Embed Size (px)
Citation preview

Science of the Total Environment 568 (2016) 118–123
Contents lists available at ScienceDirect
Science of the Total Environment
j ourna l homepage: www.e lsev ie r .com/ locate /sc i totenv
Effects of salt stress on eco-physiological characteristics in Robiniapseudoacacia based on salt-soil rhizosphere
Peili Mao a, Yujuan Zhang a, Banghua Cao a,⁎, Longmei Guo a, Hongbo Shao b,c,⁎, Zhenyu Cao a,Qiankun Jiang a, Xuan Wang a
a Key Laboratory of Agricultural Ecology and Environment, Shandong Agricultural University, Taian 271018, Chinab Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, Chinac Institute of Agro-biotechnology, Jiangsu Academy of Agricultural Sciences, Nanjing 210014,China
H I G H L I G H T S G R A P H I C A L A B S T R A C T
• Salt stress is the key limited factor ofplantation construction in the YellowRiver Delta.
• The growth of Robinia pseudoacaciaseedlings was significantly inhibited bysaline–alkali degrees.
• Eco-physiological adaptability of R.pseudoacaciawas determined by above-ground parts and root.
• To control water uptake of R.pseudoacacia seedling by rhizospherewas an important way under salt stress.
⁎ Corresponding authors.E-mail addresses: [email protected] (B. Cao), shao
http://dx.doi.org/10.1016/j.scitotenv.2016.06.0120048-9697/© 2016 Published by Elsevier B.V.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 5 May 2016Accepted 3 June 2016Available online 9 June 2016
Robinia pseudoacacia is the main arbor species in the coastal saline–alkali area of the Yellow River Delta. Becausemost studies focus on the aboveground parts, detailed information regarding root functioning under salinity isscare. Root traits of seedlings of R. pseudoacacia including morphological, physiological and growth propertiesunder four salinity levels (CK, 1‰, 3‰ and 5‰ NaCl) were studied by the pot experiments to better understandtheir functions and relationships with the shoots. The results showed that seedling biomass decreased by the re-duction of root, stem and leaf biomasswith the increase of salinity levels.With increasing salinity levels, total rootlength (TRL) and total root surface area (TRSA) decreased, whereas specific root length (SRL) and specific rootarea (SRA) increased. Salt stress decreased root activity (RA) and the maximum net photosynthetic rate(Amax) and increased the water saturation deficit (WSD) significantly in the body. Correlation analyses showedsignificantly correlations between root morphological and physiological parameters and seedling biomass andshoot physiological indexes. R. pseudoacacia seedlings could adapt to 1‰ salinity by regulating the root morphol-ogy and physiology, but failed in 5‰ salinity. How to adjust the water status in the body with decreasing wateruptake by roots was an important way for R. pseudoacacia seedlings to adapt to the salt stress.
© 2016 Published by Elsevier B.V.
Keywords:Salt toleranceBiomassRoot morphologyRoot physiologyMaximum net photosynthetic rateSalt-soil
[email protected] (H. Shao).

119l Environment 568 (2016) 118–123
1. Introduction
A comprehensive understanding of the pattern of forest growth isimportant from both the economic and the ecological points of view.In an even-aged plantation, the above-ground net primary productivityusually decreases with age following the canopy closure (Ryan et al.,1997). The mechanisms and potential causes conducive to the declinein above-ground net primary productivity have attracted considerableattentions from forest ecologists and resource managers during the re-cent decades, and have become a hot topic of research in recent years(Bond et al., 2007; Martínez-Vilalta et al., 2007; He et al., 2012; Baretet al., 2015). A large number of studies have attempted to explain thisdecline by photosynthetic physiology and the effects of stand age onsoil nutrient availability, on biomass allocation, and on stand structure(Mao et al., 2011). However, how below-ground biomass dynamicswith stand development is not clear at present (Ryan et al., 1997;Yuan and Chen, 2012).
Root production accounts for 33–67% of the global terrestrial net pri-mary production (Abramoff and Finzi, 2015), which plays a fundamen-tal role in terrestrial carbon cycling (Jackson et al., 1997). For anindividual tree, root biomass is roughly 18–45% of the total biomass(Fogel, 1983). Roots are predominantly responsible for the uptake ofwater and mineral nutrients from the soil (Norby et al., 2004), nutrientstorage and transport (Ouimet et al., 2008), etc. Through the same scioncultivar of Prunus persica growing on the rootstocks with differing size-controlling potentials, Solari et al. (2006) found that rootstocks had apronounced effect on shoot growth and photosynthetic capacity. Qi etal. (2012) thought Zea mays with greater root systems would lead tohigher yield. Moreover, root systems are highly dynamic over varioustemporal and spatial scales (Brassard et al., 2009), and are responsiveto changes in environmental conditions, such as soil nutrients(Kochsiek et al., 2013), moisture (Pedroso et al., 2014), salinity (Hill etal., 2013), etc.
