Embed Size (px)
Citation preview

SCIENCE FOR CONSERVATION 325
A classification of New Zealand’s terrestrial ecosystemsNicholas J.D. Singers and Geoffrey M. Rogers

Cover: A contiguous sequence of terrestrial indigenous ecosystems at the mouth of the lower Hollyford River/Whakatipu Kā Tuka at Martins Bay, north Fiordland. A combination of shifts in the course of the river superimposed on a prograding coastline have produced an array of fossil dune ridges and intervening wet depressions. A complex array of beach ( just out of frame lower right), dune, wetland, forest, and low- and high-alpine ecosystems stretch from the coast to the mountaintops. Lake McKerrow/Whakatipu Waitai is at centre-left, the Skippers Range is at upper left and the central May Hills subtend the distant Darran Mountains. Photo: Geoff Rogers
Science for Conservation is a scientific monograph series presenting research funded by New Zealand Department of Conservation (DOC). Manuscripts are internally and externally peer-reviewed; resulting publications are considered part of the formal international scientific literature.
This report is available from the departmental website in pdf form. Titles are listed in our catalogue on the website, refer www.doc.govt.nz under Publications, then Science & technical.
© Copyright May 2014, New Zealand Department of Conservation
ISSN 1177–9241 (web PDF)ISBN 978–0–478–15013–1 (web PDF)
This report was prepared for publication by the Publishing Team; editing and layout by Amanda Todd. Publication was approved by the Deputy Director-General, Science and Capability Group, Department of Conservation, Wellington, New Zealand.
Published by Publishing Team, Department of Conservation, PO Box 10420, The Terrace, Wellington 6143, New Zealand.
In the interest of forest conservation, we support paperless electronic publishing.

CONTENTS
Abstract 1
1. Introduction 2
1.1 Defining the terrestrial environment 5
2. Methods 5
2.1 Structure of the classification 5
2.2 Classification and mapping of zonal environments and ecosystems 8
2.3 Classification and mapping of azonal environments and ecosystems 9
2.4 Scale and boundaries of ecosystems 10
3. Results 11
3.1 Zonal ecosystems 113.1.1 Subtropical forest (Unit SF1) 113.1.2 Warm temperate forest (Units WF1–14) 113.1.3 North and South Island mild forests (Units MF1–25) 143.1.4 Cool forest and scrub (Units CLF1–12) 143.1.5 Cold forest and scrub (Units CDF1–7) 153.1.6 Low alpine and subantarctic (Units AL1–9) 153.1.7 High alpine (Units AH1–4) 163.1.8 Cold temperature inversion (Units TI1–6) 16
3.2 Azonal ecosystems 17 3.2.1 High water tables—permanently or seasonally wet soils (Units WL1–22) 173.2.2 Frequent geomorphic disturbance (Units DN1–5; EP1; CL1–11; SC1; BR1–3) 183.2.3 Extreme soil and/or atmospheric chemistry (Units SA1–11; UM1–3) 203.2.4 Geothermal activity (Units GT1–2) 213.2.5 Absence of light—subterranean caves and deep depressions in karst (Unit CV1) 213.2.6 Combustion and/or volcanic activity (fire) (Units VS1–14) 21
4. Discussion 23
4.1 Conceptual rigour 23
4.2 Intuitive and comprehensible 234.2.1 Aligning the physical environment with the biotic 234.2.2 Accommodating previous woody vegetation classifications 244.2.3 Dealing with human-modified vegetation states 244.2.4 Challenges of the alpine zone 254.2.5 Inferences about ecosystem relationships 25
4.3 Comprehensiveness and scale explicitness 25
4.4 Relationships between seral and steady states 26
4.5 Spatial explicitness and mappability 27
5. Conclusions 28

6. Acknowledgements 29
7. References 29
Appendix 1
Glossary of common and scientific names of plant species 36
Appendix 2
Ecosystem framework 41
Appendix 3
A classification of New Zealand terrestrial ecosystems 51

1Science for Conservation 325
A classification of New Zealand’s terrestrial ecosystems
Nicholas J.D. Singers1,2 and Geoffrey M. Rogers3
1 Department of Conservation, Private Bag 2, Turangi 3353, New Zealand2 Present address: 44 Raukura St, Turangi 3334, New Zealand3 Department of Conservation, PO Box 5244, Dunedin 9058, New Zealand Email: [email protected]
AbstractThe need to classify New Zealand’s diverse and complex ecosystems is driven not only by scientific curiosity, but also by increased land use planning activity. The classification of ecosystems, as opposed to vegetation, involves the description of relationships between the abiotic environment and its biotic overlay. Although the use of modern quantitative approaches to ecosystem classification would be a preferred option, these have had limited development and application across the full range of biodiversity in New Zealand. Therefore, recognising the need to incorporate physical variables, process variables and biota, we initially constructed a subjective, theoretical framework of environmental or physical drivers in New Zealand. This resulted in a three-variable hierarchy of temperature, moisture availability, and landform and soil gradients, which were divided sequentially down into categories or environmental classes. Vegetation classification literature and expert opinion were then used to align vegetation communities and ground cover classes with each of these environmental zones. A primary division within this classification was the conceptual distinction between zonal and azonal ecosystems, where zonal ecosystems are driven primarily by the physical, macroclimatic variables of temperature and moisture availability, and azonal ecosystems are primarily the product of process variables producing edaphic extremes (e.g. extreme rock and soil chemistry, extreme heat, and frequent disturbance). Thus, for the azonal section of the classification, the three-variable abiotic framework was applied after these additional process variables, which included geomorphic disturbance, frequent fire, geothermal heat and extreme soil chemistry. In total, this classification led to 152 ecosystems being recognised—78 zonal and 74 azonal. This is just one thematic scale of classification, with coarser or finer levels possible depending on purpose. Transition from this mainly qualitative ecosystem classification to quantitative approaches will occur as biodiversity databases and statistical modelling permit improved fits with natural geographic patterns.
Keywords: ecosystem classification, climate variables, New Zealand, conservation planning, vegetation maps
© Copyright May 2014, Department of Conservation. This paper may be cited as:Singers, N.J.D.; Rogers, G.M. 2014: A classification of New Zealand’s terrestrial ecosystems. Science for Conservation 325.
Department of Conservation, Wellington. 87 p.

2 Singers & Rogers—Classification of terrestrial ecosystems
1. Introduction
The ecosystem concept is at the centre of international agreements, New Zealand legislation, and modern policy and planning systems that aim to sustainably manage natural resources. All definitions of ecosystems include the concept of the physical environment being integrated with its biotic components. Functionally, the concept embodies disturbance cycles, and flows of energy, nutrients and non-living materials (Tansley 1935), with these processes underpinning the concept of ecosystem health or integrity (Whittaker 1975). Since these processes operate at variable spatio-temporal scales, and species and communities intergrade variably along environmental gradients, there is no single optimal scale at which to apply the ecosystem concept. Rather, the openness and hierarchical nature of ecosystem processes lead to any one classification scale being viewed as nested within coarser and finer scale components (Gauch & Whittaker 1981).
One of the goals of the New Zealand Biodiversity Strategy is to ‘maintain and restore a full range of remaining habitats and ecosystems …’ (DOC & MfE 2000: goal 3). However, although many environmental agencies and individuals can contribute to this goal, any investment decisions are currently being made in the absence of a comprehensive list of ecosystems or a systematic threat ranking. Therefore, classification of the full range of ecosystem types for New Zealand is overdue.
For any such classification to be useful to land managers, ecosystem types need to be discrete enough to map. Therefore, the question is how best to delineate ecosystems, measure them quantitatively and evaluate them qualitatively (Park 2000). The continuous variation in ecosystem composition, structure and processes, and our limited understanding of that complexity makes it extremely difficult to describe their distinguishing properties and map their distributional limits—which may explain the scarcity of comprehensive classifications that are suitable for providing lists for threat evaluation, despite progress in developing frameworks to guide classification (Nicholson et al. 2009).
Ideally, an ecosystem classification needs to meet the following broad criteria:
• Incorporates the four key elements of an ecosystem: the characteristic biota, the abiotic environment, the key processes and interactions, and its spatial distribution (Keith et al. 2013).
• Is intuitive and comprehensible, i.e. it has a conceptual foundation, a framework and a typology that is accessible to specialist and lay audiences alike.
• Is comprehensive and scale-explicit in terms of the level of detail it includes.
• Incorporates seral as well as intact vegetation states across environmental space because value can be attached to all biodiversity. Although successional relationships or potential trajectories are difficult to include in classifications, the variation within states of dynamic equilibria need to be recognised.
• Ultimately, results in individual ecosystems that are mappable.
Several plant community and ecosystem classifications dealing with particular New Zealand geographic areas or biomes have been produced in the past:
• The New Zealand Protected Natural Area Programme (PNAP) (Kelly & Park 1986; Myers et al. 1987) used an ecological unit concept that incorporated abiotic (landform) and biotic components to identify and name ecosystems. However, its geographical application focused on ecosystem types with high ecological integrity, overlooking more modified types and thus failing to produce a comprehensive inventory (Park 2000).
• Forest Class Maps were compiled at a scale of 1:250 000 (New Zealand Forest Service Mapping Series 6), with a small part of the North Island mapped at 1:63 360 (New Zealand Forest Service Mapping Series 5). However, the 1:250 000 series used the qualitative

3Science for Conservation 325
classification of McKelvey & Nicholls (1957), which was later modified in Nicholls (1976). Furthermore, although McKelvey (1984) quantitatively classified South Island forests, this was not used for mapping.
• The Vegetative Cover Map of New Zealand (Newsome 1987) provided coverage of vegetation communities at the relatively coarse scale of 1:1 000 000, with a minimum map unit area of 500 ha.
• The New Zealand Land Cover Database 2 (LCDB2) digitally classified and mapped 33 classes of land cover and land use from satellite imagery (Thompson et al. 2004). This classification placed all indigenous forest within one class, whereas there were eight shrubland classes. Although the minimum mapping unit was 1 ha, LCDB2 is known to have errors (sometimes substantial errors) in its class boundaries (Brockerhoff et al. 2008).
• The EcoSat woody vegetation layer used satellite imagery to classify woody vegetation by representing the proportions of beech, conifer and broadleaved tree species (Dymond & Shepard 2004, cited in Wiser & Hurst 2008).
• Several authors have produced maps of potential forest cover. Leathwick (2001) mapped 20 potential forest types using regression analysis to examine the relationship between the distribution of 37 canopy tree species and environmental variables (namely climate, landform and parent material). The modelling approach took account of the spatial discontinuities of the five species of beech (Leathwick 1998), and their competitive interactions with each other and with other tree species (Leathwick & Austin 2001). However, Leathwick (2001) discussed the limitations of this classification, as it used National Vegetation Survey (NVS) data, which were mainly compiled to record merchantable forest trees and thus lacked information on deforested, dry eastern environments. Hall & McGlone (2006) produced a forest biome map using the ecosystem process model LINKNZ, which used climate surfaces, landforms, slope and soil types in combination with 78 individually parameterised vascular plant species.
• At a national scale, Meurk (1984) compiled a bioclimatic zone classification primarily based on mean annual temperature and secondarily moisture balance, mainly to make global comparisons of New Zealand’s climatic and biogeographic profile. This work formed the basis for describing climatic envelopes for 33 representative forest tree species, which together cover the range of forest conditions in New Zealand (Meurk 1995).
• Land Environments of New Zealand (LENZ) numerically classified and mapped New Zealand’s physical environment at four nested spatial scales using an amalgamation of 11 climatic, landform and soil variables (Leathwick et al. 2003b). The selected variables had high statistical correlations with tree distributions and strong functional relevance to the physiology of trees. Further, terrestrial bioregions (termed environmental domains) have been classified based on climate, landform and vegetation types to represent patterns of potential vegetation (Leathwick et al. 2003a).
• Several authors have produced classifications specifically for wetland areas. Dobson (1979) produced a hierarchical classification of wetlands based primarily on fertility, pH and temperature variables, and the related broad plant community patterns. Johnson & Gerbeaux (2004) defined nine broad wetland classes based on substrate variables, water regime, and the consequent nutrient status and pH. The latter work formed the framework for a national map of wetlands (WONI—Waters of National Importance; www.landcareresearch.co.nz/services/informatics/ecosat/applications.asp#Wetland). In addition, Chadderton et al. (2004) divided New Zealand into 29 catchment units to produce a freshwater biogeographic framework for riverine biodiversity assessment; and Leathwick et al. (2007) used the same 29 biogeographic units to assess the importance of all freshwater wetlands for conservation planning.
• An alpine ecosystem classification was produced by Mark & Dickinson (1997) by combining climate, landform and habitat variables with field data to describe structural classes of vegetation.

4 Singers & Rogers—Classification of terrestrial ecosystems
• There have been various vegetation classifications for New Zealand’s major offshore islands: Sykes (1977) described broad vegetation types on the Kermadec Islands; Kelly (1983) described 17 primeval vegetation types on the Chatham Islands; Meurk et al. (1994) classified Campbell Island/Motu Ihupuku vegetation; and Wilson (1987) described coarse- and fine-scale vegetation communities on Stewart Island/Rakiura.
• Williams et al. (2007) produced a physiognomic framework for 72 historically rare ecosystems based on their perceived individual national extent of < 0.05% of New Zealand’s total land area. The framework used combinations of seven descriptors of the abiotic environment spanning its edaphic, climatic and disturbance components. Given the extremes of some environmental variables, non-forest ecosystems dominated the list. This framework responded to a lack of comprehensive treatment of these extreme environments and ecosystems by previous classifications.
• Wiser et al. (2011) classified New Zealand’s woody vegetation into 24 forest and shrubland alliances, each with estimated extents of 144 000–794 000 ha, using an 8-km-grid square network of plots.
All parts of New Zealand’s biological environment have been covered by one or other of the above classifications. However, these have been applied at variable spatial scales and have incorporated selective vegetation attributes, and none of these classifications meet all of the criteria listed previously—i.e. providing national-scale, mappable coverage of a full range of terrestrial ecosystems, with alignment to an abiotic framework of environmental drivers and associated processes, and scale-versatility. LENZ provides the most comprehensive treatment of the abiotic environment and can also be used to predict the ecological potential of cleared sites by cross-referencing to comparable environments with steady-state vegetation. However, there is a conspicuous absence of a classification that attempts to biotically ‘truth’ numerical classifications of physical environments.
Joseph et al. (2008) and Moilanen et al. (2009) provided an account of modern quantitative approaches to spatial conservation prioritisation, in which the benefits of explicitness, repeatability, error quantification and, therefore, scientific credibility were emphasised. In New Zealand, Leathwick et al.’s (2003b) numerically-based classification of the physical environment offered objectivity, consistency, error quantification and understanding of hierarchical relationships between units. However, not all quantitative approaches to spatial conservation prioritisation are explicit (Ferrier & Wintle 2009). Expert opinion and intuition can play a key role in the design of systems for spatial choice in conservation, particularly where quantitative spatial data are limited, e.g. when defining ecologically extreme environments that typically have poorly understood and/or deficient numerical data for their depiction. Such a deficiency of LENZ was recognised by Williams et al. (2007), who adopted a modular or multi-scale qualitative (sensu Corner et al. 2003) scheme that subjectively combined unweighted abiotic and biotic descriptors to depict extreme environments.
In the present study, we aimed to produce a comprehensive terrestrial ecosystem classification by subjectively constructing a hierarchy of perceived key environmental drivers, which included climatic variables, as well as substrates, soils and landforms. This provided an abiotic framework within which we could subjectively place selected biotic communities drawn from the various classifications listed above. Our biotic component of ecosystems involved subjective selection, amalgamation and limited modification of existing classifications, as well as, in some instances, the creation of novel communities. The placement of forest communities within the abiotic framework was aided by GIS comparisons of climatic patterns with forest class distributions (see section 2.2), but the placement of predominantly non-forest communities within environments that are determined principally by process variables was an entirely subjective process. One benefit of utilising existing vegetation classifications is that end-users are likely to be familiar with the conventions and typologies of existing schemes, thus providing a conceptual and geographical context for names.

5Science for Conservation 325
1.1 Defining the terrestrial environmentIn this study, we define the terrestrial environment as all land in New Zealand’s territorial region that is temporarily or permanently above mean low water spring, including the foreshore (as defined in the Conservation Act 1987). This aligns with estuarine communities of eelgrass (Zostera muelleri subsp. novozelandica)1, which is the ecological limit of flowering plants in the marine environment in New Zealand (Wardle 1991). Freshwater wetland types that are dominated by water-emergent plants rooted in soil are included, but aquatic macrophytes are excluded (see also Williams et al. 2007). Thus, largely permanent water bodies such as ponds, lakes, lagoons and rivers below the depth of rooted plants (littoral zone) are excluded. An exception to this rule is where geothermal and cave systems have aquatic components. We do not classify nival environments above the upper altitudinal limit of vascular and non-vascular plants and cryptograms (lichens and mosses), as ecosystem conservation management does not occur beyond this limit.
2. Methods
2.1 Structure of the classificationAny description of an ecosystem should encompass the four elements that define its identity: the abiotic environment, the characteristic biota, the key processes and interactions, and its spatial distribution. In addition, emphasis should be placed on its salient and unique features, and its distinctions and relationships with other associated units (after Keith et al. 2013).
Our abiotic framework was constructed using up to four hierarchically arranged environmental variables or processes. It is important that an abiotic framework incorporates a qualitative understanding and description of processes that govern ecosystem dynamics, as these are essential for assessing risks related to functional declines (Keith et al. 2013), and ideally it should be spatially definable. However, there are no national-scale datasets of some key variables, such as landforms, and we are limited to a conceptual or qualitative understanding of some key processes and their interactions, such as cold air ponding, flooding, sedimentation, waterlogging and disturbances such as landsliding. Climatic and edaphic variables (and even structurally important biota), particularly their extreme perturbations, can be viewed as processes affecting ecosystem function and dynamics. However, our use of process variables has been limited to those extreme physical forces that disrupt ecosystem expression by the principal climatic and edaphic variables. This is the azonal section of our classification (see below).
For a large part of the classification, temperature was the primary variable, moisture availability was the secondary variable, and lithology or landforms and soils (equivalent to fertility) was the tertiary variable. These variables are recognised as major determinants of plant species distribution at a regional scale (Gaston 1990) and have been used extensively in previous studies examining environmental predictors of biodiversity patterns (e.g. Belbin 1993; Pressey et al. 2000; Araujo et al. 2001; Faith et al. 2001; Leathwick et al. 2003b; Trakhtenbrot & Kadmon 2005). The temperature and moisture availability variables were divided into classes, the boundaries of which were perceived as corresponding to previously recognised bioclimatic boundaries (e.g. see Meurk 1984). Sequentially, the moisture availability classes were nested within the temperature classes and, in some instances, fertility classes were nested within the moisture availability classes. This approach can be viewed as a type of discrete classification (sensu Ferrier
1 See Appendix 1 for a glossary of common and scientific names of all plant species referred to in this report.

6 Singers & Rogers—Classification of terrestrial ecosystems
et al. 2009: 99) in that multiple environmental variables are selected first, following which expert opinion is used to divide the range of each variable into a small number of discrete classes.
We used the data of Leathwick et al. (2003b) to produce the mean summer temperature gradient. We then applied the following class or isotherm limits along that gradient:
• Subtropical: > 22.5°C
• Warm: > 17.5°C to < 22.5°C
• Mild: > 15°C to < 17.5°C
• Cool: > 12.5°C to < 15°C
• Cold: > 10°C to < 12.5°C
• Low alpine: > 5°C to < 10°C
• High alpine: < 5°C
These temperature classes were then further subdivided according to a gradient of moisture availability that was derived from the rainfall : potential evapotranspiration data of Leathwick et al. (2003b). Using the Thornthwaite (1948) typology of humid, sub-humid and semi-arid, the class divisions were as follows:
• Humid: Areas where rainfall is at least four times greater than evapotranspiration (cf. Thornthwaite’s (1948) definition of > 200 mm per annum) and which support predominantly mesophytic plants (Daubenmire 1974).
• Sub-humid: The humidity zone that lies between the humid and semi-arid class boundaries.
• Semi-arid: Areas where the average Penman annual water deficit is greater than 270 mm per annum (following Rogers et al. 2005; Walker et al. 2009) (cf. Thornthwaite’s (1948) definition of > 200 mm per annum deficit).
For forest ecosystems, the sub-humid/semi-arid boundary is quite closely aligned with the eastern limit of kāmahi (Weinmannia racemosa), which is strongly limited by edaphic and atmospheric moisture stress (Wardle 1966; Leathwick & Whitehead 2001). Kāmahi is more or less continuously distributed along the entire length of the North and South Island axial ranges, with limited extension into eastern parts where the height and breadth dimensions of the ranges permit the spill-over of westerly, rain-bearing airstreams, where southwesterly and easterly rain is influential in the summer, and where edaphic variables ameliorate extreme evapotranspiration. Therefore, in general, kāmahi is considered a good biological proxy for the sub-humid/ semi-arid boundary.
The third environmental gradient—landforms and soils—was considered to be less continuously influential on biological patterns than temperature and moisture availability, as most New Zealand soils have low availability of essential plant nutrients such as phosphorus and cations, irrespective of large-scale variability in substrate (Meurk 1995; Leathwick et al. 2003b). Accordingly, only perceived influential extremes of these edaphic variables were included at the tertiary level in the classification.
Using this approach, the spatial distribution of the temperature and nested moisture availability classes produced 15 zonal (see below) climatic classes across the three main islands and all inshore islands of New Zealand (Fig. 1). Because the distant offshore islands of New Zealand are not included in Fig. 1, the subtropical climatic zone covering the Kermadec Islands group is not included. Data limitations prevented the tertiary level influence of landforms and soils and a further temperature-driven climatic zone (cold air inversion topography) from being overlayed on the map.

7Science for Conservation 325
Figure 1. Macroclimatic zones of New Zealand.
E

8 Singers & Rogers—Classification of terrestrial ecosystems
Leathwick & Austin (2001) showed that the distribution of New Zealand’s beech species was poorly modelled by traditional environmental variables, and that all five2 species competitively excluded other broadleaved and conifer tree species from their potential environmental ranges. Therefore, to recognise this effect, the presence of beech-dominated communities was periodically included as an additional variable in the third level fertility gradient of the classification’s environmental framework.
That part of the classification where the vegetation is driven primarily by macroclimatic variables (principally temperature and moisture availability) was named the zonal section, producing a predictable ‘zonal’ pattern of communities (Thorp & Smith 1949; Tedrow & Cantlon 1958; New Zealand Soil Bureau 1968: 30; Walter 1985; Peinado et al. 2007). By contrast, in azonal environments, the vegetation is primarily the product of edaphic extremes, such as extreme rock and soil chemistry, extreme heat, and frequent disturbance by water, gravity and wind, which override the otherwise dominant influence of macroclimate. Azonal ecosystems often occur at a small scale and can appear anomalous within the context of the national and regional patterns of vegetation. For the azonal section of the classification, the extreme edaphic or process variables preceded the macroclimatic variables of temperature, moisture availability and fertility in the hierarchy of abiotic variables.
2.2 Classification and mapping of zonal environments and ecosystemsNicholls (1976) and McKelvey (1984) classified New Zealand forests into 18 North Island and ten South Island classes, respectively, each of which was further divided into many additional and more detailed forest types. For Stewart Island/Rakiura, Wilson (1987) described nine broad and 160 finer community types (both forest and non-forest). However, we found that some of the Nicholls (1976) and McKelvey (1984) forest classes were too broad or too general, and that most of their forest types were of too fine a level of detail to be used in our classification. Further, we found much overlap between the three aforementioned classifications. Therefore, we used ArcGIS 9.0–ArcMap version 9.1 (ESRI, Redlands, CA, USA) to compare the distribution of our 15 climatic zones (Fig. 1) with the mapped occurrences of 18 forest classes across New Zealand’s three main islands and inshore islands. The distribution of these forest classes was determined using digitised Forest Service Mapping Series 6 (1:250 000) maps, which are a distillation of the Nicholls (1974) and McKelvey (1984) classes. Three of the forest classes used in the Forest Service Mapping Series 6 (1:250 000) (Tawa (N), Tawa-taraire (S) and Tawa-beeches (O)) were amalgamated with their perceived steady-state counterpart classes that incorporated their previous podocarp component (e.g. Tawa (N) was incorporated in the Rimu tawa (D) class), as they were deemed to have been compositionally modified by logging of their merchantable podocarp element. For the parts of the southwest of the South Island that were not covered by the 1:250 000 data, we digitised the Te Anau, Hauroko and Mataura Forest Service Mapping Series 15 (1:1 000 000) maps. For Stewart Island/Rakiura, we used Wilson’s (1987) polygons for podocarp/hardwood forest (his Types A1 and A2 combined) and rātā-kāmahi hardwood forest (his Type B3). We then computed the percentage concordance between the forest classes and the climatic zones.
Forest classes that were predominantly found in a single climatic zone were adopted as zonal ecosystem units. For example, 86% of the Taraire, tawa, podocarp forest class occurred within the warm and sub-humid climatic zone in Fig. 1. Conversely, where forest classes were distributed across multiple climatic zones, had poor conformity with the zones, or were too broad or general,
2 Note that the taxonomy of New Zealand beeches has recently been revised, resulting in five species now being recognised (Heenan & Smissen 2013).

9Science for Conservation 325
we used the greater detail provided by the embedded forest types to create ecosystem units that were perceived as having a better representative fit with our climatic zones. For example, the Softwoods forest class, which was mapped across eight climatic zones and contained 18 gymnosperm forest tree species of potential diagnostic importance, was divided into several units containing diagnostic gymnosperm and hardwood tree species. Similarly, the General Hardwoods class of the 1:250 000 series of Nicholls (1976) contained what were perceived to be both steady-state forest types (e.g. P1, P2 and P3) and anthropogenically modified, secondary vegetation types (e.g. P4–P17); therefore, the steady-state types were included within the classification, while the secondary vegetation types were omitted.
Finally, distinct forest classes were assigned to the forests of some of the largest offshore islands that were not covered by Nicholls (1976) and McKelvey (1984), and which contain several narrow-range trees and shrubs (e.g. the Kermadec Islands, following Sykes (1977); and the Chatham Islands, following Kelly (1983)).
2.3 Classification and mapping of azonal environments and ecosystemsAzonal ecosystems form at sites that are exposed to extremes of process (principally edaphic) variables:
• Soil temperatures > 20°C, as occur in geothermally heated sites (Burns 1997).
• Temporary or permanent waterlogging, i.e. ephemeral and permanent wetlands (Johnson & Gerbeaux 2004).
• Geomorphic disturbance of a frequency that is sufficient to prevent the development of late successional vegetation. Recognised disturbance variables include gravity-induced erosion and accretion; freeze-thaw action; wind ablation; and flooding with associated sedimentation, including lahars on the central North Island volcanoes. Less frequent volcanic disturbances, such as pyroclastic, tephra, and lava flows and ash showers, are not included because their return intervals generally permit the return of primary successional forest. These disturbances lead to the formation of cliffs, screes, erosion pavements, beaches, active sand dunes and braided riverbeds.
• Edaphically extreme chemistry, resulting in ultramafic (ultrambasic) substrates, and atmospherically- and edaphically-derived extreme salinity.
• Insufficient solar energy for photosynthesis to occur, with energy being derived from cave entrance sources of air, water, organic matter and, locally, lithotrophic microorganisms.
• High frequency and intensity of (primarily) anthropogenic fire.
We perceived these six categories of process variables to be hierarchically superior to the macroclimatic variables as ecosystem drivers in some environmental space. Accordingly, they form the basis of six sections of the azonal section of the classification, where these variables precede temperature, moisture availability and fertility (Appendix 2). Our azonal hierarchical framework of environmental drivers builds upon the unweighted, multifactorial framework of drivers that was used by Williams et al. (2007) for identifying and describing 72 historically rare, predominantly azonal ecosystems.

10 Singers & Rogers—Classification of terrestrial ecosystems
2.4 Scale and boundaries of ecosystemsThere is no optimum or universal scale at which ecosystems should be depicted and delineated (Park 2000: 80). Functionally, ecosystem boundaries are open and dynamic, so their classification and mapping is an arbitrary and simplified depiction of ecological and historical complexity. Ecosystem boundaries may be biotically discrete (e.g. some wetlands) or diffuse (e.g. lowland forests). Ecosystem mosaics at fine spatial scales result from inconstant or fluctuating environmental gradients and/or regular cycles of disturbance that generate mixed-age seral vegetation. Some examples are variably-shallow water tables, flooding-induced disturbance in braided rivers, mobile sand dunes, and juxtaposed acid and alkaline soils on the tops and sides of karst blocks, respectively. In recognition of these principles, we focused less on the distributional limits of perceived indicator or faithful species, preferring to consider the distributional limits of sets of structurally important species or functional types of species as arrayed along primary, secondary and tertiary environmental gradients. Accordingly, in our ecosystem unit descriptions, we sometimes use the terms transitional, discrete and mosaic to highlight this variability or imprecision in scale and boundaries.
We used the following conventions in our ecosystem labels and descriptions:
1. Atkinson’s (1985) structural classes were used to label vegetation communities and ground cover types. Structural class terms were prefaced with binomial, genus, or species and taxonomic group tag names, e.g. ‘Podocarp, broadleaved forest’.
2. The species or taxonomic group terms preceding the structural class terms were those of structural or physiognomic dominance. Individual species and taxonomic groups followed a generalised order of dominance, and notes on their abundance often used terms from McKelvey & Nicholls (1957).
3. Some azonal ecosystems included multiple structural class terms to cover their mosaic patterns. For instance, braided rivers are mosaics of disturbance-related bare ground cover classes and early successional vegetation communities.
4. The descriptions of ecosystem ranges sometimes included notes on greater historical distributional extents in relation to today’s truncated environmental ranges.
5. Reference literature is provided for all ecosystems.

11Science for Conservation 325
3. Results
The application of the current classification resulted in 152 ecosystems being recognised, 78 of which are zonal and 74 of which are azonal (predominantly non-forest) (Appendices 2 & 3). The zonal ecosystems are further separated into eight broad groups according to temperature: five groups ranging from subtropical to cold, which predominantly include forest ecosystems below the treeline; two alpine groups for environments that predominantly lie above regional treelines; and a somewhat anomalous group that recognises cold air ponding or extreme frost in inversion topography. The azonal ecosystems are separated into 11 major groups according to edaphic extremes, as these were considered the most important abiotic drivers, with temperature and humidity included at secondary levels.
Appendix 2 outlines the abiotic framework of the classification, and provides ecosystem codes and names, while Appendix 3 gives a description of the composition, variability and distribution of each ecosystem. Appendix 3 also lists the references that guided the recognition and characterisation of each ecosystem, indicating equivalence with community or ecosystem names in the sourced literature. Table 1 shows the perceived range of temperature and moisture availability classes for each forest ecosystem, along with the class listing for each in Appendices 2 and 3.
3.1 Zonal ecosystems 3.1.1 Subtropical forest (Unit SF1)
We recognise a Kermadec pōhutukawa forest (Metrosideros kermadecensis) as the only subtropical forest ecosystem in New Zealand, with wet and dry climate variants (Table 1).
3.1.2 Warm temperate forest (Units WF1–14)Fourteen forest ecosystems occur within the warm temperate zone (17.5–22.5°C mean summer temperature), which is predominantly frost-free (Appendices 2 & 3). The southern and inland distributional limits of kohekohe (Dysoxylum spectabile) are an approximate biological proxy for the boundary of this zone. In terms of moisture availability, the warm temperate zone is divided into humid, sub-humid and semi-arid zones. Various landform types then provide further discrimination, including coastal and inland hillslopes and hillcrests, alluvial floodplains, and stable coastal dunes. Two ecosystem units—Tōtara, mataī, ribbonwood forest (WF2) and Pūriri forest (WF7)—occupy high-fertility alluvium and basaltic substrates, the latter of which has melanic granular soils (Meurk 1995; Conning 2001).
In the northern part of this zone, Kauri, podocarp, broadleaved forest (WF11) is the most widespread but sparse ecosystem unit because it is so relictual. It predominantly occurs in the warm, sub-humid climatic zone, with a smaller proportion also being found in the warm, humid zone (Table 1). Kauri forest (WF10) also predominantly occurs within the warm, sub-humid climatic zone, with a smaller representation in the warm humid zone (Table 1); and with only 6239 ha remaining of an estimated coverage of > 200 000 ha in the 1840s, this forest type is also highly relictual (Halkett 1978, cited in Conning 2001), with most extant patches occurring within Northland and the Waitakere Ranges.
By contrast, the Pōhutukawa, pūriri, broadleaved forest (WF4) and Taraire, tawa, podocarp forest (WF9) ecosystems are well represented. Although beech (predominantly hard beech Fuscospora truncata) is also present within the warm climatic zone, its limited distribution and generally low abundance precludes it from forming an ecosystem unit that is separate from the Kauri, podocarp, broadleaved, beech forest (WF12) and the Kāmahi, tawa, podocarp, hard beech forest

12 Singers & Rogers—Classification of terrestrial ecosystems
ECOSYSTEM UNIT CODE AND NAME CLIMATIC ZONE
SUB-
TROPICAL
WARM MILD COOL COLD
SE
MI-A
RID
SU
B-H
UM
ID
√H
UM
ID
SE
MI-A
RID√
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SF1: Kermadec pōhutukawa forest 4
WF1: Tītoki, ngaio forest 4 3
WF2: Tōtara, mataī, ribbonwood forest 4 3 3 3
WF3: Tawa, tītoki, podocarp forest 4 3 3 3
WF4: Pōhutukawa, pūriri, broadleaved forest [Coastal broadleaved forest]
3 4 3
WF5: Tōtara, kānuka, broadleaved forest [Dune forest]
4
WF6: Tōtara, mataī, broadleaved forest [Dune forest]
4 3
WF7: Pūriri forest 4 3
WF8: Kahikatea, pukatea forest 3 4 3 3 3 3
WF9: Taraire, tawa, podocarp forest 4 3
WF10: Kauri forest 4 3 3 3
WF11: Kauri, podocarp, broadleaved forest
4 3 3
WF12: Kauri, podocarp, broadleaved, beech forest
4 3 3 3
WF13: Tawa, kohekohe, rewarewa, hīnau, podocarp forest
3 4 3
WF14: Kāmahi, tawa, podocarp, hard beech forest
4 3
MF1: Tōtara, tītoki forest 3 4
MF2: Rimu, mataī, hīnau forest 4 3
MF3: Mataī, tōtara, kahikatea, broadleaved forest
4 3
MF4: Kahikatea forest 3 4 3 3
MF5: Black beech forest 3 4 3
MF6: Kohekohe, tawa forest 4 4
MF7: Tawa, kāmahi, podocarp forest 4 4
MF8: Kāmahi, broadleaved, podocarp forest
4 4 3 3
MF9: Tānekaha forest, locally with beech 4 4
MF10: Tōtara, mataī, kahikatea forest 4 4
MF11: Rimu forest 4 3
MF12: Rātā, hard beech, kāmahi forest 4 4
MF13: Kahikatea, northern rātā, kāmahi forest
3 4
MF14: Kahikatea, silver pine, kāmahi forest
4 3
MF15: Kahikatea, mataī, Westland tōtara, rimu forest 4 3
Table 1. The perceived range of temperature and moisture avai labi l i ty c lasses for each forest ecosystem. Further detai ls of each ecosystem are provided in Appendices 2 & 3. See Appendix 1 for a glossary of p lant names. ECOSYSTEM UNIT CODE AND NAME CLIMATIC ZONE
SUB-
TROPICAL
WARM MILD COOL COLD
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
MF16: Rimu forest 4 3
MF17: Rimu, kāmahi, tāwheowheo forest 4 3
MF18: Yellow silver pine, mānuka forest 4 3
MF19: Kāmahi, rimu, miro, southern rātā forest
4 3 3
MF20: Hard beech forest 3 4 3 3
MF21: Tawa, kāmahi, rimu, northern rātā, black beech forest
3 3 3 4
MF22: Tawa, rimu, northern rātā, beech forest
3 4 3 3
MF23: Chatham Island akeake, karamū, māhoe, ribbonwood forest
4
MF24: Rimu, tōwai forest 4
MF25: Kauri, tōwai, rātā, montane podocarp forest
4
CLF1: Hall’s tōtara, mountain celery pine, broadleaf forest
3 3
CLF2: Hall’s tōtara forest [Dune forest] 4
CLF3: Podocarp, ribbonwood, kōwhai forest
3 4
CLF4: Kahikatea, tōtara, mataī forest 3 4
CLF5: Mataī, Hall’s tōtara, kāmahi forest 4 3
CLF6: Kāmahi, southern rātā, podocarp forest
4 3
CLF7: Rimu, kāmahi, beech forest 4
CLF8: Silver beech, kāmahi, southern rātā forest
4
CLF9: Red beech, podocarp forest 3 3 3 4
CLF10: Red beech, silver beech forest 3 3 3 3 4 3 3 3
CLF11: Silver beech forest 3 3 4 3 3
CLF12: Silver beech, mountain beech forest
3 4 3 3
CDF1: Pāhautea, Hall’s tōtara, mountain celery pine, broadleaf forest
4 3
CDF2: Dracophyllum, Phyllocladus, Olearia, Hebe scrub [Sub-alpine scrub]
4 3
CDF3: Mountain beech forest 3 3 3 4 3 3
CDF4: Hall’s tōtara, pāhautea, kamahi forest
3 4
CDF5: Hall’s tōtara, pāhautea, kāmahi, southern rātā forest
4
CDF6: Olearia, Pseudopanax, Dracophyllum scrub [Sub-alpine scrub]
4
CDF7: Mountain beech, silver beech, montane podocarp forest
3 4
Continued on next page

13Science for Conservation 325
ECOSYSTEM UNIT CODE AND NAME CLIMATIC ZONE
SUB-
TROPICAL
WARM MILD COOL COLD
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
SE
MI-A
RID
SU
B-H
UM
ID
HU
MID
MF16: Rimu forest 4 3
MF17: Rimu, kāmahi, tāwheowheo forest 4 3
MF18: Yellow silver pine, mānuka forest 4 3
MF19: Kāmahi, rimu, miro, southern rātā forest
4 3 3
MF20: Hard beech forest 3 4 3 3
MF21: Tawa, kāmahi, rimu, northern rātā, black beech forest
3 3 3 4
MF22: Tawa, rimu, northern rātā, beech forest
3 4 3 3
MF23: Chatham Island akeake, karamū, māhoe, ribbonwood forest
4
MF24: Rimu, tōwai forest 4
MF25: Kauri, tōwai, rātā, montane podocarp forest
4
CLF1: Hall’s tōtara, mountain celery pine, broadleaf forest
3 3
CLF2: Hall’s tōtara forest [Dune forest] 4
CLF3: Podocarp, ribbonwood, kōwhai forest
3 4
CLF4: Kahikatea, tōtara, mataī forest 3 4
CLF5: Mataī, Hall’s tōtara, kāmahi forest 4 3
CLF6: Kāmahi, southern rātā, podocarp forest
4 3
CLF7: Rimu, kāmahi, beech forest 4
CLF8: Silver beech, kāmahi, southern rātā forest
4
CLF9: Red beech, podocarp forest 3 3 3 4
CLF10: Red beech, silver beech forest 3 3 3 3 4 3 3 3
CLF11: Silver beech forest 3 3 4 3 3
CLF12: Silver beech, mountain beech forest
3 4 3 3
CDF1: Pāhautea, Hall’s tōtara, mountain celery pine, broadleaf forest
4 3
CDF2: Dracophyllum, Phyllocladus, Olearia, Hebe scrub [Sub-alpine scrub]
4 3
CDF3: Mountain beech forest 3 3 3 4 3 3
CDF4: Hall’s tōtara, pāhautea, kamahi forest
3 4
CDF5: Hall’s tōtara, pāhautea, kāmahi, southern rātā forest
4
CDF6: Olearia, Pseudopanax, Dracophyllum scrub [Sub-alpine scrub]
4
CDF7: Mountain beech, silver beech, montane podocarp forest
3 4
Table 1—continued.

