Embed Size (px)
Citation preview

Journal of Ethology© Japan Ethological Society and Springer-Verlag 2003DOI 10.1007/s10164-003-0100-4
Article
Semilunar courtship rhythm of the fiddler crab Ucalactea in a habitat with great tidal variationTae Won Kim · Kil Won Kim · Robert B. Srygley · Jae C. Choe (✉)
T. W. Kim · K. W. Kim · R. B. Srygley · J. C. ChoeLaboratory of Behavior and Ecology, School of Biological Sciences, Seoul National University,Seoul 151–742, South Korea
✉ J. C. ChoePhone: +82-2-8808157Fax: +82-2-8827195E-mail: [email protected]
Received: 9 December 2002 / Accepted: 30 April 2003 / Published online: 26 July 2003
Abstract Semilunar courtship rhythm is a widely distributed phenomenon among fiddler crabs
in the genus Uca (Decapoda, Ocypodidae). Typically, synchronous courtship has been reported
to peak near spring tides. To determine whether a region of large tidal variation shifts reproductive
activity, we measured the frequency of specific courtship behaviors including claw-waving and
semidome building for U. lactea males on Kanghwa Island, Korea. We found that synchronized
courtship for U. lactea peaked near neap tides, whereas near the spring tides, seawater flooded
the habitat and males predominantly fed on the mudflat. Although active females, which hold their
burrows and usually feed on the mudflat, are abundant near to spring tides, males rarely claw-waved
to attract females. This pattern is atypical for the species because other populations of U. lactea
on Japan and Taiwan are synchronous around spring tides. We suggest that males invest most of
their time in feeding during spring tides because foraging is limited during neap tides. During neap
tides, males feed infrequently and thus expend stored energy on courtship signals. We conclude
that patterns of reproductive synchrony may be dependent on food availability in periodically
changing environments.
Keywords Fiddler crab · Semilunar rhythm · Synchronous courtship · Uca lactea · Waving
1

IntroductionMany marine animals show reproductive rhythms following the semilunar or lunar tidal cycle
(Palmer 1974; Ali 1992; Morgan and Christy 1994, 1995; Mizushima et al. 2000). Some fiddler
crabs in the genus Uca are also known to have synchronous reproductive cycles (Christy 1978,
1986; Morgan and Christy 1994, 1995). Reproductive synchrony in fiddler crabs constitutes male
courtship activity (Zucker 1978; Greenspan 1982; Salmon and Hyatt 1983; Salmon 1987), or the
synchronous release of larvae from the female which follows the semilunar tidal rhythm (Christy
1978, 1982; Morgan and Christy 1994, 1995; Kellmeyer and Salmon 2001).
Previous studies of male courtship in fiddler crabs suggested that male courtship behavior is
synchronized with female receptivity, which leads to simultaneous larval release (Christy 1978;
Greenspan 1982; Salmon and Hyatt 1983; Salmon 1987). Coordinating this reproductive rhythm
with the tidal cycle is assumed to be an adaptation that increases larval survival (Christy 1978,
1982; Morgan 1987; Morgan and Christy 1994, 1995; Kellmeyer and Salmon 2001). Exact causes
of synchronized courtship, however, still remain largely unknown. No studies have found clear
evidence that female receptivity synchronizes courtship.
Uca lactea has a particularly strong semilunar rhythm of activity (see Yamaguchi 1971; Crane
1975). For those populations studied to date (Murai et al. 1987; Severinghaus and Lin 1990;
Yamaguchi 2001a, 2001b, 2001c; Kim and Choe 2003), the mating behavior of U. lactea is
synchronous and peaks near to spring tide, a pattern which is consistent with other species of the
genus. However, courtship cycles may vary depending on the environmental conditions. We were
particularly interested in a population on Kanghwa Island, Korea, because the variation in the tides
is greater (tidal amplitude ranges from 4 to 10 m) than other sites where populations of Uca have
been studied (which are, in general, 50–200 cm). We predicted that a large tidal variation would
affect the courtship rhythm of the species because it may affect foraging opportunities. We found
that the courtship rhythm of the population in Kanghwa Island was different from other populations
of U. lactea. We discuss the environmental and behavioral factors that could lead to changes in
courtship rhythm within the species.
