Embed Size (px)
Citation preview

REVIEW
Roles of the Ras/Raf/MEK/ERK pathway in leukemia therapy
LS Steelman1,2, RA Franklin1, SL Abrams1, W Chappell1, CR Kempf2, J Basecke3, F Stivala4, M Donia4, P Fagone4, F Nicoletti4,M Libra4, P Ruvolo5, V Ruvolo5, C Evangelisti6, AM Martelli6,7 and JA McCubrey1
1Department of Microbiology and Immunology, Brody School of Medicine at East Carolina University, Greenville, NC, USA;2Department of Physics, East Carolina University, Greenville, NC, USA; 3Department of Medicine, University of Gottingen,Gottingen, Germany; 4Department of Biomedical Sciences, University of Catania, Catania, Italy; 4Department of Leukemia,University of Texas MD Anderson Cancer Center, Houston, TX, USA; 6Dipartimento di Scienze Anatomiche Umane eFisiopatologia dell’Apparato Locomotore, Universita di Bologna, Bologna, Italy and 7IGM-CNR, Sezione di Bologna, C/o IOR,Bologna, Italy
The Ras/Raf/mitogen-activated protein kinase (MEK)/extracel-lular signal-regulated kinase (ERK) pathway is often implicatedin sensitivity and resistance to leukemia therapy. Dysregulatedsignaling through the Ras/Raf/MEK/ERK pathway is often theresult of genetic alterations in critical components in thispathway as well as mutations at upstream growth factorreceptors. Unrestricted leukemia proliferation and decreasedsensitivity to apoptotic-inducing agents and chemoresistanceare typically associated with activation of pro-survival path-ways. Mutations in this pathway and upstream signalingmolecules can alter sensitivity to small molecule inhibitorstargeting components of this cascade as well as to inhibitorstargeting other key pathways (for example, phosphatidylinosi-tol 3 kinase (PI3K)/phosphatase and tensin homologue deletedon chromosome 10 (PTEN)/Akt/mammalian target of rapamycin(mTOR)) activated in leukemia. Similarly, PI3K mutations canresult in resistance to inhibitors targeting the Ras/Raf/MEK/ERKpathway, indicating important interaction points between thepathways (cross-talk). Furthermore, the Ras/Raf/MEK/ERKpathway can be activated by chemotherapeutic drugs com-monly used in leukemia therapy. This review discusses themechanisms by which abnormal expression of the Ras/Raf/MEK/ERK pathway can contribute to drug resistance as well asresistance to targeted leukemia therapy. Controlling theexpression of this pathway could improve leukemia therapyand ameliorate human health.Leukemia (2011) 25, 1080–1094; doi:10.1038/leu.2011.66;published online 15 April 2011Keywords: resistance; therapeutic sensitivity; targeted therapy;Ras; Raf; ERK
Introduction
The Ras/Raf/mitogen-activated protein kinase (MEK)/extracellularsignal-regulated kinase (ERK) signaling pathway has key roles inthe transmission of proliferative signals from membrane-boundreceptors.1–12 Mutations can occur in the genes encodingpathway constituents (for example, Ras and Raf), upstreamreceptors (for example, Kit, Fms and Fms-like tyrosine kinase(Flt)-3) or chromosomal translocations (for example, BCR–ABLand TEL–platelet-derived growth factor receptor (PDGFR)),which activate this pathway. Chemotherapeutic drugs
frequently used in leukemia therapy often activate thispathway.1,2 This pathway relays this information throughinteractions with various other proteins to the nucleus to controlgene expression.1–12 This review will discuss how these path-ways may be aberrantly regulated in leukemia and contribute totherapeutic sensitivity/resistance and in some cases poorprognosis.4–5,13 Inhibition of Ras (or Ras-related molecules),Raf, MEK and ERK may prove useful in leukemia treatment.These observations have propelled the pharmaceutical industryto develop inhibitors that target key components of this pathwayand many have been evaluated or are currently being evaluatedin clinical trials.
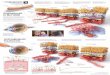
The Ras/Raf/MEK/ERK signaling pathway consist of a kinasecascade that is regulated by phosphorylation and de-phosphory-lation by specific kinases, phosphatases as well as GTP/GDPexchange proteins, adaptor proteins and scaffolding proteins. Anoverview of this pathway and common sites where mutatedupstream receptors and products of chromosomal translocationmay activate the pathway is presented in Figure 1. The sites ofintervention of signal transduction inhibitors are also shown inthis diagram.
The membrane localization of these components is oftencritical for their activity, although some members of thesepathways can function in other cellular regions (for example,mitochondrion (Raf-1), nucleus (ERK)). Indeed, one emergingobservation in ERK1/2 signaling is the realization that thispathway generates specific biological responses dependent onwhere in the cell the signal originates. Ras subcellularlocalization can determine substrate specificity through distinctutilization of scaffold proteins.14 This substrate selectivity isgoverned by the participation of different scaffold proteins thatdistinctively couple ERK1/2, activated at defined subcellulardomains, to specific substrates. Clearly, the subcellular localiza-tion of pathway components and the presence of variousadaptor and scaffolding molecules are critical for the activity ofthese pathways. The localization of various signaling moleculesalso becomes important in determining the effects of certaininhibitors. The regulation and function of this pathway will beconcisely reviewed as well as the effects of genetic mutations inthese pathways that are important in human cancer andleukemia.
Overview of the Ras/Raf/MEK/ERK pathway
After growth factor/cytokine/mitogen stimulation of the appro-priate (cognate) receptor, a Src homology 2 domain containingprotein (Shc) adaptor protein becomes associated with the
Received 14 January 2011; revised 13 February 2011; accepted10 March 2011; published online 15 April 2011
Correspondence: Dr JA McCubrey, Department of Microbiology andImmunology, Brody School of Medicine at East Carolina University,600 Moye Boulevard, 5th Floor Brody Building 5N98C, Greenville,NC 27858, USA.E-mail: [email protected]
Leukemia (2011) 25, 1080–1094& 2011 Macmillan Publishers Limited All rights reserved 0887-6924/11
www.nature.com/leu

C-terminus of activated specific growth factor receptor (forexample, FMS, Flt-3, PDGFR, insulin-like growth factor-1receptor and many others).3–6 Shc recruits the growth factorreceptor-bound protein 2 and the son of sevenless homologprotein, resulting in the loading of membrane-bound Raswith GTP.15
Ras can also be activated by growth factor receptor tyrosinekinases (RTK), such as insulin-like growth factor-1 receptor, viaintermediates like insulin receptor substrate proteins that bindgrowth factor receptor-bound protein 2.15,16 Ras:GTP thenrecruits Raf to the membrane where it becomes activated, likelyvia a Src-family tyrosine (Y) kinase.2,17–19 At this point, we willbe somewhat generic, although it should be pointed out thatboth Ras and Raf are members of multi-gene families and thereare three Ras members (Ki-Ras, N-Ras and Ha-Ras)15 and threeRaf members (B-Raf, Raf-1 (a.k.a C-Raf) and A-Raf).18,19 Raf is
responsible for serine/threonine (S/T) phosphorylation ofMEK1.20 MEK1 phosphorylates ERK1 and 2 at specific T and Yresidues.21 Activated ERK1 and ERK2 serine/threonine kinasesphosphorylate and activate a variety of substrates, including p90ribosomal six kinase-1 (p90Rsk-1).22 ERK1/2 has many down-stream and even upstream substrates (see below). The number ofERK1/2 targets is high (460). Thus, suppression of MEK and ERKactivities will have profound effects on cell growth.
Downstream of ERK is p90Rsk-1. p90Rsk-1 can activate thecAMP response element-binding protein transcription factor,which can influence gene expression.22 Activated ERK can alsotranslocate to the nucleus and phosphorylate additionaltranscription factors, such as Elk-1, cAMP response element-binding protein, Fos and globin transcription factor 1 andothers,23 which bind promoters of many genes, includinggrowth factor and cytokine genes that are important inpromoting growth and preventing apoptosis in hematopoieticcells. Thus, we have described in a simple fashion how theRas/Raf/MEK/ERK cascade can transmit a signal from the cellmembrane to the nucleus. Under certain circumstances,aberrant regulation of this pathway can contribute to abnormalcellular proliferation, which may lead to many abnormalitiesincluding: abrogation of cytokine dependence, secretionof autocrine cytokines, leukemic transformation and drugresistance.4,5,10–12,24–31
Activated ERK can also phosphorylate B-Raf, Raf-1 and MEK1,which alter their activity (Figure 1). Depending on the sitephosphorylated on Raf-1, ERK phosphorylation can eitherenhance32 or inhibit33 Raf-1 activity. In contrast, when B-Raf34
or MEK1 (ref. 35) are phosphorylated by ERK, their activitydecreases. ERK can also exert negative feedback by interferingwith Ras activation by phosphorylating son of sevenless.36 Thesephosphorylation events serve to alter the stability and/oractivities of the proteins. It is important that the reader realizesthat certain phosphorylation events either inhibit or repress theactivity of the affected protein. This often depends on theparticular residue phosphorylated on the protein, which canconfer a different configuration to the protein or target theprotein to a different subcellular localization that may result inits proteasomal degradation. Furthermore, as previously men-tioned, certain phosphorylation events will actually serve to shutoff or slow down the pathway. Thus, protein phosphorylation bythe Ras/Raf/MEK/ERK pathway is a very intricate process, whichserves to finely tune the signal often originating either from agrowth factor or mitogen.
There are many phosphatases that remove key phosphates onmolecules in this pathway.37 These include protein phosphatase1 (PP1), PP2a and PP5. PP1 and/or PP2a remove the phosphatefrom S259 on Raf-1. PP5 removes the phosphate from S338 onRaf-1.38 The activity (velocity) of the phosphatases has beenproposed to be important in carcinogenesis and interactionswith other signaling pathways.39
There are also various scaffolding proteins that regulate theactivity of this pathway. These include: Raf kinase inhibitoryprotein (RKIP), kinase suppressor of Ras, 14-3-3 and Mp-1.40–43
Many of these scaffolding proteins serve to modulate the activityof key components. Some scaffolding proteins (for example, Mp-1)serve to promote a complex between MEK and ERK to enhancesignal transduction. In contrast, in some cases they mayprevent activation (for example, RKIP), or keep the complexin an inactive state. The scaffolding proteins may be phos-phorylated by other proteins (for example, ERK), which promotetheir removal from Raf-1 and result in enhanced signaling,which in some cases may promote tumor progression andmetastasis.42,43
AAA
ERK
Raf-1
Elk CREB
P
P
PP
P
Grb2Shc
SOS
P
MEK
Ras
Regulation of gene transcription(Proliferation, Leukemic Transformation, Drug Resistance)
Sutent,Nexavar,
Midostaurin,Semaxianib,Lestaurtinib,Tandutinib
GF
PLX-4720,SB590885
Selumetinib,PD0329501,
XL-518,RDEA119
NegativeFeedback
Loopsof ERK
Inhibitors inclinical trialsor in therapy
GrowthFactor
AutocrineTransformation
Zarnesta,Sarasar
AP-2
P
eIF4B
P
ProteinTranslation
rpS6
P
B-Raf
NexavarZM336372,
RAF265
BCR-ABLInhibitors:Imatinib,Nilotinib,
Bosutinib,Dasatinib
Interactions withPI3K/Akt/mTOR
Pathway
MutatedGF Receptors
ChromosomalTranslocations
MEKInh
PanRafInh
GFRInh
GFInh
FTInh
B-RafSelective.
Inh
Figure 1 Overview of the Ras/Raf/MEK/ERK pathway and potentialsites of therapeutic intervention with small molecule membrane-permeable inhibitors. The Ras/Raf/MEK/ERK pathway is regulated byRas, as well as various upstream growth factor receptors and non-receptor kinases. The downstream transcription factors regulated bythis pathway are indicated in diamond shaped outlines. This drawingdepicts a relative common, yet frequently overlooked phenomenon inhuman cancer, autocrine transformation. Sites where various smallmolecule inhibitors suppress this pathway are indicated by blackoctagons. Drugs that have been evaluated to suppress this pathway(many in clinical trials, see below) are indicated in open boxes. Drugsthat have been approved to treat cancer patients (not necessarily inleukemia patients) include: Sutent, Nexavar, Imatinib, Nilotinib, andDasatinib. Drugs that have been in clinical trials to treat cancerpatients (not necessarily in leukemia patients and not necessarily arethey continuing to be evaluated in clinical trials) include: Midostaurin,Semaxanib, Lestaurtinib, Tandutinib, Zarnestra, Sarasar, PLX-4720,RAF265, Selumetinib, XL-518, RDEA119 and Bosutinib. GF, growthfactor; GFR, growth factor receptor.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1081
Leukemia

