Embed Size (px)
Citation preview

Root Water Uptake and Soil Water Distribution: Test of an Availability Concept1
RJ. LASCANO AND C.H.M. VAN BAVEI?ABSTRACT
An experimental test was made of the hypothesis that the frac-tional water uptake by the root system from a soil layer is propor-tional to the root density and to the difference between the waterpotential in that layer and the mean leaf water potential. This theoryadequately explained the water uptake by a cotton root system thatwas divided over a wet and a dry soil compartment. It was alsoestablished that the absolute rate of water uptake from each com-partment could be accurately calculated from the measured potentialdifference between soil and plant leaf, and from a separately mea-sured plant hydraulic resistance, which was shown to have a valueof 0.67 X 10' s, independent of transpiration rate.
Additional Index Words: plant resistance, split root, drought stress,modeling, transpiration.
Lascano, R. J., and C. H. M. van Bavel. 1984. Root water uptakeand soil water distribution: test of an availability concept. Soil Sci.Soc. Am. J. 48:233-237.
PROGRESSIVE DECLINE OF soil water supplies in theroot zone causes a corresponding lowering of the
water potential in the plant and symptoms collectivelydescribed as drought stress. Similar symptoms can becaused by other factors such as high transpiration rates,diseases of the root system, or unfavorable tempera-tures or aeration in the root zone. Drought stress isrelated only to unavailability of soil water in the rootzone in this study.
Originally a given quantity of water per unit soilvolume would be considered totally available, if thesoil water potential was above —1.5 MPa (Veihmeyerand Hendrickson, 1927). In current thought, soil wateravailability is believed to be more closely related toplant water potential. Hence, water availability variesover a wide range of values of the soil water potential(Slatyer, 1957; Gardner and Nieman, 1964; Van Bavelet al., 1968; Hsiao et al., 1976).
Furthermore, there is a need to describe water up-take by root systems in terms of the interaction be-tween the atmospheric demand and the actual distri-butions of root density and of water content in theroot zone, because a single value cannot describe thesoil water potential, nor the root density in the rootzone.
Taylor (1952) recognized the necessity of "integrat-ing" the availability of water over the entire root zoneon the basis of soil water potential. However, themethod proposed did not account for the rate of tran-spiration or the nature of the root system. Van Bavel(1953) proposed that an effective or average soil waterpotential threshold, at which drought stress would oc-cur, should be derived from experimental evidence.Slatyer (1957), and Gardner and Nieman (1964) pro-posed that such a limiting soil water potential should
1 Contribution from the Texas Agric. Exp. Stn., Texas A&M Univ.,College Station, TX 77843.2 Research Associate and Professor, respectively, Dep. of Soil andCrop Sciences, Texas Agric. Exp. Stn. Received 16 May 1983. Ap-proved 11 Oct. 1983.
be seen as a dynamic value, depending on the tran-spiration rate, an idea demonstrated experimentallyby Denmead and Shaw (1962). Cowan (1965) pro-posed the concept of a crop hydraulic resistance, andargued that its value per m2 of land area ought to bearound 5000 d (0.4 X 109 s). A number of experi-ments, with different crop plants and using variousmethods, were summarized by Lascano (1982), whoconcluded that the average measured value in 16 sep-arate studies, made between 1909 and 1981, was 1.0X 109s, with a CV of 50%.
Assuming plant hydraulic resistance (SRPL) can betaken as constant, the soil water and the plant waterpotential are related as follows:
<A, = fa - T-SRPL [1]in which fa is the plant water potential, fa the waterpotential at the root periphery, both in meters watercolumn, T the transpiration rate per unit leaf area inm3/(rn2-s), and SRPL in 5. The use of volume unitsis practical (100 m equals approximately 1 MPa), be-cause it simplifies equations.
