Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 92, pp. 2701-2705, March 1995Plant Biology
Use of Saccharomyces cerevisiae for patch-clamp analysis ofheterologous membrane proteins: Characterization of Katl, aninward-rectifying K+ channel from Arabidopsis thaliana, andcomparison with endogeneous yeast channels and carriers
(gene expression/membrane permeability/whole-cell recording/channel current)
ADAM BERTL*, JULIE A. ANDERSONt, CLIFFORD L. SLAYMAN*, AND RICHARD F. GABERt*Department of Cellular and Molecular Physiology, Yale University, New Haven, CT 06510; and tDepartment of Biochemistry, Molecular Biology, andCell Biology, Northwestern University, Evanston, IL 60206
Communicated by Emanuel Epstein, University of California, Davis, CA, November 21, 1994
ABSTRACT Transport-deficient strains of the yeast Sac-charomyces cerevisiae have recently proven useful for cloning,by functional complementation, of cDNAs encoding heterol-ogous membrane transporters: specifically, H+-amino acidsymporters and K+ channels from the higher plant Arabidop-sis thaliana. The present study uses whole-cell patch-clampexperiments to show that yeast strains which grow poorly onsubmillimolar K+ due to the deletion of two K+-transportergenes (TRKI and TRK2) are in fact missing a prominent K+inward current present in wild-type cells. Rescue of suchstrains for growth on low K+ by transformation with a gene(KATI) encoding an inward-rectifying K+ channel from Ara-bidopsis is accompanied by the appearance of an inwardcurrent whose characteristics are in qualitative agreementwith previous studies in the Xenopus oocyte system, but differin quantitative details. The ability to make such measure-ments directly on Saccharomyces should facilitate structure-function studies of any electrogenic or electrophoretic iontransporters which can be expressed in the plasma membrane(or tonoplast) of that organism.
Over the past 10 years applied cellular genetics and molecularbiology have revolutionized the study of biological mem-branes. The concepts of channel, carrier, and receptor-originally deduced from physical, chemical, and physiologicalmeasurements-have metamorphosed into pictures of familiesof protein molecules with defined primary structure. And acentral topic of membrane research has become the functionalanalysis of mutant proteins whose structures are altered bysystematic amino acid changes. Such structure-function stud-ies lean heavily on techniques for expressing the proteins ofinterest in foreign membranes, where altered function can beexamined more easily than in the native membranes (1-3).For proteins which convey ionic currents through mem-
branes-i.e., ion channels and most ion pumps or ion-substrate symporters-a prime tool of functional analysis hasbecome the patch clamp, and the most widely adopted expres-sion system has been mature oocytes of the African clawedfrog Xenopus laevis, whose large size and easy handling havegenerally outweighed their prime disadvantage, that onlytransient expression is obtained.On the other hand, an attractive stable expression system
which is excellent for primary genetic manipulation is the yeastSaccharomyces cerevisiae. Until recently, however, the smallsize of Saccharomyces cells and the peculiar mechanical prop-erties of their membranes have made their electrophysiologicalstudy trying, despite the early demonstration (4) of the feasi-bility of patch-clamping yeast. Routine methods for recording
from isolated patches of yeast membrane had also beendescribed (5-7), but repeated attempts to identify heterolo-gously expressed transporters in such patches had failed (A.B.,unpublished work). This failure led us to extend the patch-clamp technique with Saccharomyces to a stable whole-cellrecording geometry, whose usefulness when applied in com-bination with molecular genetics is demonstrated below, bycharacterization of a heterologous K+ channel, Katl fromArabidopsis.
METHODS
General methods of growing, protoplasting, and handlingyeast for patch-clamp experiments were as described (5-7).The four strains of S. cerevisiae used are listed in Table 1. Mostexperiments were carried out on a haploid strain (Katl,CY162-pKAT1) transformed with the cDNA for KAT1 fromArabidopsis thaliana, and results were directly controlled bymeasurements on a strain (VEC, CY162-pR316) transformedwith the plasmid vector alone. A diploid K+ double-deletionstrain (TrklATrk2A, CY152/CY162) was used as a null con-trol, and a tetraploid wild-type strain [WT, Y588; adopted forisolated-patch studies (6, 7)] was used for background exper-iments. WT and TrklATrk2A were grown on 2% glucose, butKatl and VEC were grown on 1% galactose, since the trans-forming plasmid carried a galactose promoter.
Patch recording was done according to Hamill et al. (11) asmodified for S. cerevisiae by Bertl et al. (5, 6). Explicit methodsfor single-channel recording are detailed in ref. 7, and thewhole-cell configuration was obtained by combining pulses ofsuction and voltage (-600 mV), after tight seal formation.Enlarged cells (diploid and tetraploid) proved easier to handlethan normal yeast (haploid), but the difference became lesscrucial as skill developed. On a "good day," tight seals on yeastplasma membrane were obtained in "80% of trials, and cleantransfer to the isolated-patch or whole-cell configurationsoccurred with "80% of tight seals.