Rationale for targeting the Ras/Raf/MEK/ERK pathway toimprove leukemia therapy
Effective targeting of signal transduction pathways activated bymutations, gene amplification or various leukemia therapiesmay be an appropriate approach to limit leukemia growth anddrug resistance. The Ras/Raf/MEK/ERK pathway can be activatedby mutations/amplifications of upstream growth factor recep-tors. The abnormal production of growth factors can frequentlyresult in receptor activation, which in turns activates theRas/Raf/MEK/ERK cascade (Figure 1). Furthermore, chemother-apeutic drugs used in leukemia therapy may induce the Ras/Raf/MEK/ERK cascade, which may contribute to drug resistance(Figure 2).
Activation of the Ras/Raf/MEK/ERK pathway by drugs used inleukemia therapy
Doxorubicin (a.k.a. adriamycin) is a commonly prescribed anti-leukemia drug. Doxorubicin exerts its chemotherapeutic effectsthrough multiple mechanisms. One mechanism is through itsinteractions with DNA and inhibition of topoisomerase II.4 Theother mechanism of action is due to the generation of reactiveoxygen species (ROS) that occurs via the interaction ofdoxorubicin with iron.1,2,4,44,45 Doxorubicin treatment resultsin the intracellular generation of superoxide anion, hydrogenperoxide and hydroxyl radicals.1,2,44,45 ROS appear to beimportant for some of the therapeutic effects of doxorubicin asscavenging oxygen radicals using anti-oxidants decreases theability of doxorubicin to induce apoptosis.1,2,4,44,45 AlthoughROS are important for some of the activities of doxorubicin theyare also the cause of some of the undesirable side effects of thisdrug1,2,4,44,45 (see Figure 2).
ROS induce activation of Ras/Raf/MEK/ERK, c-Jun N-terminalkinase, p38, big mitogen-activated protein (MAP) kinase/ERK5and phosphatidylinositol 3 kinase (PI3K)/phosphatase and tensinhomologue deleted on chromosome 10 (PTEN)/Akt/mammaliantarget of rapamycin (mTOR) signaling pathways. Oxidativestress-induced ERK1/2 activation is reported in a variety of celltypes.1,10,45 In some cases, ROS can act directly on cytokine andgrowth factor receptors and induce the Ras/Raf/MEK/ERKsignaling pathway.1,10,45 ROS can induce the ligand-indepen-dent activation of some cytokine and growth factor receptorsand a subsequent increase in Ras and Raf/MEK/ERK activity.1,10
ROS are also known to inhibit PPs.1,4,45 Inhibition ofphosphatase activity can result in activation of the Ras/Raf/MEK/ERK signaling pathway.1,4 The Ras/Raf/MEK/ERK kinase signalingcascade can be activated at multiple points by ROS. Thus, thispathway is important in drug resistance. Targeting this pathwaymay be a novel therapeutic approach for drug resistant leukemia,which is often cross-resistant to multiple chemotherapeutic drugs.
ROS can also result in increases in intracellular Ca2þ , butROS are also able to activate the calcium calmodulin-dependentkinases (CaM-Ks) in the absence of increases in intracellularCa2þ .45 CaM-KI and CaM-KII are expressed in many tissues,whereas the expression of CaM-KIV is more restricted. Multiplegenes encode different isoforms of CaM-KII, which aredesignated CaM-KII-a, -b, -d and -g.45 Maximal activity ofCaM-KI, CaM-KII and CaM-KIV requires phosphorylation. Themechanisms by which these enzymes are phosphorylateddiffers.45 CaM-KII undergoes autophosphorylation whereasCaM-KI and CaM-KIV are phosphorylated by CaM-KK. Oncephosphorylated, these kinases retain catalytic activity even inthe absence of increased intracellular Ca2þ . PP1 and PP2A
cleave the phosphate group from CaM-KII and CaM-KIVrendering them inactive.45 Inactivation of these phosphatasesis one mechanism by which ROS can activate these kinases.45
ROS and Ca2þ /calmodulin can activate the CaM-K cascade,which in turn can activate the Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR cascades, often via Ras (see Figure 2). An illustration ofthe effects of induction of Ras/Raf/MEK/ERK by leukemiatherapies on cell cycle and apoptotic regulatory moleculesand protein translation is presented in Figure 3. In thesubsequent sections, we will discuss how the Ras/Raf/MEK/ERK pathway may be activated by mutations as well aschemotherapeutic drugs used in leukemia therapy.
Mutations affecting the Ras/Raf/MEK/ERK pathway inleukemia
Mutations that affect activation of the Ras/Raf/MEK/ERK pathwayin leukemia have been frequently detected. In acute myeloid
p90Rsk
Raf
MEKP
CREBP P
P
Ras
AktP
ERK
P
PTEN
P
PI3K
PDK
PIP2 PIP3
Ca2+/Calmodulin
I�K
P
P
I�B
P
NF�B
P
NF�B
CaM-KK
CaM-KIP
CaM-KIVP
CaM-KII
AAA AAA
ROS
Doxorubicin
P
p53
ATM
P
MDM2
DDR
Gene Transcription
DNA Damage
DNA Repair
Gene Mutation PABP
7mGpppGeIF4E
eIF
4G
eIF4A
AAAAAAeIF4B
Protein Translation
P
p53
Chemotherapeutic Drug Resistance
Figure 2 Induction of the CaM-K, Ras/Raf/MEK/ERK, PI3K/PTEN/Akt/mTOR and p53 pathways by chemotherapy. Chemotherapeutic drugssuch as doxorubicin can generate ROS, which in turn can induce theCaM-K cascade and can result in activation of both the Ras/Raf/MEK/ERK and Ras/PI3K/PTEN/Akt/mTOR pathways. Inappropriate orprolonged stimulation of these pathways can result in altering geneexpression leading to chemotherapeutic drug resistance. In addition,ROS can mobilize the ATM/p53 pathway, which in turn can haveeffects on the DNA damage response, DNA repair and lead to genemutations, which further contribute to chemotherapeutic drugresistance. Induction of p53 by chemotherapeutic drugs can alsoinduce the expression of the discoidin domain receptor (DDR), whichcan result in Ras activation and subsequent stimulation of the Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways. Akt can phosphorylateand inactivate murine double minute 2 (MDM2), preventing its effectson p53 stability.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1082
Leukemia

AAA
p70S6K
Foxo
P
CREB
P
rpS6
P
4E-BP1
P
eIF-4E
Translation
Transcription
PABP
7mGpppGeIF4E
eIF
4G
eIF4A
AAAAAAeIF4B
Pp53
eIF4B
PPP
Mitochondrion
Raf-1Bad
Bcl
-2
BimPP
Bad
PP
Caspase9Mcl-1
P P
Bcl
-XL
M
S
G2
G1
E2F
E2F
E2F CDK2
Rb
Rb
cyclin D
CDK4/6
Rb
Rb
cyclin E
CDK2
cyclin B
CDK 1 P
P
ContinuousGrowth
P
ERK
Raf
P
P
CaM-K
MEK
Ras
ATM
P
Chemotherapyor Radiotherapy
AKT
PDK
PI3K
p90Rsk
P
P
p53
P
mTOR
Rheb
Raptor
TSC2
TSC1
PP
mLST8
PP
mTORC1PRAS40
PDEPTOR
P
PP
DDR1
Apoptosis
Bax
PUMA
Noxa
Foxo
CellCycle
MNK1/2
P
E2F
P
p21Cip1
p27Kip1 ROS
MDM2
P
P
P
P
P
cyclin A
Figure 3 Induction of the Ras/Raf/MEK/ERK pathway after leukemia therapy and subsequent effects on cell cycle progression, survival pathwaysand protein translation. After chemotherapy or radiotherapy there can be activation of signaling pathways, which can actually promote cellsurvival and may lead to therapy resistance. Chemotherapeutic drug treatment (shown in irregular black oval) frequently results in the induction ofROS (shown in black square). ROS can induce the calcium calmodulin kinase (CaM-K) cascade that can induce Ras, which can subsequentlyactivate both the Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR cascades (most components of the two cascades, which promote signaling are show inwhite ovals, transcription factors are shown in white diamonds). Induction of the Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways can resultin the activation of many survival pathways, and regulate both cell cycle progression as well as protein translation. An emerging field in leukemiatherapy is the role of these pathways in regulation of protein translation. These two pathways control the assembly of the eIF4 translation apparatus,downstream of mTORC1, which regulates the translation of many ‘weak’ mRNAs that are often involved in cell survival (for example, Mcl-1,c-Myc and vascular endothelial growth factor (VEGF)). Many of the phosphorylation events on apoptotic regulatory molecules such as Bad, Bimand caspase 9 may affect the activity or stability of the proteins, which can alter their subcellular distribution and target them to proteasomes orresult in their degradation. These molecules are depicted in black as they are inactivated or degraded after phosphorylation by the Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways. In contrast, some molecules such as Mcl-1 may be stabilized by phosphorylation by these cascades,although there are other pathways (for example, c-Jun N-terminal kinase (JNK)), which result in their inactivation. Also Raf-1 can be localized tothe mitochondrial membrane and affect apoptotic regulatory molecules such as Bcl-2, Bcl-XL and Bad. Some of the phosphorylation eventsmediated by Akt actually serve to inhibit the activities of key proteins such as the Foxo transcription factors, the Tuberous sclerosis protein (TSC1)and (TSC2) tumor-suppressor proteins and the murine double minute (MDM2) ubiquitin ligase (depicted in black ovals). MDM2 serves to regulatep53 protein stability by ubiquitination, however, when it is phosphorylated by Akt it is inactivated. Moreover chemotherapeutic drugs andradiotherapy can induce the ataxia telangiectasia mutated (ATM) protein shown in a black oval, which can in turn phosphorylate and regulate p53.p53 can have complex positive and negative effects on cell growth (depicted in white diamond); it can regulate the expression of p21 cyclin-dependent kinase inhibitory protein-1 (p21Cip1), which controls cell cycle progression. p53 can also control the transcription of genes such asPuma, Noxa and Bax, which are involved in apoptosis (all of these molecules are shown in black ovals, as they tend to inhibit cell cycleprogression or promote apoptosis). Alternatively, p53 can regulate the expression of the discoidin domain receptor (DDR) (shown in white oval),which in turn can regulate Ras and stimulate activation of the Ras/MEK/ERK pathway. Both Akt and ERK can phosphorylate p21Cip1 that alters itsactivity and ability to inhibit cell growth (shown as black phosphorylation sites) and subsequently influence cell growth and therapy resistance.p27Kip1 can also be phosphorylated by both Akt and ERK; however, the effects of these phosphorylation events are unclear. Akt phosphorylation ofp27Kip1 may result in its cytoplasmic localization, while ERK phosphorylation of p27Kip1 may result in elevated levels of the protein. Hence,phosphorylation of proteins by ERK and Akt can have dramatic effects on cell proliferation and contribute to the therapy resistance of leukemias.This figure serves as an introduction as to how activation of these pathways by chemotherapy and radiotherapy may contribute to therapeuticresistance in leukemia, however, there are numerous other important proteins also regulated by these pathways, which contribute to therapyresistance.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1083
Leukemia

leukemia (AML) and myelodysplastic syndrome (MDS), theseare often called class I mutations as they are believed tostimulate proliferation, whereas class II mutations consist ofmutations (often chromosomal translocations) of transcriptionfactors and the novel chimeric transcription factors preventdifferentiation of the cells. In some cases, there are mutations/gene rearrangements, which result in receptor gene activation(for example, FLT3, KIT, PDGFR and other receptors).46,47
The FLT3 gene is mutated in approximately 20–25% of AMLpatients48 and in o5% of MDS patients. It has been observedthat one-third of MDS patients acquire mutations at either FLT3or NRAS during the evolution into AML.49 Another studyexamined the mutations present in the NRAS, FLT3, KIT andMLL genes in 381 patients with MDS and 4130 patients withAML.50 Mutations at NRAS, FLT3, KIT and MLL were reported tobe more frequent in secondary (s) AML compared with primaryAML and MDS. The incidence of NRAS mutations increasedfrom about 6.3% in MDS to about 12% in sAML. KIT816mutations are rarer, in less than 2% of the patients examined.The presence of FLT3 length mutations increased in AMLpatients compared with MDS patients. Detection of theFLT-length mutation at initial diagnosis of MDS is associatedwith leukemic transformation and poorer prognosis.51
In addition, in this large study, the incidence of FLT3-lengthmutation was higher in relapsed AML than in all other classes ofAML and MDS examined, documenting the significance ofFLT3-length mutation in AML progression. NRAS mutationswere among the most frequent mutation detected in MDS. Theseinvestigators demonstrated that NRAS mutations were asso-ciated with karyotype evolution, either the acquisition ofmonosomy 7 occurring during MDS transformation andassociated with a poorer survival.
In childhood acute lymphoblastic leukemia, there aremutations in FLT3, Ras (NRAS and KRAS), BRAF and genesencoding proteins, which modulate pathways activity (forexample, mutations in the Shp2 phosphatase, a.k.a.PTPN11).52–55 It has been recently estimated that 35% of anunselected cohort of childhood acute lymphoblastic leukemiapatients contained mutations in NRAS/KRAS2/PTPN11 orFLT3.56 These mutations were mutually exclusive, an event thatthe investigators state the importance of these mutations inleukemogenesis. Furthermore, the investigators observed anincidence of 21% mutations at this pathway in relapsed patients.In contrast, in these and other studies, the investigators didnot observe mutations at BRAF.57 Overall, BRAF is notfrequently mutated in childhood acute lymphoblastic leukemia,although there are some reports documenting activating BRAFmutations.54,58
Furthermore, many chromosomal translocations (for example,BCR–ABL, TEL–PDGFR) will activate the Ras/Raf/MEK/ERKpathway. Thus, there is a strong rational for developinginhibitors, which suppress Raf/MEK/ERK pathway activation.
Mutations in therapy-induced AML and MDS
There are mutations in RAS, BRAF, CRAF and PTPN11 detectedin t-AML and t-MDS. Previously, these patients were oftentreated with alkylating agents, topoisomerase II inhibitors orradiotherapy.59–61
An additional study has documented mutated alleles of CRAFin t-AML.62 These t-AMLs arose after chemotherapeutic drugtreatment of breast cancer patients. The mutated CRAF geneswere transmitted in the germ line, thus, they were notspontaneous mutations in the leukemia, but they may be
associated with the susceptibility to induction of t-AML in thebreast cancer patients studied. Also some cases of childhoodacute lymphoblastic leukemia have arisen in cardio-facio-cutaneous syndrome patients who have a germline BRAFmutation.63
Novel RAF mutations in human cancers
Recently, it has been determined that there are genetictranslocations of BRAF and CRAF in prostate and gastric cancerand melanoma.64,65 These translocations were detected usingpaired-end transcriptome sequencing to screen ETS rearrange-ment-negative prostate tumors for targetable gene fusions. Theyare currently believed to be associated with later stages of thecancer. Although these chromosomal translocations are rare,they offer a new mode of attacking their activity as they aresensitive to Raf and MEK inhibitors in in vitro studies. Whethersimilar translocations will be detected in leukemia remains to bedetermined.
FLT3 mutations and leukemia therapy
Inhibitors (for example, midostaurin, sutent, semaxanib, lestaur-tinib, tandutinib and others), which target Flt-3 have beendeveloped and examined in the treatment of leukemia patientswith FLT3 mutations.46 Some of these inhibitors may haveadditional targets besides Flt-3 (for example, protein kinase C,vascular endothelial growth factor receptor, Kit, PDGFR andothers). Some of these inhibitors may induce Bim that promotesapoptosis.46 Sorafenib is a multi-kinase inhibitor, which alsoinhibits Flt-3. Sorafenib has many targets including Flt-3, Raf, Kitand PDGFR.66–67 Previously, it was shown that sorafenib-induced apoptosis in AML cells through Bim.68
RKIP as a therapeutic target in cancer including leukemia
RKIP was originally identified as a scaffolding protein, whichinhibits the function of Raf-1.43,69,70 Overexpression of RKIPinhibits MEK, ERK and Ap-1 transcription factor activity.Similarly, inhibition of RKIP stimulates MEK, ERK and Ap-1activity. Phosphorylation of RKIP on S153 by protein kinase C71
or ERK72 results in dissociation of RKIP from Raf-1 and relief ofRKIP’s suppressive effects on Raf-1.
RKIP can bind to the N-terminal region of Raf-1 preventingbinding of MEK to Raf-1 and downstream activation.73,74
Although RKIP can regulate ERK activation, ERK can alsoregulate RKIP expression, potentially via phosphorylation onS99 and indirectly by regulating the transcription factor Snail byphosphorylation, which can suppress RKIP transcription.75,76
RKIP has been subsequently shown to interact with otherproteins (for example, nuclear factor-binding immunoglublin-kchain enhancer in B cells and inhibitory kB kinase) and beregulated by other proteins (for example, Snail), which areinvolved in regulation of the epithelial–mesenchymal transi-tion.77,78 RKIP can also antagonize nuclear factor-bindingimmunoglublin-k chain enhancer in B cell activation inresponse to tumor necrosis factor-1 and interleukin-1b stimula-tion.78–80 RKIP expression is characterized as a metastasis-suppressor protein. The transcription factor Snail is a repressor ofRKIP transcription in metastatic prostate cancer.78 Furthermore,ERK can phosphorylate and regulate Snail expression, thusdocumenting the complicated feedback loops between Snailand Raf/MEK/ERK expression. Low levels of RKIP are frequently
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1084
Leukemia

