Embed Size (px)
Citation preview

2666 C. Peters et al. Eur. J . Immunol. 1997.27: 2666-2672
Christopher Peters', Masaya Kawakami*, Marcus Kau13, Thomas I1gl, Peter Overath' and Toni Aebischer'
' Max-Planck-Institut fur Biologie, Abteilung Membranbiochemie, Tubingen, Germany ' Basic Research Center, Kitasato
Institute, Tokyo, Japan Institut fur Medizinische Mikrobiologie und Hygiene, Johannes Gutenberg Universitat, Mainz, Germany
Secreted proteophosphoglycan of Leishmania mexicana amastigotes activates complement by triggering the mannan binding lectin pathway
Cutaneous lesions induced by infection of mice with the protozoan parasite, Leishmania mexicana, contain abundant amounts of a high molecular mass pro- teophosphoglycan (PPG), which is secreted by the amastigote stage residing in phagolysosomes of macrophages and can then be released into the tissue upon rupture of the infected cells. Amastigote PPG forms sausage-shaped but soluble particles and belongs to a novel class of serine-rich proteins that are extensively 0-glycosylated by phosphooligosaccharides capped by mannooligosaccharides. The purified molecule is shown here to efficiently activate complement (C) and deplete hemolytic activity of normal serum and may prevent the opsonization of L. mexicana amastigotes. Complement activation is Ca2+ dependent but does not depend on antibodies or the complement component C1. PPG binds to serum mannan binding protein (MBP), thus activating the MBP-associated serine protease, P100. Subsequently, the C cascade is triggered through C4 lead- ing to covalent modification probably of carbohydrate hydroxyls of PPG by C3 fragments. Thus, PPG is able to activate C via the mannan binding lectin path- way which is unusual for secreted, soluble products of microbial origin. The proteophosphoglycan-induced complement activation is postulated to con- tribute to the lesion development and pathology caused by the parasite.
1 Introduction
Leishmania are protozoan parasites with a digenetic life cycle. The flagellated, extracellular promastigotes reside in the gut of the sandfly vector and are released into the wound of the vertebrate host during the insect's blood meal. Once in the skin, the promastigotes are phagocy- tosed by macrophages and transform into amastigotes in phagolysosomes, termed parasitophorous vacuoles. After replication in the vacuole, the amastigotes are released by rupture of the macrophage and can then enter new host cells [l]. Exposure to humoral defense systems of the host is therefore limited to two short phases during the para- site's life cycle: the time between injection of promastigo- tes and entry into primary host cells and the time needed for amastigotes to reinfect secondary host cells.
In vertebrates, immediate humoral defense is afforded by the complement (C) system [2,3]. Activation of C, i.e. the formation of the C3 convertase, can occur via three path- ways: 1) the classical pathway (CCP), in which antibody- dependent activation of C1 activates C4 and leads to the formation of the C4b2a complex as the initial C3 conver- tase; 2) the mannan binding (MB) lectin pathway, in which
[I 171181
Received May 28, 1997; in revised form July 2.5, 1997; accepted July 28, 1997.
Correspondence: Toni Aebischer, Max-Planck-Institut fur Biolo- gie, Abteilung Membranbiochemie, Corrensstrasse 38, D-72076 Tubingen, Germany Fax: +49-7071-62971; e-mail: [email protected]
Abbreviations: PPG: Proteophosphoglycan LPG: Lipophos- phoglycan MBP: Mannan binding protein P1W MBP- associated serine protease GVB: Gelatin-containing veronal buffer EA: Antibody-coated sheep erythrocytes
Key words: Mannan binding protein / Glycoconjugate
specific carbohydrate binding proteins such as the man- nan-binding protein (MBP) associate with CCactivating proteases, e.g. the MBP-associated serine proteases in man [4, 51 or the protease PlOO in the mouse [6, 71. Bind- ing of these complexes by polyvalent ligands activates these proteases and this leads to the formation of the C4b2a complex without the participation of C1. 3) The alternative pathway (ACP), in which spontaneously hydro- lyzed C3 (C3H20) is the starting convertase. C activation by the first and second but not the third pathway is depen- dent on Ca" .
Interaction of Leishmania with C has been extensively studied (for review see [S-111). Promastigotes of all Leish- mania species tested, including L. major and L. mexicana, activate C. Both Ca2' -dependent and -independent path- ways of C activation have been described for promasti- gotes [12-141 and two major surface components, lipo- phosphoglycan (LPG) and the surface protease, gp63, were shown to activate C and to be the major C3b acceptor molecules [ 14-16]. In vitro, amastigotes of either species also activate C ([ 171 and C. P., unpublished results). In the case of L. major, C3 fragments were detected on the sur- face of amastigotes derived from infected mice indicating that C activation by parasites released by host cell lysis appears to occur in vivo [l8, 191. In contrast, C3 fragments were undetectable on lesion-derived L. mexicana amasti- gotes [20]. This observation suggested that in lesions formed by L. mexicana but not by L. major C fixation may somehow be prevented.
Amastigotes of L. mexicana residing in the parasitopho- rous vacuole of macrophages secrete a high molecular mass proteophosphoglycan (PPG) [21], which appears to cause the characteristic and species-specific enlargement of the vacuole [22]. Ultrastructurally, PPG purified from lesions has been shown to represent irregular sausage- shaped particles with dimensions up to 70 x 15 nm [22].
