Embed Size (px)
Citation preview

Serotonergic Systems in the Spinal Cordof the Amphibian Urodele
Pleurodeles waltl
PASCAL BRANCHEREAU,1* JOSE JULIO RODRIGUEZ,2 ISABELLE DELVOLVE,3
DJOHER NORA ABROUS,2 MICHEL LE MOAL,2AND JEAN-MARIE CABELGUEN3
1Laboratoire de Neurobiologie des Reseaux, C.N.R.S. et Universite Bordeaux 1, U.M.R.5816, F-33405 Talence, France
2INSERM U259, F-33077 Bordeaux, France3INSERM E9914, F-33077 Bordeaux, France
ABSTRACTThe role of the monoamine serotonin (5-HT) in modulating the neural networks underlying
axial locomotor movements was studied in an adult amphibian urodele, Pleurodeles waltl. 5-HTwas applied to an in vitro brainstem–spinal cord preparation of P. waltl, which displayed fictiveaxial locomotor patterns following bath application of N-methyl-D-aspartate (5 mM) withD-serine (10 mM). Our results showed that 5-HT (1–25 mM) produces a reversible increase in thecycle duration and the duration of rhythmic bursting activity recorded extracellularly fromventral roots innervating the axial musculature. When applied alone, 5-HT does not trigger axiallocomotor activity. The distribution pattern of 5-HT immunoreactive (5-HT-ir) cells along thespinal cord was investigated both in intact and in chronic spinal animals. The number of 5-HT-ircell bodies is higher at brachial levels and decreases through crural levels. Sparse oval or fusiform5-HT-ir somata are present within the gray matter, just ventrolateral to the central canal.Longitudinal fibers were detected throughout the entire white matter, except in the medial partof the dorsal funiculi. Two columns of intensely labeled and profusely branching thick and thinfibers associated with numerous varicosities run continuously along the ventrolateral surface ofthe spinal cord. Three weeks following full spinal cord transection at the level of the second spinalroot, all longitudinal processes had disappeared, indicating their supraspinal origin, whereas theventrolateral plexes remained, suggesting that they originated from intraspinal 5-HT-ir cellbodies. Our data showing that spinal 5-HT is organized according to a rostrocaudal gradientsuggest that the 5-HT systems of P. waltl are not related to the presence of limb motor pools butmore likely are related to axial central pattern generators (CPGs) networks down the length ofthe spinal cord. The possible involvement of these two sources (descending vs. intraspinal) of5-HT innervation in the modulation of the axial CPGs is discussed. J. Comp. Neurol. 419:49–60,2000. © 2000 Wiley-Liss, Inc.
Indexing terms: serotonin; immunohistochemistry; spinal cord; salamander; locomotion
The vertebrate spinal cord contains networks called cen-tral pattern generators (CPGs) that are capable of gener-ating the complex muscular synergies producing locomo-tor movements (Grillner and Wallen, 1985). The activity ofthese locomotor networks is continuously modulated byseveral neuroactive substances in order to adapt theircentrally generated output patterns to external require-ments (Rossignol and Dubuc, 1994). Among these sub-stances, serotonin (5-hydroxytryptamine; 5-HT) has beenshown to play an important role in modulation of limblocomotor CPGs in cat (Barbeau and Rossignol, 1990),rabbit (Viala and Buser, 1969), adult rat and mice (Vol-
icier, 1969), neonatal rat (Cazalets et al., 1992; Cowleyand Schmidt, 1994; Kiehn and Kjaerulff, 1996; Bracci etal., 1998), and mudpuppy (Jovanovic et al., 1996) and
Grant sponsor: Universite Bordeaux 1; Grant sponsor: Conseil Regionald’Aquitaine.
*Correspondence to: Dr. Pascal Branchereau, Laboratoire de Neurobi-ologie des Reseaux, C.N.R.S. et Universite Bordeaux 1, U.M.R. 5816,avenue des Facultes, F-33405 Talence cedex, France.E-mail: [email protected]
Received 11 May 1999; Revised 24 November 1999; Accepted 29 Novem-ber 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 419:49–60 (2000)
© 2000 WILEY-LISS, INC.

axial locomotor CPGs in tadpole (Sillar et al., 1992; Wool-ston et al., 1994) and lamprey (Harris-Warrick and Cohen,1985; Christenson et al., 1989; Wallen et al., 1989b; Mat-shushima and Grillner, 1992).
We have begun to investigate the axial CPG for locomo-tion in a limbed vertebrate, the adult urodele P. waltl. Wechose this experimental model because this animal canspontaneously exhibit two distinct locomotor modes(swimming and overground stepping), which involve twodistinct patterns of activation of axial muscles with differ-ent intersegmental coordination (Delvolve et al., 1997).Moreover, we have recently developed an isolated in vitrobrainstem–spinal cord preparation of P. waltl in whichbath-applied N-methyl-D-aspartate (NMDA) induces twodistinct fictive axial motor patterns, one displaying themain features of forward swimming and the other those ofstruggling (Delvolve et al., 1999).
The present study concerns the modulatory actions of5-HT on the axial CPG for locomotion in P. waltl. In a firststep, we have studied the effects of bath-applied 5-HT onNMDA-induced axial locomotor activities in the in vitrobrainstem–spinal cord preparation. Then, we performedan immunohistochemical analysis of 5-HT systemsthroughout the spinal cord. This anatomical study wasperformed both in intact and in chronic spinal animals inorder to identify 5-HT-immunoreactive (5-HT-ir) fibersdescending from supraspinal structures.
Our results indicate that 5-HT profoundly modulatesfictive axial locomotor activity and that spinal 5-HT-containing systems have two components, descending andintraspinal. The latter is organized according to a rostro-caudal gradient along the cord without preferential distri-bution at the level of limb motor pools. These data there-fore suggest that, in P. waltl, 5-HT plays an important rolein the operation of the axial CPG for locomotion.
MATERIALS AND METHODS
Fourteen adult salamanders were used for the presentstudy. Animals were obtained from the Centre de Biologiedu Developpement (CNRS UMR 9925, France) and kept inan aquarium at room temperature. Surgical procedures,pre- and postoperative care, and handling and housing ofthe animals were according to protocols approved by theEuropean Communities Council and conformed to NIHguidelines.
Electrophysiology
Animals (n 5 4) were anesthetized by immersion in a0.1% aqueous solution of tricaine methanesulfonate MS-222 (Sigma, St. Louis, MO). After removal of the lowerpart of the tail, limbs, intestines, and skin, the craniumand the remainder of the vertebral column were placed ina Petri dish and superfused with cooled (13°C) and oxy-genated amphibian Ringer’s (130 mM NaCl, 2.1 mM KCl,2.6 mM CaCl2, 0.2 mM MgCl2, 4 mM HEPES, 5 mMglucose, 1 mM NaHCO3, pH 7.5). The entire brain wasthen exposed and the part rostral to the mesencephalonremoved. A dorsal laminectomy was performed to exposethe first 16 segments of the spinal cord. The remainingmuscles and bones caudal to the fourth spinal segmentwere completely removed. The preparation (see Fig. 1A)was then pinned down, ventral side up, in a Sylgard-linedchamber and superfused (3 ml/min) with cooled and oxy-genated amphibian Ringer’s to which was added the irre-
versible neuromuscular blocking agent a-bungarotoxin(a-BGTX; Sigma; 1 mM). The preparation was then keptunder such conditions at 13°C for ;15 hours before exper-imentation began.
Axial motor activity was recorded extracellularly in aconventional way from the 9th ventral roots with glasssuction electrodes, digitized, and stored on computer harddisk (Cambridge Electronic Design 1401). Fictive locomo-tor activity was induced by adding 5 mM NMDA (Sigma) tothe superfusate together with 10 mM D-serine (D-ser; Sig-ma; see Wheatley and Stein, 1992; Delvolve et al., 1999).Interactive software (Spike2 for Windows) was used todetermine the onset and cessation of ventral root bursts ofactivity. The cycle duration of motor rhythmicity was mea-sured between successive bursts of ventral root discharge.The phase difference between bilateral bursting activity,expressed as percentage cycle duration, was calculated bydividing the interval between midbursts of ipsi- and con-tralateral ventral roots by the duration of the correspond-ing fictive locomotor cycle.
