Embed Size (px)
Citation preview

Seasonal Variation of the Oviduct of the AmericanAlligator, Alligator mississippiensis (Reptilia: Crocodylia)
April Bagwill,1* David M. Sever,2 and Ruth M. Elsey3
1Department of Zoology, Oklahoma State University, Stillwater, Oklahoma 740782Department of Biological Sciences, Southeastern Louisiana University, Hammond, Louisiana 704023Louisiana Department of Wildlife and Fisheries, Rockefeller Wildlife Refuge, Grand Chenier, Louisiana 70643
ABSTRACT The annual oviductal cycle of the Americanalligator, Alligator mississippiensis, is described using lightand electron microscopy. Previous work done by Palmerand Guillette ([1992] Biol Reprod 46:39–47) shed somelight on the reproductive morphology of the female alliga-tor oviduct; however, their study was limited and did notreport details relating to variation across the reproductiveseason. We recognize six variable regions of the oviduct: in-fundibulum, tube, isthmus, anterior uterus, posterioruterus, and vagina. Each area shows variation, to somedegree, in the histochemistry and ultrastructure of oviduc-tal secretions. Peak secretory activity occurs during themonths of May and June, with the greatest variation occur-ring in the tube and anterior uterus. During the month ofMay, high densities of neutral carbohydrates and proteinsare found within the tubal and anterior uterine glands.The epithelium of the entire oviduct secretes neutral carbo-hydrates throughout the year, but many regions lack pro-tein secretions, and the posterior uterine glands show littlesecretory activity of any type throughout the year. Afteroviposition, secretory activity decreases drastically, and theoviduct resembles that of the premating season. This studyalso provides evidence to support the homology between al-ligator and bird oviducts. Sperm were observed in glandsat the tubal-isthmus and utero-vaginal junctions inpreovulatory, postovulatory and postovipository females.J. Morphol. 270:702–713, 2009. � 2009 Wiley-Liss, Inc.
KEY WORDS: alligator; reproductive, oviduct, spermstorage, morphology
INTRODUCTION
Crocodilians are grouped with both birds anddinosaurs to comprise the Archosauria (Gauthier,1984, 1986). This grouping has traditionally beenbased upon morphological evidence (Gauthieret al., 1988; Hutchinson and Gatesy, 2000; Ras-skin-Gutman and Buscalioni, 2001), but moleculardata have more recently been used to confirm thisrelationship (Brochu, 2001; Chang et al., 2002).The 23 extant species of crocodilians occur circum-tropically. Currently, they are divided into threefamilies: Alligatoridae (four genera, eight species),Crocodylidae (four genera, 14 species), and Gavia-lidae (one genus, one species). Research on thebiology of crocodilians is limited, and the moststudied species is the American alligator, Alligatormississippiensis.
The range of the American alligator spans thesoutheastern coastal plain from central Texas eastto North Carolina, with small populations reachingthe southeast corner of Oklahoma and the south-ern third of Arkansas in the most northern part oftheir range (Conant and Collins, 1998). Alligatorswere close to extinction during the 1970s due tooverhunting; however, effective management planswere put into place and now the population is wellover several million. Louisiana has the highestpopulation at around 1.5 million wild alligators(Louisiana Department of Wildlife and Fisheries,2007), followed by Florida with around 1 million(Florida Fish and Wildlife Conservation Commis-sion, 2007). Alligators typically inhabit freshwaterswamps and marshes, and occasionally are foundin brackish water. In the more temperate regionsof their range, they create burrows for shelter/hibernation during the cooler months.
Reproductive characteristics of both male and femalealligators in southern Louisiana were described byLance (1989) using specimens mostly obtained fromspring to late summer, the period thought to encom-pass the breeding season. Reports also exist on femalecycles of populations in Florida (Guillette et al., 1997;Uribe and Guillette, 2000) and South Carolina(Wilkinson, 1983). Reproduction can occur annu-ally, although some individuals may not reproducein consecutive years. Courtship usually starts inlate April to early May, with mating beginning inmid-late May, and oviposition commences in lateJune–July (Joanen and McNease, 1980; Lance,1989). The reproductive season varies, however,between populations across the range distribution
Contract grant sponsor: National Science Foundation; Contractgrant number: DEB-0809831.
*Correspondence to: April Bagwill, Department of Zoology, Okla-homa State University, Stillwater, OK 74078.E-mail: [email protected]
Received 14 May 2008; Revised 21 October 2008;Accepted 25 October 2008
Published online 10 February 2009 inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/jmor.10714
JOURNAL OF MORPHOLOGY 270:702–713 (2009)
� 2009 WILEY-LISS, INC.

as well as with the temperature of the given year(Klause, 1984; Jacobsen and Kushlan, 1986; Joa-nen and McNease, 1989).
Female alligators in Florida have been shown tobegin vitellogenesis in October, slowing over thewinter, and continuing at the beginning of spring(Guillette et al., 1997). Populations from Louisianahave been reported to begin vitellogenesis inMarch (Joanen and McNease, 1980; Lance, 1989),but sample sizes from fall are small. Less is knownabout folliculogenesis and fertilization, but imma-ture follicles grow from 1 to 45 mm at the time ofovulation (Lance, 1989).
Seasonal variation within the oviduct of theAmerican alligator was previously described byPalmer and Guillette (1992). They reported onlyon late-vitellogenic and gravid females from a pop-ulation in Florida. This study aims to further theknowledge of the histological and ultrastructuralvariations in the oviduct of the American alligatorpresented by Palmer and Guillette (1992), as wellas to extend their previous study by including femalealligators across the entire reproductive season.
MATERIALS AND METHODSSpecimens
Adult female alligators were collected in southern Louisianafrom October 2005 to June 2007 at three main locations: Rock-efeller Wildlife Refuge, Grand Chenier, LA; Manchac WildlifeManagement Area, Manchac, LA; and Vermilion Gator Farm,Abbeville, LA. The alligators were captured by either settingbaited hooks or using a four-foot snare. Once collected, thespecimens were killed by severing the spinal cord. Specimenswere collected under permit number WL-Research-2006-03issued by Louisiana Department of Wildlife and Fisheries. Pro-tocols were approved by the Institutional Review Board ofSoutheastern Louisiana University.Dissection of the reproductive tract was performed, and the
left oviduct was injected with and placed in Trump’s fixative(2.5% glutaraldehyde and 2.5% formaldehyde in 0.1 M sodiumcacodylate buffer at pH 7.4; Electron Microscopy Sciences, Fort
Worth, PA) for ultrastructural investigation, whereas the rightoviduct was placed in 10% neutral buffered formalin for histo-logical investigation. The size and reproductive condition ofeach of the specimens are listed in Table 1. Adobe Photoshop7.0 was used for editing of all micrographs (Adobe Systems, SanJose, CA).
