Embed Size (px)
Citation preview

Selective Changes in HippocampalNeuropeptide Y Neurons Following
Removal of the Cholinergic Septal Inputs
TERESA A. MILNER,1* RONALD G. WILEY,2 OLIVER S. KURUCZ,1
SABRINA R. PRINCE,1 AND JOSEPH P. PIERCE1
1Division of Neurobiology, Department of Neurology and Neuroscience, Cornell UniversityMedical College, New York, New York 10021
2Laboratory of Experimental Neurology, VAMedical Center, Nashville, Tennessee 37212
ABSTRACTThe number and distribution of subpopulations of hilar interneurons containing neuropeptide
Y (NPY), somatostatin (SOM), or g-aminobutyric acid (GABA) immunoreactivities were examinedin the hilus of the dentate gyrus following removal of the cholinergic septal inputs. One, 2, 4, 8, 12,and 24 weeks after intracerebroventricular injections of immunotoxin, consisting of antibody to thelow-affinity nerve growth factor receptor conjugated to saporin (192 IgG-saporin), lesioned ratswereprocessed simultaneously with controls for NPY, SOM, or GABA immunolabeling. Across all timepoints, the number of NPY-labeled neurons was reduced to a statistically significant level (pairedt-test, P 5 0.001) in the injected rats (73% of control values, on average). The decrease in thenumber of NPY-labeled neurons was not limited to any particular subregion rostrally butappeared greater in the central region caudally. The size of NPY-labeled neurons did not differstatistically between control and immunolesioned rats examined at 1, 2, and 24 week timepoints. In contrast, the number of both SOM- and GABA-immunoreactive neurons in injectedrats did not appear to be affected in any consistent manner. Examination of the hilus inadjacent Nissl-stained sections with the optical dissector revealed that although the totalnumber of small nonprincipal cells (5–15 µm in diameter) did not appear affected at the4-week time point, there was a statistically significant (P 5 0.03) reduction across the8–24-week time points (to 80% of control values, on average). Dual-labeling studies onseparate rats showed that a small subpopulation of the NPY- and SOM-labeled neurons,primarily in the infragranular hilus, were colocalized with neurons containing GABAimmunoreactivity (18% and 5%, respectively). These studies demonstrate that removal of thecholinergic septal inputs (1) can cause relatively rapid, selective decreases in the number ofNPY-immunoreactive hippocampal interneurons and (2) appears to lead to the death ofhippocampal interneurons over a longer time course. The changes in NPY immunoreactivityseem to occur in the portion of interneurons that probably does not contain either SOM orGABA immunoreactivity. J. Comp. Neurol. 386:46–59, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: 192 IgG-saporin; somatostatin; dentate gyrus; septohippocampal afferents;
deafferentation
The major subcortical projection to the hippocampalformation arises from the septal complex (i.e., the medialseptal nucleus and the nucleus of the diagonal band;Swanson and Cowan, 1979). A number of studies usingacetylcholinesterase (AChE) histochemistry and immuno-cytochemical localization of the acetylcholine syntheticenzyme, choline acetyltransferase (ChAT), have shownthat septal neurons that project to the hippocampal forma-tion are both cholinergic and noncholinergic (Baisden etal., 1984; Amaral and Kurz, 1985; Wainer et al., 1985;Woolf et al., 1986; Senut et al., 1989). The noncholinergic
neurons contributing to this pathway are primarily g-ami-nobutyric acid (GABA)-ergic, based on the presence of thesynthetic enzyme glutamic acid decarboxylase (GAD;
Grant sponsor: NIMH; Grant number: MH42834; Grant sponsor: NIH;Grant number: 18974; Grant sponsor: Medical Research Service, DVA;Grant sponsor: NIDA; Grant number: DA08259.*Correspondence to: Dr. Teresa A. Milner, Division of Neurobiology,
Cornell University Medical College, 411 East 69th Street, New York, NY10021. E-mail: [email protected] 24 October 1996; Revised 21April 1997; Accepted 22April 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 386:46–59 (1997)
r 1997 WILEY-LISS, INC.

Kohler et al., 1984). Within the hippocampal formation,septal afferent fibers are topographically organized, withmost fibers terminating in the hilar area of the dentategyrus and strata oriens and radiatum of the CA3 region ofthe hippocampus and subiculum; few terminate within themolecular layer of the dentate gyrus, presubiculum, para-subiculum, and entorhinal cortex (Swanson and Cowan,1979; Milner et al., 1983; Gaykema et al., 1990).Recent ultrastructural studies have demonstrated that
septal afferent terminals directly contact a variety ofhippocampal GABAergic nonprincipal cells (interneurons)including subpopulations that contain immunoreactivityfor neuropeptide Y (NPY), somatostatin (SOM), and GABA(Freund and Antal, 1988; Yamano and Luiten, 1989;Frotscher, 1991; Milner and Veznedaroglu, 1993); thesubpopulations of NPY- and SOM-immunoreactive neu-rons are mostly in the central hilus and are extensivelycolocalized (Kohler et al., 1987), whereas the majority ofGABA-immunoreactive neurons are located in the infra-granular hilus (Milner and Bacon, 1989b). These studieshave also demonstrated that the cholinergic and GABAer-gic portions of the septohippocampal pathway have differ-ent patterns of connectivity within the hippocampal forma-tion, especially the dentate gyrus. Specifically, cholinergicseptal afferent terminals contact both granule cells andinterneurons, forming both asymmetric and symmetricsynapses (Clarke, 1985; Frotscher and Leranth, 1986). Anumber of the interneurons contacted by the cholinergicseptal afferents contain NPY and SOM immunoreactivityand are located in the central region of the hilus (Leranthand Frotscher, 1987;Yamano and Luiten, 1989;Milner andVeznedaroglu, 1993). GABAergic septal afferent terminalsdisplay a higher degree of selectivity; they terminate ononly GABAergic interneurons (especially those located inthe infragranular hilus) and form almost exclusively sym-metric synapses (Freund and Antal, 1988; Gulyas et al.,1991a).The different patterns of connectivity of the cholinergic
and GABAergic components of septohippocampal afferentssuggest that these two afferent components may differen-tially regulate subpopulations of hippocampal neurons,especially interneurons. This notion is supported in partby the previous observation that transection of the fornixresults in transient increases in the detectable number ofNPY-immunoreactive neurons located in the central hilus(Bayer and Milner, 1993), but not GABA-immunoreactiveneurons located in the infragranular hilus (Milner, unpub-lished observation). However, because lesions of the fornixdisrupt both cholinergic and GABAergic septal afferentpathways as well as those arising from the brainstem, morerigorous evidence to support this hypothesis is needed.A number of light microscopic studies have demon-
strated that more than 90% of the cholinergic neurons inthe septal complex contain both low-affinity (p75NTR) andhigh-affinity (p140protrk or trkA) nerve growth factor (NGF)receptors (Davies, 1979; Dawbarn et al., 1988; Mufson etal., 1989; Woolf et al., 1989; Pioro and Cuello, 1990; Gibbsand Pfaff, 1994) and retrogradely transport NGF (Seilerand Schwab, 1984; Johnson et al., 1987). In contrast,almost none of the GABAergic septohippocampal neuronscontain the low- and high-affinity NGF receptors or areresponsive to NGF (Batchelor et al., 1989; Koliatsos et al.,1994). An immunotoxin that destroys cells that expressp75NTR (a monoclonal antibody to p75NTR [192 IgG] chemi-cally linked to a ribosome-inactivating toxin, saporin; 192
IgG-saporin) has been developed, which can be used toremove selectively the cholinergic septohippocampal neu-rons without any apparent damage to the GABAergicseptohippocampal neurons (Wiley et al., 1991; Book et al.,1992, 1994; Heckers et al., 1994; Lee et al., 1994; Torres etal., 1994). Thus, it is now possible to determine whetherselective destruction of the cholinergic septal afferentterminals can result in cytoarchitectural changes in hippo-campal interneurons and whether these changes, if any,are selective.In the present study, we examined whether removal of
cholinergic septal afferent terminals following 192 IgG-saporin treatment results in changes in the number and/ordistribution of subpopulations of interneurons containingNPY, SOM, and GABA immunoreactivities in the hilus ofthe dentate gyrus. The size of the soma was also examinedto control for how changes in this parameter might affectimmunoreactive cell counts. To determine whether anypotential changes in these immunoreactivities could beassociated with changes in the total number of smallnonprincipal cells, specific subfields in adjacent Nisslsections were examined with the optical dissector. BecauseGABA, NPY, and SOM immunoreactivities are expressedin subpopulations of hippocampal GABAergic interneu-rons, a second series of experiments examined the amountof overlap of these populations by using dual-labelingimmunofluorescence.
