Embed Size (px)
Citation preview

742 The American JournalofClinicalNutrition 30: MAY 1977, pp 742-757. Printed in U.S.A.
Short-term and long-term components in theregulation of food intake: evidence for amodulatory role of carbohydrate status13Theodore B. Van itallie, M. D. , Nicole Schupf Smith, Ph . D. , and
David Quartermain, Ph.D.
The traditional hypothalmus: a point of de.parture
Most models that attempt to explain how
food intake is regulated take as their pointof departure a series of classical observa-tions suggesting that there are at least tworelated systems in the brain concerned with
feeding behavior (1-4). One system isthought to be present in the ventromedialhypothalamus, and appears to function byexercising inhibitory control over a secondsite of activity located laterally (5). The yen-tromedial area has been said to contain a
“satiety system” because electrical, or ap-propniate pharmacological, stimulation of
this region can inhibit feeding in hungryanimals. Similarly, the lateral area has beenthought to embody a “feeding system” be-
cause direct stimulation of this locus can
impel apparently satiated animals to eat.Some of the functional characteristics ofthese two systems are summarized in Figure
In view of the configuration of this tradi-tional model, it is not surprising that thenotion evolved that food intake in manyanimals is principally regulated by satietyrather than hunger. In other words, such
animals are “programmed” to seek food,with food-seeking activity being interruptedby the intermittent periods of satiety thatoccur after meal ingestion. As the durationof deprivation and strength of drive for foodincrease, a stronger satiety signal is requiredto interrupt food-seeking behavior.
Because the eating process in meal-takinganimals slows and stops after a certain
amount of food has been ingested, it seemslogical to postulate that signals generated bythe presence of food in the stomach andupper small intestine are responsible for mi-
tial activation of the satiety system. Also, itcould be reasoned that subsequent signals
derived from absorbed food might reinforceand sustain satiety. Finally, the processesresponsible for the initiation and termina-tion of individual meals are, in turn, modi-fied by the nutritional state of the animal.Thus, in the regulation of food intake, thereappears to be both a short-term componentthat varies principally in relation to the na-ture and quantity of food consumed, and along-term component that is dependent
upon the nutritional status of the animal. Innormal animals, these two components in-teract reciprocally so as to maintain energy
balance and consequently a stable body
composition .
Short-term satiety signals
The short-term component is divisibleinto at least two phases, an early “preab-
sorptive” response to food ingestion and asomewhat delayed “absorptive” phase (Ta-ble 1). There is an early decrease in hungerthat occurs when food is introduced into thestomach . This effect was originally thoughtto take place before any significant chemicalchanges could occur in the blood; however,it now appears that gastric emptying and
I From the Department of Medicine and Institute of
Human Nutrition, Columbia University College of
Physicians and Surgeons and St. Luke’s Hospital Cen-
ter, and Department of Neurology, Milbank Research
La�boratories, New York University School of Mcdi-
cine, New York, New York.
2 Supported in part by Grants AM-17624, and AM-
17259 from the National Institutes of Health and the
Weight Watchers Foundation.
3 Address reprint requests to: Dr. Theodore B. Van
Itallie, St. Luke’s Hospital Center, Ar�isterdam Aye-
nue at 1 14th Street, New York, New York 10025.
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

pisDute ACTflITT
FOOD INTAKE REGULATION 743
intestinal absorption may begin within mm-
utes after food is ingested (6, 7). Accord-ingly, there is a clear temporal overlap be-tween the occurrence of afferent responsesassociated with the arrival of food in the
upper gastrointestinal tract, with release ofputative “satiety hormones” from the gut,and buildup in the circulation of the ab-sorbed products of digestion.
