Embed Size (px)
Citation preview

1521-0103/350/3/624–634$25.00 http://dx.doi.org/10.1124/jpet.114.214890THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 350:624–634, September 2014Copyright ª 2014 by The American Society for Pharmacology and Experimental Therapeutics
Searching for Combinations of Small-Molecule Correctors toRestore F508del–Cystic Fibrosis Transmembrane ConductanceRegulator Function and Processing
Clément Boinot, Mathilde Jollivet Souchet, Romain Ferru-Clément, and Frédéric BecqLaboratoire Signalisation et Transports Ioniques Membranaires, Université de Poitiers, Centre National de la RechercheScientifique (CNRS), Poitiers, France
Received March 24, 2014; accepted June 23, 2014
ABSTRACTThe mutated protein F508del–cystic fibrosis transmembraneconductance regulator (CFTR) failed to traffic properly as a re-sult of its retention in the endoplasmic reticulum and functionsas a chloride (Cl2) channel with abnormal gating and endocytosis.Small chemicals (called correctors) individually restore F508del-CFTR trafficking and Cl2 transport function, but recent findingsindicate that synergistic pharmacology should be considered toaddress CFTR defects more clearly. We studied the function andmaturation of F508del-CFTR expressed in HeLa cells using acombination of five correctors [miglustat, IsoLAB (1,4-dideoxy-2-hydroxymethyl-1,4-imino-L-threitol), Corr4a (N-[2-(5-chloro-2-methoxy-phenylamino)-49-methyl-[4,59]bithiazolyl-29-yl]-benzamide),VX-809 [3-(6-(1-(2,2-difluorobenzo[d][1,3]dioxol-5-yl)cyclo-propanecarboxamido)-3-methylpyridin-2-yl)benzoic acid], andsuberoylamilide hydroxamic acid (SAHA)]. Using the whole-cellpatch-clamp technique, the current density recorded in responseto CFTR activators (forskolin 1 genistein) was significantly in-creased in the presence of the following combinations: VX-8091IsoLAB; VX-809 1 miglustat 1 SAHA; VX-809 1 miglustat 1
IsoLAB; VX-809 1 IsoLAB 1 SAHA; VX-809 1 miglustat 1IsoLAB 1 SAHA. These combinations restored the activity ofF508del-CFTR but with a differential effect on the appearance ofmature c-band of F508del-CFTR proteins. Focusing on theVX-8091 IsoLAB cocktail, we recorded a level of correction higherat 37°C versus room temperature, but without amelioration ofthe thermal instability of CFTR. The level of functional rescuewith VX-809 1 IsoLAB after 4 hours of incubation was maximaland similar to that obtained in optimal conditions of use for eachcompound (i.e., 24 hours for VX-809 1 4 hours for IsoLAB).Finally, we compared the stimulation of F508del-CFTR by forskolinor forskolin 1 VX-770 [N-(2,4-di-tert-butyl-5-hydroxyphenyl)-4-oxo-1,4-dihydroquinoline-3-carboxamide] with cells correctedby VX-809 1 IsoLAB. Our results open new perspectives forthe development of a synergistic polypharmacology to rescueF508del-CFTR and show the importance of temperature on theeffect of correctors and on the level of correction, suggestingthat optimized combination of correctors could lead to a betterrescue of F508del-CFTR function.
IntroductionCystic fibrosis (CF) [Mendelian Inheritance in Man (MIM)
no. 219700] is an autosomal-recessive, lethal disorder thatresults from mutations introduced in the gene encoding themultifunctional cAMP-dependent chloride ion channel CFtransmembrane conductance regulator (CFTR), ATP-bindingcassette subfamily C, member 7. Physiologically, CFTR con-trols salt and fluid transport in response to cAMP-dependentagonists across the apical membrane of most epithelial tis-sues, including the airway epithelia (Riordan, 1993). In CF
patients, this function is greatly reduced or absent. For theseindividuals, the consequences are severe, in particular in therespiratory and gastrointestinal tracts, where viscous secre-tions favor the development of inflammatory injury (Flumeet al., 2010).More than 1900 different mutations in the CFTR gene (MIM
no. 602421) have been identified in CF patients worldwide.These mutations are divided into six classes according to theirdegree of severity in CF disease and the mechanism that dis-rupts CFTR function (Welsh and Smith., 1993; Haardt et al.,1999). About 90% of people with CF have at least one copy ofthe F508del mutation, and more than 70% are homozygousfor it. This deletion leads to a CFTR misfolding that inducesendoplasmic reticulum (ER) retention (Cheng et al., 1990), andexported mutated proteins suffer from abnormal plasma mem-brane trafficking, endocytosis, and gating. The half-life of
This work was supported by grants from Vaincre La Mucoviscidose (Paris,France) and Université de Poitiers. C.B. was supported by a Ph.D. fellowshipfrom Vaincre La Mucoviscidose. M.J.S. was supported by a donation from thecharities “Mucovie” (Perpignan, France) and “Amandine contre la mucovisci-dose” (Dijon, France).
dx.doi.org/10.1124/jpet114.214890.
ABBREVIATIONS: CF, cystic fibrosis; CFTR, cystic fibrosis transmembrane conductance regulator; Corr4a, N-[2-(5-chloro-2-methoxy-phenyl-amino)-49-methyl-[4,59]bithiazolyl-29-yl]-benzamide; DIDS, 4,49-diisothiocyanatostilbene-2,29-disulfonic acid disodium salt hydrate; ER, endoplasmicreticulum; F, forskolin; G, genistein; HDAC, histone deacetylase; IsoLAB, 1,4-dideoxy-2-hydroxymethyl-1,4-imino-L-threitol; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; NBD, nucleotide binding domain; PBS, phosphate-buffered saline; RT, room temperature; SAHA, suberoylamilidehydroxamic acid; VX-770, N-(2,4-di-tert-butyl-5-hydroxyphenyl)-4-oxo-1,4-dihydroquinoline-3-carboxamide; VX-809, 3-(6-(1-(2,2-difluorobenzo[d][1,3]dioxol-5-yl)cyclopropanecarboxamido)-3-methylpyridin-2-yl)benzoic acid; WT, wild-type.
624
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from

F508del-CFTR proteins in the plasma membrane is alsoshorter than that of wild-type (WT) CFTR (Haardt et al., 1999;Heda et al., 2001), and the open probability of the channel isreduced as the result of an increase in the closed time overthat of the WT channel (Dalemans et al., 1991; Wang et al.,2000). Thermal inactivation or instability of the mutantF508del-CFTR at 37°C was recently noted in mammaliancells (Wang et al., 2011; He et al., 2013) and Xenopus oocytes(Liu et al., 2012) (i.e., F508del-CFTR channel activity inac-tivated rapidly at physiologic temperature).A number of therapeutic discovery and development efforts
are under way to develop such drugs, named CFTR correctors.These efforts have been encouraged by the fact that F508del-CFTR protein retains some functionality as a Cl2 channel atthe plasma membrane of epithelial cells (Dalemans et al.,1991; Denning et al., 1992). However, several lines of evidencealso indicate that it might not be possible to reverse fully theF508del-CFTR trafficking defect by using only a single compound.Recent findings suggest that a combination of a corrector anda potentiator of CFTR would have better efficacy than a cor-rector alone; these findings are progressively advancing towarda polypharmacology to address the CFTR defects, in particular,defective trafficking and gating (Okiyoneda et al., 2013).It is important to note that all the small chemical compounds
identified so far individually increase the amount of functionalproteins at the plasma membrane but only partially repair thetrafficking defect of F508del-CFTR (Zeitlin, 2000; Becq, 2010).Some of themmay work as pharmacologic chaperones by directbinding to F508del-CFTR, which favors its folding and mat-uration. Alternatively, correctors may also act as proteostasisregulators by modifying the cell proteome to ameliorate thematuration of F508del-CFTR (Wang et al., 2000;Mu et al., 2008;Hutt et al., 2010). Because none of these correctors, used alone,can correct fully the F508del-CFTR abnormal trafficking, thepresent study was conducted to explore the hypothesis that acombination of different correctors could be more efficaciousthan a single corrector. Among these chemical compounds, wefocused our attention on correctors with a different mechanismof action: the proteostasis regulators suberoylamilide hydroxamicacid (SAHA) (Hutt et al., 2010), miglustat (Norez et al., 2006),and IsoLAB (1,4-dideoxy-2-hydroxymethyl-1,4-imino-L-threitol)(Ardes-Guisot et al., 2011; Best et al., 2011), in addition to VX-809[3-(6-(1-(2,2-difluorobenzo[d][1,3]dioxol-5-yl)cyclopropanecarbox-amido)-3-methylpyridin-2-yl)benzoic acid] (VanGoor et al., 2011;Loo et al., 2013) and Corr4a (N-[2-(5-chloro-2-methoxy-phenyl-amino)-49-methyl-[4,59]bithiazolyl-29-yl]-benzamide) (Pedemonteet al., 2005), proposed to interact directly with CFTR. We alsodiscuss the different classes of CFTR correctors based on theirmolecular mechanism of action.