Light Microscopy
Tissues were taken from the right oviduct at 10 areas: infun-dibulum, anterior tube, posterior tube, tubal-isthmus junction,isthmus, isthmus-uterine junction, anterior uterus, posterioruterus, uterovaginal junction, and vagina. The tissues wererinsed for 30 min under running water and dehydrated in agraded series of ethanol (70% for 30 min, 95% for 30 min, andtwo cycles of 100% at 30 min each). After dehydration, the tis-sues were placed in toluene for two changes at 30 min each.The tissues were subsequently placed in paraffin overnightunder vacuum and embedded in paraffin blocks the followingmorning. Sections 10-lm thick were cut using a MR3 microtome(Research and Manufacturing Co., Tucson, AZ) and adhered toalbuminized slides. Alternating slides were stained with hema-toxylin and counterstained with eosin (H&E) for initial histolog-ical examination. The remaining slides were alternately stainedwith either bromphenol blue (BB) for proteins or treated withperiodic acid-Schiff ’s (PAS) for neutral carbohydrates with analcian blue at pH 2.5 (AB) counterstain for carboxylated glycos-aminoglycans. The slides were viewed with a Leica DM2000microscope (Leica Microsystems, Wetzlar, Germany), andimages were acquired with a Leica DFC420 camera (LeicaMicrosystems, Wetzlar, Germany).
Transmission Electron Microscopy
Tissues from the left oviduct were rinsed with deionizedwater for 30 min and postfixed with osmium tetroxide for 90min. After postfixation, the tissues were rinsed again (30 min)and dehydrated in a graded series of ethanol (70%, 95%, and100%) for 30 min each. The tissues were subsequently placed ina 1:1 ratio of 100% ethanol and propylene oxide for 30 min andcleared in pure propylene oxide for 30 min. The cleared tissueswere placed in a 1:1 mixture of propylene oxide and epoxy resin(EmBed 812, Electron Microscopy Sciences, Fort Washington,PA), a 2:1 mixture of resin and propylene oxide, and finally intopure epoxy resin and placed under vacuum for 12 h. The tissuesand fresh resin were then placed into molds to be hardened at
TABLE 1. Reproductive condition and the time of year specimens were collecteda
Date SVL (m) TL (m) Sperm Reproductive condition
8 Feb 0.99 2.08 no enlarged follicles30 Mar 0.91 1.88 no enlarged follicles6 Apr 0.99 2.08 no enlarged follicles10 May 0.95 1.96 no enlarged follicles10 May Not taken 2.08 no enlarged follicles10 May Not taken 1.98 no enlarged follicles17 May Not taken 1.8 one 2-cm follicle31 May 1.13 2.27 TUBAL-ISTHMUS JUNCTION,
UTERO-VAGIONAL JUNCTIONpreovulatory, 36 ovarian follicles �3 cm
31 May 1.04 2.18 TUBAL-ISTHMUS JUNCTION,UTERO-VAGIONAL JUNCTION
postovulatory, 13 eggs left, 11 eggs right
13 Jun 1.04 2.12 UTERO-VAGIONAL JUNCTION postovipository, at nest 40 eggs13 Jun 0.94 1.96 UTERO-VAGIONAL JUNCTION postovipository, at nest 20 eggs14 Jun 0.89 1.88 no enlarged follicles19 Jun 0.82 1.64 one 2-cm follicle21 Jun 0.99 2.01 preovulatory, 25 ovarian follicles 2–3.5-cm10 Oct 1.3 2.54 no enlarged follicles
aSperm present in either the tubal-isthmus junction (TUBAL-ISTHMUS JUNCTION) and the utero-vaginal junction (UTERO-VAGIONAL JUNCTION).
ALLIGATOR OVIDUCT 703
Journal of Morphology

608C. The tissues were thick-sectioned at 0.5-lm using a RMCMT7 ultramicrotome (Research and Manufacturing Co., Tucson,AZ) and thin-sectioned at 70 nm with a Leica Reichert UltracutS ultramicrotome (Leica Microsystems, Wetzlar, Germany)using a DiATOME diamond knife (DiATOME, Biel, Switzer-land). The sections were placed on copper grids, stained withuranyl acetate and lead citrate, and viewed with a JEOL JEM100s transmission electron microscope (TEM) (JEOL USA, Pea-body, MA). Images were taken on Kodak film, and negativeswere scanned using a Microtek ScanMaker i900 (MicrotekInternational, Carson, CA).
Scanning Electron Microscopy
Through dehydration, procedures followed those for TEM.Tissues were critically point dried using a DCP-1 critical pointdrier (Denton Vacuum, Moorestown, NJ). The tissues wereadhered to stubs and sputter-coated with gold, using a DentonDesk IV XLS (Denton Vacuum, Moorestown, NJ), and viewedon a Philips XL-20 scanning electron microscope (SEM) (PhilipsElectronics N.V., Eindoven, the Netherlands).
RESULTS
We distinguish six regions of the American alli-gator oviduct (cranial to caudal): infundibulum,tube (tuba uterina), isthmus, anterior uterus, pos-terior uterus, and vagina (see Fig. 1). We do notdistinguish a posterior infundibulum (Palmer andGuillette, 1992; Perez and Pinilla, 2002), which we
recognize simply as the transition between theinfundibulum and the tube.
The entire oviduct is lined by a simple epithe-lium composed of ciliated cells and nonciliated se-cretory cells. Throughout the year, the epitheliumof the tube and uterus remains active, whereas theproduction of secretory material in the glandsvaries, being absent in some regions in certainmonths (Tables 2, 3). During the inactive periods,neutral carbohydrates and acidic mucoids oftenare still abundant mostly in the apical portions ofthe cells. Overall, a peak in the amount of secre-tory material occurs during the months of Mayand June and decreases after oviposition to thepremating season levels. This trend also corre-sponds to the peaks of hormone activity reportedby Lance (1989) and Guillette et al. (1997).
This study recognizes two distinct regions of theoviduct which attract aggregates of sperm cellsinto the tubular glands that occupy the mucosa.These areas are the tubal-isthmus junction andthe utero-vaginal junction. Both are at the mostposterior end, respectively, of the tube and theuterus, and they possess glands that do not differfrom those in adjoining areas except for the pres-ence of sperm. These regions contain sperm withinthe glands subsequent to the alligator mating sea-son, which suggests that the purpose of theseglands is to store sperm. Gist et al. (2008) providea detailed description of the annual cycle of spermstorage in female Alligator mississippiensis.
Infundibulum
The infundibulum is the most anterior portion ofthe oviduct and lacks tubular glands (Figs. 1A, 2).The tissue layers are very thin, and the infundibu-lum appears translucent at the gross morphologi-cal level. The inner lining consists of many longi-tudinal folds that create a large amount of surfacearea. The epithelial cells are squamous to cuboidaland stain basophilic with H&E. The epithelial cellsare glandular, and throughout the reproductivecycle variations occur in their secretions (Table 2).During the inactive periods slight amounts of neu-tral carbohydrates are detected, as well as protein.However, during the reproductive months, theselevels increase in density and secretions also stainpositively for acidic mucoids. After nesting, thelevels of all secretions diminish.