Today, more than 7% of the world's land is salt-affected (Pitman andLäuchli, 2004). Salinization has become amajor concern throughout theworld (Liang et al., 2005; Setia et al., 2013) and especially in China (Liuet al., 2015). Roots are first affected by excess salinity (Córdoba et al.,2001; Cavalcanti et al., 2007). In general, plant root growth has beenfound to be reduced under salinity (Li et al., 2016; Farissi et al., 2013;Yan et al., 2016). Furthermore, many plant species decreased theirtotal root length under salinity (Gómez-Bellot et al., 2013; Álvarez andSánchez-Blanco, 2014; Li et al., 2016). However, some plants could par-tially compensate the decrease of root growth by increasing specificroot area (Rewald et al., 2011; Hill et al., 2013). Plant photosynthesisand growth are largely determined by their ability to take up waterunder salt stress (Farissi et al., 2013). Increasing root/shoot ratio has
P. Mao et al. / Science of the Tota
Fig. 1. Effect of salt treatment on biomass in seedlings of Robinia pseudoacacia. Values are thsignificant differences among salinity levels by LSD's test.
been found in many salt-stressed woody seedlings/saplings(Fernández-García et al., 2014; González et al., 2012; Álvarez andSánchez-Blanco, 2014), whichwas probably associatedwith the osmot-ic effect of salt stress (Fernández-García et al., 2014). Ranjit et al. (2016)suggested that roots and shoots had distinct mechanisms of responsesto salt stress. However, root performance under salt stress has rarelybeen conducted in tree species (Hill et al., 2013).
The YellowRiver Delta is the fastest growing delta in theworld and anotably land–ocean interaction region. Secondary salinization is one ofthe most important characteristics in this region and other coastalareas (Cruz-Fuentes et al., 2014). Moreover, salinization is a crucial fac-tor determining the vegetation distribution in the Yellow River Delta(Xiong et al., 2008). Vegetative bioremediation is an important reclama-tion approach on saline–alkali soils (Qadir and Oster, 2004). Robiniapseudoacacia plantation is themaximum area arbor forest in the YellowRiver Delta, exerting significant protective function in reducing naturaldisasters (Cao et al., 2012). However, R. pseudoacacia plantation gradu-ally showed obvious degradation for large areas and even the sheetdeath in the recent decades. Zhang and Xing (2009) thought that highsalt content of soil was an important factor for the degradation of R.pseudoacacia plantation. Many researchers have studied the salt toler-ance of R. pseudoacacia from seed germination (Cao et al., 2005), seed-ling growth (Meng et al., 2011), adult tree characteristics (Du et al.,2014a), etc. However, root performance under salt stress has rarelybeen addressed (Du et al., 2014b), which has been playing an importantrole in soil improvement (Qadir andOster, 2004). In this paper, wemea-sured the aboveground and underground characteristics of R.pseudoacacia seedlings under salt stress by pot experiments. The studyaimed at exploring the morphological and physiological responses ofseedling roots andwhether there were significant correlations betweenroot and shoot changes under salt stress.
2. Materials and methods
2.1. Study area
The study was carried out at the Forestry Experimental Station ofShandong Agricultural University, China (36°10′09″N, 117°09′24″E). Itis a warm temperate continental monsoon climate in East Asia. Themean annual temperature is 12.8 °C, and the minimal and maximumtemperature varied from −6.9 °C to 37.5 °C. The frost-free period is186.6 days with an effective accumulated temperature of about4283.1 °C. The average annual precipitation varies from 600 mm to700 mm, whereas the precipitation falls seldom in spring and mainlyin summer. The mean annual relative humidity is 65%.
e mean of three replicates and Bars represent SD. Different lowercase letters represent

Fig. 2. Effect of salt treatment on root/shoot ratio in seedlings of Robinia pseudoacacia.Values are the mean of three replicates and Bars represent SD. Different lowercaseletters represent significant differences among salinity levels by LSD's test.
120 P. Mao et al. / Science of the Total Environment 568 (2016) 118–123
2.2. Experimental design
This experiment was conducted at the greenhouse in the ForestryExperimental Station. On April 20, 2010, fifty current-year seedlings(mean height, 15 cm and ground diameter 2 cm) were selected andtransplanted to the plastic pots (one seedling per pot) of 0.26 m heightand 0.28 m diameter. The pots were filled with sand loam soil and pHwas 7.05. Then, these seedlings revived until they grew well on May20. All seedlings were watered well during this period.
Fig. 3. Effect of salt treatment on root characteristics in seedlings of Robinia pseudoacacia. Varepresent significant differences among salinity levels by LSD's test.
Thepots (one plant)were arranged according to a randomized blockdesign, with 10 individual replicates per group. Salt treatment wasstarted onMay 21 and ended on June 7. We used 1000mL of NaCl solu-tions containing 0mM(control, CK), 50mM, 150mMand 200mMNaClonce every 9 days to each pot, a total of 3 times until the final soil saltcontent of the 4 treatments was approximately equal to 0, 1‰, 3‰and 5‰. To avoid osmotic shock, the treatments of 150 mM and200 mM NaCl solutions were increased gradually by 50 mM every3 days in the first 9 days. On July 10, the experiment ended. Seedlingswere harvested and morphological and physiological parameters weremeasured.