Materials and MethodsCell Culture. HeLa cells expressing wild-type CFTR (spTCF-WT)
or F508del-CFTR (spTCF-DF) were cultured in Dulbecco’s modifiedEagle’smedium1GlutaMAX-I (Invitrogen, Carlsbad, CA) supplementedwith 8% (v/v) fetal bovine serum, 1% (v/v) penicillin/streptomycin, andwere selected using Zeocin (50 mg/ml) (Invitrogen/Life Technologies,Cergy Pontoise Cedex, France). Both cell lines were grown in standardculture conditions (37°C, 5% CO2). Cells were plated in 35-mm plasticdishes for whole-cell patch-clamp recordings and Western blot analysisand in 96-well plates for cytotoxicity assay. For all cell cultures,culture media were renewed every 2 days.
Western Blot Analysis. Cell lysates (10 mM Tris, 1% NonidetP-40, 0.5% sodium deoxycholate, pH 7.5) were separated 72 hoursafter seeding for HeLa cells by 5% SDS-PAGE (50 mg of protein/well).After saturation, nitrocellulose membranes were incubated overnightat 4°C in phosphate-buffered saline (PBS), 0.1% Tween 20 with 1 mg/mlmouse anti-CFTR monoclonal antibody (clone MAB3480; ChemiconInternational, Millipore Bioscience Research Reagents, Temecula, CA).After washing, goat peroxidase–conjugated anti-mouse IgG (1:10,000;Sigma-Aldrich) was used as secondary antibody. CFTR was visualizedby chemiluminescence with enhanced chemiluminescence Westernblotting detection reagent (GE Healthcare, Buckinghamshire, UK).
Cell Viability Assay. The cell viability of HeLa-F508del-CFTRcells exposed to different concentrations of compounds was evalu-ated using the mitochondrial-dependent reduction of MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; Sigma-Aldrich]colorimetric assay, as described elsewhere (Routaboul et al., 2007). Cellswere incubated at 37°C with the test compound at the concentration andtime of incubation previously defined. At the end of incubation, mediawere removed and cells were washed with PBS before a 4-hour-longincubation at 37°Cwith 100ml of MTT solution (0.5mg/ml in PBS). Afterincubation, the supernatantwas removed, the purple formazan crystalswere dissolved in 100 ml dimethylsulfoxide (Sigma-Aldrich), and theoptical density was measured in a microplate reader (SpectraCountmicroplate photometer; Packard, Meriden, CT). Values are the mean ofthree replicates and are expressed as percentage of the control values.
Electrophysiologic Studies Using theWhole-Cell Patch-ClampTechnique. Ionic currents were recorded using the patch-rupturedwhole-cell variant of the patch-clamp technique and measured using anAxopatch 200B amplifier (Molecular Devices, Union City, CA). Currentswere filtered at 5 kHz (23 dayB; four-pole Bessel filter). The holdingpotential was 240 mV in all whole-cell experiments. Current/voltage(I/V) relationships were built by clamping the membrane potential to240 mV and by pulses from 2100 to 1100 mV in 20 mV increments.Histograms of current densities in response to activation were obtainedby summarizing the maximal value of current density reached duringthe time course of activation. For time-course experiments, currentamplitude measured at 140 mV was plotted each 10 seconds. Pipetteswere prepared by pulling borosilicate glass capillary tubes (GC150-T10;Harvard Apparatus, Edenbridge, UK) using a two-step vertical puller(Narishige, Tokyo, Japan). They were filled with the following solution(in mM): 120 NMDG, 86 L-aspartic acid, 3 MgCl2, 1 CsEGTA, 5 TES,and 3 MgATP ex temporane, (titrated with NaOH to pH 7.2, theosmolarity was 285 6 5 mOsmol). The pipette solution was alwayshypotonic (with respect to the bath solution) to prevent cell swelling andactivation of the volume-sensitive chloride channels. The stilbenederivative DIDS (4,49-diisothiocyanatostilbene-2,29-disulfonic acid diso-dium salt hydrate) was also used to block non-CFTR Cl2 conductance.Pipettes were connected to the head of the patch-clamp amplifierthrough an Ag–AgCl pellet (pipette resistance of 10–20 MV). The ex-ternal bath solution contained (in mM): 140 NaCl, 1.2 CaCl2, 1 MgSO4,10 dextrose, and 10 TES (titrated with NaOH to pH 7.4; the osmolaritywas 315 6 5 mOsmol). Pipette capacitance was electronically compen-sated in cell-attached mode. Membrane capacitance and series resis-tances were measured in the whole-cell mode by fitting capacitancecurrents obtained in response to a hyperpolarization of 40 mV with afirst-order exponential and by integrating the surface of the capacitancecurrent. Voltage-clamp signalswere recorded via amicrocomputer equip-ped with an analog/digital-digital/analog conversion board (Digidata1440A interface; Molecular Devices). Experiments were performedeither at room temperature (RT; 20–25°C) or at 37°C. For experimentsat 37°C, from the seal formation to the end of experiment, the temper-ature in the cell bath was controlled through the external solution andmaintained at 37°C using a designed homemade temperature controllerconnected on the stage of our inverted microscope. A thermoprobepositioned in the patch chamber was used to monitor the temperature.Drugs were applied by a gravity-fed perfusion system. All Cl2 currentswere analyzed with the pCLAMP version 10.2-package software(Molecular Devices).
Combination of F508del-CFTR Correctors 625
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from
Pharmacology, Chemicals, and Reagents. The CFTR activatorforskolin (noted F) and potentiator genistein (noted G) (Illek et al.,1995) were purchased from LC Laboratories (PKC Pharmaceuticals,Woburn, MA). The selective CFTR inhibitor CFTRinh-172 (Ma et al.,2002) was from Calbiochem (Nottingham, UK). We purchased DIDSand SAHA from Sigma-Aldrich (St. Louis, MO), miglustat [(2R,3R,4R,5S)-1-butyl-2-(hydroxymethyl)piperidine-3,4,5-triol] from TorontoResearch Chemicals (Toronto, ON, Canada), and VX-809 and VX-770from Selleckchem (Houston, TX). Corr4a was provided by Dr. RobertBridges from the Rosalind Franklin University (North Chicago, IL) viathe Chemical Compound Distribution Program of the Cystic FibrosisFoundation. Stock solutions of F (10 mM), G (30 mM), CFTRinh-172(10 mM), VX-809 (10 mM), VX-770 (10 mM), SAHA (10 and 1 mM), andCorr4a (10 mM) were prepared in dimethylsulfoxide. We preparedstock solution of iminosugars, miglustat, and IsoLAB dissolved inwater(100 mM) before further dilution.
Statistical Analysis. Results are expressed as means 6 S.E. ofn observations. Statistical analysis was carried out using GraphPadPrism (GraphPad Software, La Jolla, CA) version 5.0 for Windows. Tocompare sets of data, we used one-way analysis of variance followedby Dunnett’s multiple-comparison test or Student’s t-test. Differenceswere considered statistically significant when P , 0.05.
ResultsIn this section, until experiments investigating the effect of
temperature and thermal instability on CFTR function, allpatch-clamp experiments were performed at RT.
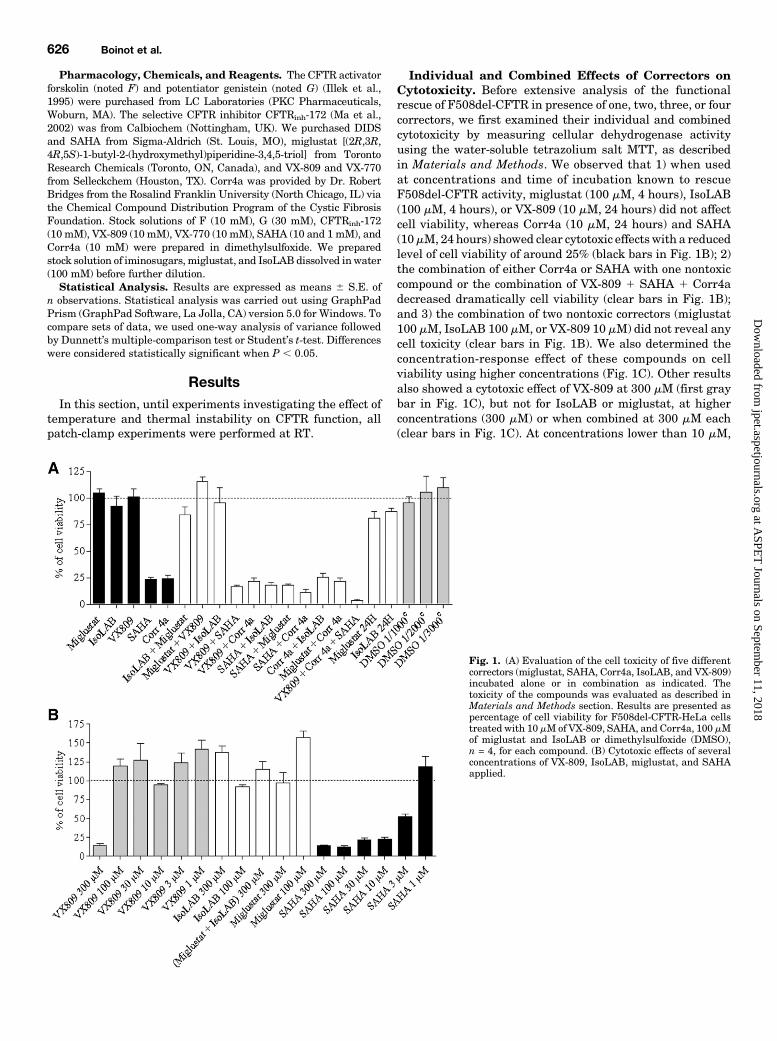
Individual and Combined Effects of Correctors onCytotoxicity. Before extensive analysis of the functionalrescue of F508del-CFTR in presence of one, two, three, or fourcorrectors, we first examined their individual and combinedcytotoxicity by measuring cellular dehydrogenase activityusing the water-soluble tetrazolium salt MTT, as describedin Materials and Methods. We observed that 1) when usedat concentrations and time of incubation known to rescueF508del-CFTR activity, miglustat (100 mM, 4 hours), IsoLAB(100 mM, 4 hours), or VX-809 (10 mM, 24 hours) did not affectcell viability, whereas Corr4a (10 mM, 24 hours) and SAHA(10mM, 24 hours) showed clear cytotoxic effects with a reducedlevel of cell viability of around 25% (black bars in Fig. 1B); 2)the combination of either Corr4a or SAHA with one nontoxiccompound or the combination of VX-809 1 SAHA 1 Corr4adecreased dramatically cell viability (clear bars in Fig. 1B);and 3) the combination of two nontoxic correctors (miglustat100 mM, IsoLAB 100 mM, or VX-809 10 mM) did not reveal anycell toxicity (clear bars in Fig. 1B). We also determined theconcentration-response effect of these compounds on cellviability using higher concentrations (Fig. 1C). Other resultsalso showed a cytotoxic effect of VX-809 at 300 mM (first graybar in Fig. 1C), but not for IsoLAB or miglustat, at higherconcentrations (300 mM) or when combined at 300 mM each(clear bars in Fig. 1C). At concentrations lower than 10 mM,
Fig. 1. (A) Evaluation of the cell toxicity of five differentcorrectors (miglustat, SAHA, Corr4a, IsoLAB, and VX-809)incubated alone or in combination as indicated. Thetoxicity of the compounds was evaluated as described inMaterials and Methods section. Results are presented aspercentage of cell viability for F508del-CFTR-HeLa cellstreated with 10 mM of VX-809, SAHA, and Corr4a, 100 mMof miglustat and IsoLAB or dimethylsulfoxide (DMSO),n = 4, for each compound. (B) Cytotoxic effects of severalconcentrations of VX-809, IsoLAB, miglustat, and SAHAapplied.