MATERIALS AND METHODS
192 IgG-saporin experiments
Surgical procedures. Atotal of 26 adultmale Sprague-Dawley rats (250–300 g) were used in this portion of thestudy. All methods were approved by the CUMC Institu-tional Animal Care and Use Committee. All rats wereanesthetized with chloral hydrate (0.42 g/kg, i.p.) andplaced in a stereotaxic apparatus. For experimental rats(n 5 15), single injections of 192 IgG-saporin (10 µl of a0.33–0.42 mg/ml saline solution yielding a final dose of3.3–4.2 µg) were placed into the lateral ventricle, aspreviously described (Wiley et al., 1991). Control rats (n 58) received injections of either saline alone or nothing.Methods for the production of the 192 IgG-saporin solutionhave been detailed previously (Wiley and Lappi, 1994).Matched sets of rats (i.e., 1 or 2 192 IgG-saporin injectedrats and 1 control rat) were killed on the same daypostsurgery at the following time points: 1 week (n 5 3), 2weeks (n 5 3), 4 weeks (n 5 3), 8 weeks (n 5 3), 12 weeks(n 5 3), and 22–24 weeks (n 5 8).Immunocytochemistry. Rats injected with 192 IgG-
saporin were always killed simultaneously with theircontrols by using the same batches of fixatives and buffers.At each time point, sets of rats were deeply anesthetizedwith Nembutal (150 mg/kg, i.p.) and perfused through theascending aorta sequentially with solutions of (1) 10–15mlof normal saline (0.9%) containing 1,000 units/ml of Hepa-rin, (2) 55 ml of 3.75% acrolein (Polysciences, Warrington,PA) and 2% paraformaldehyde in 0.1 M phosphate buffer(PB; pH 7.4), and (3) 200 ml of 2% paraformaldehyde inPB. The regions of the forebrain containing the septalcomplex and hippocampal formation were removed andcut into 5-mm-thick coronal blocks that were stored in thelatter fixative for an additional 30 minutes. After markingthe blocks with coded hole punches, sections (40 µm thick)were then cut on the same automatic Vibratome and
CHANGES IN HIPPOCAMPAL NPY NEURONS 47

collected in 0.1 M PB. To maximize uniform immunocyto-chemical labeling conditions for the purposes of quantita-tive comparisons (Auchus and Pickel, 1992), sections fromeach brain (3 brains maximum) were pooled into singlecontainers prior to treatment with 1% sodium borohydridein PB for 30 minutes (Milner and Veznedaroglu, 1992).Polyclonal antibodies raised in rabbits to NPY (Penin-
sula, Belmont, CA), SOM 28 (Immunonuclear, Stillwater,MN) and GABA (Sigma, St. Louis, MO) were used in thesestudies. The NPY and SOM 28 antibodies were tested forspecificity to their respective peptides by using both immu-nodot blots and preadsorption techniques (Sloviter andNilavier, 1987; Milner and Veznedaroglu, 1992; Bayer andMilner, 1993). GABA antibody labeling was identical tothat reported previously (Milner and Bacon, 1989a).Mousemonoclonal antibodies to the rat p75NTR, either from thesame batch as to which the saporin was conjugated or fromBoehinger-Manheim (Indianapolis, IN), were employed.The p75NTR antibodies have been characterized previously(Chandler et al., 1984) and yielded identical patterns oflabeling in the basal forebrain.Antibodies to NPY, SOM, and GABA (diluted 1:6,000,
1:2,000, and 1:35,000, respectively) were localized immuno-cytochemically in the hippocampal formation by using theavidin-biotin complex (ABC), as described previously (Mil-ner and Veznedaroglu, 1992). For lesion verification, anti-bodies to the rat p75NTR were similarly localized in theseptal complex. Primarily antibody diluents for NPY,SOM, and the rat p75NTR antibodies contained 0.25%Triton X-100. An additional 1-in-10 series of sectionsthrough the hippocampal formation and septal complexwere processed for AChE histochemistry (Hedreen et al.,1985) or thionin (Nissl) stain. The labeled sections weremounted on acid-cleaned slides previously coated with 1%gelatin, air dried, dehydrated, and coverslipped with DPX(Aldrich Chemical, Milwakee, WI). The final light micro-scopic preparations were examined and photographedwith Kodak T-Max 100 film by using brightfield optics on aNikonMicrophotmicroscope. Final photomicrographsweregenerated from photographic prints captured with anArcus II Desktop Scanner attached to a Power Macintosh8500/120 by usingAdobe Photoshop 3.0 andQuarkX-Press.The final line drawing in Figure 4 was made from print-outs generated from images captured from the microscope(see below) by usingAdobe Illustrator.In addition, an alternate series of hippocampal sections
immunolabeled for NPY, SOM, or GABAwas prepared forelectron microscopy. For this series, the sections wereimmunolabeled identically to that described above exceptthat the diluent for the NPY and SOM antibodies con-tained 0.035% Triton X-100. Sections were embedded inEpon 812, and ultrathin sections were prepared as de-scribed previously (Milner and Veznedaroglu, 1992). Finalpreparations were examined on a Philips CM10 electronmicroscope.
Data analysis
Immunohistochemical cell counts. For each postle-sion time point, matched sets of sections (i.e., in terms ofnumber of sections and rostrocaudal level) of the dorsalhilus of the dentate gyrus of paired experimental andcontrol rats were compared. The sets were examined as tothe (1) number of neurons immunoreactive for NPY, SOM,or GABA containing visible nuclei, (2) cross-sectional hilararea (to measure potential shrinkage), and (3) size of
individual NPY-immunoreactive cells (measured in termsof both soma profile area and the minor axis diameter). Todetermine cell number, an observer blind to the experimen-tal conditions first captured images of the dentate hilarregion by using a Dage CCD72S video camera and NIHImage 1.50 software. The number of labeled neurons withnuclei then were counted from a printout of the image.Questionable elements were checked directly with a Nikonlight microscope. The cross-sectional area of the hilus wasdetermined by using NIH Image. Captured images werealso qualitatively examined for potential differences in thedistribution of labeled cells. For purposes of clarity, totallabeled cell numbers from lesioned animals have beengraphically expressed as percentages of the total cellnumber of matched controls (see Fig. 6). Apparent differ-ences in the total number of NPY-labeled cells counted peranimal and dorsal hilar volume (extrapolated from cross-sectional areas) between pairs of matched control andimmunolesioned animals across time points were exam-ined by using paired t-tests (each pair being comprised of 1randomly chosen experimental rat and 1 control rat from agiven time point). Hippocampal nomenclature employed inthis study is that recently reviewed by Patton and Mc-Naughton (1995). The term ‘‘infragranular hilus’’ denotesthat region of the hilus approximately 50 µm below thegranule cell layer.Two approaches were employed to determine the distri-
bution of both the soma profile area and minor axisdiameter of NPY-immunoreactive cells. First, by using theelectron microscope, all profiles of immunoreactive somacontaining identifiable nuclear portions were photo-graphed from single ultrathin sections (2 blocks each) fromthe 2- and 24-week time points from control and experimen-tal groups. Uniformly scaled prints of each profile werethen placed on a graphics tablet, and the profile borderswere traced into NIH Image to determine profile area andminor axis diameter. In the second approach, NPY-immunoreactive cells from the central and crest regions of5 sections, each from those sections previously analyzedfrom control and immunolesioned rats at the 1- and24-week time points, were viewed with a 203 objective ona Nikon Microphot microscope and captured by using NIHImage. These gray-scale images were then converted tobinary images by using a constant threshold level. Theanalyzed particle function was then used to determinesoma profile area and the minor axis diameter for all cellimages that had contained an identifiable nucleus. Todetermine whether there was significant cell skrinkagethat could compromise total cell counts, differences be-tween control and experimental values were examined byusing unpaired t-tests.Stereological counts of Nissl-stained sections. To
determine whether there were differences in the number ofsmall nonprincipal cells (5–15 µm in diameter; readilydistinguishable from the larger cells, which were .20 µmin diameter), the optical dissector (Gundersen, 1986) wasapplied in a blind manner to matched sets of sections (bothin terms of number and rostrocaudal position) from pairedcontrol and 192 IgG-saporin injected rats. For each sectionthrough the hilus, 7 matched test fields (with forbiddenlines) from defined subregions (including the hilar tip,center, inner, and outer blades) were examined with a1003 oil objective and a grid reticule. Numerical density,NV, was determined by using the equation: NV 5 Q2/(A*d),where Q2 is the number of cells in the reference plane that
48 T.A. MILNER ET AL.

are not in the look-up plane (taken separately moving upand down between the planes and then averaged),A is areaof the examined fields (96 µm2), and d is the distancebetween the reference and look-up planes (3-µm distancesperpendicular to the plane of section were determined byusing calibrated fine-focus knob graduations). Because the40-µm sections had shrunk to a thickness of 8 µm duringdehydration on average, a correction factor of 53 wasapplied to the numerical density measurements. Finally,to control for the potential effect of differential changes inhilar volume, the volumes of the portions of the dorsalhilus examined were determined by measuring cross-sectional hilar area and extrapolating to a 400-µm blockfor each section (1 in 10 sampled sections), and the averagenumerical density (for all test fields in a given animal) wasconverted to total cell number (for either 4 3 400-µmblocks or 6 3 400-µm blocks, depending on the time point).Differences in total cell number across time points werestatistically examined by using a paired t-test.
Dual-labeling immunofluorescence
Three adult male Sprague-Dawley rats (275–325 g) wereused in this portion of the study. The rats were deeplyanesthetized with Nembutal (50 mg/kg, i.p.) and perfusedthrough the ascending aorta with either 4% paraformalde-hyde in 0.1 M PB for 6 minutes (n 5 2) or 3.75% acroleinand paraformaldehyde, as described above (n 5 1). Thehippocampal formation was blocked and sectioned asdescribed in the previous experiment.Sections through the hippocampal formation were incu-
bated in (1) 0.5% bovine serum albumin (BSA) in 0.1 MTris saline (TS), pH 7.6, for 30 minutes; (2) a combinationof GABA antiserum (1:1,000 dilution; Bayer and Pickel,1990) and the antisera to either NPY (1:2,000 dilution) orSOM (1:500 dilution); and (3) a combination of a 1:100dilution of goat anti-rat fluorescein isothiocyanate (FITC)-conjugated immunoglobulin (IgG) and goat anti-rabbitTexas Red IgG (both from Jackson Laboratories, WestGrove, PA) for 30 minutes at room temperature. Controlsections were incubated with only one primary antiserum,followed by both secondary IgGs. The primary and second-ary antibodies were diluted in 0.1% BSA in TS. Intermedi-ate washes were carried out in TS at room temperaturewith constant mild agitation. The labeled sections weremounted on acid-cleaned slides previously coated with 1%gelatin, air dried, and coverslipped with Citifluor glycerin(Ted Pella, Redding, CA).The hilus of the dentate gyrus was photographed from at
least 7 sections from each of the 2 paraformaldyhyde-fixedrats under epifluorescence illumination on a Zeiss micro-scope. FITC labeling was visualized by using a blue filterwith an excitation range of 45–49 nm, and Texas Redlabeling was viewed with a green filter with an excitationrange of 51–56 nm. Single- and dual-labeled neurons werecounted from all sections.