14 Singers & Rogers—Classification of terrestrial ecosystems
(WF14) units. Beyond the southern limits of kauri (Agathis australis), the tawa (Beilschmiedia tawa)-, broadleaved-, and podocarp-dominated units (WF3, WF13 and WF14) dominate, occurring in humid, sub-humid and semi-arid climatic zones.
Northland forest ecosystems within the warm climatic zone are the most species rich in terms of trees in New Zealand, with up to 36 species present (Meurk 1995).
3.1.3 North and South Island mild forests (Units MF1–25)Twenty-five forest ecosystems occur in areas where mean summer temperatures range from 15°C to 17.5°C (Appendices 2 & 3). The upper elevational limits of tawa across its entire latitudinal range are a useful biological proxy for the altitudinal limit of this zone. This temperature zone is split into humid, sub-humid and semi-arid moisture availability zones; and further division separates coastal, inland and alluvial (with recent soils) landform zones. Unusual landforms and their associated soils also distinguish further units, namely welded volcanic ignimbrites and glacial outwash terraces. Soil type is an additional tertiary driver in this zone, where high rainfall and/or low potential evapo-transpiration have led to the formation of podzols and organic soils, or where relatively recent soils of volcanic origin have been derived from Taupo Pumice and other volcanic ashes.
At the broadest compositional level, almost all of these 25 ecosystems are mixes of podocarp and broadleaved tree species that are environmentally filtered according to climate and edaphic fertility (Appendices 2 & 3). The often emergent podocarp element can be broadly divided into mataī (Prumnopitys taxofolia), tōtara (Podocarpus totara) and kahikatea (Dacrycarpus dacrydioides) on the higher fertility sites (with the first two being sub-humid and semi-arid specialists); rimu (Dacrydium cupressinum) and miro (Prumnopitys ferruginea) on more weathered soils and in humid and sub-humid climates; and Hall’s tōtara (Podocarpus cunninghamii) also on lower fertility soils at higher altitudes, albeit across a wide moisture availability gradient.
In terms of the broadleaved component, tawa is prominent in the North Island in both the sub-humid and semi-arid climatic zones (MF6, MF7 and MF21). Kāmahi is co-dominant with tawa in the humid climatic zone, e.g. MF7, but not in the semi-arid zone of the two main islands. In the humid zone of the South Island, rimu and kāmahi are co-dominant or individually dominant, depending on landform (e.g. MF16, MF17 and MF19). Beech species (mainly hard beech and black beech Fuscospora solandri) occur in several mild forest units, especially on steep and/or thin soil landforms within the humid zone.
3.1.4 Cool forest and scrub (Units CLF1–12)Twelve forest ecosystems occur in areas where mean annual temperatures range from 12.5°C to 15°C. These are found across the three moisture zones (Appendices 2 & 3). Coastal sand dunes, hillslopes and hillcrests, and alluvial plains (with recent soils) landform zones are recognised, and podzol soils on stable landforms that are subject to high rainfall are also influential. This zone covers some of the interior North Island, and is extensive across the montane and southern lowland zone of the South Island (Fig. 1). Cool, humid zone forests often occur in tectonically active uplands that are prone to extreme storm events, within which land-sliding produces mosaics of different-aged soils and attendant forest communities. CLF1 would once have been prominent in much of the semi-arid eastern South Island east of the sub-humid beech forests that are found on the eastern flanks of the Southern Alps/Kā Tiritiri o te Moana. However, only tiny and/or fragmented relicts of this forest type remain in the driest parts of Canterbury and Otago due to its vulnerability to fires in these regions since the arrival of humans (Wells 1972). Units CLF3–5 are predominantly central North Island types that contain a strong podocarp element that is tolerant of frosts and summer dryness. CLF7–12 are predominantly montane central North Island and widespread South Island types, many of which contain a mix of beech and non-beech elements.

15Science for Conservation 325
3.1.5 Cold forest and scrub (Units CDF1–7)These seven ecosystems predominantly occur in the treeline zone and are dominated by either beech, conifer or subalpine scrub (Appendices 2 & 3). Units CDF1, 4 and 5 are upland conifer forests that have a prominent pāhautea (Libocedrus bidwillii) component and are compatible with high cloud cover. CDF2 and 6 consist of species-rich and compositionally variable subalpine scrub, which tend to be found in the North Island and central Westland beech gap zones, as well as in a highly fire-modified state in Otago. CDF3 is a climatically widespread type that occurs along the axial ranges of both main islands (Table 1). CDF7 is compositionally quite variable in its central North Island and southern South Island strongholds.
3.1.6 Low alpine and subantarctic (Units AL1–9)Nine ecosystems occur between the mean summer isotherms of 5°C and 10°C, which are referred to as the low alpine zone by Meurk (1984) and Mark & Dickinson (1997) (Appendices 2 & 3). The 10°C summer isotherm correlates with the treeline in New Zealand and many other countries (Daubenmire 1954; Wardle 1971). However, there are some exceptions to this (Mark & Dickinson 1997). For example, in the Kaimanawa Mountains of the North Island, warmer summer temperatures than this are experienced above the treeline (Körner & Paulsen 2004), perhaps reflecting the influence of strong insularity; and on the subantarctic islands, where the entire temperature environment is marine-influenced and there is extreme wind-chill, the treeline is lowered to below what would be predicted by a latitudinal gradient (Meurk 1984). New Zealand treelines may be clearly evident where intact beech forest remains. However, where fire has disrupted montane forest and scrub, or in beech-free regions where other dicotyledonous and coniferous shrubs and trees form a gradual transition from forest to shrubland, delineation of the treeline is problematic (Wardle 1964). Indeed, in beech-free regions, the altitudinal limit of trees is lower than latitudinally-equivalent beech treelines. Moreover, the biological reality is that in addition to the 10°C mean summer isotherm, infrequent extreme low temperature events also influence the treeline, with seedlings of New Zealand’s highest ascending tree species (mountain beech Fuscospora cliffortioides or silver beech Lophozonia menziesii) being killed at temperatures between –9.0 ± 0.8 and –12.4 ± 0.1°C, depending on the provenance of local seedlings (Sun & Sweet 1996). Therefore, for convenience, we define the lower altitudinal limit of the low alpine zone as being the natural treeline or, alternatively, the upper altitude of continuous scrub or shrubland in beech-free regions. The upper altitudinal limit of the low alpine zone is at the natural tussock-line, where tall tussock species (Chionochloa spp.) give way to sub-shrubs, short grasses and turf-forming, often cushion-shaped herbs (after Mark & Dickinson 1997). The delineation and description of low alpine ecosystems was principally influenced by the distribution, composition and abundance of Chionochloa tussock species, following Wardle (1991) and Connor (1991).
The low alpine zone contains sub-humid and humid zones, which include two and seven ecosystem units, respectively. Most low-alpine areas seldom experience edaphic moisture deficits due to frequent precipitation and low potential evapotranspiration rates (Mark & Dickinson 1997; Molloy 1998). However, the combination of raw and thin soils, steep slopes, northern aspects, high solar radiation, and wind exposure can result in site-specific periodic drought events (Mark & Dickinson 1997), especially east of the Main Divide in the South Island, which experiences drought in > 25% of years (Coulter 1966). Therefore, we have included a sub-humid zone in our classification, specifically for areas east of the Main Divide in the South Island. It is difficult to define the boundary between the humid and sub-humid zones due to imprecision in alpine climatic data, and the complexity of fine-scale topographic and edaphic variation. Consequently, we used a biological proxy in the form of the western distributional limit of Chionochloa macra (see Meurk 1982; Connor 1991). This tussock occurs exclusively in the east, almost along the entire length of the South Island from Southland to South Marlborough. Beyond the northern distributional limits of C. macra, the sub-humid zone extends northwards along the

16 Singers & Rogers—Classification of terrestrial ecosystems
Wairau River, with mountains to the west being humid and to the east being sub-humid. Within these two moisture zones, further environmental subdivision is based on landform, soil and plant biogeographic patterns.
3.1.7 High alpine (Units AH1–4)Our high alpine zone extends above the mean summer 5°C isotherm (after Meurk 1984; Mark & Dickinson 1997). Biologically, the lower limit of the high alpine zone is marked by the limits of tall tussock vegetation (after Mark & Dickinson 1997). For our purpose, we defined the upper limit by the altitudinal threshold of vascular and non-vascular plants (mainly mosses and lichens), effectively incorporating the nival zone or the area above the lower limits of permanent snow and ice. Two moisture availability classes are then recognised. High alpine ecosystems predominantly have raw or recent soils; however, despite the apparent uniformity of these soils in this zone, we have singled out the andesitic scoria and ash soils of the central North Island, and the poorly-drained, acidic brown soils on the summits of Central Otago’s schist mountains (Brumley et al. 1986; Dickinson 1988, 1989) as diagnostic of two comparatively restricted ecosystem units.
The four ecosystems occurring in this zone may seem small in number for an environment of such apparent botanical (Brumley et al. 1986; Dickinson 1988, 1989; Fagan & Pillai 1992; Mark 2013) and ecological diversity. The high alpine zone usually contains complex mosaics of plant communities at varying spatial scales, reflecting sharp changes in physiographic gradients such as those of wind exposure, snow accumulation and duration, snow and rock avalanching, stormwater and frost-heave erosion, and soil and substrate type. Consequently, three of our four ecosystem labels contain a mix of structural, mostly bare ground cover classes, reflecting that mosaic complexity and the sparseness of high alpine plant cover. Indeed, even where the Dracophyllum muscoides cushionfield (AH2) label might be applied, diverse bare ground and plant community mosaics can predominate.
3.1.8 Cold temperature inversion (Units TI1–6)Below the regional treeline, there are several scrub or shrubland communities that characteristically consist of phenolic-rich and schlerophyllus species that occupy cold air inversion-prone topography, such as valley floors, intermontane basins and montane plateaus. Colloquially, such inversion communities are often referred to as frost flats or frost-hollow vegetation (Williams et al. 2007). These communities, along with fire-induced grassland derivatives (Walker & Lee 2000), contrast with the surrounding hillslopes, which support taller forest or potential forest in fire-modified districts (Wardle 1991: 81, fig. 5.4). While intense frost (probably at temperatures of < –9°C (Wardle 1971, 1991: 81; Sun & Sweet 1996)) is likely to be a critical and, in many instances, a primary driver that excludes tall trees from this low-lying topography, other physical stressors are also likely to be at play, including summer drought on stony or pumice substrates, soil nutrient impoverishment, and high water tables. Consequently, it is difficult to assign a theoretical or representative hierarchy of environmental influences for all examples in this frost-prone environment. Nevertheless, we have selected cold temperature inversion as the primary driver because, in the instances where fire has not disrupted catchment-scale vegetation patterns, there is a characteristically abrupt margin to peripheral tall forest encircling a scrub- or shrubland-covered depression floor, with no discernible change in edaphic profiles across the transition. Physiologically, ponded heavy frost probably selects against the survival of tree seedlings and saplings, rather than trees themselves, within the hollows (P. Wardle 1971). Such recruitment inhibition is manifest as a characteristically abrupt margin to peripheral tall trees encircling the depressions abutting basin floor scrub or shrubland. Corroborating evidence that the peripheral limits of the tall trees are due to the frost-intolerance of the seedlings of these species comes from a subalpine treeline study (another extreme cold temperature forest boundary), which concluded that although frosts may cause some foliar damage, especially in spring, the effects of frosts on mature trees are unlikely to control the position of the New Zealand treeline (Cieraad et al. 2012).

17Science for Conservation 325
The six ecosystems occurring in this zone span the three moisture availability classes (semi-arid through to humid) and, in broad terms, span an eastern dry/cold to western wet/cold, lowland to montane environment (Appendices 2 & 3). Contrasts in soil fertility then produce wide woody compositional variation between the six ecosystem types. On the coldest and most nutrient-poor, often glacially-derived landforms, we can confidently ascribe the coniferous woody communities of mainly bog pine (Halocarpus bidwillii) and mountain celery pine (Phyllocladus alpinus) (TI1) primarily to frost inversion and secondarily to nutrient impoverishment (Wardle 1991: 193; McGlone & Moar 1998; Rogers et al. 2005), with the two species tolerating –23°C and –20°C, respectively (Sakai & Wardle 1978). Drought may play an equally important role to frost in the derivation of these communities because they occur predominantly in the South Island’s dry interior. Where there is relatively higher fertility in interior South Island sites on colluvial fans, alluvial terraces and floodplains, diverse woody communities of small-leaved (often divaricate/filiramulate and sclerophyllous) shrubs, short broadleaved trees and liane species dominate (TI2, TI4) (Wardle 1991: 207–211; Rogers et al. 2005; Walker et al. 2006). The ignimbrite landforms with tephra soils of the central North Island mostly contain fire-modified shrublands dominated by the endemic shrub Dracophyllum subulatum (Smale 1990; Smale et al. 2011), although bog pine and mountain celery pine are often equally abundant in the least fire-impacted locations (TI3) (Elder 1962; Bishop 2005). Humid valleys west of the axial mountains contain diverse scrub and short forest of conifers and wire shrubs (otherwise referred to as divaricate, filiramulate, small-leaved and lianoid shrubs), and/or red tussock (Chionochloa rubra) grasslands on poorly drained, frosty sites, making it difficult to apply a hierarchy of environmental drivers (TI5, TI6). Indeed, in many depressions, wetland ecosystems interfinger with other communities on only slightly elevated, better-drained interfluves, making it difficult to determine the hierarchical relationship between cold air inversion and poor drainage as drivers in vegetation pattern. The characteristically phenolic-rich and schlerophyllus foliage of much of the shrubland and scrub that occurs on inversion topography renders this vegetation fire-prone. Consequently, anthropogenic fire has extensively fostered grassland over the woody vegetation that once occurred in these ecosystems, and has variously modified the margins of the fringing forest, fragmented or entirely removed hillslope forests, and removed the cold-adapted woody indicators of the topographic limits to the extreme frost (see Wardle 1991: 193–195).
3.2 Azonal ecosystemsWe recognise 11 broad divisions of azonal ecosystems across the process drivers of high water tables (1), geomorphic disturbance (5), light limitation (1), extreme soil and/or atmospheric chemistry (2), geothermally extreme heat and chemistry (1), and anthropogenic fire (1). In the abiotic framework of the azonal section, these environmental variables hierarchically precede temperature, moisture availability and landform/soil variables (Appendices 2 & 3).
3.2.1 High water tables—permanently or seasonally wet soils (Units WL1–22)Twenty-two wetland ecosystems are identified for which water-inundation, principally through impeded drainage, is the primary ecosystem driver (WL1–22); this includes areas that are either temporarily or permanently saturated. Within this zone, the abiotic environment is first subdivided according to water fertility, following Dobson (1979) and Johnson & Gerbeaux (2004), which yields three broad groups: oligotrophic systems known as bogs (WL1–9), mesotrophic systems known as fens and marshes (WL10–17), and eutrophic systems mainly known as swamps and marshes (WL18–22). These three fertility zones are then further subdivided according to mean annual temperature (after Dobson 1979), followed by substrate and landform type (after Johnson & Gerbeaux 2004). The broadest community types are used for the biotic labels, especially for fens and swamps, because almost all of these communities have been homogenised as a result of fire-induced loss of their woody steady-states and herbaceous diversity (McGlone 2009).

18 Singers & Rogers—Classification of terrestrial ecosystems
3.2.2 Frequent geomorphic disturbance (Units DN1–5; EP1; CL1–11; SC1; BR1–3)We have aggregated into one section of the classification all of the ecosystem groups that have resulted from catastrophic and/or frequent incremental erosion that produces sparsely-vegetated rock surfaces or successionally young vegetation on raw or recent soils. The different forms of erosion are all types of geomorphic disturbance, whether they result from the effects of gravity, water movement or wind. Geomorphic disturbance—in terms of its magnitude and/or frequency—is given primacy ahead of temperature, moisture and landform variables because it alone is responsible for the arrested successional potential of these ecosystems. Most of these ecosystems contain mosaics of secondary vegetation communities, some as advanced as short woody systems; however, their overriding physiognomy is of successionally early vegetation states. The ecosystem groups within the disturbance section are based on the physical disturbance agent and the broad landform types that result, namely active coastal sand dunes (DN1–5), erosion pavements (EP1), cliffs (CL1–11), screes and boulderfields (SC1), and braided rivers (BR1–3).
Active coastal sand dunes (Units DN1–5)
Active or mobile coastal sand dunes are aeolian (wind-formed) landforms that are distributed along 1100 km of the New Zealand coastline (Hesp 2000). Coastal dunes accumulate on coasts where abundant sand is delivered by persistent onshore winds and may host a sparse cover of specialised sand-binding plants (e.g. see Esler 1970). Fixed or relict dunes that are covered in mid- to late successional vegetation are excluded from this part of the classification. Sand dunes are mosaic ecosystems within which vegetation patterns are strongly related to landform mobility and age. We use the development of scrub vegetation and the appearance of incipient soil horizons as a guide to the transitional threshold from active to stable sand dunes. Five ecosystem classes are recognised in this category, which are separated according to temperature and biogeographic pattern (Cockayne 1909, 1911; Oliver 1910; Esler 1970; Wardle 1991). We have amalgamated dune plains (Logan & Holloway 1933; Esler 1969; Sykes & Wilson 1987) with finer-scale, low-relief features, such as deflation hollows, dune slacks, and small hummocks that are both dry-ground and temporary wetlands (Johnson & Rogers 2006; Williams et al. 2007), in the ‘active sand dune systems’ category—although there is an ecosystem process argument for affording them individual ecosystem status, despite their often small spatial extents.
Erosion pavements (Unit EP1)
Erosion pavements on wind-exposed topography are considered to take one of two geomorphic forms: pavements of massive and erosion-resistant rock that are largely bereft of soil; and accumulations of lag gravels on ablation-prone landforms. Igneous intrusive rocks (e.g. granite and diorite), strongly metamorphosed sediments (e.g. quartzose or arkosic sandstones and marble) or erosion-prone limestone all have rock pavement or lag gravels on wind- and frost-prone exposures. Some of these occur above the regional treeline, e.g. the quartz-rich granodiorite pavements on Mt Titiroa south of Lake Manapouri, and so should really be included as ‘fellfield’ in the alpine zone. However, their comparative lack of soil as a result of wind-stripping and the erosion-resistant basement lithology means that they greatly differ from the freeze and thaw origins of alpine fellfield gravels, making it more appropriate to include them in the disturbance section of the classification.
Coastal and inland cliffs (Units CL1–11)
Eleven ecosystems are recognised on vertical or very steep slopes that are exposed to gravity-induced erosion (which are variously termed cliffs, tors, scarps and escarpments in the literature, but referred to as cliffs hereafter). The talus accumulations that occur immediately beneath cliffs as a result of incremental fretting of rock fragments or debris avalanches are also included where they remain primarily unvegetated or in early successional vegetation states. The surface geomorphology of cliffs varies according to rock type and tectonic forces

19Science for Conservation 325
(Kennedy & Dickson 2007). Although some cliff substrates such as limestone, marble and dolomite have endemics amongst their regional and local floras, in the absence of a published literature, our perception is that early successional floras of cliffs are similar within the broad temperature and moisture zones demarcated here.
Cliffs can host lichen, bryophyte, herb, grass, fern, shrub and small tree species on the limited microhabitats where soil forms or where their roots are able to penetrate bedding and jointing fractures. The predictable, sequential vegetation change from lichens, bryophytes and herbs to shrubs and small trees is often not a feature of cliffs; rather, a heterogeneous mix of plant forms that reflects the microhabitat pattern is more usual. On Banks Peninsula, it has been found that slope, aspect, soil fertility and the intactness of the surrounding native forest are the primary potential drivers of high compositional heterogeneity on cliffs (Wiser & Buxton 2009). Applying that insight at a national scale, we would add rock texture to that list, which drives fissure provision and soil accumulation. Some iconic species such as Clianthus maximus, Pachystegia spp., Carmichaelia spp., Helichrysum spp. and Hebe spp. are diagnostically important in labelling some of our regional ecosystem types. Small seeps on cliff faces can support a distinct and often pendulous flora; and waterfall and cataract-derived spume also selects for a flora that requires an equable humidity. However, we have not included these as a separate ecosystem due to their typically small spatial expression and wide compositional variability.
In this classification, we recognise five coastal and six inland cliff ecosystems, with the former group being moderated at a quaternary level by salt spray. A wide range of structural vegetation classes are included within several of the ecosystem names to embrace the variable cover of plants and the compositional heterogeneity that occurs at micro- and meso-scales as a result of both environmental stress and substrate variability.
Screes and boulderfields (Unit SC1)
Screes are an accumulation of broken rock fragments of a size that ranges from gravel to boulders and which mantle hillslopes with a slope greater than c. 35°. Functionally, screes result from cliff outcrops that are composed of erodible rock of mainly greywacke, basalt and limestone with abundant bedding and jointing planes that are prone to freeze-thaw fragmentation. These continuously supply talus, producing a veneer of rock fragments on the steep slopes below, which can lead to a conveyor belt-like creep of rock particles veneering the hillslopes. Such a coarsely textured rock veneer and/or its instability have evolutionarily selected for a specialist, predominantly montane to alpine, vascular scree flora that invariably is at sparse cover and occurs across a wide climatic range Due to the lack of cover and the wide geographic range across which screes occur, we have not included a vegetation component in the ecosystem label. Most screes at high altitude in New Zealand are natural; however, anthropogenic vegetation clearance at montane to subalpine elevations has contributed to local expansion of screes and boulderfields in Canterbury (Whitehouse 1982; McSaveney & Whitehouse 1989).
Boulderfields are composed of coarser debris than screes, and the majority occur in the alpine zone. They have two predominant origins: gravity-induced rock avalanches, which are often seismically-triggered; and coarse moraine or in situ fragmentation debris. The poorly vegetated flow-basalt boulderfields of north Otago are a notable example of considerable antiquity.
Braided rivers (Units BR1–3)
Braided rivers result from an abundant supply of sediment being transported by frequent floods, producing wide and unstable river channels interspersed with small gravel terraces across flood plains. They are largely restricted to the South Island, with smaller examples in the central–eastern and southern North Island (Williams & Wiser 2004). Since braided channels migrate across flood plains over short geological timescales, the abandoned channels can eventually

20 Singers & Rogers—Classification of terrestrial ecosystems
support steady-state vegetation of tall forest or scrub, depending on the particle size and soil development of the substrate. In this classification, we recognise three braided river ecosystems, based on moisture availability and plant biogeography: two widespread (eastern BR1 and western BR2) and one narrow-range (the Mt Ruapehu–Whangaehu BR3). The Mt Ruapehu–Whangaehu braided channel has been singled out because it experiences frequent (decadal to multidecadal) acidic lahars from Mt Ruapehu’s crater lake (H. Keys, Department of Conservation, pers. comm. 2011). Braid bars can be made up of raw silts, sands, gravels and boulders, and can also support early successional communities of lichens, herbs, grasses, lianes and sub-shrubs in less frequently disturbed areas.
3.2.3 Extreme soil and/or atmospheric chemistry (Units SA1–11; UM1–3)We recognise two manifestations of extreme chemistry as the primary ecosystem driver: extreme salinity derived from both oceanic and edaphic sources, and extreme rock chemistry.
Saline environments (Units SA1–11)
In this classification, we recognise 11 saline ecosystems. In saline ecosystems, high concentrations of alkaline salts (predominantly sodium chloride) select for halophytic plants (which are tolerant of high concentrations of salt) and against glycophytic plants (which can only tolerate low concentrations of salt). In maritime environments, sea water delivers salt either tidally or from wave-splash, selecting for predominantly estuarine vegetation (Partridge & Wilson 1987, 1988; Thannheiser & Holland 1994; Johnson & Gerbeaux 2004). By contrast, in dry terrestrial maritime environments, salt is delivered aerially from spume and spray-drift, which, combined with extreme wind exposure, selects for prostrate herbaceous communities called coastal turfs (Johnson 1993; Rogers 1999; Rogers & Wiser 2010) or sparsely vegetated communities on gravel beaches (Wiser et al. 2010). This classification includes one forest and scrub and two herbaceous estuarine ecosystems (SA1–3), and two dry terrestrial maritime ecosystems (SA4–5).
In addition to the primary, marine-derived salinity, a further five of the 11 saline ecosystems (SA6–10) are affected by disturbance in the form of ground- and burrow-nesting seabirds and marine mammal wallowing, which is included as a tertiary level influence (see also Williams et al. 2007). Concentrated guano can also increase soil fertility and thus productivity (Fukami et al. 2006; Towns et al. 2009; Bellingham et al. 2010). These animal influences can lead to the inland extension of a normally coastal community (Gillham 1960a, b). They can also cause arrested vegetation successions, principally as a result of surface-nesting birds—although burrowing birds combined with high exposure and/or drought stress may also cause this (Wright 1980; de Lange et al. 1995). These arrested successions can range from bare guano-encrusted soil or rock (loamfield) through to a variety of herbaceous communities and scrub (sometimes referred to as mutton-bird scrub in southern New Zealand). These five ecosystems span a wide latitudinal range across which this combination of drivers occurs, ranging from the Kermadec Islands in the north to the subantarctic islands in the south. The structural vegetation that occurs in each abiotic zone can vary, reflecting the dynamic and sometimes inter-annual cyclical patterns of roosting and nesting seabirds and storm surges. During low stress and disturbance periods, these five ecosystems may succeed to short forest, in which case they would be classed as zonal forest, irrespective of the presence of seabirds and marine mammals (Stewart Island/Rakiura and the Titi/Muttonbird Islands provide pertinent extant examples). For instance, our description of some coastal forest ecosystems notes the common presence and abundance of seabirds (e.g. Kermadec pōhutukawa forest, SF1).
The final ecosystem in this category represents an inland saline ecosystem (SA11). Such ecosystems develop where semi-arid climates produce high edaphic concentrations of efflorescences of soluble salts on soil surfaces (Allen et al. 1997; Rogers et al. 2000). In New Zealand, they are confined to Otago, including the upper Waitaki valley, and support a sparse cover of halophytic plants that also tend to occur in estuarine ecosystems. Although,

21Science for Conservation 325
we have settled on just one inland saline unit, a previous classification using the concept of the topographic control on pedogenesis, and the saline and sodic expression of surface salts identified essentially four inland saline ecosystems—salt pans, salt plains, salt meanders and salt knolls (Rogers et al. 2000).
Ultramafic (ultrabasic) rocks (Units UM1–3)
In ultramafic ecosystems, soils have formed on parent materials that have low concentrations of most major nutrients, a wide Mg/Ca quotient, high concentrations of nickel, chromium and cobalt, and a high pH (Lee 1992). Ultramafic substrates weather very slowly, have raw or recent soils, and encompass a wide variety of landforms, including cliffs, scarps, tors, hillslopes, screes, erosion pavements and boulderfields. Ultramafic rocks across New Zealand have been mapped at a scale of 1:250 000 (www.gns.cri.nz/Home/Our-Science/Energy-Resources/Geological-Mapping/Geological-Maps/1-250-000-QMAP/The-QMAP-Project), and their communities and habitats have been comprehensively described and mapped by Lee (1992). In this classification, we recognise three ultramafic ecosystems based on temperature and landform.
3.2.4 Geothermal activity (Units GT1–2)Two ecosystem units (GT1–2) are driven primarily by extreme geothermal heat, which principally occurs within the Taupo Volcanic Zone (Given 1980; Burns 1997; Williams et al. 2007). This class is subdivided into well-drained terrestrial or wetland/aquatic systems (Appendices 2 & 3). Geothermal terrestrial includes heated and hydrothermally-altered soils (> 20°C) that are predominantly acidic, contain low amounts of organic matter and phosphorus, and may contain high concentrations of metals and trace elements, e.g. arsenic and titanium (Given 1980; Burns 1997). Geothermal wetlands and aquatic ecosystems include geyser mounds, hot springs, pools, streams, fumaroles and mud pools of a variety of temperature, pH and chemical compositions. Communities associated with wetland and aquatic geothermal ecosystems are dominated by extremophilic microorganisms, including thermophilic fungi, cyanobacteria, archaebacteria, algae and invertebrates, and occur where temperatures are less than 55°C. Microstromatolites occur around geyser mounds, with communities composed of bacteria, cyanobacteria, diatoms, tasmantids and amoeba, and algae (Boothroyd 2009).
3.2.5 Absence of light—subterranean caves and deep depressions in karst (Unit CV1)All caves have been assigned to a single subterranean ecosystem unit (see also Williams et al. 2007). In caves, the energy flow is primarily derived from external organic inputs via water and air flow, and is locally combined with the biotic component of lithotrophic microorganisms (organisms that use inorganic substrates for biosynthesis). Caves have near-constant temperatures, equably high humidities (> 95%), and include both aquatic and terrestrial components. Caves predominantly form within subterranean calcareous rocks (e.g. limestone and marble) in combination with subterranean water-flow that facilitates both chemical weathering and physical erosion, but can also form within basalt rock (Williams et al. 2007). The biotic component is dominated by invertebrate and microbial species and communities.
3.2.6 Combustion and/or volcanic activity (fire) (Units VS1–14)Although the classification primarily focuses on the steady-state or late successional composition of ecosystems, it also needs to accommodate the substantial area and wide environmental and compositional variability of fire-modified vegetation that occurs below the treeline. Therefore, the vegetation succession section of the classification principally applies to early to mid-successional or seral states of vegetation, rather than to advanced states. These secondary communities can be extensive in areas where old-growth woody communities have been most fire-prone, especially on low-fertility substrates where repeat fire has been the main method of land management. In most cases, our ecosystem labels incorporate just a small number of structural dominants of what

22 Singers & Rogers—Classification of terrestrial ecosystems
might be viewed as iconic species of seral communities, e.g. bracken (Pteridium esculentum), mānuka (Leptospermum scoparium), kānuka (Kunzea ericoides), and red (Chionochloa rubra) or copper (C. rubra subsp. cuprea) tussock. Several of the ecosystem labels use a small number of species; however, these are often labels of convenience that distil or represent a wide range of potential co-dominant species. These compositionally-simple labels also belie the successional variability and complexity of their subsequent compositional states. For instance, mānuka shrubland or scrub is a successional nurse or precursor to widely divergent communities across its full edaphic and climatic range—it can give way variously to beech forests, broadleaved forests and upland conifer-dominated forests.
We have not included any exotic plant-dominated seral ecosystems in the fire classification, even though they are also antecedents of many types of native woody vegetation, including forests (e.g. see Sullivan et al. 2007). Indeed, exotic plants are a substantial component of many seral communities, especially in rain shadow regions. The tall tussock community is effectively physiognomically equivalent to some alpine tall tussock grassland types (AL1–9), albeit in an entirely different climatic environment, with different herbaceous associates, and hosting different assemblages of invasive herbs, shrubs and trees. Repeat fire disturbance of seral communities can reset successions to the predictable physiognomic sequence of colonising non-vascular plants, herbs and grasses that eventually give way to woody species, including lianes. In other instances, the wind-dispersed seed of woody species such as mānuka, kānuka and Dracophyllum spp. permit their primary colonisation of fired-landscapes. Although our primary ecosystem driver emphasises anthropogenic fire rather than lightning-strike fire, the latter is also a source of less frequent and spatially-confined fires in New Zealand. It should also be noted that although all 14 ecosystems can have fire origins, there are specific instances where environmental stress and/or herbivory by domestic and feral ungulates can lead to similar communities; for example, the pest animal-induced tree-fernland or bush tussock grassland of montane southern Ruahine Range is a case in point (Rogers & Leathwick 1997). Consequently, the two universal influences on vegetation condition—environment and landscape history—need to be integrated before some seral states can be placed in this class.
We have included vulcanism alongside fire as a driver of seral vegetation. In renewing soils, volcanic disturbance tends to elicit similar seral vegetation states as local fire within regional contexts in both primary and secondary successions. Further, despite speculation to the contrary, there is no evidence that vulcanism caused long-term deforestation in New Zealand, except in very close proximity to active vents (McGlone 1989).

23Science for Conservation 325
4. Discussion
In the Introduction, we stated that the ideal ecosystem classification should be conceptually rigorous, be intuitive and comprehensible, be comprehensive and scale explicit, span seral and steady states, and be mappable. In the following sections, we discuss these challenges and how successful the present classification was in each of these areas in terms of its design and interpretation.
4.1 Conceptual rigourAlthough we commenced from a somewhat limited theoretical rationale, we have succeeded in establishing a classification based on a hierarchy of high-order physical and functional variables. At the structural and detailed design levels, our scheme has variably embraced many of the inter-related ecological concepts on which ecosystem classification and risk evaluation rely, such as continua, resilience, ecological integrity and succession. Furthermore, our description has established reference states and appropriate proxies of definable features that can be used to diagnose the loss of both ecosystem processes and structurally important or keystone biota. We have also cross-referenced our ecosystem units to existing community and habitat classifications to aid understanding of the inter-relationships between the two. One quite novel, generic concept we have adopted is to consider the extreme expression of some ecosystem processes to be the primary drivers in the hierarchy of abiotic ecosystem drivers. Prominent among these is disturbance, whether it be a function of gravity, water, wind or temperature extremes (freeze-thaw).
4.2 Intuitive and comprehensibleThere were many challenges in producing a comprehensible and accessible scheme that would be suitable for specialist and lay audiences alike. We discuss some of these below.
4.2.1 Aligning the physical environment with the bioticAlthough the macroclimatic zones depicted in Fig. 1 are an intuitive way of understanding the spatial interplay between temperature and moisture availability at a national scale, a large number of forest ecosystems span multiple temperature-moisture zones (Table 1), making it difficult to ascribe a representative or signature abiotic-biotic relationship or position in the classification for some ecosystems. This is particularly true for a few beech types—particularly red beech, silver beech (CLF10), silver beech (CLF11), and silver beech, mountain beech (CDF3)—and this problem is exacerbated by the difficulty in predictably modelling their climatic envelopes (Leathwick 1998).
Classification can be problematic where there is a spatially gradual transition in composition from one distinct community to another, which particularly occurs along elevational (temperature) or precipitation gradients in the forests of humid zones. Compounding this issue is perceived spatial unconformity between physical, functional and compositional turnover in the landscape. In some instances, perceived gradual or diffuse transition zones have been judged to constitute separate ecosystems in their own right that are intermediate between two widely divergent types. By contrast, many non-forest ecosystems, such as wetlands, cliffs and geothermal sites, have sharp, unequivocal discontinuities in composition that facilitate their classification and mapping.

24 Singers & Rogers—Classification of terrestrial ecosystems
4.2.2 Accommodating previous woody vegetation classificationsWhen Nicholls (1976) and McKelvey (1984) classified North and South Island forests, respectively, they placed a greater emphasis on species of commercial value, especially podocarps, rather than many commercially unimportant broadleaved species, as the data they used were collected primarily for the volumetric assessment of timber. Podocarp trees have much larger ranges (McGlone et al. 2010) and therefore are more tolerant of a wider range of climatic conditions than many broadleaved trees. Therefore, in modifying those previous forest classifications, we recognised the structural importance of broadleaved species alongside that of the podocarp element, and also acknowledged the valuable climatic and edaphic inferences that broadleaved species can provide. For example, kohekohe is restricted to the warm (predominantly frost-free) climatic zone, tawa provides a good biological boundary between the mild and cool climatic zones in the North Island and Marlborough as it is frost sensitive (Kelly 1987), and kāmahi and silver beech are indicative of the humid climatic zones as they are strongly limited by moisture stress (Wardle 1964, 1991). Further, although the classifications of Nicholls (1976) and McKelvey (1984) provided a strong basis for most mainland forest types, we supplemented them with more detail for North Island coastal forests, for the relict podocarp forests on alluvial soils and for forests of the larger offshore islands (e.g. the Kermadec and Chatham Islands).
Wiser et al. (2011) used data from 1177 systematically located vegetation plots to quantitatively describe 24 woody vegetation alliances, which we have cross-referenced in the present classification (Appendix 3). Many of their vegetation alliances had a notably high abundance of ungulate-avoided species, however (see Forsyth et al. 2002; Husheer et al. 2003, 2006), especially those that are characteristic of the subcanopy and lower tiers (e.g. horopito (Pseudowintera colorata), crown fern (Blechnum discolor), Myrsine divaricata, Leucopogon fasiculatus, Coprosma pseudocuneata and kātote (Cyathea smithii and C. dealbata)). By contrast, these species feature little in the present classification, where emphasis is instead placed on canopy dominants. Similarly, possum (Trichosurus vulpecula)-preferred northern rātā (Metrosideros robusta) and, in some communities, Hall’s tōtara were unimportant in the Wiser et al. (2011) alliances as they have greatly declined in recent decades, but are included as historically important forest components influencing the present classification.
4.2.3 Dealing with human-modified vegetation statesNew Zealand’s extremes of climate, lithology and catastrophic disturbance provide abundant opportunities for the formation of azonal ecosystems that are mostly spatially restricted and often show strong regional distinctions. Despite these extreme environments, azonal systems are/were not consistently non-forest or even non-woody, and several azonal types are known to have supported at least short forest communities in prehuman times. For instance, many types of wetlands originally supported short (and sometimes tall) forest, but that component was lost to fire in the post-settlement period (McGlone 2009); and consequently, our wetland section includes just one short forest community (a Chatham Island example). However, whilst there are no opportunities to accommodate kahikatea-forested swamps or silver pine (Manoao colensoi)-forested fens within the wetland section, they are provided for in the zonal section. Further, whilst the majority of the bog pine, mountain celery pine community of cold air inversion basins and valleys of the eastern South Island exists as fire-modified scrub, the rare, non-fired forest examples are included in our scrub/forest structural term. Finally, because the cliffs, braided rivers and dunes divisions are all characteristically geomorphically disturbed, their vegetation terms reflect the dominantly open or short stature vegetation that seldom progresses beyond shrubland.
The placement of today’s secondary derivatives of dryland woody communities in the classification was problematic. Our zonal section does not recognise humidity-limitation (both atmospheric and edaphic) as the primary ecosystem driver for today’s open vegetation of dryland environments for two reasons. Firstly, these environments can partly be accommodated

25Science for Conservation 325
in the cold air inversion section, which sometimes environmentally overlaps with the driest environments. Secondly, because fire has comprehensively modified this dry zone, its pervasive secondary shrubland and grassland communities can be accommodated within the frequent fire (secondary vegetation) section. Moreover, in the heavily fire-modified landscape that currently exists, it would be very difficult to establish a threshold level of moisture-stress within the semi-arid zone that was causally correlated with a transition from dry forest to short treeland or scrub; and some modification of rainshadow weather patterns has likely accompanied the extensive deforestation of this zone (e.g. see Betts et al. 2007; Sampaio et al. 2007).
4.2.4 Challenges of the alpine zoneThe depiction of the low and high alpine zones in Fig. 1 is at best a theoretical model of their spatial arrangement based on temperature lapse rate and a generalised rainfall gradient in an environment that has few data sources from weather stations. Wind exposure in relation to terrain confounds altitudinal lapse rate predictions of temperature, with high-alpine temperature environments sometimes prominent within the low-alpine altitudinal zone; for instance, high-alpine AH2 Dracophyllum muscoides cushionfield occurs in the low-alpine altitudinal zone on Otago schist mountains due to terrain-related exposure and edaphic influences. Further, the low-altitude limit of low alpine may not accord well with the treeline in areas where there is a gradual physiognomic vegetation transition from forest to scrub and grassland, especially in non-beech regions. Also, in the North Island, some treelines deviate from a progressive latitudinally-correlated decline southwards (Körner & Paulsen 2004). Finally, the scale of Fig. 1 is not conducive to reliably showing the small alpine island pattern that is evident in most upland zones, especially in the North Island, west Nelson and Fiordland.
4.2.5 Inferences about ecosystem relationshipsSince the present classification is mostly qualitative, it offers no valid opportunity to judge the quantitative similarity or difference between ecosystems. However, inferences on the hierarchical relationship between ecosystems are possible via the classification’s abiotic framework, especially for zonal types—in the azonal section, that opportunity is compromised because there are wide functional differences between the ecosystem processes that underpin the 11 broad divisions.
4.3 Comprehensiveness and scale explicitnessAs this classification is applied and tested, there will inevitably be questions over its comprehensiveness or coverage of a ‘full range’ of New Zealand’s terrestrial ecosystems (as referred to in goal 3 of the New Zealand Biodiversity Strategy; DOC & MfE 2000). An ecosystem’s extent is a function of the operational scale of the vital processes that sustain it and the scale of human perception (Park 2000: 81), meaning that the classification of an ecosystem is essentially a conceptual exercise (Johnson & Gerbeaux 2004) and that ecosystem units are an arbitrary construct at any particular scale. For this classification, our choice of scale was strongly influenced by those used in pre-existing classifications, which we acknowledge as being biased towards a plant community view rather than a process or functional view. Consequently, the full range of ecosystems reflects the levels used previously for different biomes, and our capacity to modify those and adequately fill any perceived gaps at an arbitrarily chosen scale.
In the last 750 years, the clearance of forest has been so comprehensive across parts of the semi-arid and sub-humid zones of New Zealand that it has been necessary to draw upon historical accounts, ecological modelling and descriptions of seral remnants (e.g. McGlone & Neall 1994; Leathwick 2001; Rogers et al. 2005; Hall & McGlone 2006) to supplement insights from the sparse distribution of relict stands when classifying their forest ecosystems. Further,

26 Singers & Rogers—Classification of terrestrial ecosystems
it is thought that most relict stands in these areas have been modified by fire, so that originally subordinate or narrow-niche elements now dominate. Thus, the inevitable imprecision of classifying this zone has led to just nine warm to cool semi-arid forest ecosystems being defined, which are applicable to approximately 19% of New Zealand’s total land area. By contrast, this zone contains 13 of 20 LENZ Level 1 and 181 of 500 LENZ Level IV environments of Leathwick et al. (2003b). There may be no broadly representative extant examples of some pre-settlement semi-arid forest ecosystems; for instance, there may have been a podocarp-broadleaved forest type of semi-arid lowland hill country in Hawke’s Bay and Marlborough that was devoid of drought-sensitive tawa, but this idea is not supported by relicts and modelling. There are much greater areas of extant forest in the warm to cool humid and sub-humid zones, as reflected by the 42 ecosystems that have been identified in these regions. Inevitably, humid and sub-humid forests have also received the bulk of forest research investigation and classification (e.g. see McKelvey & Nicholls 1957; Nicholls 1976; McKelvey 1984).
In this classification, some fine-scale community mosaics were arbitrarily homogenised or aggregated into one ecosystem that represented the spatially-dominant matrix ecosystem. An example of this is valley and basin floors that predominantly support frost-determined scrub or grassland, but are interspersed with poor-drainage-determined wetlands or fire-induced seral derivatives. Similarly, alpine landscapes can have rich compositional variation at fine spatial scales, reflecting variable exposure, snow lie and edaphic influences. Also, the predominantly raw soils of alpine environments can be lithologically quite diverse, leading to high community distinctiveness at a regional scale. By contrast, the more deeply weathered soils of forested environments are less edaphically influential. Fine-scale community mosaics and regional distinctiveness have probably influenced the quite generalised ecosystem classifications that have previously been used for alpine environments, such as those of Wardle (1991) and Mark & Dickinson (1997). For similar reasons of convenience, we have included a comparatively small number of shrubland, tussock grassland and cushionfield ecosystems in this classification, along with a few bare ground-based units, such as gravelfield, stonefield and fellfield. Likewise, the fine-scale community patterns of cliffs and braided rivers are conveniently represented by broad ecosystem types. However, wetlands, especially ephemeral wetlands, have been much studied, characterised and classified (e.g. Dobson 1979; Johnson & Rogers 2002; Johnson & Gerbeaux 2004), and so represent 15% of all ecosystems, reflecting their high physiographic and compositional variability. Thus, overall, the wide differences in the numbers of ecosystems within and between the abiotic sections of the classification reflect the combined influence of genuine geographic variability, the different amounts of scientific study across biome or bioclimatic zones, and scale-related classification convenience.
This classification may also have shortcomings for multi-scale conservation planning, which incorporates more detailed, finer scale levels than this classification is pitched at. For instance, it would be unsuitable for distinguishing red beech (Fuscospora fusca) forest on colluvial toeslopes from red beech forest on colluvial hillslopes. Rather, the classification might be most useful for land use planning at national and regional scales, and the ecological unit concept of Myers et al. (1987) may help to discriminate subtle landform-driven variations in vegetation composition and ecosystem function at a finer scale.
4.4 Relationships between seral and steady statesFor several reasons, our description of community composition concentrates on steady states and, to a lesser extent, potential states, whilst also recognising that disturbance (both natural and anthropogenic) produces compositional variation, which is often labelled as a state of dynamic equilibrium. Therefore, our forest unit descriptions often incorporate the compositional variability that arises from catastrophic or local disturbances of a climatic and tectonic nature. There are three reasons for this approach. First, it provides insights into ecosystem dynamics

27Science for Conservation 325
within particular environmental disturbance regimes. Second, it encourages interpretation and improved understanding of the successional trajectories and relationships of secondary communities with their developmental steady states, particularly where the two are spatially intermixed. Third, it provides conservation managers with compositional goals for ecosystem restoration.
For landscapes in which secondary communities are extensive and/or only changing slowly, or where steady states have been comprehensively lost, the secondary vegetation section of the classification is best applied. Here, predictive modelling of community potentiality based on species-environment relationships (e.g. see Leathwick 2001) can offer at least coarse-scale insights into future community composition, assuming that propagule sources for its late successional potential are present. However, it must also be accepted that in today’s pyrophylic dryland environments that contain a pernicious and persistent exotic flora, the testing of model predictions of fire-lost states may be a distant proposition. Similarly, where herbivory by introduced animals has greatly transformed structural and compositional states leading to unclear potential trajectories, it will be difficult to determine the potential of the extant secondary communities.
In some areas, seral vegetation resulting from fire or land clearance masks the previous community boundaries that reflected discontinuities in ecosystem processes, making it difficult to recognise azonal ecosystems that are nested within larger surrounding zonal systems. For example, vegetation homogenisation or the loss of landscape processes can inhibit recognition of the limits of active versus stable dunes (the latter potentially supported tall forest), the margins of wetlands, frost flats and saline patches, and long-stabilised river braids.
4.5 Spatial explicitness and mappability Spatially explicit ecosystem classifications are required for many conservation planning tasks. For instance, biodiversity offsetting requires classifications to explore spatial opportunities for offsetting or, conversely, avoiding land clearance, by enabling the accurate assessment of irreplaceability and preventing inappropriate exchanges of dissimilar biodiversity. Spatially explicit classifications may also aid the measurement of vulnerability (as a measure of how much has been lost and the degree of threat), by comparing loss in the form of clearance and the extent of secondary or derivative communities with the remaining cover of the original or steady-state community. While some national-scale ecosystem and vegetation classifications are likely to be too coarse for many offsetting applications, the present classification is potentially useful.
Due to its subjectivity, the present classification is not spatially explicit in terms of the quantitative discrimination of ecosystems. However, it is possible to map some types using a combination of remote sensing, aerial photographs, existing maps, digital climatic databases and expert knowledge. Digitisation of the New Zealand Forest Service Map Series 6, which describes 18 forest classes that form the basis of parts of the zonal section, is a useful foundation for mapping. In addition, useful mapping data also exist for some broad groups of azonal ecosystems, e.g. geothermal (Beadel 2004) and ultramafic (Lee 1992) ecosystems. Vegetation community maps also exist for some districts or discrete conservation reserves that could be adapted for this use, e.g. the Chatham Islands (Kelly 1983), Tongariro National Park (Atkinson 1981), and those provided by the four publications in the National Parks Scientific Series (Wilson 1976; Mark 1977; Wardle 1979; Clarkson 1986). Furthermore, the Department of Conservation is undertaking digital mapping of many of the historically rare ecosystems of Williams et al. (2007), predominantly by combining digital environmental data, satellite imagery and expert knowledge.