2.3. Plant harvest and determination of morphological and physiologicalparameters
The mature and fine leaves of five seedlings were used to measurephotosynthesis. According to the light response curves, the saturatedlight intensity was 1400 μmolm−2·s−1. The maximum rate of photo-synthesis (Amax) was measured before the harvest using CIRAS-2 por-table photosynthesis system under the saturated irradiance. Then, theseedlings were harvested and separated into roots, stems and leaves.Roots were washed with distilled water, blotted dry on filter paper.
Subsequently, rootswere arranged and floated on shallowwater in aglass tray (24 cm × 32 cm), scanned (on an HP Scanjet 8200), and ana-lyzed with an image analyzer (Delta-T Area Meter Type AMB2; Delta-T
lues are the mean of three replicates and Bars represent SD. Different lowercase letters
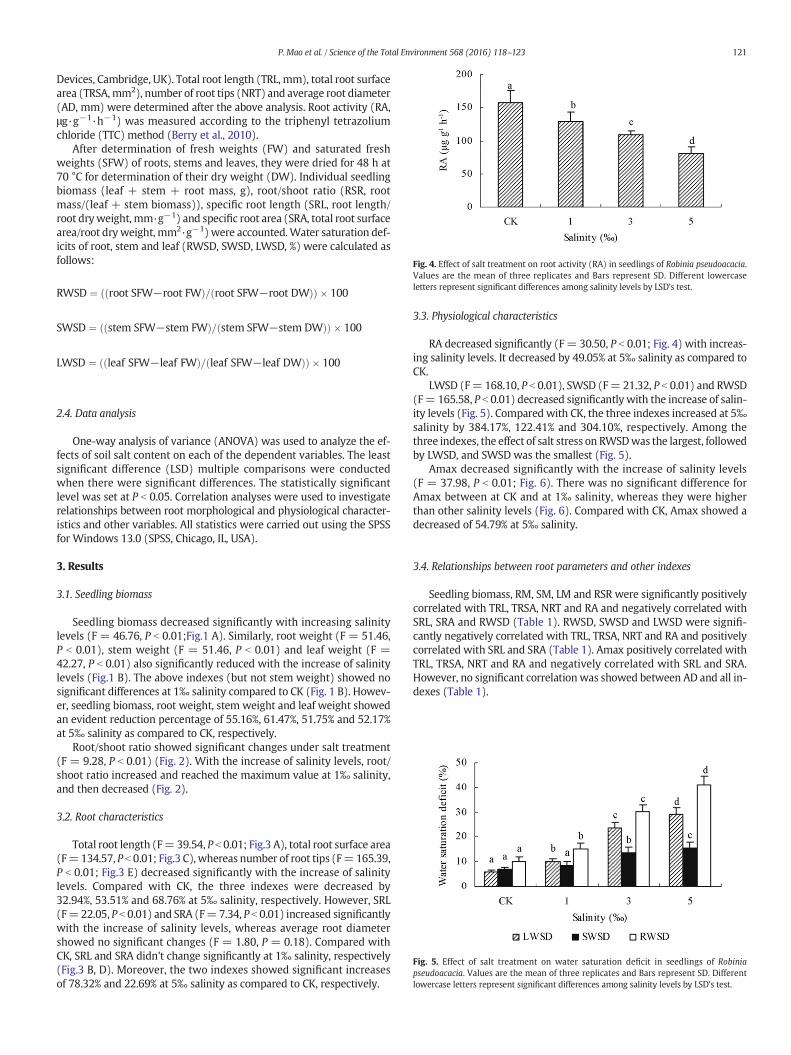
Fig. 5. Effect of salt treatment on water saturation deficit in seedlings of Robiniapseudoacacia. Values are the mean of three replicates and Bars represent SD. Differentlowercase letters represent significant differences among salinity levels by LSD's test.
Fig. 4. Effect of salt treatment on root activity (RA) in seedlings of Robinia pseudoacacia.Values are the mean of three replicates and Bars represent SD. Different lowercaseletters represent significant differences among salinity levels by LSD's test.
121P. Mao et al. / Science of the Total Environment 568 (2016) 118–123
Devices, Cambridge, UK). Total root length (TRL, mm), total root surfacearea (TRSA,mm2), number of root tips (NRT) and average root diameter(AD, mm) were determined after the above analysis. Root activity (RA,μg·g−1·h−1) was measured according to the triphenyl tetrazoliumchloride (TTC) method (Berry et al., 2010).