When food enters into and distends the
stomach, stretch receptors in the stomachwall are activated and thereby may contnib-ute to the satiety mosaic via the vagus nerve
(8). However, a more important determi-nant of the onset of satiety appears to beactiviation of the proximal small intestine.Studies on food-deprived rats with open gas-tric fistulas have demonstrated that wheningested food fails to enter the small intes-tine, satiety does not occur (9). Smith et al.(9) believe that such observations “ . .
appear to exclude taste, other oral stimuli,pharyngeal and esophageal movements, andthe contact of food with the gastric mucosa
A naamouai. ia�L 1St T �aAL �ua�s. or e�nmc
�Al� $AT�TT � � �
I.�tbLttas
(5) ,#{174}:::::::::x:�
(2) � �#{163}b..�. at Uttaty St�1ipasal 5.sto.
(a) ah�iaa5* GTG. �m 4.(5) �,taa1: alstrolysl, etc.LecaS ta).ctts .1 atpha-.�et.t�
(3) #{174}it��;t�;t �
ch�.t*�1atte.
(4) #{174} &�Vocal 1..t... (.lattrtty.Ls. #{149}tc.)
Easel tajeettos of bsta..asslet(-tee...’)
FIG. 1 . A traditional model for the neural control of
feeding. Activation of the satiety system (condition 1)
causes inhibition of the feeding system, with conse-
quent suppression of food intake. Conversely,inactiva-tion of the satiety system (condition 2) disinhibits the
feeding system and thereby induces feeding. When the
feeding system is directly activated (condition 3) or
inactivated (condition 4), feeding activity is, respec-
tively, evoked or terminated.
as potent stimuli for satiety. The results
also seem to exclude the possibility that theend of a test meal (is) determined byhabit . . . .“
In contrast to the absence of satiety in ratswhen ingested food is permitted to drain outthrough an open gastric fistula is the “intes-tinal satiety” that occurs when small vol-umes of liquid food are infused directly intothe duodenum (9). Entrance of food intothe duodenum may stimulate release intothe circulation of intestinal hormones suchas secretin , enterogastrone , ‘ ‘glucagon,”and cholecystokinin (CCK). Recently, An-tin and Smith (10) reported that in rats, theparenteral administration of CCK in ratherlarge doses elicits a behavioral responsewhich is indistinguishable from that shownby the same animals at the end of a normal
meal. This observation lends support to theview that, directly or indirectly, CCK maybe an important satiety signal. In like man-
ner enterogastrone has been reported to in-
hibit food intake in fasted mice (11),whereas secretin and gastnin have beenfound relatively ineffective as satiety-induc-ing agents (9). The duodenum also contains
osmoreceptors that inhibit gastric emptyingwhen they are exposed to hypertonic solu-tions (12). This anatomical fact is consistentwith the substantial series of observations
showing that a bolus of food can be tempo-rarily satiating in proportion to its tonicity
(13-15).During the absorptive state, “postinges-
tive” signals generated by absorbed nutni-
ents are thought to augment and reinforceafferent responses elicited by the presenceof food in the gut. As used in this context,postingestive signals are those generated di-rectly or indirectly by calorigenic substratesabsorbed from the digestive tract. It is, ofcourse, impossible at this time to explainexactly how such food derived signals areperceived and acted upon by putative recep-tors in the satiety system; however, asBooth (6) has suggested, the functionalstimulus need not reflect an absolute quan-tity but could be the speed at which energyis supplied.