28 Singers & Rogers—Classification of terrestrial ecosystems
5. Conclusions
Classifying ecosystems is a priority both in New Zealand and globally as the science and application of spatial conservation prioritisation and ecosystem threat ranking gathers pace (Joesph et al. 2008; Moilenan et al. 2009; Holdaway et al. 2012; Keith et al. 2013). There is a need for such classifications to use consistent quantitative methodologies that are grounded in theoretical concepts of biodiversity organisation, because the valuation of ecosystems must incorporate quantitative estimates of ecosystem collapse (Keith et al. 2013). Foremost among the difficulties of ensuring classification consistency is the uncertainty about ecosystem boundaries, or the divergence between physical, functional and compositional turnover in the landscape.
In New Zealand, ecosystem taxonomy is in its infancy. Presently, there is a lack of integration of physical variables (e.g. LENZ), functional/process variables (e.g. flooding and sedimentation) and biotic composition. Most of New Zealand’s terrestrial vegetation is covered by existing classifications; however, most of these are conceptually community and habitat schemes. The PNAP was the strongest scheme in terms of integrating at least physical variables and biotic composition, as this was based on an ecological unit concept—essentially an ecosystem label linking landforms, soils and vegetation within an ecological district geographic setting (Kelly & Park 1986).
In this study, we have tried to make progress in designing a genuine ecosystem classification for terrestrial New Zealand by engaging with all three theoretical components. Our solution to incorporating both physical and functional drivers as abiotic components with minimal bias was two-fold. First, we used the principle drivers of temperature, moisture and lithology as physical variables to construct a zonal or macroclimate- and soil-driven section of the classification. Second, we used the extremes of other variables such as disturbance, soil chemistry, and water tables that disrupt or curtail the full macroclimatic and edaphic potential of an environment as our functional or process drivers. Accordingly, these were afforded primacy ahead of the physical variables to form the azonal section of the classification. Elsewhere, qualitative approaches have used an understanding of the multiscale regulation of ecosystems across climate, landforms, lithology and vegetation to produce nested hierarchical classifications of land (e.g. the Netherlands (Klijn & Udo de Haes 1994), Canada (Pojar et al. 1987) and Italy (Blasi et al. 2000)). These are conceptually similar approaches to ours but differ methodologically.
In testing our classification, there will be tension between the need for specificity for district and regional planning, and the need for generality to support national-scale application in an archipelago of steep environmental gradients. The application and testing of the present scheme is proceeding at pace within both national and local government agencies, with Auckland, Waikato, Bay of Plenty and Wellington regional authorities using it to gain an improved understanding and awareness of local ecosystem patterns. Indeed, the integrated application of the abiotic and biotic components has facilitated understanding of the relationships between ecosystem patterns and processes across modified and more natural indigenous vegetation. Mapping has also facilitated an understanding of historic and current ecosystem representation, enabling the identification of priority areas for protection and management of both semi-natural and natural ecosystems.
The most challenging environment to classify was rain shadow New Zealand, where the modification and loss of vegetation greatly compromises our understanding of ecosystem patterns and processes. Topographic and edaphic influences on temperature and moisture availability in dryland environments would have produced high ecosystem heterogeneity at fine spatial scales that were largely masked or only hinted at by today’s relatively homogeneous seral and exotic vegetation. Therefore, it is there, as well as in the alpine and seral sections of the classification with their pragmatic generic treatments, that testing is likely to produce some welcome refinement.

29Science for Conservation 325
6. Acknowledgements
First, we warmly thank John Leathwick for preparing Figure 1, for helpful discussion and for refereeing the manuscript. As another referee, Mark Smale also provided insightful comments that led to many improvements, as did Amanda Todd with editorial input. Further, many provincial and biome experts generously contributed their knowledge of community type and pattern in the early stages of this study, but they are too numerous to acknowledge individually; and, with an eye to history, we especially recognise the contribution provided by the publications of the field ecologists of the now disbanded New Zealand Forest Research Institute and the Botany Division of the Department of Scientific and Industrial Research (DSIR). Additionally, in recognition of his conceptual thought on the topic of New Zealand ecosystems, we especially acknowledge the late Geoff Park and his narrative ‘New Zealand as ecosystems’. Geoff also established the Biological Resources Centre of the DSIR, the agency that pioneered a systematic framework upon which geographic data could be assembled. This was portrayed in the survey manual of their Protected Natural Areas Programme, which introduced to New Zealand the ecological unit concept that united the abiotic and biotic components of life in geographic description.
7. ReferencesAllen, R.B.; MacIntosh, P.D.; Wilson, J.B. 1997: The distribution of plants in relation to pH and salinity on inland saline-
alkaline soils of Central Otago. New Zealand Journal of Botany 35: 517–523.
Araujo, M.B.; Humphries, C.J.; Densham, P.J.; Lampinen, R.; Hagemeijer, W.J.M.; Mitchell-Jones, A.J.; Gasc, J.P. 2001: Would environmental diversity be a good surrogate for species diversity? Ecography 24: 103–110.
Atkinson, I.A.E. 1981: Vegetation map of Tongariro National Park, North Island, New Zealand. Scale 1:50 000. New Zealand Department of Scientific and Industrial Research, Wellington. 27 p.
Atkinson, I.A.E. 1985: Derivation of vegetation mapping units for an ecological survey of Tongariro National Park, North Island, New Zealand. New Zealand Journal of Botany 23: 361–378.
Beadel, S. 2004: Geothermal vegetation of the Waikato Region—revised edition. Unpublished contract report by Wildland Consultants Ltd for Environment Waikato.
Belbin, L. 1993: Environmental representativeness: regional partitioning and reserve selection. Biological Conservation 66: 223–230.
Bellingham, P.J.; Towns, D.R.; Cameron, E.K.; Davis, J.J.; Wardle, D.A.; Wilmshurst, J.A.; Mulder, C.P.H. 2010: New Zealand island restoration: seabirds, predators, and the importance of history. New Zealand Journal of Ecology 34: 115–136.
Betts, R.A.; Falloon, P.D.; Goldewijk, K.K.; Ramankutty, N. 2007: Biogeophysical effects of land use on climate: model simulations of radiative forcing and large-scale temperature change. Agricultural and Forest Meteorology 142: 216–233.
Bishop, C.D. 2005: The nature and stability of frost flat heathland/forest ecotones in central North Island, New Zealand. Unpublished PhD thesis, University of Auckland, Auckland. 235 p.
Blasi, C.; Carranza, M.L.; Frondoni, R.; Rosati, L. 2000: Ecosystem classification and mapping: a proposal for Italian landscapes. Applied Vegetation Science 3: 233–242.
Boothroyd, I.K.G. 2009: Ecological characteristics and management of geothermal systems of the Taupo Volcanic Zone, New Zealand. Geothermics 38: 200–209.
Brockerhoff, E.G.; Shaw, W.B.; Hock, B.; Kimberley, M.; Paul, T.; Quinn, S.; Pawson, S. 2008: Re-examination of recent loss of indigenous cover in New Zealand and the relative contributions of different land uses. New Zealand Journal of Ecology 32: 115–126.

30 Singers & Rogers—Classification of terrestrial ecosystems
Brumley, C.F.; Stirling, M.W.; Manning, M.S. 1986: Old Man Ecological District. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme, Survey Report No. 3. Department of Conservation, Wellington. 174 p.
Burns, B.R. 1997: Vegetation change along a geothermal stress gradient at the Te Kopia steamfield. Journal of the Royal Society of New Zealand 27: 279–294.
Chadderton, W.L.; Brown, D.J.; Stephens, R.T. 2004: Identifying freshwater ecosystems of national importance for biodiversity: criteria, methods, and candidate list of nationally important rivers. Discussion document. Department of Conservation, Wellington. 112 p.
Cieraad, E.; McGlone, M.; Barbour, M.M.; Huntley, B. 2012: Seasonal frost tolerance of trees in the New Zealand treeline ecotone. Arctic, Antarctic, and Alpine Research 44: 332–342.
Clarkson, B.D. 1986: Vegetation of Egmont National Park, New Zealand. National Parks Scientific Series Number 5. Department of Scientific and Industrial Research, Wellington. 98 p.
Cockayne, L. 1909: Report on a botanical survey of Stewart Island, New Zealand. Department of Lands, Government Printer, Wellington. 68 p.
Cockayne, L. 1911: Report on the dune areas of New Zealand, their geology, botany and reclamation. Department of Lands, Government Printer, Wellington. 74 p.
Conning, L. 2001: Northland Protection Strategy. Nature Heritage Fund, Wellington. 134 p.
Connor, H.E. 1991: Chionochloa Zotov (Gramineae) in New Zealand. New Zealand Journal of Botany 29: 219–282.
Corner, P.; Faber-Langendoen, D.; Evans, R.; Gawler, S.; Josse, C.; Kittel, G.; Menard S.; Pyne, M.; Reid, M.; Schulz, K.; Snow, K.; Teague, J. 2003: Ecological systems of the United States. A working classification of U.S. terrestrial systems. NatureServe, Arlington, VA.
Coulter, J.D. 1966: Dry spells in New Zealand as a factor in plant ecology. Proceedings of the New Zealand Ecological Society 13: 4–8.
Daubenmire, R. 1954: Alpine timberlines in the Americas and their interpretation. Butler University Botanical Studies 11: 119–136.
Daubenmire, R.F. 1974: Plants and environment: a textbook of plant autecology. 3rd edition. John Wiley & Sons, New York. 422 p.
de Lange, P.J.; Cameron, E.K.; Taylor, G.A. 1995: Flora and fauna of Tatapihi (Groper) Island, Mokohinau Islands. Tane 35: 69–94.
Dickinson, K.J.M. 1988: Umbrella Ecological District. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme, Survey Report No. 7. Department of Conservation, Wellington. 179 p.
Dickinson, K.J.M. 1989: Nokomai Ecological District. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme, Survey Report No. 9. Department of Conservation, Wellington. 139 p.
Dobson, A.T. 1979: Mire types in New Zealand. Pp. 89−98 in: Proceedings of the International Symposium on Classification of Peat and Peatlands, Hyytlaia, Finland. International Peat Society, University of Helsinki, Helsinki.
DOC (Department of Conservation); MfE (Ministry for the Environment) 2000: The New Zealand Biodiversity Strategy. A draft strategy for public consultation. Department of Conservation and the Ministry for the Environment, Wellington. 146 p.
Elder, N.L. 1962: Vegetation of the Kaimanawa Ranges. Transactions of the Royal Society of New Zealand, Botany 2: 1–37.
Esler, A.E. 1969: Manawatu sand plain vegetation. Proceedings of the Ecological Society of New Zealand 16: 32–35.
Esler, A.E. 1970: Manawatu sand dune vegetation. Proceedings of the Ecological Society of New Zealand 17: 41–46.
Fagan, B.; Pillai, D. 1992: Manorburn Ecological District. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 22. Department of Conservation, Wellington. 117 p.
Faith, D.P.; Walker, P.A.; Margules, C.R. 2001: Some future prospects for systematic biodiversity planning in Papua New Guinea—and for biodiversity planning in general. Pacific Conservation Biology 6: 325–342.
Ferrier, S.; Faith, D.P.; Arponen, A.; Drielsma, M. 2009: Community level approaches to spatial conservation prioritisation. Pp. 94–109 in Moilanen, A.; Wilson, K.A.; Possingham, H.P. (Eds): Spatial conservation prioritisation: quantitative methods and computational tools. Oxford University Press, New York.

31Science for Conservation 325
Ferrier, S.; Wintle, B.A. 2009: Quantitative approaches to spatial conservation prioritisation: matching the solution to the need. Pp. 1–15 in Moilanen, A.; Wilson, K.A.; Possingham, H.P. (Eds): Spatial conservation prioritisation: quantitative methods and computational tools. Oxford University Press, New York.
Forsyth, D.M.; Coomes, D.A.; Nugent, G.; Hall, G.M.J. 2002: The diet and diet preferences of ungulates (Order: Artiodactyla) in New Zealand. New Zealand Journal of Zoology 29: 323–343.
Fukami, T.; Wardle, D.A.; Bellingham, P.J.; Mulder, C.P.H.; Towns, D.R.; Yeates G.W.; Bonner, K.I.; Durrett, M.S.; Grant-Hoffman, M.N.; Williamson, W.M. 2006: Above- and below-ground impacts of introduced predators in seabird-dominated island ecosystems. Ecology Letters 9: 1299–1307.
Gaston, K.J. 1990: Patterns in the geographical ranges of species. Biological Reviews 65: 105–129.
Gauch, H.G.; Whittaker, R.H. 1981: Hierarchical classification of community data. Journal of Ecology 69: 537–557.
Gillham, M.E. 1960a: Destruction of indigenous heath vegetation in victorian sea-bird colonies. Australian Journal of Botany 8: 277–317.
Gillham, M.E. 1960b: Vegetation of New Zealand shag colonies. Transactions of the Royal Society of New Zealand 88: 363–380.
Given, D.R. 1980: Vegetation on heated soils at Karapiti, central North Island, New Zealand, and its relation to ground temperature. New Zealand Journal of Botany 18: 1–13.
Hall, G.M.J.; McGlone, M.S. 2006: Potential forest cover of New Zealand as determined by an ecosystem process model. New Zealand Journal of Botany 44: 211–232.
Heenan, P.B.; Smissen, R.D. 2013: Revised circumscription of Nothofagus and recognition of the segregate genera Fuscospora, Lophozonia, and Trisyngyne (Nothofagaceae). Phytotaxa 146: 1–13.
Hesp, P.A. 2000: Coastal sand dunes: form and function. Coastal Dune Vegetation Network Technical Bulletin No. 4. Forest Research, Rotorua. 29 p.
Holdaway, R.J.; Wiser, S.K.; Williams, P.A. 2012: Status assessment of New Zealand’s naturally uncommon ecosystems. Conservation Biology 26: 619–629.
Husheer, S.W.; Allan, R.B.; Robertson, A.W. 2006: Suppression of regeneration in New Zealand mountain beech forests is dependent on species of introduced deer. Biological Invasions 8: 823–834.
Husheer, S.W.; Coomes, D.A.; Robertson, A.W. 2003: Long-term influences of introduced deer on the composition and structure of New Zealand Nothofagus forests. Forest Ecology and Management 181: 99–117.
Johnson, P.N. 1993: Dry coastal ecosystems of New Zealand. Pp. 197–221 in van der Maarel, E. (Ed.): Ecosystems of the world 2B. Dry coastal ecosystems. Elsevier, Amsterdam.
Johnson, P.N.; Gerbeaux, P. 2004: Wetland types in New Zealand. Department of Conservation, Wellington. 184 p.
Johnson, P.N.; Rogers, G.M. 2002: Ephemeral wetlands and their turfs in New Zealand. Science for Conservation 230. Department of Conservation, Wellington. 109 p.
Johnson, P.N.; Rogers, G.M. 2006: Dune wetlands and their vegetation dynamics. Unpublished Landcare Research Contract Report LC0405/157. Landcare Research, Lincoln. 88 p.
Joseph, L.N.; Maloney, R.F.; Possingham, H.P. 2008: Optimal allocation of resources among threatened species: a project prioritization protocol. Conservation Biology 23: 328–338.
Keith, D.A.; Rodríguez, J.P.; Rodríguez-Clark, K.M.; Nicholson, E.; Aapala, K.; Alonso, A. et al. 2013: Scientific foundations for an IUCN red list of ecosystems. PLoS ONE 8(5): e62111. doi: 10.1371/journal.pone0062111.
Kelly, D. 1987: Slow recovery of Beilschmiedia tawa after severe frosts in inland Taranaki, New Zealand. New Zealand Journal of Ecology 10: 137–140.
Kelly, G.C. 1983: Distribution and ranking of remaining areas of indigenous vegetation in the Chatham Islands (a map and accompanying notes). Botany Division, Department of Scientific and Industrial Research, Lower Hutt. 88 p.
Kelly, G.C.; Park, G.N. 1986: The New Zealand Protected Natural Areas Programme. A scientific focus. Biological Resources Centre, Department of Scientific and Industrial Research, Lower Hutt. 68 p.
Kennedy, D.M.; Dickson, M.E. 2007: Cliffed coasts of New Zealand: perspectives and future directions. Journal of the Royal Society of New Zealand 37: 41–57.
Klijn, F.; Udo de Haes, H.A. 1994: A hierarchical approach to ecosystems and its implications for ecological land classification. Landscape Ecology 9: 89–104.

32 Singers & Rogers—Classification of terrestrial ecosystems
Körner, C.; Paulsen, J. 2004: A world-wide study of high altitude treeline temperatures. Journal of Biogeography 31: 713–732.
Leathwick, J.R. 1998: Are New Zealand’s Nothofagus species in equilibrium with their environment? Journal of Vegetation Science 9: 719–732.
Leathwick, J.R. 2001: New Zealand’s potential forest pattern as predicted from current species-environment relationships. New Zealand Journal of Botany 39: 447–464.
Leathwick, J.R.; Austin, M.P. 2001: Competitive interactions between tree species in New Zealand’s old-growth indigenous forests. Ecology 82: 2560–2573.
Leathwick, J.R.; Collier, K.; Chadderton, L. 2007: Identifying freshwater ecosystems with nationally important natural heritage values: development of a biogeographic framework. Science for Conservation 274. Department of Conservation, Wellington. 30 p.
Leathwick, J.R.; Overton, J.M.; McLeod, M. 2003a: An environmental domain classification of New Zealand and its use as a tool for biodiversity management. Conservation Biology 17: 1612–1623.
Leathwick, J.R.; Whitehead, D. 2001: Soil and atmospheric water deficits and the distribution of New Zealand’s indigenous tree species. Functional Ecology 15: 233–242.
Leathwick, J.R.; Wilson, G.; Rutledge, D.; Wardle, P.; Morgan, F.; Johnston, K.; McLeod, M.; Kirkpatrick, R. 2003b: Land Environments of New Zealand. David Bateman Ltd, Auckland. 183 p.
Lee, W.G. 1992: New Zealand ultramafics. Pp. 375−418 in Roberts, B.A.; Proctor, J. (Eds): The ecology of areas with serpentinized rocks. A world view. Kluwer Academic Publishers, Netherlands.
Logan, M.C.; Holloway, J.E. 1933: Plant succession on the Oreti River sand dunes. Transactions of the New Zealand Institute 61: 122–139.
Mark, A.F. 1977: Vegetation of Mount Aspiring National Park. National Parks Scientific Series Number 2. Department of Lands and Survey, Wellington. 79 p.
Mark, A.F. 2013: Above the treeline: a nature guide to the New Zealand mountains. Craig Potton Publishing, Nelson. 472 p.
Mark, A.F.; Dickinson, K.J.M. 1997: New Zealand alpine ecosystems. Pp. 311–345 in Wielgolaski, F.E. (Ed.): Ecosystems of the world, volume 3: polar and alpine tundra. Elsevier, Amsterdam.
McGlone, M.S. 1989: The Polynesian settlement of New Zealand in relation to environmental and biotic changes. New Zealand Journal of Ecology 12 (Supplement): 115–129.
McGlone, M.S. 2009: Postglacial history of New Zealand wetlands and implications for their conservation. New Zealand Journal of Ecology 33: 1–23.
McGlone, M.S.; Moar, N.T. 1998: Dryland Holocene vegetation history, Central Otago and the Mackenzie Basin, South Island, New Zealand. New Zealand Journal of Botany 36: 91–111.
McGlone, M.S.; Neall, V.E. 1994: The late Pleistocene and Holocene vegetation history of Taranaki, North Island, New Zealand. New Zealand Journal of Botany 32: 251–269.
McGlone, M.S.; Richardson, S.J.; Jordan, G.J. 2010: Trees in New Zealand: heights, species richness and range size. New Zealand Journal of Ecology 34: 137–151.
McKelvey, P.J. 1984: Provisional classification of South Island virgin indigenous forests. New Zealand Journal of Forestry Science 14: 151–178.
McKelvey, P.J.; Nicholls, J.L. 1957: A provisional classification of North Island forests. New Zealand Journal of Forestry 7: 84–101.
McSaveney, M.J.; Whitehouse, I.E. 1989: Anthropic erosion of mountain land in Canterbury. New Zealand Journal of Ecology 12 (Supplement): 151–163.
Meurk, C. 1982: Alpine phytoecology of the rainshadow mountains of Otago and Southland, New Zealand. Unpublished PhD thesis, University of Otago, Dunedin. 774 p.
Meurk, C. 1984: Bioclimatic zones for the Antipodes—and beyond? New Zealand Journal of Ecology 7: 175–181.
Meurk, C. 1995: Evergreen broadleaved forests of New Zealand and their bioclimatic definition. Pp. 151–197 in Box, E.O.; Peet, R.K.; Masuzawa, T.; Yamada, I.; Fujiwara, K.; Maycock, P.F. (Eds): Vegetation science in forestry. Kluwer Academic Publishers, The Hague, Netherlands.

33Science for Conservation 325
Meurk, C.; Foggo, M.N.; Bastow-Wilson, J. 1994: The vegetation of subantarctic Campbell Island. New Zealand Journal of Ecology 18: 123–168.
Moilanen, A.; Wilson, K.A.; Possingham, H.P. 2009: Spatial conservation prioritisation: quantitative methods and computational tools. Oxford University Press, New York. 304 p.
Molloy, L.R. 1998: Soils of the New Zealand landscape: the living mantle. New Zealand Society of Soil Science, New Zealand Soil Bureau and Mallinson Rendel Publishers, Wellington. 239 p.
Myers, S.C.; Park, G.N.; Overmars, F.B. (comps) 1987: The New Zealand Protected Natural Areas Programme—a guidebook for the rapid ecological survey of natural areas. New Zealand Biological Resources Centre Publication 5. Department of Conservation, Wellington. 113 p.
Newsome, P.F.J. 1987: The vegetation cover of New Zealand. Water & Soil Miscellaneous Publication No. 112. Ministry of Works and Development, Wellington. 153 p.
New Zealand Soil Bureau 1968: Soils of New Zealand. Part 1. New Zealand Soil Bureau Bulletin 26(1). Soil Bureau, Department of Scientific and Industrial Research, Wellington. 142 p.
Nicholls, J.L. 1976: A revised classification of the North Island indigenous forests. New Zealand Journal of Forestry 28: 105–132.
Nicholson, E.; Keith, D.A.; Wilcove, D.S. 2009: Assessing the threat status of ecological communities. Conservation Biology 23: 259–274.
Oliver, R.B. 1910: The vegetation of the Kermadec Islands. Transactions and Proceedings of the New Zealand Institute 42: 118–175.
Park, G.N. 2000: New Zealand as ecosystems: the ecosystem concept as a tool for environmental management and conservation. Department of Conservation, Wellington. 98 p.
Partridge, T.R.; Wilson, J.B. 1987: Salt tolerance of salt marsh plants of Otago, New Zealand. New Zealand Journal of Botany 25: 559–566.
Partridge, T.R.; Wilson, J.B. 1988: Vegetation patterns in salt marshes of Otago, New Zealand. New Zealand Journal of Botany 26: 497–510.
Peinado, M.; Aguirre, J.L.; Delgadillo, J.; Macias, M.A. 2007: Zonobiomes, zonoecotones and azonal vegetation along the Pacific coast of North America. Plant Ecology 191: 221–252. doi 10.1007/s11258-006-9239-8.
Pojar, J.; Klinka, K.; Meidinger, D.V. 1987: Biogeoclimatic ecosystem classification in British Columbia. Forest Ecology and Management 22: 119–154.
Pressey, R.L.; Hagar, T.C.; Ryan, K.M.; Schwarz, J.; Wall, S.; Ferrier, S. 2000: Using abiotic data for conservation assessments over extensive regions: quantitative methods applied across New South Wales, Australia. Biological Conservation 96: 55–82.
Rogers, G.M. 1999: Coastal turfs of mainland New Zealand: their composition, environmental character, and conservation needs. Science for Conservation 107. Department of Conservation, Wellington. 83 p.
Rogers, G.M.; Hewitt, A.; Wilson, J.B. 2000: Ecosystem-based conservation strategy for Central Otago’s saline patches. Science for Conservation 166. Department of Conservation, Wellington. 38 p.
Rogers, G.M.; Leathwick, J.R. 1997: Factors predisposing forests to collapse in the southern Ruahine Range, New Zealand. Biological Conservation 80: 325–338.
Rogers, G.M.; Walker, S.; Lee, W.G. 2005: The role of disturbance in dryland New Zealand: past and present. Science for Conservation 258. Department of Conservation. Wellington. 122 p.
Rogers, G.M.; Wiser, S.K. 2010: Environment, composition, and conservation of coastal turfs in mainland New Zealand. New Zealand Journal of Botany 48: 1–14.
Sakai, A.; Wardle, P. 1978: Freezing resistance of New Zealand trees and shrubs. New Zealand Journal of Ecology 1: 51–61.
Sampaio, G.; Noble, C.; Satyamurty, P.; Soares-Filho, B.S.; Cardosa, M. 2007: Regional climate change over eastern Amazonia caused by pasture and soybean cropland expansion. Geophysical Research Letters 34: L17709. doi:10.1029/2007GL030612.
Smale, M.C. 1990: Ecology of Dracopyllum subulatum heathland on frost flats at Rangitaiki and north Pureora, central North Island, New Zealand. New Zealand Journal of Botany 28: 225–248.
Smale, M.C.; Fitzgerald, N.B.; Richardson, S.J. 2011: Resilience to fire of Dracophyllum subulatum (Ericaceae) frost flat heathland, a rare ecosystem in central North Island, New Zealand. New Zealand Journal of Botany 49: 231–242.

34 Singers & Rogers—Classification of terrestrial ecosystems
Sullivan, J.J.; Williams, P.A.; Timmins, S.M. 2007: Secondary forest succession differs through naturalised gorse and native kānuka near Wellington and Nelson. New Zealand Journal of Ecology 31: 22–38.
Sun, O.J.; Sweet, G.B. 1996: Comparison of frost tolerance of Nothofagus solandri var. cliffortioides (Hook.f.) Poole and Nothofagus menziesii (HooW.i.) Oerst. New Zealand Journal of Botany 34: 273–278.
Sykes, M.T.; Wilson, J.B. 1987: The vegetation of a New Zealand dune slack. Vegetatio 71: 13–19.
Sykes, W.R. 1977: Kermadec Islands flora. An annotated check list. Bulletin 219. Department of Scientific and Industrial Research, Christchurch. 216 p.
Tansley, A.G. 1935: The use and abuse of vegetational concepts and terms. Ecology 16: 284–307.
Tedrow, C.F.; Cantlon, J.E. 1958: Concepts of soil formation and classification in arctic regions. Arctic 11: 166–179.
Thannheiser, D.; Holland, P. 1994: The plant communities of New Zealand salt meadows. Global Ecology and Biogeography Letters 4: 107–115.
Thompson, S.; Gruner, I.; Gapare, N. 2004: New Zealand Land Cover Database Version 2. Illustrated guide to target classes. Report for the Ministry for the Environment, Wellington. 126 p.
Thornthwaite, C.W. 1948: An approach toward a rational classification of climate. Geographic Review 38: 55–94.
Thorp, J.; Smith, G.D. 1949: Higher categories of soil classification: order, suborder, and great soil groups. Soil Science 67: 117–126.
Towns, D.R.; Wardle, D.A.; Mulder, C.P.H.; Yeates, G.W.; Fitzgerald, B.M., Parrish, G.R.; Bellingham, P.J.; Bonner, K.I. 2009: Predation of seabirds by invasive rats: multiple indirect consequences for invertebrate communities. Oikos 118: 420–430.
Trakhtenbrot, A.; Kadmon, R. 2005: Environmental cluster analysis as a tool for selecting complementary networks of conservation sites. Ecological Applications 15: 335–345.
Walker, S.; King, N.; Monks, A.; Williams, S.; Burrows, L.; Cieraad, E.; Meaurk, C.; Overton, J.McC.; Price, R.; Smale, M. 2009: Secondary woody vegetation patterns in New Zealand’s South Island dryland zone. New Zealand Journal of Botany 47: 367–393.
Walker, S.; Lee, W.G. 2000: Alluvial grasslands in south-eastern New Zealand: vegetation change, long-term and post-pastoral change. Journal of the Royal Society of New Zealand 30: 69–103.
Walker, S.; Rogers, G.M.; Lee, W.G.; Rance, B.; Ward, D.; Rufaut, C.; Conn, A.; Simpson, N.; Hall, G.; Larivière, M.C. 2006: Consequences to threatened plants and insects of fragmentation of Southland floodplains forests. Science for Conservation 265. Department of Conservation, Wellington. 86 p.
Walter, H. 1985: Vegetation of the earth and ecological systems of the geobiosphere. 3rd edition. Springer-Verlag, Berlin. 318 p.
Wardle, P. 1964: Facets of the distribution of forest vegetation in New Zealand. New Zealand Journal of Botany 2: 352–366.
Wardle, P. 1966: Biological flora of New Zealand. 1. Weinmannia racemosa Linn f. (Cunoniaceae) Kamahi. New Zealand Journal of Botany 4: 114–131.
Wardle, P. 1971: An explanation for alpine timberline. New Zealand Journal of Botany 9: 371–402.
Wardle, P. 1979: Plants and landscape in Westland National Park. National Parks Scientific Series Number 3. Department of Lands and Survey, Wellington. 168 p.
Wardle, P. 1991: Vegetation of New Zealand. Cambridge University Press, Cambridge. 672 p.
Wells, J.A. 1972: Ecology of Podocarpus hallii in Central Otago, New Zealand. New Zealand Journal of Botany 10: 399–426.
Whitehouse, I.E. 1982: Numerical assessment of erosion from old and recent photographs: a case study from a section of Highway 73, Canterbury, New Zealand. Journal of the Royal Society of New Zealand 12: 91–101.
Whittaker, R.H. 1975: Communities and ecosystems. Macmillan, New York. 158 p.
Williams, P.A.; Wiser, S.K. 2004: Determinants of regional and local patterns in the floras of braided riverbeds in New Zealand. Journal of Biogeography 31: 1355–1372.
Williams, P.A.; Wiser, S.; Clarkson, B.; Stanley, M. 2007: New Zealand’s historical rare terrestrial ecosystems set in a physical and physiognomic framework. New Zealand Journal of Ecology 31: 119–128.
Wilson, H.D. 1976: Vegetation of Mount Cook National Park. National Parks Authority Scientific Series Number 1. Department of Lands and Survey, Wellington. 138 p.

35Science for Conservation 325
Wilson, H.D. 1987: Plant communities of Stewart Island. New Zealand Journal of Botany 25 (supplement): 1–81.
Wiser, S.K.; Buxton, R.P. 2009: Montane outcrop vegetation of Banks Peninsula, South Island, New Zealand. New Zealand Journal of Ecology 33: 164–176.
Wiser, S.K.; Buxton, R.P.; Clarkson, B.R.; Richardson, S.J.; Rogers, G.M.; Smale, M.C.; Williams, P.A. 2010: Climate, landscape and microenvironment interact to determine plant composition in naturally discrete gravel beach communities. Journal of Vegetation Science 21: 657–671.
Wiser, S.K.; Hurst, J.M. 2008: Classification of New Zealand’s forest and shrubland communities based on a national plot sampling on an 8-km grid. Landcare Research Contract Report for the Department of Conservation (unpublished). 71 p.
Wiser, S.K.; Hurst, J.M.; Wright, E.F.; Allen, R.B. 2011: New Zealand’s forest and shrubland communities: a quantitative classification based on a nationally representative plot network. Applied Vegetation Science 14: 506–523.
Wright, A.E. 1980: Vegetation and flora of Fanal Island, Mokohinau group. Tane 26: 25–43.