The intercellular canaliculi between the epithe-lial cells are usually very labyrinthine and rela-tively wide (Fig. 2A,B), although they narrow inJune in females that have oviposited (Fig. 2C).The nuclei are euchromatic with abundant mito-chondria surrounding them in inactive months(Fig. 2A) and become more heterochromatic duringthe reproductive period (Fig. 2B,C). The secretorymaterials are released into the lumen via an apoc-rine process (Fig. 2B).
Fig. 1. Gross morphology of a reproductively active oviductof Alligator mississippiensis. (A) Various regions of the oviduct.(B) The transitional region between the tube and uterus, theisthmus. (C) The intramural portion of the vagina via an inci-sion through the center of the cloaca. An asterisk (*) indicatesareas where tubular glands possess sperm in reproductivelyactive females. Aut, anterior uterus; Inf, infundibulum; Isth,isthmus; Put, posterior uterus; Ut, uterus, Vag, vagina.
704 A. BAGWILL ET AL.
Journal of Morphology

SEM shows the relationships between ciliatedand secretory cells (Fig. 2D). The quantity of cili-ated cells increases during inactive months.
Tube
The tube is just posterior to the infundibulumand is highly convoluted when the mesentery isintact, but flat once separated. The tube isinvolved in the synthesis of albumin (Lance, 1989).The tube contains secretory products in the epithe-lium lining the lumen as well as in compound tu-bular glands (Figs. 3, 4), and the products of thesetwo secretory areas differ (Tables 2, 3). The colum-nar epithelium lining the lumen of the tube is usu-ally PAS-positive, AB-positive, and BB-negativewhereas the tubal glands are PAS-positive, AB-negative, and BB-positive (Fig. 3A,B). SEM showsthat secretory activity continues throughout thereproductive season, although more secretory ma-terial is evident in recently mated specimens (Fig.3C) than one sacrificed after oviposition (Fig. 3D).
Secretory material is heterogeneously electron-dense in the tubal lining of recently mated females(Fig. 4A) and homogeneous in the tubal glands(Fig. 4B). In females collected after oviposition, se-cretory vacuoles appear heterogeneous in both thetubal lining (Fig. 4C) and tubal glands (Fig. 4D).The intercellular canaliculi are narrow and havefewer interdigitations before oviposition (Fig.4A,B) than after (Fig. 4C,D). The abundance ofsecretory material during the breeding seasonobscures the presence of other cellular organelles.
Isthmus
The isthmus is the transition area between thetube and the uterus (Fig. 1B). The isthmus con-
tains ciliated and secretory columnar cells butlacks tubular glands. On the gross morphologicalscale, it appears very flat and somewhat translu-cent, averaging around 2.5 cm in length in repro-ductively active females. Before mating, the apicalportion of the epithelium is full of AB-positive ma-terial, which becomes absent during peak matingin May when the entire epithelium stains highlyPAS-positive (Table 2). After oviposition, the apicalportions of the cells secrete both neutral carbohy-drates (PAS-positive) and acidic mucosubstances(AB-positive). Protein secretion is absent or scantthroughout the year (Table 2).
Ultrastructurally, secretory vacuoles of mediumdensity are present in the apical portions of cells
TABLE 2. Abundance of secretions found within the epithelium of the oviducta
DateInf
PAS/ABInfBB
TubePAS/AB
TubeBB
IsthPAS/AB
IsthBB
AutPAS/AB
AutBB
PutPAS/AB
PutBB
VagPAS/AB
VagBB
8 Feb 0/2 0 1/1 0 0/2 0 1/0 0 1/0 0 2/2 030 Mar 0/0 0 2/1 0 2/2 0 1/0 0 0/1 0 N/A N/A6 Apr 0/0 0 0/1 0 0/2 0 1/0 22 2/2 2 N/A N/A10 May 0/0 2 2/2 22 1/1 0 1/0 22 0/0 22 N/A N/A10 May 0/2 0 1/1 0 2/1 0 1/2 0 1/2 0 N/A N/A10 May 2/0 0 1/1 0 0/0 0 1/1 0 1/1 0 N/A N/A17 May 2/2 0 2/2 0 2/11 0 2/2 0 2/0 0 N/A N/A17 May 2/0 0 11/1 0 2/2 0 11/2 0 1/1 0 N/A N/A31 May 1/0 1 11/2 0 1/0 2 1/1 0 1/2 2 1/0 231 May 1/0 2 11/1 0 11/2 0 11/2 0 1/2 2 2/1 213 Jun 2/0 0 11/2 0 1/2 0 11/0 0 11/0 0 2/22 113 Jun 2/2 0 1/1 0 1/2 2 11/0 0 1/2 22 11/1 114 Jun 2/2 2 1/1 0 1/2 0 11/0 0 1/0 0 N/A N/A19 Jun 2/0 2 11/1 0 2/1 2 11/0 2 2/2 0 N/A N/A21 Jun 2/2 2 11/11 0 2/2 2 1/1 0 2/2 2 N/A N/A10 Oct 2/2 0 11/11 0 N/A N/A 11/0 0 1/2 0 2/0 0
aAut, anterior uterus; Inf, infundibulum; Isth, isthmus; Put, posterior uterus; Tube, tube; Vag, vagina. 0, no secretion; 2, scantsecretion; 22, very scant secretion; 1, abundant secretion; 11, very abundant secretion; N/A, tissue not available; BB, bromphe-nol blue (proteins); PAS/AB, periodic acid-Schiff/alcian blue (neutral carbohydrates/glycosaminoglycans).
TABLE 3. Abundance of secretions found within the glandsof the oviducta
DateTgl
PAS/ABTglBB
AuglPAS/AB
AugBB
PuglPAS/AB
PugBB
8 Feb 2/0 1 2/2 2 0/0 030 Mar 1/2 1 2/0 0 2/0 06 Apr 2/0 2 0/0 0 0/0 010 May 2/0 2 0/0 0 0/0 010 May 1/2 22 22/22 0 22/0 010 May 2/2 0 2/2 0 2/22 017 May 2/2 0 2/0 0 2/2 017 May 11/0 11 2/2 1 2/0 031 May 11/0 11 2/1 11 1/2 031 May 11/0 11 2/1 1 1/2 013 Jun 11/0 1 0/0 1 2/0 013 Jun 11/0 1 2/22 1 2/0 014 Jun 11/0 1 0/0 2 0/0 019 Jun 11/0 1 2/2 1 2/0 021 Jun 11/0 11 1/2 11 2/2 210 Oct 11/0 0 2/0 0 2/0 0
aAugl, anterior uterine gland; Pugl, posterior uterine gland;Tgl, tubal gland BB, bromphenol blue; PAS/AB, periodic acid-Schiff/alcian blue (neutral carbohydrates/glycosaminoglycans);0, no secretion; 2, scant secretion; 22, very scant secretion; 1,abundant secretion; 11, very abundant secretion.
