Embed Size (px)
Citation preview

Agrin expresszió primer májtumorban
Doktori értekezés
Dr. Batmunkh Enkhjargal
Semmelweis Egyetem
Patológiai Tudományok Doktori Iskola
Témavezető: Dr. Kiss András, egyetemi docens
Hivatalos bírálók: Prof. Dr. Tímár József, egyetemi tanár
Dr. Abonyi Margit, egyetemi docens
Szigorlati bizottság elnöke: Prof. Dr. Jeney András, egyetemi tanár
Szigorlati bizottság tagjai: Prof. Dr. Szentirmay Zoltán, egyetemi tanár
Dr. Moldvay Judit, egyetemi docens
Budapest
2007

2
1.TARTALOMJEGYZÉK 1.TARTALOMJEGYZÉK ............................................................................................... 2 2. RÖVIDÍTÉSEK JEGYZÉKE....................................................................................... 4 3. BEVEZETÉS ÉS IRODALMI HÁTTÉR .................................................................... 6
3.1. Primer májtumorok................................................................................................ 6 3.1.1. Hepatocellularis carcinoma ............................................................................ 6 3.1.2. Cholangiocarcinoma....................................................................................... 9 3.1.3. Hepatocellularis adenoma ............................................................................ 10 3.1.4 Cholangiocellularis adenoma ........................................................................ 10 3.1.5. Fokális Noduláris Hyperplasia ..................................................................... 11
3.2. Az extracelluláris mátrix (ECM) ......................................................................... 12 3.2.1. ECM: szintézis, lebontás, szabályozás ......................................................... 13 3.2.2. Az ECM szerepe a szöveti működés szabályozásában................................. 14 3.2.3. Az ECM és a tumorprogresszió.................................................................... 15 3.2.4. Az ECM összetétele humán májszövetben................................................... 16 3.2.5. Proteoglikánok mint ECM komponensek .................................................... 16
3.2.5.1. A proteoglikánok szerkezete ................................................................. 17 3.2.5.2. A proteoglikánok osztályozása.............................................................. 19 3.2.5.3. Az agrin mint BMHSPG ....................................................................... 24
3.2.5.3.1. Az agrin szerkezete................................................................................. 24 3.2.5.3.2. Az agrin típusai....................................................................................... 25 3.2.5.3.3. Az agrin funkciói .................................................................................... 26
3.3. A proteoglikánok szerepe májbetegségekben...................................................... 29 3.4. A BMHSPG-k szerepe az angiogenesisben ........................................................ 33
4. CÉLKITŰZÉSEK....................................................................................................... 36 5. ANYAG ÉS MÓDSZER............................................................................................ 37
5.1. Beteganyag ......................................................................................................... 37 5.2. Módszerek ........................................................................................................... 38
5.2.1. Fehérjeexpressziós vizsgálatok .................................................................... 38 5.2.1.1. Immunhisztokémiai vizsgálatok............................................................ 38 5.2.1.2. Immunfluoreszcens vizsgálat ................................................................ 40 5.2.1.3. Morfometriai elemzés............................................................................ 40 5.2.1.4. Proteoglikán izolálása............................................................................ 41 5.2.1.5. Western blot analízis ............................................................................. 41
5.2.2. mRNS szintű expressziós vizsgálatok .......................................................... 42 5.2.2.1. RNS izolálás .......................................................................................... 42 5.2.2.2. Primertervezés és grádiens PCR............................................................ 43 5.2.2.3. mRNS expresszió vizsgálata real-time RT-PCR módszerrel ................ 45 5.2.2.4. Statisztikai kiértékelés ........................................................................... 46
6. EREDMÉNYEK......................................................................................................... 48 6.1. Agrin fehérje expresszió vizsgálata immunhisztokémiai módszerrel ................. 48
6.1.1. Agrin expresszió ép és cirrhotikus humán májszövetben............................. 48 6.1.2. Agrin expresszió hepatocellularis carcinomában ......................................... 48 6.1.3. Az agrin expressziója más kötőszöveti fehérjékkel kolokálizációban. ........ 50 6.1.4. Agrint termelő sejtek májelváltozásokban ................................................... 51 6.1.5. Az agrin expressziójának vizsgálata a cholangiocarcinomában................... 53 6.1.6. Az agrin expresszió vizsgálata hepatocellularis adenomában...................... 55

3
6.1.7. Az agrin expresszió vizsgálata fokális noduláris hyperplasiában ................ 56 6.2. Az agrin fehérje expresszió vizsgálata Western Blot technikával....................... 57 6.3. Az agrin mRNS kimutatása real time RT-PCR-vel............................................. 58
7. MEGBESZELÉS........................................................................................................ 60 8. KÖVETKEZTETÉSEK.............................................................................................. 66 9. ÖSSZEFOGLALÁS ................................................................................................... 68 10. SUMMARY ............................................................................................................. 70 11. IRODALOMJEGYZÉK ........................................................................................... 72 12. SAJÁT KÖZLEMÉNYEK JEGYZÉKE................................................................. 88 13. KÖSZÖNETNYILVÁNÍTÁS................................................................................. 93

4
2. RÖVIDÍTÉSEK JEGYZÉKE ABC avidin - biotin komplex
AchR acetylcholine receptor
AEC 3-amino-9 ethyl-carbazole
AFP alfa-fetoprotein
ATP adenozin-trifoszfát
BM bazális membrán
BMHSPG bazális membrán heparán szulfát proteoglikán
bFGF basic fibroblast growth faktor, bázikus fibroblaszt növekedési faktor
BSA bovine serum albumin, borjú szérum albumin
CC cholangiocarcinoma
CCA cholangiocelullaris adenoma
CK cytokeratin
CS kondroitinszulfát
CSPG kondroitinszulfát proteoglikán
DAB 3,3’-diaminobenzidin tetrahidroklorid
DSPG dermatánszulfát proteoglikán
ECL enhanced chemiluminescence, erősített kemilumineszcencia
ECM extracelulláris mátrix
EDTA ethylene -diamine-tetraacetic acid
EGF epidermal growth factor, epidermális növekedési faktor
EHS Engelbreth-Holm-Swarm
GAG glükózaminoglikán
GAPDH glyceraldehyde-3 phosphate dehydrogenase, glicerinaldehid-3-foszfát
dehidrogenáz
GBM gromeruláris bazális membrán
HBV hepatitis B vírus
HCA hepatocellularis adenoma
HCC Hepatocellularis carcinoma
HCV Hepatitis C vírus
HE hematoxilin –eozin
HS heparánszulfát

5
HSPG heparánszulfát proteoglikán
HRP horseradish peroxidase, tormaperoxidáz
IR idegrendszer
KSPG keratánszulfát proteoglikán
MuSK Muscle-specific tyrosine kinase, izom specifikus tirozin kináz
MMP matrix metalloproteinase, matrix metalloproteináz
NCAM neural-cell adhesion molecule, idegsejt adhéziós molekula
NEM N-etil- maleimid
NMJ neuromuszkuláris junkció
PBS phosphate-buffered saline, foszfát-pufferolt sóoldat
PCR polymerase chain reaction, polimeráz láncreakció
PDGF platelet-derived growth faktor, vérlemezke eredetű növekedési faktor
PMSF fenil-metil-szulfonil-fluorid
PG proteoglikán
PTK protein tirozin kináz
RT reverse transcriptase, reverz transzkriptáz
SCG superior cervical ganglion, felső cervikális ganglion
SD standard deviation, standard deviancia
SDS-PAGE sodium-dodecil-szulfát -poliakrilamid gél elektroforézis
SMA smooth muscle actin, simaizom aktin
TBE tris-borát-EDTA
TBS tris-buffered saline, tris-pufferolt sóoldat
TGF transforming growth factor, transzformáló növekedési faktor
TIMP tissue inhibitor of metalloprotease, metalloproteázok szöveti inhibítora
TNF tumor necrosis factor, tumor nekrózis faktor
VEGF vascular endothelial growth factor, vaszkuláris endotél eredetű növekedési faktor

6
3. BEVEZETÉS ÉS IRODALMI HÁTTÉR
3.1. Primer májtumorok
3.1.1. Hepatocellularis carcinoma
A daganatok okozta halálozások száma évente világszerte növekszik. Az utóbbi
15 év során a daganatos halálozások száma a férfiak esetében átlagosan 12-, a nők
esetében pedig 9 százalékkal emelkedett az Európai Unióban. A Nemzetközi Rákkutató
Intézet napokban közzétett tanulmánya szerint tavaly 39 európai országban mintegy 3,2
millió új daganatos megbetegedést diagnosztizáltak, ami 300. 000-rel több mint 2004-
ben volt. 2006-ban Európa-szerte 1,7 millióan haltak meg rákos megbetegedés
következtében. (http://www.aerzteblatt.de/v4/news/news.asp). A daganatos elhalálozás 21,6%-
át teszi ki az összes halálozásnak. A hepatocellularis carcinoma (HCC) a világszerte
előforduló rosszindulatú daganatoknak mintegy 4%-át teszi ki, férfiak között a hetedik,
nők között a kilencedik helyen áll gyakoriságban. Világviszonylatban körülbelül
félmillió új beteggel kell számolni évente (Bosch és mtsai, 2004).
Magyarországon a rosszindulatú daganatos megbetegedések halálozási aránya
rendkívül kedvezőtlen, a szív- és keringési rendszer-eredetű halálokok mögött a
második helyet foglalja el, kb. 25%-os gyakorisággal. A nagy európai, illetve
nemzetközi halálozási felmérésekben férfiaknál az első, nőknél pedig a második helyen
állunk (Otto és Kasler, 2005).
Az emésztőrendszer betegségeinek tulajdonítható halálozások gyakorisága a
világon a második legmagasabb mortalitással jellemzi Magyarországot. Szembetűnő
növekedés tapasztalható az elmúlt évtizedekben a krónikus májbetegségek, elsősorban
májzsugor és májrák okozta halálozásban. 100 ezer lakosra számítva 66 haláleset jut,
ami hétszerese az EU átlagnak (www.datanet.hu).
Az ázsiai országokban az európai adatokkal szemben a daganatok okozta halálok
között elsők között említendő a primer májrák. Ezen országok között különös helyet
foglal el Mongólia, ahol a HCC legnagyobb előfordulási aranyát dokumentálták,
100.000 lakosra 99 esetet (www-dep.iarc.fr). Az utóbbi évek adatai szerint itt a primer

7
májrák a malignus tumorok okozta halálozások mintegy 44%-át teszi ki (Oyunsuren és
mtsai, 2006).
Etiológia
A májdaganatok az utóbbi években növekvő számban kerülnek felismerésre.
Ennek oka részben az, hogy a detektálás módszerei érzékenyebbé válnak, elsősorban a
képalkotó eljárások fejlődése következtében. Másrészt viszont az emelkedés oka
összefügghet egyes etológiai tényezők, elsősorban a hepatitis C-vírus (HCV) és a
hepatitis B-vírus (HBV) fertőzés, az ipari mérgekkel és egyéb toxikus anyagokkal való
expozíció nagyobb arányával (Schaff és mtsai, 2004).
A krónikus vírusfertőzések gyakran vezetnek cirrhosishoz, ami önmaga is fontos
kockázati tényező a májrák kialakulásában (Kiss és mtsai, 2002).
A HBV és a HCC kapcsolatát azok a szeroepidemiológiai vizsgálatok vetették fel
először, amelyek igazolták a szoros összefüggést a HBV-infekció és a HCC
előfordulása között Ázsiában. A HCC előfordulása 100-szoros volt HBV- fertőzött
személyekben a HBV negatívakhoz képest (Beasley és mtsai, 1981).
Európában, ahol a HBV nem hiperendémias, mint Ázsiában, a HCV
előfordulását sokkal gyakoribbnak találták HCC-ben, mint a HBV jelenlétet (el Serag,
2001). A HCV-infectiót követően a HCC kifejlődésére kb. 25-30 év elteltével lehet
számítani.
Bármilyen okból eredő krónikus májsejtkárosodás és a kísérő regeneráció
kapcsán a mitózisok a DNS-t fogékonnyá teszik a genetikai alterációkra. Ezért a
krónikus májbetegség más formái is kockázati tényezőt jelentenek májrákra, és e
tekintetben az alkoholos májbetegség, egyéb toxikus tényezők, valamint
anyagcserezavarok, így az α1-antitrypsin hiány, a haemochromatosis és a tyrosinaemia
jön szóba.
Afrikában és Dél-Kínában az Aspergillus flavusból és Aspergillus parasiticusból
származó aflatoxin nagy közegészségügyi probléma (Henry és mtsai, 2002). Ez a
mycotoxin potenciálisan carcinogen, a májrák gyakori előfordulásában ugyancsak
jelentős, mégpedig azáltal, hogy a tumorszuppresszor gén p53 259-es kodonjában
specifikus mutációt indukál (Smela és mtsai, 2001).

8
Hormonális tényezőkre utal a HCC gyakoribb előfordulása férfiakban, valamint
az a tény, hogy a hosszan tartó androgen terápiával is kapcsolatos a tumor előfordulása.
Egyébként orális anticoncipiensek, tórium-dioxid és vinil-klorid-expozíció is kockázati
tényezőnek tekinthető a májdaganatok keletkezésében.
A fent említett oki tényezők közül Mongóliában a vírusfertőzések és az
alkoholfogyasztás szerepe említendő (Oyunsuren és mtsai, 2006).
Pathologia
A HCC makroszkóposan szoliter, multiplex nodularis vagy diffúz formában
mutatkozhat (Schaff és mtsai, 2004). Az esetek közel 70%-ában cirrhosis talaján alakul
ki. A tumor nagyságától függően vérzés, necrosis gyakori. A kisebb méretű HCC-k
tokkal rendelkezhetnek. A progresszió mértékét a tokinfiltráció, a fiókdaganatok
jelenléte és az érinvázió jelzi.
Hisztológiailag trabecularis, acinaris (pseudoglandularis) és szolid (kompakt)
szerkezetű formákat különíthetünk el. Emellett speciális szöveti szerkezetű tumorok is
észlelhetőek, így a világossejtes, óriássejtes, eozinofil sejtes forma. Külön entitás a
fibrolamellaris típusú HCC. Ezen jellegzetes szöveti szerkezetű daganat cirrhosis
mentes májban, fiatalabb életkorban fordul elő, fokális noduláris hyperplasia (FNH)-hoz
hasonló centrális heg és jobb prognózis jellemzi.
A HCC TNM klasszifikációjának elve hasonló az egyéb tumorokéhoz. A T1-T3
kategóriák közötti differenciálást a tumorátmérő, a daganat szoliter vagy multiplex
volta, a lebenyek érintettsége határozza meg.
A HCC klinikai detektálásában a képalkotó módszerek mellett az alfa-
fetoprotein (AFP) meghatározás jelentős, bár bizonyos korlátokkal. Az AFP
hisztológiailag is detektálható, és fontos marker a HCC diagnosztikájában. Újabb
adatok alapján a HCC-k két jól elhatárolható csoportra oszthatók az AFP termelés
alapján (Lee és Thorgeirsson, 2002). Magasabban differenciált daganatokban az AFP
mRNS expressziója alacsonyabb, mint a differenciálatlanokban.
A HCC differenciáltsági fokozatai követik a I-IV beosztást, a magasan
differenciált formától a differenciálatlan formáig, mely egyes szerzők szerint a
prognózissal is összefügg (Nagy és mtsai, 2006).

9
3.1.2. Cholangiocarcinoma
Kiindulhat az epeutak bármely szakaszából, a kis epecapillarisoktól a közös
epevezetékig. A daganat elhelyezkedése alapján megkülönböztetünk intrahepatikus (5-
10%) és extrahepatikus epeúti tumorokat (20-30%), valamint az Altemeier és Klatskin
által leírt ún. hiliaris tumorokat (60-70%) (Klatskin-tumor) (Nakeeb és mtsai, 1996).A
cholangiocarcinomák (CC) több mint 90%-a adenocarcinoma, a sejtek egyszerű
duktularis elrendeződésével és fibrosus stromával.
Epidemiológiájára jellemző, hogy ritkábban fordul elő, mint a HCC és a primer
máj malignus tumorok 5-10%-át képviseli, kivéve a Távol-Keleten, ahol ez az arány
20% is lehet (Vauthey és Blumgart, 1994). Általában idősebbekben észlelhető, mint a
HCC, jellemzően a 6. és 7. évtizedben levő férfiakban (Shaib és el-Serag, 2004).
Az epeútrák kialakulásában különböző kémiai anyagok szerepéről számoltak be.
A tórium-dioxid (thorotrast), mint korábban jól ismert röntgen-kontrasztanyag, bár
ritkán, de oka lehet a CC-nek (Sahani és mtsai, 2003).
Epeúti paraziták szerepe is igazolódott. Távol-Keleten a Clonorchis sinensis,
hátsó-Indiában az Opisthorchis viverrini nevű intrahepaticus parazita szabadgyök-
termelés révén stimulálja a sejtproliferációt (Watanapa és Watanapa, 2002).
Colitis ulcerosában megnő a CC kockázata (10-szeres az átlag népességhez
viszonyítva), főleg azokban a betegekben, akiknek több mint 10 éves a kórlefolyása
(Broome és mtsai, 1996). Primer szklerotizáló cholangitisben is leírták a tumor
gyakoribb előfordulását (Shaib és el Serag, 2004). Az epekő szerepét az extrahepatikus
formák keletkezésében felvetették, de e téren az oki kapcsolat nem bizonyított (Su és
mtsai, 1997).
Makroszkóposan a CC szürkésfehér, tömött, szoliter vagy multiplex göb
formájában mutatkozik. Esetenként mucin termelés, nagyobb daganatokban centrális
necrosis észlelhető (Kopper L, Schaff Zs. 2004).
Az epeúti tumorok szövettana 90%-ban adenocarcinoma. Lehet papillaris, ill.
sclerosus, diffúzan terjedő. A papillaris típus a ritkább, de kevésbé agresszíven
viselkedő forma (Albores-Saavedra és mtsai, 2000). A tumor esetek kb. 35%-a
lokalizálódik a ductus choledochusra (Tompkins és mtsai, 1991).
Immunhisztokémiailag a daganatsejteket a CK7 pozitivitás jellemzi.

10
3.1.3. Hepatocellularis adenoma
A hepatocytákból kiinduló, ritka, jóindulatú daganat. Leggyakrabban fiatal
nőkben fordul elő; számos adat szól az orális fogamzásgátlókkal és az androgén
szteroidokkal való kapcsolat mellett (Trotter és Everson, 2001a). A hepatocellularis
adenoma (HCA) kialakulásának oka lehet a glikogéntárolási betegségek (Biecker és
mtsai, 2003). Általában nem cirrhoticus májban, egyesével alakul ki, ritkán multiplex.
Színe eltér a környezettől, halványabb, a felhalmozott zsír miatt többnyire sárgás,
homogén szerkezetű, de az érgazdagság miatt vérzésekkel tarkított lehet.
Mikroszkóposan szabályos, de az átlagos hepatocytáknál nagyobb sejtekből épül
fel, amelyek két-három sorban helyezkednek el, trabeculákat és/vagy tubulusokat
képezve. A tumorban nincsenek epeutak és portális mezők, vannak viszont
szinuszoidok és az ezeket bélelő Kupffer-sejtek. Néha cytoplasmaticus eperögök, illetve
canalicularis epedugaszok arra utalnak, hogy a májsejtekben megtörténik a bilirubin
konjugációja. Egyes esetekben a daganaton belül kifejezett érhálózat van. Ezek
vékonyabb-vastagabb falúak, bennük gyakoriak a frissebb és régebbi thrombusok. A
HCA nem tartalmaz epeutakat és ebben különbözik a fokális noduláris hyperplasiától
(Grazioli és mtsai, 2001). Biopsziás vizsgálattal többnyire biztonsággal
diagnosztizálható, kivéve a dysplasiával járó eseteket, melyek elkülönítése a magasan
differenciált HCC-től nehéz lehet (Schaff Z és Nagy P, 1997). A HCA legnagyobb
veszélye az esetleges malignus transzformáció lehetősége, mely különösen az
anyagcserezavarok talaján kialakult formákban gyakori.
3.1.4 Cholangiocellularis adenoma
Epeutak hámsejtjeiből indul ki. Szürkésfehér, tömött, szívós, szoliter vagy
multiplex, nem ritkán szubkapszulárisan elhelyezkedő daganat. Változó nagyságú lehet,
de többnyire 1 cm-nél kisebb. Szövettanilag a proliferáló epeutakra jellemző köb- vagy
hengerhámmal bélelt dúctusok láthatók, tömött kötőszövetbe ágyazva. A makroszkópos
képen adenocarcinoma metastasisával vagy cholangiocelluláris carcinomával
téveszthető össze. Ez különösen akkor merül fel, ha az 1 cm körüli tumor multiplex és a
tok alatt helyezkedik el.

11
3.1.5. Fokális Noduláris Hyperplasia
A daganatszerű elváltozások közé tartozik. A fokális noduláris hyperplasia
(FNH) gyakori jóindulatú fokális májelváltozás, az összes primer májtumor 8%-át teszi
ki (Biecker és mtsai, 2003a). A kialakult sejttömeg inkább reaktív elváltozás (a
cirrhosishoz hasonlóan) és nem rendelkezik a daganatok progresszív proliferációs
sajátosságával, valamint a daganatokra jellemző génalterációkkal, beleértve a p53
mutációt is (Schaff és mtsai, 1995). Általában szoliter, ritkábban multiplex; változó
nagyságú lehet, tok nem határolja. Jellegzetessége az igen kifejezett centrális, gyakran
csillag alakú heg a lézió központjában, amely az atípusos vaskos falú kanyargós ereket
tartalmazza. Az éranomáliák következtében a vérzés és ruptura gyakori. Szövettanilag a
hepatocytákra igen hasonlító sejtek trabeculákat képeznek, köztük portális traktusoknak
megfelelő képleteket láthatók. Az epeút proliferáció igen kifejezett (Hussain és mtsai,
2004). A nőkben gyakrabban fordul elő; orális contraceptivumok szedésével is
összefüggésbe hozzák, ám sokan tagadják ezt a kapcsolatot (Trotter és Everson, 2001).
A májtumor rizikócsoportok szűréséhez és a betegkövetéshez, a kiújulás és a
progresszió megelőzéséhez megfelelően érzékeny és specifikus markerekre van
szükség. Elsődleges fontosságú lenne a betegek kiszűrése minél korábbi stádiumban,
amikor a tumor a helyi kezelésre még jobban reagál. Emellett olyan szisztémás
kezelések kidolgozása is fontos cél, melyek hatékonyan gyógyítanák a betegeket
előrehaladott stádiumban, lehetővé téve ezáltal a máj funkciójának megtartását és
elkerülhetőek lennének a kiterjedt műtétek. A primer májtumorok kutatásának
elsődleges célja, hogy a különböző biológiai viselkedésű daganatok kialakulásának,
progressziójának genetikai hátterét megértsük és erre alapozva új, célzott terápiás
eljárások kerüljenek kidolgozásra.
A májtumorok patogenezisében a vírusfertőzések, illetve más etológiai tényezők
különböző elváltozásokon keresztül vezetnek a máj tumoros elváltozásához. Ez is, mint
általában a tumorgenesis, többlépcsős folyamat eredménye. Különböző
szignáltranszdukciós útvonalak aktivizálásán keresztül a stromának is jelentős szerep jut
a daganat progressziójában. A daganatos szövetté fejlődés egyik alapfeltétele a

12
daganatos stroma kialakítása, így az utóbbi időben egyre inkább előtérbe került az
extracelluláris mátrix kutatása, mely új megvilágításba helyezi a májban zajló
patológiás elváltozásokat, többek közt a fibrogenesis, a cirrhosis valamint a
carcinogenesis mechanizmusát.
3.2. Az extracelluláris mátrix (ECM)
Az ECM elválasztja a különböző szöveti részeket, biztosítja a sejtek
kapcsolódását és meghatározza a szöveti architektúrát. Az ECM, illetve annak egyes
komponensei nemcsak a szövetspecifikus szerkezet kialakításáért felelősek és az
intercelluláris kapcsolatok megteremtésében vesznek részt, hanem újabb megfigyelések
szerint közreműködnek a sejtek proliferációjában, differenciációjában, szövetspecifikus
működésében, életképességének fenntartásában, valamint a biokémiai és a celluláris
szerkezetet szabályozó jelátvitelben. Az ECM-sejt interakció eltérése több kóros állapot,
így a malignus transzformáció és progresszió jellemzője (Liotta, 1982). Az ECM
mennyisége, struktúrája szervenként, szövetenként változó. Hámszövetekben normális
körülmények között az ECM aránya nagyon alacsony, mindössze a sejtek bazális
felszíne alatt végighúzódó bazális laminára korlátozódik, azonban a mesenchymális
eredetű kötő- és támasztószövetekben – a sejtes elemekhez viszonyítva – nagy
mennyiségben van jelen.
Az ECM alkotórészek termelésére számos sejt képes, ezek közül a fibroblasztok,
oszteoblasztok, simaizomsejtek, endothelsejtek érdemelnek említést. Az ECM-et alkotó
fehérjék főbb csoportjai a strukturális fehérjék (elasztin, kollagén), adhéziós
tulajdonságokkal rendelkező fehérjék (fibronektin, tenascin), proteoglikánok és bazális
membrán fehérjék (kollagén IV, laminin, entactin) (Alberts és mtsai, 1989). Az ECM
szinte minden fehérjéje képes a sejtek biológiai viselkedésének a befolyásolására. Főleg
a proteoglikánok alkalmasak növekedési faktorok kötésére, tárolására, aktiválására és
inaktiválására, esetleg ko-receptorként felkínálni a tumorsejtek felszíni receptorai
számára.