626 Boinot et al.
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from
SAHA appears less cytotoxic (see black bars in Fig. 1C), anobservation that leads us to use SAHA at 1 mM. Based onthese results, we focused our study on VX-809, SAHA (1 mM),miglustat, and IsoLAB.Properties of F508del-CFTR Cl2 Currents. We ex-
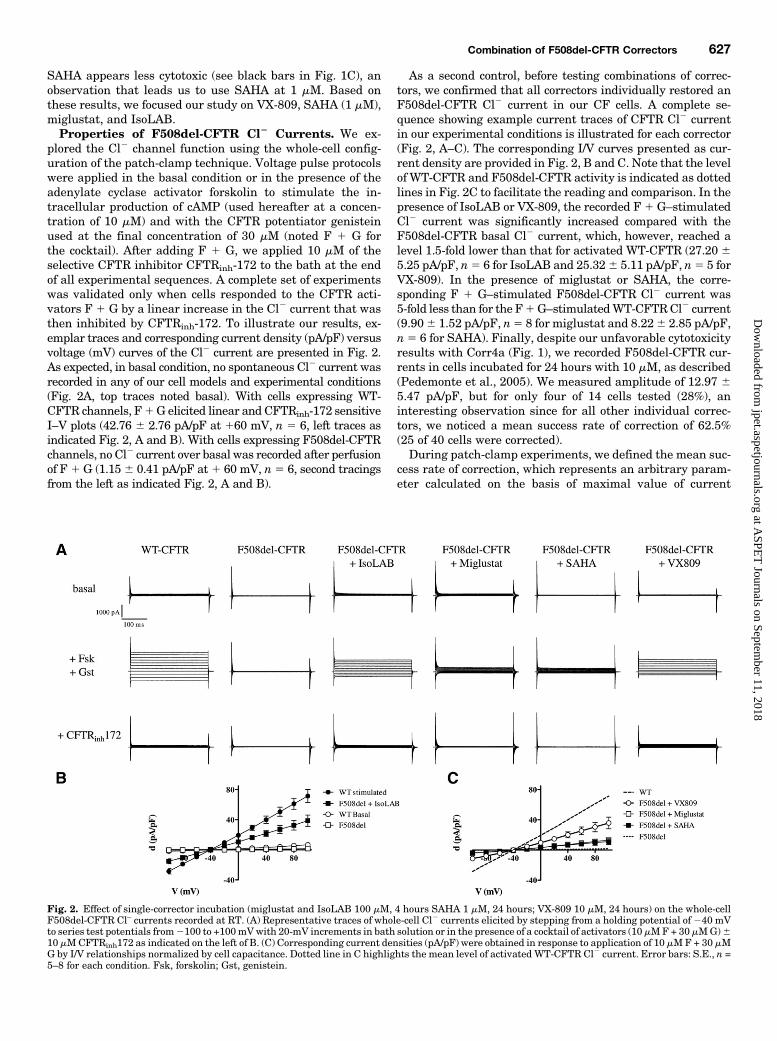
plored the Cl2 channel function using the whole-cell config-uration of the patch-clamp technique. Voltage pulse protocolswere applied in the basal condition or in the presence of theadenylate cyclase activator forskolin to stimulate the in-tracellular production of cAMP (used hereafter at a concen-tration of 10 mM) and with the CFTR potentiator genisteinused at the final concentration of 30 mM (noted F 1 G forthe cocktail). After adding F 1 G, we applied 10 mM of theselective CFTR inhibitor CFTRinh-172 to the bath at the endof all experimental sequences. A complete set of experimentswas validated only when cells responded to the CFTR acti-vators F 1 G by a linear increase in the Cl2 current that wasthen inhibited by CFTRinh-172. To illustrate our results, ex-emplar traces and corresponding current density (pA/pF) versusvoltage (mV) curves of the Cl2 current are presented in Fig. 2.As expected, in basal condition, no spontaneous Cl2 current wasrecorded in any of our cell models and experimental conditions(Fig. 2A, top traces noted basal). With cells expressing WT-CFTR channels, F1G elicited linear and CFTRinh-172 sensitiveI–V plots (42.76 6 2.76 pA/pF at 160 mV, n 5 6, left traces asindicated Fig. 2, A and B). With cells expressing F508del-CFTRchannels, no Cl2 current over basal was recorded after perfusionof F 1G (1.156 0.41 pA/pF at 1 60 mV, n5 6, second tracingsfrom the left as indicated Fig. 2, A and B).
As a second control, before testing combinations of correc-tors, we confirmed that all correctors individually restored anF508del-CFTR Cl2 current in our CF cells. A complete se-quence showing example current traces of CFTR Cl2 currentin our experimental conditions is illustrated for each corrector(Fig. 2, A–C). The corresponding I/V curves presented as cur-rent density are provided in Fig. 2, B and C. Note that the levelof WT-CFTR and F508del-CFTR activity is indicated as dottedlines in Fig. 2C to facilitate the reading and comparison. In thepresence of IsoLAB or VX-809, the recorded F1 G–stimulatedCl2 current was significantly increased compared with theF508del-CFTR basal Cl2 current, which, however, reached alevel 1.5-fold lower than that for activated WT-CFTR (27.20 65.25 pA/pF, n5 6 for IsoLAB and 25.326 5.11 pA/pF, n5 5 forVX-809). In the presence of miglustat or SAHA, the corre-sponding F 1 G–stimulated F508del-CFTR Cl2 current was5-fold less than for the F1G–stimulatedWT-CFTRCl2 current(9.906 1.52 pA/pF, n5 8 for miglustat and 8.226 2.85 pA/pF,n5 6 for SAHA). Finally, despite our unfavorable cytotoxicityresults with Corr4a (Fig. 1), we recorded F508del-CFTR cur-rents in cells incubated for 24 hours with 10 mM, as described(Pedemonte et al., 2005). We measured amplitude of 12.97 65.47 pA/pF, but for only four of 14 cells tested (28%), aninteresting observation since for all other individual correc-tors, we noticed a mean success rate of correction of 62.5%(25 of 40 cells were corrected).During patch-clamp experiments, we defined the mean suc-
cess rate of correction, which represents an arbitrary param-eter calculated on the basis of maximal value of current
Fig. 2. Effect of single-corrector incubation (miglustat and IsoLAB 100 mM, 4 hours SAHA 1 mM, 24 hours; VX-809 10 mM, 24 hours) on the whole-cellF508del-CFTR Cl– currents recorded at RT. (A) Representative traces of whole-cell Cl2 currents elicited by stepping from a holding potential of240 mVto series test potentials from2100 to +100mVwith 20-mV increments in bath solution or in the presence of a cocktail of activators (10 mMF+ 30 mMG)610 mMCFTRinh172 as indicated on the left of B. (C) Corresponding current densities (pA/pF) were obtained in response to application of 10 mMF + 30 mMG by I/V relationships normalized by cell capacitance. Dotted line in C highlights the mean level of activated WT-CFTR Cl2 current. Error bars: S.E., n =5–8 for each condition. Fsk, forskolin; Gst, genistein.