2

Materials and methods
Study area
Fieldwork was conducted at the intertidal mudflat in Kanghwa Island off the west coast of South
Korea (37°35′N, 126°32′E) during the breeding season from 2 July to 2 August 2000. Mean air
temperature during the observation period was 31.4°C (range: 28–38°C), and the mean surface
temperature was 31.6°C (range: 27–37°C). Mean relative humidity was 67% (range: 49–82%).
Monsoon rains fell on approximately 1 out of every 4 days (data obtained from the Kanghwa
meteorological observatory).
The study site was located within the intracoastal waterway (downstream of the Han River),
which is 1 km away from Daemyong port, in the Kimpo region (Fig. 1). Maximum tidal amplitude
was approximately 10 m at spring tides and minimum amplitude was approximately 4 m at neap
tides (Fig. 2A). Uca lactea lives on the upper intertidal mudflat, 700–850 cm above datum line,
covering 400–500 m2. The habitat is not inundated for 6–8 days around neap tides per semilunar
cycle.
Fig. 1. Map of the study area on Kanghwa Island
3
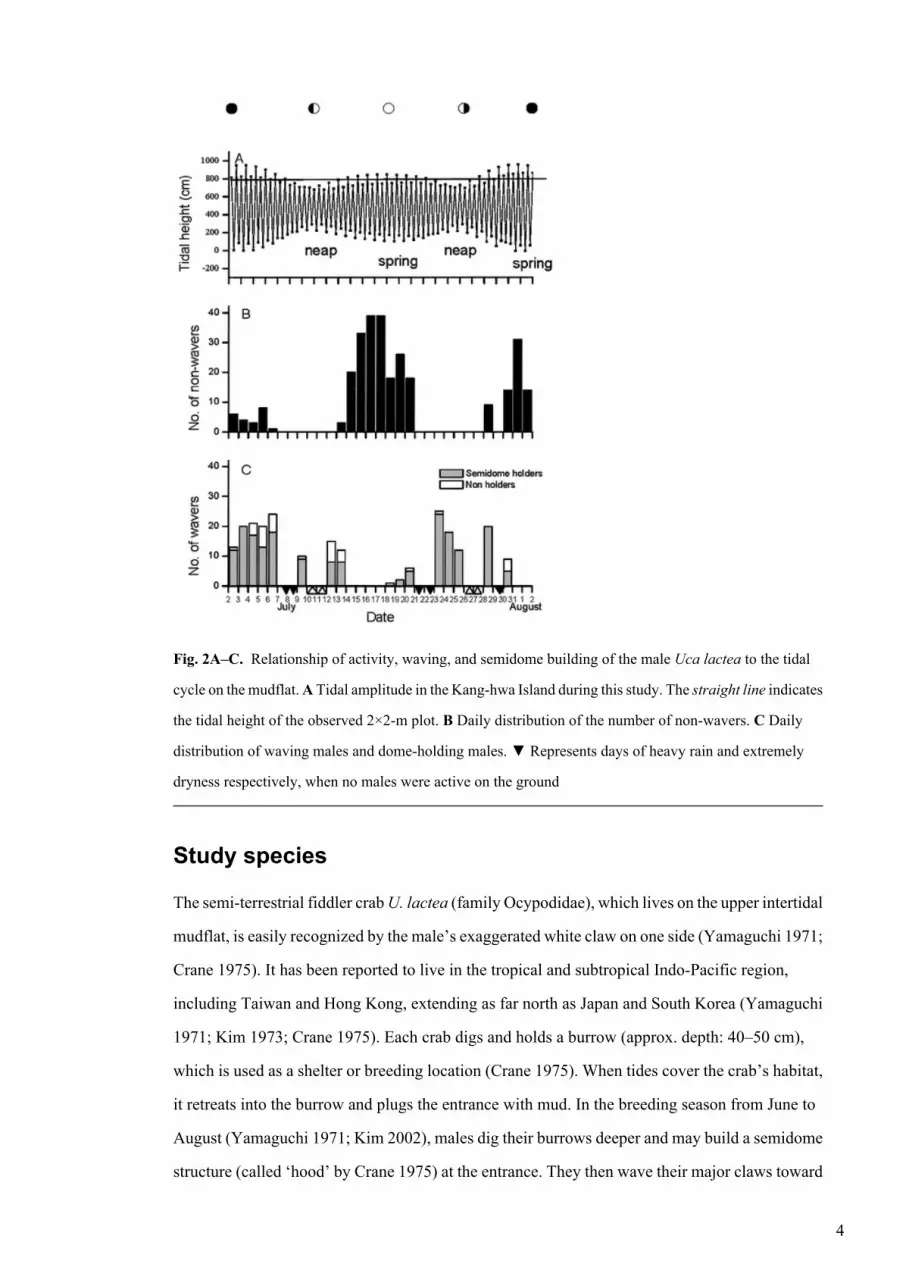
Fig. 2A–C. Relationship of activity, waving, and semidome building of the male Uca lactea to the tidal
cycle on the mudflat. A Tidal amplitude in the Kang-hwa Island during this study. The straight line indicates
the tidal height of the observed 2×2-m plot. B Daily distribution of the number of non-wavers. C Daily
distribution of waving males and dome-holding males. ▼ Represents days of heavy rain and extremely
dryness respectively, when no males were active on the ground
Study species
The semi-terrestrial fiddler crab U. lactea (family Ocypodidae), which lives on the upper intertidal
mudflat, is easily recognized by the male’s exaggerated white claw on one side (Yamaguchi 1971;
Crane 1975). It has been reported to live in the tropical and subtropical Indo-Pacific region,
including Taiwan and Hong Kong, extending as far north as Japan and South Korea (Yamaguchi
1971; Kim 1973; Crane 1975). Each crab digs and holds a burrow (approx. depth: 40–50 cm),
which is used as a shelter or breeding location (Crane 1975). When tides cover the crab’s habitat,
it retreats into the burrow and plugs the entrance with mud. In the breeding season from June to
August (Yamaguchi 1971; Kim 2002), males dig their burrows deeper and may build a semidome
structure (called ‘hood’ by Crane 1975) at the entrance. They then wave their major claws toward
4

wandering females (see Murai et al. 1987 for details). The females choose the mating partner by
sequentially entering (‘visiting’) the burrows of several males (Yamaguchi 1971; Kim 2002). The
crabs copulate in the male’s burrow or on the surface near the female’s burrow (Yamaguchi 1971,
2000b; Murai et al. 1987; Severinghaus and Lin 1990).
