Embed Size (px)
Citation preview

SEMS Year 3 project Literature Review Text 1 1.1 Stem cells and cell potency
Stem cells are cells that have potential to commit to a more specialized cell through
what is termed differentiation. Cells become more committed, specialized, as they
undergo differentiation, which is characterized by changes to a cells phenotype due
to the alteration of the expression or repression of certain genes (C. S. Potten, 1990).
Most lineage commited cells will proliferate a finite number of times before they
reach senescence, stem cells have the capacity for prolonged proliferation (Lanza, 2005), and can either divide reproducing a copy of itself, or to a differentiated daughter
cell (Catherine M. Verfaillie, 2002). In mature organisms reserves of stem cells are
maintained within tissues meaning that when needed they can differentiate to replace
damaged cells, or those being replaced in normal tissue turnover.
In mammals there are a few main types of stem cells: Embryonic stem cells
(ESCs), fetal stem cells and adult stem cells. Cells of these different sources vary in
the diversity of cells that they can give rise to, and the measure of this
differentiation potential is termed ‘cell potency’. Adult stem cells found in mature
tissues are often limited to one potential lineage, effectively meaning that their ‘stem
cell’ state is a means of maintaining the homeostasis of cells in a tissue (Turksen,
2004). Stem cells of limited potential, such as unipotent cells that can only
specialize to one cell type, are often termed progenitor cells. Cells such as these
are important in maintaining tissues such as muscle where cells are regularly
damaged through mechanical stimulation (Turksen, 2004).
Cell potency varies from unipotent progenitor cells to totipotnent cells such as the
zygote in humans that can produce all cell types of the mature organism as well as
the extraembryonic tissues. By some definitions stem cells must be capable of
deriving multiple cell types, meaning progenitor cells are sometimes seen as
descendents of stem cells; further along the differentiation pathway but not yet
committed to a mature lineage (Lanza, 2005).
1.2 Embryonic stem cells and pluripotency

Pluripotency describes a cell’s ability to give rise to lineages of all three germ layers
(ectoderm, mesoderm and endoderm), and to the germline, Figure 1. Stem cells can
7

be derived from several stages of the developing embryo, and these cells have a
range of differentiation potentials including many with pluripotent potential (Amy
Ralston, 2010). ESCs are derived from the inner cell mass (ICM) from the blastocyst.
These cells remain un-committed at a stage of development where individual tissues
have not yet started to form separately.
Figure 1 showing a simple schematic of pluripotency, taken
from http://tigger.uic.edu/~merrillb/pluripotency.html
Also characteristic of ESCs, as well as prolonged self renewal and multi-lineage
differentiation potential, is the formation of cell colonies and a rapid proliferation rate (Lanza, 2005).
Human ESCs are typically derived in vitro from fertilized embryos; the ICM is isolated
from the blastocyst-stage human embryo at 5 to 7 days after fertilization. After
isolation and washing in serum supplemented medium the ICM is seeded for culture
on a mitotically inactivated mouse embryonic fibroblast (MEF) feeder layer. The cells
will first reach confluence at roughly 10 to 14 days, ready to be passaged for the first
time (Elena Notarianni, 2006).
Mouse ESCs are isolated from blastocysts explanted onto a mitotically inactivated MEF
feeder layer. From this set up the ICM can be surgically removed after 2 to 5 days of
culture, then seeded onto another feeder layer, which provides the cultures with
leukaemia inhibitory factor (LIF), which can also be supplemented to the culture
medium. From this initial culture the cells are either trypsinized and passaged, or
individual colonies can be isolated and expanded (Elena Notarianni, 2006).
8

In a mature organism adult stem cells exist in specific environments, suggesting that
the sensitive conditions present are important in maintaining potency. This has
stimulated much debate on how stem cells being studied in vitro should be cultured.
1.3 Genetics of pluripotency and induced pluripotency
In the past differentiation was considered to be a one-way process; however the
work of Shinya Yamanaka confirmed that mature cells could be reprogrammed into a
pluripotent state (cells termed induced pluripotent stem cells (iPS cells)) with the
forced expression of three transcription factors Oct4, Sox2 and Klf4 and an
oncogene, c-Myc. These factors have been shown along with others to be important
in the maintenance of pluripotency. Following on from the earliest work on
reprogramming by Gurdon et al in the 1970s, major breakthroughs came from Sir Ian Wilmut’s work on ‘Dolly’, an adult sheep derived from an enucleated oocyte that had
the nucleus of a mature mammary gland cell transplanted into it (I. Wilmut, 1997). This somatic cell nuclear transfer work showed that the enucleated oocyte contained
factors that could reprogram the somatic nucleus.
Yamanka’s work identifying the factors that drive reprogramming has spurned a major
area of ongoing research into the functional relevance of the factors shown to induce a
stem-cell-like state in somatic cells. Recent work has shown that c-Myc can be
substituted. Work in neural stem cells that already express Sox2, Klf4 and c-Myc has
led to the regard in some quarters that Oct4 is a central factor of interest in both
inducing and maintaining pluripotency, supported by it’s expression profile in the
embryo (Bosnali M, 2009). Recent work also suggests that a combination of Oct4, Sox2 and NANOG are sufficient to reprogram somatic cells (Bosnali M, 2009)
(Amy Ralston, 2010). These factors are likely to represent some of the core factors
in a large array of factors that maintain the pluripotent state (Bosnali M, 2009).
Oct4 was observed as having elevated expression in embryo cells of the mouse,
and of ESCs before its discovery as being a reprogramming factor. It is thought that
Oct4 acts to repress the development of trophoblast cells in the early embryo.
Blastocytes lacking in Oct4 have an ICM that degenerates, with no endoderm
9

