / A 2 0 and the columns in non-str iate visual areas of both hemispheres . The Nissl s tained sections are adjacent to A20/B20. MB = medial bank, LS ~ lateral sulcus, PSS ~ posterior suprasylvian sulcus

W. Singer et al.: Development of Orientation Columns 207
Fig. 5. Montages from horizontal serial sections through occipital cortex of kitten C4. For each of the four prints three successive serial sections were superimposed. Sections All/B10 and A16/B20 are from a depth of 2 and 3 mm from the dorsal convexity of striate cortex. Raising and stimulation conditions as well as exposure times of the individual contact prints are indicated as in previous figures; sections A are from the adequately and sections B from the inadequately stimulated hemisphere. The Nissl-stained sections correspond to montages A16/B20. Note that the incomplete translaminar extent of columns on the inadequately stimulated hemisphere is more pronounced in superficial (B10) than in deep (B20) sections. Further description is given in the text

208 W. Singer et al.: Development of Orientation Columns
Fig. 6. Computer-processed activity distributions in individual horizontal sections through the occipital cortex of kitten C3. Sections A2/B2 and Al l /Bl l are adjacent to the respective sections used for the montages in Figs. 2 and 4. For technical reasons the computer reproductions are mirror images of the direct prints and in addition, dark zones on the X-ray film appear as bright zones. A2/B2 run tangentially through supragranular layers. A and B correspond to adequately and inadequately stimulated hemispheres, respectively. The left pair A2/B2 shows the hard copy reconstruction of the digitalized activity distributions. The fight pair A2/B2 and the bottom pair A11/ B l l are reconstructions following a threshold operation in which 6 activity ranges were defined and associated with different gray levels (scale on left margin of bottom pair). MB = medial bank, LS = lateral sulcns. Further explanation is given in the text
Width and Spacing of Columns Within Layer IV. In kit tens C3 and C4, where the co lumnar organiza t ion was par t icular ly well d is t inguishable , a statistical analysis was carr ied out on the dis tance be t w e e n columns and their widths. Because of the i r regular extent of co lumns into n o n - g r a n u l a r layers (see
below) all m e a s u r e m e n t s were restr icted to layer IV, selecting sections that were o r thogona l to the cortical laminat ion . F r o m each hemisphe re two samples of 6-8 successive sections were analyzed. These samples were taken at a dep th of 1.8 m m and again at 3 m m from the dorsal crest of the lateral gyrus.

W. Singer et al.: Development of Orientation Columns 209
Fig. 7. Computer-processed activity profiles in individual horizontal sections from kitten C4. Sections A10/B11 are from a depth of 2 mm and are orthogonal to the cortical lamination of striate cortex in the medial bank of the inadequately (A10) and adequately (Bll) stimulated hemisphere, respectively, The same six thresholds as in Fig. 6 were used (scale on left margin)
Table 2
Kitten Hemisphere No. of No. of distance between width of columns sections columns columns (~tm) (~tm)
14 71 adequately C3 ~ ~ ' ~ s t i m u l a t e d
~ inadequaely stimulated
12 75
adequately 14 73 C4 ~ . . ~ ' ~ s t i m u l a t e d
~ i n a d e q u a t e l y 9 68 stimulated
1015+170q
-1000+ 176 y n . s .
- - r l .S .
- n . s .
-1038-+197-]
_1016+222..J -n's"
5 6 3 - + 9 4 ;
396-85 -a p<O.05
681_+15;
432+87_7 p<0.05
The width of columns and the interpeak distances were strikingly similar in different samples taken at various dep ths from the s a m e hemisphere. In the one-tailed t-test, all p-values for this intrahemispheric comparison were above 20% and so for Table 2 all data from a particular hemisphere were pooled.