With normal root densities, in nonsaline soils, andnonlimiting water potentials, the transpiration streamdoes not induce differences between the bulk soil waterpotential fa and fa, that are comparable to those be-tween fa and fa (Newman, 1969 and Hillel et al., 1975).This can be explained, in part, by the fact that stomataclose at low values of 0,. Therefore, we assumed thatthe resistance to water flow in the soil is negligiblecompared to that in the plant.
A number of plant water uptake models have beenproposed based on Eq. [1]. In an early model, VanBavel (1974a) neglected the inhomogeneity of the rootsystem, but introduced environmental and stomatalcontrol of transpiration. Molz and Remson (1970) andNimah and Hanks (1973a, 1973b) proposed a depth-dependent root extraction term. A number of similarmodels that deal with root water extraction from alayered soil, and its effect on canopy transpiration andcrop performance, have been reported in Feddes(1981), in Molz (198 la, 1981b), and in Lascano (1982).These later models did not deal adequately with thedependence of canopy transpiration upon the atmos-pheric environment and the surface energy balance,although methods for doing so had been proposed byLemon et al. (1971).
HYPOTHESISAn integration of multilayered models of both canopy and
root zone was proposed by Van Bavel (1974b) through adefining equation and a hypothesis. The denning equationformulated an effective leaf water potential, <t>e,, in m, as theaverage leaf water potential of the canopy weighted by thetranspiration rate in each canopy layer. The hypothesis cal-culates the partition of the total transpiration (water uptake)rate over the root zone, by stating
Rj = fa - faWDj/SKPL . [2]In any one root zone layer, or compartment (/). the rate ofwater extraction from the root zone (RJt in m/s) equals the
233
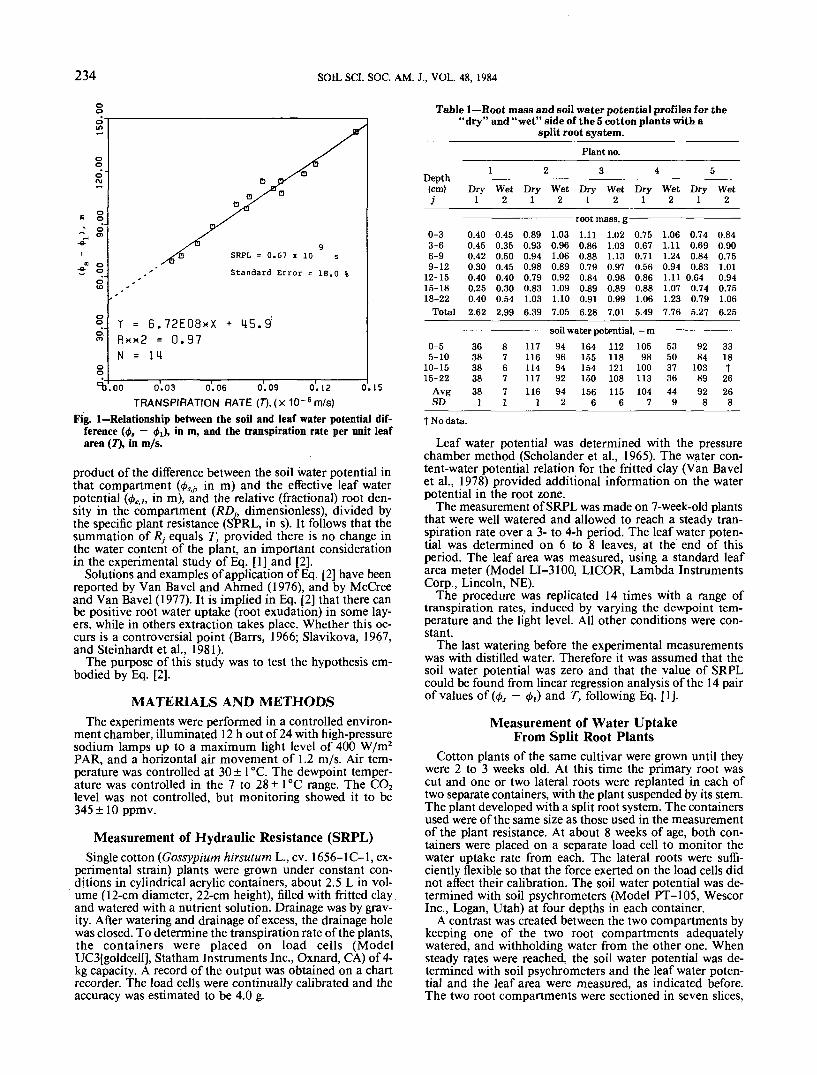
234 SOIL SCI. SOC. AM. J., VOL. 48, 1984
SRPL = 0.67 x 10 s
Standard Error = 18.0 %
Y = 6 . 7 2 E 0 8 x XR**2 = 0.97N = 14
) 0.03 0.06 0.09 0.12 0.15
TRANSPIRATION RATE (7"),(X 1Q-6m/S)Fig. 1—Relationship between the soil and leaf water potential dif-
ference (<t>, — <t>i), in m, and the transpiration rate per unit leafarea (T), in m/s.