observed in metastatic prostate and breast cancer sampleswhereas higher levels are observed in non-metastatic cells.Increased expression of RKIP can sensitize therapy-resistant cellsto various therapeutic approaches (for example, tumor necrosisfactor-related apoptosis-inducing ligand and chemotherapeuticdrugs).81 In contrast, suppression of RKIP renders therapy-sensitive cells, therapy-resistant.82 Proteasome inhibitors cansuppress the therapy resistance by downregulation of nuclearfactor-binding immunoglublin-k chain enhancer in B cell andSnail and increased expression of RKIP.83 Although there havenot been many studies published regarding RKIP and leukemia,it was recently observed that RKIP had a role in some cases oft-AML containing mutant CRAF. Blast cells from patients withthe CRAF (Raf-1) germline mutations also had loss of RKIP.84
The importance of RKIP was determined by transfectionexperiments with either small interfering RNA directed againstRKIP or expression vectors overexpressing RKIP.84 The levelsof RKIP were determined to influence the levels of CRAF-mediated transformation as high levels of RKIP suppressedCRAF-mediated transformation, while low levels enhancedCRAF-mediated transformation.84
Control of apoptotic regulatory molecules by the Ras/Raf/MEK/ERK pathway
The Ras/Raf/MEK/ERK pathway regulates the activity of manyproteins involved in apoptosis (Figure 3). Deregulated expres-sion of apoptotic regulatory molecules can lead to drugresistance.10–12,30,31 ERK phosphorylates transcription factorsthat influence the transcription of the Bcl-2 family of genes aswell as other important genes involved in the regulation ofapoptosis.27,85 Furthermore, increased expression of Bcl-2 andBcl-XL and decreased expression of Bax is observed in somedrug-resistant leukemias.86–89 Many of the effects of the Ras/Raf/MEK/ERK pathway on apoptosis are mediated by ERK phosphor-ylation of key apoptotic effector molecules (for example, Bcl-2,Mcl-1, Bad, Bim, cAMP response element-binding protein,caspase-9 and many others).90–93
Certain Bcl-2 inhibitors (for example, Abt-737) may sensitizechronic lymphocytic leukemia and chronic myeloid leukemiacells to chemotherapy, implicating the Bcl-2 family members,which are sensitive to Abl-737 (for example, Bcl-2 and Bcl-XL
but not Mcl-1) in their drug resistance.94–95 Bcl-2 inhibitorssensitize B lymphoma cells to rituximab, a chimeric monoclonalantibody, which targets CD20.96 Bcl-2 inhibitors also rendervarious lymphoid malignancies susceptible to proteasomeinhibitors.97
Resistance to the chemotherapeutic drug fludarabine, in somesituations, may also be mediated by other Bcl-2 family memberssuch as increased expression of Mcl-1, which is associated witha poor prognosis.98 This may be regulated by microRNAs. Someof the effects of the microRNAs are on the suppression of genes,such as PTEN and Bim.99 Development of specific Mcl-1inhibitors is in progress based on the crystal structure ofMcl-1.100,101 Mcl-1 confers less protection against chemo-therapy than Bcl-2 in certain experimental situations. This hasbeen examined with Em-Bcl-2 and Em-Mcl-1 mice.102 This maybe due to the effects of ERK as well as mTOR and theirphosphorylation of the apoptotic molecules as well as regulationof protein translation (Figure 3).
A key molecule downstream of the Ras/Raf/MEK/ERK cascadeis the BH3-domain containing anti-apoptotic protein Bim. Bim’sactivity is regulated by phosphorylation at different residues byERK, Akt and c-Jun N-terminal kinase.92,103–105 When Bim is
phosphorylated by ERK and Akt, it is targeted for proteosomaldegradation and also inhibits Bim’s interaction with Bax, a deathexecutioner protein. In contrast, when Bim is phosphorylated byc-Jun N-terminal kinase, it has enhanced pro-apoptotic activ-ity.106 Bim is also transcriptionally regulated by Foxo-3a,(a transcriptional factor), which is also a target of Akt. Normally,when Akt is active, it phosphorylates and inhibits the activity ofFoxo-3a (Figure 3). On cytokine withdrawal of hematopoieticcells, Foxo-3a is not phosphorylated and enters the nucleus andstimulates the transcription of genes, such as Bim and other,which results in apoptosis.107
Bim levels can also be regulated by epigenetic silencing. Thismay be important in the sensitivity of ALL cells to glucocor-toids.108 The glucocorticoid resistance observed in thexenografts and patient biopsies in this study correlated withdecreased histone H3 acetylation. The investigators demon-strated that the histone deacetylase inhibitor vorinostat relievedBim repression and also exerted anti-leukemic effects whencombined with dexamethasone in vitro and in vivo. Thesestudies suggest a new approach to overcome glucocorticoidresistance and improve therapy for high-risk pediatric ALLpatients.
Bim protein levels are also regulated by stromal cellinteractions. When leukemia cells were attached to stromaand b1 integrin was activated, suppression of Bim expressionand apoptosis occurred and drug resistance increased.109 Bimwas targeted to the proteasome and degraded.109 Proteasomalinhibitors suppressed Bim degradation and rendered the cellssensitive to therapy. The investigators of this study suggested thatb1 integrin-mediated stromal interactions with leukemia cellsand subsequent Bim protein degradation may contribute tominimal residual disease. The tumor microenvironment canregulate drug resistance, perhaps due to the presence ofcytokines, which stimulate anti-apoptotic Bcl-2 and otherfactors preventing prevent apoptosis.110–112
Regulation of Bim and Foxo3A by the Raf/MEK/ERK pathway
BCR–ABL-transformed hematopoietic precursor cells have beenshown to have low levels of Bim and to be refractory to theinduction of apoptosis after cytokine withdrawal.113–114 BCR–ABL induces ERK activation and hence suppression of ERK byeither imatinib or dasatinib results in prevention of Bimphosphorylation and Bim is not targeted to the proteosomeand is active. Thus, these drugs are proposed to exert some oftheir inhibitory effects by induction or enhancement of Bimactivity. Knockdown of Bim by small interfering RNA abrogatesimatinib-induced cell death in chronic myeloid leukemiacells.115 Imatinib will induce Foxo-3a in BCR–ABL-transformedcells, which subsequently induces Bim transcription andapoptosis.116 Imatinib induces the transcription of Bim andBmf and also induces posttranslational modifications of Bim andBad favoring apoptosis. Bcl-2 inhibitors can overcome resis-tance to imatinib because of decreases in Bim and Bad levels.115
Mutant receptors such as Flt-3 may induce Foxo-3a inactivation,which can lead to resistance to Flt-3 inhibitors and result in apoor prognosis.51,117,118
Recently, it was demonstrated that Foxo-3a is inactive in AMLcells and is localized in the cytoplasm.119 Treatment of AMLswith MEK and PI3K/Akt inhibitors did not result in the nucleartranslocation of Foxo-3a where it could potentially induce cellcycle inhibitory and apoptotic genes. In contrast, it was shownthat inhibitory kB kinase controlled Foxo-3A expression.Treatment of cells with the inhibitory kB kinase-specific
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1085
Leukemia

inhibitor NEMO resulted in the nuclear translocation of Foxo-3aand induced the expression of p21Cip-1 and Fas by inhibitory kBkinase.119
Regulation of the eIF4F translation complex by the Ras/Raf/MEK/ERK pathway
Certain mRNAs encoding genes implicated in survival (forexample, Mcl-1, c-Myc, Bcl-XL, survivin, cyclin D, vascularendothelial growth factor, ornithine decarboxylase, matrixmetalloproteinase 3 and -9) have 50untranslated regions (UTRs)possessing tertiary structures that are difficult to translate and arehence considered weak mRNAs.120,121 These 50UTR are GþCrich and highly structured and can affect cellular transformationand metastatic progression. Efficient translation of these survivalmRNAs requires the assembly of a translation complex, whichattaches to the 50UTR.122 eIF4E is a component of the eIF4Fcomplex, which stimulates ribosome recruitment. This transla-tion complex is comprised of eIF4E, the RNA helicase eIF4A andits activator eIF4B that serves to unwind the 50UTR. Otherproteins are also involved, eIF4G serves as a scaffold necessaryfor assembly of the complex and eIF2a is a negative regulator,which can block translation if it is phosphorylated by proteinkinase R. eIF4E is the critical rate-limiting factor in the formationof the complex. Ectopic overexpression of eIF4E can transformcertain cells in culture and cooperate with Em-Myc in theinduction of lymphomatogenesis and induce chemotherapeuticdrug resistance.123 However, overexpression of eIF4E, which isdownstream of mTOR and also regulated by ERK does notconfer sensitivity to the mTOR inhibitor rapamycin, in fact, thetumors are rapamycin-resistant. Furthermore, overexpression ofeIF4E in cancers, which were previously rapamycin-sensitivemade them rapamycin-resistant.123
Another kinase downstream in the Ras/Raf/MEK/ERK pathwaythat regulates the translation complex is p90Rsk-1, whichphosphorylates ribosomal protein S6 (rpS6) at S235/236, thesame site as p70S6K.124 Phosphorylation of rpS6 at these sitespromotes its association with the 7-methyguanosine capcomplex. eIF4B exerts a dominant role in recruiting the 40Sribosomal subunit to the mRNA. On mitogen stimulation, eIF4Bis phosphorylated on S422 by p70S6K in a rapamycin-sensitivemanner. It turns out that p90Rsk1 also phosphorylates eIF4B onthe same residue.125 However, the phosphorylation by the twodifferent kinases occurs by very different kinetics.125 Bothp70S6K and p90Rsk-1 are members of the cyclic adenosinemonophosphate kinase, cyclic guanosine monophosphatekinase, protein kinase C family (AGC) of protein kinasesfamily, and require phosphotidylinositide-dependent kinase 1phosphorylation for activation.125 These studies further demon-strate the importance of the Ras/Raf/MEK/ERK cascade in thetranslational machinery and also the redundancy in regulationof rpS6 and eIF4B phosphorylation, which is critical for thetranslation of many critical RNAs involved in growth control.
The ability of eIF4e to enter the eIF4F complex is regulated bythe eIF4E-binding proteins, which are regulated by mTORphosphorylation. On phosphorylation of 4-BP by mTOR, eIF4Edisassociates from the eIF4E-binding proteins and eIF4E canenter the eIF4F complex and stimulate transcription of thesurvival mRNAs with the unique 50UTRs. Recently, it has beenshown that cyclopenta[b]benzofuran flavagines such as silves-trol can modulate eIF4A activity and inhibit translationinitiation.125 Silvestrol and related compounds may be able toenhance chemosensitivity in cancers, which grow in response toderegulated PTEN and downstream eIF4E.125
One of the key targets of the eIF4F complex is Mcl-1.126 Highlevels of Mcl-1 and eIF4E are detected in myelomas, which maycontribute to their drug resistance.127 ERK and p38MAPK canincrease phosphorylation of eIF4E via MAP kinase-interactingserine/threonine-protein kinase1/2 (MNK1/2). ERK can phos-phorylate and activate MNK1/2.128 Activated MNK1 canphosphorylate and activate eIF4E on S209.129 Constitutivelyactivated MNK1 can induce lymphatogenesis in a fashionsimilar to eIF4E.130–131 MNK1/2 is reported to be dispensable fornormal mammalian development.132 Recent studies haveindicated that combined MNK1 and MNK2 deficiency (inmouse crosses lacking both genes) delays tumor progression inboth PTEN-deficient T cells and in glioblastoma.133 Thus,MNK1/2 is potentially an important target for cancer therapyand drug resistance.
The ras homology protein enriched in brain (Rheb) is aGTPase and an activator of mTOR. Rheb is highly expressed insome human cancers.134–138 Increased Rheb expression isassociated with a poor prognosis in breast and head and neckcancers.138 Rheb activity is dependent on farnesylation. Theeffects of enhanced expression of Rheb on the induction oflymphomas in the Em-Myc mouse system were examined.139,140
Introduction of Rheb into the Em-Myc mouse model enhancedthe development of lymphomas. These lymphomas were moreresistant to chemotherapeutic drugs such as doxorubicin. Rhebexpression induced cellular senescence and treatment withrapamycin prevented the induction of senescence.139 Incontrast, c-Myc expression blocked the induction of senescence.However, the combination of c-Myc expression and Rheb,prevented the induction of senescence induced by Rheb and theapoptosis induced by c-Myc, which may explain the increasedincidence of lymphomas in the Rheb/EmMyc mice than inEm-Myc mice.140
This group has shown that the abilities of Ras, Akt and Rheb toactivate cellular senescence are dependent at least in part bytheir ability to activate mTOR. This effect may be due to eIF4Bactivation as eIF4B also induces cellular senescence. MEKinhibitors also inhibit cellular senescence in some systems.139
Thus, in some scenarios mTOR and MEK inhibitors maypromote cell growth by preventing cellular senescence.
Similar to previous experiments performed with introductionof eIF4E into Em-Myc mice, the Rheb-Em-Myc mice alsoexpressed high levels of the Mcl-1 protein. Lymphomas thatoverexpress Rheb are sensitive to both farnesyl transferaseinhibitors and rapamycin.140 The effects of farnesyl transferaseinhibitors on PTEN-deficient tumor cells are dependent onfunctional Rheb activity. In summary, in some cases, Rheb canbe an oncogenic regulator of mTORC1 and eIF4E and Rheb is adirect target of farnesyl transferase inhibitors in cancer.
By performing high-throughput screening assays, 4EGI-1 wasidentified as an eIF4E-binding protein-1 mimetic and a potentinhibitor of the interactions between eIF4E and eIF4G. 4EGI-1 hasbeen shown in some systems to inhibit the growth ofBCR–ABL-transformed Ba/F3 cells but not the parental interleu-kin-3-dependent cells line (Ba/F3).141 The specificity of4EGI-1 was also examined on AML blasts and normal CD34þ
hematopoietic precursor cells and it was demonstrated that 4EGI-1-induced apoptosis in the AML blasts but had much less effectson the differentiation and survival of normal CD34þ cells.142
The antiviral drug ribavirin can block the binding of eIF4E tomRNA.143–144 The effects of this compound were examined insome patients with relapsed M4/M5 AML who were no longereligible for chemotherapy. Although the number of patients inthis preliminary trial were low (n¼ 11), one complete and twopartial responders were observed.145
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1086
Leukemia