formation suggesting that Oct4 is involved in early lineage determination (Amy Ralston, 2010). This idea that Oct4 is a key regulator of pluripotency is supported by
RNAi work where Oct4 knockdown ESCs caused trophectodermal differentiation (Bosnali M, 2009).
Chip-Chip and ChipPET analysis are used to look at the interactions between
proteins and DNA in a cell (Rizzino, 2009). This analysis on Oct4 and Sox2 showed
that the pair co-ocupied a large number of genes in ES cells, this result also supports
the idea that Oct4 and Sox2 are key regulators in pluripotency as the co-occupied
genes turned off at the RNA level after the ES cells had differentiated (Rizzino, 2009).
Sox2 belongs to the DNA binding group of proteins termed the High Mobility Group,
and is a cofactor of Oct4. Similar experiments as above carried out focusing on
Sox2 show that Sox2 null embryos cannot form ES cells, and ES cells depleted of
Sox2 also derive trophoblast-like cells. These observations support Sox2’s role in
maintaining pluripotency, however Sox2 mutants develop defects at a later stage
than Oct4 mutants (Amy Ralston, 2010). One suggestion is that one of Sox2’s major
roles is to activate the expression of Oct4, and this is supported by experiments
where Oct4 up-regulated cells were able to continue to exhibit pluripotency in the
absence of Sox2 (Amy Ralston, 2010).
NANOG is seen currently as the third key member of a network of transcription
factors that are central in regulating pluripotency. Enhancer segments of genes that
have been considered to play a role in pluripotency and preventing differentiation
show co-occupation by these three factors (Amy Ralston, 2010) (Bosnali M, 2009).
NANOG has been shown to play roles in early lineage decision such as the
commitment to embryonic or extraembryonic endoderm; in the absence of NANOG
only extraembryonic endoderm survives. ES cells depleted of NANOG still appear to
exhibit pluripotency however develop morphological changes, and differentiate more
readily than ES controls (Amy Ralston, 2010). In mouse ES cells NANOG
overexpression can overcome the cell’s dependency, in relation to pluripotency, on
the self-renewal cytokine LIF (Amy Ralston, 2010) (Bosnali M, 2009).
10