After determination of fresh weights (FW) and saturated freshweights (SFW) of roots, stems and leaves, they were dried for 48 h at70 °C for determination of their dry weight (DW). Individual seedlingbiomass (leaf + stem + root mass, g), root/shoot ratio (RSR, rootmass/(leaf + stem biomass)), specific root length (SRL, root length/root dryweight, mm·g−1) and specific root area (SRA, total root surfacearea/root dryweight, mm2·g−1) were accounted.Water saturation def-icits of root, stem and leaf (RWSD, SWSD, LWSD, %) were calculated asfollows:
RWSD ¼ root SFW−root FWð Þ= root SFW−root DWð Þð Þ � 100
SWSD ¼ stem SFW−stem FWð Þ= stem SFW−stem DWð Þð Þ � 100
LWSD ¼ leaf SFW−leaf FWð Þ= leaf SFW−leaf DWð Þð Þ � 100
2.4. Data analysis
One-way analysis of variance (ANOVA) was used to analyze the ef-fects of soil salt content on each of the dependent variables. The leastsignificant difference (LSD) multiple comparisons were conductedwhen there were significant differences. The statistically significantlevel was set at P b 0.05. Correlation analyses were used to investigaterelationships between root morphological and physiological character-istics and other variables. All statistics were carried out using the SPSSfor Windows 13.0 (SPSS, Chicago, IL, USA).
3. Results
3.1. Seedling biomass
Seedling biomass decreased significantly with increasing salinitylevels (F = 46.76, P b 0.01;Fig.1 A). Similarly, root weight (F = 51.46,P b 0.01), stem weight (F = 51.46, P b 0.01) and leaf weight (F =42.27, P b 0.01) also significantly reduced with the increase of salinitylevels (Fig.1 B). The above indexes (but not stem weight) showed nosignificant differences at 1‰ salinity compared to CK (Fig. 1 B). Howev-er, seedling biomass, root weight, stem weight and leaf weight showedan evident reduction percentage of 55.16%, 61.47%, 51.75% and 52.17%at 5‰ salinity as compared to CK, respectively.
Root/shoot ratio showed significant changes under salt treatment(F = 9.28, P b 0.01) (Fig. 2). With the increase of salinity levels, root/shoot ratio increased and reached the maximum value at 1‰ salinity,and then decreased (Fig. 2).
3.2. Root characteristics
Total root length (F=39.54, P b 0.01; Fig.3 A), total root surface area(F=134.57, P b 0.01; Fig.3 C), whereas number of root tips (F=165.39,P b 0.01; Fig.3 E) decreased significantly with the increase of salinitylevels. Compared with CK, the three indexes were decreased by32.94%, 53.51% and 68.76% at 5‰ salinity, respectively. However, SRL(F=22.05, P b 0.01) and SRA (F=7.34, P b 0.01) increased significantlywith the increase of salinity levels, whereas average root diametershowed no significant changes (F = 1.80, P = 0.18). Compared withCK, SRL and SRA didn't change significantly at 1‰ salinity, respectively(Fig.3 B, D). Moreover, the two indexes showed significant increasesof 78.32% and 22.69% at 5‰ salinity as compared to CK, respectively.
3.3. Physiological characteristics
RA decreased significantly (F= 30.50, P b 0.01; Fig. 4) with increas-ing salinity levels. It decreased by 49.05% at 5‰ salinity as compared toCK.
LWSD (F= 168.10, P b 0.01), SWSD (F= 21.32, P b 0.01) and RWSD(F= 165.58, P b 0.01) decreased significantly with the increase of salin-ity levels (Fig. 5). Compared with CK, the three indexes increased at 5‰salinity by 384.17%, 122.41% and 304.10%, respectively. Among thethree indexes, the effect of salt stress on RWSDwas the largest, followedby LWSD, and SWSD was the smallest (Fig. 5).
Amax decreased significantly with the increase of salinity levels(F = 37.98, P b 0.01; Fig. 6). There was no significant difference forAmax between at CK and at 1‰ salinity, whereas they were higherthan other salinity levels (Fig. 6). Compared with CK, Amax showed adecreased of 54.79% at 5‰ salinity.
3.4. Relationships between root parameters and other indexes
Seedling biomass, RM, SM, LM and RSR were significantly positivelycorrelated with TRL, TRSA, NRT and RA and negatively correlated withSRL, SRA and RWSD (Table 1). RWSD, SWSD and LWSD were signifi-cantly negatively correlated with TRL, TRSA, NRT and RA and positivelycorrelated with SRL and SRA (Table 1). Amax positively correlated withTRL, TRSA, NRT and RA and negatively correlated with SRL and SRA.However, no significant correlationwas showed between AD and all in-dexes (Table 1).

Fig. 6. Effect of salt treatment on the maximum net photosynthetic rate (Amax) inseedlings of Robinia pseudoacacia. Values are the mean of three replicates and Barsrepresent SD. Different lowercase letters represent significant differences among salinitylevels by LSD's test.