13
3.2.1. ECM: szintézis, lebontás, szabályozás
Sokáig az ECM-et csak támasztó szöveti komponensnek tekintették és passzív
szerepet tulajdonítottak neki. Mai ismereteink szerint a stroma igen aktív
kölcsönhatások színtere, jeltovábbító rendszere.
A tumorok csupán növekedésük kezdeti szakaszában (az avaszkuláris fázisban)
épülnek fel csak daganatos sejtekből. A szövetté fejlődés alapfeltétele a daganatos
stroma kialakítása. A daganatos stroma nagymértékben eltér a normál szövet
stromájától. A daganatszövet ezt újonnan szintetizálja vagy termelteti a nem tumoros
mesenhymális eredetű sejtekkel (fibroblasztok, myofibroblasztok, histiocyták, endothel
és gyulladásos sejtek). Egyes kutatók szerint a stroma termelése a tumorsejtek genetikus
kontrollja alatt áll és a tumorprogresszió elősegítése a cél. Bizonyos változások
jellegzetesek lehetnek egy tumortípusra. Számos proteoglikán, illetve a cukorláncok
magas expressziója kedvez a tumorprogressziónak. Ezt az elképzelést támasztja alá az
az eredmény is, hogy a magas metasztázis-hajlamú tumorok más proteoglikán
expresszióval rendelkeznek, mint a nem metasztatizáló típusok (Timár és mtsai, 1995).
A mátrix-szintézis fő stimulátora a transforming growth faktor β1 (TGFβ1),
mely számos mátrixfehérje (fibronektin, I, III, IV típusú kollagén, elasztin) fokozott
termelését indukálja (Cheng és Grande, 2002; Ronnov-Jessen és Petersen, 1993).
Emellett más növekedési faktorok, többek között a tumor necrosis faktor (TNF),
epidermal growth faktor (EGF), basic fibroblast growth faktor (bFGF), platelet-derived
growth faktor (PDGF), valamint az interleukinek is részt vesznek a szintézis
szabályozásában, részben a mátrix fehérjék termelésének stimulálásával, részben az
ezeket termelő sejtek számának növelésével. Vérellátás nélkül a daganatszövet
életképtelen, ezért a tumor növekedése és terjedése szempontjából lényeges növekedési
faktorok a bFGF és a vascular endothelial growth faktor (VEGF), melyek az
endothelsejtek proliferációját, a daganatos érújdonképződést stimulálják. Fontosságukat
támasztja alá, hogy gátlásuk akadályozza a daganatnövekedést (Angelucci és Bologna,
2007).

14
Mivel a tumorprogresszió számos eseménye megköveteli a daganatsejtek és a
környező mátrix kapcsolatát, feltételezhető, hogy az ECM minősége hatással van a
sejtek biológiai viselkedésére. A pontos molekuláris mechanizmus ma még nem ismert.
Az invazív tumorsejtekben számos mátrixbontó enzim expressziója fokozódik.
A proteolitikus enzimek három csoportját különítik el: cisztein/aszpartil proteinázok,
mátrix metalloproteinázok (MMP), szerin proteázok (Metayer és mtsai, 2002).
Előfordul, hogy az invazív fenotípus kialakulásakor nem a mátrixbontó enzimek
expressziója fokozódik, hanem a gátlófehérjék expressziója csökken. Mindhárom fő
mátrixbontó enzimrendszernek megvan az inhibitora. Az MMP-9/2 kollagénáz IV gátlói
például a TIMP1/2 fehérjék (tissue inhibitor of metalloprotease), a katepszinek gátlói a
stefinek, míg a plazminogén aktivátor rendszer gátlói a plazminogén inhibitorok.
A kollagenázok jelentőségét igazolja az a kísérletes adat, mely szerint TIMP-t
tumorsejtekbe transzfektálva azok elvesztik invazív karakterüket (Gallegos és mtsai,
1995).
3.2.2. Az ECM szerepe a szöveti működés szabályozásában
A szöveti kötelékben a sejteket egymástól az ECM választja el. Passzív,
térkitöltő funkcióját is beleértve a szerv alakját, méretét, felépítését szabja meg,
valamint különböző szignálok, növekedési faktorok, enzimek, hormonok tároló és
továbbító helye. Ezek kötését, aktiválását, inaktiválását feltehetően a proteoglikánok
végzik, befolyásolva a sejt-sejt, sejt-mátrix kölcsönhatásokat, valamint a
sejtproliferációt. Napjaikban az ECM aktív szerepét a regenerációs folyamatokban is
egyre inkább hangsúlyozzák. A bazális membrán molekulaszűrőként szerepel. Negatív
töltésű cukorláncai révén szelektíven ereszt át bizonyos molekulákat a vesetubulusokba,
vízmolekulákat kötve befolyásolja a szöveti hidratációt, és szövetspecifikus
génexpresszió-regulátorként serkentő vagy gátló funkciót tölthet be.
A normális ECM fehérjéinek gyakorlatilag mindegyike megtalálható a daganatos ECM-
ben is: a szerkezetet biztosító fehérjék, mint a kollagének és az elasztin; az adhéziós és
antiadhéziós molekulák mint a laminin, fibronektin, vitronektin, entaktin, tenascin,
proteoglikánok (decorin, versican, syndecan), és szinte minden fehérje képes a sejtek
biológiai viselkedésének a befolyásolására. Normális és daganatos sejtek egyaránt

15
rendelkeznek mátrixfehérje receptorokkal, melyek megfelelő ligand jelenlétében jeleket
közvetítenek a sejt számára. Legismertebb receptorcsalád az integrinek családja, ezek
főleg Arg-Gly-Asp-Ser (RGDS)-régiokon keresztül a laminint, fibronektint, kollagént
képesek megkötni.
Az ECM szerepei közé tartozik a mikrokörnyezet megváltoztatása. Egyes ECM
fehérjék, elsősorban a proteoglikánok, nagy vízkötő képességgel rendelkeznek, így
fokozott termelődésük laza mátrixot eredményez, mely a tumorsejtek terjedését segíti
elő (Madri és Marx, 1992). Számos ECM fehérje multidomain szerkezetet mutat. A
tenascin, laminin, trombospondin és az egyes proteoglikánok (pl. az agrin, perlecan)
bizonyos domainjai növekedési faktorokkal mutatnak homológiát. Kísérletileg
igazolták, hogy ezek a domainok képesek a növekedési faktor receptorához kötődni és
stimuláló hatást kiváltani.
Az ECM-mel kapcsolatban lévő növekedési faktorok közül a legismertebbek a heparint
és heparánszulfátot kötők. Ilyenek a TNF és TGF család, melyek fontos szerepet
játszanak a tumorok növekedésében.
3.2.3. Az ECM és a tumorprogresszió
A tumorprogresszió alapvető feltétele a daganatos stroma kialakítása. Ez a
neostroma támasztékot nyújtva lehetővé teszi a tumorsejtek jellemző elrendeződését,
biztosítja a tumorszövet vérellátását, modulálja a biológiailag aktív molekulák
transzportját.
A daganatos stroma kedvezhet a tumor terjedésének és növekedésének, vagy tükrözheti
a környező normál szövet védekezési, valamint elhatárolási törekvéseit, tehát lehet
serkentő vagy gátló (Mueller és Fusenig, 2002).
A tumorsejtek kapcsolódása az ECM molekuláihoz membránreceptorokon
keresztül történik (integrinek), melyek szintézise és sejtfelszíni expressziója jellegzetes
az adott tumorra ill. tumor régióra (Eble és Haier, 2006). A malignusan transzformált
sejtek képesek olyan proteázokat (pl. mátrix metalloproteázok) ill. proteáz aktivátorokat
termelni, melyek a tumorsejtek migrációját, invázióját a mátrix degradálásával teszik
lehetővé. Továbbá a tumorsejtek képesek a gazdaszervezet sejtjeit (fibroblasztok)

16
mátrix molekulák termelésére késztetni és ezáltal a maguk számára megfelelő miliőt
kialakítani.
A tumorsejtek túlélésének, proliferációjának másik lényeges mozzanata a
tumorsejteket tápláló tumorérhálózat létrehozása, amely folyamat elengedhetetlen, ha a
tumor mérete 1-2 mm-nél nagyobbra nő és amikor már nem képes a környező
szövetekből biztosítani az anyagcsere működését. A tumor angiogenesis elősegítésében
a számos tumorszövetből izolált és tumorsejtek által szekretált vascular endothelial
growth faktor / vascular permeability faktor szerepe tűnik kulcsfontosságúnak, amely
növekedési faktorok serkentik az endothelialis sejtek proliferációját és fokozzák a
permeabilitást (Timár és mtsai, 2006).
A daganatsejtek a környező stromával állandó, dinamikus kapcsolatban vannak.
Részben saját létükhöz új, ún. „neostromát” termelnek, vagy erre késztetik a kötőszöveti
sejteket, másfelől növekedésük és terjedésük során a már levő stromát lebontják. A
stroma felől szignálok jutnak el a daganatsejtek felé, melyek azok viselkedését
befolyásolják.
3.2.4. Az ECM összetétele humán májszövetben
A normál májban az ECM makromolekulák dinamikus komplexe, mely a máj
rostos vázát képezi és részben a Disse-térben foglal helyet. Az ECM fehérjék
termelődésének fokozódása vagy lebontásuk csökkenése az ECM felszaporodásához
vezet, mely a Disse-tér lezárásával mechanikusan akadályozza a makromolékulák
szabad cseréjét a hepatocyták és plazmatér között. Újabb ismereteink szerint a
strukturális szerep mellett aktívan részt vesz a szervet alkotó sejtek működésének
szabályozásában és nélkülözhetetlen a hepatocyták differenciált funkcióinak
ellátásához. Az ECM-et alkotó makromolekulák három csoportba sorolhatóak, 1.
kollagének, 2. nem kollagén glikoproteinek és 3. proteoglikánok (PG).
3.2.5. Proteoglikánok mint ECM komponensek

17
3.2.5.1. A proteoglikánok szerkezete
A proteoglikánok olyan ECM komponensek, melyek a sejtfelszínen, de a sejten
belül is előforduló makromolekulák. Jellegzetességük, hogy vázfehérjéikhez savas
karakterű cukorláncok, a glükózaminoglikánok (GAG) kapcsolódnak. A GAG-ok olyan
elágazás nélküli poliszacharidok, amelyekben egy diszacharid egység ismétlődik,
melynek egyik tagja mindig egy amino-cukor (N-acetil-glükózamin vagy N-acetil-
galaktózamin), ami igen gyakran szulfatálódik is. A másik cukor általában egy uronsav
(Gallagher és mtsai, 1986). Mind a szulfát-, mind a karboxil-csoport negatív töltést
hordoz, így a GAG-ok az emlős sejtek által termelt leganionosabb molekulák. Ezáltal
sok kationt képesek kötni, amiket pedig vízmolekulák kísérnek. Jellemző, hogy nagyon
kiterjedt konformációt vesznek fel, miáltal nagy terek kitöltésére képesek. Négy nagy
csoportra bonthatók a GAG-ok az őket felépítő cukorkomponensek alapján: 1)
hialuronan (hialuronsav), 2) chondroitin-szulfát (dermatánszulfát), 3) heparánszulfát, 4)
keratánszulfát (Taylor és Gallo, 2006).
A glikoproteinekre általában 1-60 tömeg% szénhidráttartalom jellemző, míg a
proteoglikánoknak hosszú (átlagosan 80), elágazásmentes szénhidrátláncai a teljes
molekula tömegének 95%-át is kitehetik. Két példa proteoglikánokra: a porcszövet
zömét adó aggrecan több mint 100 GAG lánccal rendelkező fehérjemolekula, míg a
kollagénrostokhoz kapcsolódó decorinnak csak egyetlen GAG lánca van. A
proteoglikán vázát adó fehérjemolekula a durva felszínű endoplazmatikus retikulumon
szintetizálódik, annak üregébe kerül. A szénhidrát oldalláncok ezt követően a Golgi
apparátusban kapcsolódnak e fehérjéhez. Első lépésként egy tetraszacharid csatoló
(link) alakít ki kovalens kötést egy szerin oldallánccal. Egy fehérje sok szerint
tartalmazhat és nem ismert az a mód, ami meghatározza, hogy ezek közül melyek
kapcsolódjanak a poliszacharid lánccal. (Feltételezhetően a fehérje konformációja
hordozza az információt.) Ezt követően a glikozil-transzferáz enzim a tetraszacharid
lánchoz egyesével újabb monoszacharidokat kapcsol, úgy, hogy az újonnan létrejövő
lánc leginkább ismétlődő diszacharid egységekből fog állni. Még mindig a Golgi
apparátushoz kötött állapotban zajlik le a polimerizált cukormolekulák esetleges további
kovalens módosítása (epimerizáció, szulfatálás). Figyelembevéve a vázfehérjeként
szóba jövő proteinek és a GAG-ok felépítésére rendelkezésre álló cukorkomponensek

18
számát, a lehetséges proteoglikánok száma csaknem végtelen (Silbert és Sugumaran,
2002; Sugahara és Kitagawa, 2002).
A proteoglikánok sokféleségét látva nem meglepő, hogy szerepük nem
korlátozódik a sejtek közötti hidratált közeg létrehozására. A GAG oldalláncok
variálásával pl. eltérő pórusméretű, töltésű géleket alakíthatnak ki, ami megszabja a
proteoglikánon belüli diffúziós folyamatokat. Erre példa a vese glomerulusainak bazális
laminájában található agrin és perlecan (heparánszulfátok közé tartoznak), amely
megszabja, hogy mi juthat a vérből a szűrletbe (Rabenstein, 2002). A proteoglikánok
kémiai jelátviteli folyamatokba is bekapcsolódnak. A fibroblast növekedési faktorok
(FGF) számos sejtféleség osztódását serkentik. Ez a faktor heparán-szulfát
oldalláncokhoz kötődve oligomerizálódik, ami szükséges receptorának aktiválódásához
(Kato és mtsai, 1998). Egy másik növekedési faktor, a TGF-β nem GAG oldallánchoz,
hanem a protein komponenshez kapcsolódik és ez a kötődés meggátolja a faktor
működését (Hildebrand és mtsai, 1994). A proteoglikánokon nem csak növekedési
faktorok, hanem egyéb fehérjék is megkötődnek. Ilyen módon szabályozódik az
aktivitása számos szekretált fehérjének, köztük sok proteolítikus enzimnek (proteáznak)
is.
A GAG-ok és a proteoglikánok hatalmas, rendezett struktúrákat is felépíthetnek
az ECM-ben. Például sok aggrecan molekula kapcsolódhat egyetlen hialuronsav
molekulához, így egy baktérium méretű komplex keletkezik. De a proteoglikánok egyéb
mátrixfehérjékkel is kapcsolódhatnak, pl. kollagénnel a bazális lamina felépítésekor.
Magukhoz a kollagén fibrillumokhoz is kapcsolódik egy proteoglikán, a decorin, ami
nélkülözhetetlen a kollagénrostok kialakulásához. Ezt bizonyítja, hogy a decorinhiányos
egerek bőre törékeny, azaz nyújtással szemben kevéssé ellenálló (Reed és Iozzo, 2002).
Az újabb eredmények szerint proteoglikánok befolyásolják a sejtszaporodást, az
embriogenezist, a morfogenezist, a karcinogenezist, és jelentőségük lehet a betegségek
kialakulásának megértésében (Iozzo, 2005). A daganatokban általában megnövekszik a
proteoglikánok termelése, mennyiségük akár a normális szövetben mértnek 10-20-
szorosa is lehet. Vizsgálataink részletesen érintették a májbetegségekben - elsősorban a
májrákokban - bekövetkező egyes proteoglikánok, elsősorban az agrin minőségi és
mennyiségi változását.

19
3.2.5.2. A proteoglikánok osztályozása
1. táblázat. Proteoglikánok osztályozása.
A) Cukorláncaik alapján (Tímár és mtsai, 1995)
A proteoglikánokat osztályozzák a fehérjevázhoz kapcsolódó cukorlánc alapján.
1. Egyszerű proteoglikánok: a fehérjevázhoz egy vagy több azonos típusú
cukorlánc kapcsolódik. Ilyen módon heparánszulfát proteoglikán (HSPG),
kondroitinszulfát proteoglikán (CSPG) és keratánszulfát proteoglikán (KSPG)
Cukor láncai Intracellularis PG
Sejtfelszíni PG ECM PG
Keratán szulfát Fibromodulin Keratocan Lumican PRELP Osteoadherin Osteoglycin
Chondroitin szulfát/ Dermatán szulfát
Chromogranin-A
i-chain of HLA-DR CD44 Melanoma PG Neurocan NG2 Phosphocan Thrombomodulin
Aggrecan Biglycan Decorin Versican Collagen IX, XII, XIV Embrionalis mesenchyma PG Epiphycan
Egy
szerű
PG
Heparán szulfát Glypican 1,2,3,4,5,6 Syndecan 2,3, 4 Transferrin-receptor
Perlecan Agrin Collagen XVIII
Összetett PG-k
Serglycin Betaglycan Syndecan 1 Amphiglycan

20
különíthető el. Ezektől eltérően a hyaluronsav soha nem kapcsolódik kovalens kötéssel
a fehérjéhez. Az ECM-ben szabadon foglal helyet, vagy a porc esetén speciális
kötőfehérjével óriás CSPG molekulák kapcsolódnak hozzá.
2. Összetett proteoglikánok: a fehérjevázhoz különböző típusú GAG láncok
kapcsolódnak.
B) Szerkezet, funkció és lokalizáció alapján
Az ismeretek gyarapodásával kísérletek történnek arra, hogy a felosztásnál a molekulák
funkcióját, lokalizációját is figyelembe vegyék. Ezt a törekvést tükrözi Gallagher 1989-
es felosztása.
I. Intracelluláris proteoglikánok
A serglycinek felelősek a bazofil és hízósejtek szekretoros granulumainak
metakromáziás festődéséért. Hosszan ismétlődő Ser-Gly dipeptidjei miatt a molekulát
így nevezték el. A kis, kb. 10 kDa méretű fehérjevázhoz HS vagy CS kapcsolódhat. Ez
a cukorkötő régió, melyhez számos cukorlánc kapcsolódhat, rezisztenssé téve a
molekulát proteázokkal szemben. Sajátos vonása a fehérjeváznak, hogy sejttípustól
függően más cukorlánc kapcsolódik hozzá, így a hízósejtben heparin, az NK sejtben CS,
a mucosa hízósejtjeiben CS a kapcsolódó GAG. A molekula feltehető funkciója
proteolitikus enzimek tárolása a granulumokban, azok kötése által. A sejtekből kijutva
szabályozza a bontó enzimek aktiválódását is.
II. Sejtfelszíni proteoglikánok
1. CSPG-k
A sejt-sejt interakciókban játszanak szerepet. A melanoma proteoglikán a melanomák és
neuroektodermalis eredetű tumorok sejtfelszínén található (Nishiyama és mtsai, 1991).
A thrombomodulin a hemosztázisban vesz részt, alvadásgátló aktivitásának jelentős
hányadát a fehérjeváz határozza meg, a CS lánc kapcsolódása esetén azonban a
molekula antikoaguláns kapacitása fokozódik (Sadler, 1997). A TAP-1 az idegrostok
terminális membránján található, szerepe az axonok lokalizálásában van az izomsejtek
felszínén (Arenson és Bissell, 1987). A HNK-1 a gliasejtek és idegsejtek
interakciójának egyik letéteményese. A CD44 fakultatív proteoglikán, eredetileg a

21
lymphocyták felszínén kimutatott fehérje, mely fontos szerepet játszik a lymphocyták
nyirokcsomókba történő recirkulációjában. Epithelialis sejtek felszínén is megtalálható,
CS és HS lánc glikanálhatja. A variációk szövetspecifikusak. Feltételezhetően a
sejtspecifikus felismerésben és adhézióban játszik szerepet (Naor és mtsai, 1997).
2. HSPG-k
Syndecan 1-4, a név a görög syndein - összekötni - szóból származik. A molekula 3 HS
és 1 CS láncot tartalmazhat, transzmembrán domain köti a felszínhez. A HS láncain
keresztül képes kapcsolódni az I, III, V típusú kollagénhez, fibronektinhez,
trombospondinhoz. Az epiteliális sejtek típusos proteoglikánja. Az elmúlt években
számos, az eredetivel homológ molekulát szekvenáltak, melyek vázfehérjéinek mérete
31-90 kD között van (Woods, 2001). A syndecan csoporthoz tartozó syndecan-2-t
(fibroglycan) embrionális tüdő és bőr fibroblasztokon mutatták ki, 125, 90, 60 és 45
kDa fehérjevázzal (Heremans és mtsai, 1988). A 90 és 45 kD fehérjeláncok a
peptidanalízis alapján rokon struktúráknak bizonyultak. Feltehetően ez is
transzmembrán proteoglikán, amelyhez valószínűleg három cukorlánc kapcsolódik. A
fibroglikánnal nagyfokú szekvencia-homológiát találtak a patkány hepatocyták
sejtfelszíni HSPG-jének klónozása során, így feltételezhető, hogy ezek felszínén is
fibroglikán található. A betaglycan a TGBβl III típusú receptora. 110 kD fehérjevázához
HS és CS lánc kapcsolódik. A GAG láncok nem szükségesek a ligandkötési funkcióhoz
(Esko és mtsai, 1987). Feltehetően a növekedési faktor receptorkötődése által kiváltott
fokozott mátrix-szintézis negatív regulációjában van szerepe (Cheifetz és mtsai, 1987).
A vastagbélrák sejtek HSPG-je nagyfokú homológiát mutat a perlecannal (Dodge és
mtsai, 1991). Transzmembrán fehérje, mely hidrofób domaint tartalmaz (Iozzo és mtsai,
1990). Northern blot analízissel hasonló proteoglikán messenger RNS szintű
expresszióját igazolták normál vastagbél szövetben is.
A proteoglikánok különböző módon kapcsolódhatnak a sejtfelszínhez, ennek egyik
lehetséges módja a transzmembrán hidrofób domain segítségével történő kapcsolódás,
egy másik lehetőség a beékelődés a lipid kettősrétegbe a proteoglikánon levő
inozitolfoszfát segítségével, vagy egy specifikus membránreceptorhoz való kötődés a
HS láncon vagy inozitolfoszfáton keresztül.

22
A sejtfelszíni HSPG molekulák szerepét a cukorláncok vagy a vázfehérjék
határozhatják meg. Igen változatos szerepet játszhatnak, megszabják a sejt-sejt, sejt-
ECM interakciókat, enzimeket, növekedési faktorokat, hormonokat képesek kötni,
azokat serkentve, vagy éppen inaktiválva. Bizonyos rendszerekben a fehérjéről leváló
HS láncok a sejtmagba jutva a sejtproliferáció szabályozásában vesznek részt.
Felvetődik a lehetőség, hogy egyes HS láncok információt is hordoznak pl. specifikus,
bázikus FGF és thrombospondin kötő régiót sikerült kimutatni rajtuk.
III. Az ECM proteoglikánjai
1. Térkitöltő CSPG-k az ECM-ben
Az aggrecan a porc proteoglikánja (Doege és mtsai, 1990), a human fibroblaszt
proteoglikán versican, az embrionalis mesenchyma proteoglikán (Kimata és mtsai,
1986) és az agy CSPG-i a legismertebbek. Jellegzetességük, hogy egységekből épülnek
fel, így pl. a versican EGF, lektinszerű, complement regulatory protein (CRP) és hyalin
kötő régiókat tartalmaz. A sejtek adhézióját általában gátolják (Cattaruzza és mtsai,
2002).
2. Kollagén rostokhoz kapcsolódó CSPG-k
A proteoglikán-2/decorin, kb. 40 kD fehérjevázzal rendelkező molekula a kollagén I d
és e csíkjaihoz kapcsolódik, gátolja a kollagénrost képződést. Csak 1 cukorkötő helye
van, DS vagy CS kapcsolódik ehhez. A kötőszövetben csaknem mindenütt megtalálható
(Krusius és Ruoslahti, 1986). A proteoglikán-1/biglycan szintén kis CS- vagy DSPG,
mely DNS szinten nagyfokú szekvenciahomológiát mutat a decorinnal, azonban bi-
zonyíthatóan elkülöníthető gén terméke (Fisher és mtsai, 1989). Nevét azért kapta, mert
két cukorlánc kapcsolódik hozzá. Kollagénhez való kötődését eddig még nem
bizonyították, ugyanakkor erősen kötődik az ECM egyéb komponenseihez. A biglycan
és decorin együtt a rostos kötőszövet proteoglikánjainak a 95%-át teszik ki. A kollagén
IX-ről az utóbbi időben tisztázódott, hogy ez a molekula is proteoglikán, 1 CS lánc
kapcsolódik hozzá (Konomi és mtsai, 1986). A hyalin porcban található rostos hálózat
része, a cukorlánc különösen alkalmassá teszi, hogy egyéb mátrix proteinekhez
kapcsolódjon.