Pecularities of Raf kinase inhibitors
Most of the described experiments below have been performedin non-hematopoietic cells. However, they are presented in thisreview as Raf and MEK have also been considered as targets inleukemia and this information should be kept in mind whenconsidering targeting this pathway. It has been known for quitesome time now, that treatment with certain inhibitors (forexample, Raf inhibitors), can result in hyper-activation of wild-type (WT) Raf.8,146–148 It has recently become clear that it willbe essential to determine the genetic status at both BRAF andRAS before treatment with B-Raf selective inhibitors pro-ceeds.149 Class I B-Raf inhibitors (active conformation inhibi-tors) such as PLX4720 and 885-A (a close analog of SB590885)will inhibit BRAF mutants, however, these ATP-competitiveB-Raf inhibitors will not inhibit WT BRAF or mutant RAS. In factthese B-Raf inhibitors can activate Raf-1 in these cells in thepresence of active Ras. 885-A could induce B-Raf binding toRaf-1 and to a lesser extent PLX-4720 also could when the ERK-mediated negative feedback loop on B-Raf was inhibited with aMEK inhibitor. These binding events were determined to bedependent on the presence of oncogenic or growth factoractivated Ras, which may be required for translocation from thecytoplasm to the membrane and assembly into the signalingcomplex. Thus, this has therapeutic implications as in cells withmutant RAS, if the patient is treated with certain B-Raf inhibitors,B-Raf can bind and activate Raf-1 and promote the oncogenicpathway. In fact, even kinase-dead BRAF mutations, which arefound in human cancer, can dimerize with Raf-1, whenstimulated by the mutant Ras protein and activate the Raf/MEK/ERK cascade.
Also kinase-dead BRAF mutants form a constitutive complexwith Raf-1 in mutant RAS D04 cells, which activated MEKconstitutively. This activation was independent of inhibitortreatment. Thus, it is inactive B-Raf that forms a complex withRaf-1 and results in MEK activation. Additional experimentswere also performed with various conditional mouse mutantsthat demonstrated that only inactive B-Raf-bound Raf-1 in amutant Ras background. These studies provide a model of howresistance to these inhibitors could develop in patients treatedwith these B-Raf selective inhibitors. Such inhibitor treatmentcould result in additional cancers if the drug treatment select forcells in the patients, which have either a RAS mutation or amutation in an upstream receptor or signaling molecule thatactivates Ras. Hence, B-Raf selective inhibitors should not beadministered to patients with RAS mutations or mutations inupstream signaling components.
Also these results indicate a model where cells could becomeresistant to pan Raf inhibitors. If there was a gatekeeper mutationin Raf-1, which prevented inhibitor binding, the mutant B-Rafcould stimulate the mutant Raf-1 to allow inhibitor-resistantgrowth.
Pan-Raf inhibitors such as ZM336372, and RAF265 alsoinduced B-Raf binding to Raf-1 but did not activate ERK. This islikely because they also inhibit Raf-1. Sorafinib induced B-Rafbinding to Raf-1 and Raf-1 activation. However, sorafinibinhibited ERK activity.
Other studies on B-Raf inhibitors documented that other B-Rafinhibitors could induce Raf activity and that Raf activity can bestimulated in B-Raf-null tumor lines though Raf-1 dimeriza-tion.150 However, activation of MEK/ERK by the Raf inhibitorswas inhibited in cells containing the CRAF gatekeeper mutation,which cannot bind the Raf inhibitors. Finally, a third studysuggested that the binding of the Raf inhibitors to one Rafprotein results in the transactivation of the other Raf protein in
the dimer.151 These observations are dependent on Ras beingactive.151 Overall these results document the complexity withRaf inhibitor therapy, clearly for B-Raf selective inhibitors to betherapeutic useful previous screening of patients for Rasmutations will be required, as well as perhaps additionalscreening during treatment. Otherwise resistance may developand in some cases further stimulation of the Raf/MEK/ERKcascade.
Genes involved in sensitivity and resistance to MEK inhibitors
MEK inhibitors such as PD0329501 inhibit both MEK1 andMEK2 by locking the enzymes in a closed and catalyticallyinactive conformation.152 In MEK inhibitor-sensitive cells, MEKinhibitor treatment results in a dose-dependent decrease inactivated ERK1,2 levels as well as cyclin D expression. Incontrast, there is an increase in p27Kip�1 levels, RB hypospho-sphorylation and accumulation of cells in G1 phase of the cellcycle.153
By analyzing transcriptional pathway signatures, a panel of 18genes has been determined to be important in terms of MEKaddiction and response to the MEK inhibitor selumetinib(AZD6244, ARRY-142886).154 Importantly, this transcriptionalsignature was determined to be reproducible across differentcell panels of diverse tumor origins, even when the profiling wasperformed in different laboratories using different technologyplatforms. The genes they determined to be transcriptionalcommon events to MEK function are termed MEK-functional-activation signature and include: dual-specificity phosphatases(DUSP4/6), sprouty homologue 2 (SPRY2), pleckstrin homology-like domain family A member 1 (PHLD1), all the above threegenes were previously identified to be transcriptional targets ofMEK/ERK and involved in the downregulation of the pathway.Other transcriptional targets in the signature included the ETSvariant transcription factors ETV4, ETV5 and ELF1, and otherMEK family members such as MAP3K3. Finally, some genes inthe MEK inhibitor responsive signature were linked to regulationof MAPK singling, cell cycle and tumor progression wereidentified which included tribbles 2 (TRIB2) galectin 3 (LGALS3)and the transcription factors KANK1 (ANKRD15) and TS1. Theinvestigators also identified a 13-gene signature of genes, whichare involved in resistance to the MEK inhibitors when MEK isactive other than phosphatidyl inositol 3 kinase catalytic subunit(PIK3CA). These genes were determined to be reproduciblypredictive of resistance and entitled compensatory-resistancegenes. Some of these genes have a common linkage totransforming growth factor-b signaling. Others are known toregulate signaling pathways and may offer alternative routes tocell proliferation (for example, Frizzled homolog 2 (FZD2)).Some genes may enhance cell survival and chemoresistance bycontrolling hypoxia and angiogenesis (serine proteaseSERPINE1), lysyl oxidase LOX, collagens (COL5A1 andCOL1A2), cell cycle (GOG1 switch 2 (GOS2) proliferation/apoptosis (cysteine-rich transmembrane BMP regulator 1(CRIM1)) and immune evasion (CD274). Although most of thesestudies have been performed in non-hematopoietic cells, certainleukemias are known to be sensitive to MEK inhibitors althoughthe genetic mechanisms for the sensitivities have not been aswell elaborated. We present this information on MEK inhibitorsensitivity in non-hematopoietic cancers as an example of whatmay in the future be determined to result in MEK inhibitorsensitivity in leukemia.
In a different genomic screening analysis, genes associatedwith either BRAF mutations and sensitivity to MEK inhibitors or
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1087
Leukemia