RESULTS
192 IgG-saporin experiments
Effectiveness of immunolesions. To determine theselectivity of the 192 IgG-saporin immunolesion, immuno-cytochemical localization of p75NTR in the septal complexand AChE histochemistry in the hippocampal formationwere performed on all rats. The number of detectable p75NTR-labeled neurons in the septal complex was substantially
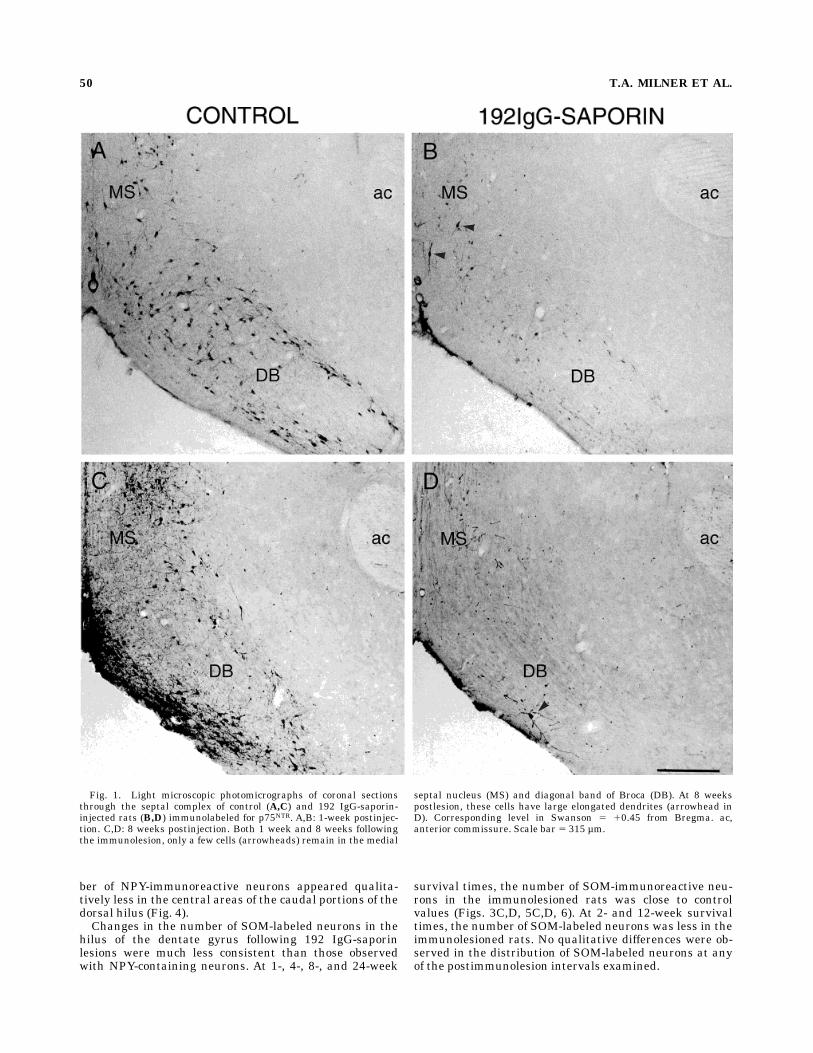
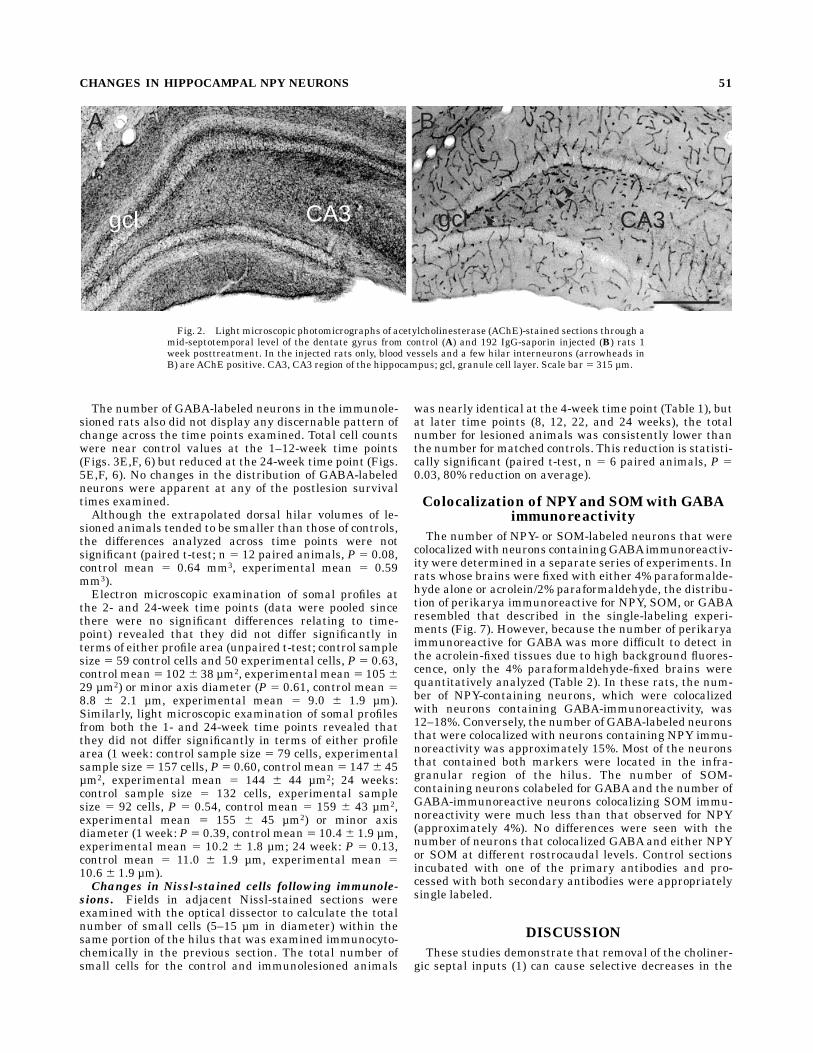
diminished in immunolesioned rats as compared withcontrols as early as 1 week postinjection (Fig. 1A,B). Theseverity of the decrease in p75NTR-immunoreactive neu-rons was similar at all postlesion time intervals examinedexcept for one 4-week rat, in which the number of p75NTR-labeled cells was reduced to only about half that of thecontrol. Moreover, at 8 weeks postlesion, the remainder ofthe p75NTR-containing neurons appeared to have longer andthicker dendritic branches (Fig. 1C,D). AChE-stained fi-bers also weremostly absent in all lamina of the hippocam-pal formation following injections of 192 IgG-saporin at alltime points examined (except for the one 4-week animal).In the dentate gyrus, AChE staining was observed in bloodvessels and in a few hilar interneurons (Fig. 2).In the course of staining for NPY, intense NPY-like
immunoreactivity (NPY-LI) was observed in the mossyfiber pathway of 1 rat at the 12-week survival time pointand of 3 rats at the 24-week1 survival time point. Becausethis pattern of labeling is indicative of the occurrence ofstrong seizure activity (Beninato and Spencer, 1988; Tonderet al., 1994; Schwarzer et al., 1995), these animals werenot used in the final analysis.Changes in NPY, SOM, and GABA immunoreactivi-
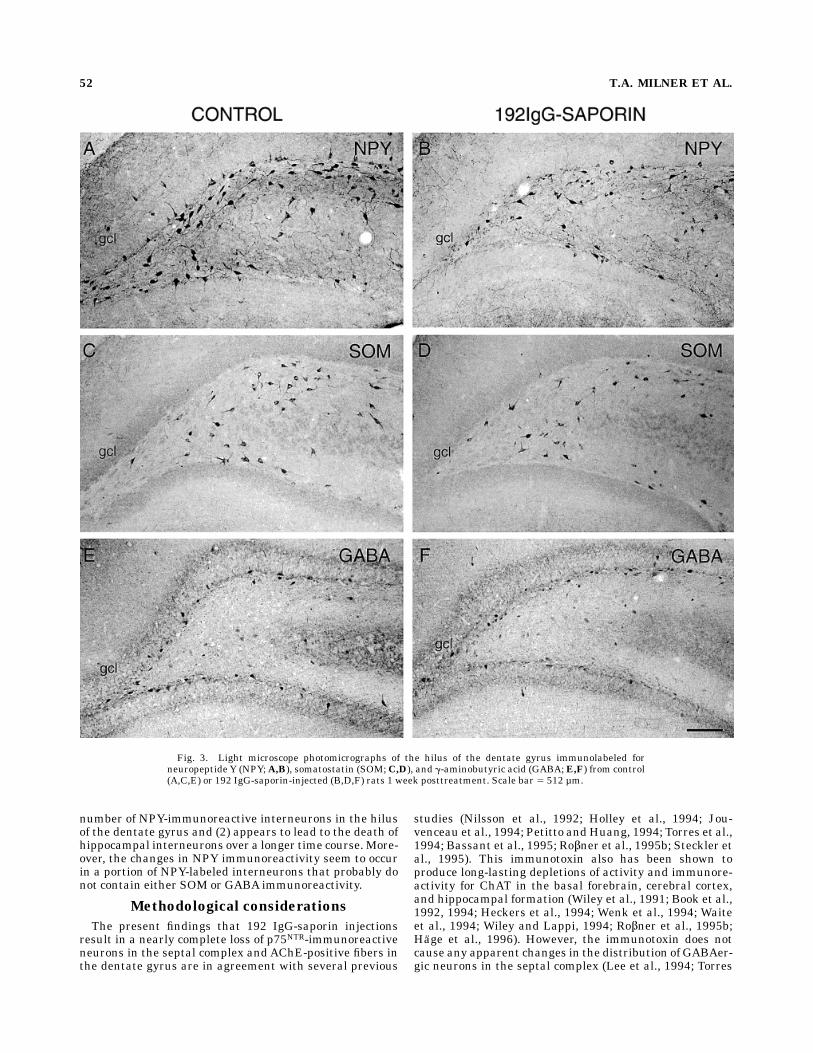
ties following 192 IgG immunolesions. The distribu-tion pattern of neuronal cell bodies immunolabeled forNPY, SOM, and GABA in the dentate gyrus of normal andcontrol rats was identical to that previously reported(Milner and Bacon, 1989a,b; Deller and Leranth, 1990;Milner and Veznedaroglu, 1992). In brief, perikarya withNPY-LI were observed most frequently in the central andinner blades of the hilus and to a lesser extent in the outerblade of the hilus (Figs. 3A, 5A). NPY-labeled perikaryawere rarely observed in the molecular layer. Perikaryawith SOM-like immunoreactivity (SOM-LI) were seenmost frequently in the inner blade and central region ofthe hilus; they were occasionally observed in the outerblade but usually not found in the infragranular region ofthe hilus (Figs. 3C, 5C). Unlike perikarya with NPY-LI orSOM-LI, GABA-immunoreactive perikarya were most nu-merous in the infragranular region of both the inner andouter blades of the hilus (Figs. 3E, 5E). Cells with GABAimmunoreactivity (GABA-I) were rarely observed in thecentral regions of the hilus.At all time points examined, the detectable number of
NPY-labeled neurons was noticeably less in injected rats(to 73% of control values, on average; Figs. 3A,B, 5A,B, 6).Because the apparent reduction in the number of NPY-containing neurons between matched immunolesionedand control rats appeared to be relatively consistent at alltime points, the data were collapsed across time points forstatistical analysis. The total number of cells counted peranimal was significantly lower in experimental animalsthan in matched control animals (paired t-test, n 5 12paired animals, P 5 0.0012, control mean 5 546, experi-mental mean 5 401). The decrease in the number ofdetectable NPY-immunoreactive neurons did not appear tobe selective for any particular subregion of the rostralportions of the dorsal hilus, although the detectable num-
1The 22- and 24-week time points had no differences in the extent of thelesion as judged by 192 IgG immunoreactivity in the septal complex andAChE histochemistry in the hippocampal formation and in the changes inthe number of NPY-, SOM- and GABA-immunoreactive cells in the hilus.Thus, for simplicity, these animals are referred to as 24-week rats, althoughthey were analyzed as separate 22- and 24-week pairings in the pairedt-tests.