Seal resistances varied between 15 and 20 GQl, and allwhole-cell recordings were corrected by subtracting a linearleak of 66.7 pS (15 Ga). Currents were recorded in responseto 2.5-s voltage steps progressing from + 100 mV to -200 mVin 20 mV decrements, with a 300-ms holding interval at -40mV between each pair. Data were filtered at 1 kHz andsampled at 200 Hz.
Standard compositions of solutions for whole-cell experi-ments were as follows. Cytosolic (pipette) solution was 175mMKCI/5 mM MgCl2/4 mM ATP/100 nM Ca2+/1 mM EGTA,brought to pH 7.0 with KOH. Extracellular (bath) solution was150 mM KCl/5 mM MgCl2/10 mM CaCl2, buffered to pH 7.5
Abbreviations: TEA+, tetraethylammonium; CrX, charybdotoxin;DTX, dendrotoxin.
2701
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad Sci. USA 92 (1995)
Table 1. Strains of S. cerevisiae used
Strain Mating type Markers Name Ref.Y588 a, a, a, a ura3-52, ade2 WT 8CY152/CY162 a, a ura3-52, his4-15, trklAtrk2A TrklATrk2A 9CY162-pRS316 a ura3-52, his4-15, trklArk2A + pRS316 VEC 10CY162-pKAT1 a ura3-52, his4-15, trklAtrk2A + pKAT1 Katl 10
with 0.1 mM Tris/Mes. According to the standard electro-physiological sign convention, cation flow from the cytoplas-mic side of the plasma membrane to the extracellular side hasbeen designated outward current and drawn positive (upward).
RESULTSStandard Properties ofthe Yeast Plasma Membrane. Under
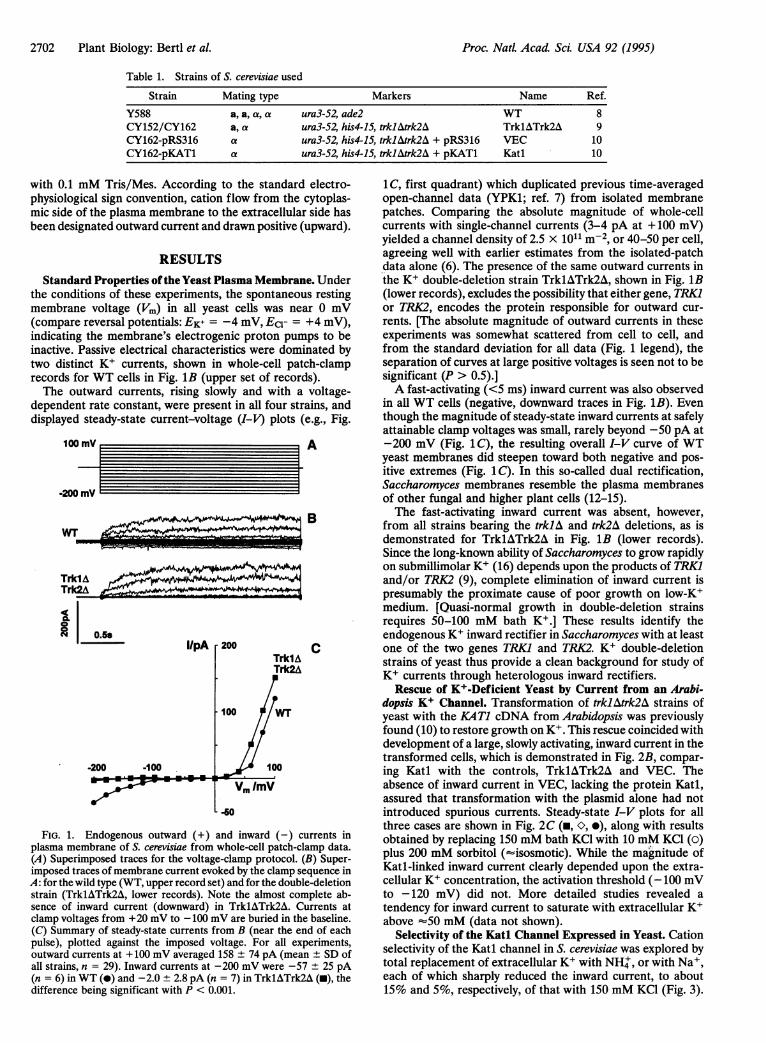
the conditions of these experiments, the spontaneous restingmembrane voltage (Vm) in all yeast cells was near 0 mV(compare reversal potentials: EK+ = -4 mV, EC- = +4 mV),indicating the membrane's electrogenic proton pumps to beinactive. Passive electrical characteristics were dominated bytwo distinct K+ currents, shown in whole-cell patch-clamprecords for WT cells in Fig. 1B (upper set of records).The outward currents, rising slowly and with a voltage-
dependent rate constant, were present in all four strains, anddisplayed steady-state current-voltage (I-V) plots (e.g., Fig.