00 14-2980/97/ 1 0 10-2666$17. SO + . SO/O 0 WILEY-VCH Verlag GmbH, D-69451 Weinheim, 1997

Eur. J . lrnrnunol. 1997.27: 2666-2672 Secreted L . rnexicana proteophosphoglycan: a complement activator 2667
Chemically, amastigote PPG belongs to a novel class of previously described serine-rich proteins secreted by pro- mastigotes that are extensively 0-glycosylated by phos- pho-oligosaccharides (repeat units P04-6Galpl-3Manal- and PO,-6[Glc~1-3]Gal~1-4Manal- in the case of L. mexi- cana) capped by mannooligosaccharides ([Manal-2],. sMan-) [23-251. A so far unique feature of amastigote PPG is the presence of several unusual branched phospho- oligosaccharides (T. Ilg, manuscript in preparation), which may be responsible for the compact structure of the parti- cle. In culture and, by inference, in lesions, PPG can be released from infected macrophages by exocytosis via vesi- cular traffick from the membrane of the parasitophorous vacuole to the host cell plasma membrane [21, 261 or by rupture of vacuole plus host cell. Notably, attempts to purify a similar PPG from lesions of L. major-infected mice were unsuccessful (T. Ilg, unpublished results).
Since PPG formation is a characteristic feature of L. mexi- cana amastigotes, we reasoned that this molecule may be a C activator. Depletion of C in lesions may explain why lesion-derived amastigotes are not opsonized with C3. It will be shown in this study that PPG, a soluble yet micro- particulate molecule, is indeed a very effective C activator by the MB lectin pathway.
2 Materials and methods
2.1 Mice and parasites
CBNJ mice were purchased from Charles River, Sulzfeld, Germany, and maintained in a conventional animal facil- ity. L. mexicana mexicana (strain MNYC/BZ/62/M379, obtained originally from James Alexander, Glasgow) pro- mastigotes were used to infect mice. Amastigotes isolated from lesions were grown in short-term culture in Schnei- der's Drosophila medium (pH 5.5; Serva, Heidelberg, Germany) supplemented with 20 YO heat-inactivated FCS as described in [20].
2.2 Antibodies and reagents
The mouse mAb AP3 and LT22 [27] directed against man- nooligosaccharide cap structures and glucosylated phos- phosaccharides of LPG and related glycoconjugates such as PPG, respectively, and the polyclonal rabbit anti-mouse MBP-P100 have been described [28]. The anti-mouse IgG antiserum and protein G beads were from Pharmacia, Freiburg, Germany, goat anti-mouse C3 antiserum from Cappel Laboratories, Eppelheim, Germany, purified rab- bit anti-sheep erythrocyte antibodies and human C4 from Sigma, Deisenhofen, Germany, and sheep erythrocytes from Difco, Augsburg, Germany. C4-deficient, MBP- P100-depleted guinea pig serum was prepared as in [28]. Mouse sera were prepared by retroorbital bleeding, co- agulation on ice followed by centrifugation. Immunoglobulin-deficient sera from RAG knockout mice were a kind gift of Dr. F. Brombacher, Max-Planck-Institut fur Immunbiologie, Freiburg, Germany. To deplete C1, mouse serum was passed over a 2-ml mouse IgG-agarose column (Sigma) at a flow rate of 2 mlh and eluted with Veronal-buffered saline containing 0.1 YO gelatin (GVB) supplemented with 2 mM CaCI, and 2 mM MgClz (Mg-
Ca-GVB) at 4 "C. Thereafter, the serum was unable to lyse antibody-coated sheep erythrocytes (EA).
2.3 Purification of PPG
Dorsal lesions of L. mexicana-infected CBA/J mice were processed and the PPG was purified as described [21]. Briefly, the supernatant of a tissue homogenate was frac- tionated first over a DE52 cellulose and then over an octyl- Sepharose column. The concentrated PPG-containing fractions were deproteinized by extraction with phenol. The aequous phase was then passed over a Superose-6 column (Pharmacia, Freiburg, Germany) instead of Superose-12 as originally described [21] and the PPG- containing fractions were pooled, concentrated and treated with hyaluronidase and benzonnuclease (both from Sigma). The mixture was again extracted with phenol and re-run over the Superose-6 column. Quantitation and purity control of Centricon-30 (Amicon, Witten, Germany)-concentrated material were performed as described [21] by determining the phosphorus content and by SDS-PAGE. PPG preparations used in this study did not contain any protein contaminant detectable by SDS- PAGE and the phenol treatment assured the inactivation of any contaminating proteases. Finally, PPG was recon- centrated and resuspended in PBS at 3 mg/ml.
2.4 Rabbit erythrocyte lysis assay
Sera (100 pI) supplemented with the indicated amounts of PPG diluted in GVB or controls without PPG were incu- bated for different lengths of time at 37°C. Thereafter, 10 pl of rabbit erythrocytes (1.5 x 109/ml) were added and the mixture was incubated at 37°C for 30 min. Samples were diluted with GVB containing 10 mM EDTA (GVB- EDTA) to a final volume of 1 ml, centrifuged and the absorbance at 530 nm of the supernatant was measured.
2.5 Estimation of MBP-P100 binding by ELISA and C4 consumption test
PPG was adsorbed overnight at 4°C from a solution containing 10 pq PPG/ml in PBS onto 96-well Immuno- Plate Maxi-Sorb (NUNC, Wiesbaden, Germany). Plates were blocked with PBS/2% BSA and washed twice with PBS. Purified MBP-P100 at the indicated amounts was added in 100 p1 Mg-Ca-GVB. Plates were incubated for 2 h at 4°C and washed five times with Mg-Ca-GVB. MBP- PlOO bound was detected by adding anti-MBP-P100 antise- rum diluted 1 : 500, followed by goat anti-rabbit antibodies coupled to alkaline phosphatase (Dianova, Hamburg, Germany).