Surgery
To identify 5-HT-ir fibers descending from supraspinalstructures, the spinal cords of animals (n 5 3) weretransected at the level of the second spinal roots accordingto the following procedure. Animals were first anesthe-tized with MS-222. A small cut was made in the skin andmuscle overlying the spinal cord 1 cm rostral to the fore-limbs, and then the spinal cord was exposed by laminec-tomy. The number of vertebrae removed was sufficient toallow the underlying spinal cord to be picked up betweenthe blades of microdissecting scissors and cut. When thetransection was complete, the two ends of the spinal cordretracted, producing a 2 mm gap. The cavity was flushedwith Ringer’s to allow visualization of both ends of the cutspinal cord, and a ball of sterile cotton was used to swabthe vertebral canal to remove any remaining pieces oftissue. The wound was then closed with sutured. Postop-erative time (3 weeks) was sufficient to ensure maximaldegeneration of sectioned descending fibers (Davis et al.,1989).
Immunohistochemistry
Control animals (n 5 7) and transected animals (n 5 3)were anesthetized and perfused transcardially with 1.5%glutaraldehyde. The nervous tissue was exposed, and thepreparation was postfixed for 3 hours. After washing inphosphate buffer (PB; pH 7.4), the spinal cord was re-moved and partitioned into segments that were cryopro-tected and sectioned on a freezing microtome (HM 500M;Microm, Germany) at 20 mm in the frontal or horizontalplan.
Sections were treated with 1% NaBH4 for 30 minutesand washed extensively with PB. They were then incu-bated in a humid chamber in 1) 0.1 M phosphate-bufferedsaline (PBS; pH 7.4) containing 0.3% Triton X-100 with3% normal goat serum for 45 minutes; 2) a rabbit anti-serum directed against 5-HT (gift from Dr. M. Geffard; seeGeffard et al., 1985) diluted 1:10,000 in 0.1 M PBS plus0.3% Triton X-100 with 0.3% normal goat serum over-night; 3) 0.1 M PBS (three changes); 4) Biotin conjugategoat anti-rabbit antiserum (Dako, France) diluted 1:200 in0.1 M PBS for 90 minutes; 5) 0.1 M PBS (three changes);6) streptavidin-biotinylated horseradish peroxidase com-plex diluted 1:200 (Dakopatts, France) for 60 minutes; 7)0.1 M PBS (three changes); and 8) 0.1 M Tris (pH 7.4).
50 P. BRANCHEREAU ET AL.

Finally, the immune complex was visualized by develop-ment with 0.1% 3,39-diaminobenzidine tetrahydrochloride(DAB; Sigma) and 0.01% H2O2 in 0.1 M Tris for approxi-mately 8 minutes. Sections were then rinsed, dehydrated,and coverslipped with Entellan. As a control for the spec-ificity of the immunochemical reactions, some sectionswere processed with normal goat serum diluted in PBSinstead of the specific 5-HT antibody. No labeled elementswere observed under the latter condition.
Quantification
Cell bodies. In control animals, the number of 5-HT-ir–labeled perikarya was taken as the mean numbercounted in serial sections of spinal cords from three ani-mals from segments 1–28. In animals with spinal transec-tion, 5-HT-ir somata were counted in representative seg-ments taken from the three operated animals. Becausemethodological considerations made it difficult to use dis-sector methods (see Coggeshall and Lekan, 1996), weadopted the Abercrombie method (Abercrombie, 1946) as away of comparing populations. Numbers of counted5-HT-ir profiles were then multiplied by a correction fac-tor, F, previously defined by Abercrombie (F 5 T/T 1 S,with T being the thickness of the section [20 mm] and S themean size of the somata [12.4 6 0.1 mm, mean 6 s.e.; n 5160]). On this basis F was calculated as 0.62. To testwhether the sizes of the 5-HT-ir somata differ in differentportions of the cord, which would present a major obstacleto using the Abercrombie correction method, we measuredcell diameters in horizontal sections of selected segmentsof a control animal cord. The following means 6 s.e. werefound: S2, 13.71 6 0.31 mm; S3, 13.89 6 0.30 mm; S10,12.30 6 0.44 mm; S11, 12.14 6 0.46 mm; S18, 12.60 6 0.36mm; S19, 13.03 6 0.70 mm. A Kruskal-Wallis test appliedto these values did not reveal any significant difference(H 5 14.96, df 5 5) between segments.
Varicosities. For analysis of the distribution of5-HT-ir varicosities in the ventrolateral plexus, spinalcords were examined at four different segmental levels: 1)a cervical level (S3) including innervation of the forelimbs;2) a thoracic level (S10); 3) a lumbar level (S16) includinginnervation of the hindlimbs; and 4) a sacral level (S24),which includes innervation of the tail. Microscope focuswas adjusted in order to visualize the largest part of theplexus, and all visible varicosities were counted manually.This was performed at high magnification on 10 represen-tative sections at every level of the cord in both control andchronic animals.
Statistical analysis
All values were expressed as mean 6 s.e. Statisticalsignificance of variance was assessed by a paired two-tailed Student’s t-test or by one-way analysis of variancefollowed by a pairwise multiple comparison procedure(Tukey test).
Photomicrography
An optical microscope (Leica) was used to observe andphotograph immunoreactive structures. The contrast ofthe brown diaminobenzidine reaction product was in-creased by the use of interference filters. All photomicro-graphs were printed, scanned, and imported into CorelDraw software, with which figures were made.
RESULTS
Modulatory action of 5-HT
It was recently shown that two distinct axial motorprograms can be generated by the in vitro brainstem spi-nal cord preparation of P. waltl after adding NMDA (5mM) with D-Ser (10 mM) to the bath (Delvolve et al., 1999).The first, “fast” pattern is characterized by cycle periodsranging from 3 to 7 seconds; the second, “slow” patterndisplays longer cycle periods (10–16 seconds). In addition,the fast pattern exhibits a rostrocaudal propagation ofrhythmic activity down the spinal cord, whereas the slowpattern is characterized by a caudorostral propagation ofrhythmic activity up the cord (Delvolve et al., 1999). Bothaxial motor patterns are characterized by an alternationbetween activity on the two sides.
Bath application of 5-HT (1–25 mM; n 5 4) profoundlymodulated both fast and slow NMDA-induced motor pat-terns (Fig. 1B). The mean cycle duration was significantlyincreased (t-test, P , 0.001) from 4.53 6 0.21 seconds (n 512) to 7.54 6 0.26 seconds (n 5 13) and from 13.31 6 0.52seconds (n 5 12) to 17.79 6 0.26 seconds (n 5 24) for fastand slow motor patterns, respectively (Fig. 1C). The aver-age burst duration was also significantly increased (t-test;P , 0.001) by 5-HT from 1.50 6 0.13 seconds (n 5 12) to3.43 6 0.36 seconds (n 5 14) during the fast motor patternand from 5.08 6 0.36 seconds (n 5 13) to 8.84 6 0.19seconds (n 5 26) during the slow motor pattern (Fig. 1D).All these effects were fully reversible after washing outthe 5-HT with normal Ringer’s (not illustrated).
With three preparations, we tested the ability of 5-HT toelicit fictive locomotion when applied in the absence ofNMDA and D-Ser. In all trials 5-HT tonically activatedventral root firing without eliciting rhythmic motor pat-terns. This result suggests that in P. waltl 5-HT is morelikely to be involved in modulating ongoing rhythmic ac-tivity than in activating the axial CPG for locomotion.
Spinal 5-HT immunoreactivity incontrol animals
Neuronal profiles. Intraspinal 5-HT-containing perikaryawere found intermixed in the gray matter and just ventro-lateral to the central canal of the spinal cord (Fig. 2A).Transverse sections revealed that these 5-HT-ir somata sentprocesses in the lateral direction in the white matter andalso in the ventral direction along the ependymal lining (Fig.2B). Horizontal sections further revealed that 5-HT-ir so-mata could be either oval (type I neurons) or fusiform inshape (type II neurons; Fig. 3; see also Jovanovic et al.,1996). Although a comparative quantification of these twoneuronal types was not attempted because of the difficulty ofaccurately classifying all 5-HT-ir somata, we found that typeI neurons were about 10 times more numerous than type IIneurons in every segment of the spinal cord.