Unfortunately, the available evidenceabout the role of postingestive signals inthe satiety process remains confusing and
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

TABLE 1Possible short-term satiety signals
Phase” Food�derived stimulus Peripheral transducers Satiety signalt
Ingestive (pre- Gastric distention Intragastric stretch receptors Neural (vagus)
absorptive)Tonicity of gastric-duo-
denal contents
Substrates entering
duodenum
Duodenal osmoreceptors
Gut chemoreceptors
Neural (vagus) (inhibition of
gastric motility and empty-
ing)
Gut secretion into circulation
of cholecystokinin and other
“satiety hormones”
Postingestive Portal circulation(absorptive)
GlucoseAmino acids
Glucose
Glucose
Amino acids
/3-Cells of pancreas
Hepatic-vagal glucoreceptors
Rate of hepatic metabolism of
food-derived substrate
Hepatic thermoreceptors
Insulinemia
Neural (vagus)
Neural or “liver-derived hu-
moral signals”
Systemic circulation Tonicity of blood
Temperature of blood
(Thermic response to food)
Glucosemia
Aminoacidemia
Chylomicronemia
Insulinemia“Other humoral satiety sig-
nals”Hormone ratios (e .g. , insulin/
glucagon)
�-a The ingestive and postingestive phases are sequential but may overlap temporally. b Some “postingestive”satiety signals (i .e ., the thermic response to food and the concentrations in plasma of various hormones) may also
serve as “long-term” signals reflecting nutritional status.
744 VAN ITALLIE ET AL.
contradictory. On the one hand, numer-
ous studies have been reported in support ofthe notion that a postprandial increase inplasma amino acids (16-18) or glucose (19-23) can induce or prolong satiety. In thisregard, it may not be necessary for a signal
to build up rapidly in the systemic circula-tion . Increased activity has been reported inthe vagus nerve in response to a rise in
glucose concentration in portal venousblood (24). Russek (25) has described in-duction of “anorexia” in fasted dogs when
relatively small quantities of glucose are ad-ministered intraportally . This observationsuggests that glucoreceptors in the liver mayindirectly promote satiety . On the otherhand, induction of systemic hyperglycemiaby means of glucose infusion does not read-
ily elicit satiety (26), although parenteraladministration of various amino acid mix-tunes has been reported to cause anorexia(27).
Recently, Koopmans (28, 29) has devisedan ingenious experimental model that casts
further doubt upon the importance of post-
ingestive signals in short-term satiety. Heprepared pairs of parabiotic rats in which a
1 2-inch segment of each rat’s intestine wasexchanged for a 12-inch segment of its part-ner’s intestine . In this preparation , food ea-
ten by one rat passes through the first 21/2
inches of his own intestine to the intestine ofhis partner. After 1 2 inches of intestinaltransit in the second rat, which embodiesthe distal half of the duodenum and much ofthe jejunum, it returns to the intestine of the
rat that originally took the food . Using thismodel, Koopmans studied the effect offeeding one of the pair on the spontaneousintake of the other offered food 30 or 60mm later. He found that after both of theseintervals the second rat ate slightly more
and certainly not less than the rat which wasfed first. Thus, the segment of the intestinein the second rat that received the food
eaten by the first rat presumably did notrelease satiety-producing signals. It is alsonoteworthy that, when one rat was fed la-
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

FOOD INTAKE REGULATION 745
beled glucose or amino acids, the subse-quent rise in the radioactivity in the blood ofthe two animals was prompt and both curveswere very similar. Behavioral tests showedthat, at 30 and 60 mm, when approximately
equal amounts of glucose and amino acidhad been absorbed into the second intes-tines-crossed rat, one was completely sa-
tiated whereas the other was still hungry.
Thus, satiety appeared to occur independ-ently of changes in the blood levels of glu-cose and/or amino acids.
Subsequent longer-term experiments con-ducted by Koopmans showed that the intes-
tines-crossed rats increase their food intakein response to nutritive dilution but do notadjust when only one rat of the pair is al-lowed to feed over a 5-day period, meaning
that each rat obtains only about one-half asmuch food as usual. After 5 days, when
substantial weight loss has occurred , the ratthat is being fed will begin to adapt by in-creasing intake, suggesting the emergence
of a long-term regulatory response . Takentogether with the observations of Smith andassociates (9), these results suggest that the
intact stomach and/or first half of the duo-denum play a critical role in initiating short-term satiety. However, it remains unclearwhether the products of digestion, once ab-
sorbed, contribute in a significant way to the
onset of satiety. It seems more likely thatthe usual role of absorbed nutrients is to
sustain satiety, perhaps in proportion to
their potential energy content.