It can be seen that the columnar spacing is remarkably constant showing neither interhemi- spheric nor interindividual differences. Clear interhemispheric differences (p < 0.05) were, how- ever, noted for the width of the columns, active zones being broader on the adequately than on the inade-

210 W. Singer et al.: Development of Orientation Columns
quately stimulated sides. These interhemispheric dif- ferences remained significant (p < 0.05) even when the data from the two cats were pooled, in spite of the fact that the average width of the active zones was slightly larger in C4 than in C3. Laminar Extent of Columns. In the adequately stimulated hemispheres columns of increased activity consistently extended throughout all layers (Fig. 3 (B6)) as is the case in cats raised with normal visual experience (Albus 1979). In addition, we noted in numerous sections that the blurring of columnar boundaries, which is characteristic for the adequately stimulated hemispheres, tended to be particularly pronounced within layer IV and supragranular layers (Fig. 3 (B6, B9)). In area 17 of the inadequately stimulated hemispheres, by contrast, columns only rarely extended throughout all layers. As exemplified in the sections running orthogonal to the lamination of area 17 (Figs. 3-7) patches of increased activity are confined essentially to layer IV and the immediately adjacent parts of layers III and V. Only occasionally narrow bands extend from these islands of activity towards more superficial and deeper layers whereby their width and density decrease with increasing distance from layer IV (Fig. 3 (A6/A9), Fig. 4 (Al l / A20) and Fig. 5 (B10)).
The interhemispheric differences in the trans- laminar extent of columns are obvious also in tangen- tial sections running preferentially through supra- granular (Fig. 2 and Fig. 6 (A2, B2)) or infragranular layers (Fig. 3 A6/B6). In superficial layers of the adequately stimulated side columns appear as con- fluent, poorly delineated bands; in the other hemi- sphere they are reduced to small isolated dots, which correspond to the peaks of the cone shaped activity zones extending from layer IV into supragranular layers. Consistent with an incomplete extent of columns into supragranular layers is also the finding that in tangential sections from the inadequately stimulated hemisphere a regular columnar arrange- ment was consistently apparent only at depths bey- ond 500 ~tm, while on the other side patches and bands were already visible at a depth of 200 ~m. In section A6 of Fig. 3, the central part is tangential to infragranular layers V and VI. It can be seen that this region is nearly as inactive as the white matter in section B6 supporting the evidence for an incomplete extent of columns also within infragranular layers.
Within layer IV of both hemispheres there is furthermore an indication of a band of increased activity which runs parallel to the cortical surface. This band is visible best in sections from the inade- quately stimulated hemispheres because of the low activity in adjacent non-granular layers (Fig. 3 (A9), Fig. 4 (All)) .
Comparison Between Central and Peripheral Visual Field Representation
Tangential sections through the representation area of the central visual field as well as orthogonal sections taken from the superficial parts of the medial bank of the inadequately stimulated hemispheres have indicated that in these regions columns tend to be restricted to the vicinity of the granular layer. In sections taken from the medial bank at depths beyond 3 mm, however, this tendency decreases. In these deep sections most patches of increased activity extend from layer IV throughout layers III and II. They are, however, only rarely in continuity with a second row of active zones which is in register with the superficial patches and located in layer VI (Fig. 1 (B25) and Fig. 5 (B20)).
A decrease in interhemispheric differences at greater eccentricities is also apparent for the overall level of activity in A 17 and the activity gradient between striate and extrastriate cortex. The marked reduction of activity in A 17 on the inadequately stimulated side and hence the differences between striate and non striate areas were less pronounced in deeper parts of the medial bank than in the more superficial sections (cf. Fig. 1, B25 and Fig. 5, B20 with Figs. 3 and 4). These results are thus suggesting that the representation of the central visual field is more severely affected by restricted experience than the representation of the periphery.
Comparison Between A 17 and Other Visual Areas
In the non-striate areas the columnar systems appear to~'have a less orderly arrangement, active regions being broader, and less regularly spaced than in area 17 (Figs. 4 and 5). Also, in these areas overall activity is always lower on the inadequately stimulated side and columns tend to be narrower and better segre- gated from each other than in the adequately stimu- lated hemisphere. These differences are illustrated for two cats by the contact prints in Figs. 4 and 5 and by the quantified activity distributions in Fig. 6 (Al l , B l l ) and Fig. 7. Although no quantitative evaluation was attempted because of the difficulty of defining the borders of the areas to be sampled, two observa- tions indicate that interhemispheric differences are consistently more marked in the striate cortex than in the other visual areas. In the adequately stimulated hemisphere overall activity levels are high and similar in the various areas. On the inadequately stimulated side overall activity is lower but in addition there is the marked gradient between striate and non-striate areas, striate cortex being considerably less active

W. Singer et al.: Development of Orientation Columns
than the other areas (cf. sections All/B11 in Fig. 6 and sections A10/B11 in Fig. 7). Moreover, as illustrated in Fig. 4 (All/A20), the tendency for an incomplete translaminar extent of columns on the inadequately stimulated side is much less pronounced in non-striate than in striate visual areas.