C4 consumption by PPG-bound MBP-P100 was measured by the addition of 50 pl Mg-Ca-GVB containing 3.5 U human C4. Plates were incubated for 40 min at 30°C and then chilled for 15 min. C4 consumption was assessed by adding 100 pl of EA (1.5 x lo8 cells/ml in Mg-Ca-GVB) and 100 1.11 Mg-Ca-GVB, 1 % CCdeficient, MBP-P100- depleted guinea pig serum for 1 h at 37°C. The reaction mixtures were transferred to Eppendorf tubes containing
2668 C. Peters et al.
750 p1 GVB-EDTA and the absorption at 415 nm of the supernatants was determined after centrifugation.
Eur. J. Immunol. 1997.27: 2666-2672
2.6 Estimation of monosaccharide inhibition of C activation
Serum from RAG knockout mice was diluted to 50% in cold Mg-Ca-GVB and then supplemented with mannose or mannitol to a final concentration of 10 mM. The mix- ture was incubated for 1 h at 4"C, then PPG was added to different final concentrations and the samples were brought to 37 "C for 30 min. C activity was then measured in the rabbit erythrocyte lysis assay.
2.7 Immunoprecipitations and immunoblots
Normal mouse serum (100 pl) was diluted with 200 pl Mg- Ca-GVB containing 10 pg PPG and incubated for 4 h on ice. LT22 mAb bound to protein G beads was added and the mixture was rotated end-over-end for 1 h at 4°C. Beads were washed five times in Mg-Ca-GVB and then incubated in 200 pl GVB-EDTA. Eluted material was separated by SDS-PAGE under reducing conditions and blotted onto Immobilon membranes (Millipore, Eschborn, Germany). Membranes were blocked in PBS/ 5 % milk powder and incubated with anti-MBP-P100 anti- serum. Bound antibodies were detected with alkaline phosphatase-conjugated sheep anti-rabbit antibodies diluted 1: 2000 in PBS/5 % milk powder and visualized with 0.5 mM 5-bromo-4-chloro-3-indolylphosphate, 0.5 mM nitrobluetetrazolium in 1 M diethanolamine/HCl, 1 mM MgClz, pH 9.8.
3 Results
3.1 PPG activates complement
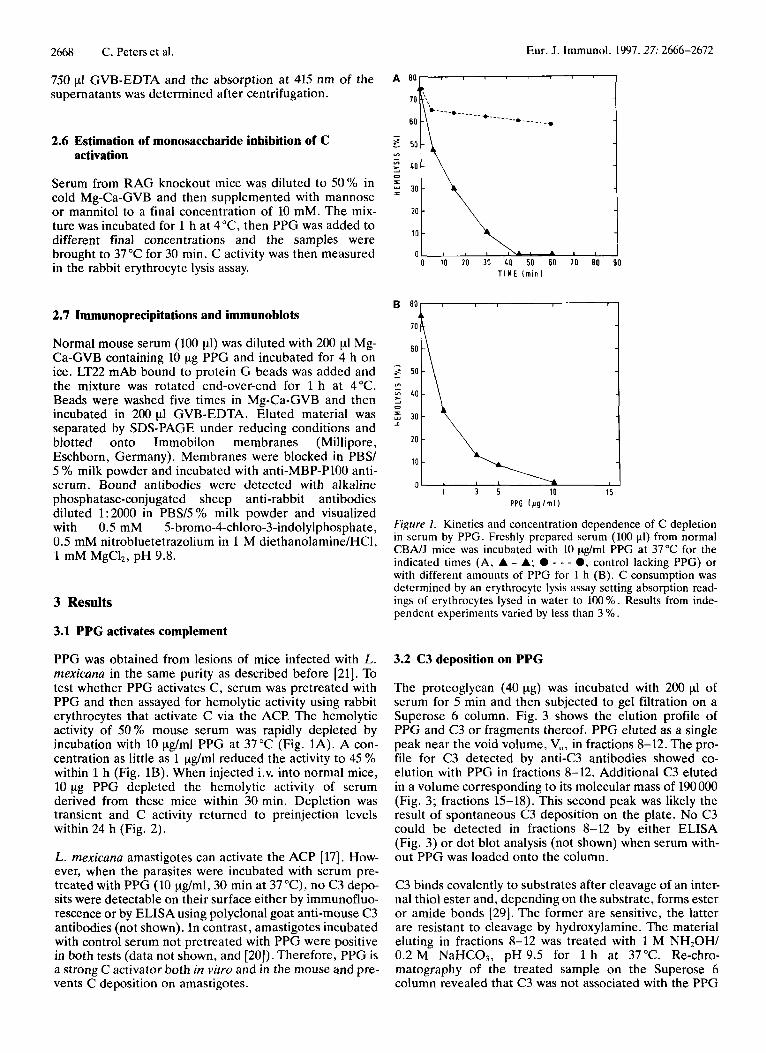
PPG was obtained from lesions of mice infected with L. rnexicuna in the same purity as described before [21]. To test whether PPG activates C, serum was pretreated with PPG and then assayed for hemolytic activity using rabbit erythrocytes that activate C via the ACP. The hemolytic activity of 50% mouse serum was rapidly depleted by incubation with 10 pg/ml PPG at 37°C (Fig. 1A). A con- centration as little as 1 pg/ml reduced the activity to 45 YO within 1 h (Fig. 1B). When injected i.v. into normal mice, 10 pg PPG depleted the hemolytic activity of serum derived from these mice within 30 min. Depletion was transient and C activity returned to preinjection levels within 24 h (Fig. 2 ) .