Long-term component
Although it is relatively easy to visualize afirst order feedback system which seeks,finds, and ingests food, shutting itself off for
a time when a certain rate of energy inflowhas been achieved, it is more difficult tounderstand how such a system can also con-
rect for previously imposed deficits and sun-feits. Nevertheless, embodied in energyhomeostasis must be a long-term compo-nent that, over a period of days or weeks,gradually adjusts food intake (and perhaps
energy output) so as to restore and thenmaintain body fat content within a fairlynarrow “preferred” range.
The evidence that most nonobese animalsdefend a characteristic body composition
seems quite firm. Physically active youngmen and women tend to maintain a rela-tively constant weight despite rather widevariations in food intake and energy ex-
penditure (30-32). When their food is di-luted with nonabsorbable carbohydrate , ratsrespond by increasing volume intake and
thereby maintain a constant weight (33).
Lean young men adapt in like manner to
covert nutritive dilution (34). Animalsmade obese by force feeding spontaneously
reduce energy intake and return to their
previous weight when permitted ad libitumfeeding (35, 36). Analogous studies of hu-
man subjects have yielded somewhat similarresults (37). After periods of semistarva-tion, both laboratory animals (36) and hu-man volunteers (38) gradually regain lost
weight by restricting energy expenditureand consuming sufficient extra calories to
replenish energy stores. However, in viewof the prevalence of obesity in affluent soci-
eties, it is evident that human subjects aremore effective in defending calorie storesagainst depletion than against excess.
During the past quarter century, many
theories have been advanced to explain howthe body can estimate the quantity of tn-glyceride in the adipose tissue stores and
thereby regulate depot fat content. Becausesome adipocytes appear to be directly inner-vated (39, 40), it is conceivable that theirstatus can be directly monitored by the cen-
tral nervous system . However, most work-ens in the field seem to have been moreattracted by the possibility that the adiposetissue might generate a signal or signalstransmitted via the circulation that is pro-pontional to the quantity of stored triglycer-ide (41, 42).
In recent years, this latter approach to theproblem has received support from observa-
tions indicating that the rate of release fromadipocytes of free fatty acids (FFA) andglycerol is, in part, a function of the quantityof triglyceride stoned within the cell (43,44). The mechanism responsible for thisphenomenon is not clearly understood;however, there is evidence that as adipo-cytes are increasingly distended with fatthey become more insulin-resistant (45).Associated with the insulin resistance ofobesity is a varying degree of hypennsulin-
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

746 VAN ITALLIE ET AL.
ism, manifested by elevated basal and stim-ulated concentrations of immunoreactive in-sulin in the circulation (46).
If it is assumed for the purposes of discus-sion that some consistent proportionality ex-
ists between circulating levels of FFA andglycerol on the one hand, and degree of
adipocyte hypentrophy on the other, theremay be a basis for regarding either of these
moieties as the signal that informs a hypo-thalamic “ponderostat” of the status of thebody’s triglyceride reserves. However, therewould still have to be a receptor system inthe brain capable of responding directly orindirectly to these particular signals. Someof the preliminary evidence in support of theexistence of appropriate receptor neuronswill be discussed subsequently. In any case,it is difficult to imagine how these substrates
by themselves could serve as reliable indicesof depot fat status, because elevated plasmaconcentrations of FFA and glycerol are con-comitants of both fasting (deprivation) andof obesity. If these moieties play any role inlong-term regulation of food intake, it mustbe as a part of a multifactorial system thatincludes other signals which, in combina-tion, can faithfully reflect nutritional status.