Observations of behavior
Prior to the observation period, we selected a focal area where U. lactea male burrows were at
relatively high density. We defined a 2 mx2 m plot with four 50-cm long PVC stakes connected
by fine nylon straps. Crabs’ burrows within the plot were numbered with flags made with colored
tape on sticks.
The crabs emerge from their burrows and are active on the surface for about 8 h each day during
the diurnal low tide. Hence, we recorded hourly behavioral patterns of the crabs for approximately
8 h after the tide receded from 2 July to 2 August. They were not active on the surface at night, in
heavy rain, nor on extremely dried sediment when the temperature was very high (Yamaguchi
1971; Crane 1975).
We recorded the behavior of each crab every hour while it was on the surface. We began with
the male at burrow number one, recorded its behavior at that moment, and subsequently recorded
the behavior of the male at the next burrow. Sample size did not exceed 50 males. Behavior was
categorized into seven classes (see Table 1). If the male was out of sight in the burrow, no data
were recorded until the next hour. We also noted the identity of the males that built semidomes.
To calculate the proportion of frequencies spent conducting each behavior, we pooled the behavioral
data for all of the males.
[Table 1. will appear here. See end of document.]
Each day, males active on the surface were categorized into three groups based on courtship
behaviors. Males that displayed claw waving at least twice during the eight daily observation
periods were called ‘wavers’ and the others ‘non-wavers.’ Wavers were divided into two additional
groups: those that had semidomes were called ‘semidome-holders’ and those that lacked a semidome
were ‘non-holders.’
The relationship between male behaviors and female activity
Each day at low tide, we counted the frequency of active females on the surface in the focal area,
most of which were burrow-holders and fed on sediment. As near to the same time as possible,
we recorded the frequencies of waving males and feeding males on the surface. In order to determine
5
whether male courtship activity or feeding activity was associated with the frequency of females,
we regressed the frequency of waving males and the frequency of feeding males on the number
of females.
ResultsMales active on the surface were more frequently observed around the spring tides than the neap
tides (Fig. 2B). Of the seven behavioral categories, males invested most of their time to waving
and feeding (Frequency of feeding and waving: 1140; Frequency of other behaviors: 131; X2=475.28,
df=6, P<0.0001; Fig. 3). On average, 52% of the observed males showed waving behavior more
than once a day and 48% did not wave at all on that particular day.