23
3. KSPG-k
A cornea állományának egyik fő alkotórésze egy KSPG, alapvető szerepe van az
optikailag átlátszó, rendezett rosthálózat kialakításában. Keratánszulfát láncot mutattak
ki ezen túlmenően az aggrecanon, az ín és az aorta proteoglikánjai között. A
fibromodulin egy KS proteoglikán, mely jelentős homologiát mutat a decorinnal és a
biglycannal (Chakravarti, 2002). Szintén képes a kollagénhez kötődni.
4. Bazális membrán HSPG-k (BMHSPG)
A perlecan (Engelbreth-Holm-Swarm (EHS) sarcoma proteoglikán) a legjobban
tanulmányozott BMHSPG (Gallai és mtsai, 1996). Fehérjeváza kb. 400 kD, ehhez 4
GAG lánc kapcsolódik. Szerkezetében a lineáris struktúrát 7 globuláris régió szakítja
meg, mint gyöngyfüzér. Innen kapta a nevét is. Számos domainje homologiát mutat
egyéb fehérjékkel, így ciszteingazdag régiója a laminin B láncával, egy többhurkos
struktúrája az NCAM-al és egyéb az immunglobulin szupercsaládba tartozó fehérjékkel.
Az endothelsejtek és a humán fibroblaszt nagy HSPG-je feltehetően hasonló struktúrák
(Heremans és mtsai, 1988). A kollagen XVIII a HS láncai miatt a bazális membrán
HSPG-k közé tartozik (Halfter és mtsai, 1998). A BMHSPG-k egyik fontos tagja az
agrin, amelyikről következő fejezetben írok részletesebben.
A BMHSPG-k elsődleges funkciója a molekulaszűrés. Ezen túlmenően az idegrostok
növekedésében a lamininnal alkotott komplexének jelentőségét írták le, úgy tűnik
szoros kapcsolata van az NCAM fehérjével is elősegítve ezáltal az idegrostok
regenerációját (Cole és Burg, 1989). Újabb adatok szerint HS láncaival növekedési
faktorok tárolására képes.
A proteoglikán kutatás felgyorsult üteme a csoportok egyre újabb tagjait deríti fel.
Nyilvánvalóvá vált, hogy méretükben, szerkezetükben és funkcióikban nagyon
heterogén molekulákról van szó. Ismereteink gyarapodásával egyre több adat támasztja
alá, hogy ezen molekulák jelentősége messze nagyobb, mint feltételeztük.
A proteoglikánok fiziológiás körülmények között a következő folyamatokban
játszanak szerepet:
1. A kötőszövet és támasztószövet alkotórészei.

24
2. Negatív töltésük miatt megszabják a szövetek hidratációját; molekulaszűrőként
funkcionálnak.
3. Meghatározóak a sejt-sejt és sejt-ECM kapcsolatokban.
4. Részt vesznek a sejtek szövetté történő szerveződésében.
5. Növekedési faktorok, enzimek, hormonok, citokinek kötése, prezentálása és
semlegesítése révén aktív részesei a sejt- és szövetműködés fiziológiás szabályozásának.
3.2.5.3. Az agrin mint BMHSPG
3.2.5.3.1. Az agrin szerkezete
Agrin, a név görög „agrein”-összegyűjteni (assemble) - szóból származik, ami
azt jelenti, hogy képes acetylcholine receptor (AChR) aggregációra. Az agrin nagy
méretű, erős negatív töltésű multidomain HSPG (Tsen és mtsai, 1995).
Az emberi agrin gén (AGRN) locusa: 1p36.1-1, ami viszonylag közel helyezkedik a
perlecan génhez (1p35-36.1) (Dodge és mtsai, 1991). Az emberi agrin mRNA
hosszában meghaladja a 7 kb-t és több mint 2,000 aminosavból álló, 212 kDa
molekuláris tömegű fehérjét kódol (Rupp és mtsai, 1992).
Szerkezetileg az agrin egy globularis laminin-kötő domaint (NtA), kilenc
follistatin-szerű proteáz inhibitor domaint (FS), két laminin-szerű epidermális
növekedési faktor ismétlődést (LE), két serine/threonine gazdag domaint (S/T), a SEA
module-t (SEA), négy EGF ismétlődést (EG) és három laminin α-lánchoz homológ
laminin globuláris domaint (LG) tartalmaz (1. ábra) (Rupp és mtsai, 1991; Tsim és
mtsai, 1992). Az intenzíven N- és O- glikolizált N-terminális miatt az agrin molekuláris
tömege oldallánccokkal együtt eléri a 600 kDa-t (Tsen és mtsai, 1995). Az
aminocsoportokhoz kapcsolódó szénhidrátok és a GAG az agrin szekvencia egész
hosszában megtalálhatók, de HS-GAG formája főként a fehérje N-terminális szakaszára
korlátózódik. Ezek a régiók ugyancsak részt vesznek az idegsejt adhéziós molekulák
(NCAM) és a heparin kötő növekedési faktorok megkötésében (Cole és Halfter, 1996).
A C-terminálisan helyezkedő, 95-kDa része (ahol az első epidermális
növekedési faktor (EGF)-szerű domain (EG) kezdődik) szerepet játszik az AChR
aggregacióban, valamint tartalmaz α-dystroglycan, heparin, egyes integrin és még nem

25
azonosított „agrin receptor” kötő szakaszokat (Ferns és mtsai, 1993; Gesemann és
mtsai, 1995; Ruegg és mtsai, 1992)
1. ábra. Agrin szerkezete: NtA-N-terminális globularis laminin-kötő domain,
TM- transzmembrán domain, FS-follistatin-szerű proteáz inhibitor domain, LE-
laminin-szerű ismétlődő szekvenciák, S/T-serine/threonine gazdag domain, a
SEA-SEA module, EG-EGF ismétlődés, LG-laminin α-lánchoz homológ
laminin globuláris domain. (Bezakova és Ruegg 2003).
3.2.5.3.2. Az agrin típusai Burger és munkatársai (2000) a patkány agrin két, különböző N-terminálissal
rendelkező izoformáját írták le: az SN-agrint (rövid izoforma, TM agrin) és LN-agrint
(hosszú izoforma, NtA agrin), amelyek a szubcelluláris lokalizációban, a szöveti
eloszlásban és a funkcióban is különböznek egymástól. Az SN-agrin elsősorban az
idegrendszerben (IR) található, míg az LN-agrin a központi IR-en kívül más szervekben
(vese, tüdő, NMJ) is előfordul (Ruegg és Bixby, 1998). Az SN-agrin sejtmembránhoz
kötődve, az LN-agrin az ECM bazális laminájához társultan fordul elő (Bezakova és
Ruegg, 2003). Knockout egerekben az LN-agrin expressziójának csökkenése a
neuromuszkularis junkció működésének csökkenéséhez vezetett (Gautam és mtsai,
1996; Lin és mtsai, 2001). Az LN-agrin a bazális lamina egyik komponenseként, fontos

26
szerepet játszik a neuromuszkuláris junkció (NMJ) jeltovábbításában, míg az SN-agrin
elsősorban az idegsejtek közötti jelátvitelben vesz részt.
Mind a LN-agrin és a SN-agrin izoformák alternatív splicing következtében
különbözhetnek még legalább két további – A/y- és B/z-ként ismert – pozícióban,
melyek a karboxi-terminális laminin-globuláris (LG) LG2, illetve LG3 tartományban
vannak. Az e helyekre való beillesztés olyan fehérje variánsokat hoz létre, melyek 0
vagy 4 aminosavat tartalmazhatnak az A/y pozícióban, és 0, 8, 11 vagy 19 (8+11)
aminosavat a B/z pozícióban. A Lys-Ser-Arg-Lys (Lys =lysin, Ser=serin, Arg=arginin)
A/y helyre történő beillesztése a heparin kötéséhez szükséges, továbbá modulálja az
agrin kötődését az α-dystroglycanhoz, egy erősen glikozilált periferiális membrán
fehérjéhez, mely számos szövetben, így az izomban is expresszálódik. Ezen beillesztés
fiziológiai fontossága az A/y helyen tisztázatlan, mivel látszólag normálisak azok az
egerek, melyekben nem található ez a bizonyos exon (Bezakova és Ruegg, 2003).
Ezzel ellentétben, az aminosav beillesztések a B/z helyen döntőek az agrin-indukált
AChR aggregáció szempontjából. Tenyésztett myotubusok esetében csak a
beillesztéssel (agrin –B/z+) rendelkező izoformok indukálnak AChR aggregációt és ez
az aktivitás nincs jelen, amennyiben hiányzik a beillesztés a B/z helyen (agrin B/z-).
Az AChR aggregációs aktivitást mutató agrin mRNS-t kódoló izoformák a
motoneuronokban expresszálódnak, míg a nem idegi szövetek sejtjei (tüdő, vese és
izom) olyan agrin transzkriptumokat szintetizálnak, melyeknél hiányoznak a
beillesztések az A/y és B/z helyeken. A nem izomszövetek esetében az agrin a
dystroglycan nagy affinitású kötő fehérjéje (Gesemann és mtsai, 1998).
3.2.5.3.3. Az agrin funkciói
AChR aggregáció:
Az agrin legfontosabb és legrészletesebben tanulmányozott funkciója az NMJ-
ben az AChR aggregáció kiváltása. A B/z+ agrin posztszinaptikus differenciálódást
kiváltó képessége bizonyított, ennek molekuláris folyamata azonban csak részlegesen
ismert. Az AChR-aggregáló képességhez az LN-agrinnak rendelkeznie kell karboxi-
terminális 95-kDa domainnal. Érdékes, hogy in vitro a B/z+ agrin LG3 domaint
tartalmazó 21-kDa fragmentje is kiváltja az AChR aggregációt (Gesemann és mtsai,
1995; Yang és mtsai, 2001), de jóval kisebb mértekben, mint a teljes hosszúságú vagy a

27
95-kDA-os fragment. Valószínűsíthető, hogy a B/z+ agrin a LG3-B/z+ domainen
keresztül kapcsolódik a receptorával (receptoraival) (McMahan, 1990), másrészt az
agrin α-dystroglycanhoz kötődve, valamint a muscle-specific receptor tyrosine kinase
(MuSK) aktiválásával részt vesz az AChR aggregálásában (Ruegg és Bixby, 1998). Az
agrin-deficiens egerek az NMJ súlyos funkciózavara következtében közvetlenül születés
után elpusztulnak (Gautam és mtsai, 1996).
Embrionalis szövetekben az agrin a motoneuronokban szintetizálódik, majd
axonalis transzport révén terminálisan szekretálódik, így az NMJ szinaptikus résben a
bazális laminában állandóan jelen van (Magill-Solc és McMahan, 1988). NMJ
kialakulásakor a motoneuronok által expresszált agrin izoformák indukálják a
posztszinaptikus membránban az AChR-ok aggregációját. Ezzel szemben az izomsejtek
által expresszált izoformáknak nincs aggregáló hatásuk (Ruegg és mtsai, 1992).
A strukturális és funkcionális vizsgálatok kimutatták, hogy az agrin különböző
sejtfelszíni és ECM fehérjékhez kötődik, például a lamininhez (Denzer és mtsai, 1997;
Denzer és mtsai, 1998; Kammerer és mtsai, 1999) és az α-dystroglycanhoz (Gesemann
és mtsai, 1998).
A laminin az LN-agrin amino-terminális részén levő NtA véghez kötődik (2.
ábra). Az agrin elsősorban a laminin 2, 4-hez kötődik és ez a két laminin jelen van az
izom bazális laminájában. Az agrin erős kötődése a laminin-4-hez megmagyarázhatja,
hogy miért marad az agrin a sejtes elemek degenerálódása után is a szinaptikus bazális
laminához társultan (Reist és mtsai, 1987). Az agrin karboxi-terminális részén található
az α-dystroglycan kötő rész (Bowe és Fallon, 1995). Az α-dystroglycan a prekurzor
fehérje poszttranszlációs hasítását követően alakul ki, erősen glikolizált perifériás
membránfehérje, a lamininhez, az agrinhoz és a transzmembrán β-dystroglycanhoz
kötődik (Gesemann és mtsai, 1998). Az α- és a β-dystroglycan különböző fehérjékkel
együtt alkotja a dystrophin- glycoprotein complexet. Igy az agrin a bazálmembránt a
cytoskeletonhoz rögzítve jeltovábbításban vehet részt (Bowe és mtsai, 1994; Bowe és
mtsai, 1994; Gee és mtsai, 1994).

28
2.ábra. Az agrin a postsynaptikus differenciálódásban:
(www.cgap.nci.nih.gov/Pathways)
Más kísérletek két további fehérje szerepét támasztják alá az agrin jelátvitelben játszott
szerepében. Az egyik az izom-specifikus tirozin kináz (MuSK) (Jennings és Burden,
1993), a másik cytoplasma membrán fehérje a rapsyn (Frail és mtsai, 1988).
Az agrin neurogenesisben játszott szerepét alátámasztja, hogy magas agrin
expressziós szintet mutattak ki a fejlődő agy ventricularis és subventicularis zónáiban és
a gerincvelőben; ezzel ellentétben a felnőtt agyból hiányzik. Ezen kívül agrin-hiányos
egerek agya sokkal kisebb volt, mint a vad típusé (Serpinskaya és mtsai, 1999).
Az agrin létfontosságú a kolinerg idegsejtek szinaptikus működésének szabályozásában
a felső cervicalis ganglionban (SCG). Az agrin-deficiens embriók a pre- és
postsynaptikus struktúrák szabálytalan illeszkedést és jeltovábbítási hibáit mutatják
(mismatch-et és synaptikus transmission hibát). (Gingras és Ferns, 2001). Az SCG-ben

29
az agrin SN izoformája expresszálódik, és a MuSK-on keresztül hatással van a
synaptogenesisre (Ip és mtsai, 2000).
Az agrin immunreakcióját mutattak ki az agy mikrovaszkulatúrájában, ahol az agrin az
α-dystroglycanhoz és a lamininhoz kötődve stabilizálja a specifikus bazális laminát, ami
a vér-agy gát felépítésében játszik szerepet (Donahue és mtsai, 1999). Az agrin β-
amyloid-hoz is kötődik (Cotman és mtsai, 2000), ami az Alzheimer-es beteg amyloid
plaque-jában felhalmozódik; ennek a kórnak egyik tipikus következménye a vér-agy gát
fokozott permeabilitása (Donahue és mtsai, 1999).
Agrin a glomerularis bazális membránban
Az agrin a vese glomeruláris bazális membránjában (GBM) is kimutatható
(Groffen és mtsai, 1998). Itt részt vesz a töltésszelektív ultrafiltrációs folyamatban.
Az agrin a GBM-ben elsősorban a lamininhoz kötődik. Ez a kötődés a laminin hosszú
kar centrális régiójára lokalizálódik.
A GBM és a sejt felszíne közötti térben az agrin fontos szerepet játszhat a sejt-
matrix adhézióban. Az agrin egyik sejtfelszíni receptora, az α-dystroglycan a vese érett
és fejlődő epitheliumában magas expressziót mutatott (Durbeej és mtsai, 1998). Az érett
glomerulus GBM-ben az α-dystroglycan az agrinhoz hasonlóan expresszálódik (Durbeej
és mtsai, 1998). Így az agrin és az α-dystroglycan kolokalizációja a GBM-ben
támogatja a sejt-mátrix kölcsönhatás funkciót. Ebben a kapcsolatban az agrinnak
kihorgonyzó szerepe lehet a citoszkeleton és BM közötti szerkezeti kölcsönhatás
biztosításában. A citoszkeletonnak a GBM-mel való szoros kölcsönhatása nem csak a
magas helyi vérnyomás fenntartásához nélkülözhetetlen, de a glomerularis filtráció
hemodinamikai változásokhoz történő alkalmazkodásában is szerepet játszik. Az agrin a
transzmembrán jelátvitelben az integrinen keresztül is részt vesz. Az agrin kötődik a
αvβ1-integrinhez (Martin és Sanes, 1997), amelynek fontos szerepe van a sejtek
differenciálódásában.
3.3. A proteoglikánok szerepe májbetegségekben
Az orvostudományban elsődleges kérdésként vetődik fel, mi a jelentősége a
proteoglikánoknak bizonyos kórfolyamatokban. Az elmondottakból nyilvánvaló, hogy a

30
proteoglikánoknak kitüntetett szerepe lehet mindenütt, ahol az ECM felhalmozódása
zajlik, pl. májcirrhosis, májfibrosis, egyéb fibrosisok, sebgyógyulás, keloid képződés,
arteriosclerosis. Ugyancsak fontos az agrin azon folyamatokban, ahol a sejtek
vándorlása történik az ECM-on keresztül, pl. gyulladás, tumorinvázió, metasztatizálás,
valamint ahol a sejtek, szövetek autoregulációja kerül előtérbe, pl. tumorok autonóm
szabályozása, valamint a szervezet védekező funkciói zajlanak, pl. citotoxicitás,
cellularis immunreakciók.
A felsoroltak közül a májra vonatkozó ismereteket szeretném röviden összefoglalni.
A máj proteoglikánjai és változásuk kóros körülmények között
A máj proteoglikánjai
Az ép májban egyéb fehérjékhez képest a proteoglikánok mennyisége elenyésző. A
cukorláncok analízise alapján 65—70%-uk HSPG. A fehérjeváz(ak) méretét, aminosav
és nukleinsav szekvenciáját még nem határozták meg teljes egészében. Feltehetően több
HSPG típus is megtalálható, amelyek vagy a hepatocyták sejtfelszínéhez kötődnek,
vagy az ECM-ben rakódnak le.
Leírtak kicsi, kb. 27 kD-os molekulát, melyhez három vagy négy, egyenként 14 kD
tömegű cukorlánc kapcsolódik (Oldberg és mtsai, 1979). Beszámoltak egy, a
hepatocyták által termelt, az ECM-be lokalizálódó HSPG-ról, melynek mérete 150—
300 kD, 40 kD tömegű fehérjevázzal. Izoláltak egy sejtfelszíni HSPG-t, melynek mérete
200 kD körüli, a fehérjeváz pedig 77 kD (Gallagher, 1989; Lyon és Gallagher, 1991).
Ez a molekula feltehetően homológ a fibroglikánnal. Az ismert proteoglikánok közül a
syndecan expressziója is kimutatható a humán májban. Megtalálhatók ezen kívül a
decorin, a biglycan, a perlecan és a versican is (Kovalszky, I és mtsai, 1997). Ezeket
nagy valószínűséggel nem a hepatocyták, hanem a nonparenchymalis májsejtek (Ito
sejt: decorin, biglycan, perlecan, Kupffer sejt: syndecan, endothelsejt: decorin, perlecan)
termelik.
A proteoglikánok változása májcirrhosisban
A proteoglikánok a kötőszövet és a bazális membrán szerves alkotórészei, így
természetes, hogy mennyiségük a cirrhotikus májban felszaporodik, összetételük

31
megváltozik. Elsősorban a kötőszöveti szeptumoknak megfelelően rakódnak le, döntő
többségük DS-CSPG (Arenson és Bissell, 1987). Előrehaladott cirrhosisban a humán
máj ötször több HS-ot, tízszer több CS-ot és tizenhatszor több DS-ot tartalmaz, mint a
normál máj (Murata és mtsai, 1985). Hasonló nagyságrendű fokozódás a teljes
proteoglikán molekulák esetében nem volt kimutatható, így feltehetően a vázfehérjék
glikanációja is növekszik (Kovalszky és mtsai, 1993). A cukorláncok szulfatáltsága
fokozódik. Az elmúlt években vált ismertté, hogy cirrhosisban a proteoglikánok
fokozott termelését, egyéb mátrixfehérjékhez hasonlóan, a nem-parenchymalis
májsejtek, elsősorban az Ito-sejtek végzik (Senoo, 2004). A fokozott szintézis TGFβl
hatására következik be (Kiss és mtsai, 1998). A perlecan és versican fokozódó
expressziója cirrhosisban szintén a nem parenchymalis sejtekben figyelhető meg
(Kovalszky, I és mtsai, 1997). A syndecan expresszióját humán biliaris cirrhosisban
real-time RT PCR vizsgálattal fokozottnak találták, míg kísérletes széntetraklorid
cirrhosisban nem észleltek érdemi változást (Sebestyen és mtsai, 2000). A
proteoglikánok felszaporodása azonban nemcsak a fokozott szintézis, hanem a
progresszióval párhuzamosan csökkenő lebontás következménye is. A lizoszómális
endohexozaminidázok aktivitása csökken, és különösen nehezítetté válik a DS láncok
degradációja. Részben ez magyarázza a DS mennyiségének kitüntetett emelkedését.
Kérdés, hogy a proteoglikánok mennyiségének és összetételének változása pusztán,
mint a szövetproliferáció egyik részjelensége értékelendő, vagy ennél többről van szó és
ezek a változások hatással vannak a cirrhosis kimenetelére is. Több adat utal arra, hogy
bizonyos proteoglikánok részesei a mátrixképződés szabályozásának. Ismert, hogy a
decorin a kollagén rostokhoz kötődve gátolja azok érését, tehát a kötőszövetképzés
gátlásának irányába hat. A decorin a TGFβl inaktiválására képes, erről a növekedési
faktorról köztudott, hogy a fibrogenezis egyik fő stimulátora (Kovalszky és mtsai,
1998).
Proteoglikánok és májtumor
A témával foglalkozó kutatók kivétel nélkül egyetértenek abban, hogy a
proteoglikánoknak fontos szerepük van a daganatos betegségek progressziójában. Nem
tudjuk, csak feltételezzük, hogy a tumorok indukciójában is szerepük lehet. Elsősorban
leíró adatok állnak rendelkezésre, bemutatva, hogy szinte nincs olyan daganat, melyben

32
a proteoglikánok mennyisége és minősége ne változna meg az eredeti szövethez
viszonyítva. Így van ez a máj esetében is. Kémiai karcinogenezis során az úgynevezett
alterált fókuszokban intenzív GAG festődést mutató anyag jelenik meg a májsejtek
citoplazmájában. A kialakult májrákban ez a felhalmozódás tovább fokozódik. A humán
hepatoma sejtek citoplazmája is alcián kék pozitív és a pozitivitás kondroitináz enzim
emésztést követően eltűnik. Immunhisztokémiával erős HSPG pozitivitás is
megfigyelhető a tumorsejtek citoplazmájában. Kémiai módszerekkel analizálva a májrá-
kokban a kondroitinszulfát kb. hússzoros felhalmozódását találták, a heparánszulfát-
tartalom kb. tízszer nagyobb, mint a normál májban, a GAG-ok egymáshoz viszonyított
aránya is megváltozik.
Nemcsak a májrákok proteoglikán tartalma és összetétele változik meg a normál májhoz
képest, hanem a tumor körüli látszólag ép májszövetben is GAG halmozódik fel
morfológiai és kémiai módszerrel vizsgálva egyaránt (Kovalszky és mtsai, 1990). A
fehérjeváz analízise arra utal, hogy nem a molekulák véletlenszerű felhalmozódásáról,
hanem szelektív eltolódásról van szó, mivel egyes proteoglikánok, mint a syndecan,
expressziója gyakorlatilag eltűnik a májrákból. Ugyanakkor a decorin és egy a bazális
membrán HSPG-nek megfelelő proteoglikán felhalmozódik.
Nem tisztázott még a felhalmozódó proteoglikánok forrása. Nagyon valószínű azonban,
hogy a tumorsejtek részben proteoglikánokat termelnek nagy mennyiségben, részben
pedig olyan faktorokat, melyek a környezet nem tumoros sejtjeit fokozott proteoglikán
szintézisre késztetik. Mi a jelentősége ezeknek a változásoknak? A tumorsejtek felszí-
nén és környezetükben felhalmozódó megváltozott összetételű mátrix, melyben a
CSPG-k dominálnak, a sejtek egymástól való elszakadását segíti elő. Megkönnyíti a tu-
morsejtek invázióját, a mátrix degradációját. Ugyanakkor mintegy körülbástyázza a
daganatot, lehetőséget nyújtva annak fokozott autonómiájára. Ezt a feltételezést
támasztja alá az a tény, hogy a proteoglikánok növekedési faktorokat, cytokineket,
hormonokat képesek kötni, tárolni és semlegesíteni (Ruoslahti és Yamaguchi, 1991). A
bFGF receptora egy syndecannal homológ HSPG (Qiao és mtsai, 2003). A TGFβl III.
típusú receptora, a betaglycan, egy HS/CSPG. A decorin a TGFβl-et semlegesíti.
Hasonló funkció más proteoglikánokról is elképzelhető. Így a megváltozott
proteoglikán-összetétel folytán teljesen más információk jutnak el a tumorsejtekhez,
mint normális proteoglikán-összetétel esetén. Ismert, hogy a TGFβ1 a hepatocyták