product of the difference between the soil water potential inthat compartment (<t>sj, in m) and the effective leaf waterpotential (<j>ej, in m), and the relative (fractional) root den-sity in the compartment (RDj, dimensionless), divided bythe specific plant resistance (SPRL, in s). It follows that thesummation of Rj equals T, provided there is no change inthe water content of the plant, an important considerationin the experimental study of Eq. [1] and [2].
Solutions and examples of application of Eq. [2] have beenreported by Van Bavel and Ahmed (1976), and by McCreeand Van Bavel (1977). It is implied in Eq. [2] that there canbe positive root water uptake (root exudation) in some lay-ers, while in others extraction takes place. Whether this oc-curs is a controversial point (Barrs, 1966; Slavikova, 1967,and Steinhardt et al., 1981).
The purpose of this study was to test the hypothesis em-bodied by Eq. [2].
MATERIALS AND METHODSThe experiments were performed in a controlled environ-
ment chamber, illuminated 12 h out of 24 with high-pressuresodium lamps up to a maximum light level of 400 W/m2
PAR, and a horizontal air movement of 1.2 m/s. Air tem-perature was controlled at 30 ± 1 °C. The dewpoint temper-ature was controlled in the 7 to 28 ± 1°C range. The CO2level was not controlled, but monitoring showed it to be345±10ppmv.
Measurement of Hydraulic Resistance (SRPL)Single cotton (Gossypium hirsutum L, cv. 1656-1C-1, ex-
perimental strain) plants were grown under constant con-ditions in cylindrical acrylic containers, about 2.5 L in vol-ume (12-cm diameter, 22-cm height), filled with fritted clayand watered with a nutrient solution. Drainage was by grav-ity. After watering and drainage of excess, the drainage holewas closed. To determine the transpiration rate of the plants,the containers were placed on load cells (ModelUC3fgoldcell], Statham Instruments Inc., Oxnard, CA) of 4-kg capacity. A record of the output was obtained on a chartrecorder. The load cells were continually calibrated and theaccuracy was estimated to be 4.0 g.
Table 1—Root mass and soil water potential profiles for the"dry" and "wet" side of the 5 cotton plants with a
split root system.Plant no.
1 2 3 4 5Depth —————— —————— —————— —————— ——————(cm) Dry Wet Dry Wet Dry Wet Dry Wet Dry Wetj 1 2 1 2 1 2 1 2 1 2
0-33-66-99-12
12-1515-1818-22
Total
0-55-10
10-1515-22
AvgSD
0.400.450.420.300.400.250.402.62
36383838381
0.450.350.500.450.400.300.542.99
876771
0.890.930.940.980.790.831.036.39
117116114117116
1
1.030.961.060.890.921.091.107.05
root mass, g1.110.860.880.790.840.890.916.28
1.021.031.130.970.980.890.997.01
0.750.670.710.560.860.881.065.49
soil water potential, — m94969492942
164155154150156
6
112118121108115
6
10598
100113104
7
1.061.111.240.941.111.071.237.76
53503736449
0.740.690.840.83
0.640.740.795.27
9284
10389928
0.840.900.751.010.940.751.066.25
3318t
26268
t No data.