122 P. Mao et al. / Science of the Total Environment 568 (2016) 118–123
4. Discussion
Salinity has detrimental effects on the plant growth. Li et al. (2016)found that the root and shoot dry weight of Rosa chinensis decreasedas the salinity in irrigation water increased. Farissi et al. (2013) foundthat salt stress reduced the biomass ofMedicago sativa by the reductionof root and shoot dry biomass. In this paper, R. pseudoacacia seedlingsbiomass also decreased significantly with the increase of salinity levels.Root dry biomass decreased more obviously than that of stem and leafunder salinity stress, which suggested that the root had significant effecton the decrease of the seedling biomass of R. pseudoacacia. However,root biomass would not reduce always. The root biomass of Lawsoniainermis was not affected by salinity at 75 mM NaCl (Fernández-Garcíaet al., 2014). R. pseudoacacia seedlings also didn't show significant de-crease at 1‰ salinity level and root/shoot ratio increased significantly.Plant species could adapt the appropriate salinity range because theycould use some salt ions as osmotic adjustment substance (Koyro,2006). The increase of root/shoot ratio is a common response undersalt stress (Fernández-García et al., 2014; González et al., 2012;Álvarez and Sánchez-Blanco, 2014). This trait may favor to limit the ac-cumulation of toxic ions in the aerial parts (Munns and Tester, 2008). At5‰ salinity level, the root/shoot ratio of R. pseudoacacia decreased sig-nificantly, which was related to the severe reduction of root biomass.The similar result was also found by Fernández-García et al. (2014)and Glaeser et al. (2016). However, Li et al. (2016) didn't find the signif-icant change in root/shoot ration for R. chinensis, which may be con-trolled by the genes in plants (Coyle and Coleman, 2005).
Root morphology determines the ability of the plant to acquire soilresources (Costa et al., 2000; Shi et al., 2015). R. pseudoacacia seedlingsdecreased TRL and TRSA, which suggested that the ability of ion andwater uptake decreased (Wang et al., 2006). The decrease of total rootlength was also found in Euonymus plants (Gómez-Bellot et al., 2013),Callistemon citrinus (Álvarez and Sánchez-Blanco, 2014) and R. chinensis(Li et al., 2016). NRT decreased in saline conditions in this study, whichindicated that the production of new roots was hampered. This result
Table 1Correlations between root parameters and other measured variables under salinity.
TPM RM SM LM
TRL 0.95** 0.91** 0.95** 0.92**TRSA 0.98** 0.93** 0.97** 0.93**NRT 0.96** 0.90** 0.97** 0.92**AD −0.23 −0.22 −0.24 −0.24SRL −0.88** −0.93** −0.82** −0.70**SRA −0.64** −0.76** −0.54* −0.44RA 0.96** 0.92** 0.95** 0.88**RWSD −0.98** −0.95** −0.97** −0.92
** Correlation is significant at the 0.01 level; * correlation is significant at the 0.05 level.
was perhaps related to the additional carbon costs of roots to sustainwater uptake in saline conditions (Buwalda, 1993). However, SRL andSRA increased with the increase of salinity levels. Olive increased SRAin saline conditions (Rewald et al., 2011; Hill et al., 2013). Echeveria etal. (2008) found that salinity increased the SRL of Lotus glaber. IncreasedSRA and SRL may partially compensate for the loss in root biomassunder salinity (Hill et al., 2013), whereas it failed to adapt to the high sa-linity level for R. pseudoacacia. Rootmorphology (except AD)was signif-icantly correlated with seedling biomass, WSD and Amax, suggestingthat they were vital for R. pseudoacacia to adapt salt stress.
It is a relatively inexpensiveway for plants to alter root physiology toadapt to the external environments (Hutchings and John, 2003). RA andAmax decreased with the increase of salinity levels, indicating thatphysiological processes of R. pseudoacacia seedling were inhibited sig-nificantly. RA was significantly negatively correlated with RWSD,SWSD and LWSD. Therefore, the decrease of water uptake of roots dueto decreasing RA led to the deficits of R. pseudoacacia seedlings. An in-crease in the resistance towater flow from soil to plant is common in sa-line conditions (Farissi et al., 2013; Kosová et al., 2014; Li et al., 2016;Ranjit et al., 2016; Álvarez and Sánchez-Blanco, 2014). In this paper,roots suffered the most severe water deficit. High WSD reflect the lowlevel in relative water content that constitutes a determinant factor forthe metabolic activity (Farissi et al., 2013). Consequently, the reductionin Amax was because of the low level in relative water content in thebody of R. pseudoacacia. However, the reduction of water content inthe body could prevent salt ion reaching the photosynthetic tissues(Ranjit et al., 2016). A reduction in stomata conductance was also oneof themain causes of photosynthesis decline, which could favor to con-serve water status and prevent absorbing salt ion under salt stress(Fernández-García et al., 2014; Álvarez and Sánchez-Blanco, 2014).Therefore, the ability of water conservationwas crucial for plant speciesto adapt salt stress (Fernández-García et al., 2014).