J mv, A
-200 mV ~~~~~~BWT
TrklATrk2A
CLN o.ss
I/pA 200 CTrklATrk2A
100 WT
-200 -100 100
_ Vm ImV
-50
FIG. 1. Endogenous outward (+) and inward (-) currents inplasma membrane of S. cerevisiae from whole-cell patch-clamp data.(A) Superimposed traces for the voltage-clamp protocol. (B) Super-imposed traces of membrane current evoked by the clamp sequence inA: for the wild type (WT, upper record set) and for the double-deletionstrain (TrklATrk2A, lower records). Note the almost complete ab-sence of inward current (downward) in TrklATrk2A. Currents atclamp voltages from +20 mV to -100 mV are buried in the baseline.(C) Summary of steady-state currents from B (near the end of eachpulse), plotted against the imposed voltage. For all experiments,outward currents at + 100 mV averaged 158 ± 74 pA (mean ± SD ofall strains, n = 29). Inward currents at -200 mV were -57 ± 25 pA(n = 6) in WT (-) and -2.0 ± 2.8 pA (n = 7) in TrklATrk2A (U), thedifference being significant with P < 0.001.
1 C, first quadrant) which duplicated previous time-averagedopen-channel data (YPK1; ref. 7) from isolated membranepatches. Comparing the absolute magnitude of whole-cellcurrents with single-channel currents (3-4 pA at +100 mV)yielded a channel density of 2.5 x 101l m-2, or 40-50 per cell,agreeing well with earlier estimates from the isolated-patchdata alone (6). The presence of the same outward currents inthe K+ double-deletion strain TrklATrk2A, shown in Fig. 1B(lower records), excludes the possibility that either gene, TRKIor TRK2, encodes the protein responsible for outward cur-rents. [The absolute magnitude of outward currents in theseexperiments was somewhat scattered from cell to cell, andfrom the standard deviation for all data (Fig. 1 legend), theseparation of curves at large positive voltages is seen not to besignificant (P > 0.5).]A fast-activating (<5 ms) inward current was also observed
in all WT cells (negative, downward traces in Fig. 1B). Eventhough the magnitude of steady-state inward currents at safelyattainable clamp voltages was small, rarely beyond -50 pA at-200 mV (Fig. 1 C), the resulting overall I-V curve of WTyeast membranes did steepen toward both negative and pos-itive extremes (Fig. 1 C). In this so-called dual rectification,Saccharomyces membranes resemble the plasma membranesof other fungal and higher plant cells (12-15).The fast-activating inward current was absent, however,
from all strains bearing the trklA and trk2A deletions, as isdemonstrated for TrklATrk2A in Fig. 1B (lower records).Since the long-known ability of Saccharomyces to grow rapidlyon submillimolar K+ (16) depends upon the products of TRKIand/or TRK2 (9), complete elimination of inward current ispresumably the proximate cause of poor growth on low-K+medium. [Quasi-normal growth in double-deletion strainsrequires 50-100 mM bath K+.] These results identify theendogenous K+ inward rectifier in Saccharomyces with at leastone of the two genes TRK1 and TRK2. K+ double-deletionstrains of yeast thus provide a clean background for study ofK+ currents through heterologous inward rectifiers.Rescue of K+-Deficient Yeast by Current from an Arabi-
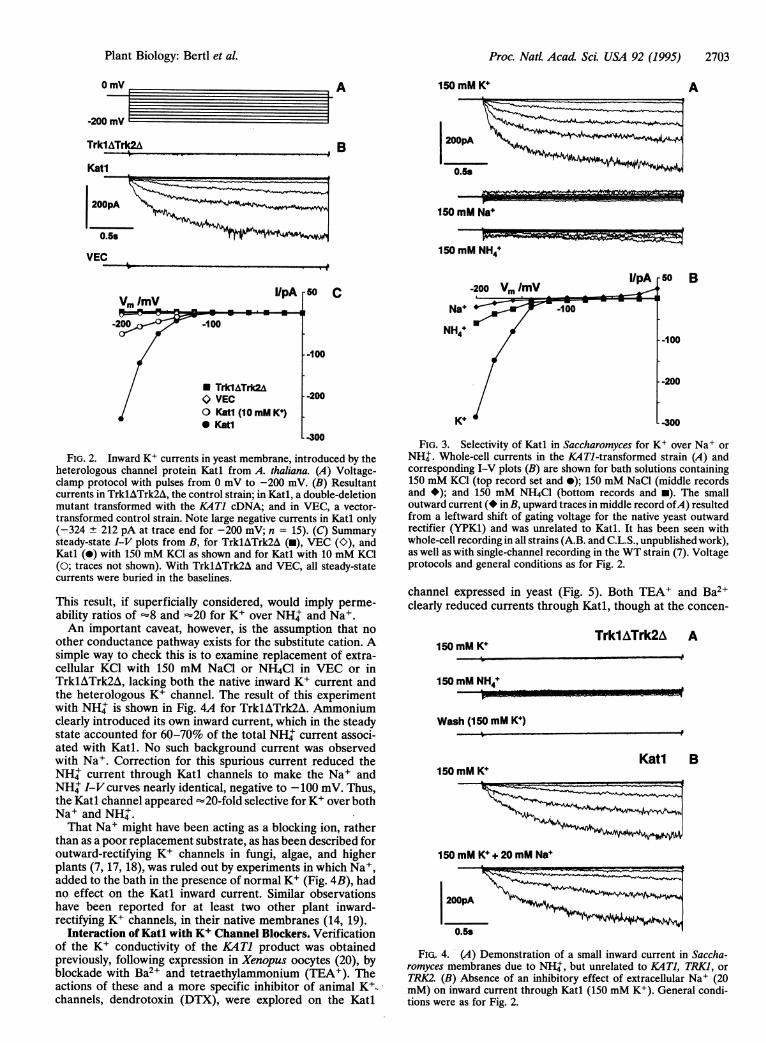