with RTK mutations that conferred resistance to MEK inhibitorswere determined.155 The importance of this study is theformulation of the concept that different oncogenic mutationsin genes in the same pathway can result in divergent sets of geneexpression. Briefly, in cells with BRAF mutations, the cellsexpress a panel of genes, which are MEK/ERK dependent andsensitive to MEK inhibitors. Whereas in cells with RTKmutations, the cells express a different panel of genes, whichare MEK inhibitor insensitive. The investigators determined thatRaf/MEK/ERK signaling is activated in BRAF-mutant cells anddownregulated in RTK-mutant cells. In the BRAF-mutant cells,the Raf/MEK/ERK pathway was resistant to feedback down-regulation by downstream phosphatases, whose transcription issometimes dependent on activation of this pathway. This resultsin two different subtypes of cellular transformation originatingfrom mutations at genes in the same pathway. In the BRAF-mutant cells, they express elevated activated ERK and these cellsare insensitive (or less sensitive) to negative feedback regulationby phosphatases as the mutant BRAF protein is resistant to thenegative feedback initiated by activated ERK. In contrast, in theRTK-mutant cells, they express a different panel of genes and theRas/Raf/MEK/ERK pathway is downregulated and sensitive tofeedback regulation.
This study identified a panel of 52 genes whose expressionwas dependent on MEK/ERK activity in BRAF-mutant (V600E)cells. Some of the genes identified included: cyclin D1 (CCN1),transcription factors (MYC, FOS, FOSL1, EGF1, IER3, ETV1,ETV5, EV4 and MAFF, which were previously identified asERK-target genes. Interestingly, this study documented theimportance of feedback regulation of the Ras/Raf/MEK/ERKpathway as some of the genes identified were phosphatasesincluding: DUSP6, DUSP4, SPRY2, SPRY4 and SPRED2. Theinvestigators also documented the presence of 36 genes in theirprofile, which had not previously been associated with Raf/MEK/ERK signaling. Thus, in RTK-mutant cells, there is modestactivation of the Ras/Raf/MEK/ERK pathway, which is regulatedby phosphatases, whereas in the BRAF-mutant cells, there is ahigh level of activation of the Raf/MEK/ERK including theinduction of phosphatase gene expression, however, the mutantBRAF is not subject to downregulation by ERK.
Recently, it has been determined that certain mutations inMEK1 will cause resistance to both MEK and B-Raf selectiveinhibitors.156 By performing a MEK1 random mutagenesisscreen in vitro, mutations in MEK1 were identified whichconferred resistance to the MEK inhibitor AZD6244 (selumetinib).The complementary DNA library containing the MEK1 muta-tions were introduced into the MEK inhibitor-sensitive line A375melanoma cells, which harbor the BRAFV600E mutation andthen cultured the cells in the presence of MEK inhibitors (eitherselumetinib or CI-1040) for 4 weeks. Over 1000 MEK inhibitor-resistant clones emerged, pooled and the MEK1 gene wascharacterized en masse by massively parallel sequencing. MEKinhibitor-resistant alleles segregated into two different classes,those with mutations either within or affecting the allostericMEK inhibitor-binding pocket or those outside of the MEKinhibitor-binding pocket. Binding of the MEK inhibitors to thebinding pocket prevents the structural reorganization of theMEK1 protein and generates a catalytically inactive MEK1conformation. Arylamine MEK inhibitors lock MEK1 in a closedinactive conformation whereby the activation loop induceshelix C to be rotated and displaced.157 However, the MEK1mutants, which prevent MEK inhibitor-binding cause resistanceby preventing this altered conformation of MEK. It has beenknown that treatment of cells with this class of MEK inhibitorsresults in the detection of activated MEK1/2 but inactivated
downstream ERK1/2. The other class of MEK inhibitor-resistantmutants contained mutations present in two different regions ofMEK1. Some were present in the N-terminal negative regulatorydomain known as helix A (for example, Q56P) while otherswere present in a proline proximal to the C helix that abuts helixA (for example, P124S), which may enhance MEK1 activity. Themutations at Q56P and P124S in MEK1 are similar in biologicalconsequences to the mutations detected in cardio-facio-cuta-neous syndrome. The investigators concluded that resistance toMEK inhibitors can arise via reduction of drug binding (firstgroup) or enhanced MEK1 activation (second group). The effectsof these mutations on resistance to MEK inhibitors wasconfirmed by introducing the specific mutations into a MEK1complementary DNA by side directed mutagenesis, introduc-tion into A375 melanoma cells (normally sensitive to MEKinhibitors) and then determining the biochemical responses toMEK inhibitor treatment.
The presence of a MEK1 mutation was determined in amelanoma patient sample from a patient treated with selume-tinib, which had undergone relapse. This mutation occurred inP124L, which is functionally similar to the P124S mutationindentified by the cell line screening approach. The P124Lmutation was only present in the patient’s post-relapse sampleand not present in the pretreatment tumor sample. Melanomacultures (M307) were derived from the relapse patient’sselumetinib-resistant melanoma from their left axillary lymphnode. These cells contained the MEK1 P124L mutation and wereindeed more resistant to selumetinib (GL50 exceed 2 mM) thantreatment-naıve melanoma cultures containing the BRAF V600Emutation (GL50 10–50 nM).
The MEK1 mutation present in M307 cells derived from themelanoma patient treated with selumetinib who had undergonerelapse was determined to confer resistance to the selectiveB-Raf inhibitor (PLX4720) as these cells were highly resistant tothe B-Raf inhibitor. Thus, MEK1 inhibitor-resistant mutationsmay also confer cross-resistance to B-Raf inhibitors. Finally,these investigators determined that administration of B-Raf andMEK1 inhibitors simultaneously prevented the emergence ofresistant clones suggesting that this might be a therapeuticoption. Furthermore, the concept of dual targeting of MEK andPI3K pathways is arising because of the cross-regulation of manytargets of this pathway (for example, transcription factors,apoptotic regulatory molecules and proteins involved in theregulation of protein translation).158–160
Mutations within the RTK/Ras/Raf/MEK/ERK pathway thatalter sensitivity to MEK inhibitors
Mutations at the BRAF, KRAS, EGFR genes or the chromosomalfusion between ALK and ROS are detected in approximately50% of non-small cell lung cancer (NSCLC). NSCLC cells withmutations at BRAF where shown to be more sensitive to MEKinhibitors than NSCLC with mutations in EGFR, KRAS, or thechimeric fusion between ALK and ROS.158,161 This wasdetermined by screening a large panel of cell lines (n¼ 87)and tumors (n¼ 916). These results support the hypothesis thatmutations at BRAF are driver mutations, which are critical forthe initiation of the tumor. In this study, cells with mutations atEGFR were resistant to MEK inhibitors. This may have resultedfrom the ability of EGFR to activate the PI3K/PTEN/Akt/mTORpathway, which has some crucial overlapping targets as the Raf/MEK/ERK pathway. As expected BRAF-mutant cells were notsensitive to EGFR inhibitors. Thus, NSCLC patients with BRAFmutations should not be treated with EGFR inhibitors and
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1088
Leukemia

NSCLC patients with EGFR mutations should not be treated withMEK (or BRAF) inhibitors as the respective therapies would beineffective. The investigators developed a mass spectrometrybased genotyping method for the detection of hotspot mutationsin BRAF, KRAS and EGFR as NSCLC patients, which harborBRAF mutations should be treated differently than NSCLCpatients with EGFR tumors. Interestingly, the investigatorsdemonstrated that EGFR mutations are not necessarily asso-ciated with cigarette smoking in NSCLC, whereas BRAFmutations are associated with either current or former cigarettesmoking. Mutations in neurofibromin-1 (NF1) or MEK1 mayresult in cells, which are dependent on MEK signaling andsensitive to MEK inhibitors. MEK1 mutations are observed at alow frequency in NSCLC.162
Mutations at PIK3CA confer resistance to MEK inhibitors inKRAS-mutant tumors
Mutations at KRAS often confer sensitivity to MEK inhibitors.MEK inhibitor-sensitive tumors are usually WT at PIK3CA. Manytumors exist that have altered expression of both the Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways. These tumorswill usually be resistant to MEK inhibitors. This was examinedexperimentally by using pairs of isogenic cell lines (HCT116 andDLD-1) with mutant KRAS and either WT or mutant PIK3CA.163
Cells containing mutant PIK3CA were insensitive to MEKinhibitors whereas the KRAS mutant cells, which had WTPIK3CA in these tumors were sensitive to MEK inhibitors.Clinically, there are tumors (for example, melanomas) that havemutations at KRAS or BRAF and also have mutations at PIK3CAor deletions at PTEN. For successful inhibition of cell growth inthese tumors, inhibition of both pathways is necessary.However, not all MEK inhibitor-resistant cells have PIK3CA orPTEN mutations. In the study by Halilovic et al., elevatedactivated Akt levels were detected in the MEK inhibitor-resistantlines in the absence of PIK3CA and PTEN mutations, suggestingthat Akt was activated by a PI3K/PTEN-independent mechanism.Clearly, there are additional genes that confer inhibitor-resistance. These results have suggested that these two pathwayshave a common set of downstream components. One down-stream component is eIF4E-binding protein-1.160 Anotherimportant downstream target that both of these pathwaysconverge on is cyclin D1. Increased cyclin D1 expression willconfer B-Raf inhibitor resistance.163
Interactions between KRAS and PIK3CA mutations can resultin conferring resistance to rapamycin
Cancers containing PIK3CA mutations are often sensitive to themTOR inhibitor rapamycin and the modified rapamycins(Rapalogs). However, PIK3CA-mutant cells that also havemutations at KRAS are resistant to rapalogs.164 This may bedue to complicated feedback loops between the Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways wherein eithermTORC1 inhibition leads to ERK1/2 activation by a p70S6K/PI3K/Ras-dependent pathway or by the KRAS mutantsactivating p90Rsk-1, which serves to activate eIF4B and rpS6thereby bypassing mTOR-dependent activation. Although manyof the examples in the previous sections were from studies withsolid cancers, they serve to illustrate the complexity ofsensitivity and resistance to targeted therapy with Raf andMEK inhibitors.
Resistance to MEK inhibitors in leukemia
Although mutations in certain upstream cytokine receptors orcomponents of the Ras/Raf/MEK/ERK pathway often alter sensitivityto MEK inhibitors in leukemic cells, recently it has been shown thatcertain mutations will confer resistance to MEK inhibitors.165
NF1 is a GTPase activating protein, which normallyterminates Ras signaling via stimulation of Ras-GTP hydrolysis.NF1 is inactivated in children with NF1 and these children havea 200- to 500-fold increase in juvenile myelomonocyticleukemia.166,167 Dr Shannon et al. have developed mousemodels to examine the genes involved in transitions from MPDto AML. MPDs initiated by inactivating NF1 in mouse bonemarrow are resistant to MEK inhibitors as are myeloid progenitorcells from WT mice; however, subsequent AMLs induced in theNF1-deficient mice by retroviral insertional mutagenesisindicated that the NF1-deficient AMLs were sensitive to MEKinhibitors. These results indicated that the NF1-mutant AMLsprobably acquired additional mutations, which made themsensitive to Raf/MEK/ERK pathway signaling. However, resis-tance to the MEK inhibitors could develop in some NF1-deficient AML cells. The resistant cells were determined to bepresent in the original NF1-deficient-sensitive clones as deter-mined by the sites of retroviral integration. That is, there weresubsequent mutations in the original retroviral-induced NF1-deficient AML, which resulted in resistant AMLs and the resistantcells were selected on culture in media containing the MEKinhibitor. The sites of retroviral insertion were subsequentlycloned in the resistant cells. The candidate genes frequentlyimplicated in resistance to the MEK inhibitors were identified.One gene identified was Rasgrp1. Some MEK inhibitor-resistantclones displayed approximately 1000-fold increase in copynumber of Rasgrp1, which resulted in an average of 10-foldincrease in mRNA specific for this gene. Ras activation wasconstitutive in the MEK inhibitor-resistant cells, which formedcytokine-independent blast colonies in methylcellulose. Infec-tion of MEK inhibitor-resistant cells with a lentivirus containinga short hairpin RNA specific for Rasgrp1 reduced the RasGRP1protein and restored sensitivity to the MEK inhibitor.
Another gene identified whose altered expression resulted inresistance to the MEK inhibitor is Mapk14, which encodes p38a.In this case, the retrovirus integrated in the opposite readingframe to Mapk14, which resulted in inactivating one Mapk14allele. Drugs such as SB202190 (a specific p38a inhibitor)antagonized the inhibitor effects of the MEK inhibitors on theMEK inhibitor-sensitive NF1-deficient AMLs, but had onlyminimal effects on the MEK inhibitor-resistant NF1-deficientAMLs. The presence of resistant mutants present in the originalleukemias has also been observed with chronic myeloidleukemias and ALLs.168,169 The genes often detected in theresistant clones included genes involved in developmentalprograms, cell cycle control and DNA damage response. Incontrast, mutations/overexpression of genes involved in drugtransport or metabolism are not commonly observed, at least inthe drug-resistant ALLs.
Switching Raf isoforms can confer resistance to Raf inhibitors
In BRAF-mutant melanoma cells that had been maintained inmedium containing the B-Raf inhibitor AZ628 shifted theirdependence from B-Raf to Raf-1.170 Some of additional geneticmutations may be preexisting in the tumor cell population and onculture of the cells or tumor in the presence of the Raf inhibitor; the‘mutant-resistant’ cells may take over the population.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1089
Leukemia