5. Conclusions
Roots were the crucial organ for R. pseudoacacia seedlings to adaptsalt stress. R. pseudoacacia seedlings showed no significant decreaseon root biomass and increased root/shoot ratio at 1‰ salinity level,which suggested that it could adapt this low salinity level. Roots de-creased TRL and TRSA, leading to the decrease of the space of absorbingwater and ion. At the same time, the significant reduction of root activitydecreased the absorption capacity of water and ions, too. Althoughthere was significant water deficit due to the decrease of absorbingwater by roots, but the photosynthetic capacity didn't decreased. How-ever, the reductions of root biomass, root/shoot ratio, root activity andphotosynthetic rate at 5‰ salinity level indicated that roots failed toadapt to the high salinity level. Therefore, the manner that roots con-trolled water status in order to reduce the accumulation of salt ions inthe body by themorphological and physiological regulation was an im-portant adaptive strategy for R. pseudoacacia under salt stress.
Because roots of all sizes are inherently difficult to sample(Berhongaray et al., 2013), the structure and function of plant under-ground part is less understood than aboveground, and the mechanisms
RSR RWSD SWSD LWSD Amax
0.52* −0.96** −0.92** −0.94** 0.93**0.53* −0.99** −0.94** −0.99** 0.95**0.46* −0.97** −0.90** −0.97** 0.91**
−0.15 0.36 0.12 0.33 −0.25−0.87** 0.85** 0.83** 0.83** −0.87**−0.94** 0.58** 0.61** 0.54** −0.67**
0.54* −0.94** −0.92** −0.92** 0.93**−0.59** 1.00 0.94** 0.99** −0.96**

123P. Mao et al. / Science of the Total Environment 568 (2016) 118–123
that control fine root dynamics are still poorly understood (Yuan andChen, 2012). Moreover, roots play an important role on soil microbialcommunity and enzyme activity (Xiao et al., 2015) and soil improve-ment. Therefore, in the future, it is needed to explore the salt toleranceof the adult trees of R. pseudoacacia based on roots and soil microbialcommunity in the coastal saline soil of the Yellow River Delta, whichprovides a theoretical basis for the vegetation restoration in this area.
Conflict of interest
The authors declare that there are no conflicts of interest.
Acknowledgments
This study was supported by the National Natural ScienceFoundation of China (30950016), the Agricultural Science and Tech-nology Achievements Transformation Fund Project of China(2011GB2C600024) and Shandong Science and Technology Devel-opment Plan (2015GNC111026), the National Basic Research Pro-gram of China (2013CB430403), Shuangchuang Talent Plan ofJiangsu Province, and Jiangsu Autonomous Innovation of Agricultur-al Science & Technology [CX(15)1005].
References
Abramoff, R.Z., Finzi, A.C., 2015. Are above- and below-ground phenology in sync? NewPhytol. 205, 1054–1061.
Álvarez, S., Sánchez-Blanco, M.J., 2014. Long-term effect of salinity on plant quality, waterrelations, photosynthetic parameters and ion distribution in Callistemon citrinus. PlantBiol. 16, 757–764.
Baret, M., Pepin, S., Ward, C., Pothier, D., 2015. Long-term changes in belowground andaboveground resource allocation of boreal forest stands. For. Ecol. Manag. 350, 62–69.
Berhongaray, G., Janssens, I.A., King, J.S., Ceulemans, R., 2013. Fine root biomass and turn-over of two fast-growing poplar genotypes in a short-rotation coppice culture. PlantSoil 373, 269–283.
Berry, J.A., Beerling, D.J., Franks, P.J., 2010. Stomata: key players in the earth system, pastand present. Curr. Opin. Plant Biol. 107, 233–240.
Bond, B.J., Czarnomski, N.M., Cooper, C., Day, M.E., Greenwood, M.S., 2007. Developmentaldecline in height growth in Douglas-fir. Tree Physiol. 27, 441–453.
Brassard, B.W., Chen, H.Y.H., Bergeron, Y., 2009. Influence of environmental variability onroot dynamics in northern forests. Crit. Rev. Plant Sci. 28, 179–197.
Buwalda, J., 1993. The carbon costs of root systems of perennial fruit crops. Environ. Exp.Bot. 33, 131–140.
Cao, B., Zhai, M., Wu, L., 2005. Effects of pre-chilling on germination of Robiniapseudoacacia under salt stress. J. Beijing For. Univ. 27, 39–42 (in Chinese).
Cao, B., Zhang, Y., Mao, P., Li, C., 2012. Formation causes of wind damage to Robiniapseudoacacia plantation in Yellow River Delta. Chin. J. Appl. Ecol. 23, 2049–2054 (inChinese).
Cavalcanti, F.R., Lima, J.P.M., Ferreira-Silva, S.L., Viegas, R.A., Silveira, J.A.G., 2007. Roots andleaves display contrasting oxidative response during salt stress and recovery in cow-pea. J. Plant Physiol. 164, 591–600.
Córdoba, A., Seffino, L.G., Moreno, H., Arias, C., Grunberg, K., Zenoff, A., Taleisnik, E., 2001.Characterization of the effect of high salinity on roots of Chloris gayana Kunth carbo-hydrate and lipid accumulation and growth. Grass Forage Sci. 56, 162–168.