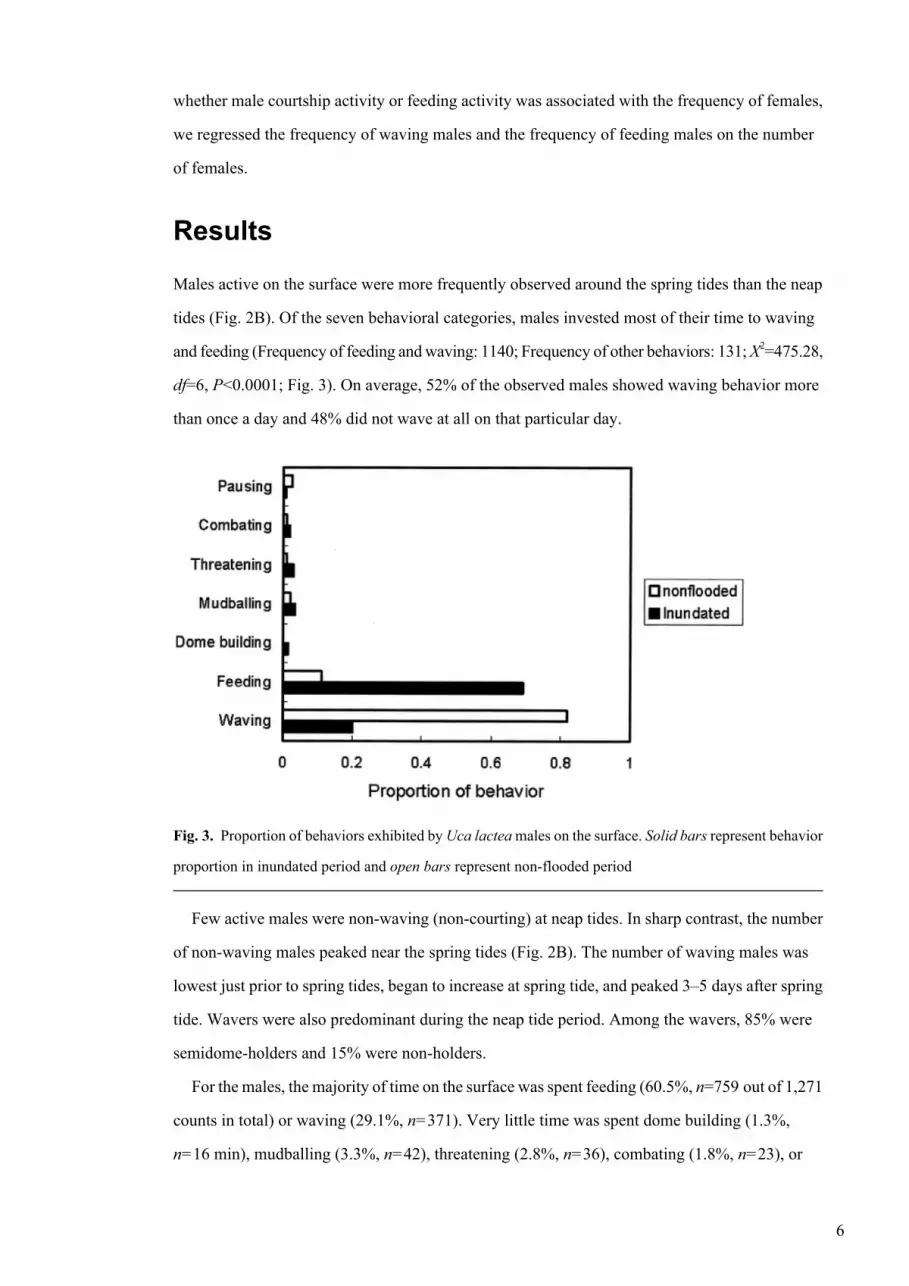
Fig. 3. Proportion of behaviors exhibited by Uca lactea males on the surface. Solid bars represent behavior
proportion in inundated period and open bars represent non-flooded period
Few active males were non-waving (non-courting) at neap tides. In sharp contrast, the number
of non-waving males peaked near the spring tides (Fig. 2B). The number of waving males was
lowest just prior to spring tides, began to increase at spring tide, and peaked 3–5 days after spring
tide. Wavers were also predominant during the neap tide period. Among the wavers, 85% were
semidome-holders and 15% were non-holders.