ALLIGATOR OVIDUCT 705
Journal of Morphology

in preovipository females (Fig. 5A,B), and secre-tions are not as abundant as in the tubal lining. Infemales collected after oviposition, the secretorymaterial is scant and secretory vacuoles are elec-tron-lucent (Fig. 5C). In a June postovipositoryfemale, electron-lucent secretory vacuoles are stillpresent (Fig. 6C). Intercellular canaliculi are dis-tinct in active females from May (Fig. 5A) whereasin June these cell junctions narrow and are diffi-cult to distinguish (Fig. 5C). SEM confirms thatciliated cells are more numerous than secretorycells, even in recently mated females (Fig. 5D).
Anterior Uterus
The anterior uterus is another highly glandulararea with branched tubular endometrial glands.This area starts the formation of the eggshellmembranes (Buhi et al., 1999). As in the tube, theepithelial cells of the anterior uterine lining differin secretory activity from the anterior uterineglands (Fig. 6A,B; Tables 2, 3). The epithelial lin-ing of the anterior uterus consists of columnarcells with basal nuclei; these cells are basophilicthroughout the entire year. The epithelium pro-duces PAS1 material during the active months,which is present throughout the entire cell (Fig.6A). During the rest of the year, this material isstill present, but is located only in the apical por-tion of the cells. The epithelium lining the anterioruterus is AB-negative and BB-negative (Fig.6A,B). In contrast, the anterior uterine glands are
Fig. 2. The infundibulum of Alligator mississippiensis. (A)Transmission electron micrograph shows the labyrinthine inter-cellular canaliculi and the abundant mitochondria in the basalportion of the cell of a reproductively inactive female collected 8February. (B) Transmission electron micrograph depicting theapocrine secretions released from the epithelium and the wideintercellular canaliculi of a postovulatory female collected 31May. (C) Transmission electron micrograph showing the ciliatedcells and narrow intercellular canaliculi of postovipositoryfemale collected 13 June. (D) Scanning electron micrograph of apreovulatory female collected 31 May illustrating the associa-tion between secretory and ciliated epithelium. Ab, apocrineblebs; Bb, basal bodies; Bl, basal lamina; Cf, collagen fibers; Ci,cilia; Ds, desmosomes; Ic, intercellular canaliculi; Lu, lumen;Mi, mitochondria; Nu, nucleus; Rbc, red blood cell; Sv, secretoryvacuoles; Tj, tight junction.
Fig. 3. The tube of Alligator mississippiensis. (A) Lightmicrograph of a postovulatory female collected 31 May treatedwith PAS for neutral carbohydrates and counterstained withAB for glycosaminoglycans. (B) Same female used in A, stainedwith BB for proteins. (C) Scanning electron micrograph throughthe tubal lining of a preovulatory female collected 31 May. (D)Scanning electron micrograph through the tubal linining of apostovipository female collected 13 June. AB1, alcian blue-posi-tive; BB1, bromphenol blue-positive; Ci, cilia; Ep, epithelium;PAS1, periodic acid/Schiff ’s reagent-positive; Sm, secretorymaterial; Sv, secretory vacuoles; Tbg, tubal glands.
706 A. BAGWILL ET AL.
Journal of Morphology

strongly BB-positive (Fig. 6B) and show mixedreactions to carbohydrates. In recently matedspecimens, much of the epithelium is lightly PAS-positive but the apical cytoplasm of some glandscan stain intensely AB-positive (Fig. 6A). SEMreveals the abundance of secretory materialthroughout the reproductive season in activefemales (Fig. 6C,D). The clusters of secretory ma-terial on the surface of the uterine lining, whichare similar to secretory vacuoles observed in thecytoplasm, indicate that an apocrine process isalso involved in release of material from thisportion of the oviduct.
Before the mating season and in early May, theepithelial lining of the anterior uterus consists oftall columnar cells with dark cytoplasm and littlesecretory material. Once the mating season beginslater in May, the amount of secretory materialgreatly increases and light cells appear (Fig. 7A).
The light cells, with less electron-dense cytoplasmthan dark cells, can contain secretory vacuoles ofvarious sizes and densities (Fig. 7A). The darkcells, however, seem to contain only smallvacuoles, which like the cytoplasm, are uniformlyelectron-dense (Fig. 7A). The anterior uterineglands contain only dark cells, and the secretoryvacuoles are quite heterogeneous in both size anddensity (Fig. 7B).
In postovipository females examined from June,the light cells are no longer observed in the uter-ine lining, and the dark cells are still filled withsmall secretory vacuoles, which are so numerous,that they distend the shape of nuclei in the cells(Fig. 7C). Occasional apical cells are observed withdisorganized cytoplasm, and we propose that theseare cells that have released their product throughapocrine processes and are undergoing apoptosis.
Fig. 4. The tube of Alligator mississippiensis. TEM. (A)Tubal lining epithelium of a preovulatory female collected 31May showing abundant heterogeneous secretory vacuoles andapocrine secretions of the epithelium. (B) Tubal gland of thefemale used in A, illustrating large homogeneous secretoryvacuoles. (C) Tubal lining of a postovipository female collected13 June showing fewer secretory vacuoles than the May speci-men. (D) Heterogeneous secretory material in the tubal glandsof the June female used in C. As, apocrine secretion; Ci, cilia;Ic, intercellular canaliculi; Lu, lumen; Mi, mitochondria; Mv,microvilli; No, nucleolus; Nu, nucleus; Sv, secretory vacuole.
Fig. 5. The isthmus of Alligator mississippiensis. (A) Trans-mission electron micrograph depicting the alternating ciliatedand microvillus-secretory cells of the epithelium lining the isth-mus of a preovulatory female collected 31 March. (B) Transmis-sion electron micrograph showing abundant ciliated cells andsecretory cells with supranuclear electron-lucent secretory ma-terial in a preovulatory female collected 21 June. (C) Transmis-sion electron micrograph of a postovipository female collected13 June showing a decrease in secretory material. (D) Scanningelectron micrograph of a postovulatory female collected 31 May.Bb, basal bodies; Ci, cilia; Ep, epithelium; Ic, intercellularcanaliculi; Mi, mitochondria; Mv, microvilli; No, nucleolus; Nu,nucleus; Sv, secretory vacuoles.
ALLIGATOR OVIDUCT 707
Journal of Morphology

In the anterior uterine glands of female after ovi-position, secretory vacuoles of various sizes anddensities are still abundant (Fig. 7D). The lucidityof secretory material in lumen indicates thattransformation of the products has occurred dur-ing the passage of eggs.
Posterior Uterus
The epithelial lining and tubular glands of theposterior uterus differ in secretory activity fromthe anterior uterus (Figs. 8, 9; Tables 2, 3). Thisregion is presumably involved in secreting the cal-careous material of the eggshell (Palmer and Guil-lette, 1992). We did not detect, however, a highdegree of secretory activity with ultrastructure
examination (see Fig. 8), although histochemicallycarbohydrate production was noted (Fig. 9A).
As in the anterior uterus, both electron-lightand electron-dense cells are present in the poste-rior uterine lining but absent in the glands (seeFig. 8). In contrast to the anterior uterus, however,dark cells persist in the posterior uterine lining af-ter oviposition (Fig. 8A,C). Only one type of secre-tory vacuole with medium density is present in thelining of the posterior uterus, and these are re-stricted to the dark cells (Fig. 8A,C). The epithelialcells of the posterior uterine have barely distin-guishable intercellular canaliculi (Fig. 8A,C).