dopsis K+ Channel. Transformation of trklAtrk2A strains ofyeast with the KATl cDNA from Arabidopsis was previouslyfound (10) to restore growth on K+. This rescue coincided withdevelopment of a large, slowly activating, inward current in thetransformed cells, which is demonstrated in Fig. 2B, compar-ing Katl with the controls, TrklATrk2A and VEC. Theabsence of inward current in VEC, lacking the protein Katl,assured that transformation with the plasmid alone had notintroduced spurious currents. Steady-state I-V plots for allthree cases are shown in Fig. 2C (-, O, 0), along with resultsobtained by replacing 150 mM bath KCl with 10 mM KCI (0)plus 200 mM sorbitol (-isosmotic). While the ma"gnitude ofKatl-linked inward current clearly depended upon the extra-cellular K+ concentration, the activation threshold (-100 mVto -120 mV) did not. More detailed studies revealed atendency for inward current to saturate with extracellular K+above =50 mM (data not shown).
Selectivity of the Katl Channel Expressed in Yeast. Cationselectivity of the Katl channel in S. cerevisiae was explored bytotal replacement of extracellular K+ with NH+, or with Na+,each of which sharply reduced the inward current, to about15% and 5%, respectively, of that with 150 mM KCl (Fig. 3).
2702 Plant Biology: Bertl et al.
nn -1fI Uu
Proc. Natl. Acad Sci. USA 92 (1995) 2703
0 mV A 150 mM K A
tmV ------- 5
B
150mMNa===4150 mM NaH
F=M.....I O ~z
150 mM NH +
Vm ImV
-200
I/pA
-100
TrklATrk2A<> VECO Katl (10 mM K)* Katl
r 50 C
Nai -10
NH4+
K+
--100
- -200
- -300
FIG. 2. Inward K+ currents in yeast membrane, introduced by theheterologous channel protein Katl from A. thaliana. (A) Voltage-clamp protocol with pulses from 0 mV to -200 mV. (B) Resultantcurrents in TrklATrk2A, the control strain; in Katl, a double-deletionmutant transformed with the KAT1 cDNA; and in VEC, a vector-transformed control strain. Note large negative currents in Katl only(-324 ± 212 pA at trace end for -200 mV; n = 15). (C) Summarysteady-state I-V plots from B, for TrklATrk2A (i), VEC (K), andKatl (0) with 150 mM KCl as shown and for Katl with 10 mM KCl(0; traces not shown). With TrklATrk2A and VEC, all steady-statecurrents were buried in the baselines.
This result, if superficially considered, would imply perme-ability ratios of -8 and =20 for K+ over NH+ and Na+.An important caveat, however, is the assumption that no
other conductance pathway exists for the substitute cation. Asimple way to check this is to examine replacement of extra-cellular KC1 with 150 mM NaCl or NH4Cl in VEC or inTrklATrk2A, lacking both the native inward K+ current andthe heterologous K+ channel. The result of this experimentwith NH' is shown in Fig. 4A for TrklATrk2A. Ammoniumclearly introduced its own inward current, which in the steadystate accounted for 60-70% of the total NH' current associ-ated with Katl. No such background current was observedwith Na+. Correction for this spurious current reduced theNH' current through Katl channels to make the Na+ andNH+ I-V curves nearly identical, negative to -100 mV. Thus,the Katl channel appeared =20-fold selective for K+ over bothNa+ and NH'.That Na+ might have been acting as a blocking ion, rather
than as a poor replacement substrate, as has been described foroutward-rectifying K+ channels in fungi, algae, and higherplants (7, 17, 18), was ruled out by experiments in which Na+,added to the bath in the presence of normal K+ (Fig. 4B), hadno effect on the Katl inward current. Similar observationshave been reported for at least two other plant inward-rectifying K+ channels, in their native membranes (14, 19).