33
proliferációját gátolja. Hatásának felfüggesztése proliferációs stimulust jelenthet. A
syndecan az egyik fő letéteményese a sejtek lehorgonyzásának (Saunders és mtsai,
1989), eltűnése a sejtfelszínről elősegíti a sejtek anyatumortól való elszakadását. A
hepatoma sejtek felszínén kimutattak egy HSPG-t, mely bizonyos ingerek hatására
internalizálódik, majd HS lánca a fehérjéről leválva a sejtmagba jut és
proliferációgátlást idéz elő (Fedarko és mtsai, 1989). Amennyiben egy ilyen HSPG
mennyisége csökken, a következmény ismét a proliferáció fokozódása lesz. Újabb
adatok szerint a magban a HS lánc az egyes gének derepressziójában, aktiválásában
fontos szerepet játszó topoizomeráz enzimet gátolja.
Ismert tény, hogy a hepatocyták sejtfelszíni HSPG-jai heparinszerű, tehát antithrombin
aktivitással rendelkeznek. Ezen proteoglikánok arányának csökkenése a
kondroitinszulfát proteoglikánok javára, a HS láncok szulfatáltságának csökkenése,
melyet számos tumor esetén leírtak és magunk májrákok vizsgálata során is
megfigyeltünk, a hemosztázis egyensúlyát az alvadás irányába billentheti el.
Egy daganat biológiai viselkedése természetesen számos, ma még nem minden
részletében ismert, tényezőtől függ. A proteoglikánok megváltozása csak egy ezek
közül, de úgy tűnik, nem jelentéktelen. Jobb megismerésük, észlelt változásaik
modellezése kísérleti rendszerekben remélhetőleg pontosabb választ fog adni arra,
hogyan képesek e fehérjék befolyásolni egy tumor aktuális progresszióját. Nagy a
valószínűsége annak is, hogy a jövőben bizonyos proteoglikánok a daganat-terápia
targetjeiként is számításba jönnek. A fehérjeváz, vagy a cukorláncok szintézisének
gátlásával kísérletes körülmények között már ma is lehet daganat-, illetve
metasztázisgátló hatást elérni.
3.4. A BMHSPG-k szerepe az angiogenesisben
A HSPG-k nélkülözhetetlenek az endothelsejtek strukturális és funkcionális
épségéhez. A HSPG-k a endothelsejtek bazális felszínen vannak jelen, és mátrix-
receptorként viselkednek a különböző bazális membrán fehérjék számára. Továbbá a
BMHSPG-k felelősek a töltésszelektív filtrációs folyamatokért az endotheliumban
(Morita és mtsai, 2005), és gátolják a simaizomsejtek proliferációját, migrációját

34
(Castellot, Jr. és mtsai, 1981). HSPG-k az endothelsejtek luminális felszínen is jelen
vannak, ahol részt vesznek a lipoprotein-lipáz kötődésében és internalizációjában
(Moon és mtsai, 2005). Ezek a luminális HSPG-k ugyancsak fontos szerepet játszanak
az erek felszíni antikoagulatív tulajdonságának a meghatározásában is az intrinsic
véralvadási kaszkád-proteázok, thrombin és proteáz inhibitorok, beleértve az
antithrombin III kötésével. Végül az endothel felszíni HSPG-k számos angiogenikus
növekedési faktor koreceptoraként is működnek, és ezzel az angiogenezis
szabályozásában is részt vesznek.
Mind a makro-, mind a mikrovaszkuláris endothelsejtek termelnek HSPG-ket.
Az endotheliális HSPG-k előfordulhatnak intracellulárisan, ECM-hez vagy
plazmamembránhoz kötődve, valamint szolubilis formában is (Yamamoto és mtsai,
2005). A syndecan-1 a leggyakrabban előforduló HSPG a mikrovaszkuláris
endothelsejtekben. A syndecan-4 is expresszálódik az endothelsejtek felszínén, míg a
perlecan az endothel ECM-ben gyakori. Az endothelsejtek tenyészetének mediumából
28-800 kD molekuláris tömegű, nehezen meghatározható szolubilis HSPG-ket izoláltak
(Kainulainen és mtsai, 1996).
Megfigyelték, hogy mikrovaszkuláris endothelsejtekben 10-15-ször nagyobb
mennyiségben vannak jelen HSPG-k, mint a makrovaszkuláris endothelsejtekben. Az
endothelsejtek felszíni HSPG-inek eltávolítása gátolja a neovaszkularizációt (Cavari és
Vannucchi, 1993).
Az angiogenezis HSPG-függő szabályozása részben összefügg az
angiogenetikus növekedési faktorok kötő kapacitásával és aktivitásuk módosításával.
Például a tisztított perlecan fokozza az FGF-2-függő angiogenezist, míg más HSPG-
knek nincs hatásuk az angiogenezisre (Knox és Whitelock, 2006). A NIH 3T3 sejtek
felszínén túlexpresszálódó syndecan-1 gátolja a FGF-2-indukált sejtproliferációt. Ezzel
ellentétben a syndecan-1 a mátrixhoz kötődve stimulálja az FGF-2-közvetített
sejtnövekedést. Azonban olyan mutáns sejteknél, melyek deficiensek sejtfelszíni HSPG-
re, a szolubilis syndecan és a glypican a heparin/HS-indukált FGF2-receptorkötést
gátolja. Ez azt jelenti, hogy a szolubilis HS eltávolítja a sejtfelszínről az FGF2-t, vagyis
vetélkedik a sejtfelszíni kötött HS-sel.
Ezek az adatok azt sugallják, hogy az angiogenesist negatívan befolyásoló

35
hatások inkább a szolubilis HSPG-k vagy GAG-ok angiogenetikus növekedési faktor-
kötésével magyarázhatók, mintsem a sejthez társult HSPG-k hatásával.
A receptorkötést módosító kapacitáson kívül a HSPG-k hatással lehetnek az
angiogenesisre azáltal, hogy megvédik az angiogenetikus növekedési faktorokat a hőtől
és a proteolitikus bomlástól, valamint növelik diffúziós hatósugarukat (Saksela és mtsai,
1988).
Végül az ECM-ben levő HSPG-k az angiogenetikus növekedési faktorok
rezervoárjaként szolgálhatnak, amelyek így magas helyi koncentrációt elérve az
endothelsejtek hosszú távú stimulálását biztosítják (Mongiat és mtsai, 2001).
Az a megállapítás, hogy az endothelsejtek HSPG-t expresszálnak az
angiogenezis során, nagy jelentőségű. Ennek a mechanizmusnak a szabályozása
különböző szinteken juthat érvényre: a HSPG molekulák típusainak kvantitatív és
kvalitatív differenciált expressziója révén, a GAG oldalláncok összetételének
változatossága révén, illetve a HSPG-k sejtfelszínről történő indukcióján és
mobilizációján keresztül érvényesül. (3. ábra)
3. ábra: HSPG-k és a növekedési faktorok közötti összefüggés az angiogenezisben.

36
4. CÉLKITŰZÉSEK
A fentiekben vázolt adatok alapján vizsgálataink a következő kérdések köré
csoportosulnak:
1. Hogyan változik mennyiségében és lokalizációjában az agrin expressziója a
cirrhotikus májban, összehasonlítva normál májszövettel?
2. Változik-e az agrin expressziója humán hepatocellularis carcinomában (HCC) a
normál májszövethez képest?
3. Kimutatható-e eltérés az agrin mRNS és fehérje expressziójában a HCC-ben és
cholangiocarcinomában (CC)?
4. Kimutatható-e agrin a jóindulatú májtumorokhoz sorolt hepatocellularis adenomában
(HCA) és fokális noduláris hyperplasiában (FNH)?
5. A májdaganatokban észlelt kifejezett neoangigenezist kíséri-e fokozott agrin
expresszió, felvetve ezáltal az agrin lehetséges szerepét neoangiogenezisben?
6. Található-e összefüggés az agrin-expresszió és a tumorok (HCC, CC)
differenciáltsága között?

37
5. ANYAG ÉS MÓDSZER
5.1. Beteganyag A Semmelweis Egyetem II. sz. Patológiai Intézetében archivált valamint újonnan
gyűjtött humán májmintákat használtuk fel vizsgálatainkhoz. Összességében 104 primer
májtumor (35 HCC, 35 CC, 14 HCA, 20 FNH) és környező normál vagy cirrhotikus
máj, valamint 10 balesetben vagy nem májbetegség miatt elhunyt emberből származó
normál májszövet került feldolgozásra (2.táblázat). A humán vizsgálatokat a betegek
írásos hozzájárulásával és az illetékes etikai bizottságok engedélyének birtokában
végeztük (TUKEB engedély 2/2004). A HCC-t illetően 19 jól differenciált, 11
közepesen differenciált, valamint 5 rosszul differenciált esetet vizsgáltunk. A
csoportosítás az Edmonson-Steiner féle klasszifikáció alapján történt (G1-G3). A 35
eset közül 15 esetben a hepatocellularis carcinoma cirrhosis talaján fejlődött ki. A
vizsgált betegek átlagéletkora 64,8 (42-82), a férfi/nő arány 27/8 volt. A CC-ket
intrahepatikus (n=7), Klatskin tumor (n = 9), valamint extrahepatikus (n = 19)
csoportokra osztottuk.
A sebészileg eltávolított tumormintákat a környező nem tumoros szövetekkel
együtt, valamint a normál máj mintákat 10% neutrális pufferolt formalinban fixáltuk 24
órán keresztül, utána dehidráltuk és paraffinba ágyaztuk. A rutin hisztológia, a
morfológiai elváltozások kimutatása és a tumor grade megítélése hematoxilin-eozinnal
(HE) festett 3-5 µm-os metszeteken történt. Diagnózis céljából a következő festéseket is
elvégeztük: alciánkék-PAS, Shikata-féle orcein, picrosirius-vörös.
Molekuláris biológiai vizsgálatokra a minták egy részet RNA-laterben (Sigma,
St. Louis, MO, USA) (13 HCC, 13 HCC tumormentes környező májszövet, 9 CC, 7 CC
tumormentes környező májszövet, 10 normál máj) fixáltuk, vagy folyékony nitrogénben
történt fagyasztás után -80°C-on archiváltuk.

38
2. táblázat. A vizsgálatokhoz használt beteganyag.
Vizsgált csoport Eset Kor Nem
CC 35 61,56 12M/23F
Intrahepatikus típus 7 53,14 3M/4F
Klatskin típus 9 61 2M/7F
Extrahepatikus típus 19 64,36 7M/12F
HCC 35 64,83 27M/8F
a. cirrhotikus talajú HCC 15
I b. nem cirrhotikus talajú HCC 20
1. jól differenciált HCC (G1) 19
2.közepesen differenciált HCC (G2) 11
II 3. rosszul differenciált HCC (G3) 5
HCA 14
FNH 20 Normál máj 10 24 6M/4F
Összesen 114
5.2. Módszerek
5.2.1. Fehérjeexpressziós vizsgálatok
5.2.1.1. Immunhisztokémiai vizsgálatok A formalinban fixált, paraffinba ágyazott anyagok antigén feltárása a deparaffinálást
követően proteáz emésztéssel (0,01% tripszin, 10 perc, szobahőn) vagy mikrohullámú
feltárással történt. Az agrin antigén-feltárása mikrohullámú sütőben, DAKO
antigénfeltáró oldatban (DAKO Cytomation, Glostrup, Dánia) történt. Ezt követően az
endogén peroxidáz aktivitásának gátlását 3%-os hidrogén peroxiddal (10 perc)
végeztük. Az antitest aspecifikus kötődését ló szérum (Novocastra, New Castle, UK)
inkubálással blokkoltuk (20 perc, szobahőn). 1:100 hígításban használtuk a
monoklonális agrin ellenes antitestet (clone 7E12, MAB 458, Chemicon International
Inc., Temecula, CA, USA), melyet egy glomerulus bazális membránból izolált HSPG
ellen termeltettek Kemeny és msai (1998). Az antitest specifitását munkacsoportunk
tömeg-spektrometriával igazolta (Tatrai és mtsai, 2006). A további antitesteket az alábbi
hígításokban használtuk: monoklonális laminin, (clone: Lam89) 1:500 (Novacastra,
New Castle, UK), monoklonális CD34 (clone QBEnd/10) 1:80 (Novocastra),
monoklonális Ki67 (clone MIB-1) 1:120 (DAKO), monoklonális CK7 (M7018, DAKO)

39
1:600, monoklonális CK19 (RCK108, DAKO) 1:100. Másodlagos antitestként
peroxidáz-jelölt IgG-t (DAKO) és biotinilált IgG-t (Vectastain Elite ABC Kit, Vector
Laboratories, USA) használtunk 1:200 hígításban, 30 percig szobahőn. Biotinilált
másodlagos antitest alkalmazása esetén ezt az avidin-biotin-komplexszel (ABC) történő
inkubálás követte (30 perc szobahőn). Az agrin esetében az immunreakció
érzékenységét biotinilált tiramiddal fokoztuk a Merz és munkatársai (1995) által leírt
módszer szerint (7mM biotinilált tiramid törzsből 1:125+0,033% H2O2, 10 min) (4.
ábra). Ismételt ABC reakció után az immunreakciót 0,05 mg/ml 3-amino-9-ethyl-
carbazol (AEC) (Biogenex HK 129-5K, San Ramon, USA) vagy diaminobenzidin
(DAB) (Genetex, San Antonio, USA) kromogénekkel hívtuk elő.
Háttérfestésként hematoxilint alkalmaztunk és glicerines fedőanyaggal fedtük a
metszeteket.
Pozitív kontrollként humán vesét használtunk, míg negatív kontrollként az elsődleges
ellenanyaggal való inkubálást kihagytuk és az előhívórendszer specifitásának
bizonyítására az elsődleges ellenanyagnak megfelelő nem immunszérumot használtunk
az elsődleges ellenanyagnak megfelelő hígításban.

40
4. ábra. A biotinilált tiramidos immunhisztokémiai reakció vázlata. Az elsődleges,
ill. biotinilált másodlagos antitestet és ABC-t követően a hozzáadott biotinilált tiramid
torma-peroxidáz hatására H2O2 jelenlétében aktiválódik. Az aktivált biotinilált tiramid
hozzákötődik a minta fehérjének tirozin oldalláncaihoz, és ABC-vel újra amplifikálható.
5.2.1.2. Immunfluoreszcens vizsgálat
A fagyasztott metszeteket metanol-aceton módszerrel fixáltuk (15 perc metanol –20°C-
on, 3 másodperc aceton), majd 20 percig blokkoltuk 5% BSA – PBS oldattal (pH:6.5).
A primer antitestek a következőek voltak: poliklonális agrin 2µg/ml (R&D Systems,
Abingdon, UK), poliklonális laminin 1:200 (DAKO), αν-integrin 10µg/ml (clone
P2W7, R&D Systems), poliklonális bFGF 1:50 (R&D Systems), smooth muscle α-actin
(SMA) 1:100 (clone 1A4, DAKO). A primer antitest reakcióját az Alexa Fluor
(Molecular Probes, Eugene, OR, USA) másodlagos antitesttel tettük láthatóvá. Az
előhívást mosási lépések követték, a vizsgálandó mintákat floureszcens fedővel
(DAKO) fedtük és BioRad MRC 1024 konfokális lézer scanning mikroszkóppal
vizsgáltuk (Bio-Rad, Hercules, CA, USA). Pozitív kontrolként humán veseszövetet
használtunk, míg negatív kontrolként primer antitest helyett 5% BSA-val inkubáltuk a
metszeteket.
5.2.1.3. Morfometriai elemzés
Az agrin immunhisztokémiai reakciókat fénymikroszkóp segítségével
fotódokumentáltuk (200X nagyítás, Olympus BX microscope). Tíz, egymást nem fedő
területet értékeltünk. Az agrin immunreakció mennyiségi meghatározására a Leica
QWin software-t (Leica Microsystems Imaging Solutions Ltd., Cambridge, UK)
használtuk fel. Az immunpozitív területet kijelölve végeztük el a méréseket. A vörös,
zöld, kék összetevők alapján állítottuk be a küszöbértékeket, amelyeket minden metszet
esetében állandónak fogadtunk el az összevethetőség érdekében.

41
5.2.1.4. Proteoglikán izolálása
A proteoglikán-izolálás azonos mennyiségű fagyasztott mintákból (2 HCC, 2
CC, 1 normál máj) történt Lyon és munkatársai (1994) módszere alapján. A mintákat
10ml homogenizáló pufferben (50 mM/L Tris-HCl, [pH 7.4], 10 mM EDTA, 5mM N-
ethylmaleimid [NEM], 0.5mM phenylmethylsulphonyl fluoride [PMSF], 3.4M NaCl,
5% Gordox) történő homogenizálás után inhibitorokat (5 mM NEM, 0.5 mM PMSF,
0,001%-os tripszin inhibitor, 5%-os Gordox) tartalmazó extraháló pufferrel (4 M
guanidin-HCl, 50 mM Na-acetát, 1% Triton X-100) felszuszpendáltuk és 2 napon át
rázattuk 4°C -on. Majd 4°C -on, 30 percig centrifugáltuk 11000 rpm-el, ezt követően a
felülúszót dializáltuk az extraháló pufferból 7 M-os urea pufferbe (10 mM Tris pH:7.4,
0.5% Triton X-100, 0.5 mM N-etilmaleimid, 0.5 mM PMSF, 0.001% tripszin inhibitor).
Minden alkalommal megmértük a vezetőképességet és összehasonlítottuk az eredeti 7
M-os urea puffer vezetőképességével. Amint az eredeti oldat vezetőképessége
megegyezett a végső oldat vezetőképességével, ioncserélő oszlopkromatográfiát
végeztünk a proteoglikán elválasztás céljából DEAE-cellulóz (diethylaminoethyl
cellulose) oszlopon. Az izolált anyagok tárolása -80°C-on történt. További analízishez a
proteoglikán frakciót abszolút alkohollal kicsaptuk, lecentrifugáltuk (12000 rpm, 4°C,
10 perc), 70% alkohollal mostuk és újra centrifugáltuk. Vákuumban történő beszárítás
után a proteoglikánt 3-15%-os poliakrilamid gradiens gélen futattuk meg. A futtatást
követően Alcian kék 8GX-el és Coomassie Brillant kék oldattal festettük meg a gélt.
5.2.1.5. Western blot analízis
A western blotthoz poliakrilamid gélen elválasztott fehérjéket elektrotranszferrel
nitrocellulóz membránra (Bio Rad, Hercules, CA, USA) blottoltuk. A blottolás 330
mA–rel 90 percig tartó elektrotranszferrel történt. A membránt TBS-ben oldott 5% (v%)
BSA-val blokkoltuk. Az agrin monoklonális antitestet 1:500 hígításban, 5%-os BSA-
ban higítva használtuk. A biotinilált másodlagos antitest és peroxidázzal jelölt ABC
inkubálást követően az előhívást kemilumineszcenciával (ECL, Amersham Biosciences,
UK) végeztük. A kiértékelést képanalizátorral (Kodak, Hemel Hempstead, UK)
végeztük.

42
5.2.2. mRNS szintű expressziós vizsgálatok
5.2.2.1. RNS izolálás RNS izolálás RNA later-ben fixált mintákból
A RNS izolálás RNA-laterben fixált (24 óra), majd –80 C°-on tárolt mintákból
történt. A kiindulási szövet mennyisége egyenként 200 mg volt (13 HCC, 13 HCC
tumormentes környező májszövet, 9 CC, 7 CC tumormentes környező májszövet, 10
normál máj). Homogenizálás után Trizol reagensben (Sigma, T 9424) inkubáltuk a
mintákat szobahőmérsékleten (15 percig). Steril Eppendorf-csövekbe pipettáztuk át a
folyadékot, majd 0,2 ml kloroform hozzáadása után vortexeltük és centrifugáltuk.
Ezután a felülúszót, amely az RNS-t tartalmazta, kloroformmal tovább tisztítottuk, majd
az RNS-t izopropanollal kicsaptuk, vortexeltük és tíz perc állás után újra centrifugáltuk.
Ezután leöntöttük a felülúszót és 75 %-os etanollal mostuk, majd centrifugáltuk. Roche
RNS tisztító kitből (Roche, Indianapolis, Indiana, USA) származó elúciós pufferben
feloldottuk az RNS-t, DNáz-zal emésztettük, majd a Roche RNS-tisztító kitjével
(Roche) tovább tisztítottuk, végül megmértük az optikai denzitását (O.D.) és az RNS
koncentrációját a kapott folyadéknak. További felhasználásig a tisztított RNS-t -80°C-
on tároltuk. A teljes RNS épségét 2%-os agaróz gélen ellenőriztük.
Reverz transzkripció
Az RNS (1µg 10µl mixben) átírása M-MulV reverz transzkriptázzal (Applied
Biosystems, Foster City, CA, USA) történt (10 perc 25ºC-on, 50 perc 42°C-on, és 5
perc 95ºC-on) RNáz inhibitor (Applied Biosystems) jelenlétében, Random Hexamerek
(Applied Biosystems) felhasználásával. Az RT-PCR reakcióhoz használt reakcióelegy
összetételét a 3. táblázat foglalja össze.

43
3. táblázat. Az RT-PCR reakcióhoz használt komponensek
Komponensek
Térfogat
A komponenst gyártó cég
RT Puffer 2 µl Boehringer 145376
Hexamer 0,5 µl Applied Biosystems, CA, USA
dNTP mix 0,5 µl Roche D 8220
20 U/ µl RN-áz gátló 0,5 µl Applied Biosystems, CA, USA
50U/ µl Reverz
Transzkriptáz
1 µl Applied Biosystems, CA, USA
dH2O 4,5 µl Eppendorf 0032006159
RNS templát 1 µg
Összesen 10 µl
5.2.2.2. Primertervezés és grádiens PCR
A kísérletek elvégzéséhez a PubMed adatbázisból rendelkezésre álló
szekvenciák alapján a Primer Expressz 2.00 program (ABI) segítségével megterveztük a
lehetőségek szerinti legideálisabb primereket. A tervezésnél figyelembe vettük, hogy
bizonyos fehérjéknek több transzkripciós variánsa is létezik; ezeknél olyan primereket
használunk, amely valamennyi transzkripciós variáns expressziójának kimutatására
alkalmas.
A PCR-hez megkerestük az optimális primer kötődési hőmérsékletet grádiens PCR-rel.
Ehhez a reakcióhoz MgCl2-at tartalmazó PCR puffert (Roche J 0051), dNTP mixet
(Applied Biosystems, D8220), forward és reverz primert, AmpliTaq Gold enzimet
(Roche, G 0448), desztillált vizet (Eppendorf, 0032006159) és a korábban előállított
cDNS-t használtuk fel. A reakció végtérfogata 25 µl. Az azonos tartalmú PCR csöveket
úgy helyeztük el a készülékben, hogy 50°C és 69,7°C közötti hőmérsékleten történjen
meg a primer kötődés. A reakció paraméterei: 95°C-on 10 perc, 95°C-on 20 másodperc,
60°C-on 60 másodperc, 72°C-on 60 másodperc, és 4°C Ciklusszám: 40. A
grádiens PCR reakcióelegy komponenseit a 4. táblázatban tüntettük fel.

44
4. táblázat. A grádiens PCR reakcióelegy komponensei
Komponensek
Térfogat
A kompnenst gyártó cég
10x PCR puffer MgCl2dal 2,5 µl Roche J 0051
10mM dNTP mix 0,5 µl Applied Biosystems, D8220
25 µM forwar primer 0,5 µl
25 µM forwar primer 0,5 µl
5U/ µl AmpliTaq Gold
Enzim
0,5 µl Roche G0448
dH2O 19,5 µl Eppendorf 0032006159
cDNS 1 µl
Összesen 25 µl
Agaróz gélelektroforézis A grádiens PCR reakciók során kapott PCR termékeket 2%-os agaróz gélben
futtattuk meg (5. ábra). A gélkészítésnél 1 g agarózhoz (GIBCO 15510-019) 50 ml 1x-
es Tris borát EDTA (TBE) puffert adtunk, majd forralással feloldottuk az agarózt és 2,5
µl etídium-bromidot (BIO-RAD 161 0433) adtunk hozzá. Gélöntő formába öntöttük,
szilárdulás után pedig a gélt 1x-es TBE pufferrel feltöltött futtatókádba tettük és a
létrehozott zsebekbe bemértünk 12 µl előre előkészített mintát. Az előkészítés úgy
történt, hogy 10 µl mintához 2 µl futtató puffert adtunk (Novagen 69180). A mintákkal
párhuzamosan a gélre 5 µl DNS létrát (Novagen 70539) vittünk fel. A futtatás 100 V-on
40 mA áramerősség mellett 20-30 percig történt. Ezután a SYNGENE MultiGenius Bio
Imaging rendszerével, a GeneSnap szoftver felhasználásával fotódokumentáltuk a gélt.
Ez alapján azt tapasztaltuk, hogy a primer optimális kötődési hőmérséklete 60°C (5.
ábra). A későbbiekben elvégzett real-time RT PCR reakcióban ezt állítottuk be primer
kötődési hőmérsékletként.