Type I neurons were bipolar (Fig. 3A) or multipolar (Fig.3B), with perikarya ranging from 10 to 18 mm in diameter.Their processes extended primarily in the lateral directionwithin the white matter, and their somata were often seenin the vicinity of processes originating from other type Ineurons (Fig. 3A), suggesting that these serotonergic neu-rons may interact along the spinal cord. The processes ofmultipolar type I neurons often bifurcated, forming a Tshape, with one collateral oriented longitudinally and theother laterally (Fig. 3B). Varicose 5-HT-ir longitudinalfibers were sometimes observed in close apposition withsomata of oval cell bodies (Fig. 3C).
Type II neurons were located in close proximity to the
51SEROTONIN IN URODELE SPINAL CORD

Fig. 1. Bath application of 5-HT modulates NMDA-induced fast andslow axial motor patterns in P. waltl. A: Schematic drawing of the invitro brainstem–spinal cord preparation. Ipsilateral (i) and contralateral(co) ventral root (VR) activity was recorded during superfusion with amixture of NMDA and D-Ser. Note that muscles and bones (shaded area)rostral to the fourth ventral root were partially preserved. Muscle con-tractions were suppressed by using a-bungarotoxin (1 mM). B: Ventralroot activity in the presence of NMDA 1 D-Ser (Control) and 20 minutes
after the onset of bath addition of 5-HT (5 mM; lower panel). Noteincreased cycle period and burst duration in the presence of 5-HT forboth fast and slow axial motor patterns. C: Histogram of mean cycleduration for fast and slow motor patterns before (gray bars) and after(black bars) bath application of 5-HT. D: Histogram of mean burstduration. Same representation as in C. Error bars in C and D indicatestandard errors, and asterisks indicate statistical significance (***P ,0.001, t-test).
52 P. BRANCHEREAU ET AL.

central canal, and their perikarya ranged from 12 to 16mm in their long axis and from 4 to 6 mm in their shortaxis. They exhibited two processes arising from each ex-tremity of the soma (Fig. 3D) and running in the rostro-caudal direction, adjacent and slightly lateral to the wallsof the central canal. One of these two processes oftencontained varicosities and was therefore considered to bethe cell’s axon (Fig. 3D). Appositions between varicose5-HT-ir longitudinal fibers and somata of type II neuronswere sometimes observed (Fig. 3D).
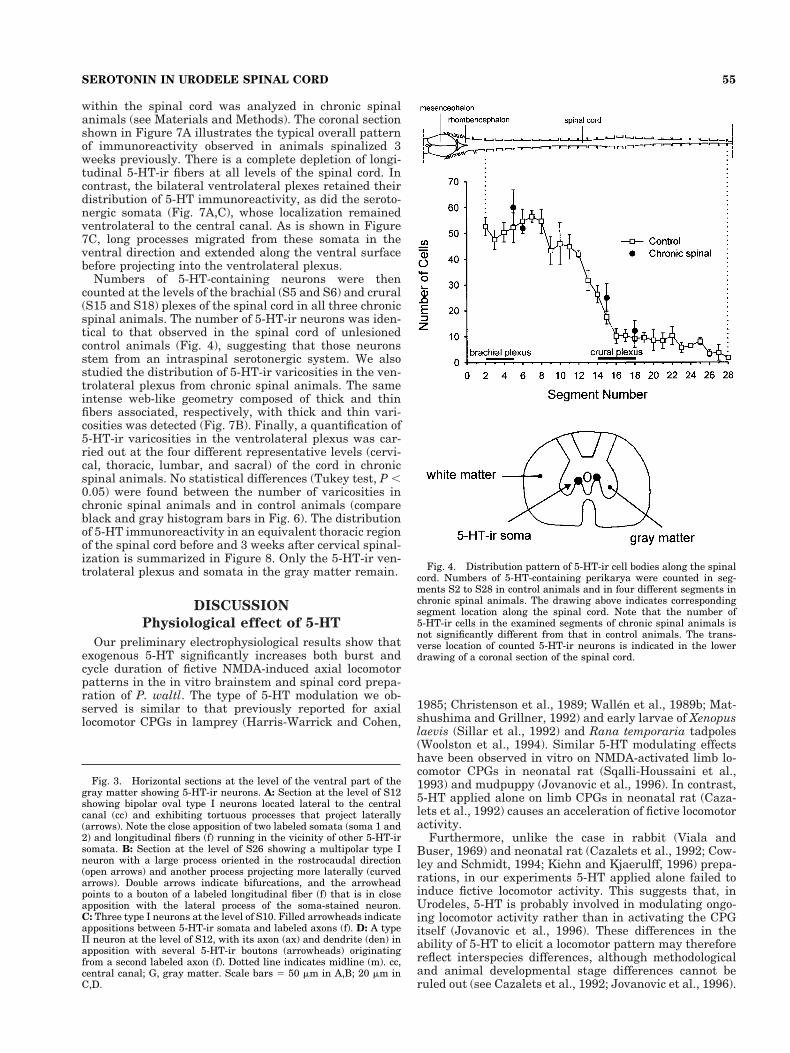
The number of 5-HT-ir perikarya was counted in hori-zontal sections of each spinal segment from the cervical(S2) to sacral levels (S28). The graph in Figure 4 illus-trates pooled data obtained from three animals. The great-est density of 5-HT-ir somata was found between S2 andS8, where 45–65 somata per segment were counted. Be-tween S8 and S16 the number of 5-HT-ir somata de-creased progressively to reach 6–16 per segment, thendecreased further at the sacral level to attain only 0–6cells per segment.