For the males, the majority of time on the surface was spent feeding (60.5%, n=759 out of 1,271
counts in total) or waving (29.1%, n=371). Very little time was spent dome building (1.3%,
n=16 min), mudballing (3.3%, n=42), threatening (2.8%, n=36), combating (1.8%, n=23), or
6
pausing (1.1%, n=14). The proportional frequency of seven types of behaviors differed significantly
between flooded and non-flooded phases (X2=310.56, df=6, P<0.0001) (Fig. 3). Especially, the
proportion of waving and feeding behavior was dramatically different between the two phases.
During inundated periods, most of the active males fed on the mudflat. On non-flooded days, most
males waved (Waving: X2=296.75, df=1, P<0.0001; feeding: X2=295.07, df=1, P<0.0001). Other
behaviors showed no significant differences following sequential Bonferroni’s correction (n=7
tests, including waving and feeding).
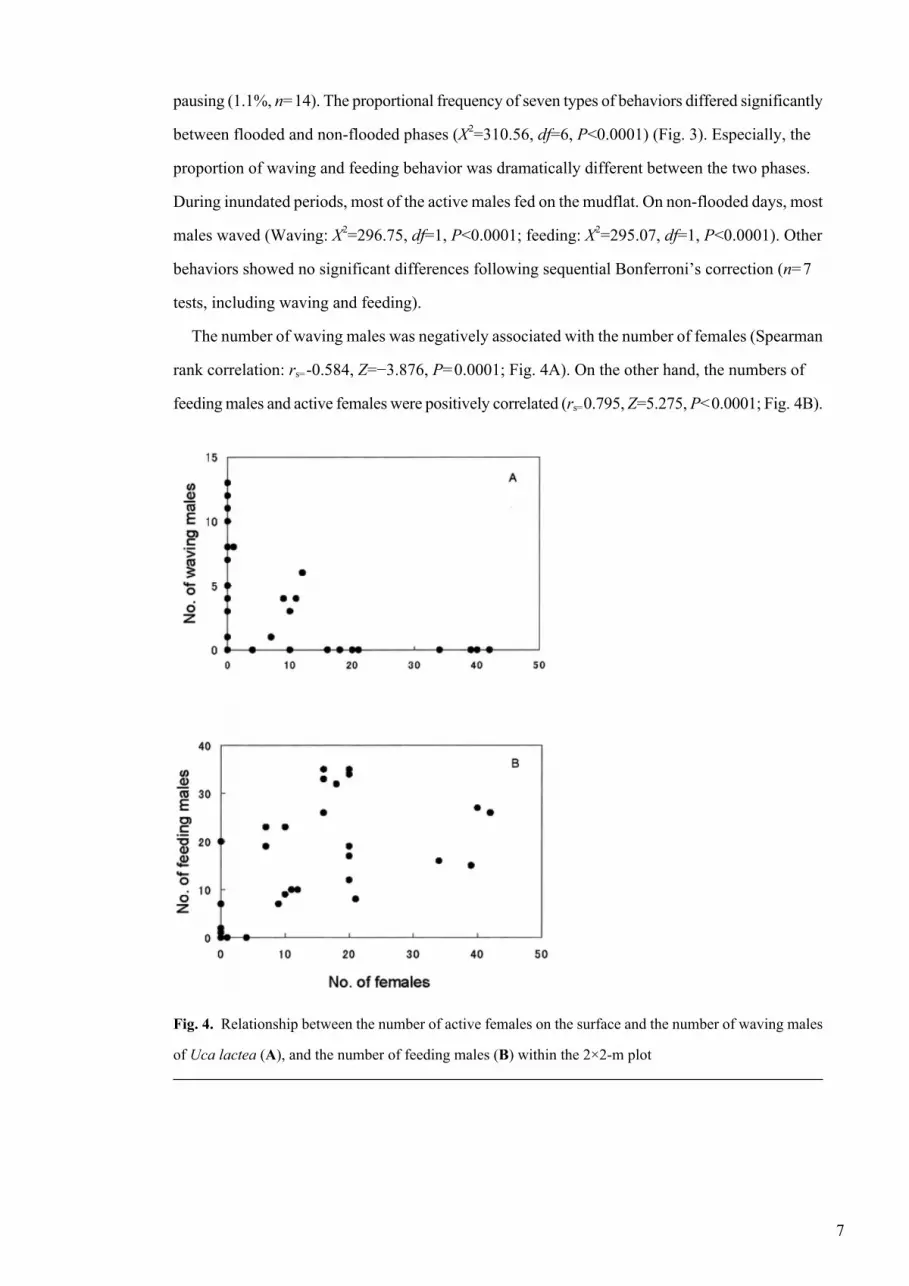
The number of waving males was negatively associated with the number of females (Spearman
rank correlation: rs=-0.584, Z=−3.876, P=0.0001; Fig. 4A). On the other hand, the numbers of
feeding males and active females were positively correlated (rs=0.795, Z=5.275, P<0.0001; Fig. 4B).