45
5. ábra. A grádiens PCR termékek 2%-os
agaróz gélben futtatva:
1. 50°C 2. 51.4°C
3. 55.5°C 4. 60.8°C
5. 66° 6. 69.7°C
5.2.2.3. mRNS expresszió vizsgálata real-time RT-PCR módszerrel
A real-time PCR reakciókhoz 2 µl cDNS-oldatot használtunk, a méréseket ABI
Prism 7000 készüléken (Applied Biosystems) végeztük. Minden egyes PCR reakciót 25
µl reakcióelegyben, SYBR Green PCR Master Mix (AB4309155, Foster City, USA)
felhasználásával végeztünk. A primer szekvenciák az alábbiak voltak: AGRN
(GI:51459422, 149bp), forward 5’-CATGGCTGTGTTCTTCATGTGTT-3’ 23, reverse
5’-GGTCATTCTGAGACAAGGACGACT-3’ 24; gliceraldehid 3-foszfát dehidrogenáz
(GAPDH) (GI:7669491, 141bp) forward 5’-CATGGGTGTGAACCATGAGAAGT-3’
23, reverse 5’-TGGACTGTGGTCATGAGTCCTT-3’ 22). A mérések 96-os plate-eken,
triplikátumokban történtek: 2 percig 95°C-on, majd 40 cikluson keresztül 95°C-on 20
másodperc, 60°C-on 30 másodperc és 72°C-on 30 másodperc. Az adatok elemzéséhez
és a relatív expresszió statisztikai kiértékeléséhez a REST-programot (relative
expression software tool, www.wzw.tum.de/gene-quantification) használtunk, a
génexpressziót minden gén esetében a GAPDH háztartásgénhez mint referenciagénhez
viszonyítottuk. A relatív expressziót az alábbi képlet alapján számítottuk ki:

46
R= a relatív expresszió
E =a PCR reakció hatékonysága (azaz ciklusonként észlelt PCR növekedés szorzója.
CP= crossing point, az a ciklusszám, ahol az RNS mennyisége meghaladja a
detektálhatóság küszöb (treshold) értékét.
t - treshold, küszöbérték: exponenciális fázisban;
Ct – treshold ciklus: hányadik ciklusban lépi át az adott minta fluoreszcenciája ezt a
szintet – a kiindulási DNS mennyiséggel arányos;
A PCR termékek méretének ellenőrzésére a reakció termékekből 10 µl-t
futtattunk 2%-os agaróz gélen, majd etídium-bromidos festéssel tettük láthatóvá az
amplikonokat (6. ábra).
A B
6. ábra. Az (A) ábrán az agaróz gélen futatott real-time RT-PCR termékek
láthatók. (B) A termékek olvadáspont görbéje látható.
5.2.2.4. Statisztikai kiértékelés

47
A statisztikai kiértékelést a GraphPad Prism 2.01 (GraphPad Software, Inc. San
Diego, CA, USA) software segítségével végeztük. A csoportok közötti
összehasonlításra a Mann-Whitney U tesztet alkalmaztuk. A különbséget p <0,05 esetén
tartottuk szignifikánsnak.
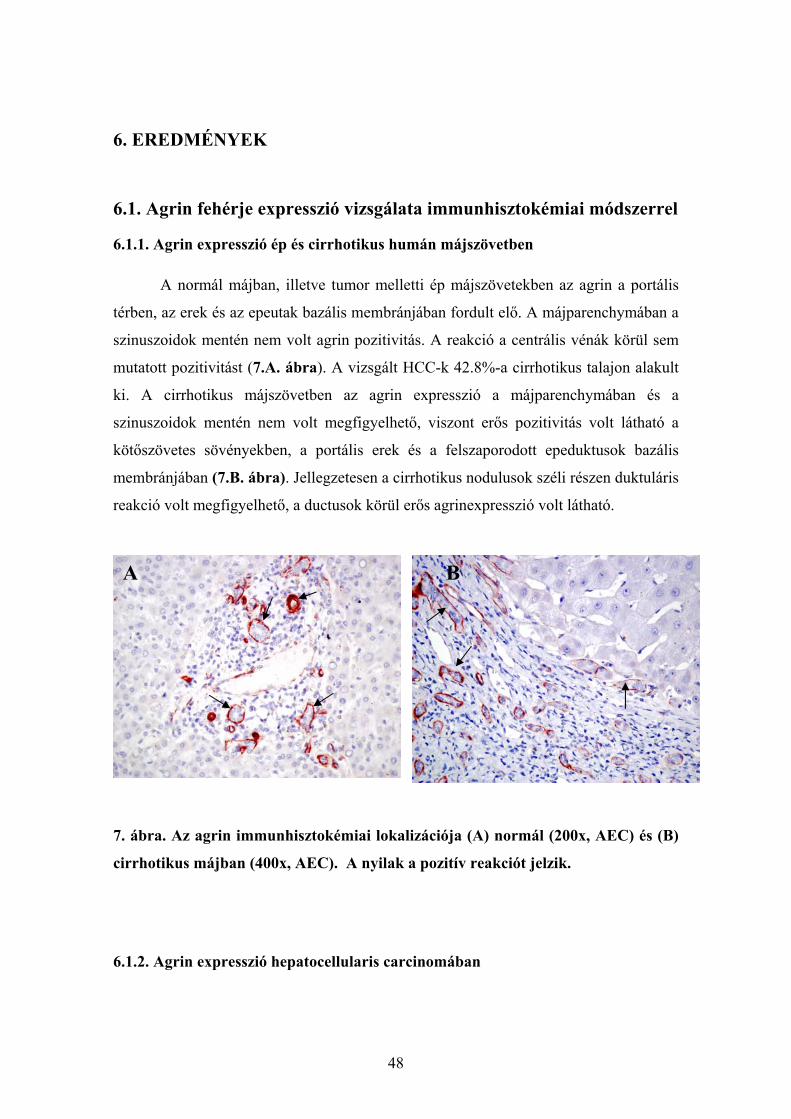
48
6. EREDMÉNYEK
6.1. Agrin fehérje expresszió vizsgálata immunhisztokémiai módszerrel
6.1.1. Agrin expresszió ép és cirrhotikus humán májszövetben A normál májban, illetve tumor melletti ép májszövetekben az agrin a portális
térben, az erek és az epeutak bazális membránjában fordult elő. A májparenchymában a
szinuszoidok mentén nem volt agrin pozitivitás. A reakció a centrális vénák körül sem
mutatott pozitivitást (7.A. ábra). A vizsgált HCC-k 42.8%-a cirrhotikus talajon alakult
ki. A cirrhotikus májszövetben az agrin expresszió a májparenchymában és a
szinuszoidok mentén nem volt megfigyelhető, viszont erős pozitivitás volt látható a
kötőszövetes sövényekben, a portális erek és a felszaporodott epeduktusok bazális
membránjában (7.B. ábra). Jellegzetesen a cirrhotikus nodulusok széli részen duktuláris
reakció volt megfigyelhető, a ductusok körül erős agrinexpresszió volt látható.
7. ábra. Az agrin immunhisztokémiai lokalizációja (A) normál (200x, AEC) és (B)
cirrhotikus májban (400x, AEC). A nyilak a pozitív reakciót jelzik.
6.1.2. Agrin expresszió hepatocellularis carcinomában
A B
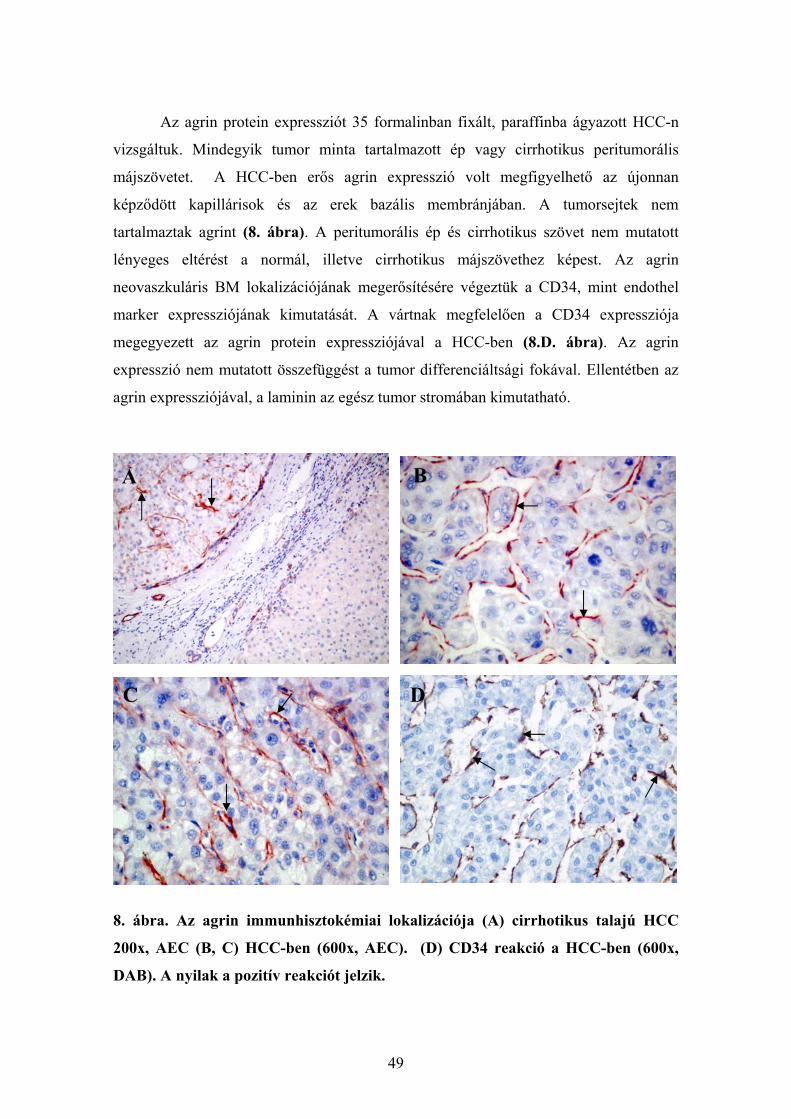
49
Az agrin protein expressziót 35 formalinban fixált, paraffinba ágyazott HCC-n
vizsgáltuk. Mindegyik tumor minta tartalmazott ép vagy cirrhotikus peritumorális
májszövetet. A HCC-ben erős agrin expresszió volt megfigyelhető az újonnan
képződött kapillárisok és az erek bazális membránjában. A tumorsejtek nem
tartalmaztak agrint (8. ábra). A peritumorális ép és cirrhotikus szövet nem mutatott
lényeges eltérést a normál, illetve cirrhotikus májszövethez képest. Az agrin
neovaszkuláris BM lokalizációjának megerősítésére végeztük a CD34, mint endothel
marker expressziójának kimutatását. A vártnak megfelelően a CD34 expressziója
megegyezett az agrin protein expressziójával a HCC-ben (8.D. ábra). Az agrin
expresszió nem mutatott összefüggést a tumor differenciáltsági fokával. Ellentétben az
agrin expressziójával, a laminin az egész tumor stromában kimutatható.
8. ábra. Az agrin immunhisztokémiai lokalizációja (A) cirrhotikus talajú HCC
200x, AEC (B, C) HCC-ben (600x, AEC). (D) CD34 reakció a HCC-ben (600x,
DAB). A nyilak a pozitív reakciót jelzik.
A B
C D
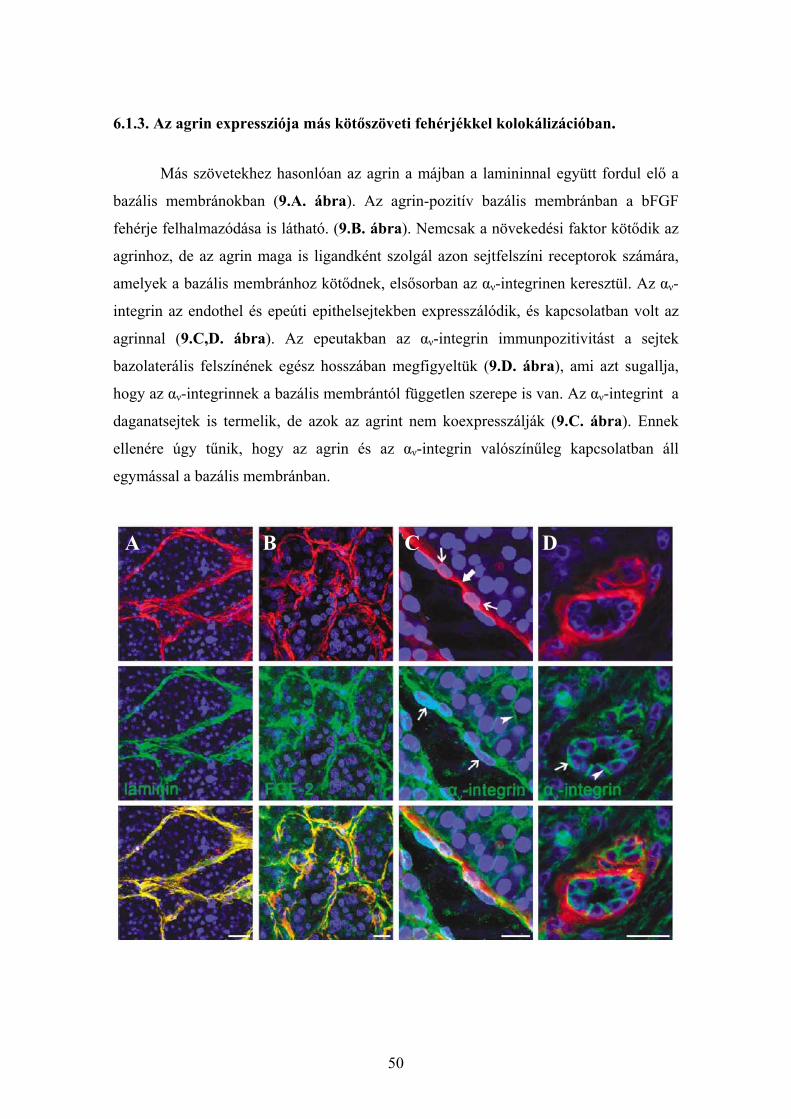
50
6.1.3. Az agrin expressziója más kötőszöveti fehérjékkel kolokálizációban.
Más szövetekhez hasonlóan az agrin a májban a lamininnal együtt fordul elő a
bazális membránokban (9.A. ábra). Az agrin-pozitív bazális membránban a bFGF
fehérje felhalmazódása is látható. (9.B. ábra). Nemcsak a növekedési faktor kötődik az
agrinhoz, de az agrin maga is ligandként szolgál azon sejtfelszíni receptorok számára,
amelyek a bazális membránhoz kötődnek, elsősorban az αν-integrinen keresztül. Az αν-
integrin az endothel és epeúti epithelsejtekben expresszálódik, és kapcsolatban volt az
agrinnal (9.C,D. ábra). Az epeutakban az αν-integrin immunpozitivitást a sejtek
bazolaterális felszínének egész hosszában megfigyeltük (9.D. ábra), ami azt sugallja,
hogy az αν-integrinnek a bazális membrántól független szerepe is van. Az αν-integrint a
daganatsejtek is termelik, de azok az agrint nem koexpresszálják (9.C. ábra). Ennek
ellenére úgy tűnik, hogy az agrin és az αν-integrin valószínűleg kapcsolatban áll
egymással a bazális membránban.
A B C D

51
9. ábra. Az agrin lehetséges molekuláris partnerei. Kettős immunfloureszcens
vizsgálat fagyasztott humán HCC mintákon (A-C) és a cirrhotikus májszövetben (D);
konfokális felvételek. Piros: agrin (R&D System), zöld: a képen látható feliratoknak
megfelelő struktúrákat jelöli, kék: sejtmag. Az összeadott képekben, a legalsó sorban a
sárga szín kolokalizációt/átfedést jelent. Az agrin a bazális membránban a lamininhoz
kötődve immobilizálódik (A); az agrin képes kötni FGF-2 szerű növekedési faktorokat
(B). (C) A tumor vérerekben az endothelsejtek által expresszált αν-integrin (vékony nyíl
a középső soron) receptorként szerepelhet a BM-ban expresszálódó agrin számára
(vastag nyíl a felső soron). (D) A biliaris epithel sejtek a bazolaterális membrán egész
hosszában expresszálják az αν-integrint (bazális felszín: vékony nyíl; laterális felszín:
nyílhegy). Az agrin a bazális felszínen látható. Hosszmértékek: a: 50µm, b:25µm, c, d:
20µm.
6.1.4. Agrint termelő sejtek májelváltozásokban
Szemmel láthatóan az agrin reakció erőssége az artériák és vénák falaiban erősen
korrelál az érfal izomrétegének vastagságával. Az érfal simaizomsejtei teljes átfedést
mutatnak az agrin felhalmozódásával (10. ábra). Ennek alapján úgy tűnik, hogy a
simaizomsejtek szekretálják az agrint a sejteket burkoló bazális membránba, és az is
elképzelhető, hogy az endothelium bazális membránjába is. Noha az epeutakat nem
veszik körül szorosan a simaizom-aktin (SMA) - pozitív sejtek, ennek ellenére az
epeutakat övező bazális membrán erős agrin- immunreakciót mutat. Ezért,
feltételezhető, hogy az epeúti hámsejtek is termelnek az agrint.
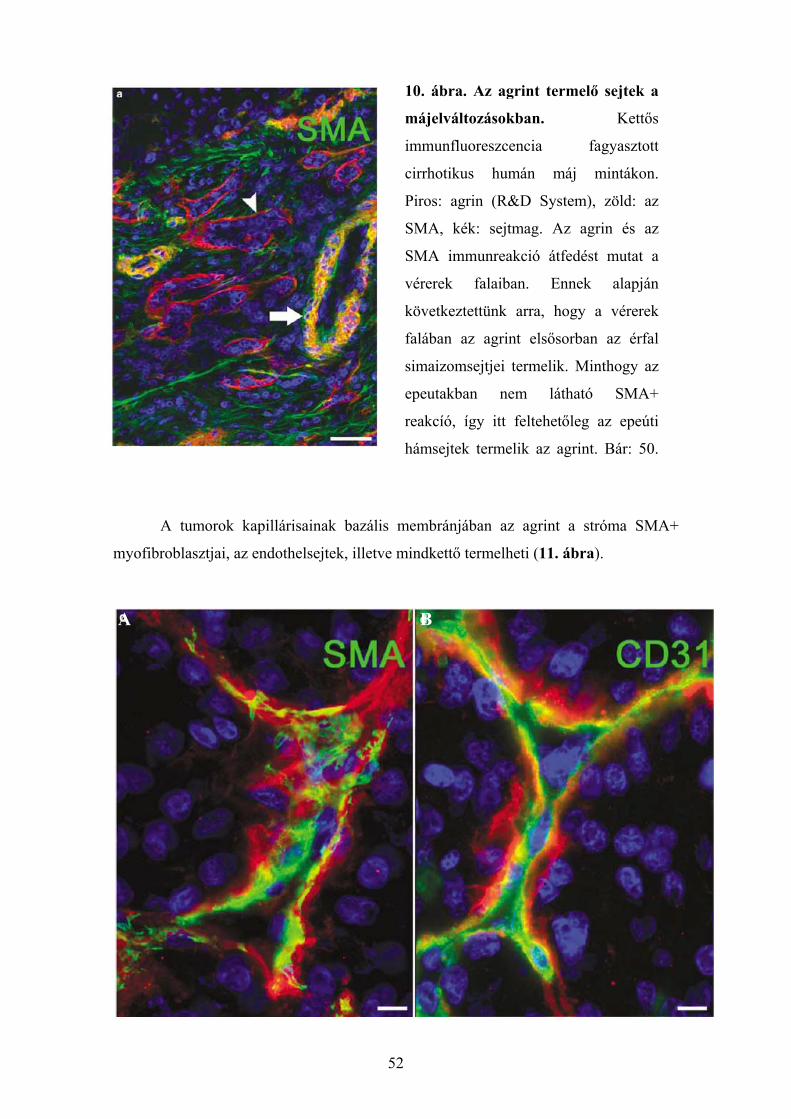
52
A tumorok kapillárisainak bazális membránjában az agrint a stróma SMA+
myofibroblasztjai, az endothelsejtek, illetve mindkettő termelheti (11. ábra).
A B
10. ábra. Az agrint termelő sejtek a
májelváltozásokban. Kettős
immunfluoreszcencia fagyasztott
cirrhotikus humán máj mintákon.
Piros: agrin (R&D System), zöld: az
SMA, kék: sejtmag. Az agrin és az
SMA immunreakció átfedést mutat a
vérerek falaiban. Ennek alapján
következtettünk arra, hogy a vérerek
falában az agrint elsősorban az érfal
simaizomsejtjei termelik. Minthogy az
epeutakban nem látható SMA+
reakcíó, így itt feltehetőleg az epeúti
hámsejtek termelik az agrint. Bár: 50.

53
11. ábra. Kettős immunfluoreszcencia humán HCC fagyasztott metszeten. A HCC
stromájában mind az SMA-pozitív mesenchymális sejtek mind a CD31-pozitív
endotheliális sejtek a bazális membránnal határosak. Hosszmérték: 10 µm
6.1.5. Az agrin expressziójának vizsgálata a cholangiocarcinomában
Immunhisztokémiával az agrin protein expressziót 35 formalinban fixált és
paraffinba ágyazott CC-n vizsgáltuk. CC-s mintákból intrahepatikus (n=7), Klatskin
(n=9), és extrahepatikus (n=19) CC-ket vizsgáltunk. Egyes minták a lokalizáció folytán
nem tartalmaztak peritumorális szövetet. Az epeúti rákokat a WHO kritériumok szerint
osztályoztuk jól differenciált (n=5), közepesen differenciált (n=24), valamint rosszul
differenciált (n=6) tumorokra.
Erős agrin expresszió figyelhető meg a tumorsejtekből álló glanduláris és
trabekuláris szövetszerkezetet körülvevő bazális membránban a jól és közepesen
differenciált CC-ákban (12.A, B, E ábra). Ezzel szemben a rosszul differenciált
tumorokban, valamint a környezetbe betörő tumorszöveteknél az agrin expresszió
fragmentálódott, csökkent, sőt el is tűnik (12.C, D ábra). A tumorsejtekben, valamint a
lamininnal ellentétben a tumor stromájában sem volt megfigyelhető agrin expresszió.
A CD34 endothel-marker expressziója megegyezett az agrin expresszióval a
tumor stroma ereiben, viszont nem volt megtalálható a tumorban a glanduláris és
trabekuláris struktúrákat körülvevő bazális membránban (12.F ábra).
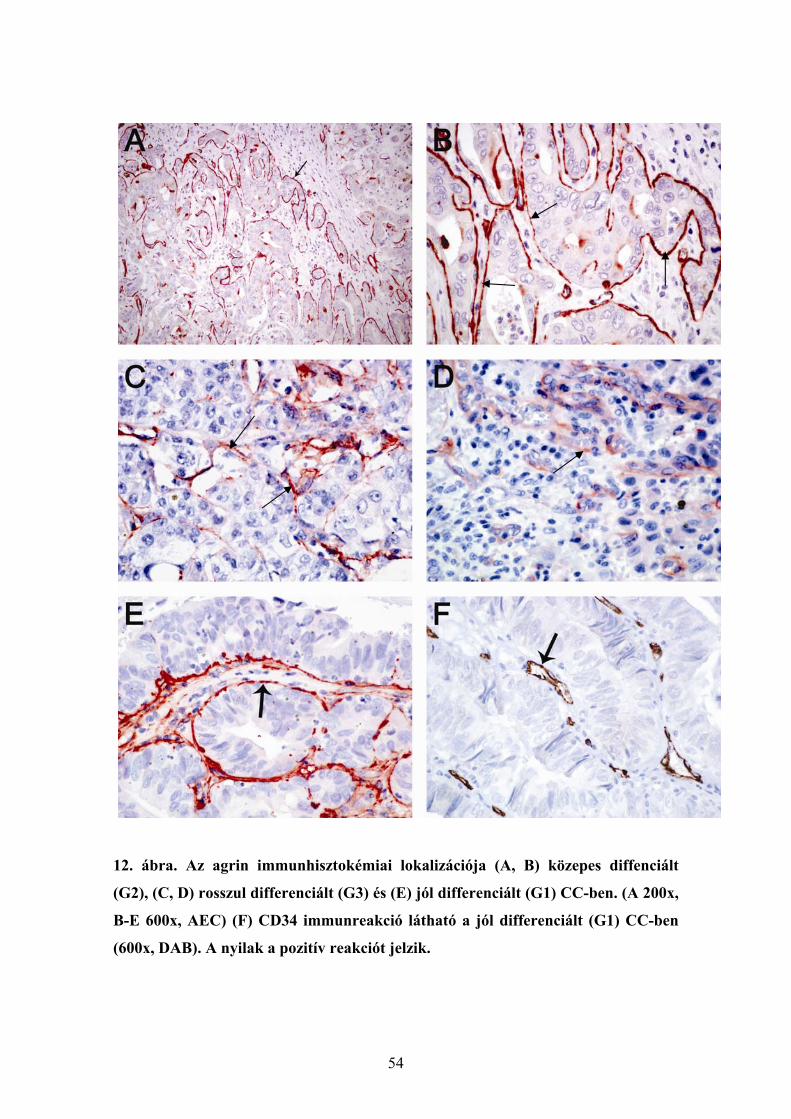
54
12. ábra. Az agrin immunhisztokémiai lokalizációja (A, B) közepes diffenciált
(G2), (C, D) rosszul differenciált (G3) és (E) jól differenciált (G1) CC-ben. (A 200x,
B-E 600x, AEC) (F) CD34 immunreakció látható a jól differenciált (G1) CC-ben
(600x, DAB). A nyilak a pozitív reakciót jelzik.