In a quite different approach to the prob-lem of long-term regulation of caloriestores, Hervey (47) has proposed that the
hypothalamus might monitor depot fat bymeasuring the concentration in blood of asteroid that is partitioned in a fixed ratiobetween the lipid and aqueous phases of thebody’s constituents. Hervey’s dilution the-ory is ingenious, but, unfortunately, lacks
experimental verification. Recently, Baileet al. (48) have suggested that one or moreprostaglandins may signal the status of thefat stores to regulatory receptors in the hy-
pothalamus. They have hypothesized thatthe production of prostaglandin occurs inadipose tissue at a rate related to the level ofadiposity . Prostaglandins are believed toleak out of adipocytes, and it is possible thatsome fraction may escape degradation in the
lungs and reach the brain . Baile and associ-ates postulate that the arterial prostaglandinlevel may vary with the degree of adiposityand thereby modulate hypothalamic regula-tion of energy stores. In support of thishypothesis, they have shown that subcuta-
neous injections of prostaglandin E1 (PGE1)can reduce the food intake of rats. More-over, when injected directly into the lateralhypothalamus of the rat, 1 �tg of PGE1 alsocauses a reduction of food intake.
Finally, it is conceivable that within the
hypothalamic system concerned with theregulation of calorie stores, there might ex-ist an entity that serves as a microcosm ofthe animal’s nutritional status. During de-pletion of depot fat this entity might experi-ence and signal a parallel depletion . Somesupport for the possible existence of such anarrangement is to be found in the studies ofPfaff (49), who has reported that the nu-cleoli in ventromedial hypothalamic neu-rons in the brains of well-fed rats are signifi-cantly larger than those in underfed ani-mals. Powley (50) has described morpho-logical changes in the ventromedial nuclei ofZucker obese (fa/fa) rats, whereas Nicholai-dis (5 1) has made preliminary observationsshowing a positive correlation between theadiposity of the rat and the lipid content ofits hypothalamus . The relative excess of hy-pothalamic lipid in obese rats appears to beaccounted for by extra triglycerides and cho-lesterol rather than by structural phospho-
lipids or glycolipids . It is also pertinent thatosmosensitive cells which participate in themechanisms by which the hypothalamusregulates water intake appear to serve asmicrocosms of the body’s state of hydration(52, 53). But perhaps the best argument in
favor of a microcosmal theory of long-termregulation of energy stores is the lack of firmevidence, to date, for a reliable communica-tions system between the peripheral fat de-pot and an appropriate central nervous sys-tem monitor.
Integration of short#{149}term and long-termcomponents
In order to have a coherent framework for
discussion about the control of energy in-take, we have assembled an elementarymodel that attempts to account, with someparsimony, for both short-term and long-term regulation. This model (Fig. 2) ac-knowledges that proximate signals derived
from food ingestion activate the satiety sys-tem. However, to explain long-term regula-tion, the model proposes that the quantity
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

FOOD INTAKE REGULATION 747
FIG. 2. A simplified model of long-term regulation of energy intake. Short-term satiety signals are generated by
food in the gastrointestinal tract (via gastric stretch receptors, duodenal osmoreceptors, gut “satiety hormones,”
etc.) and by circulating metabolites derived from digested food . The response of the satiety system to these signals
is modulated by corrective signals reflecting the status of the body’s energy stores. Thus, during depletion, a
stronger signal is required for satiety, leading to increased energy intake. When stores are replenished or
overfilled, a weaker signal can induce satiety and intake is reduced accordingly. (In an alternate theory, Brobeck
(68) proposed that “ . . . adipose tissue [hasi a predominant effect on food intake when this tissue is replete, orwhen it is depleted, not by virtue of a signal it generates but via effects lipid storage has on other possible signals.”)
of stored triglyceride (and/or carbohydrate)can somehow vary the responsiveness of thesatiety system to food-derived satiety sig-nals. Thus, during depletion, the systemwould be less responsive to short-term sa-
tiety signals, leading to a greater than usualfood intake. Conversely, after surfeit, thesame system would exhibit an enhanced sen-sitivity to satiety stimuli, with a consequentreduction in spontaneous energy intake.