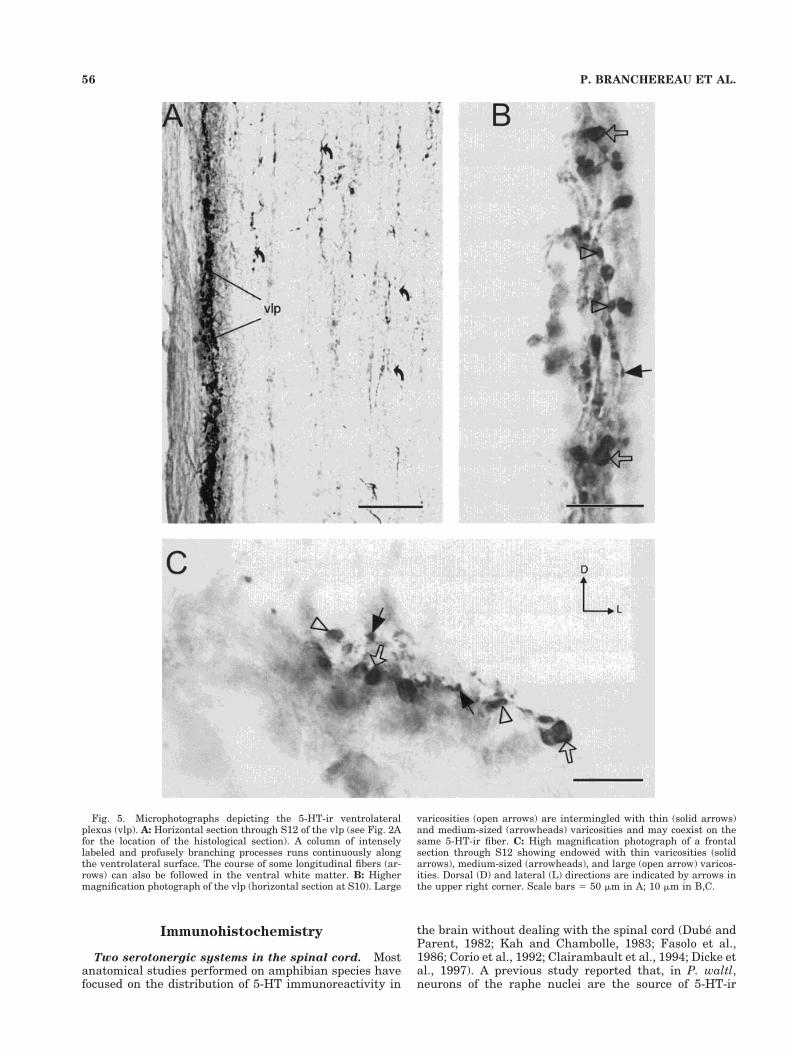
5-HT-ir ventrolateral plexus. Coronal sections (asshown in Fig. 2A) showed an intense 5-HT-containingmeshwork in two patches that were located near the ven-trolateral surface of the cord. We refer to each of thesebilateral 5-HT-ir regions as ventrolateral plexes. In hori-zontal sections, each ventrolateral plexus appeared as acolumn of intensely labeled and profusely branching pro-cesses, all running along the ventrolateral surface (Fig.5A). Observations at higher magnification from horizontal(Fig. 5B) or coronal (Fig. 5C) sections revealed that theseplexes were made up of densely packed, thin (0.5–1.5 mm),medium-sized (2–3 mm), and large (4–7 mm) 5-HT-ir var-icosities. These 5-HT-ir varicosities were counted acrossthe depth of transverse sections (20 mm) at four differentlongitudinal levels of the spinal cord as a basis for com-parison with chronic transected animals (see below). As
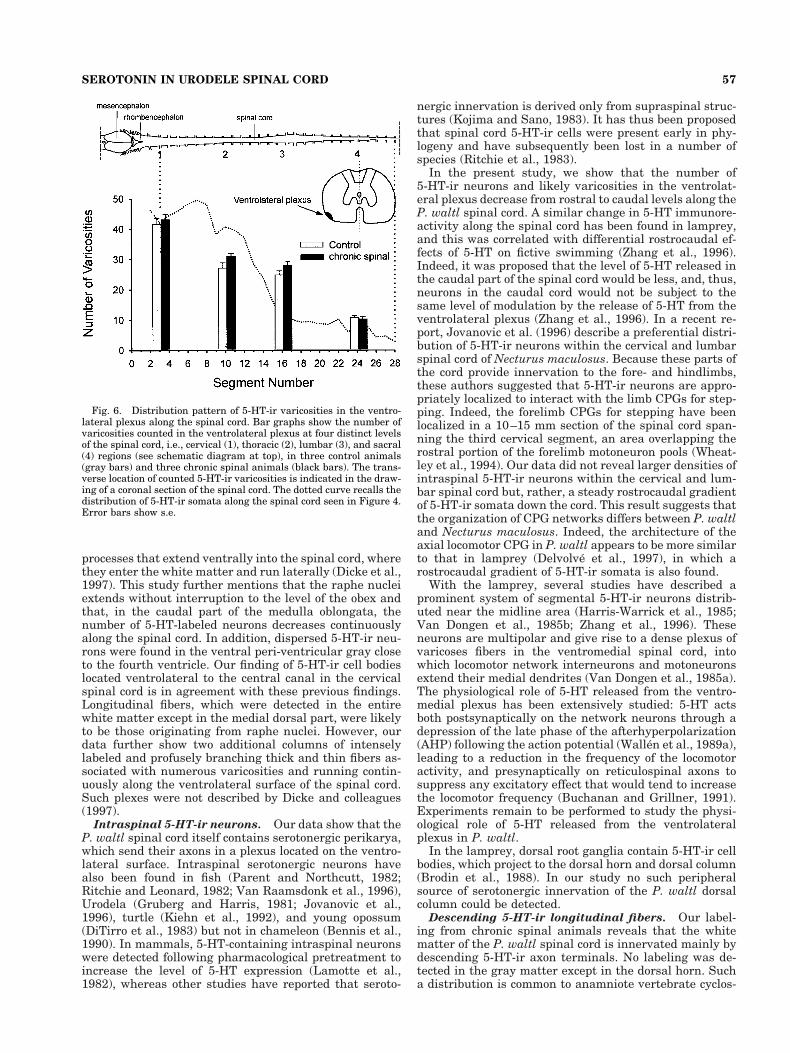
can be seen in Figure 6 (gray histogram bars), counts fromcontrol animals were 30–55 in the cervical part (S3–S4),15–35 in the thoracic (S10–S11) and lumbar (S16–S17)parts, and 5–15 at sacral level (S24–S25). Although ourregional sampling of segments does not allow conclusionsabout the continuous distribution of varicosities along thelength of the spinal cord, our counts at the four chosenlocations suggest that the number of plexus varicositiesdecreases progressively in a rostrocaudal direction. Inter-estingly, this rostrocaudal gradient in 5-HT-ir plexus var-icosities parallels the longitudinal decrease in 5-HT-ir so-mata (see dotted line in Fig. 6).
5-HT-ir fibers. Longitudinal intermixed 5-HT-ir fi-bers running in the lateral and dorsoventral plane wereobserved throughout the white matter of the spinal cord,except in a medial dorsal tract corresponding to the dorsalfuniculus (see Fig. 2A). The central gray region displayedvery few 5-HT-ir fibers, whereas the dorsal field of thegray matter was densely innervated with 5-HT-ir fibers.We did not find labeled fibers crossing the peri-ependymallining, which would have indicated a potential release sitefor 5-HT in the central canal. Horizontal sections revealedweakly varicose medium-sized fibers (0.4 mm), thin fiberswith large varicosities (2 mm), and very thin (visible onlyat 3400) varicose fibers running rectilinearly in the hori-zontal plane (see Fig. 5A). Longitudinal smooth (i.e., notvaricose) thin and medium-sized fibers running continu-ously in the horizontal plane were also detected. The me-dial ventral white matter displayed medium-sized fibersrunning horizontally and enriched with 1–5-mm-diametervaricosities.
5-HT immunoreactivity in chronicspinal animals
To determine the contribution of the brainstem to spinal5-HT innervation, the pattern of 5-HT immunoreactivity
Fig. 2. Microphotographs of 5-HT-ir distribution across theP. waltl spinal cord. A: Coronal section through the spinal cord of thetenth segment showing longitudinal 5-HT-ir processes in all fields ofthe spinal cord except for the central gray (G) and a region (arrow-heads) along the dorsal midline (m, dotted line). Note intense labelingon the ventrolateral surface corresponding to two ventrolateral plexes(vlp). Two 5-HT-ir somata located ventrolateral to the central canal(cc) are clearly distinguisable. The asterisk indicates a nonspecific
reaction product that stains the sulcus longitudinalis dorsalis that ispart of the venous plexus. The box outlines the location of the hori-zontal section illustrated in Figure 5A,B. B: Higher magnificationphotograph of the 5-HT-ir soma framed in A. Note that the cell bodylies ventrolateral to the peri-ependymal lining and sends convolutedprocesses (arrows) towards the ventral surface. Scale bars 5 50 mm inA; 20 mm in B.
53SEROTONIN IN URODELE SPINAL CORD

Figure 3
54 P. BRANCHEREAU ET AL.
within the spinal cord was analyzed in chronic spinalanimals (see Materials and Methods). The coronal sectionshown in Figure 7A illustrates the typical overall patternof immunoreactivity observed in animals spinalized 3weeks previously. There is a complete depletion of longi-tudinal 5-HT-ir fibers at all levels of the spinal cord. Incontrast, the bilateral ventrolateral plexes retained theirdistribution of 5-HT immunoreactivity, as did the seroto-nergic somata (Fig. 7A,C), whose localization remainedventrolateral to the central canal. As is shown in Figure7C, long processes migrated from these somata in theventral direction and extended along the ventral surfacebefore projecting into the ventrolateral plexus.