55
Az agrin immunhisztokémiai reakciónak morfometriai elemzése
Az agrin-pozitív területek kvantitatív morfometriai elemzése az összterület
százalékában a következő eredményeket hozta: Normál máj: 0.22 ± 0.029%, HCC: 1.55
± 0.121%, CC: 3.72 ± 0.277%. Ezek az eredmények azt mutatják, hogy CC-ben
szignifikánsan (p<0,0001) több agrin expresszálódik, mint a HCC-ben. Mind a HCC,
mind a CC szignifikánsan (p<0,0001) több agrint tartalmaz, mint a normál máj (13.A.
ábra). A jól és közepesen differenciált CC-ben az immunpozitív területek aránya
szignifikánsan (p< 0.0001; p< 0.0001) magasabb volt a rosszul differenciált tumorokhoz
viszonyítva. A jól és közepesen differenciált tumorok között nem volt szignifikáns
különbség (13.B. ábra).
13. ábra. Az agrin immunhisztokémiai reakciónak morfometriai elemzése HCC-
ben, CC-ben és normál májban. (A) Normál, CC és HCC csoportok összehasonlítása.
(B) Különböző differenciáltságú CC-k összehasonlítása. A * a csoportok közötti
szignifikáns eltérést jelöli.
6.1.6. Az agrin expresszió vizsgálata hepatocellularis adenomában Tizennégy hepatocellularis adenomában (HCA) vizsgáltuk az agrin fehérje
expressziót immunhisztokémiai módszerrel. Az agrin expressziója a tumorokon belül
eltéréseket mutat. Egyes területeken erős immunpozitivitás látható a szinuszoidoknak
megfelelő képletek mentén (14.A,C. ábra), míg máshol a tumorszövet teljesen negatív
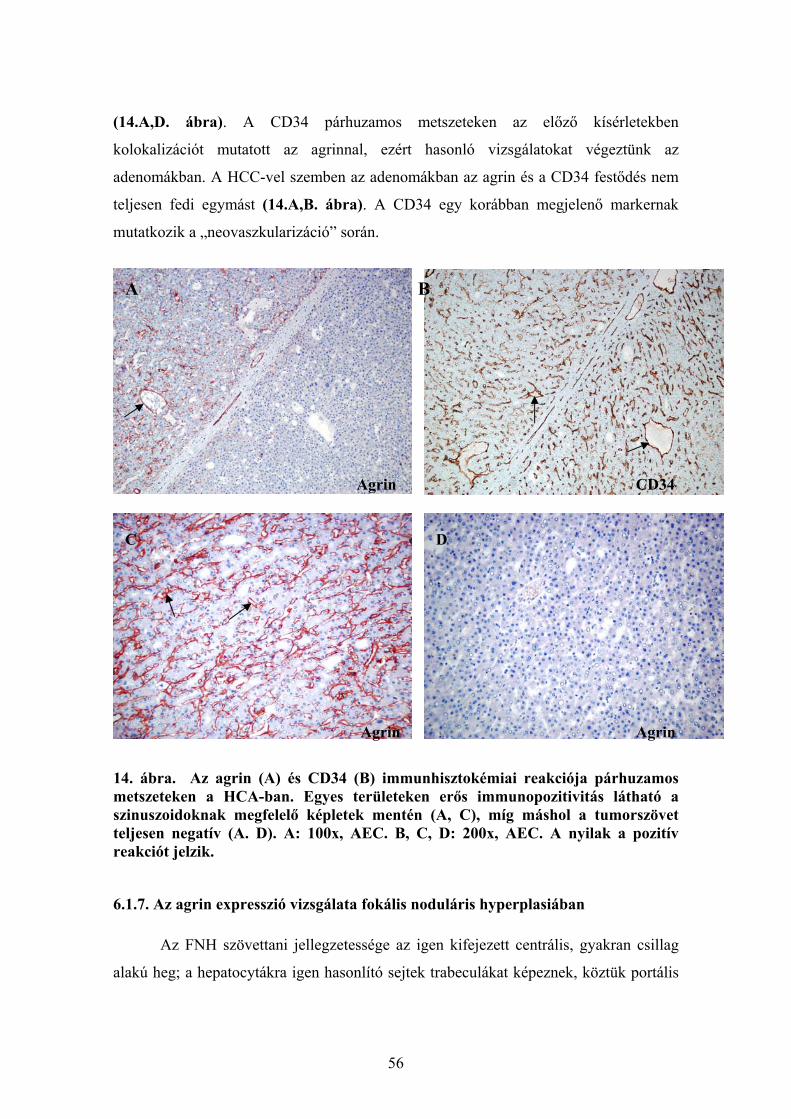
56
(14.A,D. ábra). A CD34 párhuzamos metszeteken az előző kísérletekben
kolokalizációt mutatott az agrinnal, ezért hasonló vizsgálatokat végeztünk az
adenomákban. A HCC-vel szemben az adenomákban az agrin és a CD34 festődés nem
teljesen fedi egymást (14.A,B. ábra). A CD34 egy korábban megjelenő markernak
mutatkozik a „neovaszkularizáció” során.
A B
Agrin CD34
C D
Agrin Agrin
14. ábra. Az agrin (A) és CD34 (B) immunhisztokémiai reakciója párhuzamos metszeteken a HCA-ban. Egyes területeken erős immunopozitivitás látható a szinuszoidoknak megfelelő képletek mentén (A, C), míg máshol a tumorszövet teljesen negatív (A. D). A: 100x, AEC. B, C, D: 200x, AEC. A nyilak a pozitív reakciót jelzik.
6.1.7. Az agrin expresszió vizsgálata fokális noduláris hyperplasiában
Az FNH szövettani jellegzetessége az igen kifejezett centrális, gyakran csillag
alakú heg; a hepatocytákra igen hasonlító sejtek trabeculákat képeznek, köztük portális
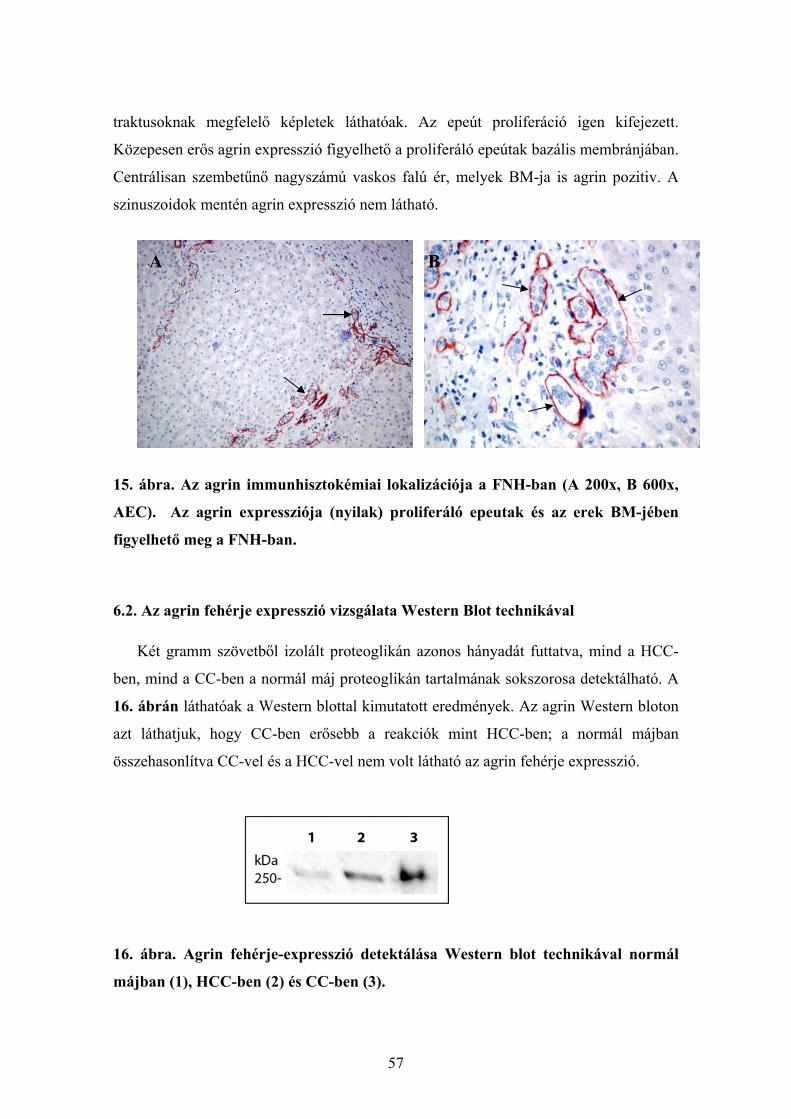
57
traktusoknak megfelelő képletek láthatóak. Az epeút proliferáció igen kifejezett.
Közepesen erős agrin expresszió figyelhető a proliferáló epeútak bazális membránjában.
Centrálisan szembetűnő nagyszámú vaskos falú ér, melyek BM-ja is agrin pozitiv. A
szinuszoidok mentén agrin expresszió nem látható.
A B
15. ábra. Az agrin immunhisztokémiai lokalizációja a FNH-ban (A 200x, B 600x,
AEC). Az agrin expressziója (nyilak) proliferáló epeutak és az erek BM-jében
figyelhető meg a FNH-ban.
6.2. Az agrin fehérje expresszió vizsgálata Western Blot technikával
Két gramm szövetből izolált proteoglikán azonos hányadát futtatva, mind a HCC-
ben, mind a CC-ben a normál máj proteoglikán tartalmának sokszorosa detektálható. A
16. ábrán láthatóak a Western blottal kimutatott eredmények. Az agrin Western bloton
azt láthatjuk, hogy CC-ben erősebb a reakciók mint HCC-ben; a normál májban
összehasonlítva CC-vel és a HCC-vel nem volt látható az agrin fehérje expresszió.
16. ábra. Agrin fehérje-expresszió detektálása Western blot technikával normál
májban (1), HCC-ben (2) és CC-ben (3).

58
6.3. Az agrin mRNS kimutatása real time RT-PCR-vel Az „Anyagok és módszerek” fejezetnek megfelelően, a real-time-RT PCR-rel
kapott adatokat egy relatív kvantifikációra alkalmas szoftver segítségével értékeltük ki.
Referencia génként a GAPDH-t használtuk.
Az 17. ábrán látható, hogy HCC-ben a normál májhoz képest az agrin mRNS
expressziója fokozott, de nem mutatott szignifikáns változást. A CC mintákban az agrin
szignifikánsan magasabb volt mind a normál májban (10,75-szer), mind a HCC-ben
(3,4-szer). CC-t környező tumormentes szövetekben az agrin mRNS szintje magasabb,
mint a normál májban. A CC-ben, annak tumormentes környezetéhez viszonyítva 2.8-
szoros emelkedést mértünk, bár ez statisztikailag nem volt szignifikáns (17. ábra).
A PCR vizsgálat megerősítette az immunhisztokémiai vizsgálat eredményeit: az
agrin fehérje- és mRNS- expressziója CC-ben szignifikánsan magasabb volt a normál
májhoz és HCC-hez képest, és emelkedést mutatott a környező tumormentes
szövetekhez viszonyítva.
A real-time RT-PCR eredmények számértékeit a 4. táblázatban foglaltuk össze.
*
*
17. ábra. Az agrin expresszió real-time RT-PCR reakció eredmények alapján
kimutatott mennyiségi eltérései különböző csoportokban. Az oszlopok az agrin
mRNS expresszió n-szeres mennyiségi különbséget ábrázolják a vizsgált csoportokban

59
a kontrollnak vett csoportokhoz képest. Minden esetben az aláhúzott csoportot
hasonlítjuk a másik csoporthoz; N: normál máj, HCC: hepatocellularis carcinoma, CC:
cholangiocarcinoma, kHCC: HCC tumormentes környező májszövet, kCC: CC
tumormentes környező májszövet. *- szignifikáns különbséget jelöli.
4. táblázat. Az agrin mRNS expresszió mennyiségi összehasonlítása real-time RT-
PCR reakció alapján.
Csoportok Agrin relatív mRNS expresszió
n-HCC U 2,271
NS, P=0,089
n-CC U 10,755
S, P=0,001
HCC-CC U 3,374
S, P=0,022
n-kHCC U 1,723
NS, P=0,238
kHCC-HCC D 1,003
NS, P=0,85
kCC-CC U 2,807
NS, P=0,0545
n-kCC U 1,838
NS, P=0,29
kHCC-kCC U 1,199
NS P=0,77 N: normál máj, HCC: hepatocellularis carcinoma, CC: cholangiocarcinoma, kHCC:
HCC tumormentes környező májszövet, kCC: CC tumormentes környező májszövet. Az
U – „ upregulált”, D- „ downregulált”, NS-nincs szignifikáns eltérés, S – szignifikáns
eltérés,

60
7. MEGBESZELÉS
A malignusan transzformált sejtek a gazdaszervezeten belül túlélésre, sőt
szaporodásra és invázióra képesek. Ez jelzi, hogy a daganatsejtek mielőtt elpusztítják,
azelőtt leigázzák a gazdaszervezetet, azaz áthangolják különböző sejtjeinek,
sejtvonalainak működését: kikapcsolják a védekező mechanizmusokat, saját
szolgálatukba állítják a gazdaszervezet sejtjeit. Utóbbira példa a daganatnövekedés
egyik domináns mozzanata, a tumormátrix-képződés folyamata, azaz a daganatsejtek
sajátos mikrokörnyezetének kialakulása, mely nem pusztán a tumorsejtek
gazdaszervezeten belüli túlélésének, de kontrollálatlan szaporodásának színtere. Ma már
egyre sokasodnak az irodalomban azok a közlések, amelyek a tumormátrixot – ezen
belül is elsősorban a proteoglikánok depozícióját – terápiás intervenciók célpontjaként
jelölik meg, hangsúlyozván, hogy a gazdaszervezet stromális sejtjeinek áthangolódását
kellene megakadályozni, vagyis gátolni a tumorsejteket túléltető mikrokörnyezet
kialakulását.
A HSPG-k alkalmasak számos szabályozó- illetve növekedési faktor kötésére,
tárolására, képesek azokat aktiválni vagy inaktiválni, prezentálni, ko-receptorként
felkínálni a tumorsejtek felszíni receptorai számára, így minőségbeli és mennyiségbeli
változásaik új, a normálistól eltérő információk közvetítését eredményezhetik. Ilyen
funkcionális változások tisztázásának előfeltétele a daganatos proteoglikán expresszió
és az azt befolyásoló faktorok pontos feltérképezése. A megváltozott proteoglikán
expressziónak mind a daganatok kialakulásában, mind pedig progressziójában szerepe
lehet.
A májbetegségekben a proteoglikánok képződésében és szerkezetében szembetűnő
változás következik be.
A bazális membrán HSPG családjába tartozó proteoglikánok közül egyesekről
már bebizonyították, hogy fontos szerepük van a különböző típusú daganatok
pathogenezisében. E csoport legnagyobb képviselőjéről, a perlecanról már leírták, hogy
részt vesz a tumor terjedésében, a metasztázis-képződésben, valamint a
neoangiogenezisben. A perlecan mRNS szintje jelentősen emelkedett a humán
metasztatizáló melanomában a normál szövethez képest. A kórosan expresszálódott
perlecan a pericelluláris mátrixban lerakódik és megjelenése a melanoma sejtek erős

61
metasztatizáló képességével összefügghet (Marchetti és mtsai, 1993). A perlecan HS
láncairól kiderült, hogy különböző növekedési faktorokkal (pl: bFGF) kölcsönhatásba
lépve stimulálják a tumornövekedést és az érképződést (Aviezer és mtsai, 1994).
Bebizonyították, hogy a perlecannak szerepe van az intrahepatikus CC (Sabit és mtsai,
2001), emlő (Iozzo és mtsai, 1994), petefészek, nyálmirigy (Kimura és mtsai, 2000),
colon (Sharma és mtsai, 1998) valamint a prosztata rák (Jiang és Couchman, 2003)
kialakulásban és terjedésben. Érdékes, hogy a perlecan C-terminális része, az
endorepellin gátolja az angiogenesist (Mongiat és mtsai, 2003).
A BMHSPG-k nemrég felfedezett másik tagjáról, a collagen XVIII-ról
kimutatták, hogy egy fragmense, az endostatin mind in vivo, mind in vitro anti-
angiogenikus hatással bír (Marneros és Olsen, 2005).
Kevesebbet tudunk az agrinnak daganatok kialakulásában és
progressziójában játszott szerepéről. Csupán a glioblastoma multiforméban a
vaszkuláris barrier érésében és differenciálódásában való részvételét írták le (Rascher és
mtsai, 2002).
Mind az agrin, mind a perlecan megtalálható a BM-ban, és mindkettőt kódoló
gén az 1. emberi kromoszoma rövid karján helyezkedik el. Az agrin C-terminális része a
perlecanéhoz hasonló, ezért feltehetőleg a perlecanhoz hasonlóan szerepe lehet
különböző sejtproliferációs folyamatokban.
Számos vizsgálat feltárta a neuronalis agrin szerepét embriógenezis során (Kim és
mtsai, 2007). Felnőttekben a non-neuronális szövetek közül a tüdőben és a vesében levő
agrinnal foglalkoztak (Groffen és mtsai, 1998). Az agrin májban, ill. májbetegségek
során bekövetkező mennyiségi és minőségi változásairól eddig nem jelent meg
közlemény.
Munkacsoportunkkal az agrin expresszióját vizsgáltuk a különböző májbetegségekben,
elsősorban a primer májtumorokban.
A normál májszövet egyik fontos jellemzője, hogy nagyon kevés proteoglikánt
tartalmaz. Ezek többsége sejtfelszíni, illetve transzmembrán HSPG, és elhanyagolható
az ECM proteoglikánok mennyisége. A formalin-fixált, paraffinba ágyazott normál
humán májakon az agrin fehérje komponensét immunhisztokémiával detektáltuk. Az
agrin az epeutak BM-jában, a portális artériák és vénák BM-jában volt erős intenzitással

62
kimutatható, míg a centrális vénában, a nyirokerek BM-jában immunohisztokémiai
módszerrel nem volt detektálható. Az agrin immunhisztokémiai megjelenése a
perlecanétól abban különbözik, hogy a perlecan periszinuszoidális pozitivitást is mutat a
normál májban.
A normál májszövetben látottakat összehasonlítva a májrák körüli, látszólag ép,
cirrhosismentes májszövettel nem volt különbség az agrin immunhisztokémiájában.
Cirrhotikus májban fokozódik az agrin depozíciója a proliferáló epeutakban. A
fibrotikus szeptumokban megfigyelhető reakció (erek, epeutak BM) mellett a
cirrhotikus göbök széli részen bizonyos sejtcsoportok körül megjelenik az agrin
pozitivitás, ami a duktularis reakciónak felelhet meg. Maga a duktularis reakció a
károsult májszövetek regenerációjának részjelensége, amelynek következtében
periportális ill. cirrhotikus nodulusok széli részen levő őssejtek aktiválódnak és
hepatocytákka differenciálódnak (Desmet és mtsai, 1995). Az agrin e nodulusok körüli
megjelenésnek feltehetőleg szerepe lehet az őssejtek differenciálódásában. Egyes
esetekben a cirrhotikus göbök közepén enyhe periszinuszoidális agrin pozitivitás jelenik
meg amely a szinuszoidalis cirrhotikus májkárosodás egyik legsúlyosabb eseménye, ez
vezet végső soron a hepatocyták elégtelen funkciójához .
Kérdés, hogy az agrin mennyiségének és összetételének változása pusztán mint a
szövetproliferáció egyik részjelensége értékelendő, vagy ennél többről van szó és ezek a
változások hatással vannak a cirrhosis kimenetelére is. Több adat utal arra, hogy
bizonyos proteoglikánok részesei a mátrixképződés szabályozásának. Ismert, hogy a
decorin a kollagén rostokhoz kötődve gátolja azok érését, tehát a kötőszövetképzés
gátlásának irányában hat. A decorin a TGFβl inaktiválására képes, mely a növekedési
faktorról köztudott, hogy a fibrogenezis egyik fő stimulátora (Hildebrand és mtsai,
1994; Kovalszky és mtsai, 1998).
Az FNH a máj daganatszerű elváltozásai közé tartozik. Jellemző, hogy a
hepatocytákra igen hasonlító sejtek trabekulákat képeznek, körülöttük kifejezett epeút-
proliferáció látható. Az agrin itt a proliferált epeutak bazális membránjában található.
Jellegzetes, hogy FNH-ban a nodulusok széli részén a duktularis reakció körül fokozott
agrin pozitivitás látható. A proliferáló epeúti sejtek eredete nem tisztázott, szerepe lehet
benne a differenciálódó őssejteknek.

63
A humán májtumorok esetében az agrin fehérje-expresszió fokozott, de
megjelenése tumor típusától függően váltózó.
A HCA-ban változatos az agrin expresszió. Egyes tumorok erős agrin pozitivitást
mutatnak a tumor-szinuszoidoknak megfelelő képletek és az erek BM-je mentén. Ez a
jelenség a tumoron belül is változó erősségű, a tumorok egyes részei teljesen negatívak.
Ezzel ellentétben az immunhisztokémiai reakció CD34-gyel kiterjedtebb szinuszoidalis
pozitivitást mutat az adenomákban. A CK7 viszont az adenomákon belül teljesen
negatív, ami arra utal, hogy nincsenek bennük epeutak. A fentiekből következik, hogy
az adenomán belüli neovaszkularizációs folyamatnak több lépcsője lehet. Ott, ahol csak
a CD34 pozitivitás mutatható ki, csak az endothelsejtek képeznek érfalat, míg az agrin-
pozitív területeken már fokozottabb strukturális átrendeződés alakulhatott ki.
A HCC-ben intenzív agrin fehérje-expressziót találtunk az újonnan képződött
kapillárisok és az erek bazális membránjában. A HCC-ben nincsenek epeutak, és
vaszkularizáció indul meg a tumor területén. Az így kialakuló erek biztosítják a tumor
tápanyagellátását. Ezen újonnan képződött kapillárisok és az erek bazális membránjában
agrin immunpozitivitás figyelhető meg. Az agrin neovaszkuláris BM lokalizációjának
megerősítésére végzett CD34 és CD31, mint endothel-markerek immunohisztokémiai
reakciója kolokalizációt mutatott az agrinéval a HCC-ben. Az agrin mint HSPG
feltehetően képes kötni különböző növekedési faktorokat, beleértve az FGF-2-t is.
Ennek bizonyítására végeztünk immunfloureszcens vizsgálatot a HCC-n FGF-2- és
agrin-ellenes antitestekkel. Elképzeléseinknek megfelelően az agrin pozitív BM-ben az
FGF-2-t találtunk.
Az agrin-expresszió a tumor differenciáltságával nem mutat összefüggést HCC-ben, bár
a jól differenciált HCC erős trabekuláltságának megfelelően nagyon erezett, ezért
erősen agrin pozitív.
Mindebből az következtethető ki, hogy az agrinnak szerepe van az
angiogenesisben. Feltehetően az agrin HS láncai különböző növekedési faktorokat
(bFGF, VEGF, stb) kötve stimulálják a neovaszkularizációt és a tumornövekedést. Ezen
kívül az agrin azáltal is segítheti a neoangiogenesist, hogy a különböző növekedési
faktorok agrinhoz való kötődése megvédi őket a proteolitikus bomlástól és a hőtől
(Saksela és mtsai, 1988). Lehetséges, hogy az agrin nem csak HS láncaival vesz részt az
angiogenetikus és proliferaciós folyamatokban, hanem közvetlenül vázfehérjéje

64
segítségével is, mivel ezzel kapcsolódni tud az integrinhez és a lamininhoz. Míg a
laminin a HCC stromájában mind az erek BM-jében, mind a kötőszövetben előfordul,
addig az agrin expresszió csak az erek BM-jére korlátozódik.
A CC-ben erős agrin immunreakciót kaptunk. Az agrin immunpozitivitás a
glanduláris szerkezetű tumorsejteket körülvevő BM-ra lokalizálódik. A tumor stromája
az erek BM-je kivételével negatív volt. Az agrin fehérjeexpresszió intenzitása a tumor
differenciáltságtól függően eltért. Magasan és közepesen differenciált tumorban, a
tumorfészkek körüli BM-ben jóval erősebb agrin expresszió volt detektálható, mint
rosszul differenciált tumorokban.
A kevésbé differenciált tumorok egyre kevesebb agrint tartalmaznak, ami arra
utal, hogy az agrin csökkenése a tumor progressziójával és nem a malignus
transzformáció tényével függ össze. Itt az agrin eltünése a bazális membrán
degradálódásával, az intraendothelialis kapcsolatok fellazulásával járhat együtt, és az
endothelsejt migrációjának elindítása, az angiogén stimulus irányában hathat.
A megfigyelt változások feltehetõen maguk után vonják a mikrokörnyezet
módosulását, eltérő szabályozó faktorok kötődését és érvényre jutását a daganatos sejtek
környezetében, elősegítve a daganatok növekedési autonómiájának kialakulását.
Mind a real-time RT-PCR, mind a Western blot, mind pedig az
immunhisztokémiai vizsgálatok arra utalnak, hogy a környező, illetve normál májhoz és
a HCC-hez képest a CC-ben az agrin expresszió jelentősen fokozott.
Az RNS- és fehérje-szintű vizsgálatok igen jól egyeztek egymással, ami arra utal, hogy
a vizsgált mintákban az expresszió elsősorban a transzkripció szintjén szabályozódik.
Mások közlése szerint előfordul, hogy viszonylag magas RNS szint mellett a fehérje
nem íródik át, tehát számítani kell a transzláció szintjén ható szabályozó
mechanizmusokra is (Benjamin és Moroni, 2007).
Még nem tisztázott az agrin fokozott expressziójának a forrása. A CC-ben
valószínű, hogy a tumorsejtek részben agrint termelnek, részben pedig olyan faktorokat,
amelyek a környezet nem tumoros sejtjeit fokozott agrin-szintézisre késztetik. A
fokozott agrin expresszió egyrészt különböző növekedési faktorok kötésével, ko-
receptorként közvetlenül segíti a tumor proliferációját, invázióját, ugyanakkor mintegy
körülbástyázza a daganatot, lehetőséget nyújtva annak fokozott autonómiájára.