The posterior uterine glands of recently matedfemales have wide lumina, wide and interdigitat-ing intercellular canaliculi, and numerous infranu-clear mitochondria (Fig. 8B). They are also charac-
Fig. 6. The anterior uterus of Alligator mississippiensis. (A)Light micrograph of the anterior uterine lining of a postovula-tory female collected 31 May treated with PAS for neutral car-bohydrates and counterstained with AB for glycosaminoglycans.(B) Same female used in A, stained with BB for proteins. (C)Scanning electron micrograph of the anterior uterine epithe-lium of the female used in A showing abundance of secretoryvacuoles. (D) Scanning electron micrograph of the anterior uter-ine lining of a postovipository female collected 13 June. AB1,alcian blue-positive; Aug, anterior uterine gland; Auglu, ante-rior uterine gland lumen; Aul, anterior uterine lining; BB1,bromphenol blue-positive; Ci, cilia; Ep, epithelium; PAS1, peri-odic acid/Schiff ’s reagent-positive; Sm, secretory material; Sv,secretory vacuoles.
Fig. 7. The anterior uterus of Alligator mississippiensis.TEM. (A) The epithelium of a May postovulatory female show-ing light cells, dark cells, and secretory vacuoles of varioussizes. (B) Tubular gland cytoplasm of the May female used in Ashowing large, heterogeneous secretory vacuoles. (C) Epithe-lium of a June postovulatory female with no light cells andabundant secretory vacuoles throughout the cells. (D) Tubulargland cytoplasm of the June female used in C with a largelumen and abundant secretory material. Apc, cell undergoingapoptosis; DcSSv, dark cells with small secretroy vacuoles; Ic,intercellular canaliculi; Lc, light cells; Lg, gland lumen; Lsv,large secretory vacuoles; Lu, lumen; Mi, mitochondria; Mv, mi-crovilli; Nu, nucleus; Smlu, secretory material in the lumen; Sv,secretory vacuole; Tj, tight junction.
708 A. BAGWILL ET AL.
Journal of Morphology
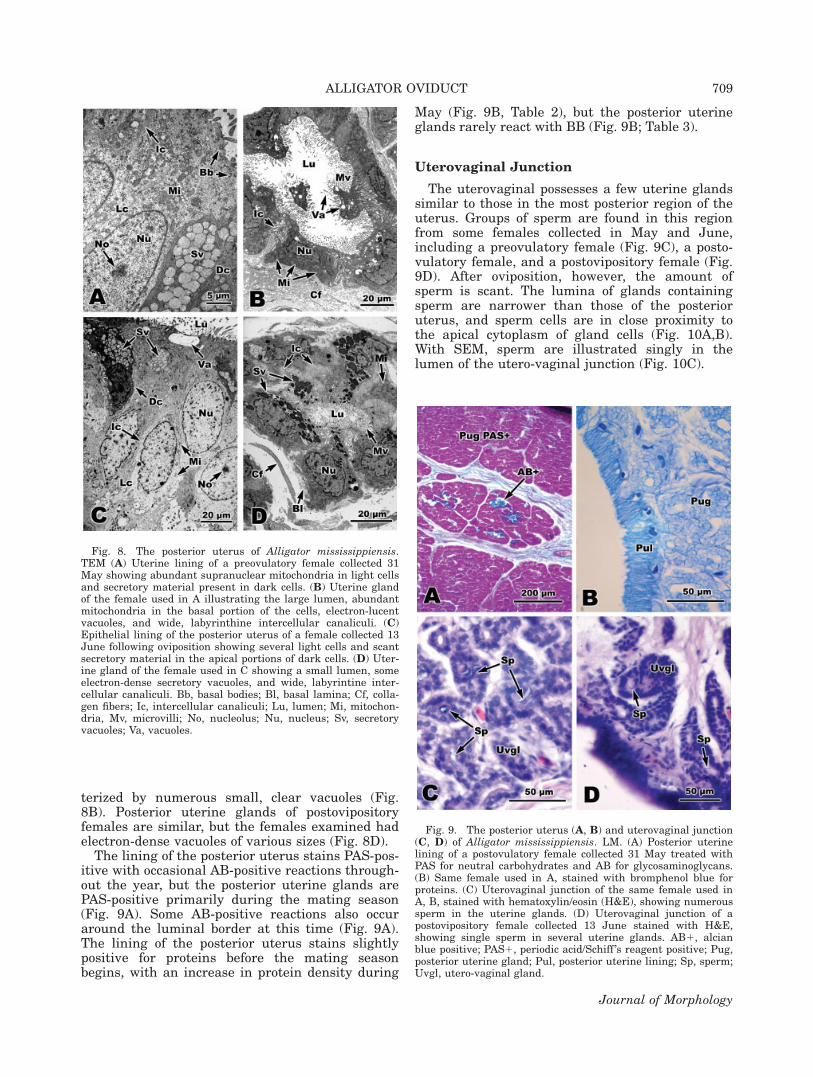
terized by numerous small, clear vacuoles (Fig.8B). Posterior uterine glands of postovipositoryfemales are similar, but the females examined hadelectron-dense vacuoles of various sizes (Fig. 8D).
The lining of the posterior uterus stains PAS-pos-itive with occasional AB-positive reactions through-out the year, but the posterior uterine glands arePAS-positive primarily during the mating season(Fig. 9A). Some AB-positive reactions also occuraround the luminal border at this time (Fig. 9A).The lining of the posterior uterus stains slightlypositive for proteins before the mating seasonbegins, with an increase in protein density during
May (Fig. 9B, Table 2), but the posterior uterineglands rarely react with BB (Fig. 9B; Table 3).
Uterovaginal Junction
The uterovaginal possesses a few uterine glandssimilar to those in the most posterior region of theuterus. Groups of sperm are found in this regionfrom some females collected in May and June,including a preovulatory female (Fig. 9C), a posto-vulatory female, and a postovipository female (Fig.9D). After oviposition, however, the amount ofsperm is scant. The lumina of glands containingsperm are narrower than those of the posterioruterus, and sperm cells are in close proximity tothe apical cytoplasm of gland cells (Fig. 10A,B).With SEM, sperm are illustrated singly in thelumen of the utero-vaginal junction (Fig. 10C).
Fig. 8. The posterior uterus of Alligator mississippiensis.TEM (A) Uterine lining of a preovulatory female collected 31May showing abundant supranuclear mitochondria in light cellsand secretory material present in dark cells. (B) Uterine glandof the female used in A illustrating the large lumen, abundantmitochondria in the basal portion of the cells, electron-lucentvacuoles, and wide, labyrinthine intercellular canaliculi. (C)Epithelial lining of the posterior uterus of a female collected 13June following oviposition showing several light cells and scantsecretory material in the apical portions of dark cells. (D) Uter-ine gland of the female used in C showing a small lumen, someelectron-dense secretory vacuoles, and wide, labyrintine inter-cellular canaliculi. Bb, basal bodies; Bl, basal lamina; Cf, colla-gen fibers; Ic, intercellular canaliculi; Lu, lumen; Mi, mitochon-dria, Mv, microvilli; No, nucleolus; Nu, nucleus; Sv, secretoryvacuoles; Va, vacuoles.