L. rnexicuna amastigotes can activate the ACP [17]. How- ever, when the parasites were incubated with serum pre- treated with PPG (10 pg/ml, 30 min at 37 "C), no C3 depo- sits were detectable on their surface either by immunofluo- rescence or by ELISA using polyclonal goat anti-mouse C3 antibodies (not shown). In contrast, amastigotes incubated with control serum not pretreated with PPG were positive in both tests (data not shown, and [20]). Therefore, PPG is a strong C activator both in vitro and in the mouse and pre- vents C deposition on amastigotes.
TIME ( m i n )
70 " i i
PPG I p g l m l )
Figure I. Kinetics and concentration dependence of C depletion in serum by PPG. Freshly prepared serum (100 P I ) from normal CBNJ mice was incubated with 10 pg/ml PPG at 37°C for the indicated times (A, A - A: 0 - - - 0, control lacking PPG) or with different amounts of PPG for 1 h (B). C consumption was determined by an erythrocyte lysis assay setting absorption read- ings of erythrocytes lysed in water to 100 %. Results from inde- pendent experiments varied by less than 3 YO.
3.2 C3 deposition on PPG
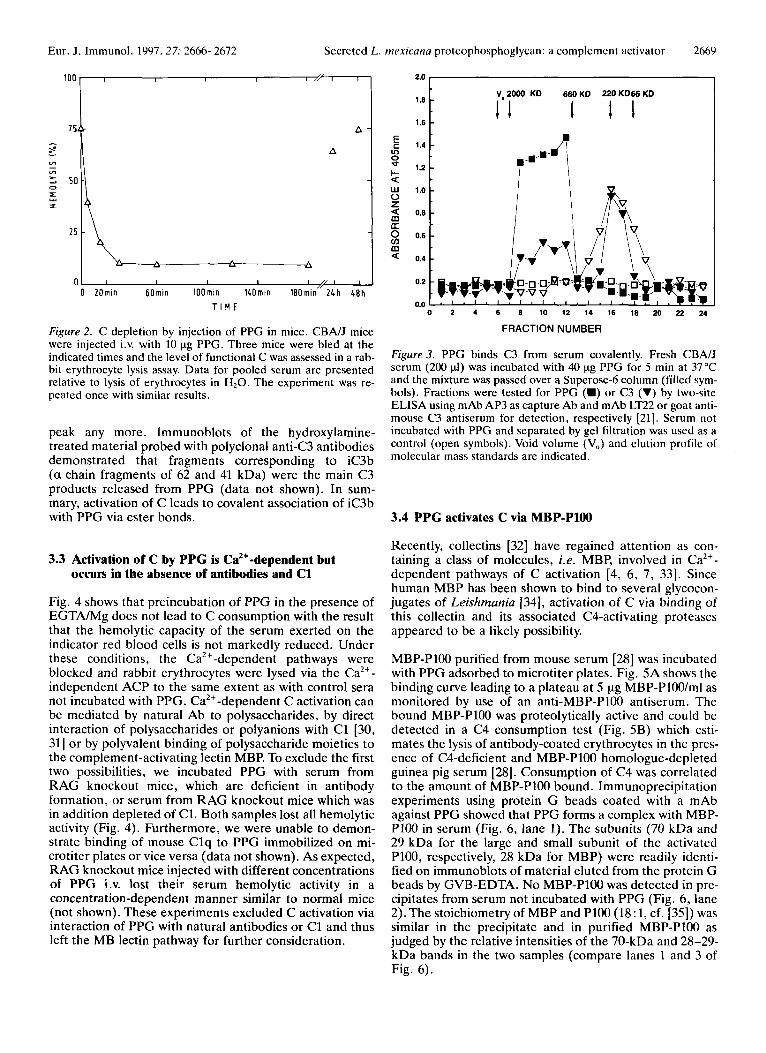
The proteoglycan (40 pg) was incubated with 200 pI of serum for 5 min and then subjected to gel filtration on a Superose 6 column. Fig. 3 shows the elution profile of PPG and C3 or fragments thereof. PPG eluted as a single peak near the void volume, V,, in fractions 8-12. The pro- file for C3 detected by anti-C3 antibodies showed co- elution with PPG in fractions 8-12. Additional C3 eluted in a volume corresponding to its molecular mass of 190 000 (Fig. 3; fractions 15-18). This second peak was likely the result of spontaneous C3 deposition on the plate. No C3 could be detected in fractions 8-12 by either ELISA (Fig. 3) or dot blot analysis (not shown) when serum with- out PPG was loaded onto the column.