Leaf water potential was determined with the pressurechamber method (Scholander et al., 1965). The water con-tent-water potential relation for the fritted clay (Van Bavelet al., 1978) provided additional information on the waterpotential in the root zone.
The measurement of SRPL was made on 7-week-old plantsthat were well watered and allowed to reach a steady tran-spiration rate over a 3- to 4-h period. The leaf water poten-tial was determined on 6 to 8 leaves, at the end of thisperiod. The leaf area was measured, using a standard leafarea meter (Model LI-3100, LICOR, Lambda InstrumentsCorp., Lincoln, NE).
The procedure was replicated 14 times with a range oftranspiration rates, induced by varying the dewpoint tem-perature and the light level. All other conditions were con-stant.
The last watering before the experimental measurementswas with distilled water. Therefore it was assumed that thesoil water potential was zero and that the value of SRPLcould be found from linear regression analysis of the 14 pairof values of (0, — <£,) and T, following Eq. [1].
Measurement of Water UptakeFrom Split Root Plants
Cotton plants of the same cultivar were grown until theywere 2 to 3 weeks old. At this time the primary root wascut and one or two lateral roots were replanted in each oftwo separate containers, with the plant suspended by its stem.The plant developed with a split root system. The containersused were of the same size as those used in the measurementof the plant resistance. At about 8 weeks of age, both con-tainers were placed on a separate load cell to monitor thewater uptake rate from each. The lateral roots were suffi-ciently flexible so that the force exerted on the load cells didnot affect their calibration. The soil water potential was de-termined with soil psychrometers (Model PT-105, WescorInc., Logan, Utah) at four depths in each container.
A contrast was created between the two compartments bykeeping one of the two root compartments adequatelywatered, and withholding water from the other one. Whensteady rates were reached, the soil water potential was de-termined with soil psychrometers and the leaf water poten-tial and the leaf area were measured, as indicated before.The two root compartments were sectioned in seven slices,

LASCANO & VAN BAVEL: ROOT WATER UPTAKE AND SOIL WATER DISTRIBUTION: AN AVAILABILITY CONCEPT 235
Table 2—Average soil water potential and measured andcalculated transpiration rates for the "dry" and "wet"side of the five cotton plants with a split root system.
Plantno.j
12345
Average soilwater potential
Dry1
3811615610492
Wet2
794
1154426
Transpiration rate
Measured
Dry1
4.441.031.131.440.99
Wet2
7.772.514.276.878.88
Calculatedt
Dry1
x 10-" ——4.030.851.201.571.50
Wet2
7.052.664.557.327.10
Table 3—Ratio of water uptake from the dry and the wet side offive plants with a split root system, as measured and
as calculated with Eq. |2|.____________Water uptake (dry/wet)
t Using the proposed model of water uptake |Eq. (2)].
and the soil water content determined in each, as well as thetotal mass and total length of the root system in each slice.The purpose of this sectioning at the end of the experimentwas to verify the homogeneity of each container with respectto root length and water content as a function of depth. Theroot length was determined by the line transect method(Newman, 1966).
Using the measured values of SRPL, of <j>Cih of $v, and ofRDj, the value of .ft, was calculated to give the water uptakefrom the "dry" (j—1) and from the "wet" (j=2) half of theroot system. The terms "wet" and "dry" were quantified bycalculating the average root zone water potential for each ofthe two compartments. The split-root experiment was rep-licated five times.