CHANGES IN HIPPOCAMPAL NPY NEURONS 49
ber of NPY-immunoreactive neurons appeared qualita-tively less in the central areas of the caudal portions of thedorsal hilus (Fig. 4).Changes in the number of SOM-labeled neurons in the
hilus of the dentate gyrus following 192 IgG-saporinlesions were much less consistent than those observedwith NPY-containing neurons. At 1-, 4-, 8-, and 24-week
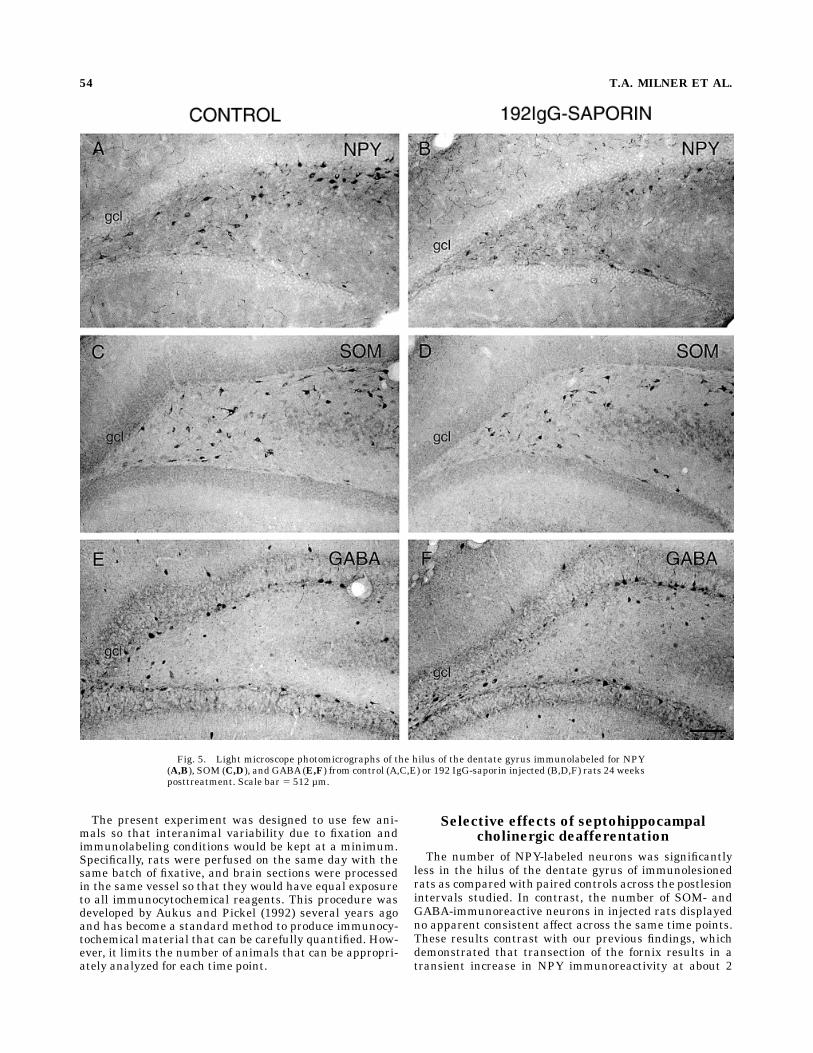
survival times, the number of SOM-immunoreactive neu-rons in the immunolesioned rats was close to controlvalues (Figs. 3C,D, 5C,D, 6). At 2- and 12-week survivaltimes, the number of SOM-labeled neurons was less in theimmunolesioned rats. No qualitative differences were ob-served in the distribution of SOM-labeled neurons at anyof the postimmunolesion intervals examined.
Fig. 1. Light microscopic photomicrographs of coronal sectionsthrough the septal complex of control (A,C) and 192 IgG-saporin-injected rats (B,D) immunolabeled for p75NTR. A,B: 1-week postinjec-tion. C,D: 8 weeks postinjection. Both 1 week and 8 weeks followingthe immunolesion, only a few cells (arrowheads) remain in the medial
septal nucleus (MS) and diagonal band of Broca (DB). At 8 weekspostlesion, these cells have large elongated dendrites (arrowhead inD). Corresponding level in Swanson 5 10.45 from Bregma. ac,anterior commissure. Scale bar 5 315 µm.
50 T.A. MILNER ET AL.
The number of GABA-labeled neurons in the immunole-sioned rats also did not display any discernable pattern ofchange across the time points examined. Total cell countswere near control values at the 1–12-week time points(Figs. 3E,F, 6) but reduced at the 24-week time point (Figs.5E,F, 6). No changes in the distribution of GABA-labeledneurons were apparent at any of the postlesion survivaltimes examined.Although the extrapolated dorsal hilar volumes of le-
sioned animals tended to be smaller than those of controls,the differences analyzed across time points were notsignificant (paired t-test; n 5 12 paired animals, P 5 0.08,control mean 5 0.64 mm3, experimental mean 5 0.59mm3).Electron microscopic examination of somal profiles at
the 2- and 24-week time points (data were pooled sincethere were no significant differences relating to time-point) revealed that they did not differ significantly interms of either profile area (unpaired t-test; control samplesize 5 59 control cells and 50 experimental cells, P 5 0.63,control mean5 1026 38 µm2, experimental mean5 105629 µm2) or minor axis diameter (P 5 0.61, control mean 58.8 6 2.1 µm, experimental mean 5 9.0 6 1.9 µm).Similarly, light microscopic examination of somal profilesfrom both the 1- and 24-week time points revealed thatthey did not differ significantly in terms of either profilearea (1 week: control sample size 5 79 cells, experimentalsample size 5 157 cells, P 5 0.60, control mean 5 147 6 45µm2, experimental mean 5 144 6 44 µm2; 24 weeks:control sample size 5 132 cells, experimental samplesize 5 92 cells, P 5 0.54, control mean 5 159 6 43 µm2,experimental mean 5 155 6 45 µm2) or minor axisdiameter (1 week: P 5 0.39, control mean 5 10.4 6 1.9 µm,experimental mean 5 10.2 6 1.8 µm; 24 week: P 5 0.13,control mean 5 11.0 6 1.9 µm, experimental mean 510.6 6 1.9 µm).Changes in Nissl-stained cells following immunole-
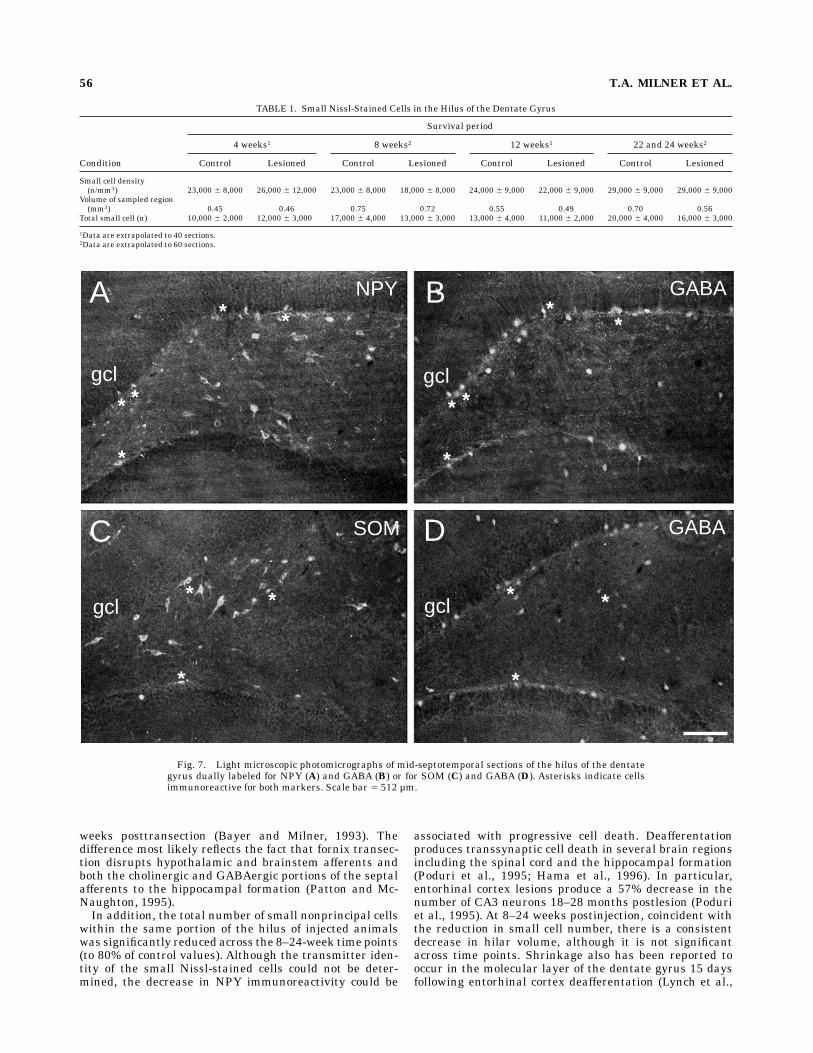
sions. Fields in adjacent Nissl-stained sections wereexamined with the optical dissector to calculate the totalnumber of small cells (5–15 µm in diameter) within thesame portion of the hilus that was examined immunocyto-chemically in the previous section. The total number ofsmall cells for the control and immunolesioned animals
was nearly identical at the 4-week time point (Table 1), butat later time points (8, 12, 22, and 24 weeks), the totalnumber for lesioned animals was consistently lower thanthe number formatched controls. This reduction is statisti-cally significant (paired t-test, n 5 6 paired animals, P 50.03, 80% reduction on average).