The expression of NANOG in pluripotent cells, observed from the ICM, has been
shown to follow a ‘salt and pepper’ distribution, where some cells express the gene,
but others do not (Janet Rossant, 2003) (Berenika Plusa, 2008). Isolation and
expansion of NANOG positive cells from the ICM gives rise to the same distribution
in the subsequent descendent cells (Claire Chazaud, 2006). One suggestion is that
this establishment of epiblast and primitive endoderm progenitors in the 3.5 day ICM (Claire Chazaud, 2006).
1.4 Induced Pluripotent Stem Cells
The process of reprogramming works at an efficiency of roughly 0.1%. iPS cells
satisfy an array of assays aimed at assessing their phenotype in comparison to
ES cells. They are remarkably similar in a number of areas including:
differentiation potential, karyotype, telomeric activity as well as similar cell surface
marker activity and particular gene expression (Shi, 2009).
The forced expression of the selected genes used to induce pluripotency is achieved
in a variety of ways, Table 1, with varying success, efficiency and reproducibility of
different methods. The first studies on inducing pluripotency used retroviral vectors
to transfect the cells. After sufficient reprogramming retroviral transgenes are
silenced, however this process can often be incomplete due to DNA and histone
methyltransferase activation (Stadtfeld M, 2010). This results in low reprogramming
efficiencies and cells that rely on exogenous factor expression. Reactivation of viral
transgenes can also pose problems in reprogramming, often leading to tumour
formation (Stadtfeld M, 2010) (Yamanaka S, 2010).
A popular system for reprogramming is the use of a doxycycline inducible lentiviral
transduction system (Hamilton B, 2009). Using inducible lentiviral vectors allows
the expression to be controlled using doxycycline, with the advantage being that
cells relying on exogenous factor expression can be selected out with the removal
of doxycycline.
11
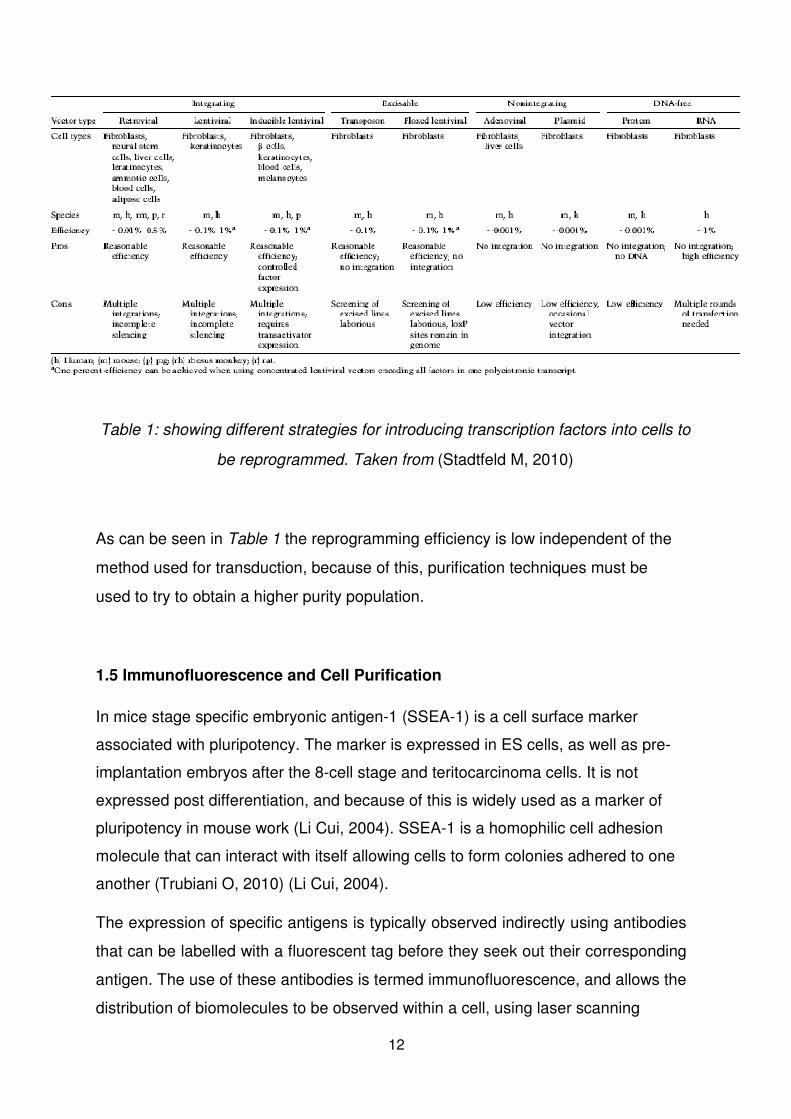
Table 1: showing different strategies for introducing transcription factors into cells to
be reprogrammed. Taken from (Stadtfeld M, 2010)
As can be seen in Table 1 the reprogramming efficiency is low independent of the
method used for transduction, because of this, purification techniques must be
used to try to obtain a higher purity population.
1.5 Immunofluorescence and Cell Purification
In mice stage specific embryonic antigen-1 (SSEA-1) is a cell surface marker
associated with pluripotency. The marker is expressed in ES cells, as well as pre-
implantation embryos after the 8-cell stage and teritocarcinoma cells. It is not
expressed post differentiation, and because of this is widely used as a marker of
pluripotency in mouse work (Li Cui, 2004). SSEA-1 is a homophilic cell adhesion
molecule that can interact with itself allowing cells to form colonies adhered to one
another (Trubiani O, 2010) (Li Cui, 2004).
The expression of specific antigens is typically observed indirectly using antibodies
that can be labelled with a fluorescent tag before they seek out their corresponding
antigen. The use of these antibodies is termed immunofluorescence, and allows the
distribution of biomolecules to be observed within a cell, using laser scanning
12
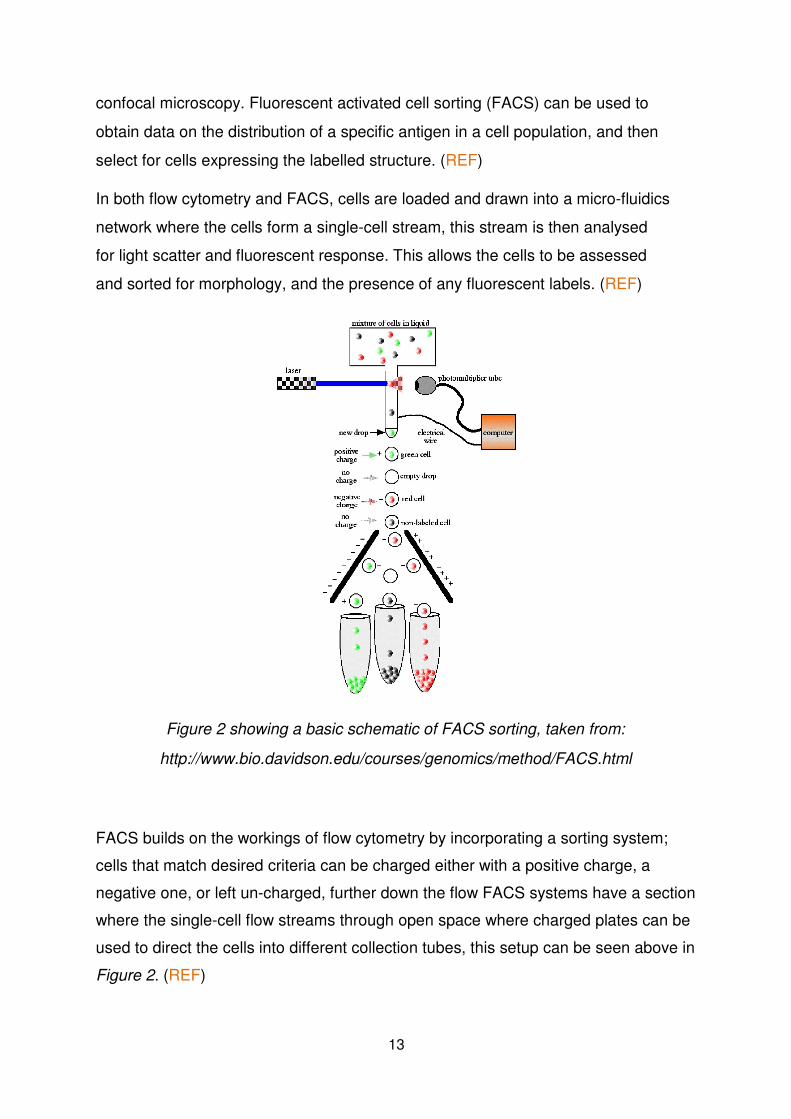
confocal microscopy. Fluorescent activated cell sorting (FACS) can be used to
obtain data on the distribution of a specific antigen in a cell population, and then
select for cells expressing the labelled structure. (REF)
In both flow cytometry and FACS, cells are loaded and drawn into a micro-fluidics
network where the cells form a single-cell stream, this stream is then analysed
for light scatter and fluorescent response. This allows the cells to be assessed
and sorted for morphology, and the presence of any fluorescent labels. (REF)
Figure 2 showing a basic schematic of FACS sorting, taken from:
http://www.bio.davidson.edu/courses/genomics/method/FACS.html
FACS builds on the workings of flow cytometry by incorporating a sorting system;
cells that match desired criteria can be charged either with a positive charge, a
negative one, or left un-charged, further down the flow FACS systems have a section
where the single-cell flow streams through open space where charged plates can be
used to direct the cells into different collection tubes, this setup can be seen above in Figure 2. (REF)
13