Interaction of Katl with K+ Channel Blockers. Verificationof the K+ conductivity of the KAT1 product was obtainedpreviously, following expression in Xenopus oocytes (20), byblockade with Ba2+ and tetraethylammonium (TEA+). Theactions of these and a more specific inhibitor of animal K+t.channels, dendrotoxin (DTX), were explored on the Katl
B
-100
-200
-300
FIG. 3. Selectivity of Katl in Saccharomyces for K+ over Na+ orNH'. Whole-cell currents in the KATJ-transformed strain (A) andcorresponding I-V plots (B) are shown for bath solutions containing150 mM KCl (top record set and *); 150 mM NaCl (middle recordsand *); and 150 mM NH4Cl (bottom records and *). The smalloutward current (*inB, upward traces in middle record ofA) resultedfrom a leftward shift of gating voltage for the native yeast outwardrectifier (YPK1) and was unrelated to Katl. It has been seen withwhole-cell recording in all strains (A.B. and C.L.S., unpublished work),as well as with single-channel recording in the WT strain (7). Voltageprotocols and general conditions as for Fig. 2.
channel expressed in yeast (Fig. 5). Both TEA+ and Ba2+clearly reduced currents through Katl, though at the concen-
150 mM K4
150 mM NH44
Wash (150 mM K+)
Trkl ATrk2A A
Katl B150 mM K+
150 mM K4+ 20 mM Na4
FIG. 4. (A) Demonstration of a small inward current in Saccha-romyces membranes due to NH+, but unrelated to KATI, TRK1, orTRK2. (B) Absence of an inhibitory effect of extracellular Na+ (20mM) on inward current through Katl (150 mM K+). General condi-tions were as for Fig. 2.
-20C
TrklATrk2A
Kati
200pA
0.5s
VEC
Plant Biology: Bertl et aL
I/pA 50':~-200 V,, /mV
Proc. Natl. Acad ScL USA 92 (1995)
Control A
+ 10 mM TEA+
Control B
V10mMBa++
+ 10 mM Ba++!Ak~
7~~~~~~~~~
Control
I
+5OnM DTX AAl
200pA
0.5s
C
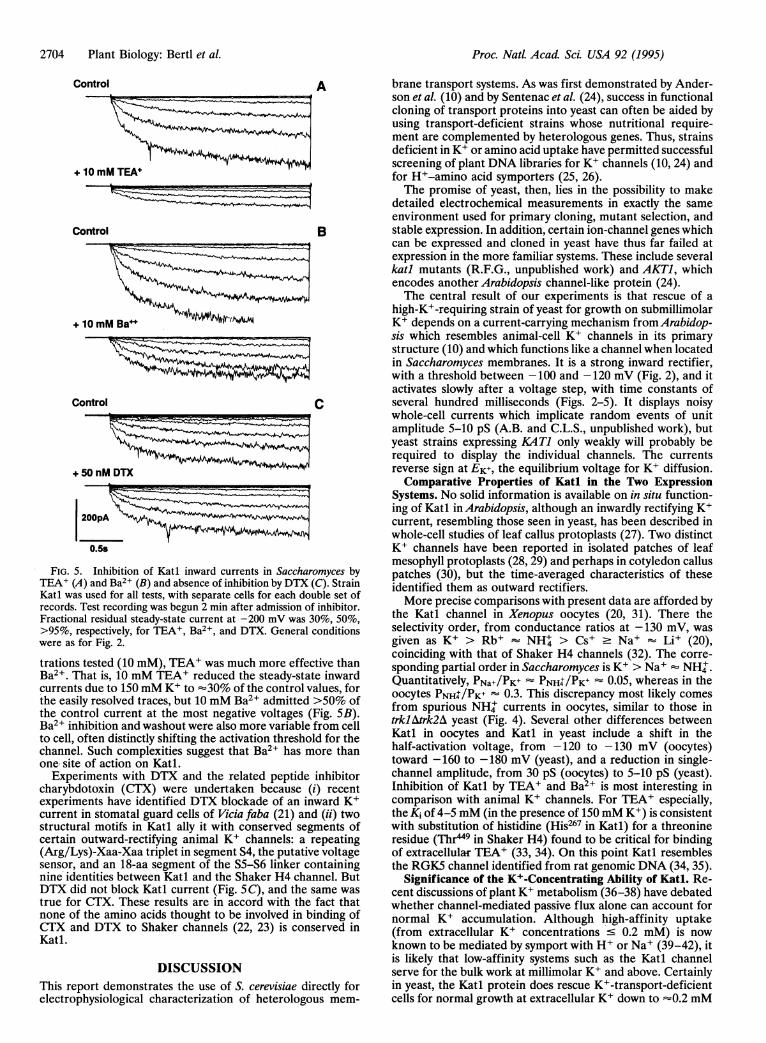
FIG. 5. Inhibition of Katl inward currents in Saccharomyces by
TEA+ (A) and Ba2+ (B) and absence of inhibition by DTX (C). Strain
Katl was used for all tests, with separate cells for each double
records. Test recording was begun 2 min after admission of inhibitor.