Combination of F508del-CFTR Correctors 627
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from
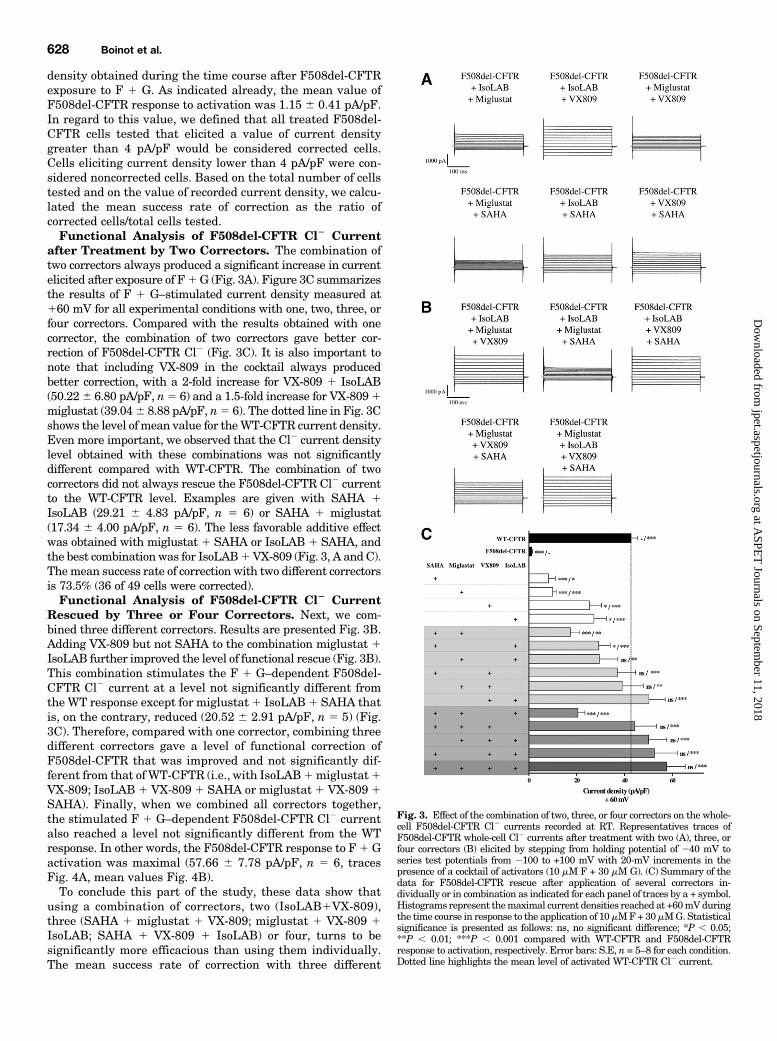
density obtained during the time course after F508del-CFTRexposure to F 1 G. As indicated already, the mean value ofF508del-CFTR response to activation was 1.15 6 0.41 pA/pF.In regard to this value, we defined that all treated F508del-CFTR cells tested that elicited a value of current densitygreater than 4 pA/pF would be considered corrected cells.Cells eliciting current density lower than 4 pA/pF were con-sidered noncorrected cells. Based on the total number of cellstested and on the value of recorded current density, we calcu-lated the mean success rate of correction as the ratio ofcorrected cells/total cells tested.Functional Analysis of F508del-CFTR Cl2 Current
after Treatment by Two Correctors. The combination oftwo correctors always produced a significant increase in currentelicited after exposure of F1G (Fig. 3A). Figure 3C summarizesthe results of F 1 G–stimulated current density measured at160 mV for all experimental conditions with one, two, three, orfour correctors. Compared with the results obtained with onecorrector, the combination of two correctors gave better cor-rection of F508del-CFTR Cl2 (Fig. 3C). It is also important tonote that including VX-809 in the cocktail always producedbetter correction, with a 2-fold increase for VX-809 1 IsoLAB(50.226 6.80 pA/pF, n5 6) and a 1.5-fold increase for VX-8091miglustat (39.046 8.88 pA/pF, n5 6). The dotted line in Fig. 3Cshows the level of mean value for theWT-CFTR current density.Even more important, we observed that the Cl2 current densitylevel obtained with these combinations was not significantlydifferent compared with WT-CFTR. The combination of twocorrectors did not always rescue the F508del-CFTR Cl2 currentto the WT-CFTR level. Examples are given with SAHA 1IsoLAB (29.21 6 4.83 pA/pF, n 5 6) or SAHA 1 miglustat(17.34 6 4.00 pA/pF, n 5 6). The less favorable additive effectwas obtained with miglustat 1 SAHA or IsoLAB 1 SAHA, andthe best combinationwas for IsoLAB1VX-809 (Fig. 3, A andC).Themean success rate of correction with two different correctorsis 73.5% (36 of 49 cells were corrected).Functional Analysis of F508del-CFTR Cl2 Current
Rescued by Three or Four Correctors. Next, we com-bined three different correctors. Results are presented Fig. 3B.Adding VX-809 but not SAHA to the combination miglustat 1IsoLAB further improved the level of functional rescue (Fig. 3B).This combination stimulates the F 1 G–dependent F508del-CFTR Cl2 current at a level not significantly different fromtheWT response except for miglustat1 IsoLAB1 SAHA thatis, on the contrary, reduced (20.52 6 2.91 pA/pF, n 5 5) (Fig.3C). Therefore, compared with one corrector, combining threedifferent correctors gave a level of functional correction ofF508del-CFTR that was improved and not significantly dif-ferent from that of WT-CFTR (i.e., with IsoLAB1miglustat1VX-809; IsoLAB 1 VX-8091 SAHA or miglustat 1 VX-8091SAHA). Finally, when we combined all correctors together,the stimulated F 1 G–dependent F508del-CFTR Cl2 currentalso reached a level not significantly different from the WTresponse. In other words, the F508del-CFTR response to F1 Gactivation was maximal (57.66 6 7.78 pA/pF, n 5 6, tracesFig. 4A, mean values Fig. 4B).To conclude this part of the study, these data show that
using a combination of correctors, two (IsoLAB1VX-809),three (SAHA 1 miglustat 1 VX-809; miglustat 1 VX-809 1IsoLAB; SAHA 1 VX-809 1 IsoLAB) or four, turns to besignificantly more efficacious than using them individually.The mean success rate of correction with three different
Fig. 3. Effect of the combination of two, three, or four correctors on the whole-cell F508del-CFTR Cl2 currents recorded at RT. Representatives traces ofF508del-CFTR whole-cell Cl2 currents after treatment with two (A), three, orfour correctors (B) elicited by stepping from holding potential of 240 mV toseries test potentials from 2100 to +100 mV with 20-mV increments in thepresence of a cocktail of activators (10 mM F + 30 mM G). (C) Summary of thedata for F508del-CFTR rescue after application of several correctors in-dividually or in combination as indicated for each panel of traces by a + symbol.Histograms represent themaximal current densities reached at +60mVduringthe time course in response to the application of 10mMF+30mMG. Statisticalsignificance is presented as follows: ns, no significant difference; *P , 0.05;**P , 0.01; ***P , 0.001 compared with WT-CFTR and F508del-CFTRresponse to activation, respectively. Error bars: S.E, n = 5–8 for each condition.Dotted line highlights the mean level of activated WT-CFTR Cl2 current.