C3 binds covalently to substrates after cleavage of an inter- nal thiol ester and, depending on the substrate, forms ester or amide bonds [29]. The former are sensitive, the latter are resistant to cleavage by hydroxylamine. The material eluting in fractions 8-12 was treated with 1 M NH20H/ 0.2 M NaHC03, pH 9.5 for 1 h at 37°C. Re-chro- matography of the treated sample on the Superose 6 column revealed that C3 was not associated with the PPG
Eur. J . Immunol. 1997.27: 2666-2672 Secreted L
I I I I
l o o l I
A
A
01 I I 1 I
0 2Omin 6 0 m i n 1OOmin 14Omin 180min’2Lh 48h T I M E
Figure 2. C depletion by injection of PPG in mice. CBNJ mice were injected i.v. with 10 pg PPG. Three mice were bled at the indicated times and the level of functional C was assessed in a rab- bit erythrocyte lysis assay. Data for pooled serum are presented relative to lysis of erythrocytes in H 2 0 . The experiment was re- peated once with similar results.
peak any more. Immunoblots of the hydroxylamine- treated material probed with polyclonal anti-C3 antibodies demonstrated that fragments corresponding to iC3b ( a chain fragments of 62 and 41 kDa) were the main C3 products released from PPG (data not shown). In sum- mary, activation of C leads to covalent association of iC3b with PPG via ester bonds.
3.3 Activation of C by PPG is Ca*+-dependent but occurs in the absence of antibodies and C1
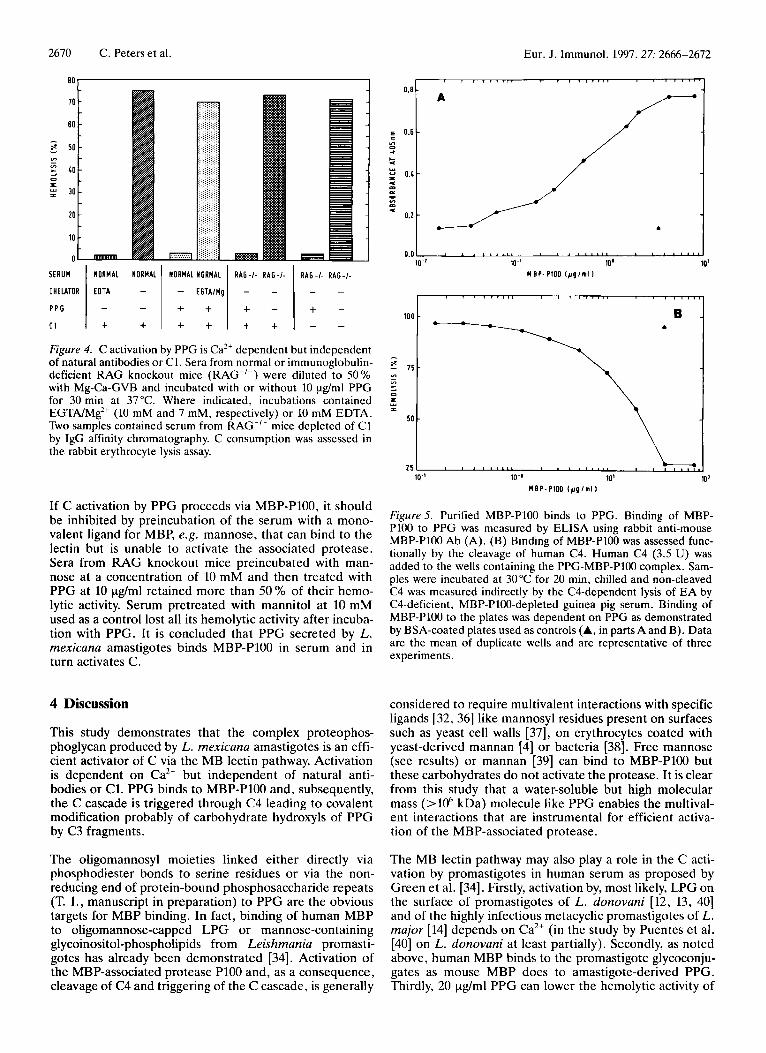
Fig. 4 shows that preincubation of PPG in the presence of EGTNMg does not lead to C consumption with the result that the hemolytic capacity of the serum exerted on the indicator red blood cells is not markedly reduced. Under these conditions, the Ca’+-dependent pathways were blocked and rabbit erythrocytes were lysed via the Ca2+- independent ACP to the same extent as with control sera not incubated with PPG. Ca2+-dependent C activation can be mediated by natural Ab to polysaccharides, by direct interaction of polysaccharides or polyanions with C1 [30, 311 or by polyvalent binding of polysaccharide moieties to the complement-activating lectin MBP. To exclude the first two possibilities, we incubated PPG with serum from RAG knockout mice, which are deficient in antibody formation, or serum from RAG knockout mice which was in addition depleted of C1. Both samples lost all hemolytic activity (Fig. 4). Furthermore, we were unable to demon- strate binding of mouse Clq to PPG immobilized on mi- crotiter plates or vice versa (data not shown). As expected, RAG knockout mice injected with different concentrations of PPG i.v. lost their serum hemolytic activity in a concentration-dependent manner similar to normal mice (not shown). These experiments excluded C activation via interaction of PPG with natural antibodies or C1 and thus left the MB lectin pathway for further consideration.
rnrxicanu proteophosphoglycan: a complement activator 2669
2.0 I Vo2000 KD 660KD 220KDbbKD
1.6 I I 1
1.2 1.41 8
a w 1.0 0 z
0.8
0 0.6 v) 03 a 0.4
0.2
0.0
I
0 2 4 6 8 10 12 14 16 18 20 22 24
FRACTION NUMBER
Figure 3. PPG binds C3 from serum covalently. Fresh CBA/J serum (200 pi) was incubated with 40 pg PPG for 5 min at 37°C and the mixture was passed over a Superose-6 column (filled sym- bols). Fractions were tested for PPG (W) or C3 (V) by two-site ELISA using mAb AP3 as capture Ab and mAb LT22 or goat anti- mouse C3 antiserum for detection, respectively [21]. Serum not incubated with PPG and separated by gel filtration was used as a control (open symbols). Void volume (VJ and elution profile of molecular mass standards are indicated.