Fig. 9. The posterior uterus (A, B) and uterovaginal junction(C, D) of Alligator mississippiensis. LM. (A) Posterior uterinelining of a postovulatory female collected 31 May treated withPAS for neutral carbohydrates and AB for glycosaminoglycans.(B) Same female used in A, stained with bromphenol blue forproteins. (C) Uterovaginal junction of the same female used inA, B, stained with hematoxylin/eosin (H&E), showing numeroussperm in the uterine glands. (D) Uterovaginal junction of apostovipository female collected 13 June stained with H&E,showing single sperm in several uterine glands. AB1, alcianblue positive; PAS1, periodic acid/Schiff ’s reagent positive; Pug,posterior uterine gland; Pul, posterior uterine lining; Sp, sperm;Uvgl, utero-vaginal gland.
ALLIGATOR OVIDUCT 709
Journal of Morphology
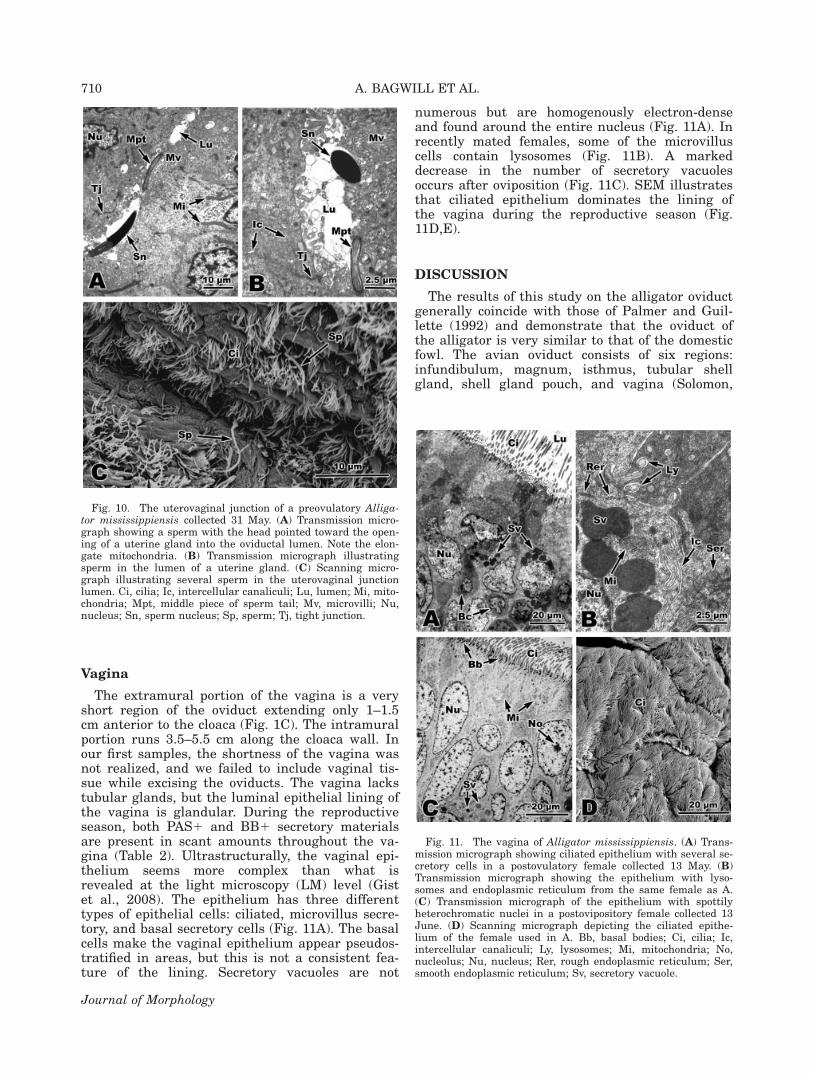
Vagina
The extramural portion of the vagina is a veryshort region of the oviduct extending only 1–1.5cm anterior to the cloaca (Fig. 1C). The intramuralportion runs 3.5–5.5 cm along the cloaca wall. Inour first samples, the shortness of the vagina wasnot realized, and we failed to include vaginal tis-sue while excising the oviducts. The vagina lackstubular glands, but the luminal epithelial lining ofthe vagina is glandular. During the reproductiveseason, both PAS1 and BB1 secretory materialsare present in scant amounts throughout the va-gina (Table 2). Ultrastructurally, the vaginal epi-thelium seems more complex than what isrevealed at the light microscopy (LM) level (Gistet al., 2008). The epithelium has three differenttypes of epithelial cells: ciliated, microvillus secre-tory, and basal secretory cells (Fig. 11A). The basalcells make the vaginal epithelium appear pseudos-tratified in areas, but this is not a consistent fea-ture of the lining. Secretory vacuoles are not
numerous but are homogenously electron-denseand found around the entire nucleus (Fig. 11A). Inrecently mated females, some of the microvilluscells contain lysosomes (Fig. 11B). A markeddecrease in the number of secretory vacuolesoccurs after oviposition (Fig. 11C). SEM illustratesthat ciliated epithelium dominates the lining ofthe vagina during the reproductive season (Fig.11D,E).
DISCUSSION
The results of this study on the alligator oviductgenerally coincide with those of Palmer and Guil-lette (1992) and demonstrate that the oviduct ofthe alligator is very similar to that of the domesticfowl. The avian oviduct consists of six regions:infundibulum, magnum, isthmus, tubular shellgland, shell gland pouch, and vagina (Solomon,
Fig. 10. The uterovaginal junction of a preovulatory Alliga-tor mississippiensis collected 31 May. (A) Transmission micro-graph showing a sperm with the head pointed toward the open-ing of a uterine gland into the oviductal lumen. Note the elon-gate mitochondria. (B) Transmission micrograph illustratingsperm in the lumen of a uterine gland. (C) Scanning micro-graph illustrating several sperm in the uterovaginal junctionlumen. Ci, cilia; Ic, intercellular canaliculi; Lu, lumen; Mi, mito-chondria; Mpt, middle piece of sperm tail; Mv, microvilli; Nu,nucleus; Sn, sperm nucleus; Sp, sperm; Tj, tight junction.
Fig. 11. The vagina of Alligator mississippiensis. (A) Trans-mission micrograph showing ciliated epithelium with several se-cretory cells in a postovulatory female collected 13 May. (B)Transmission micrograph showing the epithelium with lyso-somes and endoplasmic reticulum from the same female as A.(C) Transmission micrograph of the epithelium with spottilyheterochromatic nuclei in a postovipository female collected 13June. (D) Scanning micrograph depicting the ciliated epithe-lium of the female used in A. Bb, basal bodies; Ci, cilia; Ic,intercellular canaliculi; Ly, lysosomes; Mi, mitochondria; No,nucleolus; Nu, nucleus; Rer, rough endoplasmic reticulum; Ser,smooth endoplasmic reticulum; Sv, secretory vacuole.