628 Boinot et al.
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from
correctors is 82% (23 of 28 cells were corrected) and 75% (six ofeight cells were corrected) with the cocktail of four correctors.Effect of Varying the Duration of Incubation of
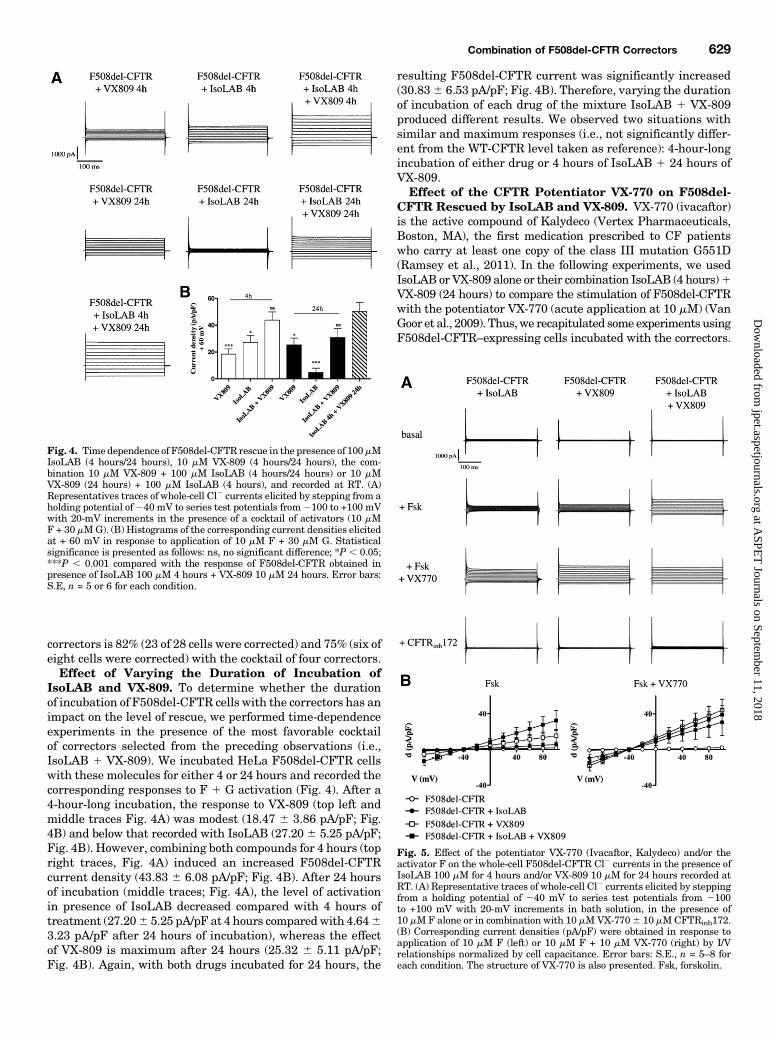
IsoLAB and VX-809. To determine whether the durationof incubation of F508del-CFTR cells with the correctors has animpact on the level of rescue, we performed time-dependenceexperiments in the presence of the most favorable cocktailof correctors selected from the preceding observations (i.e.,IsoLAB 1 VX-809). We incubated HeLa F508del-CFTR cellswith these molecules for either 4 or 24 hours and recorded thecorresponding responses to F 1 G activation (Fig. 4). After a4-hour-long incubation, the response to VX-809 (top left andmiddle traces Fig. 4A) was modest (18.47 6 3.86 pA/pF; Fig.4B) and below that recorded with IsoLAB (27.20 6 5.25 pA/pF;Fig. 4B). However, combining both compounds for 4 hours (topright traces, Fig. 4A) induced an increased F508del-CFTRcurrent density (43.83 6 6.08 pA/pF; Fig. 4B). After 24 hoursof incubation (middle traces; Fig. 4A), the level of activationin presence of IsoLAB decreased compared with 4 hours oftreatment (27.206 5.25 pA/pF at 4 hours comparedwith 4.6463.23 pA/pF after 24 hours of incubation), whereas the effectof VX-809 is maximum after 24 hours (25.32 6 5.11 pA/pF;Fig. 4B). Again, with both drugs incubated for 24 hours, the
resulting F508del-CFTR current was significantly increased(30.836 6.53 pA/pF; Fig. 4B). Therefore, varying the durationof incubation of each drug of the mixture IsoLAB 1 VX-809produced different results. We observed two situations withsimilar and maximum responses (i.e., not significantly differ-ent from the WT-CFTR level taken as reference): 4-hour-longincubation of either drug or 4 hours of IsoLAB 1 24 hours ofVX-809.Effect of the CFTR Potentiator VX-770 on F508del-
CFTR Rescued by IsoLAB and VX-809. VX-770 (ivacaftor)is the active compound of Kalydeco (Vertex Pharmaceuticals,Boston, MA), the first medication prescribed to CF patientswho carry at least one copy of the class III mutation G551D(Ramsey et al., 2011). In the following experiments, we usedIsoLAB or VX-809 alone or their combination IsoLAB (4 hours)1VX-809 (24 hours) to compare the stimulation of F508del-CFTRwith the potentiator VX-770 (acute application at 10 mM) (VanGoor et al., 2009). Thus,we recapitulated some experiments usingF508del-CFTR–expressing cells incubated with the correctors.
Fig. 4. Time dependence of F508del-CFTR rescue in the presence of 100mMIsoLAB (4 hours/24 hours), 10 mM VX-809 (4 hours/24 hours), the com-bination 10 mM VX-809 + 100 mM IsoLAB (4 hours/24 hours) or 10 mMVX-809 (24 hours) + 100 mM IsoLAB (4 hours), and recorded at RT. (A)Representatives traces of whole-cell Cl2 currents elicited by stepping from aholding potential of240 mV to series test potentials from2100 to +100 mVwith 20-mV increments in the presence of a cocktail of activators (10 mMF + 30 mMG). (B) Histograms of the corresponding current densities elicitedat + 60 mV in response to application of 10 mM F + 30 mM G. Statisticalsignificance is presented as follows: ns, no significant difference; *P , 0.05;***P , 0.001 compared with the response of F508del-CFTR obtained inpresence of IsoLAB 100 mM 4 hours + VX-809 10 mM 24 hours. Error bars:S.E, n = 5 or 6 for each condition.
Fig. 5. Effect of the potentiator VX-770 (Ivacaftor, Kalydeco) and/or theactivator F on the whole-cell F508del-CFTR Cl2 currents in the presence ofIsoLAB 100 mM for 4 hours and/or VX-809 10 mM for 24 hours recorded atRT. (A) Representative traces of whole-cell Cl2 currents elicited by steppingfrom a holding potential of 240 mV to series test potentials from 2100to +100 mV with 20-mV increments in bath solution, in the presence of10 mMF alone or in combination with 10 mMVX-7706 10 mMCFTRinh172.(B) Corresponding current densities (pA/pF) were obtained in response toapplication of 10 mM F (left) or 10 mM F + 10 mM VX-770 (right) by I/Vrelationships normalized by cell capacitance. Error bars: S.E., n = 5–8 foreach condition. The structure of VX-770 is also presented. Fsk, forskolin.
Combination of F508del-CFTR Correctors 629
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from

We stimulated the CFTR Cl2 current by forskolin 6 VX-770.Results are presented inFig. 5with, for each condition, exampleCl2 current traces in A and current density versus voltage in B.The level of F508del-CFTRCl2 current after IsoLAB or VX-809treatment was significantly stimulated by F 1 VX-770, abovethe level recorded with forskolin alone (Fig. 5A). For cells in-cubated with the cocktail IsoLAB1VX-809, the best functionalresponse was also recorded with F 1 VX-770. Of note, the re-sponse to forskolin alone is already significantly above the levelreached with forskolin and either or the two correctors usedindividually (Fig. 5A).Effect of Temperature and Thermal Instability of
F508del-CFTR Chloride Currents in the Presence ofCorrectors. In the following experiments, we investigatedthe effect of temperature on the activity of F508del-CFTR.Thermal inactivation or instability of the mutant F508del at37°C was recently shown in mammalian cells (Wang et al.,2011; He et al., 2013) and Xenopus oocytes (Liu et al., 2012).We measured the stability of WT-CFTR and corrected-F508delchloride currents using whole-cell, patch-clamp technique andcompared the effect of various correctors at 37°C. The currentdensity of CFTR currents was quantified with cells continu-ously maintained at 37°C during the complete recording ofthe whole-cell CFTR chloride currents at 140 mV (from sealformation to the end of experiment). After seal formation andcell opening, cells were continuously exposed to F 1 G for20 minutes of time recording. For each condition, the time oflatency observed before increase of current density (1 or 2minutes) represents the lapse of time for F 1 G distributionin the whole-patch chamber. WithWT-CFTR, the current den-sity remained stable for several minutes (about 14–16 minutes)at the maximum level after adding forskolin and genistein
and before decreasing slowly (rundown) without the applica-tion of any inhibitor (Fig. 6A). On the contrary, with F508del-CFTR cells corrected by 24-hour long-term incubation at 27°C(noted low temperature–corrected cells), the current densitydeclined rapidly within 6–8 minutes after maximal stimulationby forskolin and genistein (Fig. 6B). In the presence of eitherVX-809 (filled squares) or IsoLAB (black triangles), the stim-ulated F508del-CFTR current declined even more rapidly thanfor temperature-corrected cells (i.e., within 2 or 3 minutes (Fig.6C). Lastly, when F508del-CFTR cells were incubated withVX-809 1 IsoLAB, the stimulated F508del-CFTR current wasincreased (empty squares, Fig. 6C) compared with VX-809 orIsoLAB separately. However, the current density still declinedmore rapidly than for temperature-corrected cells, althoughslower than with each individual correctors (i.e., in 4–6minutes,the current density vanished) (Fig. 6C).In a second set of experiments, we compared the level of cur-
rent density corresponding to the maximal level of activationreached during time course of activation of WT-CFTR andF508del-CFTR treated with VX-809, IsoLAB, or VX-809 1IsoLAB recorded at RT or at 37°C (Fig. 6D). The activity ofWT-CFTR was significantly increased at 37°C versus RT (Fig.6D). The activity of F508del-CFTR was also increased at 37°Cversus RT but only for VX-809 1 IsoLAB (Fig. 6D). Surpris-ingly, the levels of current densities of F508del-CFTR correctedeither with VX-809 or IsoLAB were similar at 37°C and RT(Fig. 6D).Increased Mature c-Band F508del-CFTR in Cells
Treated by Multiple Correctors. Finally, the results ofthese several experiments told indicated that a combination ofat least two different chemicals could improve the functionalcorrection of F508del-CFTR Cl2 current. Our next question
Fig. 6. Effect of temperature on the activity of WT-CFTR and corrected F508del-CFTR. (A) Time coursecorresponding to thermal inactivation of WT-CFTR(d, n = 4). (B) Temperature-corrected F508del-CFTRafter 24 hours of incubation at 27°C (s, n = 5). (C)F508del-CFTR in the presence of VX-809 (j, n = 5),IsoLAB (m, n = 5), or VX-809 + IsoLAB (u, n = 6) afterstimulation with 10 mM F + 30 mM G and elicited at+40mV and continuously warmed to 37°C during thewhole experiment (horizontal pointed gray bar). (D)Summary of current densities corresponding to themaximal activation reached during the time course ofthe whole-cell current of WT-CFTR and F508del-CFTR treated with VX-809, IsoLAB, or VX-809 +IsoLAB elicited at + 40 mV at room temperature(20–25°C, noted RT, white bar) or at 37°C (black bar)after stimulation with 10mMF+ 30mMG. Statisticalsignificance is presented as follows: ns, no significantdifference; *P , 0.05; ***P , 0.001 compared withresponse of WT-CFTR or corrected F508del-CFTRobtained at RT. Error bars, S.E, n = 4–7 for eachcondition.