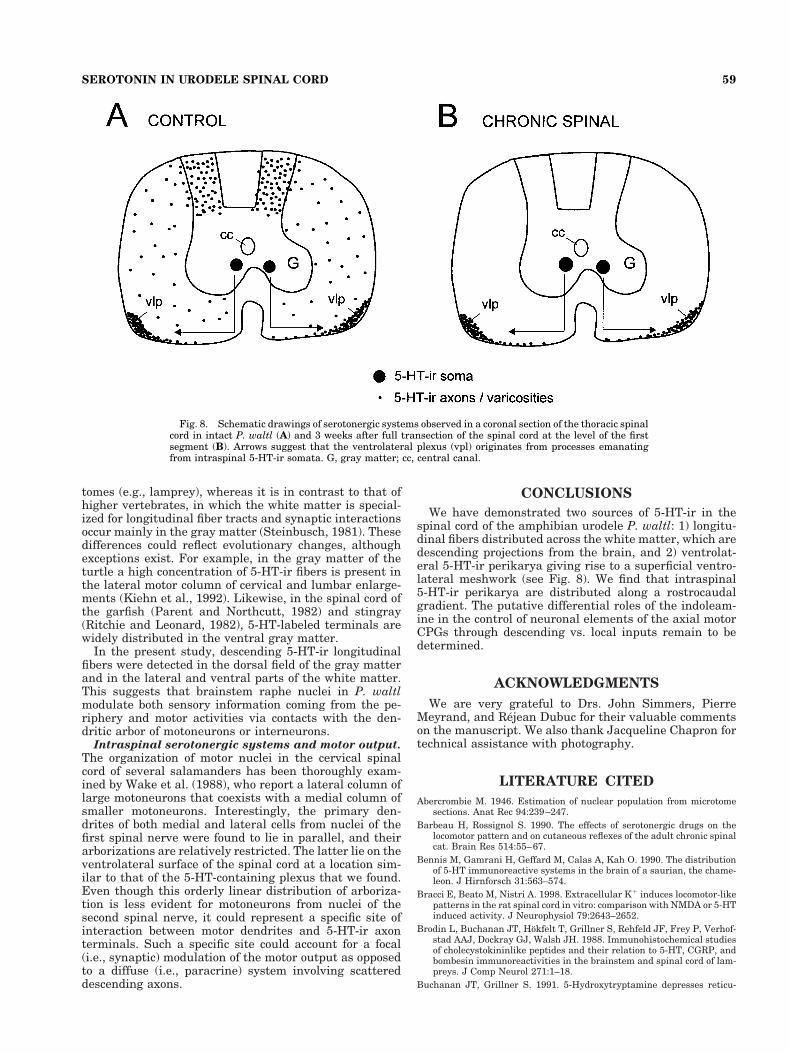
Numbers of 5-HT-containing neurons were thencounted at the levels of the brachial (S5 and S6) and crural(S15 and S18) plexes of the spinal cord in all three chronicspinal animals. The number of 5-HT-ir neurons was iden-tical to that observed in the spinal cord of unlesionedcontrol animals (Fig. 4), suggesting that those neuronsstem from an intraspinal serotonergic system. We alsostudied the distribution of 5-HT-ir varicosities in the ven-trolateral plexus from chronic spinal animals. The sameintense web-like geometry composed of thick and thinfibers associated, respectively, with thick and thin vari-cosities was detected (Fig. 7B). Finally, a quantification of5-HT-ir varicosities in the ventrolateral plexus was car-ried out at the four different representative levels (cervi-cal, thoracic, lumbar, and sacral) of the cord in chronicspinal animals. No statistical differences (Tukey test, P ,0.05) were found between the number of varicosities inchronic spinal animals and in control animals (compareblack and gray histogram bars in Fig. 6). The distributionof 5-HT immunoreactivity in an equivalent thoracic regionof the spinal cord before and 3 weeks after cervical spinal-ization is summarized in Figure 8. Only the 5-HT-ir ven-trolateral plexus and somata in the gray matter remain.
DISCUSSION
Physiological effect of 5-HT
Our preliminary electrophysiological results show thatexogenous 5-HT significantly increases both burst andcycle duration of fictive NMDA-induced axial locomotorpatterns in the in vitro brainstem and spinal cord prepa-ration of P. waltl. The type of 5-HT modulation we ob-served is similar to that previously reported for axiallocomotor CPGs in lamprey (Harris-Warrick and Cohen,
1985; Christenson et al., 1989; Wallen et al., 1989b; Mat-shushima and Grillner, 1992) and early larvae of Xenopuslaevis (Sillar et al., 1992) and Rana temporaria tadpoles(Woolston et al., 1994). Similar 5-HT modulating effectshave been observed in vitro on NMDA-activated limb lo-comotor CPGs in neonatal rat (Sqalli-Houssaini et al.,1993) and mudpuppy (Jovanovic et al., 1996). In contrast,5-HT applied alone on limb CPGs in neonatal rat (Caza-lets et al., 1992) causes an acceleration of fictive locomotoractivity.
Furthermore, unlike the case in rabbit (Viala andBuser, 1969) and neonatal rat (Cazalets et al., 1992; Cow-ley and Schmidt, 1994; Kiehn and Kjaerulff, 1996) prepa-rations, in our experiments 5-HT applied alone failed toinduce fictive locomotor activity. This suggests that, inUrodeles, 5-HT is probably involved in modulating ongo-ing locomotor activity rather than in activating the CPGitself (Jovanovic et al., 1996). These differences in theability of 5-HT to elicit a locomotor pattern may thereforereflect interspecies differences, although methodologicaland animal developmental stage differences cannot beruled out (see Cazalets et al., 1992; Jovanovic et al., 1996).
Fig. 4. Distribution pattern of 5-HT-ir cell bodies along the spinalcord. Numbers of 5-HT-containing perikarya were counted in seg-ments S2 to S28 in control animals and in four different segments inchronic spinal animals. The drawing above indicates correspondingsegment location along the spinal cord. Note that the number of5-HT-ir cells in the examined segments of chronic spinal animals isnot significantly different from that in control animals. The trans-verse location of counted 5-HT-ir neurons is indicated in the lowerdrawing of a coronal section of the spinal cord.
Fig. 3. Horizontal sections at the level of the ventral part of thegray matter showing 5-HT-ir neurons. A: Section at the level of S12showing bipolar oval type I neurons located lateral to the centralcanal (cc) and exhibiting tortuous processes that project laterally(arrows). Note the close apposition of two labeled somata (soma 1 and2) and longitudinal fibers (f) running in the vicinity of other 5-HT-irsomata. B: Section at the level of S26 showing a multipolar type Ineuron with a large process oriented in the rostrocaudal direction(open arrows) and another process projecting more laterally (curvedarrows). Double arrows indicate bifurcations, and the arrowheadpoints to a bouton of a labeled longitudinal fiber (f) that is in closeapposition with the lateral process of the soma-stained neuron.C: Three type I neurons at the level of S10. Filled arrowheads indicateappositions between 5-HT-ir somata and labeled axons (f). D: A typeII neuron at the level of S12, with its axon (ax) and dendrite (den) inapposition with several 5-HT-ir boutons (arrowheads) originatingfrom a second labeled axon (f). Dotted line indicates midline (m). cc,central canal; G, gray matter. Scale bars 5 50 mm in A,B; 20 mm inC,D.
55SEROTONIN IN URODELE SPINAL CORD
Immunohistochemistry
Two serotonergic systems in the spinal cord. Mostanatomical studies performed on amphibian species havefocused on the distribution of 5-HT immunoreactivity in
the brain without dealing with the spinal cord (Dube andParent, 1982; Kah and Chambolle, 1983; Fasolo et al.,1986; Corio et al., 1992; Clairambault et al., 1994; Dicke etal., 1997). A previous study reported that, in P. waltl,neurons of the raphe nuclei are the source of 5-HT-ir
Fig. 5. Microphotographs depicting the 5-HT-ir ventrolateralplexus (vlp). A: Horizontal section through S12 of the vlp (see Fig. 2Afor the location of the histological section). A column of intenselylabeled and profusely branching processes runs continuously alongthe ventrolateral surface. The course of some longitudinal fibers (ar-rows) can also be followed in the ventral white matter. B: Highermagnification photograph of the vlp (horizontal section at S10). Large
varicosities (open arrows) are intermingled with thin (solid arrows)and medium-sized (arrowheads) varicosities and may coexist on thesame 5-HT-ir fiber. C: High magnification photograph of a frontalsection through S12 showing endowed with thin varicosities (solidarrows), medium-sized (arrowheads), and large (open arrow) varicos-ities. Dorsal (D) and lateral (L) directions are indicated by arrows inthe upper right corner. Scale bars 5 50 mm in A; 10 mm in B,C.
