Embed Size (px)
Citation preview

ARTICLE
Backbone 1HN, 13C, and 15N resonance assignments of the tandemRNA-binding domains of human DGCR8
Braden M. Roth • Mirko Hennig
Received: 9 May 2012 / Accepted: 21 June 2012
� Springer Science+Business Media B.V. 2012
Abstract Double-stranded RNA binding domain
(dsRBD) containing proteins are critical components of the
microRNA (miRNA) pathway, with key roles in small
RNA biogenesis, modification, and regulation. DiGeorge
Critical Region 8 (DGCR8) is a 773 amino acid, dsRBD-
containing protein that was originally identified in humans
as a protein encoded in the region of chromosome 22 that is
deleted in patients with DiGeorge syndrome. Now, it is
realized that DGCR8 complements the nuclear RNase III
Drosha to initiate miRNA biogenesis by promoting effi-
cient recognition and cleavage of primary miRNAs (pri-
miRNA). A pair of C-terminal tandem dsRBDs separated
by a flexible linker are required for pri-miRNA substrate
binding and recognition. The crystal structure of the
DGCR8 core region comprising residues 493–720 revealed
that each dsRBD adopts the canonical abbba fold. How-
ever, several residues located in important flexible regions
including the b1-b2-loop implicated in canonical dsRNA
recognition are absent in the crystal structure and no RNA-
bound structure of DGCR8 has been reported. Here we
report the 1HN, 13C, and 15N backbone resonance assign-
ments of the 24 kDa, 214 amino acid human DGCR8core
(residues 493–706) by heteronuclear NMR spectroscopy.
Our assignments lay the foundation for a detailed solution
state characterization of the dynamical and RNA-binding
properties of this protein in solution.
Keywords DiGeorge Critical Region 8 (DGCR8) �Double stranded RNA binding domain (dsRBD) �Microprocessor � microRNA (miRNA)
Biological context
MicroRNAs are non-coding RNAs that regulate gene
expression through directed downregulation of targeted
messenger RNA (mRNA). MiRNA biogenesis involves two
distinct RNase III processing events. Long, noncoding pri-
miRNA transcripts are initially cleaved by the nuclear
RNase III Drosha to release a *70nt precursor (pre)-miR-
NA. Pre-miRNAs undergo a second round of processing by
the cytoplasmic RNase III Dicer before incorporating into
the RISC (RNA-Induced Silencing Complex) where the
mature miRNA serves as a guide to repress translation or
destabilize mRNAs with incomplete complementarity to the
miRNA guide. RNase III-dependent miRNA biogenesis is
facilitated by dsRBD containing protein cofactors. In
humans, the nuclear protein DGCR8 assists Drosha in the
recognition and cleavage of miRNA-containing pri-miRNA
hairpins and is required for embryonic development and
differentiation.
DGCR8 was initially identified as the essential gene
located within a 1.5-Mb chromosomal deletion (del22q11)
commonly associated with DGS. More recently, DGCR8 has
been shown to play a central role in pri-miRNA recognition
and processing (Han et al. 2004; Landthaler et al. 2004).
Drosha and DGCR8 copurify from cell extracts and together
constitute the ‘‘minimal’’ Microprocessor, responsible for
the initiation step of pri-miRNA biogenesis (Gregory et al.
2004). Through a largely unknown process, the Micropro-
cessor recognizes and binds the miRNA-containing hairpin
at its single-stranded(ss)-double stranded (ds) RNA junction
B. M. Roth � M. Hennig (&)
Department of Biochemistry and Molecular Biology, Medical
University of South Carolina, Charleston, 70 President St,
DD213, PO Box 250509, South Carolina 29425, USA
e-mail: [email protected]
123
Biomol NMR Assign
DOI 10.1007/s12104-012-9406-x

positioning Drosha’s catalytic RNase III domains to cleave
the pri-miRNA approximately one helical turn from the ss-
dsRNA junction (Han et al. 2006).
The *175 amino acid tandem dsRBD domain (511–685)
is the signature motif of RNase III-associated accessory
proteins. Crystal structures of dsRBD in complex with two
collinearly stacking, GC-rich dsRNA substrates helped to
define canonical interaction interfaces involving the a-heli-
cal and b1-b2 loop regions of a typical abbba dsRBD.