Conclusions
The Ras/Raf/MEK/ERK cascade has important roles in promotinghematopoietic cell growth, regulating apoptosis, as well as theinduction of chemotherapeutic drug resistance. Commonly usedanti-leukemia chemotherapeutic drugs can induce this pathwayand may contribute to subsequent drug resistance. Recentstudies have also indicated that ERK and p90Rsk-1 can regulatecertain aspects of protein translation in conjunction with thePI3K/PTEN/Akt/mTOR pathway. Mutations that activate thiscascade are among the most frequently detected in humancancer. Inhibitors of Ras, Raf and MEK have been developed andanalyzed clinically. Although many of these inhibitors showpromise, there are also complications. This review has discussedsome of the genes that confer resistance/sensitivity to Ras, Rafand MEK inhibitors. Sometimes mutations at other signalingpathways such as PI3K/PTEN/Akt/mTOR (for example, PIK3CA)will inhibit the sensitivity to the inhibitors, which target the Ras/Raf/MEK/ERK pathway. Similarly, mutations at KRAS willeliminate the sensitivity of PIK3CA-mutant tumors to rapamycin.Other mutations have also been identified which either conferresistance or sensitivity to MEK inhibitors. Certain inhibitors (forexample, Raf inhibitors) may activate the Raf/MEK/ERK pathwaythat may be deleterious in cells, which have KRAS mutations.Thus, although we now have more precise bullets in our arsenalto treat leukemia than classical chemotherapy, we also havediscovered mechanisms by which the leukemic cells can protectpenetration and subsequent death by these bullets.
Conflict of interest
The authors declare no conflict of interest.
Acknowledgements
This work was supported in part by grants from Fondazione delMonte di Bologna e Ravenna, MinSan 2008, ‘Molecular therapyin pediatric sarcomas and leukemias against IGF-1 receptorsystem’ (PRIN 2008 and FIRB 2010 (RBAP10447J)) to AMM.
References
1 McCubrey J, LaHair M, Franklin RA. Reactive oxygen species-induced activation of the MAP kinase signaling pathway.Antioxidants Redox Signal 2006; 8: 1745–1748.
2 Franklin R, Rodriquez-Mora O, La Hair M, McCubrey JA.Activation of the calcium/calmodulin-dependent protein kinasesas a consequence of oxidative stress. Antioxidants Redox Signal2006; 8: 1807–1817.
3 McCubrey JA, Steelman LS, Kempf CR, Chappell WH, Abrams SL,Stivala F et al. Therapeutic resistance resulting from mutations inRaf/MEK/ERK and PI3K/PTEN/Akt/mTOR signaling pathways. JCell Physiol 2011; e-pub ahead of print 1 February 2011;doi:10.1002jcp22647.
4 McCubrey JA, Steelman LS, Chappell WH, Abrams SL, Wong EW,Chang F et al. Roles of the Raf/MEK/ERK pathway in cell growth,malignant transformation and drug resistance. Biochim BiophysActa 2007; 1773: 1263–1284.
5 Steelman LS, Abrams SL, Whelan J, Bertrand FE, Ludwig DE,Basecke J et al. Contributions of the Raf/MEK/ERK, PI3K/PTEN/Akt/mTOR and JAK/STAT pathways to leukemia. Leukemia 2008;22: 686–707.
6 McCubrey JA, Steelman LS, Abrams SL, Bertrand FE, Lugwig DE,Basecke J et al. Targeting survival vascades induced by activationof Raf/Raf/MEK/ERK, PI3K/PTEN/Akt/mTOR and Jak/STAT path-
ways for effective leukemia therapy. Leukemia 2008; 22: 708–722.
7 Misaghian N, Ligresti G, Steelman LS, Bertand FE, Basecke J,Libra M et al. Targeting the leukemic stem cellFthe holy grail ofleukemia therapy. Leukemia 2009; 23: 25–42.
8 McCubrey JA, Steelman LS, Abrams SL, Chappell WH, Russo S,Ove R et al. Emerging Raf inhibitors. Exp Opin Emerging Drugs2009; 14: 633–648.
9 McCubrey JA, Steelman LS, Abrams SL, Chappell WH, Russo S,Ove R et al. Emerging MEK inhibitors. Exp Opin Emerging Drugs2010; 15: 203–223.
10 McCubrey JA, Abrams SL, Ligresti G, Misaghian N, Wong ET,Basecke J et al. Involvement of p53 and Raf/MEK/ERKpathways in hematopoietic drug resistance. Leukemia 2008; 22:2080–2090.
11 Steelman LS, Abrams SL, Shelton JG, Chappell WH, Basecke J,Stivala F et al. Dominant roles of the Raf/MEK/ERK pathway incell cycle progression, prevention of apoptosis and sensitivity tochemotherapeutic drugs. Cell Cycle 2010; 9: 1629–1638.
12 Abrams SL, Steelman LS, Shelton JG, Wong ET, Chappell WH,Basecke J et al. The Raf/MEK/ERK pathway can govern drugresistance, apoptosis and sensitivity to targeted therapy. CellCycle 2010; 9: 1781–1791.
13 Kornblau SM, Womble M, Qiu YH, Jackson CE, Chen W,Konopleva M et al. Simultaneous activation of multiple signaltransduction pathways confers poor prognosis in acute myelo-genous leukemia. Blood 2006; 108: 2358–2365.
14 Casar B, Arozarena I, Sanz-Moreno V, Pinto A, Agudo-Ibanez L,Marais R et al. Ras subcellular localization defines extracellularsignal-regulated kinase 1 and 2 substrate specificity throughdistinct utilization of scaffold proteins. Mol Cell Biol 2009; 29:1338–1353.
15 Downward J. Targeting Ras signaling pathways in cancer therapy.Nat Rev Cancer 2003; 3: 11–22.
16 Hayashi K, Shibata K, Morita T, Iwasaki K, Watanabe M,Sobue K. Insulin receptor substrate-1/SHP-2 interaction, aphenotype-dependent switching machinery of insulin-like growthfactor-I signaling in vascular smooth muscle cells. J Biol Chem2004; 279: 40807–40818.
17 Marais R, Light Y, Paterson HF, Marshall CJ. Ras recruits Raf-1 tothe plasma membrane for activation by tyrosine phosphorylation.EMBO J 1995; 14: 3136–3145.
18 Marais R, Light Y, Paterson HF, Mason CS, Marshall CJ.Differential regulation of Raf-1, A-Raf, and B-Raf by oncogenicras and tyrosine kinases. J Biol Chem 1997; 272: 4378–4383.
19 Mason CS, Springer CJ, Cooper RG, Superti-Furga G, Marshall CJ,Marais R. Serine and tyrosine phosphorylations cooperate in Raf-1, but not B-Raf activation. EMBO J 1999; 18: 2137–2148.
20 XU S, Robbins D, Frost J, Dang A, Lange-Carter C, Cobb MH.MEKK1 phosphorylates MEK1 and MEK2 but does not causeactivation of mitogen-activated protein kinase. Proc Natl AcadSci USA 1995; 92: 6808–6812.
21 Derijard B, Raingeaud J, Barrett T, Wu IH, Han J, Ulevitch RJet al. Independent human MAP kinase signal transductionpathways defined by MEK and MKK isoforms. Science 1995;267: 682–685.
22 Xing J, Ginty DD, Greenberg ME. Coupling of the Ras-MAPKpathway to gene activation by Rsk2, a growth factor regulatedCREB kinase. Science 1996; 273: 959–963.
23 Davis RJ. Transcriptional regulation by MAP kinases. Mol ReprodDev 1995; 42: 459–467.
24 McCubrey JA, Steelman LS, Hoyle PE, Blalock WL, Weinstein-Oppenheimer C, Franklin RA et al. Differential abilities ofactivated Raf oncoproteins to abrogate cytokine-dependency,prevent apoptosis and induce autocrine growth factor synthesis inhuman hematopoietic cells. Leukemia 1998; 12: 1903–1929.
25 Hoyle PE, Moye PW, Steelman LS, Blalock WL, Franklin RA,Pearce M et al. Differential abilities of the Raf family of proteinkinases to abrogate cytokine-dependency and prevent apoptosisin murine hematopoietic cells by a MEK1-dependent mechanism.Leukemia 2000; 14: 642–656.
26 Blalock WL, Pearce M, Steelman LS, Franklin RA, McCarthy SA,Cherwinski H et al. A conditionally-active form of MEK1 results inautocrine transformation of human and mouse hematopoeiticcells. Oncogene 2000; 19: 526–536.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1090
Leukemia

27 Blalock WL, Moye PW, Chang F, Pearce M, Steelman LS,McMahon M et al. Combined effects of aberrant MEK1 activityand BCL2 overexpression on relieving the cytokine-dependencyof human and murine hematopoietic cells. Leukemia 2000; 14:1080–1096.
28 Moye PW, Blalock WL, Hoyle PE, Chang F, Franklin RA,Weinstein-Oppenheimer C et al. Between Raf and BCL2 inabrogating the cytokine-dependency of hematopoietic cells.Leukemia 2000; 14: 1060–1079.
29 Blalock WL, Pearce M, Chang F, Lee J, Pohnert S, Burrows C et al.Effects of inducible MEK1 activation on the cytokine-dependencyof lymphoid cells. Leukemia 2001; 15: 794–807.
30 Weinstein-Oppenheimer CR, Henrıquez-Roldan CF, Davis J,Navolanic PM, Saleh OA, Steelman LS et al. Role of the Raf signaltransduction cascade in the in vitro resistance to the anticancer drugdoxorubicin. Clin Cancer Res 2001; 7: 2892–2907.
31 Davis JM, Weinstein-Oppenheimer CR, Steelman LS, NavolanicPN, Hu W, Konopleva M et al. Raf-1 and Bcl-2 induce distinctand common pathways which contribute to breast cancer drugresistance. Clin Cancer Res 2003; 9: 1161–1170.
32 Balan V, Leicht DT, Zhu J, Balan K, Kaplun A, Singh-Gupta Vet al. Identification of novel in vivo Raf-1 phosphorylation sitesmediating positive feedback Raf-1 regulation by extracellularsignal-regulated kinase. Mol Biol Cell 2006; 17: 1141–1153.
33 Dougherty MK, Muller J, Ritt DA, Zhou M, Zhou XZ, CopelandTD et al. Regulation of Raf-1 by direct feedback phosphorylation.Mol Cell 2005; 17: 215–224.
34 Brummer T, Naegele H, Reth M, Misawa Y. Identification ofnovel ERK-mediated feedback phosphorylation sites at theC-terminus of B-Raf. Oncogene 2003; 22: 8823–8834.
35 Catalanotti F, Reyes G, Jesenberger V, Galabova-Kovacs G,de Matos Simoes R, Carugo O et al. A Mek1-Mek2 heterodimerdetermines the strength and duration of the Erk signal. Nat StructMol Biol 2009; 16: 294–303.
36 Dong C, Waters SB, Holt KH, Pessin JE. SOS phosphorylationand disassociation of the Grb2-SOS complex by the ERK and JNKsignaling pathways. J Biol Chem 1996; 271: 6328–6332.
37 Dhillon AS, von Kriegsheim A, Grindlay J, Kolch W. Phosphataseand feedback regulation of Raf-1 signaling. Cell Cycle 2007;6: 3–7.
38 von Kriegsheim A, Pitt A, Grindlay GJ, Kolch W, Dhillon AS.Regulation of the Raf-MEK-ERK pathway by protein phosphatase 5.Nat Cell Biol 2006; 8: 1011–1016.
39 Kim D, Rath O, Kolch W, Cho KH. A hidden oncogenic positivefeedback loop caused by crosstalk between Wnt and ERKpathways. Oncogene 2007; 26: 4571–4579.
40 Schaeffer HJ, Catling AD, Eblen ST, Collier LS, Krauss A, WeberMJ. MP1: a MEK binding partner that enhances enzymaticactivation of the MAP kinase cascade. Science 1998; 281: 1668–1671.
41 O’Neill E, Kolch W. Conferring specificity on the ubiquitous Raf/MEK signallingpathway. Br J Cancer 2004; 90: 283–288.
42 Kolch W. Coordinating ERK/MAPK signalling through scaffoldsand inhibitors. Nat Rev Mol Cell Biol 2005; 6: 827–837.
43 Hagan S, Garcia R, Dhillon A, Kolch W. Raf kinase inhibitorprotein regulation of raf and MAPK signaling. Methods Enzymol2006; 407: 248–259.
44 LaHair MM, Howe CJ, Rodriguez-Mora O, McCubrey JA,Franklin RA. Molecular pathways leading to oxidative stress-induced phosphorylation of Akt. Antioxidants Redox Signal 2006;8: 1749–1756.
45 Rodriguez-Mora O, LaHair MM, Howe CJ, McCubrey JA,Franklin RA. The calcium/calmodulin-dependent proteinkinases as potential targets in cancer therapy. Expert Opin TherTar 2005; 9: 791–808.
46 Weisberg E, Barrett R, Liu Q, Stone R, Gray N, Griffin JD. FLT3inhibition and mechanisms of drug resistance in mutant FLT3-positive AML. Drug Resist Updat 2009; 12: 81–89.
47 von Bubnoff N, Gorantla SP, Engh RA, Oliveira TM, Thone S,Aberg E et al. The low frequency of clinical resistance to PDGFRinhibitors in myeloid neoplasms with abnormalities of PDGFRAmight be related to the limited repertoire of possible PDGFRAkinase domain mutations in vitro. Oncogene 2010; 30:933–943.
48 Haslam K, Chadwick N, Kelly J, Browne P, Vandenberghe E,Flynn C et al. Incidence and significance of FLT3-ITD and NPM1mutations in patients with normal karyotype acute myeloidleukaemia. Ir J Med Sci 2010; 179: 507–510.
49 Shih LY, Huang CF, Wang PN, Wu JH, Lin TL, Dunn P et al.Acquisition of FLT3 or N-ras mutations is frequently associatedwith progression of myelodysplastic syndrome to acute myeloidleukemia. Leukemia 2004; 18: 466–475.
50 Bacher U, Haferlach T, Kern W, Haferlach C, Schnittger S. Acomparative study of molecular mutations in 381 patients withmyelodysplastic syndrome and in 4130 patients with acutemyeloid leukemia. Haematologica 2007; 92: 744–752.
51 Shih LY, Lin TL, Wang PN, Wu JH, Dunn P, Kuo MC et al. Internaltandem duplication of fms-like tyrosine kinase 3 is associatedwith poor outcome in patients with myelodysplastic syndrome.Cancer 2004; 101: 989–998.
52 Armstrong SA, Mabon ME, Silverman LB, Li A, Gribben JG, FoxEA et al. FLT3 mutations in childhood acute lymphoblasticleukemia. Blood 2004; 103: 3544–3546.
53 Perentesis JP, Bhatia S, Boyle E, Shao Y, Shu XO, Steinbuch Met al. RAS oncogene mutations and outcome of therapy forchildhood acute lymphoblastic leukemia. Leukemia 2004; 18:685–692.
54 Hou P, Liu D, Xing M. The T1790A BRAF mutation (L597Q) inchildhood acute lymphoblastic leukemia is a functional onco-gene. Leukemia 2007; 21: 2216–2218.
55 Tartaglia M, Martinelli S, Cazzaniga G, Cordeddu V, Iavarone I,Spinelli M et al. Genetic evidence for lineage-related anddifferentiation stage-related contribution of somatic PTPN11mutations to leukemogenesis in childhood acute leukemia. Blood2004; 104: 307–313.
56 Case M, Matheson E, Minto L, Hassan R, Harrison CJ, Bown Net al. Mutation of genes affecting the RAS pathway is common inchildhood acute lymphoblastic leukemia. Cancer Res 2008; 68:6803–6809.
57 Davidsson J, Lilljebjorn H, Panagopoulos I, Fioretos T, JohanssonB. BRAF mutations are very rare in B- and T-cell pediatric acutelymphoblastic leukemias. Leukemia 2008; 22: 1619–1621.
58 Gustafsson B, Angelini S, Sander B, Christensson B, Hemminki K,Kumar R. Mutations in the BRAF and N-ras genes in childhoodacute lymphoblastic leukaemia. Leukemia 2005; 19: 310–312.
59 Christiansen DH, Andersen MK, Desta F, Pedersen-Bjergaard J.Mutations of genes in the receptor tyrosine kinase (RTK)/RAS-BRAF signal transduction pathway in therapy-related myelodys-plasia and acute myeloid leukemia. Leukemia 2005; 19:2232–2240.
60 Pedersen-Bjergaard J, Andersen MK, Andersen MT, ChristiansenDH. Genetics of therapy-related myelodysplasia and acutemyeloid leukemia. Leukemia 2008; 22: 240–248.
61 Christiansen DH, Desta F, Andersen MK, Pedersen-Bjergaard J.Mutations of the PTPN11 gene in therapy-related MDS and AMLwith rare balanced chromosome translocations. Genes Chromo-somes Canc 2007; 46: 517–521.
62 Zebisch A, Staber PB, Delavar A, Bodner C, Hiden K, FischerederK et al. Two transforming C-RAF germ-line mutations identified inpatients with therapy-related acute myeloid leukemia. Cancer Res2006; 166: 3401–3408.
63 Makita Y, Narumi Y, Yoshida M, Niihori T, Kure S, Fujieda K et al.Leukemia in cardio-facio-cutaneous (CFC) syndrome: a patientwith a germline mutation in BRAF proto-oncogene.J Pediatr Hematol Oncol 2007; 29: 287–290.
64 McMahon M. RAF translocations expand cancer targets. Nat Med2010; 16: 749–750.
65 Palanisamy N, Ateeq B, Kalyana-Sundaram S, Pflueger D,Ramnarayanan K, Shankar S et al. Rearrangements of the RAFkinase pathway in prostate cancer, gastric cancer and melanoma.Nat Med 2010; 16: 793–798.
66 Borthakur G, Kantarjian H, Ravandi F, Zhang W, Konopleva M,Wright JJ et al. Phase 1 study of sorafenib in patients withrefractory or relapsed acute leukemias. Haematologica 2010; 96:62–68.
67 Crump M, Hedley D, Kamel-Reid S, Leber B, Wells R, BrandweinJ et al. A randomized phase I clinical and biologic study of twoschedules of sorafenib in patients with myelodysplastic syndromeor acute myeloid leukemia: a NCIC (National Cancer Institute of
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1091
Leukemia