56 P. BRANCHEREAU ET AL.
processes that extend ventrally into the spinal cord, wherethey enter the white matter and run laterally (Dicke et al.,1997). This study further mentions that the raphe nucleiextends without interruption to the level of the obex andthat, in the caudal part of the medulla oblongata, thenumber of 5-HT-labeled neurons decreases continuouslyalong the spinal cord. In addition, dispersed 5-HT-ir neu-rons were found in the ventral peri-ventricular gray closeto the fourth ventricle. Our finding of 5-HT-ir cell bodieslocated ventrolateral to the central canal in the cervicalspinal cord is in agreement with these previous findings.Longitudinal fibers, which were detected in the entirewhite matter except in the medial dorsal part, were likelyto be those originating from raphe nuclei. However, ourdata further show two additional columns of intenselylabeled and profusely branching thick and thin fibers as-sociated with numerous varicosities and running contin-uously along the ventrolateral surface of the spinal cord.Such plexes were not described by Dicke and colleagues(1997).
Intraspinal 5-HT-ir neurons. Our data show that theP. waltl spinal cord itself contains serotonergic perikarya,which send their axons in a plexus located on the ventro-lateral surface. Intraspinal serotonergic neurons havealso been found in fish (Parent and Northcutt, 1982;Ritchie and Leonard, 1982; Van Raamsdonk et al., 1996),Urodela (Gruberg and Harris, 1981; Jovanovic et al.,1996), turtle (Kiehn et al., 1992), and young opossum(DiTirro et al., 1983) but not in chameleon (Bennis et al.,1990). In mammals, 5-HT-containing intraspinal neuronswere detected following pharmacological pretreatment toincrease the level of 5-HT expression (Lamotte et al.,1982), whereas other studies have reported that seroto-
nergic innervation is derived only from supraspinal struc-tures (Kojima and Sano, 1983). It has thus been proposedthat spinal cord 5-HT-ir cells were present early in phy-logeny and have subsequently been lost in a number ofspecies (Ritchie et al., 1983).
In the present study, we show that the number of5-HT-ir neurons and likely varicosities in the ventrolat-eral plexus decrease from rostral to caudal levels along theP. waltl spinal cord. A similar change in 5-HT immunore-activity along the spinal cord has been found in lamprey,and this was correlated with differential rostrocaudal ef-fects of 5-HT on fictive swimming (Zhang et al., 1996).Indeed, it was proposed that the level of 5-HT released inthe caudal part of the spinal cord would be less, and, thus,neurons in the caudal cord would not be subject to thesame level of modulation by the release of 5-HT from theventrolateral plexus (Zhang et al., 1996). In a recent re-port, Jovanovic et al. (1996) describe a preferential distri-bution of 5-HT-ir neurons within the cervical and lumbarspinal cord of Necturus maculosus. Because these parts ofthe cord provide innervation to the fore- and hindlimbs,these authors suggested that 5-HT-ir neurons are appro-priately localized to interact with the limb CPGs for step-ping. Indeed, the forelimb CPGs for stepping have beenlocalized in a 10–15 mm section of the spinal cord span-ning the third cervical segment, an area overlapping therostral portion of the forelimb motoneuron pools (Wheat-ley et al., 1994). Our data did not reveal larger densities ofintraspinal 5-HT-ir neurons within the cervical and lum-bar spinal cord but, rather, a steady rostrocaudal gradientof 5-HT-ir somata down the cord. This result suggests thatthe organization of CPG networks differs between P. waltland Necturus maculosus. Indeed, the architecture of theaxial locomotor CPG in P. waltl appears to be more similarto that in lamprey (Delvolve et al., 1997), in which arostrocaudal gradient of 5-HT-ir somata is also found.
With the lamprey, several studies have described aprominent system of segmental 5-HT-ir neurons distrib-uted near the midline area (Harris-Warrick et al., 1985;Van Dongen et al., 1985b; Zhang et al., 1996). Theseneurons are multipolar and give rise to a dense plexus ofvaricoses fibers in the ventromedial spinal cord, intowhich locomotor network interneurons and motoneuronsextend their medial dendrites (Van Dongen et al., 1985a).The physiological role of 5-HT released from the ventro-medial plexus has been extensively studied: 5-HT actsboth postsynaptically on the network neurons through adepression of the late phase of the afterhyperpolarization(AHP) following the action potential (Wallen et al., 1989a),leading to a reduction in the frequency of the locomotoractivity, and presynaptically on reticulospinal axons tosuppress any excitatory effect that would tend to increasethe locomotor frequency (Buchanan and Grillner, 1991).Experiments remain to be performed to study the physi-ological role of 5-HT released from the ventrolateralplexus in P. waltl.
In the lamprey, dorsal root ganglia contain 5-HT-ir cellbodies, which project to the dorsal horn and dorsal column(Brodin et al., 1988). In our study no such peripheralsource of serotonergic innervation of the P. waltl dorsalcolumn could be detected.
Descending 5-HT-ir longitudinal fibers. Our label-ing from chronic spinal animals reveals that the whitematter of the P. waltl spinal cord is innervated mainly bydescending 5-HT-ir axon terminals. No labeling was de-tected in the gray matter except in the dorsal horn. Sucha distribution is common to anamniote vertebrate cyclos-
Fig. 6. Distribution pattern of 5-HT-ir varicosities in the ventro-lateral plexus along the spinal cord. Bar graphs show the number ofvaricosities counted in the ventrolateral plexus at four distinct levelsof the spinal cord, i.e., cervical (1), thoracic (2), lumbar (3), and sacral(4) regions (see schematic diagram at top), in three control animals(gray bars) and three chronic spinal animals (black bars). The trans-verse location of counted 5-HT-ir varicosities is indicated in the draw-ing of a coronal section of the spinal cord. The dotted curve recalls thedistribution of 5-HT-ir somata along the spinal cord seen in Figure 4.Error bars show s.e.