3.4 PPG activates C via MBP-P100
Recently, collectins [32] have regained attention as con- taining a class of molecules, i .e. MBP, involved in Ca2+- dependent pathways of C activation [4, 6, 7, 331. Since human MBP has been shown to bind to several glycocon- jugates of Leishmania [34], activation of C via binding of this collectin and its associated CCactivating proteases appeared to be a likely possibility.
MBP-P100 purified from mouse serum [28] was incubated with PPG adsorbed to microtiter plates. Fig. 5A shows the binding curve leading to a plateau at 5 yg MBP-PlOO/ml as monitored by use of an anti-MBP-P100 antiserum. The bound MBP-P100 was proteolytically active and could be detected in a C4 consumption test (Fig. 5B) which esti- mates the lysis of antibody-coated erythrocytes in the pres- ence of C4-deficient and MBP-P100 homologue-depleted guinea pig serum [28]. Consumption of C4 was correlated to the amount of MBP-P100 bound. Immunoprecipitation experiments using protein G beads coated with a mAb against PPG showed that PPG forms a complex with MBP- PlOO in serum (Fig. 6, lane 1). The subunits (70 kDa and 29 kDa for the large and small subunit of the activated P100, respectively, 28 kDa for MBP) were readily identi- fied on immunoblots of material eluted from the protein G beads by GVB-EDTA. No MBP-P100 was detected in pre- cipitates from serum not incubated with PPG (Fig. 6, lane 2). The stoichiometry of MBP and PlOO (18 : 1, cf. [35]) was similar in the precipitate and in purified MBP-P100 as judged by the relative intensities of the 70-kDa and 28-29- kDa bands in the two samples (compare lanes 1 and 3 of Fig. 6).
2670 C. Peters et al.
SERUM
CHEIATOR
P P G
C I
Eur. J . Immunol. 1997.27: 2666-2672
NORMAL NORMAL NORMAL NORMAL RAG- / - RAG- / - RAG- / - R A G - / .
- - E O T A - - E G T A / H g - -
i i i - t - - -
- - t i + i i i
I I " " I I
If C activation by PPG proceeds via MBP-P100, it should be inhibited by preincubation of the serum with a mono- valent ligand for MBP, e.g. mannose, that can bind to the lectin but is unable to activate the associated protease. Sera from RAG knockout mice preincubated with man- nose at a concentration of 10 mM and then treated with PPG at 10 pg/ml retained more than 50 % of their hemo- lytic activity. Serum pretreated with mannitol at 10 mM used as a control lost all its hemolytic activity after incuba- tion with PPG. It is concluded that PPG secreted by L. mexicana amastigotes binds MBP-P100 in serum and in turn activates C.
4 Discussion
This study demonstrates that the complex proteophos- phoglycan produced by L. mexicana amastigotes is an effi- cient activator of C via the MB lectin pathway. Activation is dependent on Ca2+ but independent of natural anti- bodies or C1. PPG binds to MBP-P100 and, subsequently, the C cascade is triggered through C4 leading to covalent modification probably of carbohydrate hydroxyls of PPG by C3 fragments.
The oligomannosyl moieties linked either directly via phosphodiester bonds to serine residues or via the non- reducing end of protein-bound phosphosaccharide repeats (T. I., manuscript in preparation) to PPG are the obvious targets for MBP binding. In fact, binding of human MBP to oligomannose-capped LPG or mannose-containing glycoinositol-phospholipids from Leishmania promasti- gotes has already been demonstrated [34]. Activation of the MBP-associated protease P100 and, as a consequence, cleavage of C4 and triggering of the C cascade, is generally
0.8 -
E 0.6 - c m 0 3
c 4
Y
0.4 - U p1 e 0 m m U
0.2 -
0.0 10-1 10' 10'
M E P - P 1 0 0 lpglal I
1
lo-' 10' M B P - P I 0 0 ( p g l n l l
107
Figure5. Purified MBP-P100 binds to PPG. Binding of MBP- Pl00 to PPG was measured by ELISA using rabbit anti-mouse MBP-P100 Ab (A). (B) Binding of MBP-P100 was assessed func- tionally by the cleavage of human C4. Human C4 (3.5 U) was added to the wells containing the PPG-MBP-P100 complex. Sarn- ples were incubated at 30°C for 20 min, chilled and non-cleaved C4 was mcasured indirectly by the CCdependent lysis of EA by C4-deficient, MBP-P100-depleted guinea pig serum. Binding of MBP-P100 to the plates was dependent on PPG as demonstrated by BSA-coated plates used as controls (A, in parts A and B). Data are the mean of duplicate wells and are representative of three experiments.
considered to require multivalent interactions with specific ligands [32,36] like mannosyl residues present on surfaces such as yeast cell walls [37], on erythrocytes coated with yeast-derived mannan [4] or bacteria [38]. Free mannose (see results) or mannan [39] can bind to MBP-P100 but these carbohydrates do not activate the protease. It is clear from this study that a water-soluble but high molecular mass (>loh kDa) molecule like PPG enables the multival- ent interactions that are instrumental for efficient activa- tion of the MBP-associated protease.