Colocalization of NPY and SOM with GABAimmunoreactivity
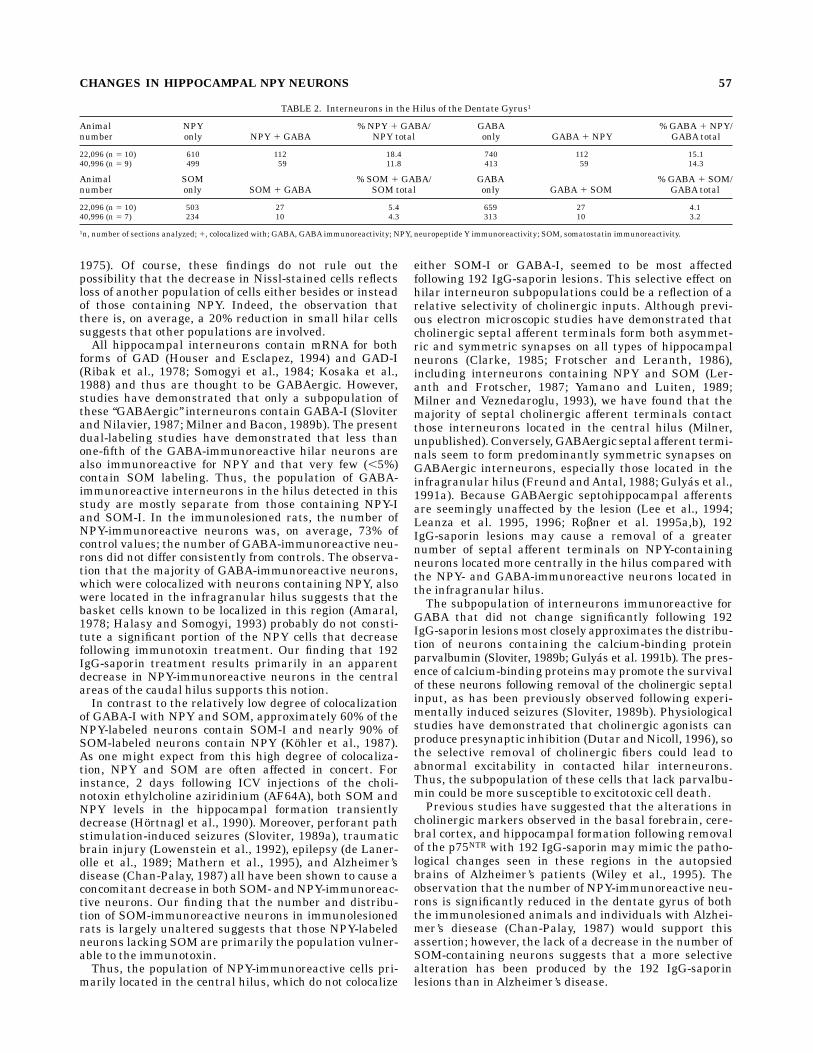
The number of NPY- or SOM-labeled neurons that werecolocalizedwith neurons containingGABAimmunoreactiv-ity were determined in a separate series of experiments. Inrats whose brains were fixed with either 4% paraformalde-hyde alone or acrolein/2% paraformaldehyde, the distribu-tion of perikarya immunoreactive for NPY, SOM, or GABAresembled that described in the single-labeling experi-ments (Fig. 7). However, because the number of perikaryaimmunoreactive for GABA was more difficult to detect inthe acrolein-fixed tissues due to high background fluores-cence, only the 4% paraformaldehyde-fixed brains werequantitatively analyzed (Table 2). In these rats, the num-ber of NPY-containing neurons, which were colocalizedwith neurons containing GABA-immunoreactivity, was12–18%. Conversely, the number of GABA-labeled neuronsthat were colocalized with neurons containing NPY immu-noreactivity was approximately 15%. Most of the neuronsthat contained both markers were located in the infra-granular region of the hilus. The number of SOM-containing neurons colabeled for GABAand the number ofGABA-immunoreactive neurons colocalizing SOM immu-noreactivity were much less than that observed for NPY(approximately 4%). No differences were seen with thenumber of neurons that colocalized GABA and either NPYor SOM at different rostrocaudal levels. Control sectionsincubated with one of the primary antibodies and pro-cessed with both secondary antibodies were appropriatelysingle labeled.
DISCUSSION
These studies demonstrate that removal of the choliner-gic septal inputs (1) can cause selective decreases in the
A B
gcl CA3 gcl CA3
Fig. 2. Light microscopic photomicrographs of acetylcholinesterase (AChE)-stained sections through amid-septotemporal level of the dentate gyrus from control (A) and 192 IgG-saporin injected (B) rats 1week posttreatment. In the injected rats only, blood vessels and a few hilar interneurons (arrowheads inB) areAChE positive. CA3, CA3 region of the hippocampus; gcl, granule cell layer. Scale bar 5 315 µm.
CHANGES IN HIPPOCAMPAL NPY NEURONS 51
number of NPY-immunoreactive interneurons in the hilusof the dentate gyrus and (2) appears to lead to the death ofhippocampal interneurons over a longer time course.More-over, the changes in NPY immunoreactivity seem to occurin a portion of NPY-labeled interneurons that probably donot contain either SOM or GABA immunoreactivity.
Methodological considerations
The present findings that 192 IgG-saporin injectionsresult in a nearly complete loss of p75NTR-immunoreactiveneurons in the septal complex and AChE-positive fibers inthe dentate gyrus are in agreement with several previous
studies (Nilsson et al., 1992; Holley et al., 1994; Jou-venceau et al., 1994; Petitto andHuang, 1994; Torres et al.,1994; Bassant et al., 1995; Ro¬ner et al., 1995b; Steckler etal., 1995). This immunotoxin also has been shown toproduce long-lasting depletions of activity and immunore-activity for ChAT in the basal forebrain, cerebral cortex,and hippocampal formation (Wiley et al., 1991; Book et al.,1992, 1994; Heckers et al., 1994; Wenk et al., 1994; Waiteet al., 1994; Wiley and Lappi, 1994; Ro¬ner et al., 1995b;Hage et al., 1996). However, the immunotoxin does notcause any apparent changes in the distribution of GABAer-gic neurons in the septal complex (Lee et al., 1994; Torres
Fig. 3. Light microscope photomicrographs of the hilus of the dentate gyrus immunolabeled forneuropeptide Y (NPY;A,B), somatostatin (SOM;C,D), and g-aminobutyric acid (GABA;E,F) from control(A,C,E) or 192 IgG-saporin-injected (B,D,F) rats 1 week posttreatment. Scale bar 5 512 µm.
52 T.A. MILNER ET AL.

et al., 1994; Baxter et al., 1995; Ro¬ner et al., 1995a,b),thus supporting the contention that 192 IgG-saporin selec-tively destroys cholinergic septohippocampal neurons.In the present study, injections of 192 IgG-saporin were
made intracerebroventricularly (ICV) rather than intrasep-tally. This method of delivery was chosen for severalreasons. First, immunolesioning by means of ICV injec-tions has been shown to be easily reproducible and resultsin similar decreases in hippocampal cholinergic markers,as seen with intraseptal injections (Wiley, 1992; Petittoand Huang, 1994; Torres et al., 1994; Zhang et al., 1996).Second, intraseptal injections of the immunotoxin maycause nonspecific tissue damage attributable to the needletrack rather than to the immunotoxin per se (Petitto andHuang, 1994; Torres et al., 1994). Third, because ourupcoming analysis of the present animals will also includethe septal complex, ICV injections of the immunotoxinwould do the least morphological damage to this region.However, because ICV injections of 192 IgG-saporin cancause destruction of the cerebellar Purkinje neurons (Wileyet al., 1991; Heckers et al., 1994), future studies will
compare the effect of the two types of immunotoxin deliv-ery on hippocampal NPY immunoreactivity.Two types of quantitative analyses were employed in
this study. For the comparison of the number of immunore-active cells in the 192 IgG-saporin-lesioned dentate gyruswith that of paired controls, total numbers of labeled cellsin matched sets of sections were used. This method ofanalysis was chosen because (1) the number of labeledcells per section was relatively low and (2) the comparisonemphasized relative cell number. Because reductions inthe average size of labeled cells in experimental tissuecould affect the apparent number of cells counted in thismanner, cell size (measured in terms of somal profile areaand minor axis diameter, at both light and electron micro-scopic levels) was examined. No significant differenceswere observed. For the quantification of the number ofsmall Nissl-stained cells in the hilus, the optical dissectorwas employed. This method of examination provides anunbiased estimate of cell number (Coggeshall and Lekan,1996) and was more appropriate for the Nissl-stainedmaterial because there were many more cells per section.
Control 192-IgG-Saporin
80
77
70
66
58
54
52
36
8 WEEKS
gclob
ib
cen
cells
Fig. 4. Camera lucida drawings showing the distribution of NPY-immunoreactive neurons in the hilusof the dentate gyrus in control and 192 IgG-saporin-injected rats 8 weeks postimmunolesion. Numbers ofNPY-labeled neurons are indicated to the right of each drawing. cen, central region of hilus; ib, inner bladeof hilus; ob, medial blade of hilus. Scale bar 5 250 µm.
CHANGES IN HIPPOCAMPAL NPY NEURONS 53
The present experiment was designed to use few ani-mals so that interanimal variability due to fixation andimmunolabeling conditions would be kept at a minimum.Specifically, rats were perfused on the same day with thesame batch of fixative, and brain sections were processedin the same vessel so that they would have equal exposureto all immunocytochemical reagents. This procedure wasdeveloped by Aukus and Pickel (1992) several years agoand has become a standard method to produce immunocy-tochemical material that can be carefully quantified. How-ever, it limits the number of animals that can be appropri-ately analyzed for each time point.
Selective effects of septohippocampalcholinergic deafferentation
The number of NPY-labeled neurons was significantlyless in the hilus of the dentate gyrus of immunolesionedrats as compared with paired controls across the postlesionintervals studied. In contrast, the number of SOM- andGABA-immunoreactive neurons in injected rats displayedno apparent consistent affect across the same time points.These results contrast with our previous findings, whichdemonstrated that transection of the fornix results in atransient increase in NPY immunoreactivity at about 2
Fig. 5. Light microscope photomicrographs of the hilus of the dentate gyrus immunolabeled for NPY(A,B), SOM (C,D), and GABA(E,F) from control (A,C,E) or 192 IgG-saporin injected (B,D,F) rats 24 weeksposttreatment. Scale bar 5 512 µm.
54 T.A. MILNER ET AL.
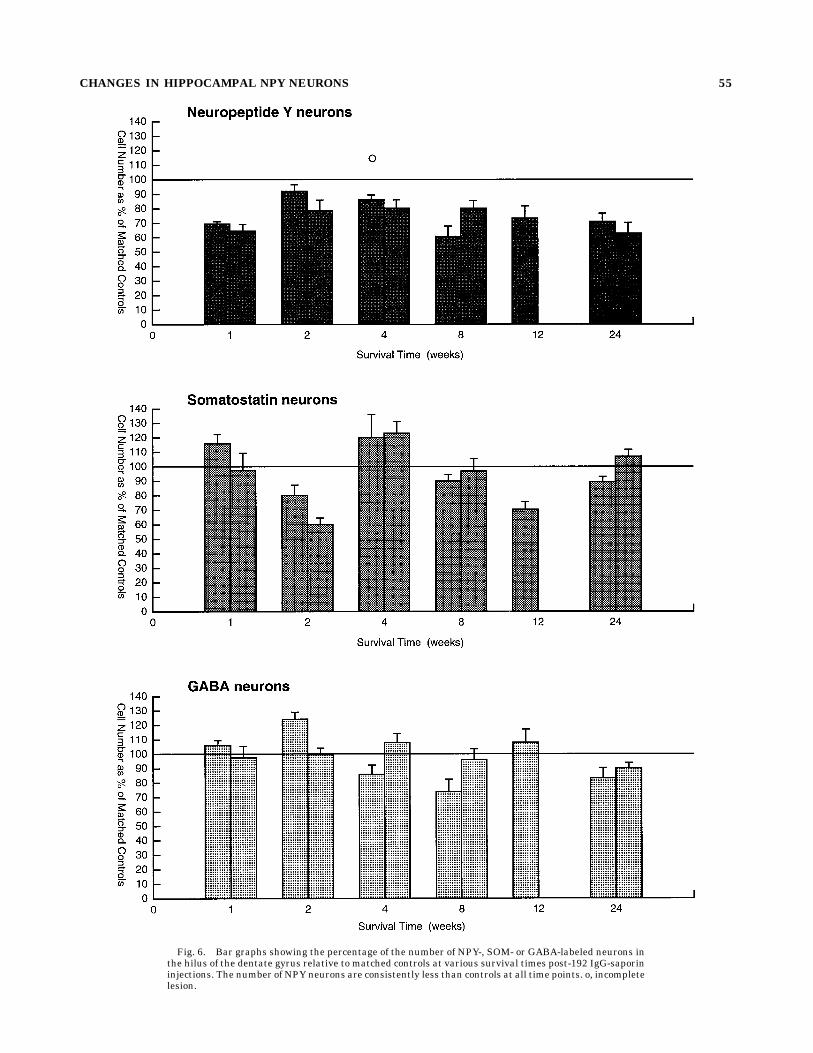
Fig. 6. Bar graphs showing the percentage of the number of NPY-, SOM- or GABA-labeled neurons inthe hilus of the dentate gyrus relative to matched controls at various survival times post-192 IgG-saporininjections. The number of NPY neurons are consistently less than controls at all time points. o, incompletelesion.