630 Boinot et al.
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from
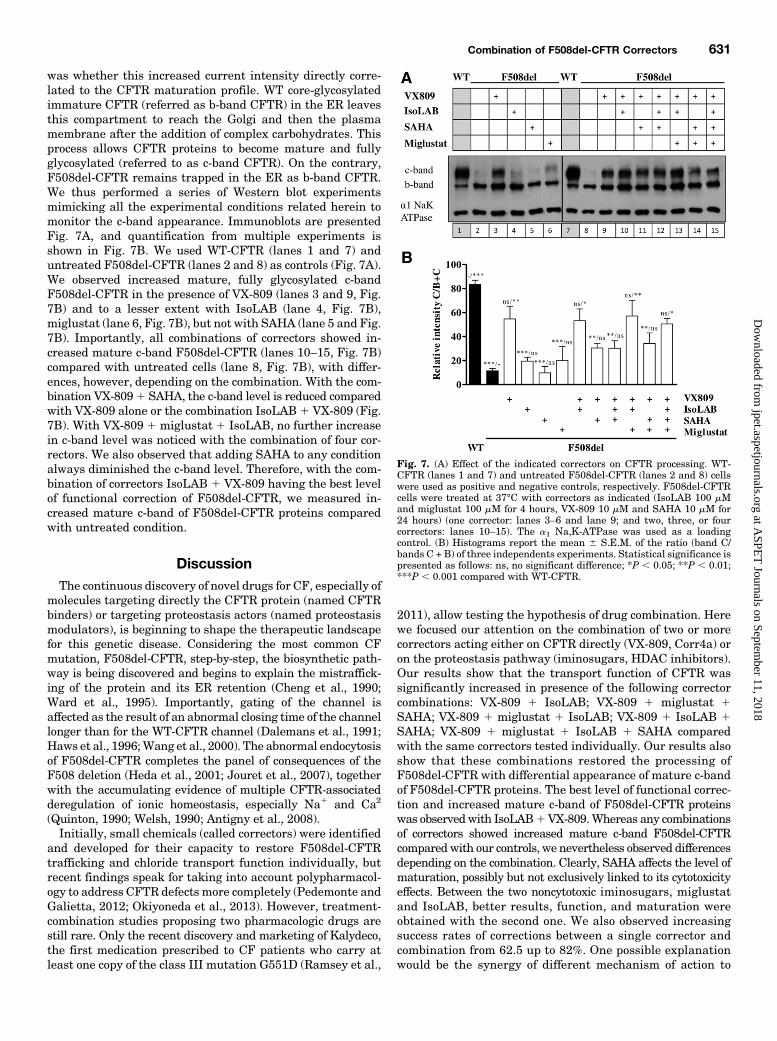
was whether this increased current intensity directly corre-lated to the CFTR maturation profile. WT core-glycosylatedimmature CFTR (referred as b-band CFTR) in the ER leavesthis compartment to reach the Golgi and then the plasmamembrane after the addition of complex carbohydrates. Thisprocess allows CFTR proteins to become mature and fullyglycosylated (referred to as c-band CFTR). On the contrary,F508del-CFTR remains trapped in the ER as b-band CFTR.We thus performed a series of Western blot experimentsmimicking all the experimental conditions related herein tomonitor the c-band appearance. Immunoblots are presentedFig. 7A, and quantification from multiple experiments isshown in Fig. 7B. We used WT-CFTR (lanes 1 and 7) anduntreated F508del-CFTR (lanes 2 and 8) as controls (Fig. 7A).We observed increased mature, fully glycosylated c-bandF508del-CFTR in the presence of VX-809 (lanes 3 and 9, Fig.7B) and to a lesser extent with IsoLAB (lane 4, Fig. 7B),miglustat (lane 6, Fig. 7B), but not with SAHA (lane 5 and Fig.7B). Importantly, all combinations of correctors showed in-creased mature c-band F508del-CFTR (lanes 10–15, Fig. 7B)compared with untreated cells (lane 8, Fig. 7B), with differ-ences, however, depending on the combination. With the com-bination VX-8091 SAHA, the c-band level is reduced comparedwith VX-809 alone or the combination IsoLAB1 VX-809 (Fig.7B). With VX-809 1 miglustat 1 IsoLAB, no further increasein c-band level was noticed with the combination of four cor-rectors. We also observed that adding SAHA to any conditionalways diminished the c-band level. Therefore, with the com-bination of correctors IsoLAB 1 VX-809 having the best levelof functional correction of F508del-CFTR, we measured in-creased mature c-band of F508del-CFTR proteins comparedwith untreated condition.
DiscussionThe continuous discovery of novel drugs for CF, especially of
molecules targeting directly the CFTR protein (named CFTRbinders) or targeting proteostasis actors (named proteostasismodulators), is beginning to shape the therapeutic landscapefor this genetic disease. Considering the most common CFmutation, F508del-CFTR, step-by-step, the biosynthetic path-way is being discovered and begins to explain the mistraffick-ing of the protein and its ER retention (Cheng et al., 1990;Ward et al., 1995). Importantly, gating of the channel isaffected as the result of an abnormal closing time of the channellonger than for the WT-CFTR channel (Dalemans et al., 1991;Haws et al., 1996;Wang et al., 2000). The abnormal endocytosisof F508del-CFTR completes the panel of consequences of theF508 deletion (Heda et al., 2001; Jouret et al., 2007), togetherwith the accumulating evidence of multiple CFTR-associatedderegulation of ionic homeostasis, especially Na1 and Ca2
(Quinton, 1990; Welsh, 1990; Antigny et al., 2008).Initially, small chemicals (called correctors) were identified
and developed for their capacity to restore F508del-CFTRtrafficking and chloride transport function individually, butrecent findings speak for taking into account polypharmacol-ogy to address CFTR defects more completely (Pedemonte andGalietta, 2012; Okiyoneda et al., 2013). However, treatment-combination studies proposing two pharmacologic drugs arestill rare. Only the recent discovery and marketing of Kalydeco,the first medication prescribed to CF patients who carry atleast one copy of the class III mutation G551D (Ramsey et al.,
2011), allow testing the hypothesis of drug combination. Herewe focused our attention on the combination of two or morecorrectors acting either on CFTR directly (VX-809, Corr4a) oron the proteostasis pathway (iminosugars, HDAC inhibitors).Our results show that the transport function of CFTR wassignificantly increased in presence of the following correctorcombinations: VX-809 1 IsoLAB; VX-809 1 miglustat 1SAHA; VX-809 1 miglustat 1 IsoLAB; VX-809 1 IsoLAB 1SAHA; VX-809 1 miglustat 1 IsoLAB 1 SAHA comparedwith the same correctors tested individually. Our results alsoshow that these combinations restored the processing ofF508del-CFTR with differential appearance of mature c-bandof F508del-CFTR proteins. The best level of functional correc-tion and increased mature c-band of F508del-CFTR proteinswas observedwith IsoLAB1VX-809.Whereas any combinationsof correctors showed increased mature c-band F508del-CFTRcomparedwith our controls, we nevertheless observed differencesdepending on the combination. Clearly, SAHA affects the level ofmaturation, possibly but not exclusively linked to its cytotoxicityeffects. Between the two noncytotoxic iminosugars, miglustatand IsoLAB, better results, function, and maturation wereobtained with the second one. We also observed increasingsuccess rates of corrections between a single corrector andcombination from 62.5 up to 82%. One possible explanationwould be the synergy of different mechanism of action to
Fig. 7. (A) Effect of the indicated correctors on CFTR processing. WT-CFTR (lanes 1 and 7) and untreated F508del-CFTR (lanes 2 and 8) cellswere used as positive and negative controls, respectively. F508del-CFTRcells were treated at 37°C with correctors as indicated (IsoLAB 100 mMand miglustat 100 mM for 4 hours, VX-809 10 mM and SAHA 10 mM for24 hours) (one corrector: lanes 3–6 and lane 9; and two, three, or fourcorrectors: lanes 10–15). The a1 Na,K-ATPase was used as a loadingcontrol. (B) Histograms report the mean 6 S.E.M. of the ratio (band C/bands C + B) of three independents experiments. Statistical significance ispresented as follows: ns, no significant difference; *P , 0.05; **P , 0.01;***P , 0.001 compared with WT-CFTR.