Tritium-labeled Sections
No columnar organization was detectable in the activity profiles of sections taken from occipital cortex. There were, however, consistent differer~ces in average activity between the visual cortices of the two hemispheres. As in the C14-1abeled sections, activity was always higher over the adequately than over the inadequately stimulated hemisphere.
Discussion
Persistence of Columnar Organization
In all four kittens labeled with C14-deoxyglucose patches of increased activity were distinguishable in all visual areas of both hemispheres. This is in agreement with the notion that, not only in striate but also in non-striate visual areas, neurons with similar response properties are clustered together (Albus 1979; Hubel and Wiesel 1965; Palmer et al. 1978). As one proceeds from areas 18 and 19 toward the visual areas of the posterior suprasylvian sulcus, these clusters become larger and less regularly spaced, suggesting a more complicated mapping of feature domains and visual space in these areas (Tusa et al. 1979)
Within layer IV of area 17 the interpeak distance between these active zones was almost identical in each of the four hemispheres analysed numerically. The modal value of i mm is the same as that found by Albus (1979) in normally reared cats. Although a spacing of about 1 mm is reported also for ocular dominance columns in cat striate cortex (Shatz et al. 1977), the following arguments suggest that the columns in Albus' work and also in the present investigation are actually related to orientation: In Albus' and also in our cats (Rauschecker and Singer 1981) cortical neurons can be assumed to be binocu- lar. Thus, even if interocular inhibition had led to the suppression of responses from one eye ocular domi- nance columns should not be resolvable. Further support for a periodicity of 1 mm in the system of orientation columns comes from a related study in monocularly deprived kittens: here the experienced eye was stimulated with horizontal and vertical
211
contours in alternation and the average peak to peak distance between adjacent columns was found to be precisely 0.5 mm (manuscript in prep.).
Accepting that the columns visualized in the present study are related to orientation preference it can be concluded that in all visual areas examined cells remain responsive to inexperienced orientations and remain grouped together in circumscribed dus- ters. For the cells in layer IV of striate cortex it can even be inferred that they remained orientation selective: Neurons within columns whose preference matches the experienced orientation can be assumed to maintain their initial preference and hence are unlikely to be activated by the novel, orthogonal orientation. The active zones on the inadequately stimulated side must, therefore, result from respon- sive cells whose position within the columnar grid corresponds to inexperienced orientations. Moreover, as suggested by the regularly spaced areas of decreased activity on the adequately stimulated side cells within columns corresponding to inexperi- enced orientations are apparently not capable of responding to the experienced orientations. This indicates that, at least within layer IV, numerous cells within the deprived orientation columns must have remained responsive and even selective for orientations corresponding to their position in the columnar system. Thus, apart from an expansion of columns encoding the experienced orientations rela- tive to the width of the deprived columns, restriction of experience to a single orientation seemed to have no effect on the development of orientation columns within the granular layer. And even in non-granular layers where the effects of restricted experience are much more pronounced, the skeleton of the colum- nar organization is still detectable. This conclusion is in good agreement with electrophysiological studies which have revealed orientation selective cells and systematic shifts of orientation preferences along tangential recording tracks despite restriction or abolition of contour vision (Hubel and Wiesel 1963b; Wiesel and Hubel 1974; Blakemore and Van Sluyters 1975; Imbert and Buisseret 1975; Buisseret and Imbert 1976; Sherk and Stryker 1976; Singer 1976; Singer and Tretter 1976; Fregnac and Imbert 1978; Stryker et al. 1978; Watkins et al. 1978; Rauschecker and Singer 1981).
The continuous band of increased activity that was visible within layer IV of both hemispheres appears to be similar to a band which is seen in layer IV C of the monkey striate cortex where it appears to result from cells lacking orientation selectivity (Hubel et al. 1977a, b, 1978). However, in cat visual cortex there is no electrophysiological evidence for the existence of a large population of non-oriented

212 w. Singer et al.: Development of Orientation Columns
cells within layer IV, although restriction of experi- ence to a single orientation does increase the fraction of these cells encountered (Rauschecker and Singer 1981). Therefore, the possibility must be considered that the continuous activity within layer IV of the cat visual cortex is reflecting the metabolism in the terminals of geniculate afferents.