Costa, C., Dwyer, L.M., Hamilton, R.I., Hamel, C., Nantais, L., Smith, D.L., 2000. A samplingmethod for measurement of large root systems with scanner-based image analysis.Agron. J. 92, 621–627.
Coyle, D.R., Coleman, M.D., 2005. Forest production responses to irrigation and fertiliza-tion are not explained by shifts in allocation. For. Ecol. Manag. 208, 137–152.
Cruz-Fuentes, T., Cabrera, M.C., Heredia, J., Custodio, E., 2014. Groundwater salinity andhydrochemical processes in the volcano-sedimentary aquifer of La Aldea, GranCanaria, Canary Islands, Spain. Sci. Total Environ. 484, 154–166.
Du, Z.,Ma, H., Liu, F.,Ma, B., Xing, S., 2014a. Growth status and leaf characteristics of long-termlocust plantations in coastal saline soil. Biol. Disaster Sci. 37, 109–113 (in Chinese).
Du, Z., Liu, F., Ma, B., Dong, H., Ma, H., 2014b. Root distribution and fine root growth inmixed plantation of Robinia pseudoacacia and Fraxinus velutina in coastal saline–alkaliarea. Sci. Silvae Sin. 50, 10–15 (in Chinese).
Farissi, M., Ghoulam, C., Bouizgaren, A., 2013. Changes inwater deficit saturation and pho-tosynthetic pigments of alfalfa populations under salinity and assessment of prolinerole in salt tolerance. Agric. Sci. Res. J. 3, 29–35.
Fernández-García, N., Olmos, E., Bardisi, E., Garma, G.D.L., López-Berenguer, C., Rubio-Asensio, J.S., 2014. Intrinsic water use efficiency controls the adaptation to high salinityin a semi-arid adapted plant, henna (Lawsonia inermis L.). J. Plant Physiol. 171, 64–75.
Fogel, R., 1983. Root turnover and productivity of coniferous forest. Plant Soil 71, 75–85.Glaeser, L.C., Vitt, D.H., Ebbs, S., 2016. Responses of the wetland grass, Beckmannia
syzigachne, to salinity andsoil wetness: consequences for wetland reclamation inthe oil sandsarea of Alberta, Canada. Ecol. Eng. 86, 24–30.
Gómez-Bellot, M.J., Álvarez, S., Castillo, M., Bañón, S., Ortuño, M.F., Sánchez-Blanco, M.J.,2013. Water relations, nutrient content and developmental responses of Euonymus
plants irrigated with water of different degrees of salinity and quality. J. Plant Res.126, 567–576.
González, P., Syvertsen, J.P., Etxeberria, E., 2012. Sodium distribution in salt-stressed citrusrootstock seedlings. Hortic. Sci. 47, 1504–1511.
He, L., Chen, J.M., Pan, Y., Birdsey, R., Kattge, J., 2012. Relationships between net primary pro-ductivity and forest stand age in U.S. forests. Glob. Biogeochem. Cycles 26, GB3009.
Hill, A., Rewald, B., Rachmilevitch, S., 2013. Belowground dynamics in two olive varietiesas affected by saline irrigation. Sci. Hort-amsterdam 162, 313–319.
Hutchings, M.J., John, E.A., 2003. Distribution of Roots in the Soil, and Root Foraging Activ-ity. In: de Kroon, H., Visser, E.J.W. (Eds.), Root Ecology. Springer, pp. 33–60.
Jackson, R.B., Mooney, H.A., Schulze, E.D., 1997. A global budget for fine root biomass, sur-face area, and nutrient contents. Proc. Natl. Acad. Sci. U. S. A. 94, 7362–7366.
Kochsiek, A., Tan, S., Russo, S.E., 2013. Fine root dynamics in relation to nutrients in oligo-trophic Bornean rain forest soils. Plant Ecol. 214, 869–882.
Kosová, K., Vítámvás, P., Hlaváčková, I., Urban, M.O., Vlasáková, E., Prášil, I.T., 2014. Re-sponses of two barley cultivars differing in their salt tolerance to moderate andhigh salinities and subsequent recovery. Biol. Plant. 59 (1), 106–114.
Koyro, H.W., 2006. Effect of salinity on growth, photosynthesis, water relations and solutecomposition of the potential cash crop halophyte Plantago coronopus (L.). Environ.Exp. Bot. 56, 136–146.
Li, X., Wan, S., Kang, Y., Chen, X., Chu, L., 2016. Chinese rose (Rosa chinensis) growth andion accumulation underirrigation with waters of different salt contents. Agric. WaterManag. 163, 180–189.
Liang, Y., Si, J., Nikolic, M., Peng, Y., Chen, W., Jiang, Y., 2005. Organic manure stimulatesbiological activity and barley growth in soil subject to secondary salinization. SoilBiol. Biochem. 37, 1185–1195.
Liu, L., Long, X., Shao, H., Liu, Z., Tao, Y., Zhou, Q., Zong, J., 2015. Ameliorants improve saline–alkaline soils on a large scale in northern Jiangsu Province, China. Ecol. Eng. 81, 328–334.