The MB lectin pathway may also play a role in the C acti- vation by promastigotes in human serum as proposed by Green et al. [34]. Firstly, activation by, most likely, LPG on the surface of promastigotes of L. donovani [12, 13, 401 and of the highly infectious metacyclic promastigotes of L. major [14] depends on Ca2+ (in the study by Puentes et al. [40] on L. donovani at least partially). Secondly, as noted above, human MBP binds to the promastigote glycoconju- gates as mouse MBP does to amastigote-derived PPG. Thirdly, 20 pg/ml PPG can lower the hemolytic activity of

Eur. J. Immunol. 1997.27: 2666-2672 Secreted L. mexicana protcophosphoglycan: a complement activator 2671
1 2 3
Figure 6. Co-immunoprecipitation of MBP-P100 with PPG after incubation with normal serum. PPG was incubated with normal CBNJ serum and then immunoprecipitated with mAb LT22 against PPG. The precipitate was washed and then treated with GVB-EDTA. The eluate was subjected to SDS-PAGE under reducing conditions, transferred to Immobilon membranes and probed with rabbit anti-mouse MBP-P100 antiserum followed by sheep anti-rabbit antibodies conjugated to alkaline phosphatase (lane 1). Precipitates from serum not containing PPG (lane 2) or affinity-purified MBP-P100 (lane 3, [28]) were included as con- trols.
human serum to 56 % of the control within 70 min (C. P., unpublished results).
We propose that PPG depletes C in lesions. The glycocon- jugate is secreted from amastigotes into the parasitopho- rous vacuole and it can then probably leave the infected macrophage by vesicular traffic from the vacuole mem- brane to the plasma membrane [21, 261. Alternatively, PPG is released upon rupture of host cells. As the yield of PPG purified from lesion material is 40-50 pg/g tissue (T. I., unpublished results), the local PPG concentration surrounding ruptured host cells in lesions will be well above the concentration required for C activation (cf. Fig. 1B). A concentration of about 5 pg PPG/ml rapidly depletes C activity in mouse blood (Fig. 2). In addition, L. mexicana amastigotes isolated from lesions do not have detectable C3 surface deposits [20] but do activate C [17, 201 and bind MBP-P100 in vitro (C. P., unpublished results). This implies that MBP and the associated C4 con- vertase are present in lesions. Indeed, studies on rheuma- toid arthritis showed that this complex is present within inflamed tissue [41]. In heavily infected mice, C depletion appears to be restricted to the lesion because sera of these animals display normal hemolytic activity. PPG may leak into the circulation but will continuously be cleared either when opsonized by CR3-positive cells, e.g. macrophages in the lung or spleen, or by direct binding to mannose receptors, e.g. by hepatic epithelial cells [42].
Cutaneous L. mexicana infections are chronic and progres- sive in most mouse strains [l]. PPG-mediated C activation may be important for the pathology of the infection. Firstly and most obviously, inactivation of C may prevent amastigote lysis, however, this parasite stage is inherently relatively resistant to C lysis [17]. Secondly and more rele- vant, C activation by PPG includes the generation of ana- phylactic peptides C3a, C4a and possibly C5a. The release of anaphylatoxins will attract infectable monocytes to the site of infection. It is of note that occasionally, mice died
within 5 min after i.v. injection of PPG showing signs of anaphylactic shock (C. P. and T. A., unpublished results). This predicts that hosts defective in C activation at the level of the C3 convertases will have a markedly retarded lesion development. Thirdly, PPG is opsonized with C3 and fragments thereof. Opsonized PPG bound to COS cells transfected with human CR3 whilst purified PPG bound to mannose receptor-transfected COS cells (C. P., unpublished observation). Therefore, PPG could engage the mannose receptor or other C-type lectin receptors able to bind mannose as well as CR on lesion-infiltrating inflammatory cells and thus modulate their functional properties. CR3 cross-linking, for example, has been shown recently to inhibit IL-12 secretion [43], a key mole- cule in the regulation of the anti- Leishmania immune response [44]. Thus, continuous activation of C by secreted parasite products such as PPG or homologous substances of other Leishmania species could make them important pathogenic factors in leishmaniases. Further definition of the function of the proteophosphoglycan will require clon- ing and deletion of the gene coding for its polypeptide.
We thank Gabriele Winternheimer, Sabine Pauls, Gerda Miiller and Jens Ruoff for expert technical assistance, Franz Perry, Institut fur Medizinische Mikrobiology & Hygiene, Johannes Cutenberg Uni- versitat, Mainz, for providing an a-mClq antibody, Frank Bromba- cher, Max-Planck-lnstitut fur Immunbiologie, Freiburg, for pro- viding the RAG knockout mice and serum and Clare Blackburn, University of Edinburgh, for stimulating comments on the manu- script.
5 References
1 Alexander, J. and Russell, D. G . , A d v . Parasitol. 1992.31: 175. 2 Farries, T. C. and Atkinson, J. P., Immunol. Today 1991. 12:
3 Rother, K. andTill, G. O., The Complement System, Springer-
4 Matsushita, M. and Fujita, T., J. Exp. Med. 1992. 176: 1497. 5 Thiel, S., Vorup-Jensen, T., Stover, C. M., Schwaeble, W.,
Laursen, S. B., Poulsen, K., Willis, A. C., Eggleton, P., Han- sen, s., Holmskov, U., Reid, K. B. M. and Jensenius, J . C., Nature 1997. 386: 506.
6 Ji, Y. H., Fujita, T., Hatsuse, H., Takahashi, A., Matsushita, M. and Kawakami, M., J. Immunol. 1993. 150: 571.
7 Takayama, Y., Takada, F., Takahashi, A. and Kawakami, M., J . Immunol. 1994. 152: 2308.
8 Joiner, K. A. in Englund, P. T. and Sher, A. (Eds.), The Biol- ogy of Parasitism, Alan R. Riss Inc., New York 1988, p. 309.