65
Összeségében eredményeink alapján elmondhatjuk, hogy az agrin expresszió
kimutatható normál májban, cirrhotikus májban, HCA-ban, FNH-ban, HCC-ben,
valamint CC-ben. A fokozott agrin expresszió HCC-ben és HCA-ban e fehérjének a
kapillarizációban és a neovaszkularizációban betöltött szerepére utalhat. Elképzelhető,
hogy CC-ben az agrin a tumor-progresszióban játszik fontos szerepet.

66
8. KÖVETKEZTETÉSEK
1. A normál humán májszövetben az agrin fehérje a portális terek ereinek és
epeútjainak bazális membránjában fordul elő. A szinuszoidok mentén agrin nem
expresszálódik.
2. A cirrhotikus májszövetben az agrin fehérje-expresszió a felszaporodott
kötőszövetben a proliferáló epeutak és az erek bazális membránjában mutatható ki. A
cirrhotikus májban - a normál májhoz hasonlóan - a parenchymában a szinuszoidok
mentén nem volt megfigyelhető agrin pozitivitás.
3. A hepatocellularis carcinomákban (HCC) erős agrin expresszió figyelhető
meg az újonnan képződött kapillárisok és az erek bazális membránjában.
4. Az endothelt jellemző CD34 fehérje expresszió a HCC-ben kolokalizációt
mutatott az agrin expresszióval.
5. A jól és közepesen differenciált cholangiocarcinomákban (CC) erős agrin
expresszió jellemzi a tumorsejtek alkotta glanduláris és trabekuláris struktúrák körüli
bazális membránt, amelyhez nem kapcsolódik CD34 pozitivitás.
6. Ezzel szemben a rosszul differenciált cholangiocarcinomákban, valamint a
környezetet infiltráló tumorszövetben az agrin expresszió csökkent, fragmentálódott, sőt
el is tűnt.
7. Sem a cholangiocarcinoma sejteiben, sem - a lamininnal ellentétben - a tumor
stromájában nem volt megfigyelhető az agrin fehérje-expresszió.
8. Összességében az agrin fehérje-expressziója a HCC-ben fokozottabb, mint a
normál májban, azonban a HCC-nél is magasabb a CC-ben. Az mRNS expresszió

67
vizsgálata hasonló eltérést mutatott és a Western blot analízis megerősítette az
immunhisztokémiai módszerrel detektált fehérje-expressziós eredményeket.
9. Az agrin expressziója a tumorokon belül változatos expressziót mutat a
hepatocellularis adenomákban (HCA). Egyes területeken erős immunopozitivitás
látható a szinuszoidoknak megfelelő képletek mentén, míg máshol a tumorszövet
teljesen negatív.
10. A fokális noduláris hyperpláziában (FNH) közepesen erős agrin expresszió
figyelhető meg a proliferáló epeútak bazális membránjában. A centrálisan szembetűnő,
nagyszámú vaskos falú ér bazális membránja is agrin pozitív, míg a szinuszoidok fala
negatív.

68
9. ÖSSZEFOGLALÁS
A heparánszulfát-proteoglikánok (HSPG) az ECM fehérjék családjába tartozó
molekulák. Részben fehérje-, részben cukorláncaik részvételével aktív részesei a
sejtműködés szabályozásának. A máj károsodása során a proteoglikánok mennyisége
igen jelentősen változik. Mennyiségi és minőségi változásuk hozzájárul a daganatos
fenotípus kialakításához. Az agrin egy multidomain HSPG, amely különböző szervek
bazális membránjában található. Jelenleg kevés irodalmi adat áll rendelkezésünkre az
agrin karcinogenezisben betöltött szerepéről. Munkánk során célul tűztük ki az agrin
mRNS- és fehérje-expressziójának tanulmányozását egészséges májban és különböző
májbetegségekben, elsősorban primer májrákokban.
Az immunohisztokémiai vizsgálatokat 114 formalinban fixált, paraffinba
ágyazott humán mintán (35 HCC, 35 CC, 14 HCA, 20 FNH, 10 normál máj) végeztük.
Az agrin mRNS-expresszóját 52 sebészetileg eltávolított és RNA laterben fixált mintán
vizsgáltuk (13 HCC, 13 HCC tumormentes környező májszövet, 9 CC, 7 CC
tumormentes környező májszövet, 10 normál máj). A fenti vizsgálatok megerősítésére
Western blot analízist végeztünk.
A normál humán májban, illetve tumor melletti ép májszövetben az agrint a
portális tér ereiben, az epeutak és a portális erek bazális membrán zónájában
detektáltuk; a szinuszoidokban nem volt kimutatható agrin fehérje expresszió. A
cirrhotikus májszövetben az agrin-expresszió a májparenchymában és a szinuszoidok
mentén nem volt megfigyelhető, viszont erős pozitivitás volt látható a kötőszövetes
sövényekben, a portális erek és a felszaporodott epekanalikulusok bazális
membránjában. A HCC-ben intenzív agrin fehérje expressziót találtunk az újonnan
képződött kapillárisok és az erek bazális membránjában, amely a tumorok
differenciáltsági fokával nem mutatott összefüggést. Erős agrin-expresszió figyelhető
meg a tumorsejtekből álló glanduláris és trabekuláris szövetszerkezetet körülvevő
bazális membránban a jól és közepesen differenciált CC-kben. Ezzel szemben a rosszul
differenciált CC-ben, valamint a környezetbe betörő tumorszövetben az agrin-
expresszió csökkent, fragmentálódott, sőt el is tűnt. A HCA-ban az agrin expressziója a
tumorokon belül változatos expressziót mutat. Egyes területeken erős immunpozitivitás
látható a szinuszoidoknak megfelelő képletek mentén, míg máshol a tumorszövet

69
teljesen negatív. A FNH-ban intenzív agrin expresszió figyelhető meg a proliferáló
epeutak bazális membránjában.
A fokozott agrin expresszió HCA-ban és HCC-ben e fehérjének a
kapillarizációban és a neovaszkularizációban betöltött szerepére utalhat. Elképzelhető,
hogy CC-ben az agrin a tumor-progresszióban játszik fontos szerepet.

70
10. SUMMARY
Heparan sulphate proteoglycans mediate cell adhesion and control the activities
of numerous growth and motility factors. They play a critical role in carcinogenesis and
tumour progression. Agrin is a large, multidomain heparan sulphate proteoglycan which
is associated with basement membranes of several tissues. Recently, the tissue
expression of agrin was described in several organs, including the liver, under
physiological conditions. So far, little is known about its role in malignancies,
especially in primary liver tumours.
Our goal, therefore, was to investigate mRNA and protein expression of agrin in
primary liver tumors (HCC, CC, HCA and FNH) to analyze its involvement in
tumorigenesis.
The mRNA expression was measured on 52 surgically resected, RNA later fixed
samples by real-time PCR method. Agrin immunostaining was carried out on 114
formalin fixed, paraffin-embedded surgical biopsy specimens (35 CC, 35 HCC, 14
HCA, 20 FNH, 10 normal liver). Western blot analysis on representative samples was
also performed.
Immunohistochemistry showed mild positivity around the bile ducts and the
blood vessels within the portal area in normal liver, however, no expression within the
hepatic lobules was found. HCC presented intense expression along the neovascular
basement membrane. CC samples showed increased mRNA expression of agrin in
comparison to both normal liver and HCC. Intense protein expression of agrin was
observed in the tumor-specific basement membrane in well differentiated areas of CCs,
which staining was fragmented, decreased or even disappeared in less differentiated
fields and sites of infiltration. Western blot analysis confirmed the presence of increased
agrin expression.
In HCA, agrin expression was variable within the tumors. In the some fields
agrin showed strong expression along the sinusoids, while elsewhere it was negative. In
FNH intensive agrin expression was detected in the basement membrane of the
proliferated bile ducts, while no agrin expression was observed along the sinusoids.

71
Agrin may play an important role in neoangiogenesis in human HCCs and
HCAs, as part of the newly formed vasculature. In CCs, agrin is also thought to play an
important role in tumour progression.

72
11. IRODALOMJEGYZÉK
Albores-Saavedra,J, L Murakata, J E Krueger, D E Henson, 2000, Noninvasive and
minimally invasive papillary carcinomas of the extrahepatic bile ducts: Cancer, 89: 508-
515.
Angelucci,A, M Bologna, 2007, Targeting vascular cell migration as a strategy for
blocking angiogenesis: the central role of focal adhesion protein tyrosine kinase family:
Curr.Pharm.Des, 13: 2129-2145.
Arenson,DM, D M Bissell, 1987, Glycosaminoglycan, proteoglycan, and hepatic
fibrosis: Gastroenterology, 92: 536-538.
Aviezer,D, D Hecht, M Safran, M Eisinger, G David, A Yayon, 1994, Perlecan, basal
lamina proteoglycan, promotes basic fibroblast growth factor-receptor binding,
mitogenesis, and angiogenesis: Cell, 79: 1005-1013.
Beasley,RP, L Y Hwang, C C Lin, C S Chien, 1981, Hepatocellular carcinoma and
hepatitis B virus. A prospective study of 22 707 men in Taiwan: Lancet, 2: 1129-1133.
Benjamin,D, C Moroni, 2007, mRNA stability and cancer: an emerging link?:
Expert.Opin.Biol.Ther., 7: 1515-1529
Bezakova,G, M A Ruegg, 2003, New insights into the roles of agrin: Nat.Rev.Mol.Cell
Biol., 4: 295-308.
Biecker,E, H P Fischer, H Strunk, T Sauerbruch, 2003, Benign hepatic tumours:
Z.Gastroenterol., 41: 191-200.
Bosch,FX, J Ribes, M Diaz, R Cleries, 2004, Primary liver cancer: worldwide incidence
and trends: Gastroenterology, 127: S5-S16.

73
Bowe,MA, K A Deyst, J D Leszyk, J R Fallon, 1994, Identification and purification of
an agrin receptor from Torpedo postsynaptic membranes: a heteromeric complex related
to the dystroglycans: Neuron, 12: 1173-1180.
Bowe,MA, J R Fallon, 1995, The role of agrin in synapse formation:
Annu.Rev.Neurosci., 18: 443-462.
Broome,U, R Olsson, L Loof, G Bodemar, R Hultcrantz, A Danielsson, H Prytz, H
Sandberg-Gertzen, S Wallerstedt, G Lindberg, 1996, Natural history and prognostic
factors in 305 Swedish patients with primary sclerosing cholangitis: Gut, 38: 610-615.
Burgess,RW, W C Skarnes, J R Sanes, 2000, Agrin isoforms with distinct amino
termini: differential expression, localization, and function: J.Cell Biol., 151: 41-52.
Castellot,JJ, Jr., M L Addonizio, R Rosenberg, M J Karnovsky, 1981, Cultured
endothelial cells produce a heparinlike inhibitor of smooth muscle cell growth: J.Cell
Biol., 90: 372-379.
Cattaruzza,S, M Schiappacassi, A Ljungberg-Rose, P Spessotto, D Perissinotto, M
Morgelin, M T Mucignat, A Colombatti, R Perris, 2002, Distribution of PG-M/versican
variants in human tissues and de novo expression of isoform V3 upon endothelial cell
activation, migration, and neoangiogenesis in vitro: J.Biol.Chem., 277: 47626-47635.
Cavari,S, S Vannucchi, 1993, Glycosaminoglycans exposed on the endothelial cell
surface. Binding of heparin-like molecules derived from serum: FEBS Lett., 323: 155-
158.
Chakravarti,S, 2002, Functions of lumican and fibromodulin: lessons from knockout
mice: Glycoconj.J., 19: 287-293.

74
Cheifetz,S, J A Weatherbee, M L Tsang, J K Anderson, J E Mole, R Lucas, J Massague,
1987, The transforming growth factor-beta system, a complex pattern of cross-reactive
ligands and receptors: Cell, 48: 409-415.
Cheng,J, J P Grande, 2002, Transforming growth factor-beta signal transduction and
progressive renal disease: Exp.Biol.Med.(Maywood.), 227: 943-956.
Cole,GJ, M Burg, 1989, Characterization of a heparan sulfate proteoglycan that
copurifies with the neural cell adhesion molecule: Exp.Cell Res., 182: 44-60.
Cole,GJ, W Halfter, 1996, Agrin: an extracellular matrix heparan sulfate proteoglycan
involved in cell interactions and synaptogenesis. [Review] [86 refs]: Perspectives on
Developmental Neurobiology.3(4):359-71.
Cotman,SL, W Halfter, G J Cole, 2000, Agrin binds to beta-amyloid (Abeta),
accelerates abeta fibril formation, and is localized to Abeta deposits in Alzheimer's
disease brain: Mol.Cell Neurosci., 15: 183-198.
Denzer,AJ, R Brandenberger, M Gesemann, M Chiquet, M A Ruegg, 1997, Agrin binds
to the nerve-muscle basal lamina via laminin: J.Cell Biol., 137: 671-683.
Denzer,AJ, T Schulthess, C Fauser, B Schumacher, R A Kammerer, J Engel, M A
Ruegg, 1998, Electron microscopic structure of agrin and mapping of its binding site in
laminin-1: EMBO J., 17: 335-343.
Desmet,V, T Roskams, P Van Eyken, 1995, Ductular reaction in the liver:
Pathol.Res.Pract., 191: 513-524.
Dodge,GR, I Kovalszky, M L Chu, J R Hassell, O W McBride, H F Yi, R V Iozzo,
1991, Heparan sulfate proteoglycan of human colon: partial molecular cloning, cellular
expression, and mapping of the gene (HSPG2) to the short arm of human chromosome
1: Genomics, 10: 673-680.

75
Doege,K, M Sasaki, Y Yamada, 1990, Rat and human cartilage proteoglycan (aggrecan)
gene structure: Biochem.Soc.Trans., 18: 200-202.
Donahue,JE, T M Berzin, M S Rafii, D J Glass, G D Yancopoulos, J R Fallon, E G
Stopa, 1999, Agrin in Alzheimer's disease: altered solubility and abnormal distribution
within microvasculature and brain parenchyma: Proc.Natl.Acad.Sci.U.S.A, 96: 6468-
6472.
Durbeej,M, M D Henry, M Ferletta, K P Campbell, P Ekblom, 1998, Distribution of
dystroglycan in normal adult mouse tissues: J.Histochem.Cytochem., 46: 449-457.
Eble,JA, J Haier, 2006, Integrins in cancer treatment: Curr.Cancer Drug Targets., 6: 89-
105.
el Serag,HB, 2001, Epidemiology of hepatocellular carcinoma: Clin.Liver Dis., 5: 87-
107, vi.
Esko,JD, J L Weinke, W H Taylor, G Ekborg, L Roden, G Anantharamaiah, A Gawish,
1987, Inhibition of chondroitin and heparan sulfate biosynthesis in Chinese hamster
ovary cell mutants defective in galactosyltransferase I: J.Biol.Chem., 262: 12189-
12195.
Fedarko,NS, M Ishihara, H E Conrad, 1989, Control of cell division in hepatoma cells
by exogenous heparan sulfate proteoglycan: J.Cell Physiol, 139: 287-294.
Ferns,MJ, J T Campanelli, W Hoch, R H Scheller, Z Hall, 1993, The ability of agrin to
cluster AChRs depends on alternative splicing and on cell surface proteoglycans:
Neuron, 11: 491-502.
Fisher,LW, J D Termine, M F Young, 1989, Deduced protein sequence of bone small
proteoglycan I (biglycan) shows homology with proteoglycan II (decorin) and several
nonconnective tissue proteins in a variety of species: J.Biol.Chem., 264: 4571-4576.

76
Frail,DE, L L McLaughlin, J Mudd, J P Merlie, 1988, Identification of the mouse
muscle 43,000-dalton acetylcholine receptor-associated protein (RAPsyn) by cDNA
cloning: J.Biol.Chem., 263: 15602-15607.
Gallagher,JT, 1989, The extended family of proteoglycans: social residents of the
pericellular zone: Curr.Opin.Cell Biol., 1: 1201-1218.
Gallagher,JT, M Lyon, W P Steward, 1986, Structure and function of heparan sulphate
proteoglycans: Biochem.J., 236: 313-325.
Gallai,M, I Kovalszky, T Knittel, K Neubauer, T Armbrust, G Ramadori, 1996,
Expression of extracellular matrix proteoglycans perlecan and decorin in carbon-
tetrachloride-injured rat liver and in isolated liver cells: Am.J.Pathol., 148: 1463-1471.
Gallegos,NC, C Smales, F J Savage, R M Hembry, P B Boulos, 1995, The distribution
of matrix metalloproteinases and tissue inhibitor of metalloproteinases in colorectal
cancer: Surg.Oncol., 4: 21-29.
Gautam,M, P G Noakes, L Moscoso, F Rupp, R H Scheller, J P Merlie, J R Sanes,
1996, Defective neuromuscular synaptogenesis in agrin-deficient mutant mice: Cell, 85:
525-535.
Gee,SH, F Montanaro, M H Lindenbaum, S Carbonetto, 1994, Dystroglycan-alpha, a
dystrophin-associated glycoprotein, is a functional agrin receptor: Cell, 77: 675-686.
Gesemann,M, A Brancaccio, B Schumacher, M A Ruegg, 1998, Agrin is a high-affinity
binding protein of dystroglycan in non-muscle tissue: J.Biol.Chem., 273: 600-605.
Gesemann,M, A J Denzer, M A Ruegg, 1995b, Acetylcholine receptor-aggregating
activity of agrin isoforms and mapping of the active site: J.Cell Biol., 128: 625-636.

77
Gingras,J, M Ferns, 2001, Expression and localization of agrin during sympathetic
synapse formation in vitro: J.Neurobiol., 48: 228-242.
Grazioli,L, M P Federle, G Brancatelli, T Ichikawa, L Olivetti, A Blachar, 2001,
Hepatic adenomas: imaging and pathologic findings: Radiographics, 21: 877-892.
Groffen,AJ, C A Buskens, T H van Kuppevelt, J H Veerkamp, L A Monnens, L P van
den Heuvel, 1998a, Primary structure and high expression of human agrin in basement
membranes of adult lung and kidney: Eur.J.Biochem., 254: 123-128.
Groffen,AJ, M A Ruegg, H Dijkman, T J van de Velden, C A Buskens, B J van den, K J
Assmann, L A Monnens, J H Veerkamp, L P van den Heuvel, 1998b, Agrin is a major
heparan sulfate proteoglycan in the human glomerular basement membrane: Journal of
Histochemistry & Cytochemistry.46(1):19-27.
Halfter,W, S Dong, B Schurer, G J Cole, 1998, Collagen XVIII is a basement
membrane heparan sulfate proteoglycan: J.Biol.Chem., 273: 25404-25412.
Henry,SH, F X Bosch, J C Bowers, 2002, Aflatoxin, hepatitis and worldwide liver
cancer risks: Adv.Exp.Med.Biol., 504: 229-233.
Heremans,A, J J Cassiman, B H Van den, G David, 1988, Heparan sulfate proteoglycan
from the extracellular matrix of human lung fibroblasts. Isolation, purification, and core
protein characterization: J.Biol.Chem., 263: 4731-4739.
Hildebrand,A, M Romaris, L M Rasmussen, D Heinegard, D R Twardzik, W A Border,
E Ruoslahti, 1994, Interaction of the small interstitial proteoglycans biglycan, decorin
and fibromodulin with transforming growth factor beta: Biochem.J., 302 ( Pt 2): 527-
534.

78
Hussain,SM, T Terkivatan, P E Zondervan, E Lanjouw, S de Rave, J N Ijzermans, R A
de Man, 2004, Focal nodular hyperplasia: findings at state-of-the-art MR imaging, US,
CT, and pathologic analysis: Radiographics, 24: 3-17.
Iozzo,RV, 2005, Basement membrane proteoglycans: from cellar to ceiling:
Nat.Rev.Mol.Cell Biol., 6: 646-656.
Iozzo,RV, I R Cohen, S Grassel, A D Murdoch, 1994, The biology of perlecan: the
multifaceted heparan sulphate proteoglycan of basement membranes and pericellular
matrices: Biochem.J., 302 ( Pt 3): 625-639.
Iozzo,RV, I Kovalszky, N Hacobian, P K Schick, J S Ellingson, G R Dodge, 1990, Fatty
acylation of heparan sulfate proteoglycan from human colon carcinoma cells:
J.Biol.Chem., 265: 19980-19989.
Ip,FC, D G Glass, D R Gies, J Cheung, K O Lai, A K Fu, G D Yancopoulos, N Y Ip,
2000, Cloning and characterization of muscle-specific kinase in chicken: Mol.Cell
Neurosci., 16: 661-673.
Jennings,CG, S J Burden, 1993, Development of the neuromuscular synapse:
Curr.Opin.Neurobiol., 3: 75-81.
Jiang,X, J R Couchman, 2003, Perlecan and tumor angiogenesis:
J.Histochem.Cytochem., 51: 1393-1410.
Kainulainen,V, L Nelimarkka, H Jarvelainen, M Laato, M Jalkanen, K Elenius, 1996,
Suppression of syndecan-1 expression in endothelial cells by tumor necrosis factor-
alpha: J.Biol.Chem., 271: 18759-18766.
Kammerer,RA, T Schulthess, R Landwehr, B Schumacher, A Lustig, P D Yurchenco,
M A Ruegg, J Engel, A J Denzer, 1999, Interaction of agrin with laminin requires a

79
coiled-coil conformation of the agrin-binding site within the laminin gamma1 chain:
EMBO J., 18: 6762-6770.
Kato,M, H Wang, V Kainulainen, M L Fitzgerald, S Ledbetter, D M Ornitz, M
Bernfield, 1998, Physiological degradation converts the soluble syndecan-1 ectodomain
from an inhibitor to a potent activator of FGF-2: Nat.Med., 4: 691-697.
Kemeny,E, H M Fillit, S Damle, R Mahabir, N A Kefalides, J D Gregory, T
Antonovych, S Sabnis, J B Zabriskie, 1988, Monoclonal antibodies to heparan sulfate
proteoglycan: development and application to the study of normal tissue and pathologic
human kidney biopsies: Connect.Tissue Res, 18: 9-25.
Kim,MJ, I H Liu, Y Song, J A Lee, W Halfter, R J Balice-Gordon, E Linney, G J Cole,
2007, Agrin is required for posterior development and motor axon outgrowth and
branching in embryonic zebrafish: Glycobiology, 17: 231-247.
Kimata,K, Y Oike, K Tani, T Shinomura, M Yamagata, M Uritani, S Suzuki, 1986, A
large chondroitin sulfate proteoglycan (PG-M) synthesized before chondrogenesis in the
limb bud of chick embryo: J.Biol.Chem., 261: 13517-13525.
Kimura,S, J Cheng, H Ida, N Hao, Y Fujimori, T Saku, 2000, Perlecan (heparan sulfate
proteoglycan) gene expression reflected in the characteristic histological architecture of
salivary adenoid cystic carcinoma: Virchows Arch., 437: 122-128.
Kiss,A, G Lotz, N P Kaposi, Z Schaff, 2002, [Hepatitis viruses and
hepatocarcinogenesis]: Orv.Hetil., 143: 83-86.
Kiss,A, A Szepesi, G Lotz, P Nagy, Z Schaff, 1998, Expression of transforming growth
factor-alpha in hepatoblastoma: Cancer, 83: 690-697.
Knox,SM, J M Whitelock, 2006, Perlecan: how does one molecule do so many things?:
Cell Mol.Life Sci., 63: 2435-2445.

80
Konomi,H, J M Seyer, Y Ninomiya, B R Olsen, 1986, Peptide-specific antibodies
identify the alpha 2 chain as the proteoglycan subunit of type IX collagen: J.Biol.Chem.,
261: 6742-6746.
Kopper L, Schaff Zs. Patológia tankönyv 2. Medicina, Budapest, 2004: 872-874
Kovalszky,I, A Jeney, K Lapis, 1993, [Proteoglycans (their structure, function and role
in liver diseases)]: Orv.Hetil., 134: 2019-2026.
Kovalszky,I, I, J O Nagy, M Gallai, A Sebestyen, Z Schaff, S Paku, A Jeney, R V
Iozzo, 1997, Altered Proteoglycan Gene Expression in Human Biliary Cirrhosis:
Pathol.Oncol.Res., 3: 51-58.
Kovalszky,I, P Nagy, B Szende, K Lapis, F Szalay, A Jeney, Z Schaff, 1998,
Experimental and human liver fibrogenesis: Scand.J.Gastroenterol.Suppl, 228: 51-55.
Kovalszky,I, G Pogany, G Molnar, A Jeney, K Lapis, S Karacsonyi, A Szecseny, R V
Iozzo, 1990, Altered glycosaminoglycan composition in reactive and neoplastic human
liver: Biochem.Biophys.Res.Commun., 167: 883-890.
Krusius,T, E Ruoslahti, 1986, Primary structure of an extracellular matrix proteoglycan
core protein deduced from cloned cDNA: Proc.Natl.Acad.Sci.U.S.A, 83: 7683-7687.
Lee,JS, S S Thorgeirsson, 2002, Functional and genomic implications of global gene
expression profiles in cell lines from human hepatocellular cancer: Hepatology, 35:
1134-1143.
Lin,W, R W Burgess, B Dominguez, S L Pfaff, J R Sanes, K F Lee, 2001, Distinct roles
of nerve and muscle in postsynaptic differentiation of the neuromuscular synapse:
Nature, 410: 1057-1064.
Liotta,LA, 1982, Tumor extracellular matrix: Lab Invest, 47: 112-113.