57SEROTONIN IN URODELE SPINAL CORD
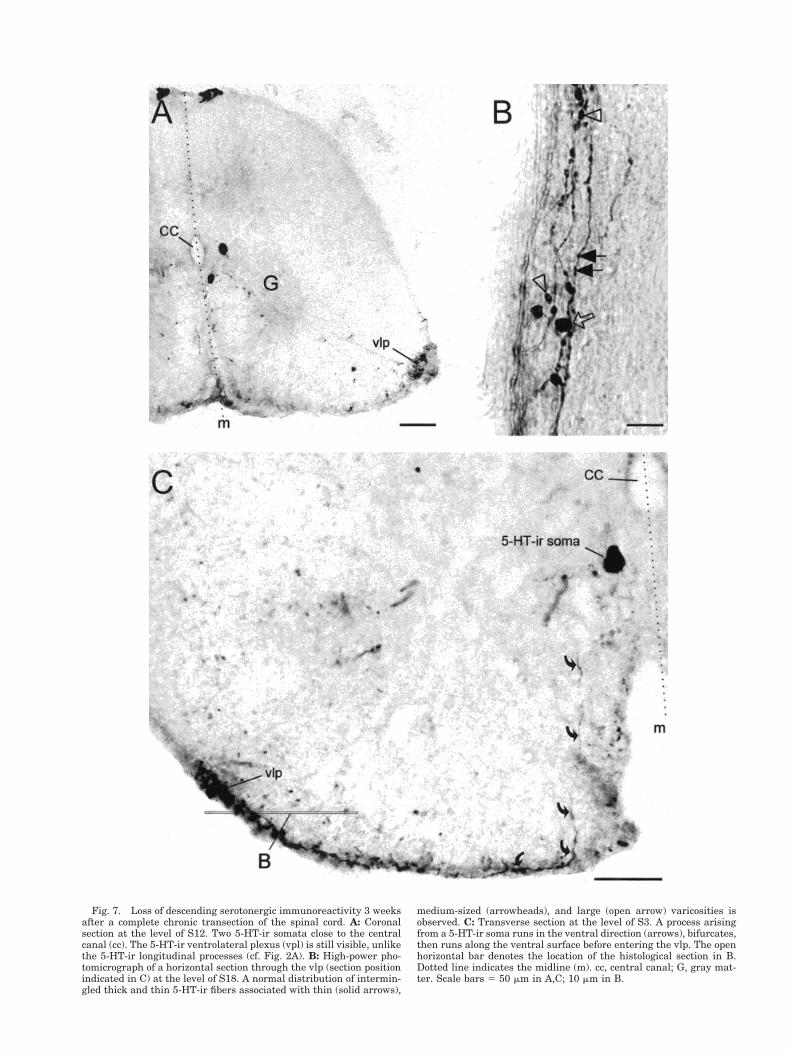
Fig. 7. Loss of descending serotonergic immunoreactivity 3 weeksafter a complete chronic transection of the spinal cord. A: Coronalsection at the level of S12. Two 5-HT-ir somata close to the centralcanal (cc). The 5-HT-ir ventrolateral plexus (vpl) is still visible, unlikethe 5-HT-ir longitudinal processes (cf. Fig. 2A). B: High-power pho-tomicrograph of a horizontal section through the vlp (section positionindicated in C) at the level of S18. A normal distribution of intermin-gled thick and thin 5-HT-ir fibers associated with thin (solid arrows),
medium-sized (arrowheads), and large (open arrow) varicosities isobserved. C: Transverse section at the level of S3. A process arisingfrom a 5-HT-ir soma runs in the ventral direction (arrows), bifurcates,then runs along the ventral surface before entering the vlp. The openhorizontal bar denotes the location of the histological section in B.Dotted line indicates the midline (m). cc, central canal; G, gray mat-ter. Scale bars 5 50 mm in A,C; 10 mm in B.
tomes (e.g., lamprey), whereas it is in contrast to that ofhigher vertebrates, in which the white matter is special-ized for longitudinal fiber tracts and synaptic interactionsoccur mainly in the gray matter (Steinbusch, 1981). Thesedifferences could reflect evolutionary changes, althoughexceptions exist. For example, in the gray matter of theturtle a high concentration of 5-HT-ir fibers is present inthe lateral motor column of cervical and lumbar enlarge-ments (Kiehn et al., 1992). Likewise, in the spinal cord ofthe garfish (Parent and Northcutt, 1982) and stingray(Ritchie and Leonard, 1982), 5-HT-labeled terminals arewidely distributed in the ventral gray matter.
In the present study, descending 5-HT-ir longitudinalfibers were detected in the dorsal field of the gray matterand in the lateral and ventral parts of the white matter.This suggests that brainstem raphe nuclei in P. waltlmodulate both sensory information coming from the pe-riphery and motor activities via contacts with the den-dritic arbor of motoneurons or interneurons.
Intraspinal serotonergic systems and motor output.
The organization of motor nuclei in the cervical spinalcord of several salamanders has been thoroughly exam-ined by Wake et al. (1988), who report a lateral column oflarge motoneurons that coexists with a medial column ofsmaller motoneurons. Interestingly, the primary den-drites of both medial and lateral cells from nuclei of thefirst spinal nerve were found to lie in parallel, and theirarborizations are relatively restricted. The latter lie on theventrolateral surface of the spinal cord at a location sim-ilar to that of the 5-HT-containing plexus that we found.Even though this orderly linear distribution of arboriza-tion is less evident for motoneurons from nuclei of thesecond spinal nerve, it could represent a specific site ofinteraction between motor dendrites and 5-HT-ir axonterminals. Such a specific site could account for a focal(i.e., synaptic) modulation of the motor output as opposedto a diffuse (i.e., paracrine) system involving scattereddescending axons.
CONCLUSIONS
We have demonstrated two sources of 5-HT-ir in thespinal cord of the amphibian urodele P. waltl: 1) longitu-dinal fibers distributed across the white matter, which aredescending projections from the brain, and 2) ventrolat-eral 5-HT-ir perikarya giving rise to a superficial ventro-lateral meshwork (see Fig. 8). We find that intraspinal5-HT-ir perikarya are distributed along a rostrocaudalgradient. The putative differential roles of the indoleam-ine in the control of neuronal elements of the axial motorCPGs through descending vs. local inputs remain to bedetermined.
ACKNOWLEDGMENTS
We are very grateful to Drs. John Simmers, PierreMeyrand, and Rejean Dubuc for their valuable commentson the manuscript. We also thank Jacqueline Chapron fortechnical assistance with photography.
LITERATURE CITED
Abercrombie M. 1946. Estimation of nuclear population from microtomesections. Anat Rec 94:239–247.
Barbeau H, Rossignol S. 1990. The effects of serotonergic drugs on thelocomotor pattern and on cutaneous reflexes of the adult chronic spinalcat. Brain Res 514:55–67.
Bennis M, Gamrani H, Geffard M, Calas A, Kah O. 1990. The distributionof 5-HT immunoreactive systems in the brain of a saurian, the chame-leon. J Hirnforsch 31:563–574.
Bracci E, Beato M, Nistri A. 1998. Extracellular K1 induces locomotor-likepatterns in the rat spinal cord in vitro: comparison with NMDA or 5-HTinduced activity. J Neurophysiol 79:2643–2652.
Brodin L, Buchanan JT, Hokfelt T, Grillner S, Rehfeld JF, Frey P, Verhof-stad AAJ, Dockray GJ, Walsh JH. 1988. Immunohistochemical studiesof cholecystokininlike peptides and their relation to 5-HT, CGRP, andbombesin immunoreactivities in the brainstem and spinal cord of lam-preys. J Comp Neurol 271:1–18.
Buchanan JT, Grillner S. 1991. 5-Hydroxytryptamine depresses reticu-
Fig. 8. Schematic drawings of serotonergic systems observed in a coronal section of the thoracic spinalcord in intact P. waltl (A) and 3 weeks after full transection of the spinal cord at the level of the firstsegment (B). Arrows suggest that the ventrolateral plexus (vpl) originates from processes emanatingfrom intraspinal 5-HT-ir somata. G, gray matter; cc, central canal.
59SEROTONIN IN URODELE SPINAL CORD

lospinal excitatory postsynaptic potentials in motoneurons of the lam-prey. Neurosci Lett 112:71–74.
Cazalets J-R, Sqalli-Houssaini Y, Clarac F. 1992. Activation of the centralpattern generators for locomotion by serotonin and excitatory aminoacids in neonatal rat. J Physiol (London) 455:187–204.
Christenson J, Franck J, Grillner S. 1989. Increase in endogenous5-hydroxytryptamine levels modulates the central network underlyinglocomotion in the lamprey spinal cord. Neurosci Lett 100:188–192.
Clairambault P, Christophe N, Pairault C, Herbin M, Ward R, Reperant J.1994. Organization of the serotoninergic system in the brain of two am-phibian species, Ambystoma mexicanum (Urodela) and Typhlonectescompressicauda (Gymnophiona). Anat Embryol 190:87–99.
Coggeshall RE, Lekan HA. 1996. Methods for determining numbers of cellsand synapses: a case for more uniform standards of reviews. J CompNeurol 364:6–15.
Corio M, Thibault J, Peute J. 1992. Distribution of catecholaminergic andserotoninergic systems in forebrain and midbrain of the newt, Triturusalpestris (Urodela). Cell Tissue Res 268:377–387.
Cowley KC, Schmidt BJ. 1994. A comparison of motor patterns induced byN-methyl-D-aspartate, acetylcholine and serotonin in the in vitro neo-natal rat spinal cord. Neurosci Lett 171:147–150.
Davis BM, Duffy MT, Simpson J. 1989. Bulbospinal and intraspinal con-nections in normal and regenerated salamander spinal cord. Exp Neu-rol 103:41–51.
Delvolve I, Bem T, Cabelguen JM. 1997. Epaxial and limb muscle activityduring swimming and terrestrial stepping in the adult newt, Pleurode-les waltl. J Neurophysiol 78:638–650.