9 Mosser, D. M., Immunol. Sem. 1994. 60: 99.
295.
Verlag: Berlin, Heidelberg, New York 1988.
10 Fuhrman, S. A. and Joiner, K. A., Exp. Parasitol. 1989. 68: 414.
11 Brittingham, A. and Mosser, D. M., Parasitol. Today 1996. 12: 444.
12 Mosser, D. M., Burke, S. K., Coutavas, E. E., Wedgwood. J . F. and Edelson, P. J., Exp. Parasitol. 1986. 62: 394.
13 Pearson, R. D. and Steigbigel, R. T., J. Immunol. 1986. 135: 2195.
14 Puentes, S. M., Sacks, D. L., Da Silva, R. P. and Joiner, K. A., J. Exp. Med. 1988. 167: 887.
15 Brittingham, A., Morrison, C. J., McMaster, W. R., McGwire, B. S., Chang, K. P. and Mosser, D. M., J. Immunol. 1995.155: 3102.
16 Russell, D. G. , Eur. J. Biochem. 1987. 164: 213. 17 Mosser, D. M., Wedgwood, J. F. and Edelson, P. J., J. Immu-
nol. 1985. 134: 4128. 18 Kelleher, M., Moody, S. F., Mirabile, P., Osborn, A. H.,
Bacic, A. and Handman, E., Infect. Immun. 1995. 63: 43.

2672 C. Peters et a1 Eur. J. Immunol. 1997.27: 2666-2672
19 Guy, R. A. and Belosevic, M., Infect. Immun. 1993. 61: 1553. 20 Peters, C., Aebischer, T., Stierhof, Y. D., Fuchs, M. and Over-
ath, P., J . CelI Sci. 1995. 108: 3715. 21 Ilg, T., Stierhof, Y. D., McConville, M. J. and Overath, P.,
Eur. J . Cell Biol. 1995. 66: 205. 22 Peters, C., Stierhof, Y. D. and Ilg, T., Infect. Immun. 1997. 65:
783. 23 Ilg, T., Overath, P., Ferguson, M. A. J., Rutherford,T., Camp-
bell, D. G. and McConville, M. J. , J. Biol. Chem. 1994. 269: 24073.
24 Wiese, M., Ilg, T., Lottspeich, E and Overath, I?, EMBO J. 1995. 14: 1067.
25 Ilg, T., Stierhof, Y. D., Craik, D., Simpson, R., Handman, E. and Bacic, A., J . Biol. Chem. 1996. 271: 21583.
26 Wolfram, M., Fuchs, M., Wiese, M., Stierhof, Y. D. and Over- ath, P., Eur. J. Immunol. 1996.26: 3153.
27 Ilg, T., Harbecke, D., Wiese, M. and Overath, P., Eur. J. Bio- chem. 1993.217: 603.
28 Ji, Y. H., Matsushita, M., Okada, H., Fujita, T. and Kawa- kami, M., J. Immunol. 1988. 141: 4271.
29 Sim, R. B., Twose, T. M., Paterson, D. S. and Sim, E., Bio- chem. J . 1981. 193: 115.
30 Loos, M. and Colomb, M., Behring Inst. Mitt. 19Y3. 93: 1. 31 Gewurz, H., Ying, S.-C., Jiang, H. and Lint, T. F., Behring
32 Epstein, J., Eichbaum, Q., Sheriff, S. and Ezekowitz, R. A. Inst. Mitt. 1993. 93: 138.
B., Curr. Opin. Immunol. 1996. 8: 29.
33 Takada, F., Takayama, Y., Hatsuse, H. and Kawakami, M., Biochem. Biophys. Res. Commun. 1993. 196: 1003.
34 Green, l? J., Feizi, T., Stoll, M. S . , Thiel, S., Prescott, A. and McConville, M. J . , Mol. Biochem. Parasitol. 1994. 66: 319.
35 Ogata, R. T., Low, P. J. and Kawakami, M., J. Immunol. 1995. 154: 2351.
36 Weis, W. I. and Drickamer, K.,Annu. Rev. Biochem. 1996.65: 441.
37 Lu, C. Y., Thiel, S., Wiedmann, H., Timpl, R. and Reid, K. B., J. Immunol. 1990. 144: 2287.
38 Kawakami, M., Ihara, I . , Ihara, S., Suzuki, A. and Fukui, K., J. Immunol. 1984. 132: 2578.
39 Ohta, M., Okada, M., Yamashima, I. and Kawasaki, T., J. B i d . Chem. 1990. 265: 1980.
40 Puentes, S. M., Dwyer, D. M., Bates, P. A. and Joiner, K. A., J. Immunol. 1989. 143: 3743.
41 Malhotra, R., Womald, M. R., Rudd, P. M., Fischer, P. B., Dwek, R. A. and Sim, R. B., Nature Med. 1995. I : 237.
42 Jansen, R. W., Molema, G., Ching, T. L., Oosting, R., Harms, G., Moolenaar, F., Hardonk, M. J . and Meijer, D. K., J. Biol. Chem. 1991. 266: 3343.
43 Sutterwala, E S., Noel, G. J., Chynes, R. and Mosser, D. M., J. Exp. Med. 1997. 185: 1977.
44 Reiner, S. L. and Locksley, R. M.,Annu. Rev. Immunol. 1995. 13: 151.





