81
Lyon,M, J A Deakin, K Mizuno, T Nakamura, J T Gallagher, 1994, Interaction of
hepatocyte growth factor with heparan sulfate. Elucidation of the major heparan sulfate
structural determinants: J.Biol.Chem., 269: 11216-11223.
Lyon,M, J T Gallagher, 1991, Purification and partial characterization of the major cell-
associated heparan sulphate proteoglycan of rat liver: Biochem.J., 273(Pt 2): 415-422.
Madri,JA, M Marx, 1992, Matrix composition, organization and soluble factors:
modulators of microvascular cell differentiation in vitro: Kidney Int., 41: 560-565.
Magill-Solc,C, U J McMahan, 1988, Motor neurons contain agrin-like molecules: J.Cell
Biol., 107: 1825-1833.
Marchetti,D, D Menter, L Jin, M Nakajima, G L Nicolson, 1993, Nerve growth factor
effects on human and mouse melanoma cell invasion and heparanase production:
Int.J.Cancer, 55: 692-699.
Marneros,AG, B R Olsen, 2005, Physiological role of collagen XVIII and endostatin:
FASEB J., 19: 716-728.
Martin,PT, J R Sanes, 1997, Integrins mediate adhesion to agrin and modulate agrin
signaling: Development, 124: 3909-3917.
McMahan,UJ, 1990, The agrin hypothesis: Cold Spring Harb.Symp.Quant.Biol., 55:
407-418.
Merz,H, R Malisius, S Mannweiler, R Zhou, W Hartmann, K Orscheschek, P
Moubayed, A C Feller, 1995, ImmunoMax. A maximized immunohistochemical
method for the retrieval and enhancement of hidden antigens: Lab Invest, 73: 149-156.
Metayer,S, F Dacheux, J L Dacheux, J L Gatti, 2002, Comparison, characterization, and
identification of proteases and protease inhibitors in epididymal fluids of domestic

82
mammals. Matrix metalloproteinases are major fluid gelatinases: Biol.Reprod., 66:
1219-1229.
Mongiat,M, J Otto, R Oldershaw, F Ferrer, J D Sato, R V Iozzo, 2001, Fibroblast
growth factor-binding protein is a novel partner for perlecan protein core: J.Biol.Chem.,
276: 10263-10271.
Mongiat,M, S M Sweeney, J D San Antonio, J Fu, R V Iozzo, 2003, Endorepellin, a
novel inhibitor of angiogenesis derived from the C terminus of perlecan: J.Biol.Chem.,
278: 4238-4249.
Moon,JJ, M Matsumoto, S Patel, L Lee, J L Guan, S Li, 2005, Role of cell surface
heparan sulfate proteoglycans in endothelial cell migration and mechanotransduction:
J.Cell Physiol, 203: 166-176.
Morita,H, A Yoshimura, K Inui, T Ideura, H Watanabe, L Wang, R Soininen, K
Tryggvason, 2005, Heparan sulfate of perlecan is involved in glomerular filtration:
J.Am.Soc.Nephrol., 16: 1703-1710.
Mueller,MM, N E Fusenig, 2002, Tumor-stroma interactions directing phenotype and
progression of epithelial skin tumor cells: Differentiation, 70: 486-497.
Murata,K, Y Ochiai, K Akashio, 1985, Polydispersity of acidic glycosaminoglycan
components in human liver and the changes at different stages in liver cirrhosis:
Gastroenterology, 89: 1248-1257.
Nagy,P, V Laszlo, Z Schaff, 2006, [Hepatocarcinogenesis in human liver]:
Magy.Onkol., 50: 107-113.
Nakeeb,A, H A Pitt, T A Sohn, J Coleman, R A Abrams, S Piantadosi, R H Hruban, K
D Lillemoe, C J Yeo, J L Cameron, 1996, Cholangiocarcinoma - A spectrum of
intrahepatic, perihilar, and distal tumors: Annals of Surgery, 224: 463-473.

83
Naor,D, R V Sionov, D Ish-Shalom, 1997, CD44: structure, function, and association
with the malignant process: Adv.Cancer Res., 71: 241-319.
Nishiyama,A, K J Dahlin, W B Stallcup, 1991, The expression of NG2 proteoglycan in
the developing rat limb: Development, 111: 933-944.
Oldberg,A, L Kjellen, M Hook, 1979, Cell-surface heparan sulfate. Isolation and
characterization of a proteoglycan from rat liver membranes: J.Biol.Chem., 254: 8505-
8510.
Otto,S, M Kasler, 2005, [Trends in cancer mortality and morbidity in Hungarian and
international statistics. Characteristics and potential outcome of public health screening
programs]: Magy.Onkol., 49: 99-7.
Oyunsuren,T, R Sanduijav, D Davaadorj, D Nansalmaa, 2006, Hepatocellular
carcinoma and its early detection by AFP testing in Mongolia: Asian Pac.J.Cancer
Prev., 7: 460-462.
Qiao,D, K Meyer, C Mundhenke, S A Drew, A Friedl, 2003, Heparan sulfate
proteoglycans as regulators of fibroblast growth factor-2 signaling in brain endothelial
cells. Specific role for glypican-1 in glioma angiogenesis: J.Biol.Chem., 278: 16045-
16053.
Rabenstein,DL, 2002, Heparin and heparan sulfate: structure and function:
Nat.Prod.Rep., 19: 312-331.
Rascher,G, A Fischmann, S Kroger, F Duffner, E H Grote, H Wolburg, 2002,
Extracellular matrix and the blood-brain barrier in glioblastoma multiforme: spatial
segregation of tenascin and agrin: Acta Neuropathol.(Berl), 104: 85-91.
Reed,CC, R V Iozzo, 2002, The role of decorin in collagen fibrillogenesis and skin
homeostasis: Glycoconj.J., 19: 249-255.

84
Reist,NE, C Magill, U J McMahan, 1987, Agrin-like molecules at synaptic sites in
normal, denervated, and damaged skeletal muscles: J.Cell Biol., 105: 2457-2469.
Ronnov-Jessen,L, O W Petersen, 1993, Induction of alpha-smooth muscle actin by
transforming growth factor-beta 1 in quiescent human breast gland fibroblasts.
Implications for myofibroblast generation in breast neoplasia: Lab Invest, 68: 696-707.
Ruegg,MA, J L Bixby, 1998, Agrin orchestrates synaptic differentiation at the
vertebrate neuromuscular junction: Trends Neurosci., 21: 22-27.
Ruegg,MA, K W Tsim, S E Horton, S Kroger, G Escher, E M Gensch, U J McMahan,
1992, The agrin gene codes for a family of basal lamina proteins that differ in function
and distribution: Neuron, 8: 691-699.
Ruoslahti,E, Y Yamaguchi. Proteoglycans as modulator of growth factor activities. 56,
867-869. 1991.
Rupp,F, T Ozcelik, M Linial, K Peterson, U Francke, R Scheller, 1992, Structure and
chromosomal localization of the mammalian agrin gene: J.Neurosci., 12: 3535-3544.
Rupp,F, D G Payan, C Magill-Solc, D M Cowan, R H Scheller, 1991, Structure and
expression of a rat agrin: Neuron, 6: 811-823.
Sabit,H, K Tsuneyama, T Shimonishi, K Harada, J Cheng, H Ida, T Saku, K Saito, Y
Nakanuma, 2001, Enhanced expression of basement-membrane-type heparan sulfate
proteoglycan in tumor fibro-myxoid stroma of intrahepatic cholangiocarcinoma:
Pathol.Int., 51: 248-256.
Sadler,JE, 1997, Thrombomodulin structure and function: Thromb.Haemost., 78: 392-
395.

85
Sahani,D, S R Prasad, K K Tannabe, P F Hahn, P R Mueller, S Saini, 2003, Thorotrast-
induced cholangiocarcinoma: case report: Abdom.Imaging, 28: 72-74.
Saksela,O, D Moscatelli, A Sommer, D B Rifkin, 1988, Endothelial cell-derived
heparan sulfate binds basic fibroblast growth factor and protects it from proteolytic
degradation: J.Cell Biol., 107: 743-751.
Saunders,S, M Jalkanen, S O'Farrell, M Bernfield, 1989, Molecular cloning of
syndecan, an integral membrane proteoglycan: J.Cell Biol., 108: 1547-1556.
Schaff Z., Nagy P. Pathology techniques and grading systems in the diagnosis of HCC.
111-133. 1997. Churchill Livingstone New York, Liver Cancer.
Schaff,Z, I Sarosi, C C Hsia, A Kiss, E Tabor, 1995, p53 in malignant and benign liver
lesions: Eur.J.Cancer, 31A: 1847-1850.
Schaff,Z, E Szekely, B Jaray, A Kiss, 2004, [Pathology of liver tumors]: Orv.Hetil.,
145: 368-374.
Sebestyen,A, M Gallai, T Knittel, T Ambrust, G Ramadori, I Kovalszky, 2000,
Cytokine regulation of syndecan expression in cells of liver origin: Cytokine, 12: 1557-
1560.
Senoo,H, 2004, Structure and function of hepatic stellate cells: Med.Electron Microsc.,
37: 3-15.
Serpinskaya,AS, G Feng, J R Sanes, A M Craig, 1999, Synapse formation by
hippocampal neurons from agrin-deficient mice: Dev.Biol., 205: 65-78.
Shaib,Y, H B el Serag, 2004, The epidemiology of cholangiocarcinoma: Semin.Liver
Dis., 24: 115-125.

86
Sharma,B, M Handler, I Eichstetter, J M Whitelock, M A Nugent, R V Iozzo, 1998,
Antisense targeting of perlecan blocks tumor growth and angiogenesis in vivo:
J.Clin.Invest, 102: 1599-1608.
Silbert,JE, G Sugumaran, 2002, Biosynthesis of chondroitin/dermatan sulfate:
IUBMB.Life, 54: 177-186.
Smela,ME, S S Currier, E A Bailey, J M Essigmann, 2001, The chemistry and biology
of aflatoxin B(1): from mutational spectrometry to carcinogenesis: Carcinogenesis, 22:
535-545.
Su,CH, Y M Shyr, W Y Lui, F K P'Eng, 1997, Hepatolithiasis associated with
cholangiocarcinoma: Br.J.Surg., 84: 969-973.
Sugahara,K, H Kitagawa, 2002, Heparin and heparan sulfate biosynthesis: IUBMB.Life,
54: 163-175.
Tatrai,P, J Dudas, E Batmunkh, M Mathe, A Zalatnai, Z Schaff, G Ramadori, I
Kovalszky, 2006, Agrin, a novel basement membrane component in human and rat
liver, accumulates in cirrhosis and hepatocellular carcinoma: Lab Invest, 86: 1149-1160.
Taylor,KR, R L Gallo, 2006, Glycosaminoglycans and their proteoglycans: host-
associated molecular patterns for initiation and modulation of inflammation: FASEB J.,
20: 9-22.
Timar,J, C Diczhazi, A Ladanyi, E Raso, W Hornebeck, L Robert, K Lapis, 1995a,
Interaction of tumour cells with elastin and the metastatic phenotype: Ciba
Found.Symp., 192: 321-335.
Timar,J, A Jeney, I Kovalszky, I, L Kopper, 1995b, Role of Proteoglycans in Tumor
Progression: Pathol.Oncol.Res., 1: 85-93.

87
Timar,J, S Paku, J Tovari, B Dome, 2006, [Rationale of antiangiogenic therapy]:
Magy.Onkol., 50: 141-151.
Tompkins,RK, B M Aizen, K D Saunders, J J Roslyn, W P Longmire, Jr., 1991,
Carcinoma of the ductus choledochus: Am.J.Surg., 161: 235-238.
Trotter,JF, G T Everson, 2001, Benign focal lesions of the liver: Clin.Liver Dis., 5: 17-
42, v.
Tsen,G, W Halfter, S Kroger, G J Cole, 1995, Agrin is a heparan sulfate proteoglycan:
J.Biol.Chem., 270: 3392-3399.
Tsim,KW, M A Ruegg, G Escher, S Kroger, U J McMahan, 1992, cDNA that encodes
active agrin: Neuron, 8: 677-689.
Vauthey,JN, L H Blumgart, 1994, Recent advances in the management of
cholangiocarcinomas: Semin.Liver Dis., 14: 109-114.
Watanapa,P, W B Watanapa, 2002, Liver fluke-associated cholangiocarcinoma:
Br.J.Surg., 89: 962-970.
Woods,A, 2001, Syndecans: transmembrane modulators of adhesion and matrix
assembly: J.Clin.Invest, 107: 935-941.
Yamamoto,C, S Shimada, Y Fujiwara, J B Lee, T Hayashi, T Kaji, 2005, Proteoglycans
released from cultured bovine aortic endothelial cell layers by sodium spirulan are both
perlecan and biglycan: Biol.Pharm.Bull., 28: 32-36.
Yang,JF, G Cao, S Koirala, L V Reddy, C P Ko, 2001, Schwann cells express active
agrin and enhance aggregation of acetylcholine receptors on muscle fibers: J.Neurosci.,
21: 9572-9584.

88
12. SAJÁT KÖZLEMÉNYEK JEGYZÉKE Az értekezés alapjául szolgáló közlemények jegyzéke: Batmunkh E., Tátrai P., Szabó E., Lódi C., Holczbauer A., Páska C., Kupcsulik P.,
Kiss A., Schaff Z., Kovalszky I. (2007) Comparison of the expression of agrin, a
basement membrane heparan sulfate proteoglycan, in cholangiocarcinoma and
hepatocellular carcinoma. Human Pathol, 38: 1508-1515
(IF: 2,810)
Tátrai P., Dudás J., Batmunkh E., Máthé M., Zalatnai A., Schaff Z., Ramadori G.,
Kovalszky L. (2006) Agrin, a novel basement membrane component in human and rat
liver, accumulates in cirrhosis and hepatocellular carcinoma. Lab Invest 86: 1149-1160,
(IF: 3.753)
Nem az értekezés alapjául szolgáló közlemények:
Szijártó A.*, Batmunkh E. *, Hahn O., Mihály Z., Kreiss A., Kiss A., Lotz G., Schaff
Z., Váli L., Blázovics A., Geró D., Szabó C., Kupcsulik P. (2007) Effect of PJ-34 Parp-
inhibitor on rat liver microcirculation and antioxidant status. J Surg Res. Sep;142(1):72-
80 ( * megosztott első szerzők)
(IF: 2.038)
Lódi C., Szabó E., Holczbauer Á., Batmunkh E., Szíjártó A., Kupcsulik P., Kovalszky
I., Paku S., Illyés G., Kiss A., Schaff Z. (2006) Claudin-4 differentiates biliary tract
cancers from hepatocellular carcinomas: Mod. Pathol 19: 460-469
(IF: 3,753)
Szabó E., Lódi C., Korpos E., Batmunkh E., Rottenberger Z., Deák F., Kiss I, Tőkés
AM., Lotz G., László V., Kiss A., Schaff Z., Nagy P. (2007) Expression of Matrilin-2 in
oval cells during rat liver regeneration. Matrix Biol, 26: 554-560
(IF: 3,679)

89
E. Szabó, É. Korpos, E. Batmunkh, G. Lotz, Á. Holczbauer, I. Kovalszky, F. Deák, I.
Kiss, Zs. Schaff, A. Kiss. Expression of matrilin-2 in liver cirrhosis and hepatocellular
carcinoma. Pathol Oncol Res. Közlésre elfogadva (2007, augusztus)
(IF: 1,241)
Szijártó A., Hahn O., Batmunkh E., Stangl R., Kiss A., Lotz G., Schaff Z., Váli L.,
Blázovics A., Gerő D.,. Szabó C, Kupcsulik P., Harsányi L. (2007) Short-term alanyl-
glutamine dipeptide pretreatment in liver ischemia-reperfusion model: effects on
microcirculation and antioxidant status in rats. Clinical Nutrition 26, 640-648.
(IF: 2,474)
Megjelent, illetve elfogadott in extensio közlemények impakt faktora összesen:
20.448
Az értekezéssel összefüggő, lektorált folyóiratban megjelent idézhető abstactok:
Batmunkh E, Lódi C, Holczbauer Á, Szabó E, Tátrai P, Páska C, Kiss A, Kupcsulik P,
Kovalszky I, Schaff Z. High expression of agrin in hepatocellular and
cholangiocellular carcinoma. 40th Annual Meeting of the European Association for the
Study of the Liver April 13-17, 2005, Paris, France. Abstract: Journal of Hepatology
Supplement No.2 Volume 42, 2005
Batmunkh E, Lódi C, Holczbauer Á, Szabó E, Tátrai P, Páska C, Kiss A, Kovalszky I,
Schaff Z. High expression of agrin in hepatocellular and cholangiocellular carcinoma.
47th Annual Meeting of the Hungarian Society of Gastroenterology June 07-11, 2005
Balatonaliga, Hungary. Abstract: Zeitschrift für Gastroenterologie; Issue 5, Volume 43,
2005
Egyéb kutatási témákból lektorált folyóiratban megjelent idézhető abstactok:
Szabó E., Páska C., Deák F., Kiss I., Batmunkh E., Lódi C., Holczbauer Á., Kiss A.,
Kovalszky I., Schaff Z. Matrilin-2, a novel extracellular matrix component in cirrhosis

90
and hepatocellular carcinoma. 40th Annual Meeting of the European Association for the
Study of the Liver April 13-17, 2005, Paris, France. J. of Hepatol 42. S2. 2005
Szabó E, Páska C, Lódi C, Batmunkh E, Holczbauer Á, Korpos É, Deák F, Kiss I, Kiss
A, Schaff Z. Expression of matrilin-2 in liver cirrhosis and hepatocellular carcinoma.
47th Annual Meeting of the Hungarian Society of Gastroenterology June 07-11, 2005
Balatonaliga, Hungary. Abstract: Zeitschrift für Gastroenterologie; Issue 5, Volume 43,
2005
Lódi Cs., Szabó E., Batmunkh E., Holczbauer Á., Paku S., Kiss A., Kupcsulik P.,
Illyés Gy, Schaff Zs. High expression of claudin-4 in biliary carcinomas. 20th European
Congress of Pathology, Paris, France, September 3-8, 2005 Virchows Arch 447: 404,
2005.
Lódi C, Szabó E, Holczbauer Á, Batmunkh E, Kovalszky I, Paku S, Kiss A, Illyés G,
Schaff Z. High expression of claudin-4 in human biliary cancers. 47th Annual Meeting
of the Hungarian Society of Gastroenterology June 07-11, 2005 Balatonaliga, Hungary.
Abstract: Zeitschrift für Gastroenterologie; Issue 5, Volume 43, 2005
Egyéb kutatási témákból készült nem idézhető abstractok:
Batmunkh E, Tátrai P, Lódi C, Holczbauer Á, Szabó E, Páska C, Kiss A, Kupcsulik P,
Kovalszky I, Schaff Z. High expression of agrin in cholangiocarcinoma. Falk
Symposium October 10-11, 2006, Freiburg, Germany
Batmunkh E, Lódi C, Holczbauer Á, Szabó E, Tátrai P, Páska C, Kiss A, Kupcsulik P,
Kovalszky I, Schaff Z. High expression of agrin in hepatocellular and
cholangiocellular carcinoma. Falk Symposium, October 3-4, 2005, Berlin, Germany
Lódi C, Szabó E, Holczbauer Á, Batmunkh E, Kiss A, Illyés G, Paku S, Schaff Z,
Nagy P. High expression of claudin-7 during liver regeneration. Falk Symposium,
October 3-4, 2005, Berlin, Germany

91
Tátrai P., Batmunkh E., Dudás J., Egedi K., Ramadori G., Schaff Z., Kovalszky L,
Agrin in the diseased liver: a multifunctional proteoglycan recently found in the liver
may promote bile duct proliferation and tumour angiogenesis FEBS 2005 Congress
Szabó E, Páska C, Lódi C, Batmunkh E, Holczbauer Á, Korpos E, Deák F, Kiss I, Kiss
A, Schaff Z. Expression of matrilin-2 in liver cirrhosis and hepatocellular carcinoma.
Falk Symposium, October 3-4, 2005, Berlin, Germany
Lódi C, Szíjártó A, Holczbauer Á, Szabó E, Batmunkh E, Kiss A, Illyés G, Schaff Z,
Kupcsulik P; High expression of claudin-4 in human cholangiocarcinomas. 6th
Congress of the European Hepato-Pancreato-Biliary Association May 25-28, 2005,
Heidelberg, Germany
Szabó E, Páska C, Deák F, Kiss I, Batmunkh E, Lódi C, Holczbauer Á, Kiss A,
Kovalszky I, Schaff Z. Matrilin-2, a novel extracellular matrix component in cirrhosis
and hepatocellular carcinoma. 40th Annual Meeting of the European Association for the
Study of the Liver April 13-17, 2005, Paris, France. Abstract: Journal of Hepatology;
Supplement No.2 Volume 42, 2005
Holczbauer Á, Halász J, Lódi C, Szabó E, Batmunkh E, Benyó G, Galántai I, Kiss A,
Schaff Zs. Expression of claudins in human hepatoblastoma. 7th Ph.D. Conference at
Semmelweis University, April 14-15, 2005, Budapest, Hungary
Lódi C, Kiss A, Holczbauer Á, Batmunkh E, Paku S, Illyés G, Kupcsulik P, Schaff Z.
High expression of claudin 4 in human cholangiocellular carcinomas. Falk Symposium
October 16-17, 2004, Freiburg, Germany
Holczbauer Á, Lódi Cs, Batmunkh E, Kiss A, Kupcsulik P, Schaff Zs. Decreased
expression of prepro-nociceptin/OFQ and nociceptin/OFQ receptor in human
hepatocellular carcinoma. 63rd Hungarian Congress of Pathology September 23-25,
2004, Siófok-Balatonszéplak, Hungary

92
Lódi C, Szabó E, Batmunkh E, Holczbauer Á, Kiss A, Illyés G, Paku S, Schaff Z,
Nagy P. High expression of claudin-7 during liver regeneration. 41th Annual Meeting
of the European Association for the Study of the Liver April 26-27, 2006, Vienna,
Austria
Lódi C, Szabó E, Batmunkh E, Holczbauer Á, Paku S, Kupcsulik P, Illyés G, Kiss A,
Schaff Z. Different claudin-4 expression on biliary and hepatocellular carcinomas. 41th
Annual Meeting of the European Association for the Study of the Liver April 26-27,
2006, Vienna, Austria

93
13. KÖSZÖNETNYILVÁNÍTÁS Köszönetemet fejezem ki mindazoknak, akik munkámhoz útmutatásaikkal és
segítségükkel hozzájárultak.
Szeretném megköszönni témavezetőmnek, Dr. Kiss Andrásnak, aki mindig nagy
türelemmel bíztatott a tudományos munkában való elmélyülésre, és aki évek óta
figyelemmel irányítja kísérletes munkámat.
Külön köszönettel tartozom Schaff Zsuzsa professzor asszonynak, aki lehetővé
tette, hogy munkámat az II. sz. Patológiai Intézetben végezhessem és mindig támogatott
a munkámban.
Hálás vagyok Dr. Kovalszky Ilona egyetemi tanár segítségéért és tanácsaiért,
mivel jelen munkám nem jöhetett volna létre korábbi ilyen tárgyú kutatásai nélkül.
Külön köszönet illeti Tátrai Pétert, akinek munkája és segítsége nélkül ez a
munka nem jött volna létre.
Külön köszönöm Szabó Erzsébetnek, PhD társamnak, aki mindig segítségemre
volt. Köszönöm a munkatársaimnak, Dr. Holczbauer Ágnesnek, Dr. Lódi Csabának,
Németh Zsuzsának, Dr. Szász Attila Marcellnak, Dr. Tőkés Anna-Máriának és Dr Páska
Csillának munkámhoz nyújtott segítségüket.
Köszönetet szeretnék mondani Szik Ágnesnek és Pekár Zoltánnénak ez idő alatt
végzett pontos, lelkes és lelkiismeretes munkájukért.
Továbbá köszönettel tartozom az II. sz. Patológiai Intézet minden tagjának, akik
munkám elvégzéséhez segítő, baráti légkört teremtettek.
Köszönetet mondok a Mongol és Magyar Oktatási Minisztériumnak, a Mongol
Nagykövetségnek, akik ösztöndíjat bocsátottak rendelkezésemre.
Végül, de nem utolsó sorban hálás vagyok szüleimnek és páromnak, akik
lehetővé tették számomra azt, hogy eljuthassak idáig, mindig mindenben mellettem
álltak és biztosították a hátteret a terveim megvalósításához.





