Fractional residual steady-state current at -200 mV was 30%,
>95%, respectively, for TEA+, Ba2 and DTX. General conditions
were as for Fig. 2.
trations tested (10 mM), TEA+ was much more effective than
Ba2+. That is, 10 mM TEA+ reduced the steady-state inward
currents due to 150 mM K+ to '30% of the control values, for
the easily resolved traces, but 10 mM Ba2+ admitted >50% of
the control current at the most negative voltages (Fig.SB).
Ba2 inhibition and washout were also more variable from cell
to cell, often distinctly shifting the activation threshold for the
channel. Such complexities suggest that Ba2 has more than
one site of action on Katl.Experiments with DTX and the related peptide inhibitor
charybdotoxin (CTX) were undertaken because (i) recent
experiments have identified DTX blockade of an inwardK+
current in stomatal guard cells of Vicia faba (21) and(ii) two
structural motifs in Katl ally it with conserved segments of
certain outward-rectifying animalK+ channels: a repeating
(Arg/Lys)-Xaa-Xaa triplet in segment S4, the putative voltage
sensor, and an 18-aa segment of the SS-S6 linker containing
nine identities between Kati and the Shaker H4 channel. But
DTX did not block Katl current (Fig. SC), and the same was
true for CTX. These results are in accord with the fact that
none of the amino acids thought to be involved in binding of
CTX and DTX to Shaker channels (22, 23) is conserved in
Katl.
DISCUSSION
This report demonstrates the use of S. cerevisiae directly for
electrophysiological characterization of heterologous mem-
brane transport systems. As was first demonstrated by Ander-son et al. (10) and by Sentenac etal. (24), success in functionalcloning of transport proteins into yeast can often be aided byusing transport-deficient strains whose nutritional require-ment are complemented by heterologous genes. Thus, strainsdeficient in K+ or amino acid uptake have permitted successfulscreening of plant DNA libraries for K+ channels (10, 24) andfor HI-amino acid symporters (25, 26).The promise of yeast, then, lies in the possibility to make
detailed electrochemical measurements in exactly the sameenvironment used for primary cloning, mutant selection, andstable expression. In addition, certain ion-channel genes whichcan be expressed and cloned in yeast have thus far failed atexpression in the more familiar systems. These include severalkatl mutants (R.F.G., unpublished work) and AKT1, whichencodes another Arabidopsis channel-like protein (24).The central result of our experiments is that rescue of ahigh-K+-requiring strain of yeast for growth on submillimolar
K+ depends on a current-carrying mechanism from Arabidop-sis which resembles animal-cell K+ channels in its primarystructure (10) and which functions like a channel when locatedin Saccharomyces membranes. It is a strong inward rectifier,with a threshold between -100 and -120 mV (Fig. 2), and itactivates slowly after a voltage step, with time constants ofseveral hundred milliseconds (Figs. 2-5). It displays noisywhole-cell currents which implicate random events of unitamplitude 5-10 pS (A.B. and C.L.S., unpublished work), butyeast strains expressing KATl only weakly will probably berequired to display the individual channels. The currentsreverse sign at EK+, the equilibrium voltage for K+ diffusion.Comparative Properties of Katl in the Two Expression
Systems. No solid information is available on in situ function-ing of Katl inArabidopsis, although an inwardly rectifyingK+current, resembling those seen in yeast, has been described inwhole-cell studies of leaf callus protoplasts (27). Two distinctK+ channels have been reported in isolated patches of leafmesophyll protoplasts (28,29) and perhaps in cotyledon calluspatches (30), but the time-averaged characteristics of theseidentified them as outward rectifiers.More precise comparisons with present data are afforded by
the Katl channel in Xenopus oocytes (20, 31). There theselectivity order, from conductance ratios at -130 mV, wasgiven as K+ > Rb+ NH' > Cs+ Na+ Li+ (20),coinciding with that of Shaker H4 channels (32). The corre-sponding partial order in Saccharomyces is K+> Na+ NH'.Quantitatively, PNa+/PK+ PNH4/PK+ 0.05, whereas in theoocytes PNH4 /PK+ 0.3. This discrepancy most likely comesfrom spurious NH+ currents in oocytes, similar to those intrklAtrk2A yeast (Fig. 4). Several other differences betweenKatl in oocytes and Katl in yeast include a shift in thehalf-activation voltage, from -120 to -130 mV (oocytes)toward -160 to -180 mV (yeast), and a reduction in single-channel amplitude, from 30 pS (oocytes) to 5-10 pS (yeast).Inhibition of Katl by TEA+ and Ba2+ ismost interesting incomparison with animal K+ channels. For TEA+ especially,theKi of 4-5 mM (in the presence of 150 mM K+) is consistentwith substitution of histidine (His267 in Katl) for a threonineresidue (Thr449 in Shaker H4) found to be critical for bindingof extracellular TEA+ (33, 34). On this point Katl resemblesthe RGK5 channel identified from rat genomic DNA (34, 35).