Modification of Orientation Columns by Selective Experience
Although the blue-print for the arrangement of orientation columns appears to be determined to a large extent by intrinsic factors, the interhemispheric differences apparent in our material indicate modifia- bility by visual experience. This conclusion is in agreement with the data of Flood and Coleman (1979), since in their kitten, the columns in the adequately stimulated hemisphere were broader than on the inadequately stimulated side. The incomplete translaminar extent of the deprived columns as found in our work indicates, in addition, that the malleabil- ity of the columnar system is much more pronounced in non-granular than in granular layers. Electrophy- siological (Toyama et al. 1974; Singer et al. 1975; Tretter et al. 1975b; Mitzdorf and Singer 1978) and anatomical evidence (Szenth~gotai 1973, 1975; Lund et al. 1979) indicates that neurons outside layer IV and in particular in supragranular layers receive their prominent excitatory drive through connections originating from monosynaptically driven cells in layer IV. The incomplete translaminar extent of deprived columns, thus, implies that activity from the responsive first order cells in layer IV is no longer relayed to second order cells in non-granular layers that are located within the same vertical column. As suggested by the following arguments some of the deafferented second order cells now appear to be responsive to the experienced orientation. In the adequately stimulated hemisphere overall activity was high and the segregation of individual columns was poor, in particular within the supragranular layers. Although interindividual comparison of co- lumnar width is fraught with considerable difficulties we believe that this reflects an expansion of ade- quately stimulated columns above the normal width. For comparison we relied on the data from Albus (1979) and also on our own unpublished data from two monocularly deprived cats that were stimulated through the normal eye either with a single orienta- tion or with horizontal and vertical gratings in alternation. The absolute values and the standard deviations for columnar width in Albus' report agree very well with our data from the monocularly de-
prived cats suggesting that the width of normal columns ranges from 300 to 450 ~tm with a standard deviation below 100 ~tm. Accepting these data as a baseline the average width of columns encoding experienced orientations is larger by a factor of between 1.4 and 1.8 while the width of the deprived columns within layer IV is in the same range as the values from normal cats,
Irrespective of these interindividual comparisons, however, the obvious interhemispheric differences warrant the conclusions that restriction of experience to a single orientation leads to marked changes in the cortical volumes reserved for particular orientations. Assuming that the number of cells per cortical volume has remained constant our data certainly imply an absolute decrease of the number of cells responding to inexperienced orientations and suggest an absolute increase in the number of cells responsive to the experienced orientation.
The Mechanism of Changes in the Columnar Arrangement
The marked shrinkage of deprived columns, particu- larly in non-granular layers and the likely expansion of adequately stimulated columns is strikingly similar to the rearrangement in the system of ocular domi- nance that occurs within layer IV after monocular deprivation (Hubel et al. 1977a). This rearrangement is attributed to competitive interactions between afferents from the two eyes, leading to the repression of the deprived pathways. As demonstrated recently (Rauschecker and Singer 1979, 1981) these competi- tive interactions are gated by the responses of the postsynaptic target cells, following rules that closely resemble those proposed by Hebb (1949) for adap- tive synapses: The efficiency of synaptic transmission increases for pathways which have a high probability of being active in contingency with the postsynaptic target cell and decreases for afferents which are inactive while the postsynaptic cell is responding to other inputs. In the absence of postsynaptic responses, however, there is no differential change in transmission and the efficiency of convergent pa th - ways decreases in an unselective way irrespective of presynaptic activity.