Fig. 4. Relationship between the number of active females on the surface and the number of waving males
of Uca lactea (A), and the number of feeding males (B) within the 2×2-m plot
7

DiscussionOn Kanghwa Island, the pattern of synchronized courtship timing of Uca lactea was different
from the courtship pattern of other fiddler crab species. Although male activity peaked at spring
tides, courtship displays such as waving and semidome building peaked at 4–5 days after spring
tides. In other species that show semilunar courtship rhythms, male courtship peaks around spring
tides (Crane 1958; Zucker 1978; Greenspan 1982; Salmon 1987; but see Christy 1978). This
general pattern has been interpreted as reproductive synchrony between males and females because
females should copulate about two weeks before spring tides in order to incubate larvae and release
them when the tidal stream is the fastest (Greenspan 1982; Salmon 1987).
Even for other populations of U. lactea courtship rhythm peaked near to spring tides. In a
Japanese population, male courtship peaked at 2–3 days before spring tides (Yamaguchi 1971,
2000b). In Taiwan, while the number of active individuals on the surface did not show a great
difference between the neap tides and spring tides, mating frequency peaked at spring tides, which
suggests that male courtship also peaked at these periods (Severinghaus and Lin 1990).
One reason for the discrepancy of behavioral rhythms in the Kanghwa Island population of U.
lactea may be due to the environmental differences between this habitat and that of the other
populations and other species. For example, the only other fiddler crab species known to have
interpopulational variation in courtship timing is U. pugilator (Christy 1978; Salmon and Hyatt
1983). In Florida, their courtship was reported to show peaks at neap tides, whereas other
populations peak at spring tides (Christy 1978). The explanation for the difference was that stream
velocity in Florida is higher in neap tides rather than in spring tides and females release larvae at
neap tides to facilitate their escape from predators (Salmon and Hyatt 1983). At Kanghwa Island,
however, stream velocity is higher at spring tides relative to neap tides. Maximum velocity is
approx. 230 cm/s at spring tides and 50 cm/s at neap tides (data from National Oceanographic
Research Institute of Korea).
The most probable cause of courtship near neap tides in the extreme tidal range at our study
site appears to be the relationship between tidal flooding and food availability. Tides are the
principal transport mechanism of food such as diatoms and other organic particles for the fiddler
crabs. Furthermore, crabs feed on the water-containing sediment because wet conditions facilitate
filtering organic materials from sediment (J. Christy, personsal communication; Reinsel and
Rittschof 1995). In our study site, the habitat of U. lactea is not flooded for 6–7 days of the
semilunar tidal cycle. In these periods, the drying of the habitat caused U. lactea to be food deprived.
8

Other populations of U. lactea probably suffer less from food deprivation. In Japan, the habitat
of U. lactea in Amakusa-Masushima (Yamaguchi 1971) was flooded almost everyday by tides
(M. Murai, personal communication). Information on the habitat in Taiwan was not available, but
there is no evidence that the habitat of the crabs is subjected to drying up, because the crabs are
generally active on the surface without interruption (Severinghaus and Lin 1990).
Food availability may thereby influence the timing of male courtship. Due to the limitation of
food supply around neap tides in the habitat, fewer males were active on the surface and active
males were not feeding. Feeding was most predominant in the few days before the spring tide.
This is the period when other species are typically observed to be courting. U. lactea probably
delays courtship toward neap tides in order to get sufficient energy during spring tides.
At the beginning of our study period in early July, many males waved their claws and relatively
few fed, although a peak in the spring tide had just occurred. The behavior of the crabs appears
to be contrary to our hypothesis that the crabs must feed when tides flood the habitat during spring
tides and court when the habitat is dry during neap tides. Two hypotheses might explain this
apparent contradiction. First, during the rainy season the study site is moistened by rains, enabling
the males to feed even at neap tides and court at spring tides. However, although rainfall provides
wet conditions for crabs to feed, it cannot provide crabs with fresh food because they only feed
on organic matter that the tide deposits on the mudflat. Only flooding can provide food by a process
of suspension and accumulation. Indeed, rainfall may delay courtship rhythm by interrupting males
from feeding (T.W. Kim, personal observation). Second, a large tidal amplitude may accumulate
more organic matter on the mudflat due to the higher floods. In early July, the spring tide was in
the new moon phase, which is higher than the spring tide in the full moon phase (see Fig. 2A).