Mao, P., Cao, B., Tian, W., Meng, F., 2011. Advances in research on the mechanisms of age-related productivity decline of planted forests. Acta Ecol. Sin. 31, 3208–3214 (inChinese).
Martínez-Vilalta, J., Vanderklein, D., Mencuccini, M., 2007. Tree height and age-related de-cline in growth in Scots pine (Pinus sylvestris L.). Oecologia 150, 529–544.
Meng, F., Pang, H., Wang, J., Li, S., Wang, Y., 2011. The ultrastructure of chloroplast in me-sophyll cell on two robinias under NaCl and Na2SO4 stress. Acta Ecol. Sin. 31, 734–741(in Chinese).
Munns, R., Tester, M., 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59,651–681.
Norby, R.J., Ledford, J., Reilly, C.D., Miller, N.E., O'Neill, E.G., 2004. Fine-root productiondominates response of a deciduous forest to atmospheric CO2 enrichment. Proc.Natl. Acad. Sci. U. S. A. 101, 9689–9693.
Ouimet, R., Camiré, C., Brazeau,M., More, J.D., 2008. Estimation of coarse root biomass andnutrient content for sugar maple, jack pine, and black spruce using stem diameter atbreast height. Can. J. For. Res. 38, 92–100.
Pedroso, F.K.J.V., Prudente, D.A., Bueno, A.C.R., Machado, E., Ribeiro, R.V., 2014. Droughttolerance in citrus trees is enhanced by rootstock-dependent changes in root growthand carbohydrate availability. Environ. Exp. Bot. 101, 26–35.
Pitman,M., Läuchli, A., 2004. Global Impact of Salinity andAgricultural Ecosystems. In: Pitman,M., Läuchli, A. (Eds.), Salinity: Environment – Plants –Molecules. Springer, pp. 3–20.
Qadir,M., Oster, J.D., 2004. Crop and irrigationmanagement strategies for saline-sodic soils andwaters aimed at environmentally sustainable agriculture. Sci. Total Environ. 323, 1–19.
Qi, W., Liu, H., Liu, P., Dong, S., Zhao, B., So, H.B., Li, G., Liu, H., Zhang, J., Zhao, B., 2012. Mor-phological and physiological characteristics of corn (Zea mays L.) roots from cultivarswith different yield potentials. Europ. J Agronomy 38, 54–63.
Ranjit, S.L., Manish, P., Penna, S., 2016. Early osmotic, antioxidant, ionic, and redox re-sponses to salinity in leaves and roots of Indian mustard (Brassica juncea L.).Protoplasma 253, 101–110.
Rewald, B., Rachmilevitchb, S., McCuec, M.D., Ephrathb, J.E., 2011. Influence of saline drip-irrigation on fine root and sap-flow densities of two mature olive varieties. Environ.Exp. Bot. 72, 107–114.
Ryan, M.G., Binkley, D., Fownes, J.H., 1997. Age-related decline in forest productivity: pat-tern and process. Adv. Ecol. Res. 27, 213–262.
Setia, R., Gottschalk, P., Smith, P., Marschner, P., Baldock, J., Setia, D., Smith, J., 2013. Soilsalinity decreases global soil organic carbon stocks. Sci. Total Environ. 465, 267–272.
Shi, F., Song, C., Zhang, X., Mao, R., Guo, Y., Gao, F., 2015. Plant zonation patterns reflectedby the differences in plant growth, biomass partitioning and root traits along a waterlevel gradient among four common vascular plants in freshwater marshes of theSanjiang Plain, Northeast China. Ecol. Eng. 81, 158–164.
Solari, L., Johnson, S., Dejong, T.M., 2006. Hydraulic conductance characteristics of peach(Prunus persica) trees on different rootstocks are related to biomass production anddistribution. Tree Physiol. 26, 1343–1350.
Wang, H., Inukai, Y., Yamauchi, A., 2006. Root development and nutrient uptake. Crit. Rev.Plant Sci. 25, 279–301.
Xiao, Y., Huang, Z., Lu, X., 2015. Changes of soil labile organic carbon fractions and theirrelation to soil microbial characteristics in four typical wetlands of Sanjiang Plain,Northeast China. Ecol. Eng. 82, 381–389.
Xiong, X., He, Q., Cui, B.S., 2008. Double principal coorclinate analysis of herbaceous veg-etation in wetlands of the Yellow River Delta, China. Chin. J. Ecol. 27, 1631–1638 (inChinese).
Yan, K., Xu, H., Zhao, S., Shan, J., Chen, X., 2016. Saline soil desalination by honeysuckle(Lonicera japonicaThunb.) dependson salt resistancemechanism. Ecol. Eng. 88, 226–231.
Yuan, Z.Y., Chen, H.Y.H., 2012. Fine root dynamics with stand development in the borealforest. Funct. Ecol. 26, 991–998.





