Significance of the K+-Concentrating Ability of Katl. Re-cent discussions of plantK+ metabolism (36-38) have debatedwhether channel-mediated passive flux alone can account fornormal K+ accumulation. Although high-affinity uptake(from extracellular K+ concentrations 0.2 mM) is nowknown to be mediated by symport with HI or Na+ (39-42), itis likely that low-affinity systems such as the Katl channelserve for the bulk work at millimolarK+ and above. Certainlyin yeast, the Katl protein does rescue K+-transport-deficientcells for normal growth at extracellularK+ down to -0.2 mM
i
2704 Plant Biology: Bertl et al.

Proc. Natl. Acad. Sci. USA 92 (1995) 2705
(10). Normal growth rates in Saccharomyces are maintained atintracellular K+ concentrations down to 130 mM (43),implying that the selective conditions for cloning KA Ti in yeastproduced intracellular/extracellular K+ ratios of 130/0.2 =650. For passive accumulation that ratio would require a Vm ofonly about -155 mV (activity corrected), well within thenormal range for Neurospora (44) and for the giant yeastEndomyces magnusii (45), and found also in pilot experimentson swollen S. cerevisiae (A.B., unpublished data).
Relation to the Endogenous Yeast K+-uptake Systems. Theoutstanding properties of the WT yeast membrane's owninward current are its small size and relatively low noise, its fastactivation following voltage steps (Fig. 1B), and its identifi-cation with proteins-Trkl and/or Trk2-whose structures(9) differ widely from those of any other known channels.These properties raise the question of whether Trkl or Trk2may be the Saccharomyces equivalent of the K+-H+ symporteralready described in Neurospora (12, 39). In this context, it isinteresting that a trklAtrk2A strain of Saccharomyces has nowmade possible cloning of a presumptive K+-H+ symportergene from wheat (HKTI; ref. 42).
We thank Drs. E. Modzydlowski and S. Goldstein for samples ofCTX and DTX, for perusal of prepublication data, and for muchhelpful advice. We thank Drs. D. Gradmann and G. Thiel for criticalreading of the manuscript. This work was supported by GrantsBE-1181/2 and BE-1181/3 from the Deutsche Forschungsgemein-schaft (to A.B.), Grant 85ER13359 from the U.S. Department ofEnergy (to C.L.S.), and Grant DCB8711346 from the National ScienceFoundation (to R.F.G.).
1. Paulmichl, M., Li, Y., Wickman, K., Ackerman, M., Peralta, E.& Clapham, D. (1992) Nature (London) 356, 238-241.
2. Ho, K., Nichols, C. G., Lederer, W. J., Lytton, J., Vassilev, P. M.,Kanazirska, M. V. & Hebert, S. C. (1993) Nature (London) 362,31-38.
3. Kubo, Y., Baldwin, T. J., Jan, Y. N. & Jan, L. Y. (1993) Nature(London) 362, 127-133.
4. Gustin, M. C., Martinac, B., Saimi, Y., Culbertson, M. R. &Kung, C. (1986) Science 233, 1195-1197.
5. Bertl, A. & Slayman, C. L. (1990) Proc. Natl. Acad. Sci. USA 87,7824-7828.
6. Bertl, A., Slayman, C. L. & Gradmann, D. (1992) J. Membr. Biol.132, 183-199.
7. Bertl, A. & Slayman, C. L. (1992) J. Exp. Biol. 172, 271-287.8. Mirzayan, C., Copeland, C. S. & Snyder, M. (1992) J. Cell Biol.
116, 1319-1332.9. Ko, C. H. & Gaber, R. F. (1991) Mol. Cell. Biol. 11, 4266-4273.
10. Anderson, J. A., Huprikar, S. S., Kochian, L. V., Lucas, W. J. &Gaber, R. F. (1992) Proc. Natl. Acad. Sci. USA 89, 3736-3740.