Combination of F508del-CFTR Correctors 631
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from
rescue more functional F508del-CFTR proteins to the plasmamembrane.CF pharmacotherapy was officially born a year ago with the
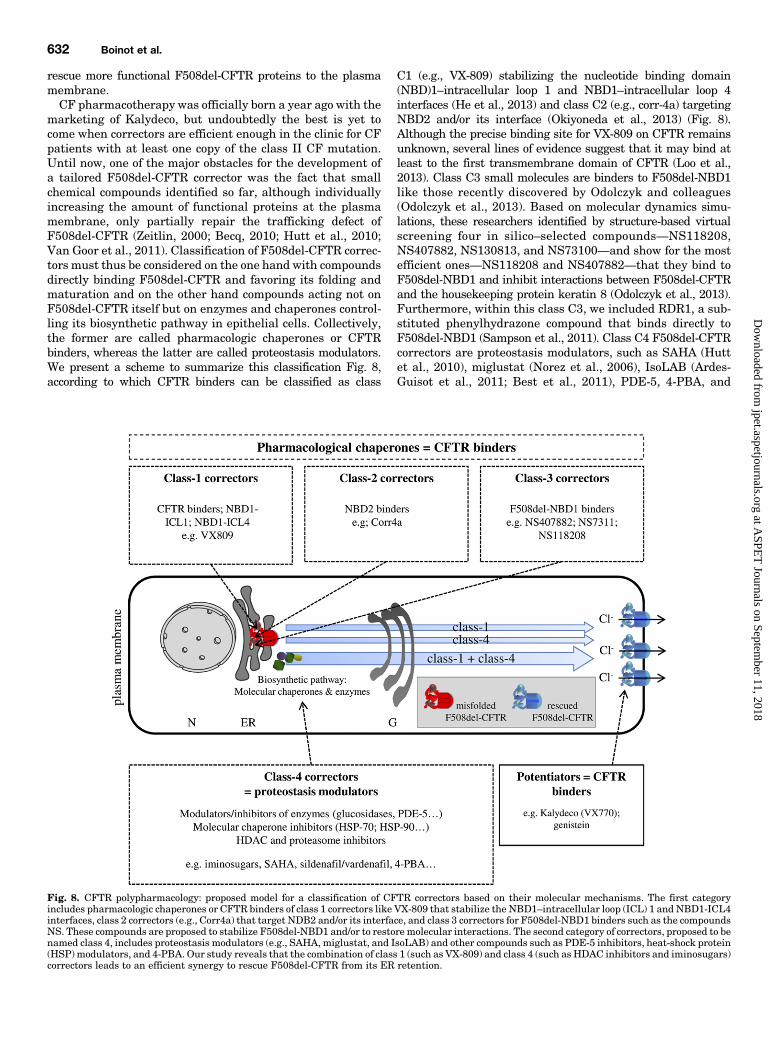
marketing of Kalydeco, but undoubtedly the best is yet tocome when correctors are efficient enough in the clinic for CFpatients with at least one copy of the class II CF mutation.Until now, one of the major obstacles for the development ofa tailored F508del-CFTR corrector was the fact that smallchemical compounds identified so far, although individuallyincreasing the amount of functional proteins at the plasmamembrane, only partially repair the trafficking defect ofF508del-CFTR (Zeitlin, 2000; Becq, 2010; Hutt et al., 2010;Van Goor et al., 2011). Classification of F508del-CFTR correc-tors must thus be considered on the one hand with compoundsdirectly binding F508del-CFTR and favoring its folding andmaturation and on the other hand compounds acting not onF508del-CFTR itself but on enzymes and chaperones control-ling its biosynthetic pathway in epithelial cells. Collectively,the former are called pharmacologic chaperones or CFTRbinders, whereas the latter are called proteostasis modulators.We present a scheme to summarize this classification Fig. 8,according to which CFTR binders can be classified as class
C1 (e.g., VX-809) stabilizing the nucleotide binding domain(NBD)1–intracellular loop 1 and NBD1–intracellular loop 4interfaces (He et al., 2013) and class C2 (e.g., corr-4a) targetingNBD2 and/or its interface (Okiyoneda et al., 2013) (Fig. 8).Although the precise binding site for VX-809 on CFTR remainsunknown, several lines of evidence suggest that it may bind atleast to the first transmembrane domain of CFTR (Loo et al.,2013). Class C3 small molecules are binders to F508del-NBD1like those recently discovered by Odolczyk and colleagues(Odolczyk et al., 2013). Based on molecular dynamics simu-lations, these researchers identified by structure-based virtualscreening four in silico–selected compounds—NS118208,NS407882, NS130813, and NS73100—and show for the mostefficient ones—NS118208 and NS407882—that they bind toF508del-NBD1 and inhibit interactions between F508del-CFTRand the housekeeping protein keratin 8 (Odolczyk et al., 2013).Furthermore, within this class C3, we included RDR1, a sub-stituted phenylhydrazone compound that binds directly toF508del-NBD1 (Sampson et al., 2011). Class C4 F508del-CFTRcorrectors are proteostasis modulators, such as SAHA (Huttet al., 2010), miglustat (Norez et al., 2006), IsoLAB (Ardes-Guisot et al., 2011; Best et al., 2011), PDE-5, 4-PBA, and
Fig. 8. CFTR polypharmacology: proposed model for a classification of CFTR correctors based on their molecular mechanisms. The first categoryincludes pharmacologic chaperones or CFTR binders of class 1 correctors like VX-809 that stabilize the NBD1–intracellular loop (ICL) 1 and NBD1-ICL4interfaces, class 2 correctors (e.g., Corr4a) that target NDB2 and/or its interface, and class 3 correctors for F508del-NBD1 binders such as the compoundsNS. These compounds are proposed to stabilize F508del-NBD1 and/or to restore molecular interactions. The second category of correctors, proposed to benamed class 4, includes proteostasis modulators (e.g., SAHA, miglustat, and IsoLAB) and other compounds such as PDE-5 inhibitors, heat-shock protein(HSP) modulators, and 4-PBA. Our study reveals that the combination of class 1 (such as VX-809) and class 4 (such as HDAC inhibitors and iminosugars)correctors leads to an efficient synergy to rescue F508del-CFTR from its ER retention.
632 Boinot et al.
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from

others (reviewed in Becq, 2010) (Fig. 8). Our study indicatesthat an efficient combination to rescue F508del-CFTR from itsER retention would be to combine class C1 (such as VX-809)and class C4 (such as iminosugars and histone deacetylase[HDAC] inhibitors) correctors (Fig. 8).A second result of our study concerns the thermal in-
stability of the mutant F508del at 37°C that was recentlydemonstrated (Wang et al., 2011; Liu et al., 2012; He et al.,2013). Wang and collaborators first showed, for example, thatlow temperature–rescued F508del-CFTR irreversibly inacti-vated in experiments in which the temperature was raisedfrom room temperature (typically 22°C) to physiologic value(36.5–37°C) with a time constant of 5 or 6 minutes (Wanget al., 2011). We obtained quite similar results, as shown Fig. 6,using comparable protocols. With the corrector VX-809, wealso found that the corrected F508del-CFTR current re-mained unstable as also shown previously (He et al., 2013).Similarly, with the iminosugar IsoLAB, we failed to stabilizethe F508del-CFTR current. However, a slower process wasobserved with the cocktail VX-809 1 IsoLAB. Also, we com-pared the level of F508del-CFTR channel activity and foundthat with the cocktail of correctors, the level of current wasstrongly augmented at 37°C compared with at RT. Our re-sults, however, also showed that despite the amelioration ofthe amplitude of corrected F508del-CFTR current at 37°C, themutant remained thermally instable. This observation wasprobably expected since we used class C4 correctors (i.e.,nonbinders to F508del). It might be predicted that stabilizingthe conformation of F508del protein with corrector binders ofclasses C1–C3 could also stabilize the F508del-CFTR currenttoward near normal value. It remains, however, to be demon-strated that a C1–C3 binders would indeed reverse the ther-mal instability of F508del-CFTR.In conclusion, given the wide possible variety of different
drug combinations and the superiority of some drug combina-tions, such as VX-809 and IsoLAB, together with evidence thatdrug combinations are a reasonable option in CF treatment (forexample, Kalydeco1VX-809), it might be important to conductmore combination studies and to include thermal stability ex-periments and comparison of the level of activation at RTversus that at 37°C. Nevertheless, our results open new per-spectives for the development of a polypharmacology to rescueF508del-CFTR and suggest that optimized combination of cor-rectors is a feasible option that could lead to a better (near wild-type level) functional rescue of F508del-CFTR.
Acknowledgments
The authors thank Caroline Norez and Isabelle Callebault forhelpful discussion on the project.
Authorship Contributions
Participated in research design: Boinot, Becq.Conducted experiments: Boinot, Jollivet Souchet, Ferru-Clément.Contributed new reagents or analytic tools: Becq.Performed data analysis: Boinot, Jollivet Souchet, Ferru-Clément,
Becq.Wrote or contributed to the writing of the manuscript: Boinot, Becq.
References
Antigny F, Norez C, Becq F, and Vandebrouck C (2008) Calcium homeostasis isabnormal in cystic fibrosis airway epithelial cells but is normalized after rescue ofF508del-CFTR. Cell Calcium 43:175–183.
Ardes-Guisot N, Alonzi DS, Reinkensmeier G, Butters TD, Norez C, Becq F, ShimadaY, Nakagawa S, Kato A, and Blériot Y, et al. (2011) Selection of the biological
activity of DNJ neoglycoconjugates through click length variation of the side chain.Org Biomol Chem 9:5373–5388.
Becq F (2010) Cystic fibrosis transmembrane conductance regulator modulators forpersonalized drug treatment of cystic fibrosis: progress to date. Drugs 70:241–259.