Delvolve I, Branchereau P, Dubuc R, Cabelguen J-M. 1999. Fictive rhyth-mic motor patterns induced by NMDA in an in vitro brain stem-spinalcord preparation from an adult urodele. J Neurophysiol 82:1074–1077.
Dicke U, Wallstein M, Roth G. 1997. 5-HT-like immunoreactivity in thebrains of plethodontid and salamandrid salamanders (Hydromantesitalicus, Hydromantes genei, Plethodon jordani, Desmognathusochrophaeus, Pleurodeles waltl): an immunohistochemical and biocytindouble-labelling study. Cell Tissue Res 287:513–523.
DiTirro FJ, Martin GF, Ho RH. 1983. A developmental study ofsubstance-P, somatostatin, enkephalin and serotonin immunoreactiveelements in the spinal cord of the North American opossum. J CompNeurol 213:241–261.
Dube L, Parent A. 1982. The organization of monoamine-containing neu-rons in the brain of the salamander, Necturus maculosus. J CompNeurol 211:21–30.
Fasolo A, Franzoni MF, Gaudino G, Steinbusch HW. 1986. The organiza-tion of serotonin-immunoreactive neuronal systems in the brain of thecrested newt, Triturus cristatus carnifex Laur. Cell Tissue Res 243:239–247.
Geffard M, Heinrich-Rock AM, Dulluc J, Seguela P. 1985. Antisera againstsmall neurotransmitter-like molecules. Neurochem Int 7:403–413.
Grillner S, Wallen P. 1985. Central pattern generators for locomotion, withspecial reference to vertebrates. Annu Rev Neurosci 18:233–261.
Gruberg ER, Harris WA. 1981. The serotonergic somatosensory projectionto the tectum of normal and eyeless salamander. J Morphol 170:55–69.
Harris-Warrick RM, Cohen AH. 1985. Serotonin modulates the centralpattern generator for locomotion in the isolated lamprey spinal cord. JExp Biol 116:27–46.
Harris-Warrick RM, McPhee JC, Filler JA. 1985. Distribution of seroto-nergic neurons and processes in the lamprey spinal cord. Neuroscience14:1127–1140.
Jovanovic K, Petrov T, Greer JJ, Stein RB. 1996. Serotonergic modulationof the mudpuppy (Necturus maculatus) locomotor pattern in vitro. ExpBrain Res 111:57–67.
Kah O, Chambolle P. 1983. Serotonin in the brain of the goldfish, Carassiusauratus. An immunocytochemical study. Cell Tissue Res 234:319–333.
Kiehn O, Kjaerulff O. 1996. Spatiotemporal charactristics of 5-HT anddopamine-induced rhythmic hindlimb activity in the in vitro neonatalrat. J Neurophysiol 75:1472–1482.
Kiehn O, Rostrup E, Moller M. 1992. Monoaminergic systems in the brain-stem and spinal cord of the turtle Pseudemys scripta elegans as re-vealed by antibodies against serotonin and tyrosine hydroxylase.J Comp Neurol 325:527–547.
Kojima M, Sano Y. 1983. The organization of serotonin fibers in the
anterior column of the mammalian spinal cord. An immunohistochem-ical study. Anat Embryol 167:1–11.
Lamotte CC, Johns DJ, deLanerolle NC. 1982. Immunohistochemical evi-dence of indoleamine neurons in monkey spinal cord. J Comp Neurol206:359–370.
Matshushima T, Grillner S. 1992. Local serotonergic modulation ofcalcium-dependent potassium channels controls intersegmental coor-dination in the lamprey spinal cord. J Neurophysiol 67:1683–1690.
Parent A, Northcutt RG. 1982. The monoamine-containing neurons in thebrain of the garfish, Lepisosteus osseus. Brain Res Bull 9:189–204.
Ritchie TC, Leonard RB. 1982. Immunocytochemical demonstration ofserotonergic cells, terminals and axons in the spinal cord of the stin-gray, Dasyatis sabina. Brain Res 240:334–337.
Ritchie TC, Livingston CA, Hughes MG, McAdoo DJ, Leonard RB. 1983.The distribution of serotonin in the CNS of an elasmobranch fish:immunocytochemical and biochemical studies in the atlantic stingray,Dasyatis sabina. J Comp Neurol 221:429–443.
Rossignol S, Dubuc R. 1994. Spinal pattern generation. Curr Opin Neuro-biol 4:894–902.
Sillar KT, Wedderburn JF, Simmers AJ. 1992. Modulation of swimmingrhythmicity by 5-hydroxytryptamine during post-embryonic develop-ment in Xenopus laevis. Proc R Soc London [Biol] 250:107–114.
Sqalli-Houssaini Y, Cazalets J-R, Clarac F. 1993. Oscillatory properties ofthe central pattern generator for locomotion in neonatal rats. J Neu-rophysiol 70:803–813.
Steinbusch HWM. 1981. Distribution of serotonin-immunoreactivity in thecentral nervous system of the rat–cell bodies and terminals. Neuro-science 6:557–618.
Van Dongen PAM, Hokfelt T, Grillner S, Verhofstad AJH, SteinbuschHWM. 1985a. Possible target neurons of 5-hydroxytryptamine fibers inthe lamprey spinal cord: immunochemistry combined with intracellu-lar staining with Lucifer yellow. J Comp Neurol 234:523–535.
Van Dongen PAM, Hokfelt T, Grillner S, Verhofstad AJH, SteinbuschHWM, Cuello AC, Terenius L. 1985b. Immunohistochemical demon-stration of some putative neurotransmitters in the lamprey spinalcord and spinal ganglia: 5-hydroxytryptamine-, tachykinin-, andneuropeptide-Y-immunoreactive neurons and fibers. J Comp Neurol234:501–522.
Van Raamsdonk W, Bosch TJ, Smit-Onel MJ, Maslam S. 1996. Organisa-tion of the zebrafish spinal cord: distribution of motoneuron dendritesand 5-HT containing cells. Eur J Morphol 34:65–77.
Viala D, Buser P. 1969. The effect of DOPA and 5-HTP on rhythmicefferent discharges in hind limb nerves in the rabbit. Brain Res 12:437–443.
Volicier L. 1969. Correlation between behavioral and biochemical effects ofp-chlorophenylalanine in mice and rats. Int J Neuropharmacol 8:361–364.
Wake DB, Nishikawa KC, Dicke U, Roth G. 1988. Organization of themotor nuclei in the cervical spinal cord of salamanders. J Comp Neurol278:195–208.
Wallen P, Buchanan JT, Grillner S, Hill RH, Christenson J, Hokfelt T.1989a. Effects of 5-hydroxytryptamine on the afterhyperpolarization,spike frequency regulation, and oscillatory membrane properties inlamprey spinal cord neurons. J Neurophysiol 61:759–768.
Wallen P, Christenson J, Brodin L, Hill R, Lansner A, Grillner S. 1989b.Mechanisms underlying the serotonergic modulation of the spinal cir-cuitry for locomotion in lamprey. Progr Brain Res 80:321–327 [discus-sion p 315–319].
Wheatley M, Stein RB. 1992. An in vitro preparation of the mudpuppy forsimultaneous intracellular and electromyographic recording during lo-comotion. J Neurosci Methods 42:129–137.
Wheatley M, Jovanovic K, Stein RB, Lawson V. 1994. The activity ofinterneurons during locomotion in the in vitro necturus spinal cord.J Neurophysiol 71:2025–2032.
Woolston AM, Wedderburn JF, Sillar KT. 1994. Descending serotonergicspinal projections and modulation of locomotor rhythmicity in Ranatemporaria embryos. Proc R Soc London [Biol] 255:73–79.
Zhang WQ, Pombal MA, ElManira A, Grillner S. 1996. Rostrocaudal dis-tribution of 5-HT innervation in the lamprey spinal cord and differen-tial effects of 5-HT on fictive locomotion. J Comp Neurol 374:278–290.
60 P. BRANCHEREAU ET AL.





