CHANGES IN HIPPOCAMPAL NPY NEURONS 55
weeks posttransection (Bayer and Milner, 1993). Thedifference most likely reflects the fact that fornix transec-tion disrupts hypothalamic and brainstem afferents andboth the cholinergic and GABAergic portions of the septalafferents to the hippocampal formation (Patton and Mc-Naughton, 1995).In addition, the total number of small nonprincipal cells
within the same portion of the hilus of injected animalswas significantly reduced across the 8–24-week time points(to 80% of control values). Although the transmitter iden-tity of the small Nissl-stained cells could not be deter-mined, the decrease in NPY immunoreactivity could be
associated with progressive cell death. Deafferentationproduces transsynaptic cell death in several brain regionsincluding the spinal cord and the hippocampal formation(Poduri et al., 1995; Hama et al., 1996). In particular,entorhinal cortex lesions produce a 57% decrease in thenumber of CA3 neurons 18–28 months postlesion (Poduriet al., 1995). At 8–24 weeks postinjection, coincident withthe reduction in small cell number, there is a consistentdecrease in hilar volume, although it is not significantacross time points. Shrinkage also has been reported tooccur in the molecular layer of the dentate gyrus 15 daysfollowing entorhinal cortex deafferentation (Lynch et al.,
TABLE 1. Small Nissl-Stained Cells in the Hilus of the Dentate Gyrus
Condition
Survival period
4 weeks1 8 weeks2 12 weeks1 22 and 24 weeks2
Control Lesioned Control Lesioned Control Lesioned Control Lesioned
Small cell density(n/mm3) 23,000 6 8,000 26,000 6 12,000 23,000 6 8,000 18,000 6 8,000 24,000 6 9,000 22,000 6 9,000 29,000 6 9,000 29,000 6 9,000
Volume of sampled region(mm3) 0.45 0.46 0.75 0.72 0.55 0.49 0.70 0.56
Total small cell (n) 10,000 6 2,000 12,000 6 3,000 17,000 6 4,000 13,000 6 3,000 13,000 6 4,000 11,000 6 2,000 20,000 6 4,000 16,000 6 3,000
1Data are extrapolated to 40 sections.2Data are extrapolated to 60 sections.
A*
B
C D
gcl
NPY
SOM
GABA
gcl
gcl gcl
GABA
***
* ** *
* *
*
** **
*
Fig. 7. Light microscopic photomicrographs of mid-septotemporal sections of the hilus of the dentategyrus dually labeled for NPY (A) and GABA (B) or for SOM (C) and GABA (D). Asterisks indicate cellsimmunoreactive for both markers. Scale bar 5 512 µm.
56 T.A. MILNER ET AL.
1975). Of course, these findings do not rule out thepossibility that the decrease in Nissl-stained cells reflectsloss of another population of cells either besides or insteadof those containing NPY. Indeed, the observation thatthere is, on average, a 20% reduction in small hilar cellssuggests that other populations are involved.All hippocampal interneurons contain mRNA for both
forms of GAD (Houser and Esclapez, 1994) and GAD-I(Ribak et al., 1978; Somogyi et al., 1984; Kosaka et al.,1988) and thus are thought to be GABAergic. However,studies have demonstrated that only a subpopulation ofthese ‘‘GABAergic’’ interneurons contain GABA-I (Sloviterand Nilavier, 1987; Milner and Bacon, 1989b). The presentdual-labeling studies have demonstrated that less thanone-fifth of the GABA-immunoreactive hilar neurons arealso immunoreactive for NPY and that very few (,5%)contain SOM labeling. Thus, the population of GABA-immunoreactive interneurons in the hilus detected in thisstudy are mostly separate from those containing NPY-Iand SOM-I. In the immunolesioned rats, the number ofNPY-immunoreactive neurons was, on average, 73% ofcontrol values; the number of GABA-immunoreactive neu-rons did not differ consistently from controls. The observa-tion that the majority of GABA-immunoreactive neurons,which were colocalized with neurons containing NPY, alsowere located in the infragranular hilus suggests that thebasket cells known to be localized in this region (Amaral,1978; Halasy and Somogyi, 1993) probably do not consti-tute a significant portion of the NPY cells that decreasefollowing immunotoxin treatment. Our finding that 192IgG-saporin treatment results primarily in an apparentdecrease in NPY-immunoreactive neurons in the centralareas of the caudal hilus supports this notion.In contrast to the relatively low degree of colocalization
of GABA-I with NPY and SOM, approximately 60% of theNPY-labeled neurons contain SOM-I and nearly 90% ofSOM-labeled neurons contain NPY (Kohler et al., 1987).As one might expect from this high degree of colocaliza-tion, NPY and SOM are often affected in concert. Forinstance, 2 days following ICV injections of the choli-notoxin ethylcholine aziridinium (AF64A), both SOM andNPY levels in the hippocampal formation transientlydecrease (Hortnagl et al., 1990). Moreover, perforant pathstimulation-induced seizures (Sloviter, 1989a), traumaticbrain injury (Lowenstein et al., 1992), epilepsy (de Laner-olle et al., 1989; Mathern et al., 1995), and Alzheimer’sdisease (Chan-Palay, 1987) all have been shown to cause aconcomitant decrease in both SOM- andNPY-immunoreac-tive neurons. Our finding that the number and distribu-tion of SOM-immunoreactive neurons in immunolesionedrats is largely unaltered suggests that those NPY-labeledneurons lacking SOM are primarily the population vulner-able to the immunotoxin.Thus, the population of NPY-immunoreactive cells pri-
marily located in the central hilus, which do not colocalize
either SOM-I or GABA-I, seemed to be most affectedfollowing 192 IgG-saporin lesions. This selective effect onhilar interneuron subpopulations could be a reflection of arelative selectivity of cholinergic inputs. Although previ-ous electron microscopic studies have demonstrated thatcholinergic septal afferent terminals form both asymmet-ric and symmetric synapses on all types of hippocampalneurons (Clarke, 1985; Frotscher and Leranth, 1986),including interneurons containing NPY and SOM (Ler-anth and Frotscher, 1987; Yamano and Luiten, 1989;Milner and Veznedaroglu, 1993), we have found that themajority of septal cholinergic afferent terminals contactthose interneurons located in the central hilus (Milner,unpublished). Conversely, GABAergic septal afferent termi-nals seem to form predominantly symmetric synapses onGABAergic interneurons, especially those located in theinfragranular hilus (Freund andAntal, 1988; Gulyas et al.,1991a). Because GABAergic septohippocampal afferentsare seemingly unaffected by the lesion (Lee et al., 1994;Leanza et al. 1995, 1996; Ro¬ner et al. 1995a,b), 192IgG-saporin lesions may cause a removal of a greaternumber of septal afferent terminals on NPY-containingneurons located more centrally in the hilus compared withthe NPY- and GABA-immunoreactive neurons located inthe infragranular hilus.The subpopulation of interneurons immunoreactive for
GABA that did not change significantly following 192IgG-saporin lesionsmost closely approximates the distribu-tion of neurons containing the calcium-binding proteinparvalbumin (Sloviter, 1989b; Gulyas et al. 1991b). The pres-ence of calcium-binding proteins may promote the survivalof these neurons following removal of the cholinergic septalinput, as has been previously observed following experi-mentally induced seizures (Sloviter, 1989b). Physiologicalstudies have demonstrated that cholinergic agonists canproduce presynaptic inhibition (Dutar andNicoll, 1996), sothe selective removal of cholinergic fibers could lead toabnormal excitability in contacted hilar interneurons.Thus, the subpopulation of these cells that lack parvalbu-min could be more susceptible to excitotoxic cell death.Previous studies have suggested that the alterations in
cholinergic markers observed in the basal forebrain, cere-bral cortex, and hippocampal formation following removalof the p75NTR with 192 IgG-saporin may mimic the patho-logical changes seen in these regions in the autopsiedbrains of Alzheimer’s patients (Wiley et al., 1995). Theobservation that the number of NPY-immunoreactive neu-rons is significantly reduced in the dentate gyrus of boththe immunolesioned animals and individuals with Alzhei-mer’s diesease (Chan-Palay, 1987) would support thisassertion; however, the lack of a decrease in the number ofSOM-containing neurons suggests that a more selectivealteration has been produced by the 192 IgG-saporinlesions than inAlzheimer’s disease.
TABLE 2. Interneurons in the Hilus of the Dentate Gyrus1
Animalnumber
NPYonly NPY 1 GABA
% NPY 1 GABA/NPY total
GABAonly GABA 1 NPY
% GABA 1 NPY/GABA total
22,096 (n 5 10) 610 112 18.4 740 112 15.140,996 (n 5 9) 499 59 11.8 413 59 14.3
Animalnumber
SOMonly SOM 1 GABA
% SOM 1 GABA/SOM total
GABAonly GABA 1 SOM
% GABA 1 SOM/GABA total
22,096 (n 5 10) 503 27 5.4 659 27 4.140,996 (n 5 7) 234 10 4.3 313 10 3.2
1n, number of sections analyzed; 1, colocalized with; GABA, GABA immunoreactivity; NPY, neuropeptide Y immunoreactivity; SOM, somatostatin immunoreactivity.
CHANGES IN HIPPOCAMPAL NPY NEURONS 57

ACKNOWLEDGMENTS
This study was supported by grant MH42834 from theNIMH, DA08259 from NIDA, and grant 18974 from theNIH (T.A.M.) and the Medical Research Service, DVA(R.G.W.). We thank Ms. Adriana Izquirdo, Mr. DanielBackenroth, and Ms. Joyce Hammel for assistance withthe quantitative analysis, Dr. Sue Aicher for statisticalconsultation, Ms. Nicole Quartarolo for help with theelectron microscopic analysis, Mr. Erol Veznedaroglu, Ms.Brenda Evans, and Ms. Zina Cappiello for technical help,and Ms. Joy Hornung for her photographic assistance.