Another strategy for purification of induced cells is through clonal selection, by
expanding individual colonies of pluripotent cells. Reprogrammed cells form round
ES-like colonies with well defined membranes that are ‘picked’, rendered into single
cells and seeded at reducing densities in different wells of a culture plate. When
seeded at low-enough densities, single clonal colonies can be tested for their
reprogramming extent, and thus expanded. The colonies are usually large enough to
be isolated between 16 and 22 days after seeding (Hamilton B, 2009).
1.6 Differentiation
From the earliest stages of embryo development cells begin to differentiate at
varying stages and to different lineages as they become more specialized in co
ordinance with the increasing complexity of the forming organism. The down-
regulation of growth activities is an indicator of differentiation (Studzinski, 1999) (Clive W Lloyd, 1981).
Cell differentiation can result in drastic changes to a cell’s morphology and size,
metabolic activity, membrane potential and a wide range of other morphological and
phonotypic changes which are the result of gene expression and gene silencing,
resulting in specific cell gene expression profiles.
In the past differentiation was thought to be controlled by a number of ‘master-
regulatory’ genes, that if expressed would commit a cell to a particular lineage
(Moody, 2004). Examples of these ‘master-regulatory’ genes include; Pax6
expression of which would cause commitment to a retinal cell, or MyoD stimulating
commitment to a muscle cell (Moody, 2004). More recently it has been established
that more information than ‘master-regulatory’ genes alone is needed for
differentiation.
This relates well to what we can observe in the development of the early embryo
where lineage commitment is determined by its spatial location in the embryo. We
now understand that different regions in the embryo provide different signalling
environments (Moody, 2004). Different signals are now being linked with different
lineages, for example FGF and Sonic hedgehog (Shh) signalling have been shown to
influence limb bud growth. These environments are created by molecular
14

signalling transduction pathways which can have direct influence on the transcription
profile of a given cell (Moody, 2004). (REF)
Work on mesenchymal stem cells (MSCs) that were seeded on poly(lactide-co-
glycolide) (PLGA) constructs yielded data showing that initial extracellular matrix
(ECM) deposition, remodelling and integrin expression profiles have an influence on
osteogenesis. MAPK and PI3K signalling pathways were shown to be involved
(Anup K. Kundu, 2009). Work like this demonstrates that differentiation is influenced
not only by ‘master-regulatory’ genes, but also by a wide range of additional factors
such as the tissue conditions surrounding cells.
It has been established that the ECM can play a role in the differentiation of cells,
previously thought to be limited to biochemical recognition of ECM ligands,
stimulating cell receptors. Biophysical properties of the ECM have been implicated in
differentiation. Nueroblastoma differentiation, which was previously identified as
being retinoic acid-mediated, involves significant morphological changes that have
been considered to be linked to mechanotransductive signalling, and has been
recently shown to be sensitive to different ECM stiffness’s (Wilbur A Lam, 2010). Neuritogenesis was stimulated by a stiffer ECM environment, with different stiffness ECM microenvironments having different effects (Wilbur A Lam, 2010). Work like
this demonstrates the complexity and variety the conditions that influence
differentiation, or conversely the maintenance of pluripotency.
Signals also need to be experienced by the cell in a particular temporal sequence,
Figure 3. It has been observed that ESCs will only differentiate into spinal motor
neurons if subjected to a sequence of signals matching in vivo conditions from the
developing embryo (Hynek Wichterle, 2002) (Moody, 2004).
Figure 3, showing a schematic of the steps in motor neuron differentiation. Taken
from: (Hynek Wichterle, 2002).
15