In the following argument we propose that the experience-dependent rearrangement of orientation columns can also be accounted for by these same rules: the only difference from the paradigm of monocular deprivation being that, in the present case, competition occurs between pathways converg- ing from cells in layer IV whose orientation selectiv- ity is prespecified onto target cells located both

W. Singer et al.: Development of Orientation Columns 213
within and outside the granular layer. In accepting this, only two conditions have to be assumed in order to account for the present findings by activity depen- dent competition. Firstly, the pathways relaying activity from the orientation selective cells in input layer IV to second order cells ought to be somewhat divergent and overlapping, spanning a distance tangential to the lamination of about one columnar cycle. Secondly, prior to experience this connectivity ought to be rather unselective, following the simple rule that coupling is strongest along lines orthogonal to the cortical lamination and decreases gradually with tangential distance. Both assumptions are com- patible with anatomical data (Szent~gothai 1973, 1975; Fisken et al. 1973; Creutzfeldt et al. 1977; Lund et al. 1979). The ensuing functional implications are (a) that the orientation selectivity of second order cells should initially be broader than that of the already committed first order neurons and (b) that even without experience the orientation preferences of all cells located within the same vertical columns should be identical. This again is in agreement with a large amount of physiological data on the tuning properties and the sequence regularity of orientation- coding cells in visually inexperienced animals (Wiesel and Hubel 1974; Sherk and Stryker 1976; Imbert and Buisseret 1975; Fregnac and Imbert 1978). It is further confirmed by the finding from deoxyglucose experiments in binocularly deprived animals, that orientation columns, although less well delineated outside layer IV, extend throughout all cortical laminae (manuscript in prep.). Accepting the validity of these rather plausible assumptions and of the rules for competitive interactions it can easily be seen that competition at the level of second order cells would lead to the distortions observed in the present study when experience is restricted to a single orientation while the same processes would lead to the consolida- tion of a regular columnar system extending through- out all layers when visual experience is normal. Because of the continuity of natural patterns contin- gency-matching assures that only those pathways remain consolidated on second order target cells which originate from first order cells sharing the same orientation preference. Second order target cells are thus becoming as sharply tuned for orienta- tion as the first order cells although they may still integrate activity not only from cells within the same vertical cylinder but also from more remote layer IV cells. From the size and the positional scatter of receptive fields along lines orthogonal to the cortical surface (Creutzfeldt et al. 1974; Hubel and Wiesel 1974b; Albus 1975a) it can be inferred that such convergence is required for the formation of second order cells with large and complex receptive fields.
The proposed mechanisms could thus serve an important role in normal development by specifying the extremely complex connections between first and second order cortical neurons according to functional criteria, thereby economizing genetic information.
/
Pathophysiological Mechanisms in Amblyopia
In analogy to humans with conatal astigmatism, the present kittens can be considered to suffer from meridional amblyopia (Freeman and Pettigrew 1973). This syndrome develops when contours of a particular orientation are consistently blurred or invisible during a critical period of early develop- ment. The consequences are perceptual disorders that (i) are specific to the range of inappropriately experienced orientations and (ii) cannot be cured with optical means (Mitchell et al. 1973; Mitchell and Wilkinson 1974; Freeman and Thibos 1975a, b). Based on the present results these perceptual disturb- ances can now be related to specific changes in the organization of orientation columns: Within the "deprived" columns of area 17 the transmission of activity from first order neurons in layer IV to second order cells is severely impaired. Since it is mainly the latter cells that mediate the output of striate cortex to other visual centers, substantial disturbances in the processing of contours with corresponding orienta- tion are to be expected. Furthermore, a close similar- ity exists between the present results and recent electrophysiological data from strabismic cats which suffered from squint amblyopia. In these animals, too, there is evidence for transmission failure from granular to supragranular layers when tested through the deviated eye (Singer et al. 1980). We would thus like to conclude that perceptual disorders in amblyopia have a common pathophysiological basis, viz., the failure to consolidate those intracortical connections which link the cells at the input stage in layer IV to the neurons mediating the cortical output.
Acknowledgements. We want to thank Susanne Zieglgfinsberger for her valuable help in the preparation of the histological sections, Margot Steinleitner for the care given to the kittens, and Mariele Kremling for the preparation of the manuscript. We further express our thanks to Dr. Platzer for providing the technical facilities for quantitative image processing and H. Bruchle for his help in adapting the software to our needs.