Canada) Clinical Trials Group Study. Leuk Lymphoma 2011; 51:252–260.
68 Zhang W, Konopleva M, Ruvolo VR, McQueen T, Evans RL,Bornmann WG et al. Sorafenib induces apoptosis of AML cellsvia Bim-mediated activation of the intrinsic apoptotic pathway.Leukemia 2008; 22: 808–818.
69 Yeung K, Seitz T, Li S, Janosch P, McFerran B, Kaiser C et al.Suppression of Raf-1 kinase activity and MAP kinase signalling byRKIP. Nature 1999; 401: 173–177.
70 Yeung K, Janosch P, McFerran B, Rose DW, Mischak H, SedivyJM et al. Mechanism of suppression of the Raf/MEK/extracellularsignal-regulated kinase pathway by the Raf kinase inhibitorprotein. Mol Cell Biol 2000; 20: 3079–3085.
71 Corbit KC, Trakul N, Eves EM, Diaz B, Marshall M, Rosner MR.Activation of Raf-1 signaling by protein kinase C through amechanism involving Raf kinase inhibitory protein. J Biol Chem2003; 278: 13061–13068.
72 Cho KH, Shin SY, Kim HW, Wolkenbauer O, McFerran B,Kolch W. Mathematical modeling of the influence of RKIP on theERK signaling pathway. Comput Methods Syst Biol 2003, 127–141.
73 Park S, Rath O, Beach S, Xiang X, Kelly SM, Luo Z et al.Regulation of RKIP binding to the N-region of the Raf-1 kinase.FEBS Lett 2006; 580: 6405–6412.
74 Rath O, Park S, Tang HH, Banfield MJ, Brady RL, Lee YC et al.The RKIP (Raf-1 kinase inhibitor protein) conserved pocket bindsto the phosphorylated N-region of Raf-1 and inhibits the Raf-1-mediated activated phosphorylation of MEK. Cell Signal 2008;20: 935–941.
75 Shin SY, Rath O, Choo SM, Fee F, McFerran B, Kolch W et al.Positive- and negative-feedback regulations coordinate thedynamic behavior of the Ras-Raf-MEK-ERK signal transductionpathway. J Cell Sci 2009; 122: 425–435.
76 Beach S, Tang H, Park S, Dhillon AS, Keller ET, Kolch W et al.Snail is a repressor of RKIP transcription in metastatic prostatecancer cells. Oncogene 2008; 27: 2243–2248.
77 Yeung KC, Rose DW, Dhillon AS, Yaros D, Gustafsson M,Chatterjee D et al. Raf kinase inhibitor protein interacts with NF-kappaB-inducing kinase and TAK1 and inhibits NF-kappaBactivation. Mol Cell Biol 2001; 21: 7207–7217.
78 Odabaei G, Chatterjee D, Jazirehi AR, Goodglick L, Yeung K,Bonavida B. Raf-1 kinase inhibitor protein: structure, function,regulation of cell signaling, and pivotal role in apoptosis.Adv Cancer Res 2004; 91: 169–200.
79 Keller ET, Fu Z, Brennan M. The role of Raf kinase inhibitorprotein (RKIP) in health and disease. Biochem Pharmacol 2004;68: 1049–1053.
80 Hagan S, Al-Mulla F, Mallon E, Oien K, Ferrier R, Gusterson Bet al. Reduction of Raf-1 kinase inhibitor protein expressioncorrelates with breast cancer metastasis. Clin Cancer Res 2005;11: 7392–7397.
81 Baritaki S, Katsman A, Chatterjee D, Yeung KC, Spandidos DA,Bonavida B. Regulation of tumor cell sensitivity to TRAIL-inducedapoptosis by the metastatic suppressor Raf kinase inhibitorprotein via Yin Yang 1 inhibition and death receptor 5 up-regulation. J Immunol 2007; 179: 5441–5453.
82 Chatterjee D, Bai Y, Wang Z, Beach S, Mott S, Roy R et al. RKIPsensitizes prostate and breast cancer cells to drug-inducedapoptosis. J Biol Chem 2004; 279: 17515–17523.
83 Baritaki S, Yeung K, Palladino M, Berenson J, Bonavida B. Pivotalroles of snail inhibition and RKIP induction by the proteasomeinhibitor NPI-0052 in tumor cell chemoimmunosensitization.Cancer Res 2009; 69: 8376–8385.
84 Zebisch A, Haller M, Hiden K, Goebel T, Hoefler G, Troppmair Jet al. Loss of Raf kinase inhibitor protein is a somatic event in thepathogenesis of therapy-related acute myeloid leukemias withc-RAF germline mutations. Leukemia 2009; 23: 1049–1053.
85 Wilson BE, Mochon E, Boxer LM. Induction of bcl-2 expressionby phosphorylated CREB proteins during B-cell activation andrescue from apoptosis. Mol Cell Biol 1996; 16: 5546–5556.
86 Thomas A, El Rouby S, Reed JC, Krajewski S, Silber R, Potmesil Met al. Drug-induced apoptosis in B-cell chronic lymphocyticleukemia: relationship between p53 gene mutation andbcl-2/bax proteins in drug resistance. Oncogene 1996; 12:1055–1062.
87 Nuessler V, Stotzer O, Gullis E, Pelka-Fleischer R, Pogrebniak A,Gieseler F et al. Bcl-2, bax and bcl-xL expression in humansensitive and resistant leukemia cell lines. Leukemia 1999; 13:1864–1872.
88 Buggins AG, Pepper CJ. The role of Bcl-2 family proteins inchronic lymphocytic leukaemia. Leuk Res 2010; 34: 837–842.
89 Capitani N, Baldari CT. The Bcl-2 family as a rational target forthe treatment of B-cell chronic lymphocytic leukaemia.Curr Med Chem 2010; 17: 801–811.
90 McCubrey JA, May WS, Duronio V, Mufson A. Serine/threoninephosphorylation in cytokine signal transduction. Leukemia 2000;14: 9–21.
91 Deng X, Kornblau SM, Ruvolo PP, Way Jr WS. Regulation of Bcl2phosphorylation and potential significance for leukemic cellchemoresistance. J Natl Cancer Inst Monogr 2001; 28:30–37.
92 Harada H, Quearry B, Ruiz-Vela A, Korsmeyer SJ. Survival factor-induced extracellular signal-regulated kinase phosphorylatesBIM, inhibiting its association with BAX and proapoptoticactivity. Proc Natl Acad Sci USA 2004; 101: 15313–15317.
93 Domina AM, Vrana JA, Gregory MA, Hann SR, Craig RW. MCL1is phosphorylated in the PEST region and stabilized upon ERKactivation in viable cells, and at additional sites with cytotoxicokadaic acid or taxol. Oncogene 2004; 23: 5301–5315.
94 High LM, Szymanska B, Wilczynska-Kalak U, Barber N, O’Brien R,Khaw SL et al. The Bcl-2 homology domain 3 mimetic ABT-737targets the apoptotic machinery in acute lymphoblastic leukemiaresulting in synergistic in vitro and in vivo interactions withestablished drugs. Mol Pharmacol 2010; 77: 483–494.
95 Kuroda J, Kimura S, Andreeff M, Ashihara E, Kamitsuji Y, YokotaA et al. ABT-737 is a useful component of combinatorychemotherapies for chronic myeloid leukaemias with diversedrug-resistance mechanisms. Br J Haematol 2008; 140:181–190.
96 Stolz C, Hess G, Hahnel PS, Grabellus F, Hoffarth S, Schmid KWet al. Targeting Bcl-2 family proteins modulates the sensitivity ofB-cell lymphoma to rituximab-induced apoptosis. Blood 2008;112: 3312–3321.
97 Paoluzzi L, Gonen M, Bhagat G, Furman RR, Gardner JR, Scotto Let al. The BH3-only mimetic ABT-737 synergizes the antineo-plastic activity of proteasome inhibitors in lymphoid malignan-cies. Blood 2008; 112: 2906–2916.
98 Pepper C, Lin TT, Pratt G, Hewamana S, Brennan P, Hiller L et al.Mcl-1 expression has in vitro and in vivo significance in chroniclymphocytic leukemia and is associated with other poorprognostic markers. Blood 2008; 112: 3807–3817.
99 Zimmerman EI, Dollins CM, Crawford M, Grant S, Nana-SinkamSP, Richards KL et al. Lyn kinase-dependent regulation of miR181and myeloid cell leukemia-1 expression: implications for drugresistance in myelogenous leukemia. Mol Pharmacol 2010; 78:811–817.
100 Stewart ML, Fire E, Keating AE, Walensky LD. MCL-1 BH3 helixis an exclusive MCL-1 inhibitor and apoptosis sensitizer.Nat Chem Biol 2010; 6: 595–601.
101 Balakrishnan K, Burger JA, Wierda WG, Gandhi V. AT-101induces apoptosis in CLL B cells and overcomes stromalcell-mediated Mcl-1 induction and drug resistance. Blood2009; 113: 149–153.
102 Brunelle JK, Ryan J, Yecies D, Opferman JT, Letai A. MCL-1-dependent leukemia cells are more sensitive to chemotherapythan BCL-2-dependent counterparts. J Cell Biol 2009; 187:429–442.
103 Ley R, Ewings KE, Hadfield K, Cook SJ. Regulatory phosphoryla-tion of Bim: sorting out the ERK from JNK. Cell Death Differ 2005;12: 1008–1014.
104 Qi XJ, Wildey GM, Howe PH. Evidence that Ser87 of BimEL isphosphorylated by Akt and regulates BimEL apoptotic function.J Biol Chem 2006; 281: 813–823.
105 Gillings AS, Balmanno K, Wiggins CM, Johnson M, Cook SJ.Apoptosis and autophagy: BIM as a mediator of tumour cell deathin response to oncogene-targeted therapeutics. FEBS J 2009; 276:6050–6062.
106 Lei K, Davis RJ. JNK phosphorylation of Bim-related members ofthe Bcl2 family induces Bax-dependent apoptosis. Proc NatlAcad Sci USA 2003; 100: 2432–2437.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1092
Leukemia