LITERATURE CITED
Amaral, D.G. (1978) AGolgi study of the cell types in the hilar region of thehippocampus in the rat. J. Comp. Neurol. 182:851–914.
Amaral, D.G., and J. Kurz (1985) An analysis of the origins of thecholinergic and noncholinergic septal projections to the hippocampalformation of the rat. J. Comp. Neurol. 240:37–59.
Auchus, A.P., and V.M. Pickel (1992) Quantitative light microscopic demon-stration of increased pallidal and striatal met5-enkephalin-like immu-noreactivity in rats following chronic treatment with haloperidol butnot with clozapine: Implications for the pathogenesis of neuroleptic-induced movement disorders. Exp. Neurol. 117:17–27.
Baisden, R.H., M.L. Woodruff, and D.B. Hoover (1984) Cholinergic andnon-cholinergic septo-hippocampal projections:Adouble-label horserad-ish peroxidase-acetylcholinesterase study in the rabbit. Brain Res.290:146–151.
Bassant, M.H., E. Apartis, F.R. Jazat-Poindessous, R.G. Wiley, and Y.A.Lamour (1995) Selective immunolesion of the basal forebrain choliner-gic neurons: Effects on hippocampal activity during sleep and wakeful-ness in the rat. Neurodegeneration 4:61–70.
Batchelor, P.E., D.M. Armstrong, S.N. Blaker, and F.H. Gage (1989) Nervegrowth factor receptor and choline acetyltransferase colocalization inneurons within the rat forebrain: Response to fimbria–fornix transec-tion. J. Comp. Neurol. 284:187–204.
Baxter, M.G., D.J. Bucci, L.K. Gorman, R.G.Wiley, andM. Gallagher (1995)Selective immunotoxic lesions of basal forebrain cholinergic cells:Effects on learning and memory in rats. Behav. Neurosci. 109:714–722.
Bayer, L.E., and T.A. Milner (1993) Transient increases in neuropeptideY-like immunoreactivity in dentate hilar neurons following fimbria/fornix transections. J. Neurosci. Res. 34:434–441.
Bayer, V.E., and V.M. Pickel (1990) Ultrastructural localization of tyrosinehydroxylase in the rat ventral tegmental area: Relationship betweenimmunolabeling density and neuronal associations. J. Neurosci. 10:2996–3013.
Beninato, M., and R.F. Spencer (1988) The cholinergic innervation of the ratsubstantia nigra: A light and electron microscopic immunohistochemi-cal study. Exp. Brain Res. 72:178–184.
Book, A.A., R.G. Wiley, and J.B. Schweitzer (1992) Specificity of 192IgG-saporin for NGF receptor-positive cholinergic basal forebrain neu-rons in the rat. Brain Res. 590:350–355.
Book, A.A., R.G. Wiley, and J.B. Schweitzer (1994) 192 IgG-saporin: I.Specific lethality for cholinergic neurons in the basal forebrain of therat. J. Neuropathol. Exp. Neurol. 53:95–102.
Chan-Palay, V. (1987) Somatostatin immunoreactive neurons in the humanhippocampus and cortex shown by immunogold/silver intensification onvibratome sections: Coexistence with neuropeptide Y neurons, andeffects inAlzheimer-type dementia. J. Comp. Neurol. 260:201–223.
Chandler, C.E., L.M. Parsons, M. Hosang, and E.M. Shooter (1984) Amonoclonal antibody modulates the interaction of nerve growth factorwith PC12 cells. J. Biol. Chem. 259:6882–6889.
Clarke, D.J. (1985) Cholinergic innervation of the rat dentate gyrus: Animmunocytochemical and electron microscopical study. Brain Res.360:349–354.
Coggeshall, R.E., and H.A. Lekan (1996) Methods for determining numbersof cells and synapses: A case for more uniform standards of review. J.Comp. Neurol. 364:6–15.
Davies, P. (1979) Neurotransmitter-related enzymes in senile dementia oftheAlzheimer type. Brain Res. 171:319–327.
Dawbarn, D., S.J. Allen, and F.M. Semenenko (1988) Coexistence of cholineacetyltransferase and nerve growth factor receptors in the rat basalforebrain. Neurosci. Lett. 94:138–144.
de Lanerolle, N.C., J.H. Kim, R.J. Robbins, and D.D. Spencer (1989)Hippocampal interneuron loss and plasticity in human temporal lobeepilepsy. Brain Res. 495:387–395.
Deller, T., and C. Leranth (1990) Synaptic connections of neuropeptide Y(NPY) immunoreactive neurons in the hilar area of the rat hippocam-pus. J. Comp. Neurol. 300:433–447.
Dutar, P., and R.A. Nicoll (1996) Classification of muscarinic responses inhippocampus in terms of receptor subtypes and second-messenger sys-tems: Electrophysiological studies in vitro. J. Neurosci. 8:4214–4224.
Freund, T.F., and M. Antal (1988) GABA-containing neurons in the septumcontrol inhibitory interneurons in the hippocampus.Nature 336:170–173.
Frotscher, M. (1991) Target cell specificity of synaptic connections in thehippocampus. Hippocampus 1:123–130.
Frotscher, M., and C. Leranth (1986) The cholinergic innervation of the ratfascia dentata: Identification of target structures on granule cells bycombining choline acetyltransferase immunocytochemistry and Golgiimpregnation. J. Comp. Neurol. 243:58–70.
Gaykema, R.P.A., P.G.M. Luiten, C. Nyakas, and J. Traber (1990) Corticalprojection patterns of the medial septum-diagonal band complex. J.Comp. Neurol. 293:103–124.
Gibbs, R.B., and D.W. Pfaff (1994) In situ hybridization detection of trkAmRNA in brain: Distribution, colocalization with p75NGFR and up-regulation by nerve growth factor. J. Comp. Neurol. 341:324–339.
Gulyas, A.I., L. Seress, K. Toth, L. Acsady, M. Antal, and T.F. Freund(1991a) Septal GABAergic neurons innervate inhibitory interneuronsin the hippocampus of the macaque monkey. Neuroscience 41:381–390.
Gulyas, A.I., K. Toth, P. Danos, and T.F. Freund (1991b) Subpopulations ofGABAergic neurons containing parvalbumin, calbindin D28k, andcholecystokinin in the rat hippocampus. J. Comp. Neurol. 312:371–378.
Gundersen, H.J.G. (1986) Stereology of arbitrary particles. A review ofunbiased number and size estimators and the presentation of some newones, in memory of William R. Thompson. J. Microsc. 143:3–45.
Hage, B., M. Frotscher, and T. Naumann (1996) Activity of cholineacetyltransferase in the rat medial septal nucleus following fimbria–fornix transection or selective immunolesioning with 192 IgG-saporin.Neurosci. Lett. 205:119–122.
Halasy, K., and P. Somogyi (1993) Subdivisions in the multiple GABAergicinnervation of granule cells in the dentate gyrus of the rat hippocam-pus. Eur. J. Neurosci. 5:411–429.
Hama, A.T., J. Sagen, and G.D. Pappas (1996) Morphological characteriza-tion of dorsal horn spinal neurons with unilateral constriction nerveinjury: a preliminary study. Neurol. Res. 16:297–304.
Heckers, S., T. Ohtake, R.G. Wiley, D.A. Lappi, C. Geula, and M.-M.Mesulam (1994) Complete and selective cholinergic denervation of ratneocortex and hippocampus but not amygdala by an immunotoxinagainst the p75 NGF receptor. J. Neurosci. 14:1271–1289.
Hedreen, J.C., S.J. Bacon, and D.L. Price (1985) A modified histochemicaltechnique to visual acetylcholinesterase-containing axons. J. Histo-chem. Cytochem. 33:134–140.
Holley, L.A., R.G. Wiley, D.A. Lappi, and M. Sarter (1994) Cortical cholinergicdeafferentation following the intracortical infusion of 192 IgG-saporin:A quantitative histochemical study. Brain Res. 663:277–286.
Hortnagl,H.,G. Sperk,G.Sobal, andD.Maas (1990)Cholinergic deficit inducedby ethylcholine aziridinium (AF64A) transiently affects somatostatinand neuropeptide Y levels in rat brain. J. Neurochem. 54:1608–1613.
Houser, C.R., and M. Esclapez (1994) Localization of mRNAs encoding twoforms of glutamic acid decarboxylase in the rat hippocampal formation.Hippocampus 4:530–545.
Johnson, E.M., M. Taniuchi, H.B. Clark, J.E. Springer, S. Koh, M.W.Tayrien, and R. Loy (1987) Demonstration of the retrograde transport ofnerve growth factor receptor in the peripheral and central nervoussystem. J. Neurosci. 7:923–929.
Jouvenceau, A., J.-M. Billard, R.G. Wiley, Y. Lamour, and P. Dutar (1994)Cholinergic denervation of the rat hippocampus by 192-IgG-saporin:Electrophysiological evidence. Neuroreport 5:1781–1784.
Kohler, C., V. Chan-Palay, and J.Y. Wu (1984) Septal neurons containingglutamic acid decarboxylase immunoreactivity project to the hippocam-pal region in the rat brain. Anat. Embryol. (Berl.) 169:41–44.
Kohler, C., L. Eriksson, S. Davies, and V. Chan-Palay (1987) Colocalization ofneuropeptide tyrosine and somatostatin immunoreactivity in neurons ofindividual subfields of the rat hippocampal region. Neurosci. Lett. 78:1–6.
Koliatsos, V.E., D.L. Price, G.K. Gouras, M.H. Cayouette, L.E. Burton, andJ.W. Winslow (1994) Highly selective effects of nerve growth factor,brain-derived neurotrophic factor, and neurotrophin-3 on intact andinjured basal forebrain magnocellular neurons. J. Comp. Neurol.343:247–262.
58 T.A. MILNER ET AL.

Kosaka, T., J.Y. Wu, and R. Benoit (1988) GABAergic neurons containingsomatostatin-like immunoreactivity in the rat hippocampus and den-tate gyrus. Exp. Brain Res. 71:388–398.
Leanza, G., O.G. Nilsson, R.G. Wiley, and A. Bjorklund (1995) Selectivelesioning of the basal forebrain cholinergic system by intraventricular192 IgG-saporin: Behavioural, biochemical and stereological studies inthe rat. Eur. J. Neurosci. 7:329–343.
Leanza, G., O.G. Nilsson, G. Nikkhah, R.G. Wiley, and A. Bjorklund (1996)Effects of neonatal lesions of the basal forebrain cholinergic system by192 immunoglobulin G-saporin: Biochemical, behavioural and morpho-logical characterization. Neuroscience 74:119–141.