36 Singers & Rogers—Classification of terrestrial ecosystems
Appendix 1
Glossary of common and scientific names of plant species Common names follow Nicol (1997).
COMMON NAME SCIENTIFIC NAME
akeake Dodonaea viscosa
akiraho Olearia paniculata
arrow grass Triglochin striata
Azolla Azolla filiculoides
bachelor’s button Cotula coronopifolia
bamboo rush Sporadanthus ferrugineus
beach morning glory Ipomoea pes-caprae subsp. brasiliensis
beech (forest or species) Fuscospora spp. and Lophozonia menziesii
black beech Fuscospora solandri
black maire Nestegis cunninghamii
blue tussock Poa colensoi
blue wheatgrass Anthosachne solandri
bog pine Halocarpus bidwillii
bracken Pteridium esculentum
bristle tussock Rytidosperma setifolium
broadleaf Griselinia littoralis
broad-leaved snow tussock Chionochloa flavescens
buggar grass Austrostipa stipoides
bush tussock Chionocloa conspicua
buttercup Ranunculus spp.
cabbage tree Cordyline australis
carpet grass Chionochloa australis
Chatham Island akeake Olearia chathamica
Chatham Island bamboo rush Sporadanthus traversii
Chatham Island karamū Coprosma chathamica
Chatham Island toetoe Austroderia turbaria
coastal tussock Chionochloa bromoides
copper tussock Chionochloa rubra subsp. cuprea
crown fern Blechnum discolor
eelgrass Zostera muelleri subsp. novaezelandica
five-finger Pseudopanax arboreus
flax Phormium spp.
geothermal kānuka Kunzea ericoides var. microflora
glasswort Sarcocornia quinqueflora
greater wire rush Empodisma robustum
gumland grass tree Dracophyllum lessonianum
half-star Selliera radicans
Hall’s tōtara Podocarpus cunninghamii
hangehange Geniostoma ligustrifolium
harakeke, flax Phormium tenax
hard beech Fuscospora truncata
hard tussock Festuca novae-zelandiae
Continued on next page

37Science for Conservation 325
COMMON NAME SCIENTIFIC NAME
hīnau Elaeocarpus dentatus
horopito Pseudowintera colorata
houhere Hoheria spp.
houpara Pseudopanax lessonii
hutu Ascarina lucida var. lucida
ice plant Disphyma australe subsp. australe
inaka Dracophyllum longifolium
kahikatea Dacrycarpus dacrydioides
kaikōmako Pennantia corymbosa
kāmahi Weinmannia racemosa
kānuka Kunzea ericoides
karaka Corynocarpus laevigatus
karamū Coprosma robusta
kāretu Hierochloe redolens
karo Pittosporum crassifolium
kātote Cyathea smithii, C. dealbata
kauri Agathis australis
kawaka Libocedrus plumosa
kawakawa Piper excelsum
Kermadec fireweed Senecio kermadecensis
Kermadec ice plant Disphyma australe subsp. stricticaule
Kermadec ngaio Myoporum rapense subsp. kermadecense
Kermadec pōhutukawa Metrosideros kermadecensis
kiekie Freycinetia banksii
kiokio Blechnum spp.
Kirk’s pine Halocarpus kirkii
Kirk’s scurvy grass Lepidium kirkii
Kirk’s tree daisy Brachyglottis kirkii
knobby clubrush Ficinia nodosa
kohekohe Dysoxylum spectabile
kōhūhū Pittosporum tenuifolium
kopoti Anisotome aromatica
korokia Corokia cotoneaster
koromiko Hebe stricta
kōtukutuku Fuchsia excorticata
kōwhai Sophora spp.
kuta Eleocharis sphacelata
lake clubrush Schoenoplectus tabernaemontani
lancewood Pseudopanax crassifolius
leatherwood Olearia colensoi
lesser wire rush Empodisma minus
long-hair plume grass Dichelachne crinita
māhoe Melicytus ramiflorus
maire Nestegis spp.
makamaka Ackama rosifolia
mangeao Litsea calicaris
mangrove Avcennia marina subsp. australasica
mānuka Leptospermum scoparium
matagouri Discaria toumatou
Appendix 1—continued
Continued on next page

38 Singers & Rogers—Classification of terrestrial ecosystems
COMMON NAME SCIENTIFIC NAME
māpou Myrsine australis
mataī Prumnopitys taxifolia
mid-ribbed snow tussock Chionochloa pallens
miro Prumnopitys ferruginea
monoao Dracophyllum subulatum
mountain beech Fuscospora cliffortioides
mountain celery pine Phyllocladus alpinus
mountain neinei Dracophyllum traversii
mountain oat grass Deyeuxia avenoides
mountain tauhinu Ozothamnus vauvilliersii
mountain tōtara Podocarpus cunninghamii
mountain tutu Coriaria plumosa
mountain wineberry Aristotelia fruticosa
mutton bird sedge Carex trifida
narrow-leaved houhere Hoheria angustifolia
narrow-leaved maire Nestegis montana
narrow-leaved snow tussock Chionochloa rigida
New Zealand spinach Tetragonia tetragonoides
ngaio Myoporum laetum
nīkau Rhopalostylis sapida
northern rātā Metrosideros robusta
oioi Apodasmia similis
pāhautea Libocedrus bidwillii
parapara Pisonia brunoniana
parataniwha Elatostema rugosum
Parkinson’s rātā Metrosideros parkinsonii
pigweed Einadia spp.
pīngao Ficinia spiralis
pink pine Halocarpus biformis
pōhutukawa Metrosideros excelsa
pōkākā Elaeocarpus hookerianus
porcupine shrub Melicytus alpinus
porokaiwhiri Hedycarya arborea
prickly mingimingi Leptocophylla juniperina subsp. juniperina
prostrate kōwhai Sophora prostrata
puka Meryta sinclarii
pukatea Laurelia novae-zelandiae
pūkio Carex secta, C. virgata
pungent snow tussock Chionochloa crassiuscula subsp. crassiuscula
pūriri Vitex lucens
putaputawētā Carpodetus serratus
rātā lianes Metrosideros spp.
raukawa Raukaua edgerleyi
raupō Typha orientalis
red beech Fuscospora fusca
red māpou Myrsine australis
red tussock Chionochloa rubra
rengarenga Arthropodium cirratum
rewarewa Knightia excelsa
Appendix 1—continued
Continued on next page

39Science for Conservation 325
COMMON NAME SCIENTIFIC NAME
ribbonwood Plagianthus regius
rimu Dacrydium cupressinum
salt marsh ribbonwood Plagianthus divaricatus
sand buttercup Ranunculus acaulis
sand coprosma Coprosma acerosa
sand daphne Pimelea villosa
sand tussock Poa billardierei
scabweed Raoulia spp.
sea blite Suaeda novae-zelandiae
seagrass Zostera muelleri subsp. novozelandica
sea primrose Samolus repens
sea rush Juncus krausii var. australiensis
shining karamū Coprosma lucida
shore bindweed Calystegia soldanella
shore celery Apium prostratum
shore groundsel Senecio lautus subsp. lautus
shore spurge Euphorbia glauca
silver beech Lophozonia menziesii
silver pine Manoao colensoi
silver tussock Poa cita
slim snow tussock Chionochloa macra
small-leaved pōhuehue Muehlenbeckia complexa
snow tōtara Podocarpus nivalis
snow tussock Chionochloa spp.
southern rātā Metrosideros umbellata
sphagnum Sphagnum cristatum
spinifex Spinifex sericeus
square sedge Lepidosperma australe
Stewart Island snow tussock Chionochloa lanea
subantarctic snow tussock Chionochloa antarctica
supplejack Ripogonum scandens
swamp akeake Olearia telmatica
swamp maire Syzygium maire
swamp millet Isachne globosa
tall mingimingi Leucopogon fasciculatus
tānekaha Phyllocladus trichomanoides
tangle fern Gleichenia dicarpa
tarahinau Dracophyllum arboreum
taraire Beilschimiedia tarairi
tarata Pittosporum eugenioides
tauhinu Ozothamnus leptophyllus
taupata Coprosma repens
tawa Beilschmiedia tawa
tawāpou Pouteria costata
tāwari Ixerba brexioides
tāwheowheo Quintinia serrata
three-finger Raukaua simplex
three-square Schoenoplectus pungens
tītoki Alectryon excelsus
Appendix 1—continued
Continued on next page

40 Singers & Rogers—Classification of terrestrial ecosystems
ReferencesNicol, E.R. 1997: Common names of plants in New Zealand. Manaaki Whenua Press, Lincoln. 115 p.
Appendix 1—continued
COMMON NAME SCIENTIFIC NAME
toatoa Phyllocladus glaucus
toetoe Austroderia spp.
tōtara Podocarpus totara
tōwai Weinmannia silvicola
tree ferns Cyathea spp. and Dicksonia spp.
tutu Coriaria spp.
twiggy tree daisy Olearia virgata
vegetable sheep Haastia spp.
water milfoil Myriophyllum spp.
weeping matipo Myrsine divaricata
Westland tōtara Podocarpus totara var. waihoensis
wharangi Melicope ternata
wharariki Phormium cookianum
white maire Nestegis lanceolata
willowherb Epilobium spp.
wineberry Aristotelia serrata
wire rush Empodisma minus
woolly moss Racomitrium lanuginosum
yellow silver pine Lepidothamnus intermedius

41Science for Conservation 325
Appendix 2
Ecosystem frameworkThe following tables outline the classification of New Zealand terrestrial ecosystems using a hierarchy of environmental gradients and factors as an abiotic framework against which vegetation community and ground cover classes are arranged. It includes zonal and azonal sections (see section 2—Methods). Common names follow Nicol (1997) and http://nzpcn.org.nz, and the scientific names for these are provided in Appendix 1.
Moisture classes are defined as follows: semi-arid—where the average Penman annual water deficit is greater than 270 mm per annum; sub-humid—the humidity zone that lies between the humid and semi-arid class boundaries; humid—where rainfall is at least four times greater than evapotranspiration.
Ecosystem unit code abbreviations are: SF, subtropical forest; WF, warm forests; MF, mild forests; CLF, cool forests; CDF, cold forests and scrub; AL, low alpine; AH, high alpine; TI, cold air inversion; WL, wetlands; DN, active coastal sand dunes; EP, erosion pavements; CL, cliffs; SC, screes and boulderfields; BR, braided rivers; SA, saline; UM, ultramafic; GT, geothermal; CV, cave; VS, fire- and volcanic activity-induced succession.
Table A2.1. Zonal ecosystems.
PRIMARY ECOSYSTEM
DRIVER:
TEMPERATURE
SECONDARY
ECOSYSTEM
DRIVER:
MOISTURE
TERTIARY ECOSYSTEM DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Subtropical: Mean summer temperature > 22.5ºC and frost free [Subtropical forests]
Sub-humid Coastal hillslopes and hillcrests SF1 Kermadec pōhutukawa forest
Warm temperate: Mean summer temperature 17.5–22.5ºC [Warm forests]
Semi-arid Coastal hillslopes and hillcrests WF1 Tītoki, ngaio forest
Recent alluvial terraces with free-draining stony and alluvial soils
WF2 Tōtara, mataī, ribbonwood forest
Hillslopes and hillcrests WF3 Tawa, tītoki, podocarp forest
Sub-humid Coastal hillslopes and hillcrests WF4 Pōhutukawa, pūriri, broadleaved forest [Coastal broadleaved forest]
Stable dunes with free-draining recent sandy soils
WF5 Tōtara, kānuka, broadleaved forest [Dune forest]
Stable dunes of the Foxton (> 100 years old), Motuiti (500 years old) and Waitarere (2000–3000 years old) phases, with free-draining recent sandy soils
WF6 Tōtara, mataī, broadleaved forest [Dune forest]
Recent alluvial terraces and rolling hillslopes with moderately fertile, free-draining recent and granular soils
WF7 Pūriri forest
Recent alluvial terraces and basins of dune plains with poor-draining recent, gley and organic soils
WF8 Kahikatea, pukatea forest
Rolling hillslopes and older alluvial surfaces with andesitic- and basaltic-derived clay loams
WF9 Taraire, tawa, podocarp forest
Hillslopes and hillcrests with moderately to strongly leached ultic soils grading into podzols
WF10 Kauri forest
Continued on next page

42 Singers & Rogers—Classification of terrestrial ecosystems
PRIMARY ECOSYSTEM
DRIVER:
TEMPERATURE
SECONDARY
ECOSYSTEM
DRIVER:
MOISTURE
TERTIARY ECOSYSTEM DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
[Warm temperate contd.] [Sub-humid contd.]
Hillslopes and hillcrests with brown grading to ultic soils
WF11 Kauri, podocarp, broadleaved forest
Hillslopes and hillcrests largely of subdued relief plus beech
WF12 Kauri, podocarp, broadleaved, beech forest
Hillslopes, hillcrests and older alluvial terraces
WF13 Tawa, kohekohe, rewarewa, hīnau, podocarp forest
Hillslopes and hillcrests plus beech WF14 Kāmahi, tawa, podocarp, hard beech forest
Mild temperate: Mean summer temperature 15–17.5ºC [Mild forests]
Semi-arid Hillslopes and older alluvial terraces with brown and pallic soils
MF1 Tōtara, tītoki forest
Hillslopes with brown soils MF2 Rimu, mataī, hīnau forest
Hillslopes and older alluvial terraces with brown, pallic and older recent soils
MF3 Mataī, tōtara, kahikatea, broadleaved forest
Recent alluvial terraces with poor-draining recent, gley and organic soils
MF4 Kahikatea forest
Rolling hillslopes and alluvial terraces plus beech
MF5 Black beech forest
Sub-humid to humid
Hillslopes, hillcrests, older alluvial surfaces and dunes, e.g. Foxton Phase
MF6 Kohekohe, tawa forest
Hillslopes, hillcrests and older alluvial terraces
MF7 Tawa, kāmahi, podocarp forest
MF8 Kāmahi, broadleaved, podocarp forest
Steep hillslopes and cliffs of welded volcanic ignimbrites, with shallow recent volcanic soils plus beech locally
MF9 Tānekaha forest, locally with hard beech
Rolling hillslopes with deep volcanic tephra soils, e.g. Taupo Pumice soils
MF10 Tōtara, mataī, kahikatea forest
MF11 Rimu forest
Coastal hillslopes and hillcrests plus beech MF12 Rātā, hard beech, kāmahi forest
Recent alluvial terraces and depressions with poor-draining recent, gley and organic soils
MF13 Kahikatea, northern rātā, kāmahi forest
MF14 Kahikatea, silver pine, kāmahi forest
Recent alluvial terraces (< 800 years old) with free-draining stony and alluvial soils
MF15 Kahikatea, mataī, Westland tōtara, rimu forest
Glaciated outwash terraces with podzols, Waiuta soil (5000–12 000 years old) and marine terraces with gley soils
MF16 Rimu forest
Glacial outwash terraces with podzol soils, Kumara soil (> 12 000 to < 22 000 years of age), and thin soils of quartzose sandstones and coal measures
MF17 Rimu, kāmahi, tāwheowheo forest
Glacial landforms (outwash terraces, roche moutonee and nunatak on gneiss and granite) with podzol soils (e.g. Okarito soil) (c. > 22 000 years old), gley podzol and organic soils
MF18 Yellow silver pine, mānuka forest
Hillslopes, hillcrests and terraces (800–2000 years old) with brown soils
MF19 Kāmahi, rimu, miro, southern rātā forest
Hillslopes and hillcrests plus beech MF20 Hard beech forest
MF21 Tawa, kāmahi, rimu, northern rātā, black beech forest
MF22 Tawa, rimu, northern rātā, beech forest
Table A2.1—continued
Continued on next page

43Science for Conservation 325
PRIMARY ECOSYSTEM
DRIVER:
TEMPERATURE
SECONDARY
ECOSYSTEM
DRIVER:
MOISTURE
TERTIARY ECOSYSTEM DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
[Mild temperate contd.] [Sub-humid to humid contd.]
Rolling hillslopes with organic soils MF23 Chatham Island akeake, karamū, māhoe, ribbonwood forest
Hillslopes, hillcrests and plateaus with leached brown and granular soils developing towards podzols
MF24 Rimu, tōwai forest
Hillslopes and hillcrests with leached brown soils developing towards podzols
MF25 Kauri, tōwai, rātā, montane podocarp forest
Cool temperate: Mean summer temperature 12.5–15ºC [Cool forests and scrub]
Semi-arid to sub-humid
Hillslopes and hillcrests CLF1 Hall’s tōtara, mountain celery pine, broadleaf forest
Sub-humid Stable dunes with free-draining recent sandy soils
CLF2 Hall’s tōtara forest [Dune forest]
Recent alluvial terraces with free-draining stony and alluvial soils
CLF3 Podocarp, ribbonwood, kōwhai forest
Hillslopes and hillcrests with brown soils CLF4 Kahikatea, tōtara, mataī forest
Hillslopes and terraces with deep volcanic tephra soils, e.g. Taupo pumice soils
CLF5 Mataī, Hall’s tōtara, kāmahi forest
Humid Moderate to steep hillslopes and terraces CLF6 Kāmahi, southern rātā, podocarp forest
Hillslopes, marine terraces and poor-draining alluvial terraces with gley and podzol soils plus beech
CLF7 Rimu, kāmahi, beech forest
Hillslopes, hillcrests and terraces plus beech with brown soils developing towards podzols on localised stable surfaces
CLF8 Silver beech, kāmahi, southern rātā forest
CLF9 Red beech, podocarp forest
CLF10 Red beech, silver beech forest
CLF11 Silver beech forest
CLF12 Silver beech, mountain beech forest
Cold: Mean summer temperature 10–12.5ºC [Cold forests and scrub]
Semi-arid Hillslopes and hillcrests CDF1 Pāhautea, Hall’s tōtara, mountain celery pine, broadleaf forest
CDF2 Dracophyllum, mountain celery pine, Olearia, Hebe scrub [Subalpine scrub]
Hillslopes, hillcrests and terraces plus beech
CDF3 Mountain beech forest
Humid Hillslopes, hillcrests and depressions with free-draining and poor-draining soils
CDF4 Hall’s tōtara, pāhautea, kāmahi forest
CDF5 Hall’s tōtara, pāhautea, kāmahi, southern rātā forest
CDF6 Olearia, Pseudopanax, Dracophyllum scrub [Subalpine scrub]
Hillslopes, hillcrests and terraces plus beech with brown soils developing towards podzols on localised stable surfaces
CDF7 Mountain beech, silver beech, montane podocarp forest
Table A2.1—continued
Continued on next page

44 Singers & Rogers—Classification of terrestrial ecosystems
PRIMARY ECOSYSTEM
DRIVER:
TEMPERATURE
SECONDARY
ECOSYSTEM
DRIVER:
MOISTURE
TERTIARY ECOSYSTEM DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Low alpine: Mean summer temperature 5–10°C [Low alpine]
Sub-humid Hillslopes, basins and ridges with brown soils
AL1 Narrow-leaved and slim snow tussock tussockland/shrubland
Hillslopes, basins and ridges with raw and recent soils
AL2 Slim and mid-ribbed snow tussock tussockland/shrubland
Humid Hillslopes, hillcrests and depressions with recent volcanic soils developing towards organic soils
AL3 Red tussock tussockland/shrubland
Hillslopes, hillcrests and depressions with raw soils developing towards recent and organic soils
AL4 Mid-ribbed and broad-leaved snow tussock tussockland/shrubland
AL5 Mid-ribbed, broad-leaved, red and carpet grass tussockland/shrubland
AL6 Mid-ribbed and narrow-leaved snow tussock tussockland/shrubland
AL7 Pungent snow tussock tussockland/shrubland
Hillslopes, hillcrests and depressions with organic soils
AL8 Stewart Island snow tussock tussockland/shrubland
AL9 Subantarctic snow tussock tussockland/shrubland
High alpine: Mean summer temperature < 5°C [High alpine]
Sub-humid Hillslopes and hillcrests with raw soils AH1 Gravelfield/stonefield [Fellfield]
Gently rolling schist hillslopes, hillcrests and plains with acidic brown soils
AH2 Dracophyllum muscoides cushionfield
Humid Hillslopes and hillcrests with raw and recent soils
AH3 Gravelfield/stonefield, mixed species cushionfield
Hillslopes and hillcrests with raw and recent soils of andesitic ash origin
AH4 Woolly moss, bristle tussock, blue tussock mossfield tussockland/stonefield
Temperature: > –9ºC frosts [Cold air inversion topography]
Semi-arid Mosaic of hummocky moraines and peri-glacial outwash, and alluvial terraces and plains with stony, semi-arid soils, e.g. Tekapo soils
TI1 Bog pine, mountain celery pine scrub/forest
Terraces with recent semi-arid alluvial and pallic soils, e.g. Fork or Molyneaux soils
TI2 Kānuka, Olearia scrub/treeland
Sub-humid Terraces and plains with recent deep pumice soils, e.g. Poronui and Kaingaroa soils
TI3 Monoao scrub/lichenfield
Terraces, hillslopes and glacial moraines with alluvial, poor- and free-draining recent soils
TI4 Coprosma, Olearia, matagouri scrub [Grey scrub]
Humid Terraces, depressions and montane plateaus with recent volcanic, alluvial and podzol soils
TI5 Bog pine, mountain celery pine, silver pine scrub/forest
Montane to subalpine terraces and montane plateaus with poor-draining and organic soils
TI6 Red tussock tussockland
Table A2.1—continued

45Science for Conservation 325
Table A2.2. Azonal ecosystems. The pr imary dr ivers for these ecosystems are A. h igh water tables, B. geomorphic disturbance, C. extreme soi l and/or atmospher ic chemistry, D. geothermal ly extreme heat and chemistry, E. l ight l imitat ion, and F. anthropogenic f i re.
PRIMARY
ECOSYSTEM
DRIVER:
HYDROLOGY
SECONDARY
ECOSYSTEM
DRIVER:
FERTILITY
TERTIARY
ECOSYSTEM
DRIVER:
TEMPERATURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Permanent or seasonally high water table [Wetlands]
Oligotrophic—low nutrient status and high acidity [Bogs]
Warm temperate Hillslopes and depressions with kauri podzols, e.g. Wharekohe or Te Kopuru soils
WL1 Mānuka, gumland grass tree, Machaerina scrub/sedgeland [Gumland]
Depressions or the lagg of raised bogs with organic soils
WL2 Mānuka, greater wire rush restiad rushland
Raised bogs on in-filled lagoons/ river oxbows with deep organic soils
WL3 Bamboo rush, greater wire rush restiad rushland
Mild to cool temperate
Glacial moraines with strongly leached and acidic gley podzols (lacking peat), e.g. Okarito soils
WL4 Mānuka, lesser wire rush, tangle fern scrub/fernland/restiad rushland [Pakihi]
Depressions and raised bogs with organic soils
WL5 Chatham Island bamboo rush restiad rushland
Cool temperate to low alpine
Depressions, raised bogs and gentle slopes with organic soils
WL6 Lesser wire rush, tangle fern restiad rushland/fernland
Gentle hillslopes, plateaus and depressions with organic soils
WL7 Tall tussock tussockland
Cold to high alpine
Gentle hillslopes and depressions with organic soils
WL8 Herbfield/mossfield/sedgeland
Shallow hillslopes, plateaus and terraces with organic soils
WL9 Cushionfield
Mesotrophic—moderate fertility and weak to neutral acidity [Fens and marshes]
Warm to cool temperate
Freshwater margins of estuaries, tidal rivers, coastal lagoons and some inland lakes
WL10 Oioi restiad rushland/reedland
Depressions, and lake and lagoon margins
WL11 Machaerina sedgeland
Depressions WL12 Mānuka, tangle fern scrub/fernland
Depressions, and lake and lagoon margins
WL13 Sphagnum mossfield
Mild temperate to cold
Depressions within a wide range of landforms
WL14 Herbfield [Ephemeral wetland]
Warm temperate to cold
Lake and lagoon margins WL15 Herbfield [Lakeshore turf]
Cool temperate to low alpine
Depressions WL16 Red tussock, Schoenus pauciflorus tussockland
Cold to high alpine
Hillslopes WL17 Schoenus pauciflorus sedgeland [Alpine seepages/flushes]
A
Continued on next page

46 Singers & Rogers—Classification of terrestrial ecosystems
Table A2.2A—continued
PRIMARY
ECOSYSTEM
DRIVER:
FREQUENT
GEOMORPHIC
DISTURBANCE
SECONDARY
ECOSYSTEM
DRIVER:
TEMPERATURE
TERTIARY
ECOSYSTEM
DRIVER:
MOISTURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Erosion and accretion of sand from wind ablation[Active coastal sand dunes]
Subtropical Humid Dunes with raw sandy soils in association with atmospheric salinity (e.g. spume and salt-spray)
DN1 Beach morning glory, knobby clubrush vineland/sedgeland
Warm to mild temperate
Semi-arid to humid
DN2 Spinifex, pīngao grassland/sedgeland
Mild to cool temperate
Semi-arid to humid
DN3 Pīngao sedgeland
Mild temperate Sub-humid DN4 Pīngao sedgeland, mega-herbfield
Warm to cool temperate
Semi-arid to humid
Dune plains (including deflation hollows, dune slacks, damp sand plains and stream terraces) and exposed coastal hill slopes with raw sandy soils
DN5 Oioi, knobby clubrush sedgeland
Erosion from wind ablation [Erosion pavements]
Mild temperate to low alpine
Sub-humid to humid
Erosion-bared rockland, especially quartzose sandstones, marble and granite
EP1 Rockland
Erosion associated with gravity and wind ablation[Cliffs]
Warm temperate Sub-humid Cliffs, bluffs, rock-stacks and their talus in association with atmospheric salinity (e.g. spume and salt-spray) [Coastal cliffs]
CL1 Pōhutukawa treeland/flaxland/rockland
Warm to mild temperate
Semi-arid to sub-humid
CL2 Ngaio, taupata treeland/herbfield/rockland
Mild temperate Sub-humid CL3 Coprosma, Muehlenbeckia shrubland/herbfield/rockland
Humid CL4 Chatham Island akeake, Hebe, māhoe treeland/herbfield/rockland
Mild to cool temperate
Humid CL5 Harakeke, Hebe elliptica flaxland/rockland
B
Continued on next page
PRIMARY
ECOSYSTEM
DRIVER:
HYDROLOGY
SECONDARY
ECOSYSTEM
DRIVER:
FERTILITY
TERTIARY
ECOSYSTEM
DRIVER:
TEMPERATURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
[Permanent or seasonally high water table contd.]
Eutrophic—high fertility and weak acidity to weak alkalinity [Swamps and marshes]
Warm to cool temperate
Depressions and terraces with recent and organic soils
WL18 Flaxland
Depressions, and lake and lagoon margins with recent and organic soils
WL19 Raupō reedland
Depressions with recent and organic soils
WL20 Coprosma, twiggy tree daisy scrub
Mild temperate Depressions with recent and organic soils
WL21 Swamp akeake, Chatham Island karamū, Coprosma propinqua var. martinii short forest/flaxland
Cool temperate to cold
Depressions, and lake and lagoon margins with recent and organic soils
WL22 Carex, Schoenus pauciflorus sedgeland

47Science for Conservation 325
PRIMARY
ECOSYSTEM
DRIVER:
FREQUENT
GEOMORPHIC
DISTURBANCE
SECONDARY
ECOSYSTEM
DRIVER:
TEMPERATURE
TERTIARY
ECOSYSTEM
DRIVER:
MOISTURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
[Erosion associated with gravity and wind ablation contd.]
Warm to mild temperate
Semi-arid to sub-humid
Cliffs, bluffs, tors and their talus [Inland cliffs]
CL6 Hebe, wharariki flaxland/rockland
Mild to cool temperate
Semi-arid to sub-humid
CL7 Pachystegia, Carmichaelia shrubland/tussockland/rockland
Sub-humid to semi-arid
CL8 Helichrysum, Melicytus shrubland/tussockland/rockland
Warm to mild temperate
Humid CL9 Parataniwha, Machaerina sinclairii herbfield/sedgeland
Mild to cool temperate
Humid CL10 Kiokio fernland/rockland
Cool temperate Humid CL11 Mountain tutu, Hebe, wharariki, Chionochloa shrubland/tussockland/rockland
Erosion associated with freeze-thaw cycles, gravity and wind ablation [Screes and boulderfields]
Cool temperate to high alpine
Semi-arid to sub-humid
Steep hillslopes with shattered, predominantly greywacke, argillite, basalt and limestone gravels and boulders
SC1 Gravelfield [Screes and boulderfields]
Erosion (and sediment accretion) associated with flooding [Braided rivers]
Mild to cool temperate
Sub-humid to semi-arid
Riverbeds and fans with raw sand, gravels and boulders
BR1 Hard tussock, scabweed gravelfield/stonefield
Humid BR2 Scabweed gravelfield/stonefield
Cool temperate Humid BR3 Bristle tussock, Raoulia, Muehlenbeckia gravelfield/sandfield
Table A2.2B—continued
Continued on next page

48 Singers & Rogers—Classification of terrestrial ecosystems
PRIMARY
ECOSYSTEM
DRIVER:
EXTREME
CHEMISTRY
SECONDARY
ECOSYSTEM
DRIVER:
TEMPERATURE
TERTIARY
ECOSYSTEM
DRIVER:
MOISTURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Salinity > 5% associated with tides or in lagoons [Estuarine hydrosystems]
Warm temperate Sub-humid River mouths, inlets, estuaries and lagoons with sulphuric gley and recent gley soils, locally with shell and/or gravel barrier beaches
SA1 Mangrove forest and scrub
Warm to cool temperate
Semi-arid to humid
SA2 Searush, oioi, glasswort, sea primrose rushland/herbfield [Saltmarsh]
Mild to cool temperate
Sub-humid SA3 Glasswort, sea primrose herbfield [Saltmarsh]
Atmospheric salinity (e.g. spume and salt-spray) associated with persistent wind
Warm to cool temperate
Semi-arid to humid
Beach ridges with raw stony and shingle soils, locally with driftwood
SA4 Shore bindweed, knobby clubrush gravelfield/stonefield
Semi-arid to humid
Marine terraces, hillslopes, rocks and cliffs rarely with marine mammal disturbance
SA5 Herbfield [Coastal turf]
Subtropical Humid Coastal hillslopes and cliffs with seabird and marine mammal disturbance, and guano-enhanced soil fertility
SA6 Kermadec ngaio scrub, mixed herbfield/loamfield
Warm to mild temperate
Semi-arid to sub-humid
SA7 Ice plant, glasswort herbfield/loamfield
Cool temperate Sub-humid SA8 Mutton bird sedge and Senecio radiolatus sedgeland/herbfield/loamfield
Humid SA9 Olearia, Brachyglottis, Dracophyllum scrub/herbfield/loamfield [Mutton-bird scrub]
Humid SA10 Poa litorosa grassland/herbfield/loamfield
Alkaline soils with pH > 8 (saline soils, saline-sodic soils and sodic soils) [Inland saline soils]
Mild to cool temperate
Semi-arid Hillslopes, terraces and floodplains with Manorburn soils
SA11 Kirk’s scurvy grass herbfield/loamfield
Parent material of high magnesium, nickel, chromium and cobalt concentrations [Ultramafic or ultrabasic]
Warm temperate Sub-humid Coastal cliffs and hillslopes with raw and recent soils
UM1 Pōhutukawa, tānekaha forest/scrub/rockland
Warm to cool temperate
Sub-humid to humid
Hillslopes and alluvial terraces plus beech with raw and recent soils
UM2 Conifer, beech, mānuka forest/scrub/rockland
Cool temperate to low alpine
Humid Hillslopes, hillcrests and boulderfields with raw and recent soils
UM3 Tussockland/stonefield/rockland
Table A2.2—continued
C
Continued on next page

49Science for Conservation 325
PRIMARY
ECOSYSTEM
DRIVER:
TEMPERATURE
SECONDARY
ECOSYSTEM
DRIVER:
HYDROLOGY
TERTIARY
ECOSYSTEM
DRIVER:
MOISTURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Soil and associated ground water with temperatures > 20°C associated with geothermal heat [Geothermal]
NA Sub-humid to humid
Hillslopes, hillcrests and depressions with hydrothermally-altered raw and recent soils with extreme chemistry, low pH, and high concentrations of Al, SO4 and Fe
GT1 Geothermal kānuka scrub
Water and steam of a range of temperature, pH and chemistry
NA Geysers, pools, springs, streams, fumaroles and sinter terraces (including their margins)
GT2 Geothermally-heated water and steam
Table A2.2—continued
D
PRIMARY
ECOSYSTEM
DRIVER:
SOLAR
RADIATION
DEFICIENCY
SECONDARY
ECOSYSTEM
DRIVER:
TEMPERATURE
TERTIARY
ECOSYSTEM
DRIVER:
MOISTURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Lacking solar energy and associated photosynthetic energy inputs— energy inputs derived from entrance processes (air, water, organic material), and lithotrophs locally [Subterranean]
Low temperature fluctuations producing near-constant atmospheric conditions (mean annual temperature dependent on geographic location)
High humidity, typically > 95%
Caves CV1 Subterranean rockland, stonefield [Caves]
E
Continued on next page

50 Singers & Rogers—Classification of terrestrial ecosystems
PRIMARY
ECOSYSTEM
DRIVER:
COMBUSTION
AND/OR
VOLCANIC
ACTIVITY
[VEGETATION
SUCCESSION]
SECONDARY
ECOSYSTEM
DRIVER:
TEMPERATURE
TERTIARY
ECOSYSTEM
DRIVER:
MOISTURE
QUATERNARY ECOSYSTEM
DRIVERS:
LANDFORMS AND SOILS
ECOSYSTEM UNIT
CODE NAME
Large-scale disturbance from fire and/or volcanic activity
Subtropical to mild Sub-humid Hillslopes, ridges and terraces VS1 Pōhutukawa scrub/forest
Warm to cool temperate
Semi-arid to sub-humid
Hillslopes, ridges, terraces and plains VS2 Kānuka scrub/forest
Sub-humid Hillslopes, ridges and terraces VS3 Mānuka, kānuka scrub
Sub-humid to humid
Hillslopes, ridges and terraces VS4 Mānuka scrub
Semi-arid to sub-humid
Hillslopes, ridges and terraces VS5 Broadleaved species scrub/forest
Semi-arid to sub-humid
Hillslopes, ridges, terraces and plains VS6 Matagouri, Coprosma propinqua, kōwhai scrub [Grey scrub]
Mild to cool temperate
Semi-arid to sub-humid
Hillslopes, ridges and terraces VS7 Mountain tauhinu, Dracophyllum rosmarinifolium scrub
Sub-humid to humid
Hillslopes, ridges and terraces VS8 Monoao scrub
Cool temperate to cold
Sub-humid to humid
Hillslopes, ridges and terraces VS9 Inaka scrub
Warm to cool temperate
Semi-arid to humid
Hillslopes, ridges, terraces and plains VS10 Bracken fernland
Semi-arid to sub-humid
Hillslopes, ridges, terraces and plains VS11 Short tussock tussockland
Warm to mild temperate
Semi-arid to humid
Terraces and plains VS12 Sward grassland
Cool temperate Semi-arid to sub-humid
Hillslopes, ridges, terraces and plains VS13 Red or copper tussock tussockland
Sub-humid to humid
Hillslopes, ridges, terraces and plains VS14 Tall tussock tussockland
Table A2.2—continued
F
ReferencesNicol, E.R. 1997: Common names of plants in New Zealand. Manaaki Whenua Press, Lincoln. 115 p.

51Science for Conservation 325
Appendix 3
A classification of New Zealand terrestrial ecosystems The following classification provides ecosystem names, unit codes, descriptions and distributions of New Zealand terrestrial ecosystems. References are also provided, with ecosystem equivalents of previous classifications listed first, followed by other relevant literature. See Appendix 2 for a complete list of ecosystem unit codes and names, and Appendix 1 for the scientific names of all plant species.

52Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
SF1: Kermadec pōhutukawa forest
Broadleaved forest of abundant Kermadec pōhutukawa of two variants: 1. dry forest with an understorey of Myrsine kermadecensis, Coprosma acutifolia and Piper excelsum subsp. psittacorum; and 2. humid forest with māhoe and an understorey of Ascarina lucida var. lanceolata in association with Rhopalostylis baueri var. cheesemanii, Homalanthus polyandrus and Pseudopanax kermadecensis.
Restricted to the subtropical and frost-free Kermadec Island group (e.g. Raoul Island). Dry forest is the dominant type, while humid forest occurs within gullies and on the upper (often cloud-covered) hillcrests of Raoul Island. Secondary derivatives occur on some islands (e.g. Macauley Island).
Sykes (1977) and Wardle (1991: 432–436). Includes rare ecosystems: recent lava flows (< 1000 years old); cloud forest; coastal cliffs on acidic rock; and seabird-burrowed soil (Williams et al. 2007).
WF1: Tītoki, ngaio forest
Broadleaved forest of tītoki, ngaio, māhoe, five-finger, red māpou, kaikōmako, kōwhai, akeake and akiraho, locally occasional mataī, tōtara and kahikatea, and locally nīkau, tawa and rewarewa in northern and central part of range.
Eastern coastal areas of the North Island and South Island from Nuhaka (Wairoa District) to Kaikoura, and in semi-arid areas south to Banks Peninsula. Largely only secondary derivatives remain, often dominated by karaka (though this is likely of anthropogenic origin; Stowe 2003).
Not covered by Nicholls (1976), but equivalent to general hardwoods class. Wardle (1971), Wardle (1991: 384), Wilson (1992: 111 & 276–277), Maxwell et al. (1993), Moore (1999) and Whaley et al. (2001).
WF2: Tōtara, mataī, ribbonwood forest
Podocarp forest of abundant tōtara and mataī, with occasional kahikatea, ribbonwood and kōwhai, and a wide range of divaricating shrubs. Locally includes occasional tawa, tītoki and maire species in northern and more humid part of range and in inland examples, with occasional riparian black beech and red beech. Early successional derivatives on younger alluvial sites include kānuka, kōwhai, cabbage tree treeland and forest.
Predominantly east of the Main Divide on recent, free-draining, stony soils in warm to mild semi-arid areas in both the North Island and South Island, from Hawke’s Bay to North Otago. Also present in warm to mild sub-humid zones west of the Main Divide in Horowhenua, Manawatu and Rangitikei Districts, and warm sub-humid areas on free-draining, water-sorted Taupo pumice alluvium (e.g. Te Maire Scientific Reserve, Whanganui River). Historically, the early successional derivatives with kānuka may have been locally abundant in areas adjoining large braided rivers, especially on the Canterbury Plains (Meurk 2008).
Esler (1978b), Maxwell et al. (1993), Lake & Whaley (1995) and Meurk (2008).
WF3: Tawa, tīitoki, podocarp forest
Podocarp, broadleaved forest with emergent kahikatea, tōtara and mataī, abundant tawa and tītoki, and occasional rewarewa and hīnau. Locally includes northern rātā, pukatea, rimu and nīkau in warm and humid microclimates, and kohekohe in the northeast of the range. Occasionally black beech occurs on dry ridges. Very few intact examples remain—and most occur as secondary kānuka-dominant derivatives.
North Island, predominantly east of the Main Divide from Ruatoria to southern Wairarapa in warm to mild semi-arid areas. Also present in warm and mild sub-humid areas within the Manawatu, Horowhenua and Rangitikei Districts. Northern rātā, rimu and pukatea are absent in the driest areas.
Maxwell et al. (1993), Lake & Whaley (1995), Leathwick et al. (1995) and Whaley et al. (2001).
WF4: Pōhutukawa, pūriri, broadleaved forest [Coastal broadleaved forest]
Broadleaved forest of several variants, with pōhutukawa and pūriri, and locally with karaka, kohekohe, tītoki, mangeao, rewarewa, tawa, puka, tawāpou, ngaio, nīkau, taraire, and occasional tānekaha and kauri in northern part of range, and locally hard beech along the Bay of Plenty coast and East Cape (also with black beech). Kānuka and kōwhai locally occur on dry, steep ridges. Includes local endemic species and varieties where present on some northern offshore islands, especially Manawatāwhi/Three Kings Islands.
Warm climatic zone from the Manawatāwhi/Three Kings Islands and Te Paki south to Mahia and New Plymouth, with inland outliers occurring around some central North Island lakes. Southern boundary conforms to the southern limits of pūriri, aligning approximately to the thermic soil temperature zone (Molloy 1998: 209).
Equivalent to B1, P1–P3 and O1–O2 types of Nicholls (1976); and type e of Ecroyd (1982). Bayfield et al. (1991), Conning (2001) and Whaley et al. (2001). Includes rare ecosystem: recent lava flows (< 1000 years) (Williams et al. 2007).
Continued on next page

53Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
WF5: Tōtara, kānuka, broadleaved forest [Dune forest]
Mosaics of kānuka forest on younger (Holocene) dunes, grading into podocarp, broadleaved forest of tītoki, tōtara, māhoe, karaka, kohekohe, tawa, pūriri and hīnau, and locally pōhutukawa, narrow-leaved maire and taraire on older dunes.
Warm climatic zone in northern dunelands from North Cape to Kawhia, Coromandel, Matakana Island and Bay of Plenty coast to East Cape (e.g. Pretty Bush and Tapu Bush). Parent material mapped for Northland by Molloy (1998: 84). Very rare, fragmented and threatened. Predominantly only secondary derivatives of kānuka-dominant forest remain (Smale 1994). Locally pōhutukawa may dominate.
Not covered by Nicholls (1976), but equivalent to softwoods class. Smale (1994), Smale et al. (1996), Ogle (1997) and Conning (2001). Includes rare ecosystem: stable sand dunes (Williams et al. 2007).
WF6: Tōtara, mataī, broadleaved forest [Dune Forest]
Podocarp, broadleaved forest of mosaics of kānuka, red māpou, korokia and akeake on very recent soils, grading into ngaio, tītoki, kōwhai, tōtara, mataī, rewarewa, maire species, māhoe, lancewood and kaikōmako. Locally includes kohekohe on older dune soils in Horowhenua District.
Stable dunes in the Foxton Ecological District from Waverley to Paekakariki (e.g. Pakipaki Bush and Himatangi Scientific Reserve) of the Motuiti Phase. Soils have been described by Molloy (1998: 108–109). A similar variant may have occurred in Nelson, e.g. Farewell Spit, Rabbit Island. Largely only secondary derivatives of kānuka-dominant forest remain. Now very rare and fragmented.
Cockayne (1911), Ravine (1991) and Ogle (1997). Includes rare ecosystem: stable sand dunes (Williams et al. 2007).
WF7: Pūriri forest Broadleaved forest of abundant pūriri of three variants determined by landform and soil type: 1. occasional tōtara, mataī, kahikatea and tītoki locally, with kōwhai and taraire on alluvial, free-draining soils; 2. locally abundant taraire and kohekohe, and occasional tōtara, mataī, pukatea, rewarewa, karaka, tawa, tītoki and northern rātā, and abundant nīkau on fertile basaltic volcanic loam soils; and 3. occasional emergent kahikatea and kohekohe, and locally taraire, tītoki, pukatea and nīkau on moderately well-drained fluvial and allophanic soils derived from basaltic ash.
Predominantly warm climatic zones and moderately fertile to fertile soils on alluvial terraces and in recent basaltic areas. Variant 2 occurs on melanic granular soils, e.g. Papakauri soil in three main areas of occurrence: Pukekohe-Auckland, Whangarei and Kerikeri-Kaikohe. Mapped by Molloy (1998: 84). Youngest basaltic examples (occurring on more recent basaltic lava flows in the Auckland volcanic field) are colloquially described as ‘lava forests’ (Lindsay et al. 2009). Variant 3 occurs on moderately well-drained soils in Northland, south Auckland, western Waikato and the East Coast (e.g. Greys Bush near Gisborne). All types now extremely rare, fragmented and generally modified.
Clarkson & Clarkson (1991), Wardle (1991: 120), Tyrell et al. (1999: 53), Emmett et al. (2000), Conning (2001) and Lindsay et al. (2009).
WF8: Kahikatea, pukatea forest
Podocarp, broadleaved forest of abundant kahikatea, with occasional to abundant pukatea, kiekie and supplejack, and locally rimu, tawa and swamp maire, particularly on organic and gley soils with a high water table.
Predominantly west of the Main Divide on poor-draining alluvial, organic and gley soils in warm to mild, humid to sub-humid areas of the North Island, from Northland to Wellington (e.g. western Egmont National Park). Also occurs in localised areas in Nelson and Blenheim. East of the Main Divide it occurs in semi-arid regions, restricted small areas in permanent wet depressions and lake margins.
Equivalent to L1 type of Nicholls (1976). Smale (1984), Clarkson (1986), Clarkson et al. (1986), Bayfield et al. (1991: 153), Ravine (1991) and Conning (2001).
WF9: Taraire, tawa, podocarp forest
Podocarp, broadleaved forest of abundant taraire, with occasional rimu, miro, northern rātā, tawa, kohekohe, hīnau and rewarewa, and with pukatea and kahikatea commonly in gullies. Locally includes tōtara, pūriri and tōwai.
Predominantly in the warm climatic zone where kauri is absent (predominantly eastern), from the lower Waikato District northwards and throughout Northland below 450 m altitude. Also found on Great Barrier Island (Aotea Island). Kohekohe can be locally abundant (e.g. Waipoua), while tawa is more common at higher altitudes. Tōwai is locally absent from the Auckland region.
Equivalent to E1–E3 and O1–O3 types of Nicholls (1976). McKelvey & Nicholls (1959) and Barton (1972).
Appendix 3—continued
Continued on next page

54Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
WF10: Kauri forest Kauri forest with occasional podocarps (miro, rimu, toatoa, Hall’s tōtara, tānekaha) and broadleaved trees (northern rātā, tawa, taraire, hīnau, rewarewa, kohekohe and tōwai).
Warm to mild climatic zones, now largely restricted to the western Auckland to Northland hill country, e.g. Warawara, Waipoua Forest, and small patches within the Waitakere and Coromandel Ranges. Also present in warm humid areas. Kauri are predominantly tall with a large basal area, while podocarp and broadleaved trees are generally stunted.
Equivalent to A1–A2 types of Nicholls (1976); and types a–b of Ecroyd (1982). Wardle (1991: 117), Burns & Leathwick (1996) and Conning (2001).
WF11: Kauri, podocarp, broadleaved forest
Kauri, podocarp, broadleaved forest with occasional rimu, miro, kahikatea, kauri, taraire, tawa, tōwai, kohekohe, pūriri and rewarewa. Altitude variants occur, with taraire and kohekohe more abundant at lower altitudes, and tawa and tōwai more common at higher altitudes.
Warm to mild climatic zones north of Hamilton and Tauranga, including Puketi-Omahuta, Waitakere, Hunua, Coromandel and northern Kaimai Ranges. Commonly a secondary derivative of kauri forest.
Equivalent to B2–B9 types of Nicholls (1976); and type f of Ecroyd (1982). Wardle (1991: 117–119), Burns & Leathwick (1996) and Conning (2001).
WF12: Kauri, podocarp, broadleaved, beech forest
Kauri, podocarp, broadleaved and hard beech forest with occasional tānekaha, Hall’s tōtara or lowland tōtara, rimu, miro, tawa, hīnau and rewarewa, and locally narrow-leaved maire, tāwari and hard beech. Generally confined to ridges.
Warm to mild climatic zones, predominantly in eastern areas south of Auckland from the Hunua Ranges to Hapuakohe Ecological District and Mt Taupiri (Waikato Region), and Coromandel Range. Now rare in Northland, where it is predominantly restricted to drier eastern sites. Also on Hauraki Gulf islands (e.g. Te Hauturu-o-Toi / Little Barrier Island). Most areas are largely secondary or modified derivatives following fire and logging. Also present in humid areas.
Equivalent to C1–C4 types of Nicholls (1976); type d of Ecroyd (1982); and type (h) of Wardle (1984). Wardle (1991: 117) and Collins & Burns (2001).
WF13: Tawa, kohekohe, rewarewa, hīnau, podocarp forest
Podocarp, broadleaved forest of occasional emergent rimu, miro, northern rātā and locally kahikatea, with abundant tawa, kohekohe, hīnau, rewarewa and pukatea. Locally includes tāwari, kāmahi, tōwai, pūriri and mangeao, although tōwai and mangeao are locally absent or rare (e.g. Auckland and East Cape).
Inland hill country and higher ground in Northland, Hunua and Coromandel where kauri is absent. More widespread in Waikato and Bay of Plenty, with southern limits at approximately New Plymouth and Mahia.
Equivalent to D1–D3, D5, D9 and D12 types of Nicholls (1976); and alliance 23 of Wiser et al. (2011). Barton (1972).
WF14: Kāmahi, tawa, podocarp, hard beech forest
Podocarp, broadleaved, beech forest with mosaics of abundant kāmahi and tawa, occasional northern rātā, rewarewa, supplejack, rimu, miro, Hall’s tōtara, tawheowheo and tāwari, and locally abundant hard beech (generally on ridges). Also, kohekohe, pukatea, nīkau and kiekie occur at low altitude.
Predominantly coastal areas from western Herangi Range to northern Taranaki and eastern Bay of Plenty, to Raukumara-Urewera. Small areas in the South Island—western Marlborough Sounds and Wanganui Inlet. Also locally present in warm humid areas inland across the above range.
Equivalent to H4–H6 types of Nicholls (1976); and type (i) of Wardle (1984). Bayfield et al. (1991).
MF1: Tōtara, tītoki forest
Podocarp, broadleaved forest of abundant emergent tōtara, occasional mataī, kahikatea and rewarewa, with tītoki and māhoe abundant in the subcanopy, and locally maire species, ribbonwood, kōwhai and tarata. Pukatea, tawa and rimu are locally present but generally restricted to gullies.
North Island from Gisborne to Wairarapa and on the driest coastal downlands from Wanganui to Rangitikei (Esler 1978b). May have historically been present in the South Island, e.g. Marlborough to Kaikoura.
Esler (1978b), Maxwell et al. (1993— e.g. RAP1), Whaley et al. (2001) and Beadel et al. (2004—e.g. RAP 29).
MF2: Rimu, mataī, hīnau forest
Podocarp, broadleaved forest of emergent rimu and mataī, and occasional miro and tōtara, with abundant hīnau, and locally kahikatea, rewarewa, black maire, white maire and tītoki. Subcanopy species include abundant māhoe, porokaiwhiri, kaikōmako, five-finger, tarata and red māpou.
Eastern Wairarapa (e.g. Rocky Hills Forest Sanctuary, Aorangi Range), southern Rimutaka and Seaward Kaikoura Range. Rewarewa, black maire and white maire are absent from the Seaward Kaikoura Range.
Equivalent to NM1 type of McKelvey (1984); and Mataī-rimu/hīnau of Franklin (1965). Wardle (1967, 1971).
Appendix 3—continued
Continued on next page

55Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
MF3: Mataī, tōtara, kahikatea, broadleaved forest
Podocarp, broadleaved forest of two regional variants: 1. a northern type of occasional emergent mataī, tōtara and kahikatea, with broadleaf, ribbonwood, narrow-leaved houhere, tarata, māhoe, five-finger, kaikōmako and locally pōkākā; and 2. a local variant with southern rātā and rimu in humid microclimates.
Kaikoura, Canterbury (including Banks Peninsula) to Otago (Taieri River) and northern Southland. Historically would have been the main forest type present on pallic soils of the Canterbury Plains.
Equivalent to MM2 type of McKelvey (1984). Holloway (1969), Molloy (1969) and Allen et al. (2003).
MF4: Kahikatea forest
Podocarp forest of abundant kahikatea locally with mataī and a sparse subcanopy of ribbonwood and houhere species, and locally kōwhai, pōkākā, māhoe and tarata on alluvial flood plains. Ribbonwood and hohere are locally absent, while pōkākā can often be more abundant. Divaricating shrubs are a common lower understorey element.
Predominantly the eastern North Island and South Island, but also present in Northland, Waikato and Manawatu. In the South Island, south to approximately Oamaru.
Equivalent to L1 type of Nicholls (1976). Maxwell et al. (1993), Moore (1999— e.g. RAP 19: 144), McGlone (2001) and Whaley et al. (2001—RAP TIN 30: 118).
MF5: Black beech forest
Beech forest and beech, broadleaved forest of abundant black beech of at least two variants: 1. with occasional podocarp and broadleaved trees, including mataī and tōtara, and locally tītoki, hīnau, black maire, kōwhai, rewarewa, hard beech and red beech with kahikatea, kāmahi and northern rātā in sub-humid to humid areas; and 2. abundant black beech with rare podocarp and broadleaved trees.
East of the Main Divide along the North Island’s east coast to the Canterbury foothills at Mt Somers. Enters mild sub-humid areas in the Rimutaka Range and Canterbury foothills. Main type occurs in the Hunderlee Ecological District to near sea level. The two types are generally altitudinally and topographically separated, with more pure black beech occurring on steeper, infertile and dry hillslopes, especially in the Canterbury foothills south to Mt Somers. Hard beech is locally co-dominant, e.g. in the eastern Rimutaka and Tararua Ranges.
Equivalent to T1–T3 types of Nicholls (1976); type (k) of Wardle (1984); NI16 type of McKelvey (1984); and alliance 11 of Wiser et al. (2011). Franklin (1965), Wardle (1971) and Moore (1999).
MF6: Kohekohe, tawa forest
Podocarp, broadleaved forest of abundant kohekohe and frequent tawa, with occasional tītoki, māhoe, porokaiwhiri and nīkau, and scattered emergent rimu, pukatea and northern rātā. Northern rātā is locally absent in the Marlborough Sounds.
Predominantly frost-free coastal areas from Taranaki through Horowhenua to the Marlborough Sounds (although absent from between the Manawatu and Patea Rivers). Now most abundant on the Kapiti Coast, Kapiti Island and the Marlborough Sounds, and nearly extinct in south Taranaki. Historically may have occurred on the eastern Cook Strait – south Wairarapa coastline.
Equivalent to D12 type of Nicholls (1976). Bayfield & Benson (1986), Bussell (1988), Wardle (1991) and McGlone & Neall (1994).
MF7: Tawa, kāmahi, podocarp forest
Podocarp, broadleaved forest of abundant tawa and kāmahi of at least three local variants: 1. Waikato/Bay of Plenty with occasional emergent rimu, miro, kahikatea, mataī, tōtara and northern rātā, abundant tawa and kāmahi, occasional mangeao, hīnau and rewarewa, and locally pukatea in the canopy; 2. central and eastern North Island with emergent rimu, miro, kahikatea, mataī, tōtara and northern rātā, and abundant tawa, kāmahi, hīnau, rewarewa and pukatea (and locally tāwari in the north of the range on non-volcanic soils); and 3. Hunua to south Taranaki, with scattered emergent rimu, kahikatea and northern rātā, abundant tawa, pukatea and māhoe, and locally kāmahi, miro, hīnau and tawheowheo.
Downland and hill country, predominantly inland. Variant 1 occurs in inland Waikato, Bay of Plenty and King Country. Variant 2 occurs from western Raukumara southern Urewera Ranges, Wanganui, and western margin of the Tararua Range; also found east of the Main Divide on higher country, particularly in the Tiniroto Ecological District and Tararua District in humid locations; and small areas occur in the Marlborough Sounds. Variant 3 occurs in Hunua and western Waikato uplands to south Taranaki.
Equivalent to D4, D10–D11, D13–D16, M1–M5 and G1–G6 types of Nicholls (1976); ND1 type of McKelvey (1984); and alliances 21 and 22 of Wiser et al. (2011). Barton (1972).
Appendix 3—continued
Continued on next page

56Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
MF8: Kāmahi, broadleaved, podocarp forest
Podocarp, broadleaved forest of abundant kāmahi of at least two main variants: 1. with rimu, mataī, miro and tōtara, occasional hīnau, rewarewa, maire species and kahikatea, and locally Hall’s tōtara and pāhautea at higher altitudes; and 2. with abundant rimu and northern rātā, and occasional miro, hīnau, rewarewa, maire species and Hall’s tōtara at higher altitude. Early successional derivatives on recent volcanic surfaces (e.g. Mt Taranaki) include abundant māhoe, kāmahi and northern rātā.
Variant 1 occurs predominantly on the central North Island volcanic plateau, Paeroa, Hauhungaroa-Rangitoto, southern Urewera, Erua, and Hauhungatahi and Ruahine uplands, plus the Rimutaka and Haurangi Ranges. Variant 2 occurs in higher rainfall areas with moderately leached soils, including the Hauhungaroa Range, Mt Taurewa, Mt Taranaki or Mt Egmont, and southern Ruahine, Tararua, Rimutaka and Haurangi Ranges. Variant 2 is also present in cool humid areas on Mt Taranaki or Mt Egmont, Central Plateau and Tararua Range. Large areas of this forest have suffered canopy collapse post the 1950s, and are now dominated by a short forest derivative of broadleaved trees, tree ferns and scattered emergent podocarp trees.
Equivalent to L4–5, M6–M7, F2–F7, G6 and P4 types of Nicholls (1976); and alliances 12 and 18 of Wiser et al. (2011). Clarkson (1986). Includes rare ecosystem: volcanic debris flows or lahars (Williams et al. 2007).
MF9: Tānekaha forest, locally with hard beech
Podocarp forest of abundant tānekaha, with stunted tōtara, Hall’s tōtara, rimu, rewarewa, kāmahi and hard beech locally on Mamaku Plateau. On very steep sites, grades into scrub of stunted tānekaha, kānuka, Olearia furfuracea, Dracophyllum strictum, tall mingimingi, prickly mingimingi and Gaultheria spp.
Localised in ignimbrite gorges and outcrops, from southern Pureora, western Taupo to Rangitoto Range and Mamaku Plateau. Ignimbrites mapped in Molloy (1998: 34). Also, a similar secondary post-fire derivative is present from Northland to the central North Island.
Equivalent to L2–L3 types of Nicholls (1976).
MF10: Tōtara, mataī, kahikatea forest
Podocarp forest on free-draining volcanic pumice soils of abundant tōtara, mataī and kahikatea, and occasional miro and rimu, with a sparse subcanopy of broadleaved trees.
Free-draining ignimbrite (Taupo pumice) soils. Large intact examples remain within Pureora and Whirinaki Forests. Extensively logged secondary derivatives occur in Whirinaki and in the west from Pureora to Tongariro Forest.
Equivalent to L4–L5 and L7 types of Nicholls (1976). McKelvey (1963) and Morton et al. (1984).
MF11: Rimu forest Podocarp forest with abundant rimu of two regional types: 1. with occasional miro and kahikatea, and scattered tawa, hīnau and rewarewa on deep volcanic tephra; and 2. with frequent miro, mataī, kāmahi and maire species, and locally Hall’s tōtara, lowland tōtara, kahikatea and hīnau.
Variant 1 occurs in the central North Island, with the largest remaining examples occurring in Whirinaki and Pureora. Variant 2 occurs in the southeastern Hauhungaroa Range south to Ohakune. Enters cool humid and sub-humid zones.
Equivalent to L5, L6 and L8 types of Nicholls (1976). McKelvey (1963), Morton et al. (1984) and Leathwick (1987).
MF12: Rātā, hard beech, kāmahi forest
Broadleaved, beech forest of northern rātā and kāmahi, locally hard beech, occasional rimu, miro and kahikatea, and abundant nīkau, with pukatea locally abundant in the northern part of the range grading into a coastal fringe of māhoe, porokaiwhiri, Olearia avicenniifolia, shining karamū, Hebe elliptica and harakeke. A local variant without hard beech and with tānekaha, kawaka and occasional mataī occurs on old (stable) dunes.
Steep coastal hillslopes from Northwest Nelson, including Abel Tasman National Park to Punakaiki. Variant on old (stable) dunes is now rare, largely only remaining at Kaihoka Lakes, although historically may have extended north from there to southern Farewell Spit.
Equivalent to NF2 and NI15 types of McKelvey (1984). Wardle (1977, 1991: 167–168, 389, 392–393).
MF13: Kahikatea, northern rātā, kāmahi forest
Podocarp, broadleaved forest of kahikatea, northern rātā and kāmahi, and abundant nīkau and tree ferns on poor-draining alluvial Holocene floodplains. Locally, kahikatea becomes more abundant with additionally rimu on very wet organic and gley soils.
Poor-draining and flood-prone alluvial terraces in Northwest Nelson-Paparoa (e.g. Heaphy River). Historically would have also been present in small areas of older dune plains near Collingwood and Karamea.
Equivalent to NF3 type of McKelvey (1984). Jelinek (1980).
Appendix 3—continued
Continued on next page

57Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
MF14: Kahikatea, silver pine, kāmahi forest
Podocarp forest with abundant kahikatea, and occasional rimu, silver pine and kāmahi. Locally includes southern rātā, pāhautea and pōkākā, as well as scattered silver beech in South Westland.
Predominantly on post-glacial, poorly drained and flood-prone alluvial terraces and swampy sites in Westland. Small areas occur on the western volcanic ringplain of Mt Tongariro and Mt Ruapehu, although without southern rātā.
Equivalent to MF5–MF7, ML2, NL3, SL2–SL3, SM1 and NI6 types of McKelvey (1984). Wardle (1974, 1980) and Smith & Lee (1984).
MF15: Kahikatea, mataī, Westland tōtara, rimu forest
Podocarp forest with abundant kahikatea, mataī and Westland tōtara, and occasional rimu. Locally includes pāhautea and/or red beech at higher altitudes on recent, free-draining, stony soils. Early successional derivatives are dominated by ribbonwood and kaikōmako.
Holocene free-draining and flood-prone alluvial terraces in south Westland. Soils described by Molloy (1998)—e.g. Harihari soil (p. 170).
Equivalent to MM2 type of McKelvey (1984). Wardle (1974, 1977).
MF16: Rimu forest Podocarp forest with abundant rimu, and occasional miro, kāmahi, tawheowheo and southern rātā, and locally Hall’s tōtara and kahikatea.
Glacial outwash terraces of Westland (e.g. southern Okarito) and localised areas on marine terraces with gley soils (e.g. Waitutu). Glacial outwash soils described by Molloy (1998: 19).
Equivalent to ML1, NL1, NI5, MF6 and MF8 types of McKelvey (1984).
MF17: Rimu, kāmahi, tāwheowheo forest
Podocarp, broadleaved forest of rimu and Hall’s tōtara, locally with kāmahi, tawheowheo, mountain celery pine, pāhautea, silver pine and pōkākā. Locally also includes occasional beech (hard, silver and mountain) outside the Westland beech gap.
Strongly leached glacial outwash terraces with well-developed podzols (soils described by Molloy (1998: 19)), or infertile thin soils on quartzose sandstones and coal measures from Northwest Nelson (e.g. Ngakawau to south Westland). Rimu and other trees are predominantly stunted.
Equivalent to ML5, NL2, NI8 and NI10 types of McKelvey (1984). Wardle (1980, 1991: 128).
MF18: Yellow silver pine, mānuka forest
Podocarp short forest of abundant yellow silver pine, locally with mānuka, pink pine, silver pine, pāhautea and rimu, as well as mountain beech or hard beech outside the Westland beech gap.
Associated with very strongly leached and/or low-fertility parent materials from Northwest Nelson (e.g. Knuckle Hill) to Fiordland. In Westland, on glacial outwash terraces (soils described by Molloy (1988: 19)). In South Westland and Fiordland (e.g. Secretary, Resolution and Coal Islands), on roche moutonnée on gneiss and granite. Enters cool, humid areas in south Westland and Fiordland.
Equivalent to ML6 type of McKelvey (1984). Wardle (1963, 1977, 1991: 127) and Wardle et al. (1986).
MF19: Kāmahi, rimu, miro, southern rātā forest
Podocarp, broadleaved forest of abundant kāmahi, with emergent rimu and miro, occasional southern rātā, and locally Hall’s tōtara and tawheowheo. Often broadleaved trees are dominant on steep sites.
Widespread on hillslopes throughout central Westland. Also from Karamea to western Paparoa Range.
Equivalent to MF1–MF4, ML3–ML4 and NF1 types of McKelvey (1984); and alliances 12, 18 and 20 of Wiser et al. (2011). Wardle (1980, 1991: 127).
MF20: Hard beech forest
Beech forest and beech, podocarp, broadleaved forest of at least three local variants, all with abundant hard beech: 1. northern, with occasional rimu, miro, Hall’s tōtara, tāwari, northern rātā, tānekaha, toatoa, tawheowheo, kāmahi and rewarewa; 2. central, with black beech, and locally kāmahi, rimu, northern rātā, hīnau and rewarewa; and 3. southern, with occasional rimu, miro, Hall’s tōtara, southern rātā and tawheowheo, and locally kāmahi, and black, red and silver beech.
Northern variant in Taranaki and eastern Bay of Plenty – northern Hawke's Bay. Central variant in Rimutaka, Tararua and Akatarawa Ranges. Southern variant most abundant in the South Island, from Northwest Nelson to Paparoa Range (e.g. Oparara River, Punakaiki).
Equivalent to I9–I14 types of Nicholls (1976); type J of Wardle (1984); and NI1–NI2, NI13–NI14 and NK3 types of McKelvey (1984). Druce & Atkinson (1958), Franklin (1965) and Jelinek (1980).
MF21: Tawa, kāmahi, rimu, northern rātā, black beech forest
Podocarp, broadleaved, beech forest with abundant tawa and kāmahi, and occasional rimu, northern rātā, kahikatea, hīnau, maire species and rewarewa, with pukatea on warmer sites, and black and/or hard beech locally abundant on ridges. Tawheowheo and tānekaha are present in northern Taranaki.
Inland Whanganui–Taranaki, especially Matemateaonga Range and north Taranaki. Black beech is dominant in the south in the Matemateaonga Ecological District and is replaced by or locally co-dominant with hard beech in the North Taranaki Ecological District.
Equivalent to I1, H1–H2 and H4 types of Nicholls (1976); and alliance 19 of Wiser et al. (2011). Fuller & Edwards (1989) and Bayfield et al. (1991).
Appendix 3—continued
Continued on next page

58Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
MF22: Tawa, rimu, northern rātā, beech forest
Podocarp, broadleaved, beech forest with abundant tawa, occasional rimu and northern rātā, and locally abundant beech on ridges (mostly red, hard, black and locally silver beech). Locally also includes toatoa, Hall’s tōtara, miro, hīnau and rewarewa.
Eastern Bay of Plenty, Raukumara Range / Gisborne hill country (Matawai) to Urewera ranges and northern Hawke’s Bay. Similar type in Marlborough Sounds, but northern rātā is absent.
Equivalent to H3 and H7–H8 types of Nicholls (1976); NH1 type of McKelvey (1984); and alliance 19 of Wiser et al. (2011).
MF23: Chatham Island akeake, karamū, māhoe, ribbonwood forest
Mixed broadleaved forest of Olearia traversiorum, Myrsine chathamica, Pseudopanax chathamicum, Coprosma chathamica and Melicytus chathamica, and locally tarahinau, Hebe barkeri and Brachyglottis huntii, with Cyathea and Dicksonia tree ferns on hillslopes. Abundant nīkau and ribbonwood also occur on more fertile soils (e.g. Pitt Island).
Chatham Islands. Largely secondary derivatives referred to as ‘tree heath’, dominated by tarahinau. Localised kōwhai and karaka of anthropogenic origin (Heenan et al. 2001; Stowe 2003).
Cockayne (1901), Kelly (1983), Wardle (1991: 436–441) and Roberts et al. (2007). Locally includes rare ecosystem: seabird burrowed soil (Williams et al. 2007).
MF24: Rimu, tōwai forest
Podocarp, broadleaved forest with occasional emergent rimu, miro and northern rātā, abundant tōwai, locally tawa and swamp maire, and occasional hīnau, rewarewa, tāwari, pukatea, mangeao, raukawa, narrow-leaved maire, makamaka and hutu.
Locally above 450 m on the Northland, Hunua and Coromandel ranges. Swamp maire is locally abundant on poorly drained soils on plateaus above 500 m in Tutamoe Range, Waima Forest and Mataraua Forest (Northland).
Equivalent to D6–D8 and F1 types of Nicholls (1976); and D6 and F2 types of McKelvey & Nicholls (1959). Wardle (1991) and Conning (2001). Includes rare ecosystem: cloud forest (Williams et al. 2007).
MF25: Kauri, tōwai, rātā, montane podocarp forest
Kauri, podocarp, broadleaved short forest of kauri, yellow silver pine, rimu, Kirk’s pine and toatoa, and locally Hall’s tōtara, tāwari, hīnau, tōwai, southern and Parkinson’s rātā, and tawheowheo.
Summits of northern Kaimai and Coromandel Ranges (e.g. Moehau), Te Hauturu-o-Toi / Little Barrier Island (> 600 m), and Mt Hobson, Great Barrier Island (Aotea Island).
Equivalent to B10–B12 and G7 types of Nicholls (1976); and type g of Ecroyd (1982). Conning (2001). Locally includes rare ecosystems: seabird burrowed soil (e.g. Te Hauturu-o-Toi / Little Barrier Island); and cloud forest (Williams et al. 2007).
CLF1: Hall’s tōtara, mountain celery pine, broadleaf forest
Podocarp, broadleaved forest with Hall’s tōtara, mountain celery pine and broadleaf. Locally includes mataī, kōwhai, tarata and five-finger at lower altitudes, as well as southern rātā in more humid locations (e.g. Mt Peel, south Canterbury).
Inland mid-slope/foothill forests from Marlborough to the western Canterbury foothills. Descends to lower altitude at the Seaward Kaikoura Range, with lowland podocarp trees present. Very small remnants still present in the dry interior of Otago, where it would have historically been common.
Equivalent to MG5 and NG1 types of McKelvey (1984). Wardle (1971), Williams (1989) and McGlone & Basher (1995).
CLF2: Hall’s tōtara forest [Dune forest]
Podocarp forest of abundant tōtara (Hall’s and lowland), locally with mataī on free-draining sandy soils.
Stable dunes in Southland, from The Catlins to Invercargill (e.g. Otatara dunes) and south Westland. Very limited areas remain. Very old dunes include broadleaved trees (pōkākā and kāmahi), rimu and kahikatea.
Norton (1996) and Harding (1999). Includes rare ecosystem: stable dunes (Williams et al. 2007).
CLF3: Podocarp, ribbonwood, kōwhai forest
Podocarp forest of abundant kahikatea, mataī and tōtara, with ribbonwood, narrow-leaved houhere, kōwhai and a wide variety of divaricating shrubs on free-draining soils.
In the North Island, largely restricted to inland areas south of the volcanic plateau, including Rangitikei District (e.g. Mataroa) and inland Hawke’s Bay (e.g. Balls Clearing). Historically more widespread in Otago and Southland, although now very rare.
Equivalent to L1 type of Nicholls (1976). Lake & Whaley (1995), Harding (1999), McGlone (2001) and Allen et al. (2003: 164).
Appendix 3—continued
Continued on next page

59Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
CLF4: Kahikatea, tōtara, mataī forest
Podocarp forest of two variants: 1. northern, with emergent kahikatea, mataī and tōtara, and occasional maire species, kōwhai and ribbonwood; and 2. southern, with occasional to common emergent mataī and tōtara, and locally Hall’s tōtara with narrow-leaved houhere, ribbonwood, kōwhai, māhoe, broadleaf and tarata. Locally includes rimu in the most humid part of the range, and ngaio and Olearia spp. in coastal Otago.
In the North Island, largely restricted to inland areas south of the volcanic plateau, including Rangitikei District (e.g. Titirangi Scenic Reserve) and inland Hawke’s Bay. In the South Island, occurs from western Southland to Otago (Moeraki) in areas with < 1000 mm of rain per annum (e.g. Hokonui Hills). Now very uncommon and only predominantly secondary derivatives remain.
Equivalent to L9 and L11 of Nicholls (1976); and SM2 type of McKelvey (1984). McGlone (2001) and Allen et al. (2003: 155–156).
CLF5: Mataī, Hall’s tōtara, kāmahi forest
Podocarp forest of abundant mataī, with occasional or local kahikatea, Hall’s tōtara, miro, tōtara, maire species, kāmahi and pōkākā, and locally (on the coldest, frost-prone sites) pāhautea, mountain celery pine and silver pine.
Central North Island, Kaingaroa Plateau (e.g. Rangitaiki), and in the west from Pureora to Ohakune, within cold air inversion basins and cool mid-altitude sites that are generally too cold for rimu. Rare examples also occur elsewhere, e.g. RAP 8 northern Wairarapa Ecological District (Sawyer et al. 1998).
Equivalent to L7 and L10–L12 types of Nicholls (1976). McKelvey (1963) and Sawyer et al. (1998).
CLF6: Kāmahi, southern rātā, podocarp forest
Podocarp, broadleaved forest of several variants: 1. abundant rimu, kāmahi and southern rātā, with occasional miro and Hall’s tōtara; 2. locally with pink pine, mountain celery pine and yellow silver pine at higher altitudes on exposed, infertile sites; 3. with mataī and kahikatea on alluvial sites; and 4. broadleaved-dominant, with abundant kāmahi and southern rātā on coastal margins and at higher altitudes.
In Southland hillslopes north to Hokonui Hills, but now uncommon. Most abundant in The Catlins and on Stewart Island/Rakiura. Southern rātā extends as a pure monoculture on the Auckland Islands on organic soils.
Equivalent to SF1–SF3 and SL4–SL5 types of McKelvey (1984); communities 1 and 2 of Wilson (1987: 10–18); and alliance 16 of Wiser et al. (2011).
CLF7: Rimu, kāmahi, beech forest
Podocarp, broadleaved, beech forest of abundant rimu, silver beech and kāmahi, with occasional Hall’s tōtara and miro, and locally kahikatea and southern rātā.
Fiordland and south Westland on poorly drained, infertile soils at lower altitudes (e.g. Waitutu). Mountain beech, mountain celery pine, yellow silver pine, silver pine and pink pine are abundant on wetland margins.
Equivalent to SI8–SI9 types of McKelvey (1984). Wardle (1963), Mark & Smith (1975) and Mark et al. (1988).
CLF8: Silver beech, kāmahi, southern rātā forest
Broadleaved-beech forest of silver beech, southern rātā and kāmahi, with Hall’s tōtara, and locally mountain beech, tawheowheo and red beech at lower altitudes.
Mid-slope forest that only occurs in the western South Island—Northwest Nelson to Fiordland.
Equivalent to type e of Wardle (1984); and NK4 type of McKelvey (1984).
CLF9: Red beech, podocarp forest
Beech, podocarp, broadleaved forest of abundant red beech of at least two types: 1. hillslope forest locally with kāmahi in humid parts of the range, occasional rimu, Hall’s tōtara, miro and mataī, and with hīnau and maire species in northern areas; and 2. on alluvial stony terraces, red beech locally with silver beech, kāmahi and southern rātā, and occasional podocarp trees (rimu, kahikatea, mataī and tōtara).
Extensive in the North Island and South Island, primarily in western districts on hillslopes and alluvial stony soils. Urewera to Ruahine ranges, Nelson, Westland and Southland. Enters sub-humid regions in the North Island.
Equivalent to I15–I17, K1–K3 and K8–K9 types of Nicholls (1976); type f of Wardle (1984); and NK11 type of McKelvey (1984).
Appendix 3—continued
Continued on next page

60Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
CLF10: Red beech, silver beech forest
Beech forest and beech, podocarp, broadleaved forest of abundant red and silver beech, and locally with podocarp and broadleaved species, with at least three local variants: 1. Raukumara–Urewera ranges, with tāwari, locally tawheowheo, kāmahi and Hall’s tōtara, and scattered rimu and miro at lower altitudes; 2. North Island and South Island locally with occasional black beech and mountain beech, Hall’s tōtara, pāhautea, kāmahi and hard beech, as well as rimu, miro and mataī at lower altitudes; and 3. with occasional black beech and mountain beech.
Variant 1 occurs on the mid-slopes of the North Island main axial ranges, from Raukumara to Akatarawa. Variant 2 occurs in the northwestern South Island and in scattered areas further south. Variant 3 occurs east of the Main Divide, from northern Southland to Marlborough (excluding beech gap zone). Enters the mild and high rainfall zone in south Westland, and the cool sub-humid zone east of the Main Divide.
Equivalent to I18–I21, J2–J13 and K4–K7 types of Nicholls (1976); NK8–9, NJ34, NK1–NK2, NK8–NK10, NI3–NI4, NI12, NJ3, SK4 and SK10 types of McKelvey (1984); type d of Wardle (1984); and alliances 10, 15 and 17 of Wiser et al. (2011). Smith & Lee (1984).
CLF11: Silver beech forest
Beech forest of abundant silver beech of at least three local variants: 1. northern, with tāwari, tawheowheo and kāmahi; 2. upper mountain slopes, locally with mountain celery pine, three-finger, Olearia spp., kōtukutuku, broadleaf and small-leaved shrubs; and 3. lower and mid-slopes, with rimu and kāmahi, occasional southern rātā, Hall’s tōtara and miro, and locally with kahikatea on alluvial terraces. Also present at Waitaanga (north Taranaki) in the mild-humid zone, with rimu, kahikatea, miro, Hall’s tōtara, tawa, kāmahi, northern rātā, hīnau and rewarewa (Nicholls 1976: O3 type).
Variant 1 occurs in the Kaimai and Raukumara–Urewera ranges. Variant 2 is widespread in humid regions, including the Raukumara and Tararua Ranges, and in the South Island from Northwest Nelson to Fiordland. Variant 3 occurs in the southern Rimutaka Range and southwestern South Island. Variants 2 and 3 are largely distributed according to altitude and topography. Stable sites develop organic and podzols soils (Molloy 1998: 130, plate 8.6).
Equivalent to alliances 7, 13 and 14 of Wiser et al. (2011); northern c type, upper slopes b type and southern g type of Wardle (1984); I23, I26, J12–J16 and K10–K14 types of Nicholls (1976); and NI11, NK5, SI3, SI5, SI7 and SK1–SK3 types of McKelvey (1984). Franklin (1965). Locally includes rare ecosystem: cloud forest (Williams et al. 2007).
CLF12: Silver beech, mountain beech forest
Beech forest of abundant silver and mountain beech, locally with kāmahi, Hall’s tōtara, mountain celery pine, red beech, three-finger, kōtukutuku, broadleaf and small-leaved divaricating shrubs.
Predominantly transitional between cool humid and cool sub-humid climatic zones. Occurs in the central North Island (e.g. Kaimanawa Mountains) and in the South Island east of the Main Divide, from Nelson (e.g. Nelson Lakes) to Fiordland (e.g. Waikaia and Eyre Mountains). Also occurs in Buller, Northwest Nelson.
Equivalent to K14 type of Nicholls (1976); SK5–SK6 and NK7 types of McKelvey (1984); and alliances 9 and 11 of Wiser et al. (2011).
CDF1: Pāhautea, Hall’s tōtara, mountain celery pine, broadleaf forest
Podocarp, broadleaved forest of pāhautea, Hall’s tōtara, mountain celery pine and broadleaf.
Montane forests in the beech gap of the eastern flanks of the Southern Alps/Kā Tiritiri o te Moana, from the Waimakariri headwaters to Aoraki/Mt Cook and outliers further south (e.g. east Matukituki valley, Mt Aspiring National Park and upper Waipara River). Also small areas on Banks Peninsula, eastern Otago uplands and The Catlins.
Equivalent to MG3–MG4 types of McKelvey (1984). Allen et al. (2003: 163).
CDF2: Dracophyllum, mountain celery pine, Olearia, Hebe scrub [Subalpine scrub]
Short forest, scrub of a wide range of local variants, with a range of xerophyllus species including species of Dracophyllum (e.g. D. acerosum, D. rosmarinifolium), Phyllocladus, Ozothamnus, Olearia, Brachyglottis, Ozothamnus, Hebe, Carmichaelia, Brachyglottis and Coprosma, and locally snow tōtara and wharariki.
Most abundant in the eastern South Island beech gap, subalpine regions of Marlborough–Kaikoura, Canterbury and Central Otago. Restricted to disturbed sites where beech forest is present. The very few primary examples that remain are largely restricted to fire refugia.
Equivalent to B.7 type of Williams (1989); and alliance 3 of Wiser et al. (2011). Wardle (1971) and Wardle (1991: 188–190).
Appendix 3—continued
Continued on next page

61Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
CDF3: Mountain beech forest
Beech forest of abundant mountain beech, with small-leaved Coprosma spp., weeping matipo, mountain celery pine, snow tōtara, broadleaf, three-finger and putaputawētā, and locally Hall’s tōtara. Locally also includes scattered silver and red beech in humid locations.
Widespread in the North Island and South Island at altitudes above 700 m, predominantly east of the Main Divide, in cool and cold climatic regions. In the North Island, occurs in the Kaimanawa, Kaweka, Mt Ruapehu and Ruahine uplands. In the South Island, very widespread, but most abundant from Nelson to Canterbury and south Canterbury to Fiordland. Where it enters cool humid areas, large transitional zones occur, often with mixes of mountain, silver and red beech.
Equivalent to K15–K17 types of Nicholls (1976); type a of Wardle (1984); SK9, NK6 and NK12 types of McKelvey (1984); and alliances 8 and 11 of Wiser et al. (2011).
CDF4: Hall’s tōtara, pāhautea, kāmahi forest
Podocarp, broadleaved forest of at least four main variants: 1. with abundant Hall’s tōtara, kāmahi and broadleaf, and locally mountain celery pine, pāhautea, tawheowheo, tāwari and miro, as well as maire species and pōkākā at lower altitudes; 2. abundant pink pine, with mountain celery pine and broadleaf, and locally Hall’s tōtara; 3. abundant pāhautea, mountain celery pine and pink pine, and locally Hall’s tōtara, broadleaf, silver pine and bog pine; and 4. abundant pink pine, with pāhautea, yellow silver pine and tāwari.
Upland areas in the beech gap. Variant 1 occurs in western areas, predominantly at altitudes between 800 and 1200 m, from Mt Pirongia to Mt Taranaki or Mt Egmont, and in the western-central North Island from Mt Pureora to Mt Taurewa, and southern Ruahine and northern Tararua Ranges; enters mild and humid zone from Herangi northwards. Variant 2 occurs on subalpine hill tops from the northern end of the Raukumara Range and Urewera, Ahimanawa, northern Ruahine and northern Tararua Ranges. Variant 3 occurs in western Ruapehu (e.g. Mt Hauhungatahi). Variant 4 occurs in the Kaimai Range (e.g. Te Hunga Ecological Area).
Equivalent to L13, G8 and G16–G20 types of Nicholls (1976). McKelvey (1973), Atkinson (1981) and Clarkson (1986, 1990). Locally includes rare ecosystems: cloud forest; and early successional examples include recent lava flows (< 1000 years) and young tephra (< 500 years) on plains and hill slopes (e.g. Mt Tarawera) (Williams et al. 2007).
CDF5: Hall’s tōtara, pāhautea, kāmahi, southern rātā forest
Broadleaved forest of abundant kāmahi and southern rātā, occasional Hall’s tōtara, and locally pāhautea, pink pine, tawheowheo, broadleaf, Olearia lacunosa, mountain neinei and three-finger.
Western side of the Main Divide in the beech gap. Kāmahi is absent from areas above 840 m a.s.l.
Equivalent to MG1–MG2 and MG6–MG7 types of McKelvey (1984). Rose et al. (1992), Stewart & Harrison (1987) and Wardle (1991: 134–136).
CDF6: Olearia, Pseudopanax, Dracophyllum scrub [Subalpine scrub]
Short forest, scrub of wide range of local variants, with a range of species of Olearia, Brachyglottis, Pseudopanax, Dracophyllum, Hebe, Coprosma, Hoheria, montane podocarp trees, mānuka and wharariki. Locally includes monocultures, such as leatherwood scrub in southern Ruahine – northern Tararua.
Most abundant and extensive in humid montane–subalpine regions in the beech gap, central North Island, Mt Taranaki or Mt Egmont, southern Ruahine – northern Tararua and in the central South Island beech gap, including Canterbury mountains east of the Main Divide, Stewart Island/Rakiura and subantarctic islands. More restricted abundance where beech forest is present.
Equivalent to S3 Sub-alpine scrub and FS7 Sub-alpine scrub and indigenous forest of Newsome (1987); and alliance 1 of Wiser et al. (2011). Cockayne (1909a, b), Wilson (1976), Clarkson (1986), Stewart & Harrison (1987) and Allan et al. (2003: 161 & 169).
CDF7: Mountain beech, silver beech, montane podocarp forest
Beech, podocarp and beech, podocarp, broadleaved forest of at least two variants: 1. North Island, with stunted mountain beech and/or silver beech, locally with montane podocarp trees (Hall’s tōtara, pāhautea, and pink, bog and silver pine), and South Island with mountain/silver beech, yellow silver pine, southern rātā and Dracophyllum traversii; and 2. lower altitude mountain/silver beech, and locally yellow silver pine, silver pine, rimu, kahikatea, miro, pāhautea, Hall’s tōtara and pōkākā.
In the North Island, restricted to high-altitude sites in the Raukumara, Ahimanawa and Ruahine Ranges, and on Mt Ruapehu. In the South Island, occurs at lower altitude sites where rimu and kahikatea can be abundant in Northwest Nelson, Fiordland (including Waitutu) and The Catlins. Stable sites develop organic and podzol soils (Molloy 1998: 130, plate 8.6; Mark et al. 1988).
Equivalent to J1 and J14–J18 types of Nicholls (1976); type l of Wardle (1984); and NI7, NI9, NJ2, SI2, SI4–SI7, SK2 and SK5–SK8 types of McKelvey (1984). Mark et al. (1988).
Appendix 3—continued
Continued on next page

62Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
AL1: Narrow-leaved and slim snow tussock tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa rigida subsp. rigida and C. macra, and species of Hebe and Dracophyllum, with areas of talus, boulderfield and bluffs. At least two regional variants: 1. Eastern/Central Otago mountains; and 2. eastern Alps.
Eastern semi-arid mountains of the South Island, from Rakaia River south, and dominant in the central and eastern Otago mountains, from the treeline to approximately 2000 m a.s.l. Western margin along the Southern Alps/Kā Tiritiri o te Moana corresponds to the boundary of Chionochloa macra (Meurk 1982).
Connor & Macrae (1969), Meurk (1982), Grove (1994) and Mark et al. (2003: 199–200). Includes rare ecosystems: screes of acidic rocks; and cliffs, scarps and tors of acidic rocks (Williams et al. 2007).
AL2: Slim and mid-ribbed snow tussock tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa macra and/or C. pallens, and species of Hebe, Brachyglottis and Dracophyllum, with areas of talus, boulderfield and bluffs. At least two regional variants: 1. northern Alps, with C. oreophila and/or C. australis at high altitudes close to the Main Divide; and 2. Kaikoura, with C. flavescens subsp. brevis, bristle tussock and blue tussock.
Eastern semi-arid mountains of the South Island, from Rakaia River north to the Marlborough and Kaikoura ranges. In Kaikoura, very little remaining in an unmodified state (Williams 1989).
Wraight (1963), Williams (1989) and Wardle (1991: 241–243). Includes rare ecosystems: screes of acidic rocks; cliffs, scarps and tors of acidic rocks; and basic cliffs, scarps and tors (Williams et al. 2007).
AL3: Red tussock tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa rubra, and species of Hebe and Dracophyllum. Locally includes C. pallens on the main axial ranges of the Kaimanawa Mountains and northern Ruahine Range.
Most abundant on the volcanic plateau on mountains of Tongariro National Park and Mt Taranaki or Mt Egmont. Also present in southern Kaimanawa Mountains and northern Ruahine Range on volcanic tephra.
Atkinson (1981), Clarkson (1986) and Wardle (1991: 244). Includes rare ecosystem: volcanic debris flows or lahars (Williams et al. 2007).
AL4: Mid-ribbed and broad-leaved snow tussock tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa pallens subsp. pallens, and species of Hebe and Dracophyllum, with areas of talus, boulderfield and bluffs. Locally includes C. rubra in the Kaimanawa Mountains and Ruahine Range, and C. flavescens subsp. flavescens in the Tararua Range.
North Island axial mountain ranges, with several regional variants from East Cape (e.g. Hikurangi to Rimutaka Range).
Williams (1975), Connor (1991) and Wardle (1991: 243). Includes rare ecosystems: screes of acidic rocks; and cliffs, scarps and tors of acidic rocks (Williams et al. 2007).
AL5: Mid-ribbed, broad-leaved, red and carpet grass tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa pallens and C. flavescens (subsp. lupeola and subsp. hirta), and locally C. rubra subsp. occulta and C. australis, and species of Hebe and Dracophyllum, with areas of talus, boulderfield and bluffs.
Northwest Nelson to Arthur’s Pass. Several local variants occur on different parent materials, with Chionochloa flavescens abundant on marble, and C. pallens and C. australis abundant on schist and granite (Bell 1973).
Bell (1973), Druce et al. (1987), Connor (1991), Wardle (1991: 223–226) and Williams (1991, 1993). Includes rare ecosystems: screes of acidic rocks; and cliffs, scarps and tors of acidic rocks (Williams et al. 2007).
AL6: Mid-ribbed and narrow-leaved snow tussock tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa pallens subsp. pilosa, locally with C. rigida subsp. amara and C. crassiuscula, and species of Hebe and Dracophyllum, with areas of talus, boulderfield and bluffs.
South of Arthur’s Pass, Westland, to northern Fiordland. Evans (1969b), Burrows (1977) and Cuddihy (1983). Includes rare ecosystems: screes of acidic rocks; and cliffs, scarps and tors of acidic rocks (Williams et al. 2007).
AL7: Pungent snow tussock tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa crassiuscula, with C. pallens and C. rigida subsp. amara, and species of Hebe and Dracophyllum, with areas of talus, boulderfield and bluffs. Locally includes C. teretifolia, C. acicularis, C. ovata and C. spiralis, and associated herbfield.
Restricted to Fiordland, Takitimu Mountains and northern Stewart Island/Rakiura, from northern Fiordland south.
Evans (1972) and Mark et al. (2003, 2008: 214–216). Includes rare ecosystems: screes of acidic rocks; and cliffs, scarps and tors of acidic rocks (Williams et al. 2007).
Appendix 3—continued
Continued on next page

63Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
AL8: Stewart Island snow tussock tussockland/shrubland
Tall tussock grassland, shrubland of abundant Chionochloa lanea and C. crassiuscula, and stunted mānuka, Olearia colensoi var. argentea and pink pine, with areas of rock pavement and bluffs.
Mt Rakeahua to Tin Range, Stewart Island/Rakiura. Wilson (1987) and Mark et al. (2003: 217).
AL9: Subantarctic snow tussock tussockland/shrubland
Tall tussock grassland, herbfield, shrubland and low scrub, with a mosaic of abundant Chionochloa antarctica, Poa foliosa and mega-herbs (species of Anisotome, Pleurophyllum and Stilbocarpa). Locally seabird nesting sites, cushionfield, sedgeland bogs (Centrolepis pallida, Carpha alpina, Oreobolus pectinatus) and areas of higher altitude basalt talus dominated by lichens, Grammitis poeppigiana, Stellaria decipiens and other small herbs, turf grasses and mosses also occur.
Subantarctic islands, especially Auckland and Campbell Island groups. Cockayne (1909a), Wardle (1991: 453–455 & 459–460) and Meurk et al. (1994). Includes rare ecosystems: seabird guano deposits; and seabird burrowed soil (Williams et al. 2007).
AH1: Gravelfield/stonefield [Fellfield]
Gravelfield/stonefield of at least two regional types (eastern Southern Alps/Kā Tiritiri o te Moana and Kaikoura), with a sparse covering of vegetable sheep, sub-shrubs (Hebe spp.), Celmisia and other herbs, with extensive areas of rock pavement, boulderfield and bluffs, and limited areas of snow banks, cushionfield and herbfield.
Restricted to greywacke mountains within the sub-humid zone of the Southern Alps/Kā Tiritiri o te Moana and Kaikoura mountains, south to approximately Ben Ohau Range in North Otago; then, in the extreme south, in the Takitimu, Eyre and Livingstone Mountains.
Wardle (1991: 416–418 & 425–429), Mark & Dickinson (1997) and Mark et al. (2003: 207 & 210). Includes rare ecosystems: screes of acidic rocks; cliffs, scarps and tors of acidic rocks; and snow banks (Williams et al. 2007).
AH2: Dracophyllum muscoides cushionfield
Cushionfield (with smaller areas of fellfield and rock tors) of prostrate shrubs and herbs dominated by Dracophyllum muscoides, and other cushion-/mat-forming sub-shrubs, herbs and snow banks dominated by Celmisia haastii and associates, on gently rolling plateaus.
Eastern and Central Otago mountains. In some locations, grazing and burning have led to cushionfield becoming more abundant, replacing tall tussock grasslands.
Mark & Bliss (1970), Wardle (1991: 419–427), Grove (1994), Dickinson et al. (1998) and Mark et al. (2008: 207). Includes rare ecosystem: snow banks (Williams et al. 2007).
AH3: Gravelfield/stonefield, mixed species cushionfield
Gravelfield/stonefield with areas of rock pavement, talus, boulderfield and bluffs, and locally cushionfield, herbfield and snow banks. At least two regional types (western Nelson and western Alps – northern Fiordland), which include a diversity of grasses, small herbs and sub-shrubs, including Aciphylla, Agrostis, Brachyscome, Brachyglottis, Celmisia, Chionochloa australis, Dolichoglottis, Epilobium, Gaultheria, Gentianella, Hebe, Ourisia, Poa, Ranunculus, Trisetum and cushion genera. Includes snow banks of Chionochloa oreophila, C. crassiuscula, C. vireta, blue tussock and Celmisia hectorii.
Most prominent on the highest mountain ranges, with a range of regional variants in relation to parent material. In the North Island, very restricted in distribution to the highest and most exposed tops of the Kaimanawa Mountains, and Ruahine and Tararua Ranges. In the South Island, western Southern Alps/Kā Tiritiri o te Moana, from Northwest Nelson to Fiordland. Small areas occur on the highest peaks of the Auckland Islands.
Wardle (1991: 229–230, 418–419 & 429–430), Mark & Dickinson (1997) and Mark et al. (2003: 211–213, 2008: 207). Includes rare ecosystems: granite gravelfields; granite sand plains; and snow banks (Williams et al. 2007).
AH4: Woolly moss, bristle tussock, blue tussock mossfield/tussockland/stonefield
Stonefield with a sparse covering of woolly moss, bristle tussock, blue tussock, species of Gaultheria and Parahebe, and kopoti, and locally mountain tōtara and Dracophyllum recurvum.
Restricted to North Island volcanic mountains of Tongariro National Park and Mt Taranaki or Mt Egmont. Also occurs largely as a secondary derivative below the natural treeline in and adjacent to Tongariro National Park on raw volcanic soils, particularly on the eastern side, where fire and erosion have extended its range.
Scott (1977), Atkinson (1981), Clarkson (1986, 1990) and Wardle (1991: 430–431). Includes rare ecosystems: recent lava flows; recent volcanic boulderfields and volcanic debris flows or lahars (Williams et al. 2007).
Appendix 3—continued
Continued on next page

64Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
TI1: Bog pine, mountain celery pine scrub/forest
Short forest, scrub of abundant bog pine and/or mountain celery pine, with species of Dracophyllum, Leucopogon, Coprosma, Hebe, Olearia, Pittosporum, Gaultheria and Pimelea, mountain tauhinu, korokia, mountain wineberry, snow tōtara and porcupine shrub. Early successional derivatives include short tussock grasslands of species of Poa, Festuca, Deyeuxia, Rytidosperma, with inter-tussock prostrate herbfield species.
Moraines and inter-montane basins of the eastern South Island dry zone, especially Otago, south Canterbury and west Marlborough (e.g. The Wilderness and Bendhu Scientific Reserves). Soils are acidic, infertile and can be seasonally waterlogged, and have been described by Molloy (1998: 150–152). Extends into sub-humid zones in tributary valleys.
Shanks et al. (1990—e.g. Cass RAP 22), Wardle (1991: 194–195), Courtney & Arand (1994—e.g. Sedgemere RAP 1), McGlone (2001) and Allen et al. (2003: 169). Includes rare ecosystems: strongly leached terraces and plains; and inland outwash gravels (Williams et al. 2007).
TI2: Kānuka, Olearia scrub/treeland
Scrub and treeland of kānuka, and species of Olearia, Carmichaelia and Melicytus, with korokia, matagouri, mountain tauhinu, prostrate kōwhai and lianes (e.g. Muehlenbeckia), and locally Leonohebe cupressiodes. Early successional derivatives include short tussock grasslands of species of Poa, Festuca, Deyeuxia and Rytidosperma, with inter-tussock prostrate herbfield species.
Driest inter-montane basins on Holocene terraces of Marlborough to Otago (e.g. Mackenzie and Heron Basins) with > 200 mm annual water deficit (Leathwick et al. 2003). Soils described by Molloy (1998: 24–25 & 157–158). Largely only highly degraded secondary derivatives remain, which are dominated by prostrate shrubland, short tussock grassland and scabweed herbfield.
McGlone & Moar (1998), McGlone (2001) and Rogers et al. (2005: 53). Includes rare ecosystem: frost hollows (Williams et al. 2007).
TI3: Monoao scrub/lichenfield
Scrub of abundant monoao and lichens, and occasional silver tussock and Pimelea prostrata. Locally with ecotone margins of mountain celery pine, bog pine and mānuka. Likely to have included mountain celery pine and bog pine in the least disturbed older successional examples, though these are now rare. Early successional derivatives include short tussock grasslands of species of Poa, Festuca, Deyeuxia and Rytidosperma, with inter-tussock prostrate herbfield species.
Kaingaroa Plateau, Kaimanawa Mountains and Hauhungaroa Range. Soils described by Molloy (1998: 38–44). Enters the humid zone in central ranges.
Smale (1990) and Wardle (1991: 193–194). Includes rare ecosystem: old tephra plains (Williams et al. 2007).
TI4: Coprosma, Olearia scrub [Grey scrub]
Scrub of two different variants: 1. on free-draining stony soils, with species including Carmichaelia, Coprosma, Olearia, Hebe, Corokia cotoneaster, mānuka, matagouri, and species of the lianes Muehlenbeckia, Rubus and Clematis; and 2. on poor-draining silty soils, with species such as Coprosma (C. propinqua, C. pedicillata), Pittosporum obcordatum and Olearia (O. polita, O. virgata). Early alluvial successions are dominated by short tussock grasslands (species of Poa, Festuca, Deyeuxia and Rytidosperma).
In the South Island, post-glacial river terraces, colluvial slopes and recent moraines west and east of the Main Divide, and in frost-prone areas further afield. In the North Island, localised in frosty hollows and river and stream margins. Often colloquially called ‘grey scrub’ or ‘frost flats’. Many species are common to both types and both types can occur together as mosaics on the different alluvial landforms/soils. Often occurs as secondary shrubland on a wide range of previously forested landforms. Enters the humid zone in central ranges.
Equivalent to Matagouri shrubland class of Wiser & Hurst (2008). Wardle (1991: 207–211), Clarkson & Clarkson (1994), Molloy et al. (1999) and Rogers et al. (2005). Includes rare ecosystem: frost hollows (Williams et al. 2007).
TI5: Bog pine, mountain celery pine, silver pine scrub/forest
Scrub and short forest with several local variants, including mountain celery pine and bog pine, locally with silver pine, pink pine, yellow silver pine, pāhautea and Westland tōtara, and often with divaricating shrubs and Dracophyllum spp.
Central North Island (e.g. Kaimanawa Mountains) and western Tongariro National Park (e.g. Waimarino Plains). In the South Island, predominantly Nelson Lakes to Westland (e.g. upper Howard River). Locally also on levee and margins of wetlands.
Equivalent to G20 type of Nicholls (1976). Wardle (1991: 193–195). Includes rare ecosystem: frost hollows (Williams et al. 2007).
Appendix 3—continued
Continued on next page

65Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
TI6: Red tussock tussockland
Tall tussock grassland of abundant red tussock with inter-tussock herbfield / short tussockland and prostrate shrub species. Early alluvial successions are dominated by short tussockland of Poa, Festuca, Deyeuxia and Rytidosperma species. Typically includes an embedded, complex mosaic of bog and fen wetlands on organic soils.
In the North Island, restricted to the volcanic plateau, from the Hauhungaroa Range south to Erua and the Kaimanawa Mountains on alluvial terraces and headwater basins, to southern Ruahine. In Northwest Nelson, occurs with wire rush (e.g. Gouland Downs and Thousand Acres Plateau) and with C. rigida in eastern Fiordland. Occurs on valley floors in Westland (e.g. Toaroha and Landsborough Rivers) and Fiordland (e.g. Takahe Valley).
Elder (1962: 22), Evans (1969a), Druce et al. (1987), Wardle (1991: 226), Grove (1994), Mark & Dickinson (1997) and Mark et al. (2003: 193 & 200–202). Includes rare ecosystem: frost hollows (Williams et al. 2007).
WL1: Mānuka, gumland grass tree, Machaerina scrub/sedgeland [Gumland]
Low scrub, sedgeland of two broad types (poor-draining and seasonally dry), dominated by mānuka with gumland grass tree and tall mingimingi, and with species of Machaerina, Schoenus, Gahnia, Tetraria, Lepidosperma sedges and, locally, tangle fern.
Palustrine wetlands in the Northland and Auckland regions, developed in association with historic kauri forest podzolised Wharekohe and Te Kopuru soils (Molloy 1998: 92–94). Poor-draining type occurs on Wharekohe soils, while seasonally dry type occurs on Te Kopuru soils. Vegetation type also occurs on fire-induced and highly leached, non-podzolised soils, and it is now difficult to determine which areas are natural or induced.
Esler & Rumball (1975), Dodson et al. (1988), Conning (2001) and Clarkson et al. (2011). Includes rare ecosystem: gumland (Williams et al. 2007).
WL2: Mānuka, greater wire rush restiad rushland
Scrub, restiad rushland, fernland, sedgeland of abundant mānuka, with greater wire rush, tangle fern, Machaerinateretifolia (e.g. M. rubignosa) and Schoenus brevifolius.
Palustrine wetlands in Northland and Waikato lowland plains (e.g. Motutangi Swamp—Northland, and Whangamarino) within bogs of approximately 1500–7000 years of age.
Elliot et al. (1983), Clarkson (1997) and Clarkson et al. (2004).
WL3: Bamboo rush, greater wire rush restiad rushland
Restiad rushland of abundant bamboo rush and locally abundant greater wire rush, with occasional scrub of mānuka, Dracophyllum lessonianum and Epacris sinclairii, and locally Lycopodiella lateralis, Machaerina teretifolia, Schoenus brevifolius and tangle fern. May include small, embedded pools with sphagnum, and species of Utricularia and Drosera.
Palustrine wetlands in lowland plains of Northland and Waikato (e.g. Kopuatai, Moanatuatua) within raised bogs of approximately > 7000 years of age.
Campbell (1964), Wardle (1991: 324–325), Clarkson (1997) and Clarkson et al. (2004). Includes rare ecosystem: domed bog (Williams et al. 2007).
WL4: Mānuka, lesser wire rush, tangle fern scrub/fernland/restiad rushland [Pakihi]
Low scrub, fernland, restiad rushland of wire rush, tangle fern, Machaerina spp., mānuka and Dracophyllum palustre, with a margin of bog pine, silver pine, pink pine and yellow silver pine. Locally includes abundant sphagnum and red tussock.
Palustrine wetlands predominantly in the West Coast occurring on podzolised glacial moraines. Soils described by Molloy (1998: 19). Large areas have been induced through logging and burning, and were likely formerly equivalent to forest units MF16 and MF17.
Equivalent to M4 Pakihi Heathland of Newsome (1987). Wardle (1977—‘Natural pakihi’, 1991: 334–337), Williams et al. (1990) and Johnson & Gerbeaux (2004). Includes rare ecosystem: pakihi (Williams et al. 2007).
WL5: Chatham Island bamboo rush restiad rushland
Restiad rushland of a mosaic of abundant Chatham Island bamboo rush, with Olearia semidentata and Dracophyllum scoparium, and locally tangle fern, square sedge and Leptecophylla robusta. On better drained slopes, merges with taller tarahinau forest, and Dracophyllum scoparium, Dicksonia squarrosa and ground ferns also occur.
Palustrine wetlands (peatlands) of the Chatham Islands, with tarahinau on easy hill country and rolling uplands (coastal to uplands), and with Chatham Island bamboo rush on flats, gentle slopes and basins with impeded drainage.
Equivalent to communities 1, 2 and 7 of Kelly (1983). Wardle (1991: 442–443). Includes rare ecosystem: domed bog (Williams et al. 2007).
Appendix 3—continued
Continued on next page

66Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
WL6: Lesser wire rush, tangle fern restiad rushland/fernland
Restiad rushland of abundant wire rush and tangle fern, with occasional sedges, including Machaerina tenax and square sedge, often with sphagnum and tussock grasses. May include pools and gradations to shrub bogs (especially small podocarp trees), mānuka, Dracophyllum spp. and mountain tauhinu or red tussock fens.
Palustrine wetlands in upland areas of the central North Island (e.g. Lake Waikareiti bogs, western Tongariro National Park to Moawhango). In the South Island, widespread in humid montane regions on the West Coast, from Mackay Downs to Stewart Island/Rakiura, with local examples to near sea level in south Westland, Southland (e.g. Awarua bog) and on Stewart Island/Rakiura. In eastern South Island, rare and restricted to headwater regions.
Burrows (1969), Burrows & Dobson (1972), Johnson (1977), Mark et al. (1979), Sandercock (1987), Wilson (1987: 48–51), Wardle (1991: 321–323 & 348–349), McQueen & Wilson (2000) and Allen et al. (2003: 168). Includes rare ecosystems: blanket mires; and string mires (Williams et al. 2007).
WL7: Tall tussock tussockland
Tussockland, restiad rushland of several variants, locally with Chionochloa acicularis, C. juncea and C. teretifolia, and abundant wire rush and tangle fern. Scattered shrubland locally includes mānuka, pink pine, yellow silver pine and Dracophyllum spp.
Palustrine wetlands and peatlands on blanket peat (irrespective of slope) throughout the humid South Island areas, on Denniston Plateau, South Westland, West Cape and the Longwood Range.
Wardle & Mark (1973), Wardle (1991: 226, 230), Mark et al. (2003: 216) and Johnson & Gerbeaux (2004: 57 & 60). Includes rare ecosystem: blanket mire (Williams et al. 2007).
WL8: Herbfield/mossfield/sedgeland
Mossfield, herbfield, sedgeland with sphagnum and other mosses, short-statured sedges (Carex, Isolepis spp. and Carpha alpina) and a range of herbs (species of Epilobium, Euphrasia and Gentianella). Often contains numerous small pools with associated aquatic species.
Palustrine wetlands predominantly locally occurring in upland areas throughout the South Island main axial range, but most abundant east of the Main Divide in the Otago and northern Southland schist mountains. These often grade to fens and seepages with an increase in slope.
Burrows (1969: 125–126), Burrows & Dobson (1972), Mark et al. (1995) and Rapson et al. (2006).
WL9: Cushionfield Cushionfield with species of Oreobolus, Donatia, Gaimardia, Centrolepis, Carpha alpina and Phyllachne, and often Androstoma empetrifolia, Pentachondra pumila and Lepidothamnus laxifolius. Locally includes scattered treeland, with mānuka, pink pine, mountain beech and yellow silver pine.
Small areas of palustrine wetlands present in montane and subalpine areas throughout. Most common in southern districts and on subantarctic islands. Occurs at sea level at Awarua and above 300 m a.s.l. at Waitutu, with scattered stunted mānuka, podocarp trees and mountain beech.
Burrows (1969), Mark & Bliss (1970), Wardle (1977, 1991: 331–334), Mark et al. (1988) and Johnson & Gerbeaux (2004). Includes rare ecosystem: cushion bog (Williams et al. 2007).
WL10: Oioi restiad rushland/reedland
Restiad rushland with abundant oioi, locally with large Machaerina, Bolboschoenus spp., kuta and lake clubrush, and often with occasional raupō and scattered harakeke grading into wetland scrub on margins.
Riverine/lacustrine wetlands in the North Island, South Island and Chatham Islands, occurring in freshwater areas of estuaries, coastal stream margins, and in some inland areas adjacent to lakes in the central North Island and Southland.
Equivalent to communities 14 and 16 of Kelly (1983). Deng et al. (2004).
WL11: Machaerina sedgeland
Sedgeland, rushland with a high water table dominated by species of Machaerina, square sedge, Eleocharis and Juncus, often with scattered harakeke and Carex spp. Locally includes oioi, tangle fern and Gahnia spp., which can be locally dominant. Lagg margins often grade into mānuka scrub fens.
Palustrine/riverine/lacustrine wetlands of a wide range of variants throughout New Zealand. Widespread in the central North Island (e.g. South Taupo Wetland), while more restricted in the South Island (e.g. Kakapo Swamp). Also occurs on the margins of inland and coastal oligotrophic/mesotrophic lakes (e.g. Lakes Waikareiti and Rotopounamu).
Burrows & Dobson (1972—‘Kakapo Mire’), Clarkson (1984), Wardle (1991: 321–335), Eser (1998) and Pegman & Ogden (2006). Includes rare ecosystems: lagoon; and lake margins (Williams et al. 2007).
WL12: Mānuka, tangle fern scrub/fernland
Scrub with abundant mānuka and occasional species of Olearia, Coprosma and Dracophyllum, and species of Machaerina, square sedge, Carex and Juncus. Locally abundant tangle fern, Schoenus pauciflorus, sphagnum, stunted harakeke, and species of Astelia and Gahnia. Locally also includes bog pine, silver pine and pink pine.
Palustrine wetlands widespread in Northland (e.g. Maitahi Scientific Reserve), central North Island (e.g. Paramanawera wetland, southern Ruapehu), West Coast of the South Island, Southland and Stewart Island/Rakiura.
Burrows & Dobson (1972), Wardle (1977—‘lowland infertile swamp’, 1991: 334–337), Clarkson (1986), Sandercock (1987), Eser (1998) and McQueen & Wilson (2000).
Appendix 3—continued
Continued on next page

67Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
WL13: Sphagnum mossfield
Mossfield of abundant sphagnum, often with a sparse canopy of stunted scrub / low treeland of mānuka, locally silver beech, bog pine and Dracophyllum spp., with abundant cover of sphagnum, and a sparse component of sedges, rushes and herbs (e.g. Drosera binata). On Chatham Islands, includes local endemics (e.g. Carex chathamica, Olearia semidentata and Dracophyllum scoparium).
Riverine/lacustrine wetlands, most abundant in Chatham Islands. Rare in both the North Island and South Island, though more abundant in humid regions. North Island examples include floating (sudd) lake edge or spring-fed systems.
Cockayne (1909b: 284–285), Bayfield et al. (1991), Ravine (1991), Wardle (1991: 312 & 444) and Lake & Whaley (1995). Includes rare ecosystem: lake margins (Williams et al. 2007).
WL14: Herbfield [Ephemeral wetland]
Herbfield and/or low sedgeland dominated by a wide range of predominantly montane, short-statured herbs, grasses and sedges. Dominants may include species of Leptinella, Lobelia, Hydrocotyle, Euchiton, Epilobium, Plantago, Ranunculus, Myriophyllum, Elatine, Glossostigma, Isolepis, Eleocharis, Carex and Deschampsia.
Riverine/lacustrine wetlands predominantly present in sub-humid or semi-arid regions, though now most abundant in cool inland regions. In the North Island, in tephra hollows of volcanic substrates (e.g. Waihora and Arahaki Lagoons). Throughout the eastern South Island, on old glacial moraines in kettle holes (e.g. Tekapo), outwash channels, and rare examples on karst, within schist depressions—including ‘Sutton Lake’, a unique saline example that includes some halophyte plants.
Shanks et al. (1990), Johnson (1991) and Johnson & Rogers (2002). Includes rare ecosystems: ephemeral wetlands; and lagoon margins (Williams et al. 2007).
WL15: Herbfield [Lakeshore turf]
Herbfield and/or low sedgeland of two broad variants (coastal and inland), which often have species in common. Coastal variant is often brackish, commonly includes Selliera radicans, and species of Isolepis, Limosella and Lilaeopsis, and grades into salt marsh with increasing salinity. Inland variant commonly includes Glossistigma elatinoides, species of Lilaeopsis, Carex, Eleocharis, Lobelia, Centrolepis, Hydrocotyle, Myriophyllum, Plantago, Ranunculus and Crassula, and other herb species.
Lacustrine wetlands associated with coastal and inland lakes in the North Island, South Island and Chatham Islands (e.g. Lakes Wairarapa and Forsyth), and inland (e.g. Lakes Manapouri, Te Anau, Waikaremoana and Taupo (Taupomoana)). Most abundant on lake edges with high seasonal water height fluctuations and moderate– high fetch.
Equivalent to community 15 of Kelly (1983). Johnson (1972), Macmillan (1979), Wardle (1991: 303), Wells et al. (1998), Champion et al. (2001) and de Lange & Murray (2008). Includes rare ecosystems: lagoon; and lake margins (Williams et al. 2007).
WL16: Red tussock, Schoenus pauciflorus tussockland
Tussockland with abundant red tussock, locally with cushionfield, sedgeland and shallow pools, with Schoenus pauciflorus, Oreobolus spp., Carpha alpina, Carex coriacea and C. sinclairii, and locally sphagnum, wire rush and scattered shrubs (e.g. Hebe odora and bog pine).
Palustrine/riverine wetlands in wet basins and gully floors locally in the central North Island (e.g. Ahukawakwa Bog, Egmont), and in the South Island within the Main Divide to Southland (e.g. Mt Hamilton flat).
Burrows (1969: 128), Dobson (1975— Mt Hamilton flat RAP, Southland), Johnson (1977, 1986), Clarkson (1986) and Shanks et al. (1990). Includes rare ecosystem: blanket mire (Williams et al. 2007).
WL17: Schoenus pauciflorus sedgeland [Alpine seepages/flushes]
Low stature sedgeland, mossfield, herbfield, with abundant mosses, liverworts and sedges, and a wide range of herbs, including Schoenus pauciflorus and Carpha alpina, and locally species of Epilobium, Montia, Ranunculus, Schizeilema, Hydrocotyle and Gentianella.
Palustrine/riverine wetlands or alpine seepages and flushes with a moderately constant flow, which are widespread in the mountain areas of the North Island and South Island, though very restricted to many small areas associated with individual springs or seepages. These often grade upslope to wet snowbank or cushionfield, or laterally downslope to large-scale fens. Often associated with other larger wetland types, especially at lower altitudes.
Burrows (1969: 131–132) and Wardle (1977, 1991: 41).
Appendix 3—continued
Continued on next page

68Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
WL18: Flaxland Flaxland of abundant harakeke, often with toetoe, species of Carex (e.g. pūkio) and Machaerina, and kiokio, occasional wetland scrub, treeland of cabbage tree, Coprosma spp. and mānuka, and locally weeping matipo and twiggy tree daisy. Areas with high water tables may be dominated by pūkio. May grade or succeed into wetland carr, with abundant emergent cabbage trees.
Palustrine/riverine/lacustrine wetlands; common in the North Island, especially coastal and riparian wetlands (e.g. Taupo Swamp, Plimmerton). Dominant type from Northwest Nelson (e.g. Mangarakau) to South Westland, often with scattered mānuka and kahikatea on margin.
Wardle (1977, 1991: 309–320), Esler (1978b), Bagnall & Ogle (1981) and Ravine (1991).
WL19: Raupō reedland
Reedland of abundant raupō, locally with species of Bolboschoenus, Schoenoplectus and Machaerinaarticulata, pūkio, harakeke, and swamp millet. A margin of scrub of Coprosma species and cabbage tree, and locally twiggy tree daisy and mānuka, with scattered kahikatea in unmodified areas. Often occurs on lake margins or includes small ponds with shallow water/pools with floating/rafted aquatics such as water milfoils, buttercups, willowherbs, species of Potamogeton, Isolepis, Azolla and Lemna, and spiked sedges (e.g. kuta).
Palustrine/riverine/lacustrine wetlands; commonly found thoughout lowlands on old river oxbows, margins of lakes and flooded valleys fom Northland to South Otago (e.g. Lake Waihola). Now abundant on farm ponds, though floristically poor.
Esler (1978b), Ogden & Caithness (1982), Sandercock (1987), Eser (1998) and Pegman & Ogden (2005). Includes rare ecosystem: lake margins (Williams et al. 2007).
WL20: Coprosma, twiggy tree daisy scrub
Scrub of species of Coprosma and locally twiggy tree daisy (which can be locally dominant), with a mosaic of a wide variety of Carex spp. and locally kiokio. May also locally include scattered harakeke, raupō, toetoe and cabbage trees.
Common type in palustrine/riverine/lacustrine wetlands in the central North Island, and southern and eastern North Island. In the South Island, largely occurs in western regions, from Northwest Nelson to Southland, often in freshwater areas of estuaries and brackish coastal lagoons. Likely to have been more abundant prior to Māori and European fires, and now largely restricted to areas with a less frequent fire history.
Lake & Whaley (1995—RAP 21) and Singers & Whyman (2001—‘Three springs wetland’).
WL21: Swamp akeake, Chatham Island karamū, Coprosma propinqua var. martinii short forest/flaxland
Short forest of Olearia telmatica, Coprosma chathamica, C. propinqua var. martinii, supplejack and kiokio, with harakeke, Chatham Island toetoe, Astelia chathamica, Carex ternaria and C. sectoides in areas with a higher water table.
Palustrine/riverine/lacustrine wetlands on main Chatham Island around larger peat lakes, wetlands, and along streams and the margins of some lowland lakes. Chatham Island toetoe is now a rare component.
Equivalent to communities 6 and 17 of Kelly (1983). Cockayne (1901: 274–275) and Wardle (1991: 444).
WL22: Carex, Schoenus pauciflorus sedgeland
Sedgeland with mosaics of a wide variety of species of Carex, including C. secta, C. virgata, C. diandra, C. coriacea, C. sinclairii and C. gauchichaudiana, and Schoenus pauciflorus, with locally small pools and lakes often with a fringe of raupō. Schoenus becomes more abundant at higher altitudes, while occasional harakeke may be present at lower altitudes. Intact examples have margins of wetland scrub.
Palustrine/riverine/lacustrine wetlands, which are relatively uncommon and restricted in distribution in the North Island. More widespread in the eastern South Island, occurring within inter-montane basins and valley floors of the Southern Alps/Kā Tiritiri o te Moana, and a dominant type at higher altitudes in inland Canterbury. Uncommon west of the Main Divide.
Wardle (1977), Partridge & Molloy (1985), Shanks et al. (1990) and Johnson (1991).
Appendix 3—continued
Continued on next page

69Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
DN1: Beach morning glory, knobby clubrush vineland/sedgeland
Vineland of abundant beach morning glory, with scattered sedgeland of knobby clubrush and Imperata cheesemanii, and locally Cyperus insularis, shore bindweed, Senecio kermadecensis, Scaevola gracilis, New Zealand spinach and shore celery, grading into Kermadec ngaio and Kermadec pōhutukawa forest.
Largely an Australian and Pacific dune type, only being found on subtropical Raoul Island within New Zealand, where it occurs in two areas: one small area on the north coast and dunes, and a gravel flat in Denham Bay. Largely in a secondary derivative state. Historically was a nesting location for Kermadec sooty tern (Onychoprion fuscatus kermadeci), which is no longer present.
Oliver (1910: 133–134). Includes rare ecosystems: active sand dunes; and seabird guano deposits (Williams et al. 2007).
DN2: Spinifex, pīngao grassland/sedgeland
Sedgeland, grassland of abundant spinifex and pīngao, with occasional shore bindweed, sand coprosma, tauhinu and sand daphne, grading into rear semi-stable dunes with open, scattered dune scrub of bracken, Muehlenbeckia complexa, toetoe, harakeke and cabbage trees. Locally includes matagouri, mānuka, kānuka, tutu and Olearia solandri.
Present from Northland to Farewell Spit, and historically scattered to Buller River in the west and the Waimakariri River in the east. Historically included Atriplex hollowayi within the strand zone, which is now restricted to the far north.
Cockayne (1911), Esler (1970) and Wardle (1991: 355–356). Includes rare ecosystem: active sand dunes (Williams et al. 2007).
DN3: Pīngao sedgeland
Sedgeland of abundant pīngao of two variants (humid and semi-arid), with occasional knobby clubrush, Carex pumila, shore bindweed, sand tussock, sand coprosma and Muehlenbeckia complexa, and locally Euphorbia glauca, Sonchus kirkii, Pimelea lyallii, Carex testacea, Acaena spp., silver tussock and kāretu, grading into rear semi-stable dunes with scattered dune scrub. Grades into abundant harakeke, mānuka and Olearia avicenniifolia in humid areas, and into matagouri, Carmichaelia spp., akeake and ngaio in semi-arid areas.
At least two regional variants, with the humid variant in western/southern parts of the range and the semi-arid variant in Canterbury (e.g. Kaitorete Spit). The humid variant occurs from Hokitika south to Stewart Island/Rakiura and coastal Otago. Mostly now very modified, with few intact examples remaining. Historically included Atriplex billardierei within the strand zone on beaches within the Foveaux Strait and on Stewart Island/Rakiura, though this is now locally extinct. The semi-arid variant is restricted to Kaitorete Spit.
Cockayne (1909b, 1911), Wardle (1977, 1991: 356–359), Wardle et al. (1986), Sandercock (1987) and de Lange et al. (2000). Includes rare ecosystem: active sand dunes (Williams et al. 2007).
DN4: Pīngao sedgeland, mega-herbfield
Sedgeland, mega-herbfield of Atriplex billardierei, sand buttercup, Carex pumila and shore bindweed above the high water line, grading into abundant pīngao with Poa chathamica, knobby clubrush, sand daphne, Myosotidium hortensium and Embergeria grandifolia; and locally Euphorbia glauca, Geranium traversii and Urtica australis, grading into scrub of Leucopogon parviflorus, and dune forest of Olearia traversiorum and Myrsine chathamica.
Dunes on main Chatham Island. Historically formed a narrow dune ridge, parallel to the coastline.
Equivalent to community types 11 and 12 of Kelly (1983). Cockayne (1901). Includes rare ecosystem: active sand dunes (Williams et al. 2007).
DN5: Oioi, knobby clubrush sedgeland
Sedgeland, herbfield of several local variants with both dry and ephemerally wet communities of a range of successional stages. Dominant species include Carex pumila, species of Gunnera, Selliera, Isolepis, Epilobium, Ranunculus, Leptinella, Lobelia, Colobanthus, Geranium and Hydrocotyle, and locally Lilaeopsis novae-zelandiae, Myriophyllum votschii, Triglochin striata, Limosella lineata and other turf-forming species. Older stages develop into oioi, knobby clubrush, toetoe and harakeke, and locally Cyperus ustulatus, Lepidosperma australe, silver tussock and Raoulia spp. Locally includes Coprosma propinqua and mānuka in older successions.
Predominantly present in larger dunelands in association with mobile dunes and rapidly accreting coastlines. In the North Island, largely occurs in Northland (e.g. Aupouri, Poutu, South Kaipara) and Foxton Ecological District (South Taranaki to Paekakariki); in the South Island, occurs on Farewell Spit, Canterbury, Otago, Southland and Stewart Island/Rakiura. Often includes a mosaic of both dry deflation hollows and seasonally wet areas (including sandy deflation hollows, sand plains, dune slacks and low mounds). Locally occurs on coastal hill slopes with sandy soils. May succeed into coastal flaxland with peat accumulation.
Cockayne (1911), Logan & Holloway (1934), Esler (1969), Sykes & Wilson (1987), Wilson (1987), Roxburgh et al. (1994), Drobner et al. (1995) and Johnson & Rogers (2002). Includes rare ecosystems: deflation hollow; damp sand plains; and dune slacks (Williams et al. 2007).
Appendix 3—continued
Continued on next page
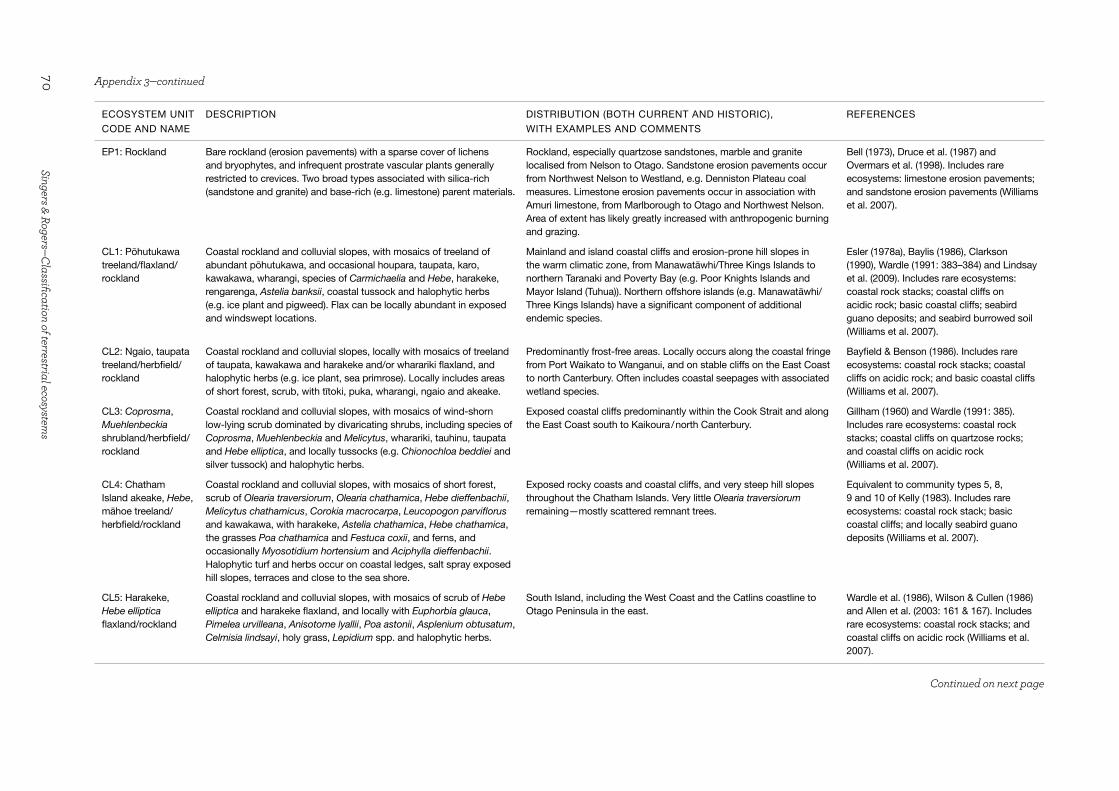
70Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
EP1: Rockland Bare rockland (erosion pavements) with a sparse cover of lichens and bryophytes, and infrequent prostrate vascular plants generally restricted to crevices. Two broad types associated with silica-rich (sandstone and granite) and base-rich (e.g. limestone) parent materials.
Rockland, especially quartzose sandstones, marble and granite localised from Nelson to Otago. Sandstone erosion pavements occur from Northwest Nelson to Westland, e.g. Denniston Plateau coal measures. Limestone erosion pavements occur in association with Amuri limestone, from Marlborough to Otago and Northwest Nelson. Area of extent has likely greatly increased with anthropogenic burning and grazing.
Bell (1973), Druce et al. (1987) and Overmars et al. (1998). Includes rare ecosystems: limestone erosion pavements; and sandstone erosion pavements (Williams et al. 2007).
CL1: Pōhutukawa treeland/flaxland/rockland
Coastal rockland and colluvial slopes, with mosaics of treeland of abundant pōhutukawa, and occasional houpara, taupata, karo, kawakawa, wharangi, species of Carmichaelia and Hebe, harakeke, rengarenga, Astelia banksii, coastal tussock and halophytic herbs (e.g. ice plant and pigweed). Flax can be locally abundant in exposed and windswept locations.
Mainland and island coastal cliffs and erosion-prone hill slopes in the warm climatic zone, from Manawatāwhi/Three Kings Islands to northern Taranaki and Poverty Bay (e.g. Poor Knights Islands and Mayor Island (Tuhua)). Northern offshore islands (e.g. Manawatāwhi/Three Kings Islands) have a significant component of additional endemic species.
Esler (1978a), Baylis (1986), Clarkson (1990), Wardle (1991: 383–384) and Lindsay et al. (2009). Includes rare ecosystems: coastal rock stacks; coastal cliffs on acidic rock; basic coastal cliffs; seabird guano deposits; and seabird burrowed soil (Williams et al. 2007).
CL2: Ngaio, taupata treeland/herbfield/rockland
Coastal rockland and colluvial slopes, locally with mosaics of treeland of taupata, kawakawa and harakeke and/or wharariki flaxland, and halophytic herbs (e.g. ice plant, sea primrose). Locally includes areas of short forest, scrub, with tītoki, puka, wharangi, ngaio and akeake.
Predominantly frost-free areas. Locally occurs along the coastal fringe from Port Waikato to Wanganui, and on stable cliffs on the East Coast to north Canterbury. Often includes coastal seepages with associated wetland species.
Bayfield & Benson (1986). Includes rare ecosystems: coastal rock stacks; coastal cliffs on acidic rock; and basic coastal cliffs (Williams et al. 2007).
CL3: Coprosma, Muehlenbeckia shrubland/herbfield/rockland
Coastal rockland and colluvial slopes, with mosaics of wind-shorn low-lying scrub dominated by divaricating shrubs, including species of Coprosma, Muehlenbeckia and Melicytus, wharariki, tauhinu, taupata and Hebe elliptica, and locally tussocks (e.g. Chionochloa beddiei and silver tussock) and halophytic herbs.
Exposed coastal cliffs predominantly within the Cook Strait and along the East Coast south to Kaikoura / north Canterbury.
Gillham (1960) and Wardle (1991: 385). Includes rare ecosystems: coastal rock stacks; coastal cliffs on quartzose rocks; and coastal cliffs on acidic rock (Williams et al. 2007).
CL4: Chatham Island akeake, Hebe, māhoe treeland/herbfield/rockland
Coastal rockland and colluvial slopes, with mosaics of short forest, scrub of Olearia traversiorum, Olearia chathamica, Hebe dieffenbachii, Melicytus chathamicus, Corokia macrocarpa, Leucopogon parviflorus and kawakawa, with harakeke, Astelia chathamica, Hebe chathamica, the grasses Poa chathamica and Festuca coxii, and ferns, and occasionally Myosotidium hortensium and Aciphylla dieffenbachii. Halophytic turf and herbs occur on coastal ledges, salt spray exposed hill slopes, terraces and close to the sea shore.
Exposed rocky coasts and coastal cliffs, and very steep hill slopes throughout the Chatham Islands. Very little Olearia traversiorum remaining—mostly scattered remnant trees.
Equivalent to community types 5, 8, 9 and 10 of Kelly (1983). Includes rare ecosystems: coastal rock stack; basic coastal cliffs; and locally seabird guano deposits (Williams et al. 2007).
CL5: Harakeke, Hebe elliptica flaxland/rockland
Coastal rockland and colluvial slopes, with mosaics of scrub of Hebe elliptica and harakeke flaxland, and locally with Euphorbia glauca, Pimelea urvilleana, Anisotome lyallii, Poa astonii, Asplenium obtusatum, Celmisia lindsayi, holy grass, Lepidium spp. and halophytic herbs.
South Island, including the West Coast and the Catlins coastline to Otago Peninsula in the east.
Wardle et al. (1986), Wilson & Cullen (1986) and Allen et al. (2003: 161 & 167). Includes rare ecosystems: coastal rock stacks; and coastal cliffs on acidic rock (Williams et al. 2007).
Appendix 3—continued
Continued on next page

71Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
CL6: Hebe, wharariki flaxland/rockland
Rockland and colluvial slopes with several local variants over a wide latitudinal/altitudinal gradient, with mosaics of short-statured herbs, grasses, short forest and scrub. Dominants include wharariki, Poa anceps, species of Hebe, Gaultheria, Pimelea, Olearia, Sophora, Carmichaelia, Leucopogon, Cyathodes and Dracophyllum, and tutu, and locally ngaio, kānuka, Chionochloa flavicans, Astelia solandri, Dianella nigra and Collospermum hastatum. Locally, subalpine species include Hebe colensoi and Pimelea spp. on inland sites, and local endemics on weakly weathered calcareous parent materials. Locally includes Machaerina sinclarii, kiokio, and rheophytic herbs, sedges, grasses and bryophytes associated with seepages, streams and rivers.
Predominantly sub-humid and semi-arid zones of the North Island: Northland–Auckland, East Cape to southern Wairarapa, Rangitikei District and the volcanic plateau. Wide range of geology in eastern and Rangitikei districts, predominantly on mudstone/siltstone/sandstone, and locally areas of limestone and greywacke (e.g. Wairarapa Taipos) and closer to the central mountains inland (e.g. Kaweka Range). Largely volcanic geology from Northland to the Taupo Volcanic Zone. Also present on exposed north-facing dry sites in the western zone.
Gardner (1989), Wardle (1991: 384–385), Lake & Whaley (1995), Whaley et al. (2001), Johnson & Gerbeaux (2004) and de Lange & Rolfe (2008). Includes rare ecosystems: cliffs, scarps and tors on acidic rocks; calcareous cliffs, tors and scarps; and cliffs, scarps and tors on basic rocks (Williams et al. 2007).
CL7: Pachystegia, Carmichaelia shrubland/tussockland/rockland
Rockland and colluvial slopes with mosaics of scrub, shrub, fern and grass species, including wharariki, and species of Pachystegia, Brachyglottis, Carmichaelia, Heliohebe, Hebe, Helichrysum, Coprosma and Coriaria, and locally tauhinu, Aciphylla aurea, and hard, silver and Chionochloa tussock grasses. Locally also includes scrub of broadleaf, Olearia paniculata, kōwhai, akeake and ferns, as well as herbs, sedges and grasses in association with seepages. Local endemics occur on weakly weathered calcareous parent materials (limestone).
Eastern South Island dryland zone, from Kaikoura to north Canterbury on greywacke and calcareous outcrops.
Wardle (1971), Druce & Williams (1989), Wardle (1991: 385 & 388), Moore (1999: 35) and Rogers et al. (2005). Includes rare ecosystems: cliffs, scarps and tors on acidic rocks; calcareous cliffs, tors and scarps; and boulderfields of calcareous rocks (Williams et al. 2007).
CL8: Helichrysum, Melicytus shrubland/tussockland/rockland
Rockland and colluvial slopes with mosaics of scrub, shrub, fern and grass species, small-leaved Olearia, Coprosma, Carmichaelia and Hebe, wharariki, tutu, kānuka, mānuka, Corokia cotoneaster, Olearia avicenniifolia, prostrate kōwhai and broadleaf. Several local altitudinal variants occur with associated subalpine species, such as species of Heliohebe, Helichrysum and Pimelea, porcupine shrub, Brachyglottis lagopus and Dracophyllum acerosum, and locally grasses, including bristle tussock, silver tussock and Chionochloa rigida. Local endemics also occur on weakly weathered calcareous parent materials (limestone).
North Canterbury to Central Otago on greywacke, and locally on limestone and basalt (e.g. Banks Peninsula and Otago Peninsula) and schist.
Wardle (1991: 394) and Wiser & Buxton (2009). Includes rare ecosystems: cliffs, scarps and tors on basic rocks; and calcareous cliffs, tors and scarps (Williams et al. 2007).
Appendix 3—continued
Continued on next page
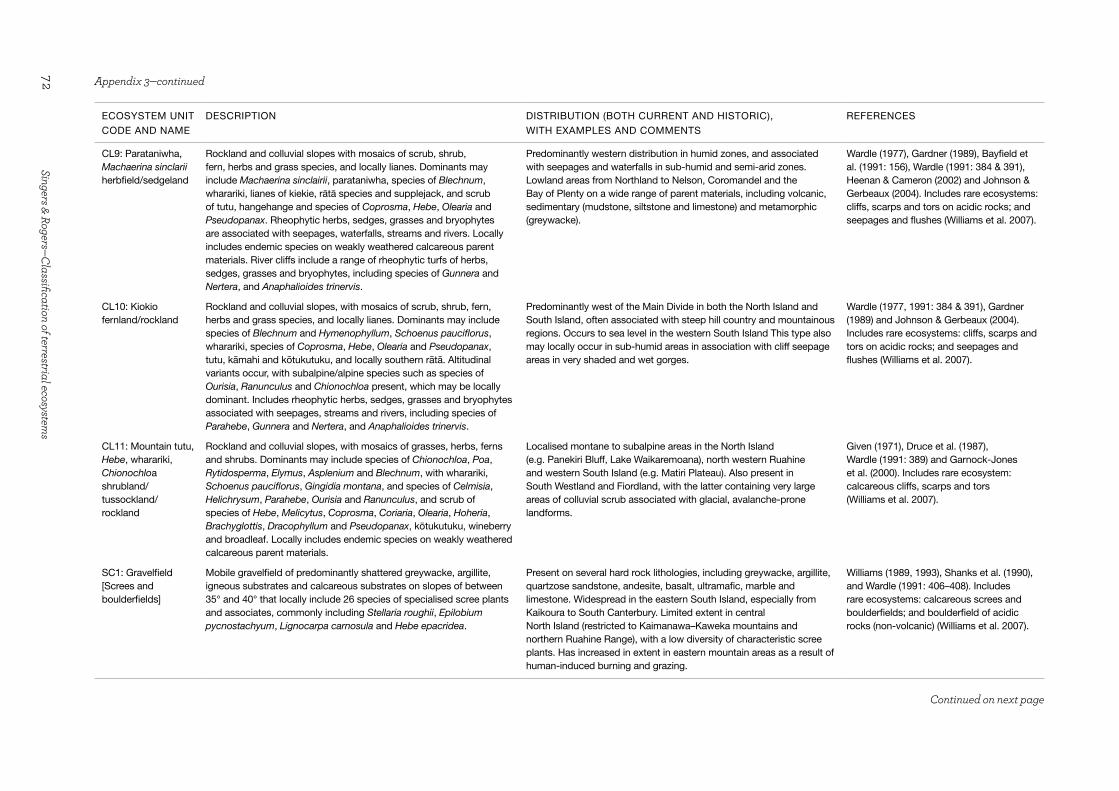
72Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
CL9: Parataniwha, Machaerina sinclarii herbfield/sedgeland
Rockland and colluvial slopes with mosaics of scrub, shrub, fern, herbs and grass species, and locally lianes. Dominants may include Machaerina sinclairii, parataniwha, species of Blechnum, wharariki, lianes of kiekie, rātā species and supplejack, and scrub of tutu, hangehange and species of Coprosma, Hebe, Olearia and Pseudopanax. Rheophytic herbs, sedges, grasses and bryophytes are associated with seepages, waterfalls, streams and rivers. Locally includes endemic species on weakly weathered calcareous parent materials. River cliffs include a range of rheophytic turfs of herbs, sedges, grasses and bryophytes, including species of Gunnera and Nertera, and Anaphalioides trinervis.
Predominantly western distribution in humid zones, and associated with seepages and waterfalls in sub-humid and semi-arid zones. Lowland areas from Northland to Nelson, Coromandel and the Bay of Plenty on a wide range of parent materials, including volcanic, sedimentary (mudstone, siltstone and limestone) and metamorphic (greywacke).
Wardle (1977), Gardner (1989), Bayfield et al. (1991: 156), Wardle (1991: 384 & 391), Heenan & Cameron (2002) and Johnson & Gerbeaux (2004). Includes rare ecosystems: cliffs, scarps and tors on acidic rocks; and seepages and flushes (Williams et al. 2007).
CL10: Kiokio fernland/rockland
Rockland and colluvial slopes, with mosaics of scrub, shrub, fern, herbs and grass species, and locally lianes. Dominants may include species of Blechnum and Hymenophyllum, Schoenus pauciflorus, wharariki, species of Coprosma, Hebe, Olearia and Pseudopanax, tutu, kāmahi and kōtukutuku, and locally southern rātā. Altitudinal variants occur, with subalpine/alpine species such as species of Ourisia, Ranunculus and Chionochloa present, which may be locally dominant. Includes rheophytic herbs, sedges, grasses and bryophytes associated with seepages, streams and rivers, including species of Parahebe, Gunnera and Nertera, and Anaphalioides trinervis.
Predominantly west of the Main Divide in both the North Island and South Island, often associated with steep hill country and mountainous regions. Occurs to sea level in the western South Island This type also may locally occur in sub-humid areas in association with cliff seepage areas in very shaded and wet gorges.
Wardle (1977, 1991: 384 & 391), Gardner (1989) and Johnson & Gerbeaux (2004). Includes rare ecosystems: cliffs, scarps and tors on acidic rocks; and seepages and flushes (Williams et al. 2007).
CL11: Mountain tutu, Hebe, wharariki, Chionochloa shrubland/tussockland/rockland
Rockland and colluvial slopes, with mosaics of grasses, herbs, ferns and shrubs. Dominants may include species of Chionochloa, Poa, Rytidosperma, Elymus, Asplenium and Blechnum, with wharariki, Schoenus pauciflorus, Gingidia montana, and species of Celmisia, Helichrysum, Parahebe, Ourisia and Ranunculus, and scrub of species of Hebe, Melicytus, Coprosma, Coriaria, Olearia, Hoheria, Brachyglottis, Dracophyllum and Pseudopanax, kōtukutuku, wineberry and broadleaf. Locally includes endemic species on weakly weathered calcareous parent materials.
Localised montane to subalpine areas in the North Island (e.g. Panekiri Bluff, Lake Waikaremoana), north western Ruahine and western South Island (e.g. Matiri Plateau). Also present in South Westland and Fiordland, with the latter containing very large areas of colluvial scrub associated with glacial, avalanche-prone landforms.
Given (1971), Druce et al. (1987), Wardle (1991: 389) and Garnock-Jones et al. (2000). Includes rare ecosystem: calcareous cliffs, scarps and tors (Williams et al. 2007).
SC1: Gravelfield [Screes and boulderfields]
Mobile gravelfield of predominantly shattered greywacke, argillite, igneous substrates and calcareous substrates on slopes of between 35° and 40° that locally include 26 species of specialised scree plants and associates, commonly including Stellaria roughii, Epilobium pycnostachyum, Lignocarpa carnosula and Hebe epacridea.
Present on several hard rock lithologies, including greywacke, argillite, quartzose sandstone, andesite, basalt, ultramafic, marble and limestone. Widespread in the eastern South Island, especially from Kaikoura to South Canterbury. Limited extent in central North Island (restricted to Kaimanawa–Kaweka mountains and northern Ruahine Range), with a low diversity of characteristic scree plants. Has increased in extent in eastern mountain areas as a result of human-induced burning and grazing.
Williams (1989, 1993), Shanks et al. (1990), and Wardle (1991: 406–408). Includes rare ecosystems: calcareous screes and boulderfields; and boulderfield of acidic rocks (non-volcanic) (Williams et al. 2007).
Appendix 3—continued
Continued on next page

73Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
BR1: Hard tussock, scabweed gravelfield/stonefield
Stonefield, gravelfield with a mosaic of prostrate herbfield and sub-shrubs of scabweed mats and short-lived herbs (e.g. species of Raoulia and Epilobium, Pimelea prostrata, and Muehlenbeckia axillaris) grading into short-tussock grasslands on fresh alluvium with silver tussock, hard tussock, Anthosachne spp. and long-hair plume grass, and matagouri scrub (inland South Island) or kānuka scrub on older, more stable soils. Locally includes inland dunes.
Adjacent to active braided rivers throughout the North Island and South Island, though most abundant in the east from Marlborough through to Canterbury and northern Otago. Also includes sub-humid areas from Waikanae to Rangitikei River in the southern North Island. Historically, small-leaved shrubs were likely common (Rogers et al. 2005).
Equivalent to Matagouri shrubland class of Wiser & Hurst (2008). Elder (1962: 22), Cox & Mead (1963: 114), Fisher (1969) and Shanks et al. (1990). Includes rare ecosystems: braided rivers; and inland dunes (Williams et al. 2007).
BR2: Scabweed gravelfield/stonefield
Stonefield, gravelfield with a mosaic of prostrate herbfield of scabweed and willowherb species, including Raoulia tenuicaulis, R. hookeri and Epilobium microphyllum on bare gravels grading into short tussock grassland at higher altitude, and/or Austroderia spp. tall tussock grasslands, with species of Hebe, Coprosma, Carmichaelia and Coriaria, and mānuka scrub on recent alluvial flood plains. Locally may also include Olearia avicenniifolia, especially at higher altitudes in Westland.
Higher rainfall areas throughout the North Island and South Island. Most abundant on the West Coast of the South Island, from the Buller River south.
Wardle (1974, 1977, 1980), Wardle et al. (1986) and Johnson & Gerbeaux (2004). Includes rare ecosystem: braided rivers (Williams et al. 2007).
BR3: Bristle tussock, Raoulia, Muehlenbeckia gravelfield/sandfield
Gravelfield, sandfield with a mosaic of prostrate herbfield of species of Raoulia and Pimelea, and Muehlenbeckia axillaris, with localised patches of bristle and blue tussock, and mountain oat grass, and with scattered snow tōtara, Gaultheria spp. and Olearia nummulariifolia. Locally includes volcanic dunes.
Unique to Mt Ruapehu on the Whangaehu River lahar fan, central North Island.
Scott (1977) and Atkinson (1985). Includes rare ecosystems: inland sand dunes; and volcanic debris flows or lahars (Williams et al. 2007).
SA1: Mangrove forest and scrub
Forest and scrub of abundant mangrove, often with areas of rushland, herbfield including sea rush and oioi, and locally species of Machaerinajuncea and Bolboschoenus, and salt marsh ribbonwood, grading to seagrass herbfield on tidal flats. May locally include shell barrier beaches with a scattered herbfield of glasswort, buggar grass, knobby clubrush, sea rush, sea primrose and sea blite.
Frost-free estuarine hydrosystems north of 38° latitude, from Raglan and Ohiwa northwards, within tidal estuaries, inlets, rivers and streams.
Ward (1967), Wardle (1991: 291–294), Conning (2001) and Deng et al. (2004). Includes rare ecosystem: shell barrier beaches (Chenier Plains) (Williams et al. 2007).
SA2: Searush, oioi, glasswort, sea primrose rushland/herbfield [Saltmarsh]
Rushland, herbfield of sea grass, glasswort and sea primrose, locally with shell barrier and/or gravel beach ridges, grading into sea rush and oioi. Locally Machaerina juncea and Schoenoplectus spp. also occur, with areas of coastal herbfield (e.g. shore celery, half-star, bachelor’s button, arrow grass) grading into a fringe of coastal scrub of salt marsh ribbonwood, Olearia solandri, Coprosma propinqua and small-leaved pōhuehue.
Estuarine hydrosystems south of 38° latitude, from Raglan and Ohiwa in the North Island to South Canterbury and the West Coast in the South Island, within tidal estuaries, inlets, lagoons, rivers and streams. Also on the Chatham Islands, with some local endemics. Very few sites have intact sequences with both wetland and dryland components.
Wardle (1977, 1991: 291–298), Esler (1978b) and Johnson & Gerbeaux (2004). Includes rare ecosystem: shell barrier beaches (´Chenier Plains´) (Williams et al. 2007).
SA3: Glasswort, sea primrose herbfield [Saltmarsh]
Herbfield of seagrass grading into glasswort and sea primrose, with half-star, sea blite, Schoenus nitens, arrowgrass, Puccinellia spp. and shore celery grading into oioi, three-square and salt marsh ribbonwood. Locally with shell barrier and/or gravel or sand beach ridges, with silver tussock, knobby clubrush and square sedge.
Otago, Southland and Stewart Island/Rakiura, within tidal estuaries, inlets, lagoons, rivers and streams.
Equivalent to community type 19 of Wilson (1987). Partridge & Wilson (1988).
Appendix 3—continued
Continued on next page

74Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
SA4: Shore bindweed, knobby clubrush gravelfield/stonefield
Stonefield, gravelfield with at least four variants. Contains halophytic herbs, sedges and vines, including glasswort, half-star, shore celery, arrow grass, shore spurge, knobby clubrush and shore bindweed grading into a coastal scrub-vineland of Coprosma and Muehlenbeckia, and locally species of Melicytus, Pimelea and Ozothamnus, and harakeke. Treeland also locally includes ngaio, taupata, akeake, kōwhai and tānekaha further inland on older beach ridges at Miranda. Dominated by local endemics, including Myosotidium hortensium and Embergeria grandifolia, on the Chatham Islands.
Most common in the South Island, including Southland, Canterbury–Marlborough and the West Coast. More local in the North Island, occurring in Wellington, Hawke’s Bay, Taranaki and Coromandel– Firth of Thames, with small examples on Chatham Island. Also occurs within accreting estuarine areas with large rivers (e.g. Whakatiwai and Miranda) and Wairau River (Rarangi–Marlborough). Treeland successional stages are extremely rare and threatened.
Equivalent to community type 12 of Kelly (1983). Bagnall (1975), Wardle (1991) and Tyrell et al. (1999). Includes rare ecosystems: shingle beaches; and stony beach ridges (Williams et al. 2007).
SA5: Herbfield [Coastal turf]
Herbfield of a wide range of prostrate species, including half-star, sea primrose, shore celery, Zoysia minima, Isolepis cernua, Centella uniflora, Colobanthus muelleri, Hydrocotyle novae-zeelandiae, and species of Leptinella, Crassula, Ranunculus, Myosotis, Epilobium, Mazus and Nertera.
Most common and well developed within Taranaki–Wanganui, Te Tai Tapu coast–Nelson, North Westland, Otago, Southland, Fiordland and Chatham Island coastlines. Occurs on a variety of landforms from cliffs to beaches.
Mark et al. (1988: 41), Rogers (1999) and Rogers & Wiser (2010). Includes rare ecosystems: coastal turf; coastal rock stacks; coastal cliffs on quartzose rocks; coastal cliffs on acidic rock; and marine mammal haul-outs (Williams et al. 2007).
SA6: Kermadec ngaio scrub, mixed herbfield/loamfield
Mosaic of scrub of Kermadec ngaio, kawakawa, parapara and wharangi, and sedgeland/herbfield/vineland with Cyperus insularis, Kermadec ice plant, Hypolepis dicksonioides, Pteris comans, Sicyos australis, Carex forsteri, Canavalia rosea, Kermadec fireweed, Lepidium castellanum and L. oleraceum, Solanum nodiflorum, and Parietaria debilis interspersed with bare ground, bird burrows and guano deposits.
Historically present throughout all of the subtropical Kermadec Islands. Now largely restricted to the coastal fringe of the main Kermadec Islands and the dominant vegetation on smaller islets (e.g. Herald, Cheeseman and Hazard). Recovering on Raoul and Macauley Islands following rat (Rattus spp.) eradication.
Oliver (1910: 130–132). Includes rare ecosystems: seabird guano deposits; and seabird burrowed soil (Williams et al. 2007).
SA7: Ice plant, glasswort herbfield/loamfield
Mosaic of herbfield of glasswort, ice plant, pigweed, shore groundsel, sea primrose, shore celery, and Lepidium spp., and locally oioi, knobby clubrush, toetoe and Poa spp., with a scattered scrub/vineland of locally taupata, houpara, flax, ngaio, shrubby Melicytus, Hebe spp. and small-leaved pohuehue interspersed with bare ground, bird burrows and guano deposits.
Formerly widespread on suitable coastal sites throughout mainland New Zealand, from warm to mild climatic zones from Northland to Otago. Now largely restricted to predator-free offshore islands. Locally occurs in Northland–Auckland, where it is colloquially known as ‘petrel scrub’ Wright (1980).
Gillham (1960), Wright (1980), Partridge (1983) and de Lange et al. (1995). Includes rare ecosystems: seabird guano deposits; and seabird burrowed soil (Williams et al. 2007).
SA8: Mutton bird sedge, Senecio radiolatus sedgeland/herbfield/loamfield
Mosaic of sedgeland/herbfield of Carex trifida and Senecio radiolatus, with locally Leptinella featherstonii, Lepidium spp., Myosotidium hortensium and Embergeria grandifolia interspersed with bare ground, bird burrows and guano deposits.
Formerly widespread on suitable coastal sites throughout the Chatham Islands, but now largely restricted to predator-free offshore islands.
Equivalent to community type 13 of Kelly (1983). de Lange and Sawyer (2008). Includes rare ecosystems: seabird guano deposits; seabird burrowed soil; coastal rock stacks; and coastal cliffs on quartzose rocks (Williams et al. 2007).
Appendix 3—continued
Continued on next page

75Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
SA9: Olearia, Brachyglottis, Dracophyllum scrub/herbfield/loamfield [Mutton-bird scrub]
Mosaic of scrub of wind-shorn southern rātā, Hebe elliptica, Dracophyllum longifolium, and several species of Olearia and Brachyglottis, and locally broadleaf and māpou. Areas of coastal herbfield and bare ground are associated with penguin and seabird nesting/burrowing sites.
Fiordland, Foveaux Strait, Stewart Island/Rakiura and smaller islands as far south as the Auckland Islands. Most abundant on very exposed coastal hill slopes and smaller islands. Asteraceae scrub component is colloquially known as ‘mutton-bird scrub’.
Equivalent to community type 8 of Wilson (1987: 28–35). Cockayne (1909a: 213–218, 1909b), Fineran (1973), Johnson (1975), Wardle (1991), Allen et al. (2003: 161) and Barker (2003: chapter 11, 167–168, 271—fig.4, & 283–292). Includes rare ecosystems: coastal rock stacks; and coastal cliffs on quartzose rocks (Williams et al. 2007).
SA10: Poa litorosa grassland/herbfield/loamfield
Mosaic of tussockland dominated by Poa litorosa, P. foliosa, and a mega-herbfield of Anisotome latifolia and Bulbinella rossii, with localised areas of bare ground associated with penguin and seabird nesting sites.
Subantarctic islands, especially Campbell, Auckland and Snares Island groups.
Cockayne (1909a: 232–235) and Wardle (1991: 453–454). Includes rare ecosystems: seabird guano deposits; seabird burrowed soil; and marine mammal haul-outs (Williams et al. 2007).
SA11: Kirk’s scurvy grass herbfield/loamfield
Herbfield, loamfield with a sparse cover of least 15 native halophytic herbs and grasses, including Kirk’s scurvy grass.
Inland saline pans of Central and northern Otago (e.g. Waitaki). Soils described by Molloy (1998: 159–160 & 163). Extremely rare and threatened, with < 100 ha of a pre-development c. 40 000 ha remaining.
Wardle (1991: 299), Rogers et al. (2000) and Mark et al. (2003: 198). Includes rare ecosystem: inland saline (salt pans) (Williams et al. 2007).
UM1: Pōhutukawa, tānekaha forest/scrub/rockland
Mosaics of short forest, scrub and rockland of pōhutukawa, tānekaha, kānuka, karo, Astelia banksii, toetoe and harakeke, with a large number of associated local endemics.
Unique to the North Cape – Surville Cliffs ultramafic belt. Druce et al. (1979), Lee (1992: 392) and Wardle (1991: 397). Includes rare ecosystems: ultrabasic sea cliffs; and ultrabasic hills (Williams et al. 2007).
UM2: Conifer, beech, mānuka forest/scrub/rockland
Mosaics of short forest, scrub and rockland of a wide variety of podocarp trees (rimu, Hall’s tōtara, yellow silver pine, pink pine, silver pine and mountain celery pine), beech, southern rātā and kāmahi, interspersed with areas of low mānuka scrub and bare ground/rock.
Lowland and montane areas of Rangitoto ki te Tonga/D’Urville Island–Nelson and Southland–Otago ultramafic belts. Altitudinal range is from sea level to the natural low alpine treeline of ultramafic vegetation.
Lee (1992: 392–393 & 397–398). Includes rare ecosystems: ultrabasic sea cliffs; and ultrabasic hills (Williams et al. 2007).
UM3: Tussockland/stonefield/rockland
Stonefield, rockland of two local variants: 1. with occasional Chionochloa defracta tall tussock grassland, shrubland with mountain tauhinu, Dracophyllum pronum and D. filifolium; and 2. with locally mānuka, D. pronum, blue tussock, bristle tussock and Poa spp.
From the natural low alpine treeline of ultramafic vegetation to the alpine zone. Red Hills (Nelson), and West Dome and Red Mountain (South Westland).
Lee (1992: 394–396), Wardle (1991: 397–400), Mark et al. (2003: 217–218) and Allen et al. (2003: 169–170). Includes rare ecosystem: ultrabasic screes (Williams et al. 2007).
GT1: Geothermal kānuka scrub
Scrub of abundant geothermal kānuka grading into Campylopus spp. mossfield on heated ground. Locally includes areas of hydrothermally altered ground, mineral deposits and sinter terraces (now cool), dry eruption craters and old stream beds, with tall mingimingi and prickly mingimingi scrub on acidic soils. Often includes a margin of geothermal-influenced (predominantly kāmahi) forest.
Taupo Volcanic Zone. Wardle (1991: 400–404), Burns & Leathwick (1995), Burns (1997), Wildland Consultants (2004) and Boothroyd (2009). Includes rare ecosystems: heated ground (dry); and hydrothermally altered ground (now cool) (Williams et al. 2007).
Appendix 3—continued
Continued on next page

76Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
GT2: Geothermally-heated water and steam
Geysers, pools, springs/streams, fumaroles and sinter terraces (including their margins), and geothermal wetlands of a range of temperature, chemical and pH conditions, with associated microbes, cyanobacteria mats and bryophytes, and locally ferns and sedges.
Taupo Volcanic Zone and rare examples elsewhere (e.g. Ngawha). Cody (2007) and Boothroyd (2009). Includes rare ecosystems: geothermal streamsides; and fumaroles (Williams et al. 2007).
CV1: Subterranean rockland, stonefield [Caves]
Cave ecosystems dominated by a range of terrestrial and aquatic invertebrate species, both epigean and troglobitic.
Found in karst and psuedokarst areas, associated (in New Zealand) with carbonate (limestone, marble, dolomite) and recent basaltic geology. Fauna can be archipelago-like with local endemism across isolated karst blocks.
Includes rare ecosystems: caves and cracks in karst; sinkholes; cave entrances; and subterranean basalt fields (Williams et al. 2007).
VS1: Pōhutukawa scrub/forest
Pōhutukawa and pōhutukawa × northern rātā scrub/forest of a range of variants. Common associates on aa lava on Rangitoto include Peperomia urvilleana, karamū, Olearia furfuracea, koromiko, Kirk’s tree daisy, red māpou and Astelia banksii. Associates on scoria include mānuka, rewarewa, māhoe, akeake, tutu and red māpou. Common associates at Lake Tarawera include northern rātā, kāmahi, rewarewa and kānuka.
Primarily coastal northern North Island and marginal areas of the Rotorua Lakes, e.g. Mt Tarawera. Especially abundant following volcanism, e.g. basaltic eruptions in Auckland and andesitic eruptions, e.g. Whakaari/White Island. Also dominant on northern offshore islands, e.g. Poor Knights Islands and Mayor Island (Tuhua) following human decolonisation. Pōhutukawa is long-lived and locally can develop into near monocultures of tall forest that are capable of living for many centuries.
Atkinson & Percy (1956), Nicholls (1963), Whiting (1986), Clarkson (1990), Newnham & Lowe (1991), Atkinson (2004), Lindsay et al. (2009) and Wilmshurst et al. (2014).
VS2: Kānuka scrub/forest
Kānuka scrub/forest of a range of variants. Later successional transitions include a wide range of broadleaved and podocarp trees.
Semi-arid and sub-humid zones, especially on free-draining soils in the northern and eastern North Island and eastern South Island to Otago. Kānuka dominates in Otago where rainfall is < 650 mm per annum. Locally succeeds VS12: Sward grassland and VS11: Short tussock tussockland.
Equivalent to P16 type of Nicholls (1976); and alliance 24 of Wiser et al. (2011). Burrows (1973), Wardle (1991: 195–205) and Smale (1993, 1994).
VS3: Mānuka, kānuka scrub
Mānuka-kānuka scrub of a range of variants. Later successional transitions include a wide range of broadleaved and podocarp trees, and tree ferns.
Predominantly semi-arid and sub-humid zones, from Northland to Southland on free-draining soils. Locally succeeds VS12: Sward grassland and VS11: Short tussock tussockland.
Equivalent to alliances 5 and 24 of Wiser et al. (2011). Burrows (1973), Esler (1983), Wardle (1991: 195–205) and Smale (1994).
VS4: Mānuka scrub Mānuka scrub of a range of variants. Later successional transitions include a wide range of broadleaved and podocarp trees, and tree ferns.
Warm to cool sub-humid and humid zones, from Northland to Stewart Island/Rakiura on free- and poor-draining soils. Restricted to gley and organic soils in the semi-arid zone.
Equivalent to P17 type of Nicholls (1976); S2 Leptospermum scrub or fern of Newsome (1987); and alliance 24 of Wiser et al. (2011). Burrows (1973), Wardle (1991: 195–205) and Stephens et al. (2005).
VS5: Broadleaved species scrub/forest
Scrub/short forest of a wide range of variants, including species of Coprosma, Coriaria, Pittosporum, Pseudopanax, Melicytus, Olearia, Hebe and Myrsine, and wineberry, and locally kōtukutuku, kāmahi, tōwai, rewarewa, northern rātā and tree ferns.
Semi-arid to humid zones, from Northland to Stewart Island/Rakiura on free- and poor-draining soils. Often succeeds bracken fernland and/or mānuka scrub in humid climatic zones. On Mt Taranaki and Mt Tarawera, kāmahi and northern rātā have established terrestrially following volcanic activity, e.g. the Maeroa debris flow. Dominant species reflect local forest composition. Tree ferns can be locally abundant in humid locations.
Equivalent to S1 Mixed indigenous scrub of Newsome (1987); and alliance 23 of Wiser et al. (2011). Wardle (1966, 1991: 531–547) and Clarkson (1990).
Appendix 3—continued
Continued on next page

77Science for Conservation 325
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
VS6: Matagouri, Coprosma propinqua, kōwhai scrub [Grey scrub]
Scrub, vineland locally dominated by matagouri and/or Coprosma propinqua in association with a wide range of divaricating, small-leaved and liane species, locally including species of Carmichaelia, Coprosma, Olearia, Hebe, Pittosporum, Sophora, Ozothamnus, Coriaria, Muehlenbeckia, Rubus and Clematis, Aristotelia fruticosa, Corokia cotoneaster, mānuka and cabbage tree. Bracken and short tussock grasses often present.
Predominantly the eastern South Island in semi-arid and sub-humid zones on hill slopes and terraces. More restricted to cool zones in the western South Island and central North Island. Locally succeeds VS11: Short tussock tussockland. Represents an expansion of the range of unit TI4 onto a wider range of previously forested landforms.
Equivalent to GS7 Grassland and matagouri of Newsome (1987); and alliance 2 of Wiser et al. (2011). Wardle (1991: 207–211).
VS7: Mountain tauhinu, Dracophyllum rosmarinifolium scrub
Mountain tauhinu and Dracophyllum rosmarinifolium scrub, often associated with short and red tussock grasses, and a wide range of shrub and scrub species, including mānuka, kānuka, inaka, Dracophyllum acerosum, matagouri, and species of Hebe, Olearia and Coprosma.
Eastern South Island, from Marlborough to Otago. Represents an expansion of the range of unit CLF7 onto a wider range of previously forested landforms. Dracophyllum acerosum is locally abundant in Canterbury. Locally succeeds VS11: Short tussock tussockland and VS14: Tall tussock tussockland.
Equivalent to GS5 Grassland and Dracophyllum scrub of Newsome (1987); and alliance 3 of Wiser et al. (2011). Shanks et al. (1990).
VS8: Monoao scrub Monoao scrub, often associated with short and red tussock grasses, and a wide range of shrub and scrub species, including mānuka and kānuka, and locally inaka, species of Ozothamnus, Hebe, Olearia and Coprosma, and mountain celery pine.
Central North Island, especially within the Moawhango Ecological District and also on the summit of Mt Tarawera on the 1886 AD tephra. Historically, monoao occupied large areas of rhyolitic tephra within the Kaingaroa, Atiamuri and Taupo Ecological Districts. Represents an expansion of the range of units TI3 onto a wider range of previously forested landforms. Locally succeeds VS11: Short tussock tussockland and VS13: Red or copper tussock tussockland. Inaka and mountain celery pine can be co-dominant in montane areas, predominantly on andesitic ash.
Equivalent to GS5 Grassland and Dracophyllum scrub of Newsome (1987). Clarkson (1990), Smale (1990) and Rogers & Leathwick (1994).
VS9: Inaka scrub Inaka scrub, often associated with tall or red/copper tussock grasses, and a wide range of shrub and scrub species, including mānuka, kānuka, species of Ozothamnus, Hebe, Olearia, Coprosma and Gaultheria, and locally mountain celery pine.
Subalpine areas of the central North Island (e.g. northwest Ruahine Range) and locally elsewhere (e.g. Tararua Range). In the South Island, occurs in Otago and Southland (e.g. Maungatua Range). Represents an expansion of the range of unit CF13 onto a wider range of previously forested landforms. Locally succeeds VS11: Short tussock tussockland and VS13: Red or copper tussock tussockland.
Equivalent to GS4 Tussock grassland and subalpine scrub, and GS5 Grassland and Dracophyllum scrub of Newsome (1987). Atkinson (1981), Wardle (1991: 552), Rogers & Leathwick (1994) and Rogers & McGlone (1994).
VS10: Bracken fernland
Bracken fernland with a range of associates, including mānuka, kānuka and tutu, locally with matagouri, Coriaria sarmentosa, kōhūhū and five-finger.
Predominantly sub-humid to humid climatic zones, from Northland to Stewart Island/Rakiura and the Chatham Islands, though locally present in semi-arid regions. Still numerous in central North Island, western Otago and the Chatham Islands. Historically has been most abundant following large rhyolitic eruptions in the central North Island (e.g. Taupo eruption) and post Māori settlement, but has declined significantly.
Equivalent to alliance 4 of Wiser et al. (2011). Wardle (1991: 213–215) and McGlone et al. (2005).
Appendix 3—continued
Continued on next page

78Singers & Rogers—Classification of terrestrial ecosystem
s
ECOSYSTEM UNIT
CODE AND NAME
DESCRIPTION DISTRIBUTION (BOTH CURRENT AND HISTORIC),
WITH EXAMPLES AND COMMENTS
REFERENCES
VS11: Short tussock tussockland
Short tussock grasslands principally of silver and hard tussock, with associated species of Poa, Festuca, Deyeuxia and Rytidosperma, often with inter-tussock prostrate shrub and herbfield species. Later successional transitions include bracken, Dracophyllum spp., mānuka, kānuka and other shrub/scrub species.
Predominantly cool climatic areas of the eastern South Island, from Marlborough to north Otago. Very restricted in the central North Island, occurring on recent volcanic surfaces, especially in Tongariro National Park. Historically occupied large areas of the Canterbury Plains (Mark & McLennan 2005).
Equivalent to G3 Short tussock grassland of Newsome (1987). Clarkson (1990), Wardle (1991: 244–263), Mark & McLennan (2005) and Rogers et al. (2005: 55–56).
VS12: Sward grassland
Sward-forming grasslands of species of Poa, Microlaena, Festuca, Echinopogon, Hierochloe, Elymus, Stenostachys, Rytidosperma and Deyeuxia, locally with a wide range of herbaceous species. Later successional transitions include bracken, mānuka, kānuka and shrub/scrub species.
In the North Island, historically (c. 1840 AD) occurred on the Wairarapa, Heretaaunga/Ahuriri and Takapau/Ruataniwha Plains, and the sand plains of the Foxton Ecological District. Limited in extent in the South Island, mainly restricted to alluvial terraces of the West Coast. Now virtually extinct, having been replaced by exotic grasses.
Hill (1963), Wardle (1991: 263–264), Rogers (1994) and Mark & McLennan (2005).
VS13: Red or copper tussock tussockland
Tall red or copper tussock grassland of a wide range of variants in association with short tussock grasses and herbaceous species, including species of Aciphylla, Celmisia and Coriaria. Later successional transitions include a wide range of shrub/scrub species.
Predominantly central North Island, Mt Taranaki and southern South Island, mainly on poor-draining soils. Represents an expansion of the range of unit AL3 onto a wider range of previously forested or scrub landforms.
Equivalent to G6 red tussock grassland of Newsome (1987). Atkinson (1981), Clarkson (1990), Rogers (1994) and Mark & McLennan (2005).
VS14: Tall tussock tussockland
Tall tussock Chionochloa dominant grassland of several variants, including C. rigida, C. pallens and C. macra. These occur in association with short tussock grasses and herbaceous species, including species of Aciphylla and Celmisia. Later successional transitions include a wide range of shrub/scrub species.
Predominantly central North Island and eastern South Island. Represents an expansion of the range of units AL1 and AL2 onto a wider range of previously forested or scrub landforms.
Equivalent to G4 snow tussock grassland and G5 short tussock–snow tussock grassland of Newsome (1987). Mark & McLennan (2005).
Appendix 3—continued

79Science for Conservation 325
ReferencesAllen, R.B.; Cree, A.; Darby, J.; Davis, L.; Patrick, P.; Spencer, H. 2003: Forests and shrublands. Pp. 153−190 in Darby, J.;
Fordyce, R.E.; Mark, A.; Probert, K.; Townsend, C. (Eds): The natural history of southern New Zealand. University of Otago Press, Dunedin.
Atkinson, I.A.E. 1981: Vegetation map of Tongariro National Park, North Island, New Zealand. Scale 1:50 000. New Zealand Department of Scientific and Industrial Research, Wellington. 27 p.
Atkinson, I.A.E. 1985: Derivation of vegetation mapping units for an ecological survey of Tongariro National Park, North Island, New Zealand. New Zealand Journal of Botany 23: 361–378.
Atkinson, I.A.E. 2004: Successional processes induced by fires on the northern offshore islands of New Zealand. New Zealand Journal of Ecology 28: 181–193.
Atkinson, I.A.E.; Percy, C.A. 1956: An account of the vegetation of Mayor Island. Tane 7: 29–34.
Bagnall, R.G. 1975: Vegetation of the raised beaches at Cape Turakirae, Wellington, New Zealand. New Zealand Journal of Botany 13: 367–424.
Bagnall, R.G.; Ogle, C.C. 1981: The changing vegetation structure and composition of a lowland mire at Plimmerton, North Island, New Zealand. New Zealand Journal of Botany 19: 371–387.
Barker, M. 2003: The coast. Pp. 272−278 in Darby, J.; Fordyce, R.E.; Mark, A.; Prubert, K.; Townsend, C. (Eds): The natural history of southern New Zealand. University of Otago Press, Dunedin.
Barton, I.L. 1972: On the vegetation of the Hunua Ranges. New Zealand Journal of Botany 10: 8–26.
Bayfield, M.A.; Benson, M.A. 1986: Egmont Ecological Region. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 2. Department of Lands and Survey, Wellington. 85 p.
Bayfield, M.A.; Courtney, S.P.; Wiessing, M.I. 1991: North Taranaki District: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 16. Department of Conservation, Wanganui. 172 p.
Baylis, G.T.S. 1986: Widened seed dispersal at the Three Kings Islands attributed to naturalised birds. Pp. 41–45 in Wright, A.E.; Beever, R.E. (Eds): The offshore islands of northern New Zealand. New Zealand Department of Lands and Survey Information Series No. 16. Department of Lands and Survey, Wellington.
Beadel, S.M.; Bibby, J.C.; Perfect, A.J.; Rebergen, A.; Sawyer, J. 2004: Eastern Wairarapa Ecological District. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 23. Wellington Conservancy, Department of Conservation, Wellington. 382 p.
Bell, C.J.E. 1973: Mountain soils and vegetation in the Owen Range, Nelson: 2. The vegetation. New Zealand Journal of Botany 11: 73–102.
Boothroyd, I.K.G. 2009: Ecological characteristics and management of geothermal systems of the Taupo Volcanic Zone, New Zealand. Geothermics 38: 200–209.
Burns, B.R. 1997: Vegetation change along a geothermal stress gradient at the Te Kopia steamfield. Journal of the Royal Society of New Zealand 27: 279–294.
Burns, B.; Leathwick, J. 1995: Geothermal vegetation dynamics. Part I: Map of the geothermal vegetation of the Te Kopia Scenic Reserve. Part II: Plant species organisation along major environmental gradients. Science for Conservation 18. Department of Conservation, Wellington. 26 p.
Burns, B.R.; Leathwick, J.R. 1996: Vegetation-environment relationships at Waipoua Forest, Northland, New Zealand. New Zealand Journal of Botany 34: 79–92.
Burrows, C.J. 1969: Bog and flush vegetation. Pp. 117–132 in Knox, G.A. (Ed.): The natural history of Canterbury. A.H. & A.W. Reed, Wellington.
Burrows, C.J. 1973: The ecological niches of Leptospermum scoparium and L. ericoides (Angiospermae: Myrtaceae). Mauri Ora 1: 5–12.
Burrows, C.J. 1977: Alpine grasslands and snow in the Arthur’s Pass and Lewis Pass regions, South Island, New Zealand. New Zealand Journal of Botany 15: 665–686.
Burrows, C.J.; Dobson, A.T. 1972: Lakes Manapouri and Te Anau. Mires of the Manapouri–Te Anau lowlands. Proceedings of the New Zealand Ecological Society 19: 75–99.