A topic of much debate in many sectors of stem cell research is how the culture
environment or any other contributing factors not being controlled, may affect cell
phenotype or stimulate differentiation. Bearing in mind that we do not yet know
definitively the breadth of mechanisms that drive differentiation, it is hard to predict how
methods of culture or even the methods of recording data, of pluripotent cells could
activate differentiation-related pathways, or potentially disturb a balanced environment
needed to maintain pluripotency. This has already been examined looking at the
significant differences of cells cultured in 3-dimensional (3D), or 2-dimensional (2D)
environments. Differences between the two conditions resulted in significant differences
in mRNA expression amongst other characteristics (Robert E. Akins, 2010). This study concluded that the 3D environment was stimulating cells to
carry out more physiologically relevant activity, and express a more physiologically
relevant phenotype (Robert E. Akins, 2010).
Mouse ESCs cultured in typical rigid constructs or monolayer in the presence of LIF
are prone to spontaneous differentiation; and the gene expression profiles of these
cells show that the expression of genes associated with pluripotency fluctuates, as
does the expression of genes associated with committed lineages. Recent work
using substrates that closely match the stiffness of the cells, found that after five
days in the absence of LIF colonies of undifferentiated cells that showed high
Alkaline Phosphatase activity, as well as high levels of Oct3/4 and NANOG
expression were present. These cells also efficiently formed teratomas and
embryoid bodies (Farhan Chowdhury, 2010). These results suggest that the use of a
soft substrate promotes homogeneously pluripotent colonies involving biophysical
signalling, and importantly that in the future better understanding and subsequent
replication of the in vivo environment that maintains cell potency will be likely to
result in improved in vitro maintenance of pluripotency.
1.7 Relevance of stem cells to medicine
There is the potential that if the differentiation pathways were fully understood, and
were possible to control, as well as the replication of an in vivo environment, that
16

stem cells could be used to generate all mature tissues. This could enable
compromised tissues to be replaced by tissues derived from stem cells, and if iPS
cells were also fully understood there is the possibility that tissues could be derived
from host somatic cells that were reprogrammed (Bosnali M, 2009) (Catherine M. Verfaillie M. F., 2002). (REF)
As well as the potential in tissue engineering stem cells could reveal the workings of
normal tissue development, as well as disease progression. The use of tissue grown
in vitro could also allow new drug testing to accelerate, as toxicity levels could be
modelled directly on the tissue they are intended for, as well as for example the
liver’s ability to remove the toxins over time. (REF)
1.8 Nuclear Structure and Chromatin
Figure 4, showing a human somatic nucleus, Taken from:
http://commons.wikimedia.org/wiki/File:Diagram_human_cell_nucleus.svg
17
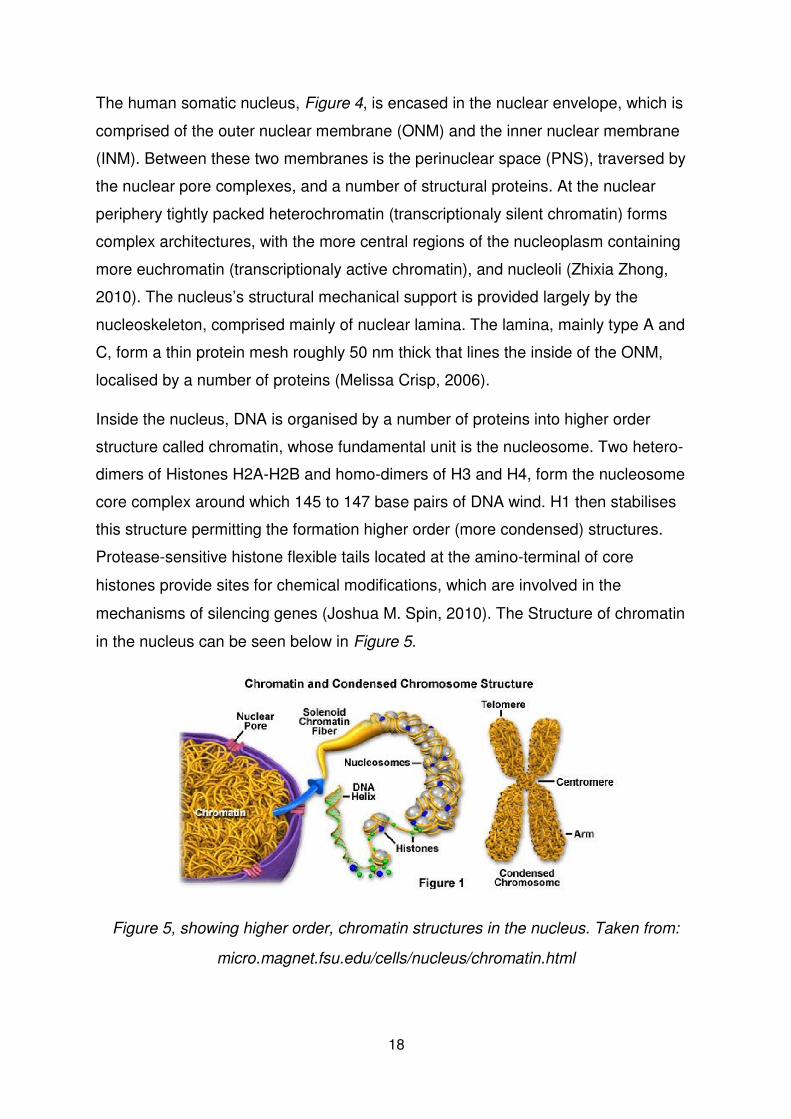
The human somatic nucleus, Figure 4, is encased in the nuclear envelope, which is
comprised of the outer nuclear membrane (ONM) and the inner nuclear membrane
(INM). Between these two membranes is the perinuclear space (PNS), traversed by
the nuclear pore complexes, and a number of structural proteins. At the nuclear
periphery tightly packed heterochromatin (transcriptionaly silent chromatin) forms
complex architectures, with the more central regions of the nucleoplasm containing
more euchromatin (transcriptionaly active chromatin), and nucleoli (Zhixia Zhong,
2010). The nucleus’s structural mechanical support is provided largely by the
nucleoskeleton, comprised mainly of nuclear lamina. The lamina, mainly type A and
C, form a thin protein mesh roughly 50 nm thick that lines the inside of the ONM,
localised by a number of proteins (Melissa Crisp, 2006).
Inside the nucleus, DNA is organised by a number of proteins into higher order
structure called chromatin, whose fundamental unit is the nucleosome. Two hetero-
dimers of Histones H2A-H2B and homo-dimers of H3 and H4, form the nucleosome
core complex around which 145 to 147 base pairs of DNA wind. H1 then stabilises
this structure permitting the formation higher order (more condensed) structures. Protease-sensitive histone flexible tails located at the amino-terminal of core
histones provide sites for chemical modifications, which are involved in the
mechanisms of silencing genes (Joshua M. Spin, 2010). The Structure of chromatin
in the nucleus can be seen below in Figure 5.
Figure 5, showing higher order, chromatin structures in the nucleus. Taken from:
micro.magnet.fsu.edu/cells/nucleus/chromatin.html
18