Females as well as male crabs could begin to feed earlier in a new moon spring tide, encounter
more organic particulate, and obtain sufficient energy to begin reproduction earlier. The data
indicate that males can even court in the latter portion of the spring tide.
In a related study, we tested whether food availability influenced courtship timing directly by
conducting food addition and removal experiments. We found that an increase in food availability
advanced the timing of male courtship and increased the courtship intensity, whereas food
deprivation had the opposite effects (Kim and Choe 2003). Hence, U. lactea males exhibit flexibility
in the expression of mating behaviors in response to food availability.
Near neap tides when males were courting, female crabs sampled burrows occupied by males,
and presumably assessed the male in each burrow. If a female stayed in the burrow for a long time
(e.g., >15 min), the male came out of the burrow and the lone female plugged the burrow entrance
9

presumably to oviposit. Then the male who lost his burrow wandered to find new burrows (T.W.
Kim, personal observations). We are not sure if mating occurred in the male’s burrow because
females can use stored sperm obtained by surface mating to fertilize eggs (e.g., Koga et al. 2000).
As a result of the females’ lone activities in the burrows, however, during male courtship activities,
a male biased sex ratio was evident on the surface. Waving males were most active when females
were least active on the surface, whereas males were feeding when females were most active and
also feeding.
In contrast, surface mating was more prevalent during spring tides. Males that mated on the
surface neither built semidomes nor waved to females (n=16 out of 17 males). Most importantly,
females mated with additional males after mating on the surface (T.W. Kim, personal observation).
Hence during spring tides, many females may not be receptive and even those that mated may not
have been fertile (Murai et al. 1987). As a result, males might not display courtship signals.
A corollary to the hypothesis that food availability influences male courtship activity is that it
also influences female reproductive receptivity. Then perhaps it is the females’ receptivity that
triggers male courtship signals. As evidence to the contrary, the periodicity for female U. lactea
reproduction in Japan was a lunar cycle, whereas that for the reproductive behavior of males was
semilunar (Yamaguchi 2001a). Therefore, we propose that factors other than female receptivity
and the female’s response to food availability are influencing male reproductive behavior. Without
experimental studies, it is difficult to determine which sex first attracts its mates. In this regard,
we need to study how reproductive synchrony may be linked to various environmental factors
such as food availability (Kim and Choe 2003), predation pressure (Christy 1978), male-male
competition (Zucker 1984), and temperatures (Henmi 1989).
Acknowledgements We are very thankful to John H. Christy at the Smithsonian Tropical Research
Institute and Minoru Murai at the University of Ryukus and two anonymous referees for valuable
comments. Delali Dovie improved this paper with valuable discussion and Michiko Sato provided
help getting information in Japan. We also thank Sanha Kim, Youna Hwang, Ho Young Suk and
other members of the Laboratory of Behavior and Ecology of Seoul National University for many
helpful discussions on the results. We thank the National Oceanographic Research Institute of
Korea for providing meteorological and tidal data cited in this study. The Korean Federation of
Science and Technology Societies supported R.B.S. with a Brain Pool Professorship. This study
was supported by the Brain Korea 21 Research Fellowship from the Ministry of Education and
Human Resources Development.
10

ReferencesAli MA (1992) Rhythms in fishes. NATO ASI series A vol 236. Plenum Press, New York
Christy JH (1978) Adaptive significance of reproductive cycles in the fiddler crab Uca pugilator: a hypothesis.Science 199:453–455
Christy JH (1982) Adaptive significance of semilunar cycles of larval release in fiddler crabs (genus Uca):test of a hypothesis. Biol Bull 163:251–263
Christy JH (1986) Timing of larval release by intertidal crabs on an exposed shore. Bull Mar Sci 39:176–191
Crane J (1958) Aspects of social behavior in fiddler crabs, with special reference to Uca maracoani (Latreille).Zoologica (NY) 43:113-130
Crane J (1975) Fiddler crabs of the world. Princeton University Press, Princeton, N.J.