80 Singers & Rogers—Classification of terrestrial ecosystems
Bussell, M.R. 1988: Mid and late Holocene pollen diagrams and Polynesian deforestation, Wanganui district, New Zealand. New Zealand Journal of Botany 26: 431–451.
Campbell, E.O. 1964: The restiad peat bogs at Motumaoho and Moanatuatua. Transactions of the Royal Society of New Zealand, Botany 2: 219–227.
Champion, P.D.; Beadel, S.M.; Dugdale, T.M. 2001: Turf communities of Lake Whangape and some potential management techniques. Science for Conservation 186. Department of Conservation, Wellington. 54 p.
Clarkson, B.D. 1986: Vegetation of Egmont National Park, New Zealand. National Parks Scientific Series Number 5. Department of Scientific and Industrial Research, Wellington. 95 p.
Clarkson, B.D. 1990: A review of vegetation development following recent (< 450 years) volcanic disturbance in North Island, New Zealand. New Zealand Journal of Ecology 14: 59–71.
Clarkson, B.D.; Clarkson, B.R. 1994: Ecology of an elusive endemic shrub, Pittosporum obcordatum Raoul. New Zealand Journal of Botany 32: 155–168.
Clarkson, B.D.; Daniel, L.J.; Overmars, F.B.; Courtney, S.P. 1986: Motu Ecological District: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 6. Department of Conservation, Wellington. 153 p.
Clarkson, B.R. 1984: Vegetation of three mountain mires west Taupo, New Zealand. New Zealand Journal of Botany 22: 361–375.
Clarkson, B.R. 1997: Vegetation recovery following fire in two Waikato peatlands at Whangamarino and Moanatuatua. New Zealand Journal of Botany 35: 167–179.
Clarkson, B.R.; Clarkson, B.D. 1991: Turanga Ecological District: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 14. Department of Conservation, Gisborne. 131 p.
Clarkson, B.R.; Schipper, L.A.; Lehmann, A. 2004: Vegetation and peat characteristics in the development of lowland restiad peat bogs, North Island, New Zealand. Wetlands 24: 133–151.
Clarkson, B.R.; Smale, M.C.; Williams, P.A.; Wiser, S.K.; Buxton, R.P. 2011: Drainage, soil fertility and fire frequency determine composition and structure of gumland heaths in northern New Zealand. New Zealand Journal of Ecology 35: 96–113.
Cockayne, L. 1901: A short account of the plant covering of the Chatham Islands. Transactions of the New Zealand Institute 34: 243–255.
Cockayne, L. 1909a: The ecological botany of the sub-antarctic islands of New Zealand. Pp. 182–235 in Chilton, C. (Ed.): The Subantarctic Islands of New Zealand. Volume 1. Philosophical Institute of Canterbury, Government Printer, Wellington.
Cockayne, L. 1909b: Report on a botanical survey of Stewart Island, New Zealand. Department of Lands, Government Printer, Wellington. 68 p.
Cockayne, L. 1911: Report on the dune areas of New Zealand, their geology, botany and reclamation. Department of Lands, Government Printer, Wellington. 74 p.
Cody, A.D. 2007: Geodiversity of geothermal fields in the Taupo Volcanic Zone. DOC Research and Development Series 281. Department of Conservation, Wellington. 70 p.
Collins, L.; Burns, B. 2001: The dynamics of Agathis australis-Nothofagus truncata forest in the Hapuakohe Ecological District, Waikato Region, New Zealand. New Zealand Journal of Botany 39: 423–433.
Conning, L. 2001: Northland Protection Strategy. Nature Heritage Fund, Wellington. 134 p.
Connor, H.E. 1991: Chionochloa Zotov (Gramineae) in New Zealand. New Zealand Journal of Botany 29: 219–282.
Connor, H.E.; Macrae, A.H. 1969: Montane and subalpine tussock grasslands in Canterbury. Pp. 167–204 in Knox, G.A. (Ed.): The natural history of Canterbury. A.H. & A.W. Reed, Wellington.
Courtney, S.; Arand, J. 1994: Balaclava, Sedgemere and Dillon Ecological Districts. Survey Report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 20. Department of Conservation, Nelson. 294 p.
Cox, J.E.; Mead, C.B. 1963: Evidence for ecologically significant changes in climate during the post-glacial period in New Zealand: soil evidence relating to post-glacial climate on the Canterbury Plains. Proceedings of the Ecological Society of New Zealand 10: 28–38.

81Science for Conservation 325
Cuddihy, M.J. 1983: Alpine grasslands of the Skippers Range. Southland Conservancy, New Zealand Forest Service, Invercargill. 15 p.
de Lange, P.J.; Cameron, E.K.; Taylor, G.A. 1995: Flora and fauna of Tatapihi (Groper) Island, Mokohinau Islands. Tane 35: 69–94.
de Lange, P.J.; Murray, B.G. 2008: Ranunculus ranceorum, a new name and rank for Ranunculus recens var. lacustris G.Simpson, an elusive, rarely seen buttercup of the Fiordland lakes, South Island, New Zealand. New Zealand Journal of Botany 46: 1–11.
de Lange, P.J.; Norton, D.A.; Crowcroft, G.M. 2000: Taxonomy, ecology, and conservation of Atriplex billardierei and A. hollowayi sp. nov. (Chenopodiaceae) in Australasia. New Zealand Journal of Botany 38: 551–567.
de Lange, P.J.; Rolfe, J.R. 2008: Hebe saxicola (Plantaginaceae)—a new threatened species from western Northland, North Island, New Zealand. New Zealand Journal of Botany 46: 531–545.
de Lange, P.J.; Sawyer, J.W.D. 2008: Flora of Western Reef, Chatham Islands, New Zealand. New Zealand Journal of Botany 46: 425–431.
Deng, Y.; Ogden, J.; Horrocks, M.; Anderson, S.H.; Nichol, S.L. 2004: The vegetation sequence at Whangapoua Estuary, Great Barrier Island, New Zealand. New Zealand Journal of Botany 42: 565–588.
Dickinson, K.J.M.; Mark, A.F.; Barratt, B.I.P.; Patrick, B.H. 1998: Rapid ecological survey, inventory and implementation: a case study from Waikaia Ecological Region, New Zealand. Journal of the Royal Society of New Zealand 28: 83–156.
Dobson, A.T. 1975: Vegetation of a Canterbury sub-alpine mire complex. Proceedings of the New Zealand Ecological Society 22: 67−75.
Dodson, J.R.; Enright, N.J.; McLean, R.F. 1988: Late Quaternary vegetation history for far northern New Zealand. Journal of Biogeography 15: 647−656.
Drobner, U.; Steel, J.B.; Smith, B.; Wilson, J.B. 1995: The sand dune vegetation of Chrystalls Beach, southern New Zealand, with particular reference to the cushion community. New Zealand Journal of Ecology 19: 143−151.
Druce, A.P.; Atkinson, I.A.E. 1958: Forest variation in the Hutt Catchment. Proceedings of the New Zealand Ecological Society 6: 41−45.
Druce, A.P.; Bartlett, J.K.; Gardner, R.O. 1979: Indigenous vascular plants of the Serpentine area of Surville Cliffs and adjacent cliff tops, north west of North Cape, New Zealand. Tane 25: 187–206.
Druce, A.P.; Williams, P.A. 1989: Vegetation and flora of the Ben More-Chalk Range area of southern Marlborough, South Island, New Zealand. New Zealand Journal of Botany 27: 167−199.
Druce, A.P.; Williams, P.A.; Heine, J.C. 1987: Vegetation and flora of Tertiary calcareous rocks in the mountains of western Nelson. New Zealand Journal of Botany 25: 41−78.
Ecroyd, C.E. 1982: Biological flora of New Zealand. 8. Agathis australis (D. Don) Lindl. (Araucariaceae) Kauri. New Zealand Journal of Botany 20: 17−36.
Elder, N.L. 1962: Vegetation of the Kaimanawa Ranges. Transactions of the Royal Society of New Zealand, Botany 2: 1−37.
Elliott, G.; Bellingham, M.; Davis, A. 1983: A biological survey of the Kaimaumau Swamp. New Zealand Wildlife Service, Wellington (unpublished). 18 p.
Emmett, D.K.; Smale, M.C.; Clarkson, B.D.; Leathwick, J.R.; Jessen, M.R.; Whaley, P.T. 2000: Indigneous vegetation of the Awhitu and Manukau Ecological Districts. Unpublished contract report prepared for Auckland Regional Council. Landcare Research, Hamilton. 181 p.
Eser, P. 1998: Ecological patterns and processes of the south Taupo wetland, North Island, New Zealand, with special reference to nature conservation management. Unpublished PhD thesis, Victoria University of Wellington, Wellington. 224 p.
Esler, A.E. 1969: Manawatu sand plain vegetation. Proceedings of the Ecological Society of New Zealand 16: 32−35.
Esler, A.E. 1970: Manawatu sand dune vegetation. Proceedings of the Ecological Society of New Zealand 17: 41−46.
Esler, A.E. 1978a: Botanical features of islands near the west coast of the Coromandel Peninsula, New Zealand. New Zealand Journal of Botany 16: 25−44.
Esler, A.E. 1978b: Botany of the Manawatu. Department of Scientific and Industrial Research Information Series No. 127. Government Printer, Wellington. 206 p.
Esler, A.E. 1983: Forest and scrubland zones of the Waitakere Range, Auckland. Tane 29: 109−117.

82 Singers & Rogers—Classification of terrestrial ecosystems
Esler, A.E.; Rumball, P.J. 1975: Gumland vegetation at Kaikohe, Northland, New Zealand. New Zealand Journal of Botany 13: 425−436.
Evans, G.R. 1969a: Alpine grasslands of the Takitimu Mountains. Protection Forestry Branch Report No. 54. New Zealand Forest Service, Forest Research Institute, Rotorua. 13 p.
Evans, G.R. 1969b: The alpine grasslands of the Taramakau River catchment. Protection Forestry Branch Report No. 61. New Zealand Forest Service, Forest Research Institute, Rotorua. 11 p.
Evans, G.R. 1972: The alpine grasslands of northern Fiordland. Protection Forestry Branch Report No. 92. New Zealand Forest Service, Forest Research Institute, Rotorua. 19 p.
Fineran, B.A. 1973: A botanical survey of seven mutton-bird islands, south-west Stewart Island. Journal of the Royal Society of New Zealand 3: 475−526.
Fisher, F.J.F. 1969: Rock, shingle-slide and riverbed. Pp. 106−116 in Knox, G.A. (Ed.): The natural history of Canterbury. A.H. & A.W. Reed, Wellington.
Franklin, D.A. 1965: The synecology of the Tararua indigenous forests. Silvicultural Report No. 54. Forest Research Institute, New Zealand Forest Service, Rotorua. 35 p.
Fuller, S.A.; Edwards, P.G. 1989: Southern Taumarunui. Biological Survey of Reserves Report No. 15. Department of Conservation, Wanganui. 9 p.
Gardner, R.O. 1989: Rheophytes in New Zealand. Auckland Botanical Society Journal 44: 22−24.
Garnock-Jones, P.J.; Bayly, M.J.; Lee, W.G.; Rance, B.D. 2000: Hebe arganthera (Scrophulariaceae), a new species from calcareous outcrops in Fiordland, New Zealand. New Zealand Journal of Botany 38: 379−388.
Gillham, M.E. 1960: Vegetation of Little Brother Island, Cook Strait, in relation to spray-bearing winds, soil salinity, and pH. Transactions of the Royal Society of New Zealand 88: 405–424.
Given, D.R. 1971: Montane-subalpine vegetation near Lake Shirley, Fiordland. New Zealand Journal of Botany 18: 1–13.
Grove, P. 1994: Hawkdun Ecological District. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 25. Department of Conservation, Dunedin. 157 p.
Harding, M.A. 1999: Southland Protection Strategy: a report to the Nature Heritage Fund Committee. Nature Heritage Fund, Wellington. 114 p.
Heenan, P.B.; Cameron, E.K. 2002: A new species of Olearia (Asteraceae) from Waima Forest, Northland, New Zealand. New Zealand Journal of Botany 40: 535–542.
Heenan, P.B.; de Lange, P.J.; Wilton, A.D. 2001: Sophora (Fabaceae) in New Zealand: taxonomy, distribution, and biogeography. New Zealand Journal of Botany 39: 17–53.
Hill, R.D. 1963: The vegetation of the Wairarapa in the mid-nineteenth century. Tuatara 11: 83–89.
Holloway, J.T. 1969: The native forests. Pp. 218–225 in Knox, G.A. (Ed.): The natural history of Canterbury. A.H. & A.W. Reed, Wellington.
Jelinek, L.R. 1980: Karamea forest association survey: 1978–1979. New Zealand Forest Service, Westport (unpublished). 118 p.
Johnson, P.N. 1972: Applied ecological studies of shoreline vegetation at Lakes Manapouri and Te Anau, Fiordland: Part 1. Vegetation of Lake Manapouri shoreline. Proceedings of the New Zealand Ecological Society 19: 102–119.
Johnson, P.N. 1975: Vegetation and flora of Solander islands, southern New Zealand. New Zealand Journal of Botany 13: 189–213.
Johnson, P.N. 1977: Vegetation at Ajax Hill, south east Otago, New Zealand. New Zealand Journal of Botany 15: 209–220.
Johnson, P.N. 1986: Manorburn wetlands: botanical report. DSIR Land Resources, Dunedin, for the Department of Lands and Survey, Dunedin (unpublished). 18 p.
Johnson, P.N. 1991: Ahuriri valley wetlands: botanical report. DSIR Land Resources, Dunedin, for the Department of Conservation, Canterbury (unpublished). 22 p.
Johnson, P.N.; Gerbeaux, P. 2004: Wetland types in New Zealand. Department of Conservation, Wellington. 184 p.
Johnson, P.N.; Rogers, G.M. 2002: Ephemeral wetlands and their turfs in New Zealand. Science for Conservation 230. Department of Conservation, Wellington. 109 p.
Kelly, G.C. 1983: Distribution and ranking of remaining areas of indigenous vegetation in the Chatham Islands (a map and accompanying notes). Botany Division, Department of Scientific and Industrial Research, Lower Hutt. 88 p.

83Science for Conservation 325
Lake, C.M.; Whaley, K.J. 1995: Rangitikei Ecological Region: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 32. Department of Conservation, Wanganui. 316 p.
Leathwick, J.R. 1987: Waipapa Ecological Area: a study of vegetation pattern in a scientific reserve. Forest Research Institute Bulletin No. 130. Ministry of Forestry, Rotorua. 82 p.
Leathwick, J.R.; Clarkson, B.D.; Burns, B.R.; Innes, J.G.; Smale, M.C. 1995: Waiapu Ecological District: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 31. Department of Conservation, Gisborne. 177 p.
Leathwick, J.R.; Wilson, G.; Rutledge, D.; Wardle, P.; Morgan, F.; Johnston, K.; McLeod, M.; Kirkpatrick, R. 2003: Land Environments of New Zealand. David Bateman Ltd, Auckland. 183 p.
Lee, W.G. 1992: New Zealand ultramafics. Pp. 375−418 in Roberts, B.A.; Proctor, J. (Eds): The ecology of areas with serpentinized rocks. A world view. Kluwer Academic Publishers, Netherlands.
Lindsay, H.; Wild, C.; Byers, S. 2009: Auckland protection priorities: prioritising the protection of biodiversity values in the Auckland Conservancy. Internal report. Department of Conservation, Wellington (unpublished). 81 p. (DOCDM-239922)
Logan, M.C.; Holloway, J.E. 1934: Plant succession on the Oreti River sand dunes. Transactions of the New Zealand Institute 61: 122−139.
Macmillan, B.H. 1979: Shoreline flora and vegetation of Lake Pukaki, south Canterbury, New Zealand. New Zealand Journal of Botany 17: 23–42.
Mark, A.F.; Bliss, L.C. 1970: The high-alpine vegetation of Central Otago, New Zealand. New Zealand Journal of Botany 8: 381−451.
Mark, A.F.; Dickinson, K.J.M. 1997: New Zealand alpine ecosystems. Pp. 311–345 in Wielgolaski, F.E. (Ed.): Ecosystems of the world, volume 3: polar and alpine tundra. Elsevier, Amsterdam.
Mark, A.F.; Grealish, G.; Ward, C.M.; Wilson, J.B. 1988: Ecological studies of a marine terraces sequence in Waitutu Ecological District of southern New Zealand. Part 1: The vegetation and soil patterns. Journal of the Royal Society of New Zealand 18: 29−58.
Mark, A.F.; Johnson, P.N.; Dickinson, K.J.M.; McGlone, M.S. 1995: Southern hemisphere patterned mires, with emphasis on southern New Zealand. Journal of the Royal Society of New Zealand 25: 23−54.
Mark, A.; Lee, W.; Patrick, B.; Cree, A.; Darby, J.; Spenser, H. 2003: Tussock grasslands and associated mountain lands. Pp. 191–235 in Darby, J.; Fordyce, R.E.; Mark, A.; Probert, K.; Townsend, C. (Eds): The natural history of southern New Zealand. University of Otago Press, Dunedin.
Mark, A.F.; McLennan, B. 2005: The conservation status of New Zealand’s indigenous grasslands. New Zealand Journal of Botany 43: 245−270.
Mark, A.F.; Porter, S.; Piggott, J.J.; Michel, P.; Dickinson, K.J.M. 2008: Altitudinal patterns of vegetation, flora, life forms, and environments in the alpine zone of the Fiord Ecological Region, New Zealand. New Zealand Journal of Botany 46: 205−237.
Mark, A.F.; Rawson, G.; Wilson, J.B. 1979: Vegetation pattern of a lowland raised mire in eastern Fiordland, New Zealand. New Zealand Journal of Ecology 2: 1−10.
Mark, A.F.; Smith, P.M.F. 1975: A lowland vegetation sequence in south Westland: Pakihi bog to mixed beech-podocarp forest. Part 1: The principal strata. Proceedings of the New Zealand Ecological Society 22: 76−92.
Maxwell, F.; Adams, J.; Walls, G. 1993: Eastern Hawke’s Bay Ecological District: survey report for the Protected Natural Areas Programme. Protected Natural Areas Programme Survey Report No. 28. Department of Conservation, Napier. 144 p.
McGlone, M.S. 2001: The origin of the indigenous grasslands of south-eastern South Island in relation to pre-human woody ecosystems. New Zealand Journal of Ecology 25: 1−15.
McGlone, M.S.; Basher, L.R. 1995: The deforestation of the upper Awatere catchment, Inland Kaikoura Range, Marlborough, South Island, New Zealand. New Zealand Journal of Ecology 19: 53–66.
McGlone, M.S.; Moar, N.T. 1998: Dryland Holocene vegetation history, Central Otago and the Mackenzie Basin, South Island, New Zealand. New Zealand Journal of Botany 36: 91−111.
McGlone, M.S.; Neall, V.E. 1994: The late Pleistocene and Holocene vegetation history of Taranaki, North Island, New Zealand. New Zealand Journal of Botany 32: 251–269.

84 Singers & Rogers—Classification of terrestrial ecosystems
McGlone, M.S.; Wilmshurst, J.M.; Leach, H.M. 2005: An ecological and historical review of bracken (Pteridium esculentum) in New Zealand, and its cultural significance. New Zealand Journal of Botany 29(2): 165–184.
McKelvey, P.J. 1963: The synecology of the West Taupo indigenous forests. Forest Research Institute, Government Printer, Wellington. 126 p.
McKelvey, P.J. 1973: The pattern of the Urewera forests. Forest Research Institute Technical Paper 59. Forest Research Institute, Rotorua. 47 p.
McKelvey, P.J. 1984: Provisional classification of South Island virgin indigenous forests. New Zealand Journal of Forestry Science 14: 151−178.
McKelvey, P.J.; Nicholls, J.L. 1959: The indigenous forest types of north Auckland. New Zealand Journal of Forestry 8(1): 29–45.
McQueen, A.A.M.; Wilson, J.B. 2000: Vegetation and environment of a New Zealand raised bog. Journal of Vegetation Science 11: 547–554.
Meurk, C. 1982: Alpine phytoecology of the rainshadow mountains of Otago and Southland, New Zealand. Unpublished PhD thesis, University of Otago, Dunedin. 774 p.
Meurk, C. 2008: Vegetation of the Canterbury plains and downlands. Pp. 195−250 in Winterbourn, M; Knox, G.; Burrows, C.; Marsden, I. (Eds): The natural history of Canterbury. Canterbury University Press, Christchurch.
Meurk, C.; Foggo, M.N.; Bastow-Wilson, J. 1994: The vegetation of subantarctic Campbell Island. New Zealand Journal of Ecology 18: 123−168.
Molloy, B.P.J. 1969: Recent history of the vegetation. Pp. 340−360 in Knox, G.A. (Ed.): The natural history of Canterbury. A.H. & A.W. Reed, Wellington.
Molloy, B.P.J.; de Lange, P.J.; Clarkson, B.D. 1999: Coprosma pedicellata (Rubiaceae), a new species from New Zealand. New Zealand Journal of Botany 37: 383−397.
Molloy, L.R. 1998: Soils of the New Zealand landscape: the living mantle. New Zealand Society of Soil Science, New Zealand Soil Bureau and Mallinson Rendel Publishers, Wellington. 239 p.
Moore, S. 1999: Hundalee Ecological District: survey report for the Protected Natural Areas Programme. Protected Natural Areas Programme Survey Report No. 43. Nelson/Marlborough Conservancy, Department of Conservation, Nelson. 257 p.
Morton, J.; Ogden, J.; Hughes, T. 1984: To save a forest—Whirinaki. The Whirinaki Forest Promotion Trust, Auckland. 111 p.
Newnham, R.M.; Lowe, D.J. 1991: Holocene vegetation and volcanic activity, Auckland isthmus, New Zealand. Journal of Quaternary Science 6: 177−193.
Newsome, P.F.J. 1987: The vegetation cover of New Zealand. Water & Soil Miscellaneous Publication No. 112. Ministry of Works and Development, Wellington. 153 p.
Nicholls, J.L. 1963: Volcanicity and indigenous vegetation in the Rotorua district. Proceedings of the New Zealand Ecological Society 10: 58–65.
Nicholls, J.L. 1976: A revised classification of the North Island indigenous forests. New Zealand Journal of Forestry 28: 105−132.
Norton, D.A.; Invercargill (N.Z.) City Council; Ernest New and Associates; Te Wai Pounamu Conservation 1996: Development and forest sustainability, Otatara, Invercargill. Te Wai Pounamu Conservation. Conservation Research. Consultancy & Management. Boffa Miskell Ltd, Christchurch. 27 p.
Ogden, J.; Caithness, T.A. 1982: The history and present vegetation of the macrophyte swamp at Pukepuke lagoon. New Zealand Journal of Ecology 5: 108−120.
Ogle, C.C. 1997: Sand movement and the protection of natural areas on Pouto Peninsula, Northland. Conservation Advisory Science Notes 145. Department of Conservation, Wellington. 24 p.
Oliver, R.B. 1910: The vegetation of the Kermadec Islands. Transactions and Proceedings of the New Zealand Institute 42: 118−175.
Overmars, F.B.; Kilvington, M.J.; Gibson, R.S.; Newell, C.L.; Rhodes, T.J. 1998: Ngakawau Ecological District. Survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 11. West Coast Conservancy, Department of Conservation, Hokitika. 177 p.

85Science for Conservation 325
Partridge, T.R. 1983: The vegetation of Wharekakahu, an offshore stack of the Otago coast. New Zealand Journal of Botany 21: 135−140.
Partridge, T.R.; Molloy, B.J.P. 1985: Botanical survey of the Wolds Wetlands, Mackenzie Basin. Botany Division, Department of Scientific and Industrial Research, Lincoln (unpublished). 31 p.
Partridge, T.R.; Wilson, J.B. 1988: Vegetation patterns in salt marshes of Otago, New Zealand. New Zealand Journal of Botany 26: 497−510.
Pegman, A.P.McK.; Ogden, J. 2005: Productivity-decomposition dynamics of Typha orientalis at Kaitoke Swamp, Great Barrier Island, New Zealand. New Zealand Journal of Botany 43: 779−789.
Pegman, A.P.McK.; Ogden, J. 2006: Productivity-decomposition dynamics of Baumea juncea and Gleichenia dicarpa at Kaitoke Swamp, Great Barrier Island, New Zealand. New Zealand Journal of Botany 44: 261−271.
Rapson, G.L.; Sykes, M.T.; Lee, W.G.; Hewitt, A.E.; Agnew, A.D.Q.; Wilson, J.B. 2006: Subalpine gully-head ribbon fens of the Lammerlaw and Lammermoor Ranges, Otago, New Zealand. New Zealand Journal of Botany 44: 351−375.
Ravine, D. 1991: Foxton Ecological District: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 19. Department of Conservation, Wanganui. 264 p.
Roberts, C.M.; Duncan, R.P.; Wilson, K.J. 2007: Burrowing seabirds affect forest regeneration, Rangatira Island, Chatham Islands, New Zealand. New Zealand Journal of Ecology 31: 208–222.
Rogers, G.M. 1994: North Island seral tussock grasslands 1. Origins and land-use history. New Zealand Journal of Botany 32: 271−286.
Rogers, G.M. 1999: Coastal turfs of mainland New Zealand: their composition, environmental character, and conservation needs. Science for Conservation 107. Department of Conservation, Wellington. 83 p.
Rogers, G.M.; Hewitt, A.; Wilson, J.B. 2000: Ecosystem-based conservation strategy for Central Otago’s saline patches. Science for Conservation 166. Department of Conservation, Wellington. 38 p.
Rogers, G.M.; Leathwick, J.R. 1994: North Island seral tussock grasslands 2. Autogenic succession: change of tussock grassland to shrubland. New Zealand Journal of Botany 32: 287−303.
Rogers, G.M.; McGlone, M.S. 1994: A history of Kaiparoro clearing and the limits of Nothofagus in the northern Tararua Range, New Zealand. New Zealand Journal of Botany 32: 463–482.
Rogers, G.M.; Walker, S.; Lee, W.G. 2005: The role of disturbance in dryland New Zealand: past and present. Science for Conservation 258. Department of Conservation, Wellington. 122 p.
Rogers, G.M.; Wiser, S.K. 2010: Environment, composition, and conservation of coastal turfs in mainland New Zealand. New Zealand Journal of Botany 48: 1−14.
Rose, A.B.; Pekelharing, C.J.; Platt, K.H. 1992: Magnitude of canopy dieback and implications for conservation of southern rata-kamahi (Metrosideros umbellata - Weinmannia racemosa) forests, central Westland, New Zealand. New Zealand Journal of Ecology 16: 23−32.
Roxburgh, S.H.; Wilson, B.; Gitay, H.; King, W.McG. 1994: Dune slack vegetation in southern New Zealand. New Zealand Journal of Ecology 18: 51−64.
Sandercock, H. 1987: Big Bay Wetlands, Pyke State Forest 25: a vegetation survey. Southland Conservancy, New Zealand Forest Service, Invercargill (unpublished). 20 p.
Sawyer, J.; Townsend, A.; Preddey, J. 1998: Eastern Wairarapa Ecological District Protected Natural Areas Programme reconnaissance survey. Wellington Conservancy, Department of Conservation, Wellington. 268 p.
Scott, D. 1977: Plant ecology above timber line on Mt Ruapehu, North Island, New Zealand. I Site factors and plant frequency. New Zealand Journal of Botany 15: 255−294.
Shanks, A.; Glenny, D.; Gibson, R.; Rosser, K.; Roozen, D.; Phillipson, S.; Steven, J.; Arand, J. 1990: Coleridge, Craigieburn and Cass Ecological Districts: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 10. Department of Conservation, Wellington. 306 p.
Singers, N.J.D.; Whyman, R.B. 2001: Natural areas survey of Winstones, Waimarino and Karioi Forests with recommendations for management in line with Forest Stewardship Accreditation. Department of Conservation, Turangi (unpublished). 95 p.
Smale, M.C. 1984: White Pine Bush—an alluvial kahikatea (Dacrycarpus dacrydioides) forest remnant, eastern Bay of Plenty, New Zealand. New Zealand Journal of Botany 22: 201−206.

86 Singers & Rogers—Classification of terrestrial ecosystems
Smale, M.C. 1990: Ecology of Dracopyllum subulatum heathland on frost flats at Rangitaiki and north Pureora, central North Island, New Zealand. New Zealand Journal of Botany 28: 225−248.
Smale, M.C. 1993: Sixth re-examination of permanent plots in secondary kanuka on Little Barrier Island. Tane 34: 107−122.
Smale, M.C. 1994: Structure and dynamics of kanuka (Kunzea ericoides var. ericoides) heaths on sand dunes in Bay of Plenty, New Zealand. New Zealand Journal of Botany 32: 441–452.
Smale, M.C.; Hall, G.M.J.; Gardener, R.O. 1996: Monitoring condition of sand dune kanuka forest at Woodhill. Science for Conservation 26. Department of Conservation, Wellington. 17 p.
Smith, S.M.; Lee, W.G. 1984: Vegetation and soil development on a Holocene river terrace sequence, Arawata Valley, South Westland, New Zealand. New Zealand Journal of Science 27: 187−196.
Stephens, J.M.C.; Molan, P.C.; Clarkson, B.D. 2005: A review of Leptospermum scoparium (Myrtaceae) in New Zealand. New Zealand Journal of Botany 43(2): 431–449.
Stewart, G.H.; Harrison, J.B.J. 1987: Plant communities, landforms, and soils of a geomorphically active drainage basin, Southern Alps, New Zealand. New Zealand Journal of Botany 25: 385−399.
Stowe, C.J. 2003: The ecology and ethnobotany of karaka (Corynocarpus laevigatus). Unpublished MSc thesis, University of Otago, Dunedin. 110 p.
Sykes, M.T.; Wilson, J.B. 1987: The vegetation of a New Zealand dune slack. Vegetatio 71: 13−19.
Sykes, W.R. 1977: Kermadec Islands flora. An annotated check list. Bulletin 219. Department of Scientific and Industrial Research, Christchurch. 216 p.
Tyrell, M.; Cutting, M.; Green, C.; Murdoch, G.; Denyer, K.; Jamieson, K. 1999: Hunua Ecological District: survey report for the Protected Natural Areas Programme. New Zealand Protected Natural Areas Programme Survey Report No. 17. Auckland Regional Council, Auckland. 256 p.
Ward, J. 1967: Studies in ecology on a shell barrier beach. Section 1: Physiography and vegetation of shell barrier beaches. Plant Ecology 14: 241−297.
Wardle, J. 1971: The forests and shrublands of the Seaward Kaikoura Range. New Zealand Journal of Botany 9: 269−292.
Wardle, J. 1984: The New Zealand beeches: ecology, utilisation and management. New Zealand Forest Service, Christchurch. 447 p.
Wardle, P. 1963: Vegetation studies on Secretary Island, Fiordland. Part 2: The plant communities. New Zealand Journal of Botany 1: 171−187.
Wardle, P. 1966: Biological flora of New Zealand. 1. Weinmannia racemosa Linn f. (Cunoniaceae) Kamahi. New Zealand Journal of Botany 4: 114−131.
Wardle, J. 1967: Vegetation of the Aorangi Range, southern Wairarapa. New Zealand Journal of Botany 5: 5−22.
Wardle, P. 1974: The kahikatea (Dacrycarpus dacrydioides) forest of south Westland. Proceedings of the New Zealand Ecological Society 21: 62−71.
Wardle, P. 1977: Plant communities of Westland National Park (New Zealand) and neighbouring lowland and coastal areas. New Zealand Journal of Botany 15: 323−398.
Wardle, P. 1980: Primary succession in Westland National Park and its vicinity, New Zealand. New Zealand Journal of Botany 18: 221−231.
Wardle, P. 1991: Vegetation of New Zealand. Cambridge University Press, Cambridge. 672 p.
Wardle, P; Johnson, P.N.; Buxton, R.P. 1986: Botany of Gorge River, South Westland. Botany Division, Department of Scientific and Industrial Research, Christchurch (unpublished). 72 p.
Wardle, P.; Mark, A.F. 1973: Vegetation and landscape of the West Cape district, Fiordland, New Zealand. New Zealand Journal of Botany 11: 599−626.
Wells, R.D.S.; Clayton, J.S.; De Winton, M.D. 1998: Submerged vegetation of Lakes Te Anau, Manapouri, Monowai, Hauroko, and Poteriteri, Fiordland, New Zealand. New Zealand Journal of Marine and Freshwater Research 32: 621−638.
Whaley, K.J.; Clarkson, B.D.; Emmett, D.K.; Innes, J.G.; Leathwick, J.R.; Smale, M.C.; Whaley, P.T. 2001: Tiniroto, Waihua, Mahia and Matawai Ecological Districts: survey report for the Protected Natural Areas Programme. East Coast/Hawke’s Bay Conservancy, Department of Conservation. 504 p.

87Science for Conservation 325
Whiting, D.C. 1986: Vegetation colonisation of Rangitoto Island: the role of crevice microclimate. Unpublished MSc thesis, University of Auckland, Auckland. 167 p.
Wildland Consultants 2004: Geothermal vegetation of the Waikato region—revised 2004. Contract report No. 896, prepared for Environment Waikato, Hamilton. Wildland Consultants, Rotorua (unpublished). 244 p.
Williams, P.A. 1975: Studies of the tall-tussock (Chionochloa) vegetation/soil systems of the southern Tararua Range, New Zealand. 2. The vegetation/soil relationships. New Zealand Journal of Botany 13: 269−303.
Williams, P.A. 1989: Vegetation of the inland Kaikoura Range, Marlborough. New Zealand Journal of Botany 27: 201−220.
Williams, P.A. 1991: Subalpine and alpine vegetation of granite ranges in western Nelson, New Zealand. New Zealand Journal of Botany 29: 317−330.
Williams, P.A. 1993: The subalpine and alpine vegetation on the central sedimentary belt of Paleozoic rocks in north-west Nelson, New Zealand. New Zealand Journal of Botany 31: 65−90.
Williams, P.A.; Courtney, S.; Glenny, D.; Hall, G.; Mew, G. 1990: Pakihi and surrounding vegetation in north Westland, South Island. Journal of the Royal Society of New Zealand 20: 179−203.
Williams, P.A.; Wiser, S.; Clarkson, B.; Stanley, M. 2007: New Zealand’s historical rare terrestrial ecosystems set in a physical and physiognomic framework. New Zealand Journal of Ecology 31: 119−128.
Wilmshurst, J.M.; Moar, N.T.; Wood, J.R.; Bellingham, P.J.; Findlater, A.M.; Robinson, J.J.; Stone, C. 2014: Use of pollen and ancient DNA as conservation baselines for offshore islands in New Zealand. Conservation Biology 28: 202−212.
Wilson, B.J.; Cullen, C. 1986: Coastal cliff vegetation of the Catlins region Otago, South Island, New Zealand. New Zealand Journal of Botany 24: 567−574.
Wilson, H.D. 1976: Vegetation of Mount Cook National Park. National Parks Authority Scientific Series Number 1. Department of Lands and Survey, Wellington. 138 p.
Wilson, H.D. 1987: Plant communities of Stewart Island. New Zealand Journal of Botany 25 (supplement): 1−81.
Wilson, H.D. 1992: Banks Ecological Region: Port Hills, Herbert and Akaroa Ecological Districts. New Zealand Protected Natural Areas Programme Survey Report No. 21. Department of Conservation, Christchurch. 342 p.
Wiser, S.K.; Buxton, R.P. 2009: Montane outcrop vegetation of Banks Peninsula, South Island, New Zealand. New Zealand Journal of Ecology 33: 164−176.
Wiser, S.K.; Hurst, J.M. 2008: Classification of New Zealand’s forest and shrubland communities based on a national plot sampling on an 8-km grid. Landcare Research Contract Report for the Department of Conservation (unpublished). 71 p.
Wiser, S.K.; Hurst, J.M.; Wright, E.F.; Allen, R.B. 2011: New Zealand’s forest and shrubland communities: a quantitative classification based on a nationally representative plot network. Applied Vegetation Science 14: 506−523.
Wraight, M.J. 1963: The alpine and upper montane grasslands of the Wairau River catchment, Marlborough. New Zealand Journal of Botany 1: 351−376.
Wright, A.E. 1980: Vegetation and flora of Fanal Island, Mokohinau group. Tane 26: 25−43.





