1.9 Heterochromatin protein 1
Heterochromatin protein 1 (HP1) is a component of transcriptionaly silent
chromatin, which binds to specific lysine residues in the histone tail which have
been methylated, and acts as a transcriptional repressor (Weihua Zeng, 2010). A
suggested model of the mechanisms of gene silencing has been devised involving
Histone Methyltransferase (HMTase), which interacts with HP1 and the modified
histone H3. It is thought that interactions between these three components are
responsible the gene silencing, and the formation of heterochromatin (Pimpinelli,
2008) (Festenstein, 2005).
As well as playing a large role in gene silencing, HP1 has also recently been shown
to have other functions such as DNA repair, telomere maintenance, transcriptional
elongation and cohesion of centromeric sister chromatin (Weihua Zeng, 2010).
1.10 The LINC complex
The LINC complex encompasses a number of proteins that form a molecular chain,
connecting the cytoskeleton to the nuclear lamina, providing a mechanical
association. At the ONM boundary to the cytoplasm the KASH (Klarsicht-Anc1- Syne1 Homolgy) domain; giant transmembrane proteins, mainly isoforms of Nesprin
(Nuclear Envelope SPectRIN repeats)-1 and -2, traverse the ONM protruding into the
PNS by ~ 30 amino acids. The KASH domain of these proteins is ~ 50 amino acids in
length, and although there are four KASH domain genes encoded in humans,
SYNE1, SYNE2, Nesprin-3 and Nesprin-4, it is mainly the transcribed and spliced
varients of SYNE1 and -2 that form the majority of the massive, some greater than
1MD, functional proteins that associate directly with the actin or plectin of the
cytoplasm (Zhixia Zhong, 2010) (Melissa Crisp Q. L., 2006) (P.J. Stewart-Hutchinson,
2008). The spectrin-like repeats (SLR) of the giant Nesprins suggest that the proteins
have elastic properties, which is also supported by the fact that the lamina network’s
mechanical properties suggest the presence of a molecular shock absorber (Kris
Noel Dahl1, 2004). Some Nesprin isoforms, smaller in size, associate with the INM
binding to nuclear lamins and emerin (Zhixia Zhong, 2010).
19

The other main components of the LINC complex are the ~ 200 residue Sad1-UNC- 84 homology (SUN) domain proteins, encoded by four genes in humans, SUN1, -2,
- 3 and SPAG4. SUN1 and SUN2 are the main transmembrane proteins localising at
the INM. Similar to the KASH domain of the giant Nesprin proteins, SUN1 and -2
proteins also have a SUN-domain region that extends into the PNS where it
associates with the KASH domain of the corresponding Nesprins (Zhixia Zhong, 2010) (P.J. Stewart-Hutchinson, 2008). At the opposite end the SUN protein’s N-
terminal domain binds nuclear lamina, completing the mechanical link from the
cytoskeleton to the nucloeskelton (Zhixia Zhong, 2010). The arrangement of the
LINC complex in the context of the nuclear envelope can be seen below in Figure 6.
Figure 6, showing the LINC complex, taken from: (P.J. Stewart-Hutchinson, 2008)
SUN2 is a protein of particular interest due to its lack of expression in the pluripotent
nucleus, and its direct association with the nuclear lamina at the INM. The SUN
proteins together with the NESPRINs provide a mechanical link between the
nucleoskelton and the cytoskeleton. Studies Found that knocking down the
expression of SUN2 affected the positioning of the nucleus during cell migration, and
the orientation of centrosomes which caused alterations in cell polarity (Ozerlat, 2010).
1.11 Gene Silencing
Chemical modifications to histones in conjunction with specific enzymes determine
determine whether a gene will be targeted by polymerase – determining whether a
gene is transcriptionally active or not. This is important during development and in
the cell cycle as different genes will be active at different times, and ultimately for a
20