RESULTS AND DISCUSSIONHydraulic Resistance of Cotton Plants (SRPL)
The total leaf area of the 14 test plants varied from800 to 2200 cm2 per plant, and the transpiration ratefrom 0.018 to 0.072 kg/h per plant. The transpirationrate per unit leaf area varied from 0.14 to 0.50 mm/h, which is comparable to well-watered fully devel-oped stands of cotton plants in representative daytimeconditions.
Figure 1 shows a plot of the transpiration rate perunit leaf area against the water potential differencebetween the soil and leaf, which in this case is theaverage leaf water potential, and the equation of thelinear regression line. The linear model is adequate,with the value of SRPL being 0.67 X 109 s ± 18%.This result shows that the hydraulic resistance of thecotton plants is independent of the transpiration rate,and that its value is within the range of values ab-stracted from the literature. The intercept, however,is not zero, but 45.9 m. For our purpose this aspectof the data is irrelevant, and we accept the value ofSRPL as a constant.
The measurement of SRPL is critical for the vali-dation of our hypothesis and has been attempted byothers in the past with conflicting outcomes (e.g.,Hailey et al., 1973; Weatherley, 1976). It is important,as shown by Bunce (1978), for example, that a steadystate first be reached in which the rate of root wateruptake equals that of transpiration. It is usually nec-essary to wait at least 3 h after a constant conditionis established in regard to the plant environment, whichexplains why this measurement cannot be done ac-curately outside, without some correction (Jones et al.,1982;Zuretal., 1982).
Plantno.
12345AvgSD
Measuredratio
0.570.410.260.210.11
Calculatedratio
0.570.320.260.200.21
Meas/Calcratio
1.001.281.001.050.520.970.28
Split-Root Plant Water UptakeIn the five split-root experiments, the leaf area var-
ied from 750 to 2200 cm2 per plant and the transpi-ration rate from 0.030 to 0.066 kg/h per plant, entirelycomparable to the plants used for the measurementofSRPL.
Table 1 gives the mass of roots in each of the sevenslices in which both the "dry" and the "wet" com-partments were divided, as well as the correspondingmeasured soil water potential values. The total andthe average value, respectively, is also shown for eachcompartment. It may be seen that a clearly distin-guishable soil water potential contrast was achievedbetween "dry" and "wet", except in plant no. 2 wherethe difference was only 22 m. The root mass was alsofairly uniform with depth, considering the limited ac-curacy of its determination. Only in plant no. 4 wasthe root density conspicuously greater in the "wet"compartment than in the "dry" one, probably moreas a result of the original root division than as a resultof the treatment. The data given in Table 1 suggestthat the root mass and soil water potential were ho-mogeneous within compartments. Hence, we used thetotal root mass (length) and average soil water poten-tial for the terms RD and & in Eq. [2].
Table 2 shows the average soil water potential ineach of the two compartments and five experiments,the transpiration rates per unit leaf area, and the val-ues calculated for the latter, using Eq. [2]. Table 2shows clearly that the treatment, in each case, resultedin a distinct "wet" vs. "dry" compartment, that thecorresponding water uptake rates differed markedly inthe expected manner, and that the calculated absolutevalues are close to the measured ones.
The last result can be seen in more detail by com-paring the "dry"/"wet" ratio calculated with Eq. [2],and as measured (Table 3). The average quotient ofthe measured over the calculated ratio is 0.97. Theagreement between the two sets is a test of Eq. [2],irrespective of the value of SRPL. The total ("dry" +"wet") transpiration rate was calculated and com-pared to the corresponding measured value, by divid-ing the latter into the former. These results are shownin Table 4. The average ratio between measured andcalculated uptake is 1.04.
We conclude that the value of SRPL, measured onwhole, well-watered plants, appears to apply to plantsthat have split-root systems and that are partiallydroughted. This is an important and practical result,because it indicates that a plant parameter of a real

236 SOIL SCI. SOC. AM. J., VOL. 48, 1984
Table 4—Comparison of the total water uptake from fiveplants with a split root system, as measured and as
calculated from Eq. [2].
Plantno.