710 A. BAGWILL ET AL.
Journal of Morphology

1983). Crocodilians have the primitive archosau-rian trait of possessing paired oviducts and ovulat-ing eggs through each simultaneously (Lance,1989; Varricchio et al., 1997; Sato et al., 2005),whereas most birds only develop one oviduct. How-ever, birds also share traits with primitive archo-saurs in that only one egg at a time is ovulatedand shelled (Solomon, 1983; Varricchio et al., 1997;Sato et al., 2005).
Buhi et al. (1999) found that a large number ofproteins are synthesized de novo in the alligatoroviduct, and these proteins have different biosyn-thetic activity correlated with location in the ovi-duct and reproductive status of the female. Theysuggested that the distribution of proteins synthe-sized and released support the proposal by Palmerand Guillette (1992) that the alligator oviduct isan intermediate form evolutionarily between birdsand ancestral oviparous amniotes.
The magnum of the bird and tube of the alliga-tor are the egg-white protein secreting regions ofthe oviduct, synthesizing albumen (Asmundsonand Baker, 1940; Buhi et al., 1999). Some proteinsincorporated into alligator egg albumin appear tobe different from those of other species (Buhiet al., 1999). The alligator tube, like the avianmagnum, is a highly glandular regions that, dur-ing the active times, contains many large secretorygranules that are uniformly electron-dense. In themost posterior region of this area (tubal-isthmusjunction), alligators possess a sperm storage site,considered the secondary site (Gist et al., 2008).Birds also possess a secondary site; however, it islocated in the posterior infundibulum. The primarysite for sperm storage in both alligators and birdsis the uterovaginal region, also called the shellgland-vaginal region in birds (Solomon, 1983;Baskt et al., 1994).
Both birds and alligators store sperm in the mostposterior portions of regions possessing tubularglands. In turtles, sperm are also stored in themost posterior portion of the albumen-secretingregion. In several species of chelonians, however,uterine gland sperm storage has also been identi-fied (Gist and Congdon, 1998). The sperm stored inthe tubal-isthmus junction are likely the first toexit the glands and are responsible for fertilizingthe descending ova. This is supported by the largenumbers of sperm present before ovulation followedby scant amounts later. In birds, the infundibulumis the site of fertilization (Bobr et al., 1964), andthis is probably true for alligators as well.
High amounts of protein (BB-positive material)were expected in the tube during the reproduc-tively active months because albumen is one of themain egg yolk components (Buhi et al., 1999). Thesecretions of tubular glands stain positively forproteins, but the epithelial lining of the tube doesnot, staining instead for neutral and acidic carbo-hydrates. The secretory products of the lining may
be serving as energy reserves at this time as wellas providing other egg yolk components, ratherthan egg protein production.
Unlike other reptiles where the egg shell fibersand the calcareous outer shell are secreted simul-taneously throughout the entire uterus (Palmerand Guillette, 1988; Girling et al., 1998), in bothbirds and crocodilians the uterus is divided intoseparate regions for egg shell formation (Solomonet al., 1975; Palmer and Guillette, 1992). The isth-mus and shell gland pouch of the avian oviductare similar to the uterus of the alligator in whichthey secrete the egg shell fibers and the calcareousouter covering (Solomon et al., 1975). In the ante-rior portions of the alligator uterus, the egg shellfibers are produced, whereas calcium is depositedin the posterior regions (Palmer and Guillette,1992). The calcareous outer egg shell is secreted inthe shell gland of the bird oviduct (Solomon et al.,1975).
The lack of secretory product found in the tubu-lar glands of the posterior uterus of the alligatormay be due in part to the limited histochemicalprocedures performed. This study tested only thepresence of neutral carbohydrates (PAS), acidicmucoids (AB), and proteins (BB) and did not testfor the presence of calcium secretions. In the birdoviduct, lipid secretion also occurs in the homolo-gous egg shell gland (Solomon et al., 1975). We didnot test for lipids, but lipid droplets were notobserved during ultrastructural examination ofthis region.
The vaginae of birds and alligators are both se-cretory regions that lack tubular glands. The thickmusculature in the posterior end of the oviduct(uterus and vagina) aids in the deposition of eggs(Jones and Guillette, 1982). The occurrence of lyso-somes is interesting, as these organelles are impor-tant for microbial defense, and the vagina isexposed to any bacteria, etc., that enter the cloaca.Buhi et al. (1999) suggested that the alligatoroviduct is part of the secretory immune system.
The only comprehensive study on oviductal mor-phology of another alligatorid was done by Perezand Pinilla (2002) on the tropical species Caimancrocodilus fuscus. In this study, they distinguishseven separate regions similar to Palmer and Guil-lette (1992): anterior infundibulum, posterior in-fundibulum, tube, isthmus, anterior uterus, poste-rior uterus, and vagina. This study does not recog-nize the posterior infundibulum as a distinctregion; rather, it is designated as the junctionbetween the infundibulum and the tube.
This study has found noticeable differencesbetween Alligator mississippiensis and Caimancrocodilus fuscus. Perez and Pinilla (2002)reported that the tubal glands contain abundanteosinophilic secretory granules, which we observedin A. mississippiensis, but they found no PAS-posi-tive and AB-positive reactions. In our study, the
ALLIGATOR OVIDUCT 711
Journal of Morphology

tubal glands in reproductively active females werehighly PAS-positive, and the epithelial lining ofthe tube was AB-positive as well. Also, Perez andPinilla (2002) stated that sperm storage is occur-ring in the vaginal folds of the anterior vagina ofC. c. fuscus, whereas alligators store sperm in thetubal-isthmus junction and the utero-vaginal junc-tion regions of the oviduct. Perez and Pinilla(2002) only found sperm in a female collected earlyin the mating season, which suggests that perhapsthe female had recently mated, and the sperm didnot have a chance to migrate into the storageregions. It is also quite possible that because theareas of storage are very short regions, Perez andPinilla (2002) may not have sampled the tissues ofthe uterovaginal and tubal-isthmus junctionswhere sperm may have been residing. Furtherstudies are needed to determine if C. c. fuscus isactually storing sperm among the vaginal folds.
We collected numerous females that were not inreproductive condition. The first year of specimencollection (2006) was preceded by hurricanesKatrina and Rita, which were then followed by aseveral month drought at the Rockefeller WildlifeRefuge collection site. The drought caused highsalinities in the marshes of the refuge, having dou-ble the concentration of sea water. Unfortunately,this drought drastically affected the alligator popu-lation and the reproductive season of 2006 causinga 100% decrease in the number of nests found onthe refuge (zero were found as opposed to severalhundred in normal years). Statewide, the numberof nests dropped to about half of that from the pre-vious year (41,392 in 2005 and 20,387 in 2006).Many of the alligators caught, which were the cor-rect size of a reproductively active female, werereleased due to their overall poor body condition.