Besides its tandem C-terminal RBDs, DGCR8 features a
WW motif overlapping with the C352-mediated heme-
binding/dimerization domain (Faller et al. 2007), and an
N-terminal nuclear localization signal. Undoubtedly, the two
C-terminal dsRBDs of DGCR8 play a key role in pri-miRNA
substrate recognition, but answering the question whether
Drosha and DGCR8 function in mutually exclusive or
cooperative fashion awaits detailed structural information on
DGCR8-pri-miRNA complexes. Sohn et al. (2007) reported
the crystal structure of the DGCR8 ‘‘core’’, (residues
493–720) comprising both dsRBDs and the flexible linker
(residues 493–720). Their analysis revealed that the dsRBDs
are arranged in a pseudo two-fold symmetry stabilized by
hydrophobic interactions and hydrogen bonding between
both dsRBDs and a C-terminal loop/helix (residues
684–698) (Fig. 2, a7). Absent from the crystal structure,
however, are residues that correspond to flexible regions,
most notably the dsRBD1-dsRBD2 linker and a canonical
RNA-binding region located in the b5–b6 loop (Fig. 2).
Although this work has confirmed the canonical abbbaarchitecture of each dsRBD, how pri-miRNA are recognized
and distinguished from the myriad of ss-dsRNA junction in
other RNA transcripts by DGCR8 and whether the dsRBDs
coordinate the binding of a single RNA substrates remains
unknown. If a single, rod-like pri-miRNA substrate would be
recognized then the tandem dsRBD1-dsRBD2 would have to
undergo a significant conformational change upon binding
suggesting that NMR dynamics investigations could help to
decipher the pri-miRNA binding mechanism.
Here we report the 1HN, 13C, and 15N backbone reso-
nance assignments and secondary structure of the 24 kDa
human DGCR8core in solution. These will serve as a
foundation for further investigation of DGCR8 in terms of
protein dynamics and DGCR8/pri-miRNA complex
interactions.
Methods and experiments
Expression and purification of deuterated 2H,15N,(13C)-
DGCR8core
We have generated a 214 amino acid (24 kDa) truncated
version of the DGCR8core consisting of residues 493–706.
DGCR8core was amplified from pFLAG/HA-DGCR8
plasmid (Addgene #10921; Landthaler et al. 2004) with the
TEV protease recognition sequence ENLYFQS incorpo-
rated in the 50 primer: (50- GAAAACCTGTACTTT-
CAGtcagtgcaagatgcacccacaaagaaag-30 and 50- tcagaccatct
tgctgctctcacggccatacatgcgc-30; uppercase represents TEV
recognition sequence). The PCR product was used to
generate a pENTR-TEV/core Gateway entry vector by
TOPO-TA cloning with Invitrogen’s pCR8/GW/TOPO
vector. pENTR-TEV/core was recombined with a cus-
tomized Gatweway p(H)GB1-GTW destination vector that
incorporated the TEV/core sequence in frame with an
N-terminal His6-GB1 protein. Positive clones were trans-
formed into E. coli BL21-CodonPlus (DE3)-RIPL compe-
tent cells (Stratagene). Deuterated cultures of E. coli
BL21-CodonPlus (DE3)-RIPL cells expressing DGCR8core
were grown from a single colony in 2 ml LB/Amp100 at
37 �C overnight. These cultures were then adapted to
growth in 500 ml of [99.9 % D2O]-15N, (13C)-enriched M9
minimal media with a succession of small-volume cultures
grown at 37 �C. Protonated glucose carbon sources were
used and expression was induced with 1 mM IPTG for 4 h
at 37 �C, then purified with the HisTrapFF affinity column
(GE Healthcare). Partially purified fusion proteins were
cleaved with TEV protease overnight at room temperature
and reapplied to the HisTrap column. The flow-through
contained highly purified protein that was subsequently
exchanged into NMR buffer.
NMR spectroscopy
Even in the absence of the previously identified dimeriza-
tion domain of DGCR8 and at modest concentration,
DGCR8core exhibits a strong tendency to dimerize in
solution. In agreement with previously published work,
size-exclusion chromatography (SEC) analysis yielded an
estimated molecular weight (MW) of 31.0 kDa for the
DGCR8core, significantly larger than the theoretical MW of
24.2 kDa (data not shown). To address the resulting line
broadening, we have expressed deuterated 15N, 13C-
DGCR8core and utilized TROSY (transverse relaxation
optimized spectroscopy)-based pulse sequences to enhance
sensitivity and resolution. NMR spectra were acquired in
500 ll of 90 % H2O/10 % D2O at 25 �C on back-
exchanged 2H,15N- as well as 2H,15N,13C-labeled
DGCR8core samples (ca. 300 lM in 20 mM Na-phosphate
buffer, pH 7.0, 150 mM KCl, and 5 mM TCEP).