12345
AvgSD
Total water uptake
Measured
————— (m/s)10-12.213.545.398.319.99
Calculated
10.813.555.808.768.57
Meas/Calcratio
1.131.000.930.951.171.040.11
physical nature exists. Also, the form of Eq. [2] is es-sentially confirmed as a concept for dividing the totalplant water uptake over a root system that has accessto regions of contrasting water "availability." In fact,it could be postulated that all the water in the rootsystem is available for plant uptake, but to a differingdegree, as stated in mathematical and quantitativeform by Eq. [2]. We present this equation as an im-proved concept of availability. That is, Eq. [2] can beused as one of the essential elements in a predictionmodel that calculates the amount and the distributionof soil water depletion in the profile, given the at-mospheric demand, a stomatal control algorithm, andthe soil water and root distribution profile.
Such a prediction method may vary in complexityand detail. It may treat the soil as a system of just afew main horizons, or as a detailed, three-dimen-sionally partioned domain. It may simply assume arepresentative consumptive water use rate, rather thantry to estimate the latter from crop and weather de-tails. It can treat the root distribution as a fixed func-tion, or as a dynamic relation, affected by the verydrought conditions it may create. The occurrence ofroot exudation, i.e., transfer of water from wet soilthrough the roots to dry soil, can be included.
Finally, one may or may not choose to add the Dar-cian flow to the root uptake function, whereby soilwater from regions not permeated by roots may alsobecome available by "capillary rise," or wherebyavailable water may be lost by "deep seepage." Theseare two concepts that are nebulous until the flow equa-tion is used to determine their true extent and signif-icance for the water supply to plants from the soil inwhich they are rooted. The availability concept pre-sented here has been tested on single plants, and can-not be applied to a field grown crop without furtherevaluation. However, in the context of our experi-ment, Eq. [2] has been verified.
Our result is limited to the availability of soil waterfor uptake by plant roots, which is the classical inter-pretation. It may not apply to an extended concept ofsoil water availability that considers plant growth andthe production of harvestable materials of the desiredquality. However, a wealth of empirical evidence(Hanks and Hill, 1980) suggests that production goalswill not be attainable with an impaired water uptake,as dry matter production rates and water use rates areoften strongly and linearly correlated, for a given cropand location.

GERMANN ET AL.: PROFILES OF BROMIDE AND INCREASED SOIL MOISTURE AFTER INFILTRATION INTO SOILS 237
9:89-109.29. Van Bavel, C.H.M. 1974b. Exploratory simulation of the de-
pletion of water reserves in the plant root zone and its conse-quences, p. 279-282. Proc. 1974 Summer Computer SimulationConf., Houston, Tx. Simulation Councils, Inc., La Jolla, CA.
30. Van Bavel, C.H.M., and J. Ahmed. 1976. Dynamic simulationof water depletion in the root zone. Ecol. Modelling 2:189-212.
31. Van Bavel, C.H.M., G.B. Stirk, and K.J. Brust. 1968. Hydraulicproperties of a clay loam soil and the field measurement ofwater uptake by roots. II. The water balance of the root zone.
Soil Sci. Soc. Am. Proc. 32:317-321.32. Van Bavel, C.H.M., R. Lascano, and D.R. Wilson. 1978. Water
relations of fritted clay. Soil Sci. Soc. Am. J. 42:657-659.33. Veihmeyer, F.J., and A.H. Hendrickson. 1927. Soil-moisture
conditions in relation to plant growth. Plant Physiol. 2:71-78.34. Weatherly, P.E. 1976. Introduction: Water movement through
plants. Philos. Trans. R. Soc. London Ser. B: 273:435-444.35. Zur, B., J.W. Janes, K.J. Bopte, and L.C. Hammond. 1982.
Total resistance to water flow in field soybeans. II. Limiting soilmoisture. Agron. J. 74:99-105.





