Greenspan BN (1982) Semi-monthly reproductive cycles in male and female fiddler crabs, Uca pugnax.Anim Behav 30:1084–1092
Henmi Y (1989) Reproductive ecology of three ocypodid crabs II. Incubation sites and egg mortality. EcolRes 4:261–269
Kellmeyer K, Salmon M (2001) Hatching rhythms of Uca thayeri Rathbun: timing in semidiurnal and mixedtidal regimes. J Exp Mar Biol Ecol 260:169–183
Kim HS (1973) Illustrated encyclopedia of fauna and flora of Korea, vol 14: Anomura Brachyura. KoreanMinistry of Education, Seoul
Kim TW (2002) Courtship and male-male competition in the fiddler crab Uca lactea. MSc thesis, SeoulNational University, Seoul
Kim TW, Choe JC (2003) The effect of food availability on the semilunar courtship rhythm in the fiddlercrab Uca lactea. Behav Ecol Sociobiol. DOI 10.1007/s00265-003-0614-3
Koga T, Murai M, Goshima S, Poovachiranon S (2000) Underground mating in the fiddler crab Ucatetragonon : the association between female life history traits and male mating tactics. J Exp Mar Biol Ecol248:35–52
Mizushima N, Nakashima Y, Kuwamura T (2000) Semilunar spawning cycle of the humbug damselfishDascyllus aruanus. J Ethol 18:105–108
Morgan SG (1987) Adaptive significance of hatching rhythms and dispersal patterns of estuarine crab larvae:avoidance of physiological stress by larval export? J Exp Mar Biol Ecol 113:71–78
Morgan SG, Christy JH (1994) Plasticity, constraint, and optimality in reproductive timing. Ecology75:2185–2203
Morgan SG, Christy JH (1995) Adaptive significance of the timing of larval release by crabs. Am Nat145:457–479
Murai M, Goshima S, Henmi Y (1987) Analysis of the mating system of the fiddler crab, Uca lactea. AnimBehav 35:1334–1342
Palmer JD (1974) Biological clocks in marine organisms. Wiley, London
Reinsel KA, Rittschof D (1995) Environmental regulation of foraging in the sand fiddler crab Uca pugilator(Bosc 1802) J Exp Mar Biol Ecol 187:269–287
Salmon M (1987) On the reproductive behavior of the fiddler crab Uca Thayeri, with comparisons to U.pugilator and U. vocans: evidence for behavioral convergence. J Crustac Biol 7:25–44
Salmon M, Hyatt GW (1983) Spatial and temporal aspects of reproduction in North Carolina fiddler crabs(Uca pugilator Bosc) J Exp Mar Biol Ecol 70:21–43
Severinghaus LL, Lin HC (1990) The reproductive behaviour and mate choice of the fiddler crab (Ucalactea lactea) in Mid-Taiwan. Behaviour 113:292–307
Yamaguchi T (1971) The courtship behavior of a fiddler crab, Uca lactea. Kumamoto J Sci Biol 10:13–37
Yamaguchi T (2001a) The breeding period of the fiddler crab, Uca lactea (Decapoda, Brachyura, Ocypodidae)in Japan. Crustaceana 74:285-293
Yamaguchi T (2001b) The mating system of the fiddler crab, Uca lactea (Decapoda, Brachyura, Ocypodidae)in Japan. Crustaceana 74:389–399
11

Yamaguchi T (2001c) Daytime larval release of the fiddler crab, Uca lactea (Decapoda, Brachyura,Ocypodidae). Crustaceana 74:545-555
Zucker N (1978) Monthly reproductive cycles in three sympatric hood-building tropical fiddler crabs (genusUca). Biol Bull 155:410–424
Zucker N (1984) Delayed courtship in the fiddler crab Uca musica terpsichores. Anim Behav 32:735–742
12

Tab
le 1. Behavior pattern of the male
Uca
lact
ea
Description
Category
Male stands on the surface motionless for more than several min.
Pausing
Male enters its burrow and returns to the surface carrying mudball and deposits it.
Mudballing
Male drags material from sediment with ambulatory legs and stacks it at his burrow to make a
semidome.
Dome building
Male takes bits of sediment with its small chela, gets them into the mouth filtering organic materials
and leaves small pellets on the surface.
Feeding
Male rhythmically extends laterally and folds its major claw toward females.
Waving
Male directs a major claw toward other crab without physical contact.
Threatening
Two males try to push each other backwards or fight with major claws interlocked.
Combating
13