107 Myatt SS, Lam EWF. The emerging roles of forkhead box (Fox)proteins in cancer. Nat Rev Cancer 2007; 7: 847–859.
108 Bachmann PS, Piazza RG, Janes ME, Wong NC, Davies C,Mogavero A et al. Epigenetic silencing of BIM in glucocorticoidpoor-responsive pediatric acute lymphoblastic leukemia, and itsreversal by histone deacetylase inhibition. Blood 2010; 116:3013–3022.
109 Hazlehurst LA, Argilagos RF, Dalton WS. Beta1 integrin mediatedadhesion increases Bim protein degradation and contributes todrug resistance in leukaemia cells. Br J Haematol 2007; 136:269–275.
110 Meads MB, Hazlehurst LA, Dalton WS. The bone marrowmicroenvironment as a tumor sanctuary and contributor to drugresistance. Clin Cancer Res 2008; 14: 2519–2526.
111 Nair RR, Tolentino J, Hazlehurst LA. The bone marrowmicroenvironment as a sanctuary for minimal residual diseasein CML. Biochem Pharmacol 2010; 80: 602–612.
112 Podar K, Chauhan D, Anderson KC. Bone marrow microenviron-ment and the identification of new targets for myeloma therapy.Leukemia 2009; 23: 10–24.
113 Kuribara R, Honda H, Matsui H, Shinjyo T, Inukai T, Sugita Ket al. Roles of Bim in apoptosis of normal and Bcr-Abl-expressinghematopoietic progenitors. Mol Cell Biol 2004; 24: 6172–6183.
114 Shinjyo T, Kuribara R, Inukai T, Hosoi H, Kinoshita T, Miyajima Aet al. Downregulation of Bim, a proapoptotic relative of Bcl-2, isa pivotal step in cytokine-initiated survival signaling in murinehematopoietic progenitors. Mol Cell Biol 2001; 21: 854–864.
115 Kuroda J, Puthalakath H, Cragg MS, Kelly PN, Bouillet P,Huang DC et al. Bim and Bad mediate imatinib-induced killing ofBcr/Abl+ leukemic cells, and resistance due to their loss isovercome by a BH3 mimetic. Proc Natl Acad Sci USA 2006; 103:14907–14912.
116 Essafi A, Fernandez de Mattos S, Hassen YA, Soeiro I, Mufti GJ,Thomas NS et al. Direct transcriptional regulation of Bim byFoxO3a mediates STI571-induced apoptosis in Bcr-Abl-expres-sing cells. Oncogene 2005; 24: 2317–2329.
117 Scheijen B, Ngo HT, Kang H, Griffin JD. FLT3 receptors withinternal tandem duplications promote cell viability and prolifera-tion by signaling through Foxo proteins. Oncogene 2004; 23:3338–3349.
118 Weisberg E, Sattler M, Ray A, Griffin JD. Drug resistance inmutant FLT3-positive AML. Oncogene 2010; 29: 5120–5134.
119 Chapuis N, Park S, Leotoing L, Tamburini J, Verdier F, Bardet Vet al. IkB kinase overcomes PI3K/Akt and ERK/MAPK to controlFOXO3a activity in acute myeloid leukemia. Blood 2010; 116:4240–4250.
120 Graff JR, Konicek BW, Carter JH, Marcusson EG. Targeting theeukaryotic translation initiation factor 4E for cancer therapy.Cancer Res 2008; 68: 631–664.
121 Chapuis N, Tamburini J, Green AS, Willems L, Bardet V, Park Set al. Perspectives on inhibiting mTOR as a future treatmentstrategy for hematological malignancies. Leukemia 2010; 24:1686–1699.
122 Sonenberg N, Hinnebusch AG. New modes of translationalcontrol in development, behavior and disease. Mol Cell 2007;28: 721–729.
123 Wendel HG, De Stanchina E, Friedman JS, Malina A, Ray S,Kogan S et al. Survival signalling by Akt and eIF4E in oncogenesisand cancer therapy. Nature 2004; 428: 332–337.
124 Roux PP, Shahbazian D, Vu H, Holz MK, Cohen MS, Taunton Jet al. RAS/ERK signaling promotes site-specific ribosomal proteinS6 phosphorylation via RSK and stimulates Cap-dependenttranslation. J Biol Chem 2007; 282: 14056–14064.
125 Shahbazian D, Roux PP, Mieulet V, Cohen MS, Raught B,Tauton J et al. The mTOR/PI3K and MAPK pathways converge oneIF4B to control its phosphorylation and activity. EMBO J 2006;25: 2781–2791.
126 Bordeleau ME, Robert F, Gerard B, Lindqvist L, Chen SM, WendelHG et al. Therapeutic suppression of translation initiationmodulates chemosensitivity in a mouse lymphoma model. J ClinInvest 2008; 118: 2651–2660.
127 Malina A, Mills JR, Zhu H, Ueda T, Watanabe-Fukunaga R,Fukunaga R et al. Dissecting eIF4E action in tumorigenesis.Genes Dev 2007; 21: 3232–3237.
128 Waskiewicz AJ, Flynn A, Proud CG, Cooper JA. Mitogen-activated protein kinases activate the serine/threonine kinasesMnk1 and Mnk2. EMBO J 1997; 16: 1909–1920.
129 Waskiewicz AI, Hohnson JC, Penn B, Mahalingam M,Kimball SR, Cooper JA. Phosphoryaltion of the cap-bindingprotein eukaryotic translation initation factor 4E by protein kinaseMnk in vivo. Mol Cell Biol 1999; 19: 1871–1880.
130 Wendel HG, Silva RL, Malina A, Mills JR, Zhu H, Ueda T et al.Dissecting eIF4E action in tumorigenesis. Genes Dev 2007; 21:3232–3237.
131 Silva RL, Wendel HG. MNK, EIF4E and targeting translation fortherapy. Cell Cycle 2008; 7: 553–555.
132 Ueda T, Watanabe Fukunga R, Fujuyama H, Nagata S, Fukunga R.Mnk2 and Mnk1 are essential for constitutive and induciblephosphorylation of eukaryotic initiation factor 4E but not for cellgrowth or development. Mol Cell Biol 2004; 24: 6539–6549.
133 Ueda T, Sasaki M, Elia AJ, Chio II, Hamada K, Fukunaga R et al.Combined deficiency for MAP kinase-interacting kinase 1 and 2(Mnk1 and Mnk2) delays tumor development. Proc Natl Acad SciUSA 2010; 107: 13984–13990.
134 Nardella C, Chen Z, Salmena L, Carracedo A, Alimonti A, Egia Aet al. Aberrant Rheb-mediated mTORC1 activation and Ptenhaploinsufficiency are cooperative oncogenic events. Genes Dev2008; 22: 2172–2177.
135 Lassman AB, Rossi MR, Raizer JJ, Abrey LE, Lieberman FS, GrefeCN et al. Molecular study of malignant gliomas treated withepidermal growth factor receptor inhibitors: tissue analysis fromNorth American Brain Tumor Consortium Trials 01-03 and00-01. Clin Cancer Res 2005; 11: 7841–7850.
136 Leseux L, Hamdi SM, Al Saati T, Capilla F, Recher C, Laurent Get al. Syk-dependent mTOR activation in follicular lymphomacells. Blood 2006; 108: 4156–4162.
137 Peponi E, Drakos E, Reyes G, Leventaki V, Rassidakis GZ,Medeiros LJ. Activation of mammalian target of rapamycinsignaling promotes cell cycle progression and protects cells fromapoptosis in mantle cell lymphoma. Am J Pathol 2006; 169:2171–2180.
138 Lu ZH, Shvartsman MB, Lee AY, Shao JM, Murray MM, KladneyRD et al. Mammalian target of rapamycin activator RHEB Isfrequently overexpressed in human carcinomas and is critical andsufficient for skin epithelial carcinogenesis. Cancer Res 2010; 70:3287–3298.
139 Lin AW, Barradas M, Stone JC, van Aelst L, Serrano M, Lowe SW.Premature senescence involving p53 and p16 is activated inresponse to constitutive MEK/MAPK mitogenic signaling. GenesDev 1998; 12: 3008–3019.
140 Mavrakis KJ, Zhu H, Silva RL, Mills JR, Teruya-Feldstein J, LoweSW et al. Tumorigenic activity and therapeutic inhibition of RhebGTPase. Genes Dev 2008; 22: 2178–2288.
141 Moerke NJ, Aktas H, Chen H, Cantel S, Reibarkh MY, Fahmy Aet al. Small-molecule inhibition of the interaction between thetranslation initiation factors eIF4E and eIF4G. Cell 2007; 128:257–267.
142 Tamburini J, Green AS, Bardet V, Chapuis N, Park S, Willems Let al. Protein synthesis is resistant to rapamycin and constitutes apromising therapeutic target in acute myeloid leukemia. Blood2009; 114: 1618–1627.
143 Kentsis A, Topisirovic I, Culjkovic B, Shao L, Borden KL. Ribavirinsuppresses eIF4E-mediated oncogenic transformation by physicalmimicry of the 7-methyl guanosine mRNA cap. Proc Natl AcadSci USA 2004; 101: 18105–18110.
144 Kentsis A, Volpon L, Topisirovic I, Soll CE, Culjkovic B, Shao Let al. Further evidence that ribavirin interacts with eIF4E. RNA2005; 11: 1762–1766.
145 Assouline S, Culjkovic B, Cocolakis E, Rousseau C, Beslu N,Amri A et al. Molecular targeting of the oncogene eIF4E in acutemyeloid leukemia (AML): a proof-of-principle clinical trial withribavirin. Blood 2009; 114: 257–260.
146 Hall-Jackson CA, Eyers PA, Cohen P, Goedert M, Boyle FT,Hewitt N et al. Paradoxical activation of Raf by a novel Rafinhibitor. Chem Biol 1999; 6: 559–568.
147 Lee Jr JT, McCubrey JA. Targeting the Raf kinase cascade incancer therapyFnovel molecular targets and therapeutic strate-gies. Expert Opin Ther Tar 2002; 6: 659–678.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1093
Leukemia

148 Shelton JG, Moye PW, Steelman LS, Blalock WL, Lee JT, FranklinRA et al. Differential effects of kinase cascade inhibitors onneoplastic and cytokine-mediated cell proliferation. Leukemia2003; 17: 1765–1782.
149 Heidorn SJ, Milagre C, Whittaker S, Nourry A, Niculescu-Duvas I,Dhomen N et al. Kinase-dead BRAF and oncogenic RAScooperate to drive tumor progression through CRAF. Cell 2010;140: 209–221.
150 Hatzivassiliou G, Song K, Yen I, Brandhuber BJ, Anderson DJ,Alvarado R et al. RAF inhibitors prime wild-type RAF to activatethe MAPK pathway and enhance growth. Nature 2010; 464:431–435.
151 Poulikakos PI, Zhang C, Bollag G, Shokat KM, Rosen N. RAFinhibitors transactivate RAF dimers and ERK signalling in cellswith wild-type BRAF. Nature 2010; 464: 427–430.
152 Sebolt-Leopold JS, English JM. Mechanisms of drug inhibition ofsignaling molecules. Nature 2006; 441: 457–462.
153 Pratilas CA, Hanrahan AJ, Halilovic E, Persaud Y, Soh J, Chitale Det al. Genetic predictors of MEK dependence in non-small celllung cancer. Cancer Res 2008; 68: 9375–9383.
154 Dry JR, Pavey S, Pratilas CA, Harbron C, Runswick S, Hodgson Det al. Transcriptional pathway signatures predict MEK addictionand response to selumetinib (AZD6244). Cancer Res 2010; 70:2264–2273.
155 Pratilas CA, Taylor BS, Ye Q, Viale A, Sander C, Solit DB et al.V600E)BRAF is associated with disabled feedback inhibition ofRAF-MEK signaling and elevated transcriptional output of thepathway. Proc Natl Acad Sci USA 2009; 106: 4519–4524.
156 Emery CM, Vijayendran KG, Zipser MC, Sawyer AM, Niu L,Kim JJ et al. MEK1 mutations confer resistance to MEK andB-RAF inhibition. Proc Natl Acad Sci USA 2009; 106:20411–20416.
157 Ohren JF, Chen H, Pavlovsky A, Whitehead C, Zhang E, Kuffa Pet al. Structures of human MAP kinase kinase 1 (MEK1) and MEK2describe novel noncompetitive kinase inhibition. Nat Struct MolBiol 2004; 11: 1192–1197.
158 Halilovic E, She QB, Ye Q, Pagliarini R, Sellers WR, Solit DBet al. PIK3CA mutation uncouples tumor growth and cyclin D1regulation from MEK/ERK and mutant KRAS signaling. Cancer Res2010; 70: 6804–6814.
159 Joseph EW, Pratilas CA, Poulikakos PI, Tadi M, Wang W, TaylorBS et al. The RAF inhibitor PLX4032 inhibits ERK signaling andtumor cell proliferation in a V600E BRAF-selective manner. ProcNatl Acad Sci USA 2010; 107: 14903–14908.
160 She QB, Halilovic E, Ye Q, Zhen W, Shirasawa S, Sasazuki Tet al. 4E-BP1 is a key effector of the oncogenic activation of theAKT and ERK signaling pathways that integrates their function intumors. Cancer Cell 2010; 18: 39–51.
161 Pratilas CA, Solit DB. Targeting the mitogen-activated proteinkinase pathway: physiological feedback and drug response.Clin Cancer Res 2010; 16: 3329–3334.
162 Marks JL, Gong Y, Chitale D, Golas B, McLellan MD, Kasai Yet al. Novel MEK1 mutation identified by mutational analysis ofepidermal growth factor receptor signaling pathway genes in lungadenocarcinoma. Cancer Res 2008; 68: 5524–5528.
163 Smalley KS, Lioni M, Dalla Palma M, Xiao M, Desai B, Egyhazi Set al. Increased cyclin D1 expression can mediate BRAF inhibitorresistance in BRAF V600E-mutated melanomas. Mol Cancer Ther2008; 7: 2876–2883.
164 Di Nicolantonio F, Arena S, Tabernero J, Grosso S, Molinari F,Macarulla T et al. Deregulation of the PI3K and KRAS signalingpathways in human cancer cells determines their response toeverolimus. J Clin Invest 2010; 120: 2858–2866.
165 Lauchle JO, Kim D, Le DT, Akagi K, Crone M, Krisman K et al.Response and resistance to MEK inhibition in leukaemias initiatedby hyperactive Ras. Nature 2009; 461: 411–414.
166 Lauchle JO, Braun BS, Loh ML, Shannon K. Inherited predisposi-tions and hyperactive Ras in myeloid leukemogenesis. PediatrBlood Cancer 2006; 46: 579–585.
167 Le DT, Kong N, Zhu Y, Lauchle JO, Aiyigari A, Braun BS et al.Somatic inactivation of Nf1 in hematopoietic cells resultsin a progressive myeloproliferative disorder. Blood 2004; 103:4243–4250.
168 Shah NP, Nicoll JM, Nagar B, Gorre ME, Paquette RL, Kuriyan Jet al. Multiple BCR-ABL kinase domain mutations conferpolyclonal resistance to the tyrosine kinase inhibitor imatinib(STI571) in chronic phase and blast crisis chronic myeloidleukemia. Cancer Cell 2002; 2: 117–125.
169 Choi S, Henderson MJ, Kwan E, Beesley AH, Sutton R, Bahar AYet al. Relapse in children with acute lymphoblastic leukemiainvolving selection of a preexisting drug-resistant subclone.Blood 2007; 110: 632–639.
170 Montagut C, Sharma SV, Shioda T, McDermott U, Ullman M,Ulkus LE et al. Elevated CRAF as a potential mechanism ofacquired resistance to BRAF inhibition in melanoma. Cancer Res2008; 68: 4853–4861.
Roles of Ras/Raf pathway in therapeutic sensitivityLS Steelman et al
1094
Leukemia