11. Hamill, 0. P., Marty, A., Neher, E., Sakmann, B. & Sigworth,F. J. (1981) Pflugers Arch. 391, 85-100.
12. Blatt, M. R., Rodriguez-Navarro, A. & Slayman, C. L. (1987) J.Membr. Biol. 98, 169-189.
13. Hedrich, R. & Schroeder, J. I. (1989) Annu. Rev. Plant Physiol.Mol. Biol. 40, 539-569.
14. Blatt, M. R. (1991) J. Membr. Biol. 124, 95-112.15. Schroeder, J. I., Ward, J. M. & Gassmann, W. (1994) Annu. Rev.
Biophys. Biomol. Struct. 23, 441-471.16. Rothstein, A. (1964) in The Cellular Functions of Membrane
Transport, ed. Hoffman, J. F. (Prentice-Hall, Englewood Cliffs,NJ), pp. 23-39.
17. Bertl, A. (1989) J. Membr. Biol. 109, 9-19.18. Blatt, M. R. (1992) J. Gen. Physiol. 99, 615-644.19. Thiel, G. & Blatt, M. R. (1991) J. Plant Physiol. 138, 326-334.20. Schachtman, D. P., Schroeder, J. I., Lucas, W. J., Anderson, J. A.
& Gaber, R. F. (1992) Science 258, 1654-1658.21. Obermeyer, G., Armstrong, F. & Blatt, M. R. (1994) J. Membr.
Biol. 137, 249-259.22. MacKinnon, R., Heginbotham, L. & Abramson, T. (1990) Neu-
ron 5, 767-771.23. Goldstein, S. A. N. & Miller, C. (1993) Biophys J. 65, 1613-1619.24. Sentenac, H., Bonneaud, N., Minet, M., Lacroute, F., Salmon,
J.-M., Gaymard, F. & Grignon, C. (1992) Science 256, 663-665.25. Frommer, W. B., Hummel, S. & Riesmeier, J. W. (1993) Proc.
Natl. Acad. Sci. USA 90, 5944-5948.26. Hsu, L.-C., Chiou, T.-J., Chen, L. & Bush, D. R. (1993) Proc.
Natl. Acad. Sci. USA 90, 7441-7445.27. Colombo, R. & Cerana, R. (1991) Plant Physiol. 97, 1130-1135.28. Spalding, E. P., Slayman, C. L., Goldsmith, M. H. M., Grad-
mann, D. & Bertl, A. (1992) Plant Physiol. 99, 96-102.29. Spalding, E. P. & Goldsmith, M. H. M. (1993) Plant Cell 5,
477-484.30. Lew, R. R. (1991) Plant Physiol. 95, 642-647.31. Hoshi, T. (1994) Biophys. J. 66, A283 (abstr. W-Posl78).32. Heginbotham, L. & MacKinnon, R. (1993) Biophys. J. 65, 2089-
2096.33. Kavanaugh, M. P., Varnum, M. D., Osborne, P. B., Christie,
M. J., Busch, A. F., Adelman, J. P. & North, R. A. (1991)J. Biol.Chem. 266, 7583-7587.
34. Yellen, G., Jurman, M. E., Abramson, T. & MacKinnon, R.(1991) Science 251, 939-942.
35. Douglass, J., Osborne, P. B., Cai, Y.-C., Wilkinson, M., Christie,M. J. & Adelman, J. P. (1990) J. Immunol. 144, 4841-4850.
36. Maathuis, F. J. M. & Sanders, D. (1993) Planta 191, 302-307.37. Kochian, L. V. & Lucas, W. J. (1993) Plant Cell 5, 720-721.38. Gassmann, W., Ward, J. M. & Schroeder, J. I. (1993) Plant Cell
5, 1491-1493.39. Rodriguez-Navarro, A., Blatt, M. R. & Slayman, C. L. (1986) J.
Gen. Physiol. 87, 649-674.40. Smith, F. A. & Walker, N. A. (1989)J. Membr. Biol. 108,125-137.41. Maathuis, F. J. M. & Sanders, D. (1994) Proc. Natl. Acad. Sci.
USA 91, 9272-9276.42. Schachtman, D. P. & Schroeder, J. I. (1994) Nature (London)
370, 655-658.43. Jones, W. B. G., Rothstein, A., Sherman, F. & Stannard, J. N.
(1965) Biochim. Biophys. Acta 104, 310-312.44. Slayman, C. L. (1965) J. Gen. Physiol. 49, 93-116.45. Bakker, R., Dobbelmann, J. & Borst-Pauwels, G. W. F. H. (1986)
Biochim. Biophys. Acta 861, 205-209.
Plant Biology: Bertl et al.





