References
Albus K (1975a) A quantitative study of the projection area of the central and the paracentral visual field in area 17 of the cat. I. The precision of the topography. Exp Brain Res 24:159-179
Albus K (1975b) A quantitative study of the projection area of the

214 W. Singer et al.: Development of Orientation Columns
central and the paracentral visual field in area 17 of the cat. II. The spatial organization of the orientation domain. Exp Brain Res 24:181-202
Albus K (1979) 14C-deoxyglucose mapping of orientation subunits in the cats visual cortical areas. Exp Brain Res 37:60%613
Barlow HB (1975) Visual experience and cortical development. Nature 258:199-204
Blakemore C, Cooper GF (1970) Development of the brain depends on the visual environment. Nature 228:477-478
Blakemore C, Van Sluyters RC (1975) Innate and environmental factors in the development of the kitten's visual cortex. J Physiol (Lond) 248:663-716
Blasdel GG, Mitchell DE, Muir DW, Pettigrew JD (1977) A physiological and behavioural study in cats of the effect of early visual experience with contours of a single orientation. J Physiol (Lond) 265:615-636
Buisseret P, Imbert M (1976) Visual cortical cells. Their develop- mental properties in normal and dark reared kittens. J Physiol (Lond) 255:511-525
Creutzfeldt OD, Irmocenti GM, BrOoks D (1974) Vertical organi- zation in the visual cortex (Area 17). Exp Brain Res 21: 315-336
Creutzfeldt OD, Garey LJ, Kuroda R, Wolff JR (1977) The distribution of degenerating axons after small lesions in the intact and isolated visual cortex of the cat. Exp Brain Res 27: 419-440
Cynader M, Mitchell DE (1977) Monocular astigmatism effects on kitten visual cortex development. Nature 270:177-178
Fisken RA, Garey LJ, Powell TPS (1973) Patterns of degeneration after intrinsic lesions of the visual cortex (Area 17) of the monkey. Brain Res 53:208-213
Hood DG, Coleman PD (1979) Demonstration of orientation columns with [C14]-2-deoxyglucose in a cat reared in a striped environment. Brain Res 173:538-542
Freeman RD, Pettigrew JD (1973) Alteration of visual cortex from environmental asymmetries. Nature 246:359-360
Freeman RD, Thibos LN (1975a) Contrast sensitivity in humans with abnormal visual experience. J Physiol (Lond) 247: 687-710
Freeman RD, Thibos LN (1975b) Visual evoked responses in humans with abnormal visual experience. J Physiol (Lond) 247:711-724
Fregnac Y, Imbert M (1978) Early development of visual cortical cells in normal and dark-reared kittens. Relationship between orientation selectivity and ocular dominance. J Physiol (Lond) 278:27-44
Hebb DO (1949) The organization of behaviour. Wiley, New York Henry GH, Dreher B, Bishop PO (1974) Orientation specificity of
cells in cat striate cortex. J Neurophysiol 37:1394-1409 Hirsch HVB, Spinelli DN (1970) Visual experience modifies
distribution of horizontally and vertically oriented receptive fields in cats. Science 168:86%871
Hirsch HVB, Spinelli DN (1971) Modification of the distribution of receptive field orientation in cats by selective visual exposure during development. Exp Brain Res 13:509-527
Hubel DH, Wiesel TN (1962) Receptive fields, binocular interac- tion, and functional architecture in the cat's visual cortex. J Physiol (Lond) 160:106-154
Hubel DH, Wiesel TN (1963a) Shape and arrangement of columns in cat's striate cortex. J Physiol (Lond) 160:106-154
Hubel DH, Wiesel TN (1963b) Receptive fields of cells in striate cortex of very young, visually inexperienced kittens. J Neurophysiol 26:994-1002
Hubel DH, Wiesel TN (1965) Receptive fields and functional architecture in two non-striate visual areas (18 and 19) of the cat. J Neurophysiol 28:22%289
Hubel DH, Wiesel TN (1974a) Sequence regularity and geometry
of orientation columns in the monkey striate cortex. J Comp Neurol 158:267-294
Hubel DH, Wiesel TN (1974b) Uniformity of monkey striate cortex. A parallel relationship between field size, scatter and magnification factor. J Comp Neurol 158:295-306
Hubet DH, Wiesel TN, LeVay S (1977a) Plasticity of ocular dominance columns in monkey striate cortex. Phil Trans R Soc Lond [Biol] 278:377--409
Hubel DH, Wiesel TN, Stryker MP (1977b) Orientation columns in macaque monkey demonstrated by the 2-deoxyglucose autoradiographic technique. Nature 269:328-330
Hubel DH, Wiesel TN, Stryker MP (1978) Anatomical demonstra- tion of orientation columns in macaque monkey. J Comp Nenrol 177:361-380
Imbert M, Buisseret P (1975) Receptive field characteristics and plastic properties of visual cortical cells in kittens reared with or without visual experience. Exp Brain Res 22:25-36