Reproductive characteristics can be used as evi-dence to support phylogenetic hypotheses (Palmerand Guillette, 1992; Sever and Hamlett, 2002).Our data support the relationship between crocodi-lians and birds, as well as some similarities withturtles. The structure of the bird and alligator ovi-ducts, with regard to shell formation, shows dis-tinct variation from that of other reptiles. Furtherstudies on crocodilian oviductal morphology areneeded to make a proper comparison across theentire group and to determine if there are majordifferences between the more temperate and tropi-cal species of crocodilians.
ACKNOWLEDGMENTS
The authors like to thank Dr. Dan Gist for help-ful reviews and comments throughout the entiretyof this study and for providing photographs forFigure 1. They also like to extend thanks to everyemployee at Rockefeller Wildlife Refuge whohelped with collection of alligators, especiallyPhillip Trosclair.
LITERATURE CITED
Asmundson VS, Baker GA. 1940. Percentage shell as a functionof shell thickness, egg volume, and egg weight. Poult Sci19:227–232.
Bakst MR, Wishart G, Brillard J. 1994. Oviductal sperm selec-tion, transport, and storage in poultry. Poult Sci Rev 5:117–143.
Bobr LW, Lorenz FW, Ogasawara FX. 1964. Distribution ofspermatozoa in the oviduct and fertility in domestic birds. I.Residence sites of spermatozoa in fowl oviducts. J ReprodFertil 8:39–47.
Brochu CA. 2001. Progress and future directions in archosaurphylogenetics. J Paleont 75:1195–1201.
Buhi WC, Alvarez IM, Binelli M, Walworth ES, Guillete LJ Jr.1999. Identification and characterization of proteins synthe-sized de novo and secreted by the reproductive tract of theAmerican alligator, Alligator mississippiensis. J Reprod Fertil115:201–213.
Chang BSW, Jonsson K, Kazmi MA, Donoghue MJ, Sakmar TP.2002. Recreating a functional ancestral archosaur visualpigment. Mol Biol Evol 19:1483–1489.
Conant R, Collins JT. 1998. A Field Guide to Reptiles andAmphibians, Eastern and Central North America. New York:Houghton Mifflin Company. 403 p.
Florida Fish and Wildlife Conservation Commission. 2007.Available at: http://myfwc.com
Gauthier J. 1984. A Cladistic Analysis of the Higher SystematicCategories of Diapsida, Ph.D. Dissertation; University of Cali-fornia at Berkeley, 564 p.
Gauthier J. 1986. Saurischian monophyly and the origin ofbirds. Mem Cal Acad Sci 8:1–55.
Gauthier J, Kluge AG, Rowe T. 1988. Amniote phylogeny andthe importance of fossils. Cladistics 1988:105–209.
Girling JE, Cree A, Guillette LJ. 1998. Oviductal structure infour species of gekkonid lizards differing in Gist partiy modeand eggshell structure. Reprod Fertil Dev 10:139–154.
Gist DH, Congdon JD. 1998. Oviductal sperm storage as areproductive tactic of turtles. J Exp Zool 282:526–534.
Gist DH, Bagwill AL, Lance VA, Sever DM, Elsey RM. 2008.Sperm storage in the oviduct of the American alligator. J ExpZool 309A:581–587.
Guillette LJ Jr, Woodward AR, Crain DA, Masson GR, PalmerBD, Cox MC, You-Xiang Q, Orlando EF. 1997. The reproduc-tive cycle of the female American alligator (Alligator missis-sippiensis). Gen Comp Endocrinol 108:87–101.
Hutchinson JR, Gatesy SM. 2000. Adductors, abductors, andthe evolution of archosaur locomotion. Paleobiology 26:734–751.
Jacobsen T, Kushlan JA. 1986. Alligator nest flooding in thesouthern everglades: Methodology for management. Proceed-ings of 7th Working Meeting of the IUCN/SSC Crocodile Spe-cialist Group, Caracas, Venezuela, pp 153–166.
Joanen T, McNease L. 1980. Reproductive biology of the Ameri-can alligator in southwest Louisiana. Reprod Biol Dis CaptiveRep 1:153–159.
Joanen T, McNease L. 1989. Ecology and physiology of nestingand early development of the American alligator. Am Zool29:987–998.
Jones RE, Guillette LJ. 1982. Hormonal control of ovipositionand parturition in lizards. Herpetologica 38:80–93.
Klause SE. 1984. Reproductive Characteristics of the AmericanAlligator (Alligator mississippiensis) in North Carolina, Mas-ter of Science Thesis; North Carolina State University,Raleigh, North Carolina.
Lance VA. 1989. Reproductive cycle of the American Alligator.Am Zool 29:999–1018.
Louisiana Department of Wildlife and Fisheries. 2007. Availableat: http://www.wlf.louisiana.gov.
Palmer BD, Guillette LJ. 1988. Histology and functional mor-phology of the female reproductive tract of the tortoise.Gopherus polyphemus. Am J Anat 183:200–211.
712 A. BAGWILL ET AL.
Journal of Morphology

Palmer BD, Guillette LJ. 1992. Alligators provide evidencefor an archosaurian mode of oviparity. Biol Reprod 46:39–47.
Perez GR, Pinilla MPR. 2002. El tracto reproductivo de la hem-bra en Crocodyla: estudio histologico y ultraestructural deCaiman crocodilus fuscus (Crocodylia: Alligatoridae). ActaBiologica Columbiana 7:3–19.
Rasskin-Gutman D, Buscalioni AD. 2001. Theoretical morphol-ogy of the archosaur (Retilia: Diapsida) pelvic girdle. Paleobi-ology 27:59–78.
Sato T, Cheng Y, Wu X, Zelenitsky DK, Hsiao Y. 2005. A pair ofshelled eggs inside a female dinosaur. Science 308:375.
Sever DM, Hamlett WC. 2002. Female sperm storage in rep-tiles. J Exp Zool 292:187–199.
Solomon SE. 1983. Oviduct. In: Freeman BM, editor. Physiologyand Biochemistry of the Domestic Fowl. London: AcademicPress. pp 379–419.
Solomon SE, Fryer JR, Baird T. 1975. The ultrastructural localiza-tion of calcium in the avian shell gland. JMicros 105:215–222.
Uribe MCA, Guillette LJ. 2000. Oogenesis and ovarian histologyof the American alligator Alligator mississippiensis.. J Mor-phol 245:225–240.
Varricchio DJ, Jackson F, Borkowski JJ, Horner JR. 1997. Nestand egg clutches of the dinosaur Troodon formosus and theevolution of avian reproductive traits. Nature 385:247–250.
Wilkinson PM. 1983. Nesting ecology of the American alligatorin coastal South Carolina. Study completion report August1978–September 1983. Columbia: South Carolina Wildlife andMarine Resources Department. 114 p.
ALLIGATOR OVIDUCT 713
Journal of Morphology





