cell to ‘specialize’ a specific combination of genes will be expressed (Paro, 2011). Activity of the enzyme HMTase has long been implicated in the process of gene
silencing, originally highlighted in work looking at the methylation of a particular lysine
(K9) residue in the tail of histone H3. Long-term heterochromatin silencing is
characterized by histone hypoacetylation and the methylated modifications, H3K9me,
as well as the appearance of HP1. Heterochromatin exists in a condensed state for
most of the cell cycle, and only replicates late in S phase (Andrew W. Snowden,
2002) (Paro, 2011). HP1 associated transcriptionally repressed chromatin in its more
common condensed state is termed constitutive heterochromatin, the other variation
is facultative heterochromatin, which exists in the euchromatic region of the nucleus,
an example being the silent ‘X’ chromosome in female somatic mammalian cells (Paro, 2011).
Some repressed genes must be maintained in a stable repressed state for long
periods of time, this is mediated by interactions with the heterogeneous class of Polycomb group (PcG) proteins, which regulate long term, stable repression (Paro, 2011). This form of gene suppression is characterized by the methylation of histone H3 at Lysine 27 (H3K27me), mainly involved in the suppression of genes associated
with development decisions (Paro, 2011).
Methylation and acetylation are among the most well documented post-translational
modifications to the lysine residues of Histones H3 and H4 tails that influence gene
expression. Methylations, depending on the particular modification, can either
determine repression, H3K9, H3K27, H4K20, or transcriptional activation, H3K4,
H3K36, H3K79, as well as depending on the extent to which the residue is modified
(Cathérine Dupont, 2009). In contrast all the varying acetylation modifications have
been linked with transcriptional promotion. Other transcription determining
modifications include modifications to H2A and H2B (Cathérine Dupont, 2009).
Another important component of gene expression are Class 1 Histone Deacetylases (HDAC1, -2, -3, -8), enzymes that form higher order complexes which are directed to
chromatin by transcription factors (Oliver M. Dovey, 2010).
21

1.12 Chromatin Architecture
As cells move from a pluripotent state, through differentiation, to a committed lineage
the chromatin in their nucleus compacts and forms complex higher-order structures,
particularly at the nuclear periphery. These changes go hand-in-hand with the
systematic silencing of genes, forming higher-order chromatin structures, as cells
adopt a more specific gene expression profile with differentiation. In the pluripotent
nucleus there is little chromatin compaction, and it is considered to be evenly spread
and free to ‘flow’ (Kashif Ahmed., 2010).
Figure 7 Figure 8
Figure 7 showing a mouse epiblast, Figure 8 showing mouse primitive endoderm,
taken from: (Kashif Ahmed., 2010).
Figure 7 and Figure 8 are spectroscopic electron microscope images showing
phosphorous-rich structures, nucleic acids, in yellow, and nitrogen-rich structures,
proteins, in blue. Figure 7 shows a mouse epiblast (day 3-5), with Figure 8 showing a
mouse primitive endoderm (after day 5); indicated by the arrows in Figure 8 are
areas of chromatin compaction, where complex architectures have begun to form,
more so than in the epiblast in Figure 7, which is earlier along the differentiation
pathway (Kashif Ahmed., 2010).
It is now considered that there is a link between developmental potential and
chromatin architecture and compaction, and the same changes can be seen in
comparing any number of different stages of development; such as from one-cell to
two-cell embryos, or between epiblast and trophectoderm, unanimously showing an
increase in chromatin architecture complexity and compaction with differentiation.
22