Three-dimensional, clean TROSY-based (Pervushin
et al. 1997; Schulte-Herbruggen and Sorensen 2000),
constant-time (CT)-HNCA, sequential CT-HNCA, CT-
HN(CA)CB, CT-HN(COCA)CB, HNCO and HN(CA)CO
were recorded on a TCI-cryoprobe-equipped Bruker
Avance800 spectrometer to establish sequential
B. M. Roth, M. Hennig
123
connectivities based on a pairwise comparison of intra- and
inter-residue 13Ca, 13Cb and 13C’ chemical shifts. Subse-
quently, assignments were confirmed using through-space1HN and 15N correlations derived from modified 3D
HSQC-NOESY-TROSY and NOESY-TROSY experi-
ments. The NMR data were processed using nmrPipe, and
analyzed with CcpNMR Analysis software.
Assignments and data deposition
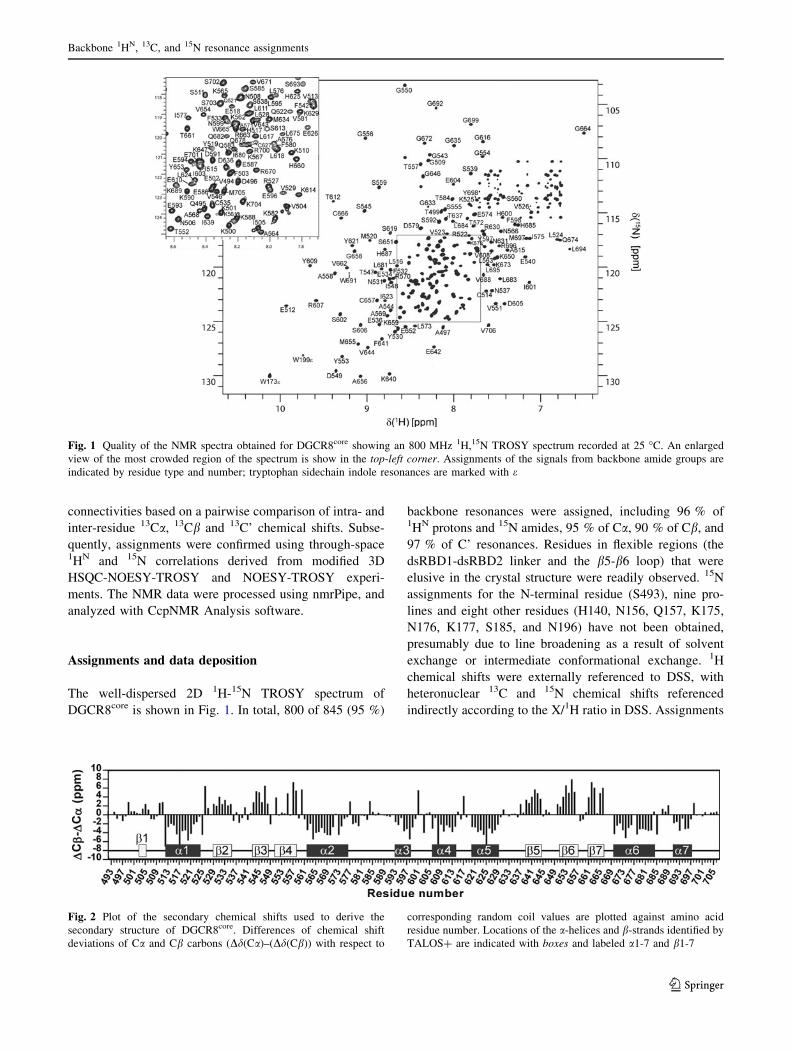
The well-dispersed 2D 1H-15N TROSY spectrum of
DGCR8core is shown in Fig. 1. In total, 800 of 845 (95 %)
backbone resonances were assigned, including 96 % of1HN protons and 15N amides, 95 % of Ca, 90 % of Cb, and
97 % of C’ resonances. Residues in flexible regions (the
dsRBD1-dsRBD2 linker and the b5-b6 loop) that were
elusive in the crystal structure were readily observed. 15N
assignments for the N-terminal residue (S493), nine pro-
lines and eight other residues (H140, N156, Q157, K175,
N176, K177, S185, and N196) have not been obtained,
presumably due to line broadening as a result of solvent
exchange or intermediate conformational exchange. 1H
chemical shifts were externally referenced to DSS, with
heteronuclear 13C and 15N chemical shifts referenced
indirectly according to the X/1H ratio in DSS. Assignments
Fig. 1 Quality of the NMR spectra obtained for DGCR8core showing an 800 MHz 1H,15N TROSY spectrum recorded at 25 �C. An enlarged
view of the most crowded region of the spectrum is show in the top-left corner. Assignments of the signals from backbone amide groups are
indicated by residue type and number; tryptophan sidechain indole resonances are marked with e
Fig. 2 Plot of the secondary chemical shifts used to derive the
secondary structure of DGCR8core. Differences of chemical shift
deviations of Ca and Cb carbons (Dd(Ca)–(Dd(Cb)) with respect to
corresponding random coil values are plotted against amino acid
residue number. Locations of the a-helices and b-strands identified by
TALOS? are indicated with boxes and labeled a1-7 and b1-7
Backbone 1HN, 13C, and 15N resonance assignments
123

have been deposited in the BioMagResBank database
(http://www.bmrb.wisc.edu) under accession number
17773.