Lund JS, Henry GH, MacQueen CL, Harvey AR (1979) Anatomi- cal organization of the primary visual cortex (Area 17) of the cat. A comparison with Area 17 of the macaque monkey. J Comp Neurol 184:59%618
Mitchell DE, Wilkinson F (1974) The effect of early astigmatism on the visual resolution of gratings. J Physiol (Lond) 243: 739-756
Mitchell DE, Freeman RD, Millodot M, Haegerstrom G (1973) Meridional amblyopia: evidence for modification of the human visual system by early visual experience. Vision Res 13:535-558
Mitzdorf U, Singer W (1978) Prominent excitatory pathways in the cat visual cortex (A17 and A18). A current source density analysis of electrically evoked potentials. Exp Brain Res 33: 371-394
Palmer LA, Rosenquist AC, Tusa RJ (1978) The retinotopic organization of lateral suprasylvian visual areas in the cat. J Comp Neurol 177:237-256
Pettigrew JD (1978) The paradox of the critical period for striate cortex. In: Cotman CW (ed) Neuronal plasticity. Raven Press, New York, pp 311-330
Pettigrew JD, Olson C, Hirsch HVB (1973) Cortical effect of selective visual experience. Degeneration or reorganization? Brain Res 51:345-351
Rauschecker JP (1979) Orientation-dependent changes in response properties of neurons in the kitten's visual cortex. In: Freeman RD (ed) Developmental neurobiology of vision. Plenum Press, New York, pp 121-133
Rauschecker JP, Singer W (1979) Changes in the circuitry of the kitten's visual cortex are gated by postsynaptic activity. Nature 280:58-60
Rauschecker JP, Singer W (1981) The effects of early visual experience on the cat's visual cortex and their possible explanation by Hebb synapses. J PhysioI (Lond) 310
Shatz CA, Lindstrom S, Wiesel TN (1977) The distribution of afferents representing the right and left eyes in the cats visual cortex. Brain Res 131:103-116
Sherk H, Stryker MP (1976) Quantitative study of cortical orientation selectivity in visually inexperienced kittens. J Neurophysiol 39:63-70
Singer W (1976) Modification of orientation and direction selectiv- ity of cortical cells in kittens with monocular vision. Brain Res 118:460-468
Singer W, Tretter F (1976) Receptive-field properties and neuronal connectivity in striate and parastriate cortex of contour-deprived cats. J Neurophysiol 39:613-630
Singer W, Tretter F, Cynader M (1975) Organization of cat striate cortex. A correlation of receptive field properties with afferent and efferent connections. J Neurophysiol 38: 1080-1098

W. Singer et al.: Development of Orientation Columns
Singer W, von Gr/inau MW, Rauschecker J (1980) Functional amblyopia in kittens with unilateral exotropia. I. Electrophy- siological assessment. Exp Brain Res 40:294-304
Skeen LC, Humphrey AL, Norton TT, Hall WC (1978) Deoxygin- cose mapping of the orientation columns system in the striate cortex of the tree shrew, Tupaia glis. Brain Res 142:538-545
Sokoloff L, Reivich M, Kennedy C, Des Rosiers MH, Patlak CS, Pettigrew KD, Sakurada O, Shinohara M (1977) The 14C- Deoxyglucose method for the measurement of local cerebral glucose utilization: Theory, procedure, and normal values in the conscious and anesthetized albino rat. J Neurochem 28: 897-916
Stryker MP, Sherk H, Leventhal AG, Hirsch HVB (1978) Physiological consequences for the cat's visual cortex of effectively restricting early visual experience with oriented contours. J Neurophysiol 41:896-909
Szent~gothal J (1973) Synaptology of the visual cortex. In: Jung R (ed) Handbook of sensory physiology, vol VII, 3 B. Springer, Berlin Heidelberg New York
Szent~igothal J (1975) The module concept in cerebral cortex architecture. Brain Res 95:475-496
215
Toyama K, Matsunami K, Ohno T, Tokashiki S (1974) An intracellular study of neuronal organization in the visual cortex. Brain Res 21:45-66
Tretter F, Cynader M, Singer W (1975a) Modification of direction selectivity of neurons in the visual cortex of kittens. Brain Res 84:143-149
Tretter F, Cynader M, Singer W (1975b) Cat parastriate cortex. A primary or secondary visual area? J Neurophysiol 38: 1098-1113
Tusa RT, Rosenquist AC, Palmer LA (1979) Retinotopic organi- zation of areas 18 and 19 in the cat. J Comp Neurol 185: 657-678
Watkins DW, Wilson JR, Sherman SM (1978) Receptive-field properties of neurons in binocular and monocular segments of striate cortex in cats raised with binocular lid suture. J Neurophysiol 41:322-337
Wiesel TN, Hubel DH (1974) Ordered arrangement of orientation columns in monkeys lacking visual experience. J Comp Neurol 158:307-318
Received March 31, 1980





