This relationship was strengthened by demonstrating that pluripotent cells stripped of Oct4 showed increased regions of compaction (Kashif Ahmed., 2010).
1.13 Alterations to Nuclear Mechanics with Differentiation
The mechanics of the nucleus undergo notable changes as cells progress towards
committed lineages. This is partly linked with chromatin compaction, as compact
chromatin is stiff, whereas if it is not compacted into architectures it flows in the
nucleus (J. David Pajerowski, 2007). This relationship works in co ordinance with the
silencing of genes as cells differentiate, and with structural reorganisations that also
occur. The contribution of chromatin to the mechanical properties of the nucleus has
been established; micropipette aspiration of different stage nuclei demonstrates that
not only is the pluripotent nucleus more deformable than mature nuclei ,Figure 9, but
also that the chromatin itself flows in the pluripotent nucleus, Figure 10 (Kris Noel Dahl, 2005) (J. David Pajerowski, 2007).
Figure 9, showing micropipette aspiration of cell nuclei, taken from: (J.
David Pajerowski, 2007)
23
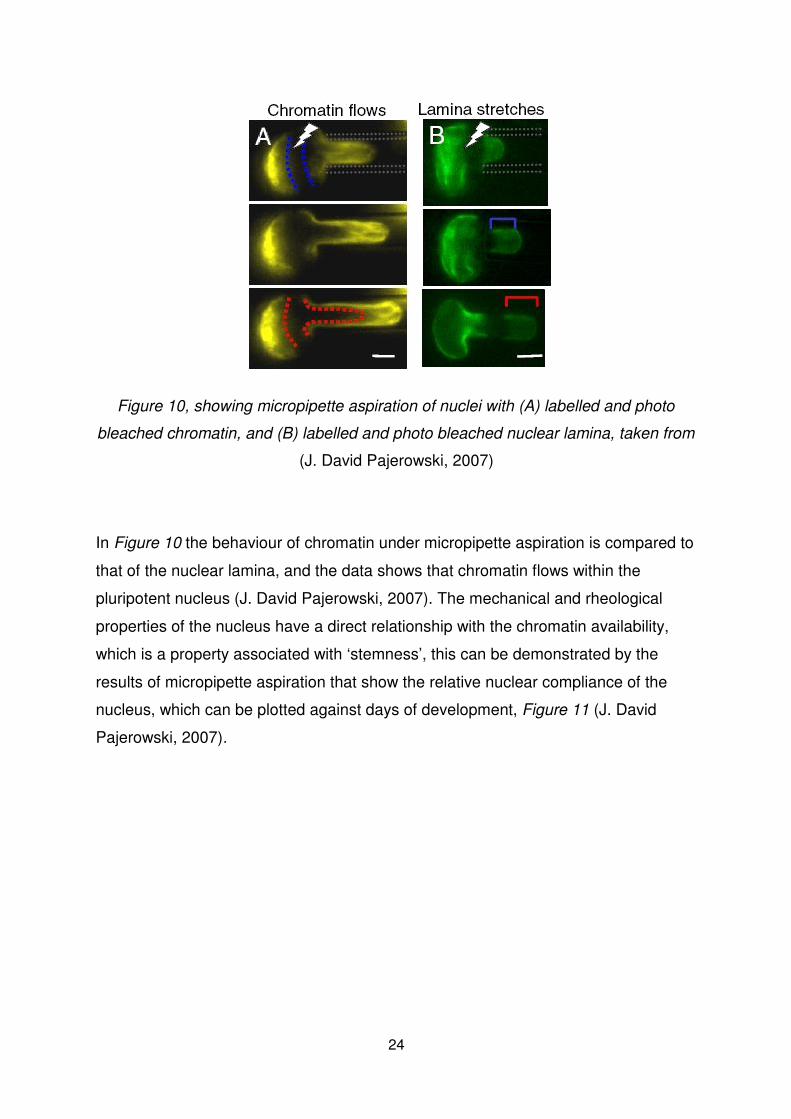
Figure 10, showing micropipette aspiration of nuclei with (A) labelled and photo
bleached chromatin, and (B) labelled and photo bleached nuclear lamina, taken from
(J. David Pajerowski, 2007)
In Figure 10 the behaviour of chromatin under micropipette aspiration is compared to
that of the nuclear lamina, and the data shows that chromatin flows within the
pluripotent nucleus (J. David Pajerowski, 2007). The mechanical and rheological
properties of the nucleus have a direct relationship with the chromatin availability,
which is a property associated with ‘stemness’, this can be demonstrated by the
results of micropipette aspiration that show the relative nuclear compliance of the
nucleus, which can be plotted against days of development, Figure 11 (J. David
Pajerowski, 2007).
24
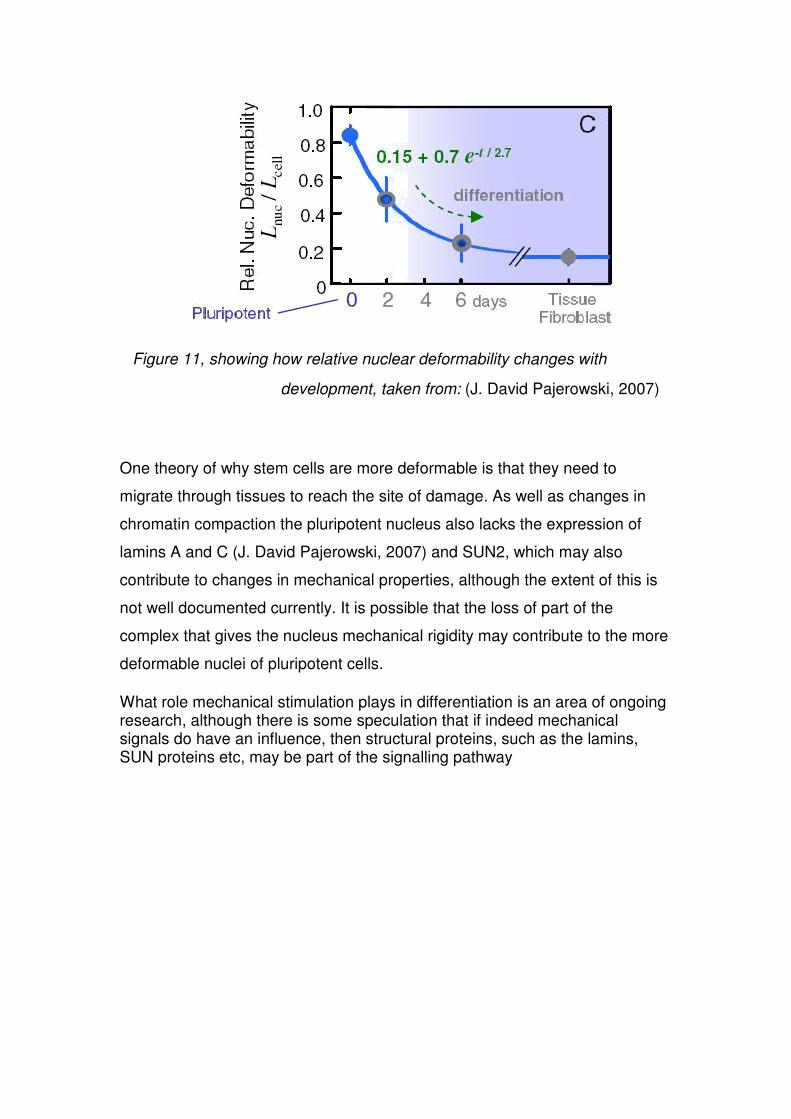
Figure 11, showing how relative nuclear deformability changes with
development, taken from: (J. David Pajerowski, 2007)
One theory of why stem cells are more deformable is that they need to
migrate through tissues to reach the site of damage. As well as changes in
chromatin compaction the pluripotent nucleus also lacks the expression of
lamins A and C (J. David Pajerowski, 2007) and SUN2, which may also
contribute to changes in mechanical properties, although the extent of this is
not well documented currently. It is possible that the loss of part of the
complex that gives the nucleus mechanical rigidity may contribute to the more
deformable nuclei of pluripotent cells.
What role mechanical stimulation plays in differentiation is an area of ongoing research, although there is some speculation that if indeed mechanical signals do have an influence, then structural proteins, such as the lamins, SUN proteins etc, may be part of the signalling pathway





