Lee, M.G., J.J. Chrobak, A. Sik, R.G. Wiley, and G. Buzsaki (1994)Hippocampal theta activity following selective lesion of the septalcholinergic system. Neuroscience 62:1033–1047.
Leranth, C. and M. Frotscher (1987) Cholinergic innervation of hippocam-pal GAD- and somatostatin-immunoreactive commissural neurons. J.Comp. Neurol. 261:33–47.
Lowenstein, D.H., M.J. Thomas, D.H. Smith, and T.K. McIntosh (1992)Selective vulnerability of dentate hilar neurons following traumaticbrain injury: A potential mechanistic link between head trauma anddisorders of the hippocampus. J. Neurosci. 12:4846–4853.
Lynch, G., G. Rose, C. Gall , and C.W. Cotman (1975) The response of thedentate gyrus to partial deafferentation. In M. Santini (ed): Golgi Centen-nial Symposium Proceedings. NewYork: Raven Press, pp. 305–317.
Mathern, G.W., T.L. Babb, J.K. Pretorius, and J.P. Leite (1995) Reactivesynaptogenesis and neuron densities for neuropeptide Y, somatostatin,and glutamate decarboxylase immunoreactivity in the epileptogenichuman fascia dentata. J. Neurosci. 15:3990–4004.
Milner, T.A., and C.E. Bacon (1989a) Ultrastructural localization of somato-statin-like immunoreactivity in the rat dentate gyrus. J. Comp. Neurol.290:544–560.
Milner, T.A., and C.E. Bacon (1989b) GABA-ergic neurons in the rathippocampal formation: Ultrastructure and synaptic relationships withcatecholaminergic terminals. J. Neurosci. 9:3410–3427.
Milner, T.A., and E. Veznedaroglu (1992) Ultrastructural localization ofneuropeptide Y-like immunoreactivity in the rat hippocampal forma-tion. Hippocampus 2:107–126.
Milner, T.A., and E. Veznedaroglu (1993) Septal efferent axon terminalsidentified by anterograde degeneration show multiple sites for modula-tion of neuropeptide Y-containing neurons in the rat dentate gyrus.Synapse 14:101–112.
Milner, T.A., R. Loy, and D.G. Amaral (1983) An anatomical study of thedevelopment of the septo-hippocampal projection in the rat. Dev. BrainRes. 8:343–371.
Mufson, E.J., M. Bothwell, L.B. Hersh, and J.H. Kordower (1989) Nervegrowth factor receptor immunoreactive profiles in the normal, agedhuman basal forebrain: Colocalization with cholinergic neurons. J.Comp. Neurol. 285:196–217.
Nilsson, O.G., G. Leanza, C. Rosenblad, D.A. Lappi, R.G. Wiley, and A.Bjorklund (1992) Spatial learning impairments in rats with selectiveimmunolesion of the forebrain cholinergic system. Neuroreport 3:1005–1008.
Patton, P.E., and B.McNaughton (1995) Connectionmatrix of the hippocam-pal formation. 1. The dentate gyrus. Hippocampus 5:245–286.
Petitto, J.M., and Z. Huang (1994) Molecular cloning of a partial cDNA ofthe interleukin-2 receptor-b in normal mouse brain: In situ localizationin the hippocampus and expression by neuroblastoma cells. Brain Res.650:140–145.
Pioro, E.P., and A.C. Cuello (1990) Distribution of nerve growth factorreceptor-like immunoreactivity in the adult rat central nervous system.Effect of colchicine and correlation with the cholinergic system—I.Forebrain. Neuroscience 34:57–87.
Poduri, A., L.L. Beason-Held, M.B. Moss, D.L. Rosene, and B.T. Hyman(1995) CA3 neuronal degeneration follows chronic entorhinal cortexlesions. Neurosci. Lett. 197:1–4.
Ribak, C.E., J.E. Vaughn, andK. Saito (1978) Immunocytochemical localiza-tion of glutamic acid decarboxylase in neuronal somata followingcolchicine inhibition of axonal transport. Brain Res. 140:315–332.
Ro¬ner, S., W. Hartig, R. Schliebs, G. Bruckner, K. Brauer, J.R. Perez-Polo,R.G. Wiley, and V. Bigl (1995a) 192IgG-saporin immunotoxin-inducedloss of cholinergic cells differentially activates microglia in rat basalforebrain nuclei. J. Neurosci. Res. 41:335–346.
Ro¬ner, S., R. Schliebs, W. Hartig, and V. Bigl (1995b) 192lgG-saporin-induced selective lesion of cholinergic basal forebrain system: Neuro-chemical effects on cholinergic neurotransmission in rat cerebral cortexand hippocampus. Brain Res. Bull. 38:371–381.
Schwarzer, C., J.M. Williamson, E.W. Lothman, A. Vezzani, and G. Sperk(1995) Somatostatin, neuropeptide Y, neurokinin B and cholecystokininimmunoreactivity in two chronic models of temporal lobe epilepsy.Neuroscience 69:831–845.
Seiler, M., and M.E. Schwab (1984) Specific retrograde transport of nervegrowth factor (NGF) from neocortex to nucleus basalis in the rat. BrainRes. 300:33–39.
Senut, M.C., D. Menetry, and Y. Lamour (1989) Cholinergic and peptidergicprojections from the medial septum and the nucleus of the diagonalband of Broca to dorsal hippocampus, cingulate cortex and olfactorybulb: A combined wheatgerm agglutinin-apohorseradish peroxidase-gold immunohistochemical study. Neuroscience 30:385–404.
Sloviter, R.S. (1989a) Chemically defined hippocampal interneurons andtheir possible relationship to seizure mechanisms. In V. Chan-Palayand C. Kohler (eds): The Hippocampus—New Vistas. NewYork:Alan R.Liss, pp. 443–461.
Sloviter, R.S. (1989b) Calcium-binding protein (calbindin-D28K) and parv-albumin immunocytochemistry: Localization in the rat hippocampuswith special reference to the selective vulnerability of hippocampalneurons to seizure activity. J. Comp. Neurol. 280:183–196.
Sloviter, R.S., and G. Nilavier (1987) Immunocytochemical localization ofGABA, cholecystokinin-, vasoactive intestinal polypeptide-, and somato-statin-like immunoreactivity in the area dentata and hippocampus ofthe rat. J. Comp. Neurol. 256:42–60.
Somogyi, P., A.J. Hodgson, A.D. Smith, M.G. Nunzi, A. Gorio, and J.Y. Wu(1984) Different populations of GABAergic neurons in the visual cortexand hippocampus of the cat contain somatostatin-or cholecystokinin-like immunoreactivity. J. Neurosci. 4:2590–2603.
Steckler, T.,A.B. Keith, R.G.Wiley, andA. Sahgal (1995) Cholinergic lesionsby 192 IgG-saporin and short-term recognition memory: Role of theseptohippocampal projection. Neuroscience 66:101–114.
Swanson, L.W., and W.M. Cowan (1979) The connections of the septalregion in the rat. J. Comp. Neurol. 186:621–656.
Tonder, N., J. Kragh, B.R. Finsen, T.G. Bolwig, and J. Zimmer (1994)Kindling induces transient changes in neuronal expression of somato-statin, neuropeptide Y, and calbindin in adult rat hippocampus andfascia dentata. Epilepsia 35:1299–1308.
Torres, E.M., T.A. Perry, A. Bjorklund, L.S. Wilkinson, R.G. Wiley, and S.B.Dunnett (1994) Behavioural, histochemical and biochemical conse-quences of selective immunolesions in discrete regions of the basalforebrain cholinergic system. Neuroscience 63:95–122.
Wainer, B.H., A.I. Levey, D.B. Rye, M.M. Mesulam, and E.J. Mufson (1985)Cholinergic and non cholinergic septohippocampal pathways. Neurosci.Lett. 54:45–52.
Waite, J.J., M.L. Wardlow, A.C. Chen, D.A. Lappi, R.G. Wiley, and L.J. Thal(1994) Time course of cholinergic and monoaminergic changes in rat brainafter immunolesioning with 192 IgG-saporin. Neurosci. Lett. 169:154–158.
Wenk, G.L., J.D. Stoehr, G. Quintana, R.G. Wiley, and S. Mobley (1994)Behavioral, biochemical, histochemical and electrophysiological effectsof 192-IgG-saporin injections into the basal forebrain of rats. J.Neurosci. 14:5986–5995.
Wiley, R.G. (1992) Neural lesioning with ribosome-inactivating proteins:Suicide transport and immunolesioning. Trends Neurosci. 15:285–290.
Wiley, R.G., and D.A. Lappi (1994) Suicide Transport and Immunolesion-ing. Austin, TX: Lawdes Medical Publishers.
Wiley, R.G., T.G. Berbos, T.L. Deckwerth, E.M. Johnson, Jr., and D.A. Lappi(1995) Destruction of the cholinergic basal forebrain using immuno-toxin to rat NGF receptor: Modeling the cholinergic degeneration ofAlzheimer’s disease. J. Neurol. Sci. 128:157–166.
Wiley, R.G., T.N. Oeltmann, and D.A. Lappi (1991) Immunolesioning:Selective destruction of neurons using immunotoxin to rat NGF recep-tor. Brain Res. 562:149–153.
Woolf, N.J., M.C. Hernit, and L.L. Butcher (1986) Cholinergic and non-cholinergic projections from the rat basal forebrain revealed by com-bined choline acetyltransferase and Phaseolus vulgaris leuccoaglutininimmunohistochemistry. Neurosci. Lett. 66:281–286.
Woolf, N.J., E. Gould, and L.L. Butcher (1989) Nerve growth factor receptoris associated with cholinergic neurons of the basal forebrain but notpontomesencephalon. Neuroscience 30:143–152.
Yamano, M., and P.G.M. Luiten (1989) Direct synaptic contacts of medialseptal efferents with somatostatin immunoreactive neurons in the rathippocampus. Brain Res. Bull. 22:993–1001.
Zhang, Z.J., T.G. Berbos, C.C. Wrenn, and R.G.Wiley (1996) Loss of nucleusbasalis magnocellularis, but not septal, cholinergic neurons correlateswith passive avoidance impairment in rats treated with 192-saporin.Neurosci. Lett. 203:214–218.
CHANGES IN HIPPOCAMPAL NPY NEURONS 59



![Neurotransmitter and neuropeptide regulation of mast cell ......triene B4 or TNF-α secretion, and nicotinic acetylcholine receptor antagonists blocked cholinergic stimulation [18]](https://img.pdfslide.net/doc/110x75/61297cd3ffa07a7e800de298/neurotransmitter-and-neuropeptide-regulation-of-mast-cell-triene-b4-or-tnf-.jpg)

