Best D, Jenkinson SF, Waldo Saville A, Alonzi DS, Wormald MR, Butters TD, NorezC, Becq F, Blériot Y, and Adachi I, et al. (2011) Cystic fibrosis and diabetes: isoLABand isoDAB, enantiomeric carbon-branched pyrrolidine iminosugars. TetrahedronLett 51:4170–4174.
Cheng SH, Gregory RJ, Marshall J, Paul S, Souza DW, White GA, O’Riordan CR,and Smith AE (1990) Defective intracellular transport and processing of CFTR isthe molecular basis of most cystic fibrosis. Cell 63:827–834.
Dalemans W, Barbry P, Champigny G, Jallat S, Dott K, Dreyer D, Crystal RG,Pavirani A, Lecocq JP, and Lazdunski M (1991) Altered chloride ion channel ki-netics associated with the delta F508 cystic fibrosis mutation.Nature 354:526–528.
Denning GM, Anderson MP, Amara JF, Marshall J, Smith AE, and Welsh MJ (1992)Processing of mutant cystic fibrosis transmembrane conductance regulator istemperature-sensitive. Nature 358:761–764.
Flume PA, Mogayzel PJ, Jr, Robinson KA, Rosenblatt RL, Quittell L, and MarshallBC Clinical Practice Guidelines for Pulmonary Therapies Committee Cystic Fi-brosis Foundation Pulmonary Therapies Committee (2010) Cystic fibrosis pulmo-nary guidelines: pulmonary complications: hemoptysis and pneumothorax. Am JRespir Crit Care Med 182:298–306.
Haardt M, Benharouga M, Lechardeur D, Kartner N, and Lukacs GL (1999)C-terminal truncations destabilize the cystic fibrosis transmembrane conductanceregulator without impairing its biogenesis: a novel class of mutation. J Biol Chem274:21873–21877.
Haws CM, Nepomuceno IB, Krouse ME, Wakelee H, Law T, Xia Y, Nguyen H,and Wine JJ (1996) Delta F508-CFTR channels: kinetics, activation by forskolin,and potentiation by xanthines. Am J Physiol 270:C1544–C1555.
He L, Kota P, Aleksandrov AA, Cui L, Jensen T, Dokholyan NV, and Riordan JR(2013) Correctors of DF508 CFTR restore global conformational maturation with-out thermally stabilizing the mutant protein. FASEB J 27:536–545.
Heda GD, Tanwani M, and Marino CR (2001) The Delta F508 mutation shortens thebiochemical half-life of plasma membrane CFTR in polarized epithelial cells. Am JPhysiol Cell Physiol 280:C166–C174.
Hutt DM, Herman D, Rodrigues AP, Noel S, Pilewski JM, Matteson J, Hoch B,Kellner W, Kelly JW, and Schmidt A, et al. (2010) Reduced histone deacetylase 7activity restores function to misfolded CFTR in cystic fibrosis. Nat Chem Biol 6:25–33.
Illek B, Fischer H, Santos GF, Widdicombe JH, Machen TE, and Reenstra WW (1995)cAMP-independent activation of CFTR Cl channels by the tyrosine kinase inhibitorgenistein. Am J Physiol 268:C886–C893.
Jouret F, Bernard A, Hermans C, Dom G, Terryn S, Leal T, Lebecque P, CassimanJJ, Scholte BJ, and de Jonge HR, et al. (2007) Cystic fibrosis is associated witha defect in apical receptor-mediated endocytosis in mouse and human kidney. J AmSoc Nephrol 18:707–718.
Liu X, O’Donnell N, Landstrom A, Skach WR, and Dawson DC (2012) Thermal in-stability of DF508 cystic fibrosis transmembrane conductance regulator (CFTR)channel function: protection by single suppressor mutations and inhibiting channelactivity. Biochemistry 51:5113–5124.
Loo TW, Bartlett MC, and Clarke DM (2013) Corrector VX-809 stabilizes the firsttransmembrane domain of CFTR. Biochem Pharmacol 86:612–619.
Ma T, Thiagarajah JR, Yang H, Sonawane ND, Folli C, Galietta LJ, and Verkman AS(2002) Thiazolidinone CFTR inhibitor identified by high-throughput screening blockscholera toxin-induced intestinal fluid secretion. J Clin Invest 110:1651–1658.
Mu TW, Ong DS, Wang YJ, Balch WE, Yates JR, 3rd, Segatori L, and Kelly JW (2008)Chemical and biological approaches synergize to ameliorate protein-folding dis-eases. Cell 134:769–781.
Norez C, Noel S, Wilke M, Bijvelds M, Jorna H, Melin P, DeJonge H, and Becq F(2006) Rescue of functional delF508-CFTR channels in cystic fibrosis epithelialcells by the alpha-glucosidase inhibitor miglustat. FEBS Lett 580:2081–2086.
Odolczyk N, Fritsch J, Norez C, Servel N, da Cunha MF, Bitam S, Kupniewska A,Wiszniewski L, Colas J, and Tarnowski K, et al. (2013) Discovery of novel potentDF508-CFTR correctors that target the nucleotide binding domain. EMBO MolMed 5:1484–1501.
Okiyoneda T, Veit G, Dekkers JF, Bagdany M, Soya N, Xu H, Roldan A, Verkman AS,Kurth M, and Simon A, et al. (2013) Mechanism-based corrector combinationrestores DF508-CFTR folding and function. Nat Chem Biol 9:444–454.
Pedemonte N, Diena T, Caci E, Nieddu E, Mazzei M, Ravazzolo R, Zegarra-Moran O,and Galietta LJ (2005) Antihypertensive 1,4-dihydropyridines as correctors of thecystic fibrosis transmembrane conductance regulator channel gating defect causedby cystic fibrosis mutations. Mol Pharmacol 68:1736–1746.
Pedemonte N and Galietta LJ (2012) Pharmacological correctors of mutant CFTRmistrafficking. Front Pharmacol 3:175.
Quinton PM (1990) Cystic fibrosis: a disease in electrolyte transport. FASEB J 4:2709–2717.
Ramsey BW, Davies J, McElvaney NG, Tullis E, Bell SC, D�revínek P, Griese M,McKone EF, Wainwright CE, and Konstan MW, et al.; VX08-770-102 Study Group(2011) A CFTR potentiator in patients with cystic fibrosis and the G551D muta-tion. N Engl J Med 365:1663–1672.
Riordan JR (1993) The cystic fibrosis transmembrane conductance regulator. AnnuRev Physiol 55:609–630.
Routaboul C, Norez C, Melin P, Molina MC, Boucherle B, Bossard F, Noel S, RobertR, Gauthier C, and Becq F, et al. (2007) Discovery of alpha-aminoazaheterocycle-methylglyoxal adducts as a new class of high-affinity inhibitors of cystic fibrosistransmembrane conductance regulator chloride channels. J Pharmacol Exp Ther322:1023–1035.
Sampson HM, Robert R, Liao J, Matthes E, Carlile GW, Hanrahan JW, and ThomasDY (2011) Identification of a NBD1-binding pharmacological chaperone that cor-rects the trafficking defect of F508del-CFTR. Chem Biol 18:231–242.
Combination of F508del-CFTR Correctors 633
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from

Van Goor F, Hadida S, Grootenhuis PD, Burton B, Cao D, Neuberger T, Turnbull A,Singh A, Joubran J, and Hazlewood A, et al. (2009) Rescue of CF airway epithelialcell function in vitro by a CFTR potentiator, VX-770. Proc Natl Acad Sci USA 106:18825–18830.
Van Goor F, Hadida S, Grootenhuis PD, Burton B, Stack JH, Straley KS, Decker CJ,Miller M, McCartney J, and Olson ER, et al. (2011) Correction of the F508del-CFTR protein processing defect in vitro by the investigational drug VX-809. ProcNatl Acad Sci USA 108:18843–18848.
Wang F, Zeltwanger S, Hu S, and Hwang TC (2000) Deletion of phenylalanine 508causes attenuated phosphorylation-dependent activation of CFTR chloride chan-nels. J Physiol 524:637–648.
Wang W, Okeyo GO, Tao B, Hong JS, and Kirk KL (2011) Thermally unstable gatingof the most common cystic fibrosis mutant channel (DF508): “rescue” by suppressormutations in nucleotide binding domain 1 an by constitutive mutations in thecytocolic loops. J Biol Chem 286:41937–41948.
Ward CL, Omura S, and Kopito RR (1995) Degradation of CFTR by the ubiquitin-proteasome pathway. Cell 83:121–127.
Welsh MJ (1990) Abnormal regulation of ion channels in cystic fibrosis epithelia.FASEB J 4:2718–2725.
Welsh MJ and Smith AE (1993) Molecular mechanisms of CFTR chloride channeldysfunction in cystic fibrosis. Cell 73:1251–1254.
Zeitlin PL (2000) Pharmacologic restoration of delta F508 CFTR-mediated chloridecurrent. Kidney Int 57:832–837.
Address correspondence to: Frédéric Becq, Laboratoire Signalisation etTransports Ioniques, Membranaires, Université de Poitiers, CNRS, Bât. B36,Pôle Biologie Santé, 1 rue Georges Bonnet, TSA 51106, 86073 Poitiers cedex 9,France. E-mail: [email protected]
634 Boinot et al.
at ASPE
T Journals on Septem
ber 11, 2018jpet.aspetjournals.org
Dow
nloaded from





