Backbone Ca Cb and C’ assignments were used in
chemical shift index (Wishart and Sykes 1994) and TA-
LOS? (Shen et al. 2009) analyses as shown in Fig. 2; the
resulting secondary structure elements closely resemble
those of the crystal structure (Sohn et al. 2007) with minor
differences in secondary structure boundaries. The pre-
dicted secondary structure of human DGCR8core in solution
includes seven a-helices (a1: residues 512–524, a2: resi-
dues 562–576, a3: residues 593–598, a4: residues 606–614,
a5: residues 620–629, a6: residues 670–684, and a7: resi-
dues 691–697) and seven b-strands (b1: residues 503–505,
b2: residues 529–535, b3: residues 544–548, b4: residues
551–558, b5: residues 639–644, b6: residues 651–657, and
b7: residues 661–666).
Acknowledgments This work was supported by the National Sci-
ence Foundation [MCB 0845512 to M.H.]. We thank T. Tuschl for the
pFLAG/HA-DGCR8 construct and members of the Hennig laboratory
for stimulating discussions and comments on the manuscript and
acknowledge the support of the Hollings Marine Laboratory NMR
Facility for this work.
References
Faller M, Matsunaga M, Yin S, Loo JA, Guo F (2007) Heme is
involved in microRNA processing. Nat Struct Mol Biol 14(1):
23–29
Gregory RI, Yan KP, Amuthan G, Chendrimada T, Doratotaj B,
Cooch N, Shiekhattar R (2004) The Microprocessor complex
mediates the genesis of microRNAs. Nature 432(7014):235–240
Han J, Lee Y, Yeom KH, Kim YK, Jin H, Kim VN (2004) The
Drosha-DGCR8 complex in primary microRNA processing.
Genes Dev 18(24):3016–3027
Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK, Sohn SY, Cho Y,
Zhang BT, Kim VN (2006) Molecular basis for the recognition
of primary microRNAs by the Drosha-DGCR8 complex. Cell
125(5):887–901
Landthaler M, Yalcin A, Tuschl T (2004) The human DiGeorge
syndrome critical region gene 8 and Its D. melanogaster
homolog are required for miRNA biogenesis. Curr Biol 14(23):
2162–2167
Pervushin K, Riek R, Wider G, Wuthrich K (1997) Attenuated T2
relaxation by mutual cancellation of dipole–dipole coupling and
chemical shift anisotropy indicates an avenue to NMR structures
of very large biological macromolecules in solution. Proc Natl
Acad Sci USA 94(23):12366–12371
Schulte-Herbruggen T, Sorensen OW (2000) Clean TROSY: com-
pensation for relaxation-induced artifacts. J Magn Reson 144(1):
123–128
Shen Y, Delaglio F, Cornilescu G, Bax A (2009) TALOS ? : a hybrid
method for predicting protein backbone torsion angles from
NMR chemical shifts. J Biomol NMR 44(4):213–223
Sohn SY, Bae WJ, Kim JJ, Yeom KH, Kim VN, Cho Y (2007) Crystal
structure of human DGCR8 core. Nat Struct Mol Biol 14(9):
847–853
Wishart DS, Sykes BD (1994) The 13C chemical-shift index: a simple
method for the identification of protein secondary structure using
13C chemical-shift data. J Biomol NMR 4(2):171–180
B. M. Roth, M. Hennig
123



























![[C9] Hennig-Thurau Hansen Book 2000](https://img.pdfslide.net/doc/110x75/56d6c0711a28ab30169a6cef/c9-hennig-thurau-hansen-book-2000.jpg)