Although there is some indirect evidenceto support the view that the responsiveness
of appropriate receptor neurons to satietysignals is altered by the nutritional state ofthe animal (54, 55), one can only speculate
on how this “long-term” effect might bemediated.
One line of reasoning that has some ap-peal is based on the growing body of evi-
dence for the existence of insulin-sensitiveglucose receptors (glucoreceptors) in the hy-
pothalamus and elsewhere (56-59).Whereas certain glucoreceptors in the cen-
tral nervous system and periphery may beconcerned principally or exclusively with themaintainance of homeostasis of blood glu-cose , it is also possible that there are otherinsulin-sensitive glucoreceptors in the brainand (perhaps) liver whose primary functionit is to detect and respond to satiety signals.
Similar receptors might be directly respon-sive to other substrates.
If one postulates the existence in the sa-tiety system of insulin-sensitive receptors ofsatiety signals, it becomes possible to con-
sider with fewer misgivings an insulin-iiie-diated mechanism of long-term regulation.
The thread of the argument would be some-
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

748 VAN ITALLIE ET AL.
what as follows: Prominent among the manymetabolic events that accompany and followfood ingestion is an increase in insulin secre-tion (60-62). Insulin secretion not only fol-lows hyperglycemia but occurs in responseto protein and fat ingestion as well (63).Because insulin is consistently secreted after
food ingestion, it could be argued that it isthe magnitude of the postprandial insulinresponse that modulates the sensitivity of
the putative insulin-sensitive satiety recep-tons. This notion is made more plausible byinformation that the insulin secretory re-sponse to food is, in fact, modified by nutri-tional state . In nutritionally depleted ani-mals the insulin secretory response to glu-cose is blunted (64); in obese animals, it isaugmented. Also, certain branched chainand aromatic amino acids are elevated inobesity, and Felig et al. (65) have suggestedthat, in obesity, hyperaminoacidemia may
serve as an important stimulus for continu-ing insulin release . Thus, the size of the fatdepot could modulate the satiety threshold
by determining the magnitude of insulin Se-cretory response to a given meal. Depletionof fat stores would then raise the satietythreshold by reducing the size of the insulin
response to the absorbed products of diges-tion, thereby favoring an increase in meal
size and, by extension, in energy intake.Unfortunately, this hypothesis is seriouslyweakened by evidence (66) that the insulinsecretory response to a meal can be drasti-cally altered by manipulating the propor-tions of fat and carbohydrate in a diet thatremains unchanged in energy content.
Other hypotheses to explain long-termregulation also can be derived from currentknowledge about adipocyte physiology . Forexample (as mentioned earlier), there ap-pears to be a relationship between the sizeof the adipocyte and the rate at which (un-
der specified conditions) it releases glyceroland FFA. Thus, it seems appropriate to con-sider whether a mechanism might exist bywhich the concentration of either of these
moieties in the circulation could be moni-tored by suitable receptor neurons. Indeed,it has already been suggested that the rate atwhich glycerol is metabolized by the livercan somehow be signaled to the satiety re-ceptors as an index of adipocyte status (42).
Also, there is indirect evidence suggestingthat a locus within the hypothalamus may becapable of monitoring directly the concen-tration of FFA in plasma (67).
Although our model invokes modulationof the satiety response to account for long-term regulation , alternative explanationsare also plausible and may ultimately proveto be more in accord with the facts. Forexample, long-term adjustment of short-
term intake could be accomplished by
changing the amplitude of satiety (or hun-ger) signals rather than the sensitivity ofsuitable receptors. Brobeck (68) has pro-posed that the strength of satiety signals isvaried by the rate of removal from the extra-cellular fluid of diet-derived substrate byliver, adipose tissue , and muscle . Thus, inthe depleted state , the rate of uptake wouldbe rapid and the signals would be corre-spondingly weaker and short-lived. In thefed or overnourished state , the rate of up-take would be slower and the signals wouldbe stronger and more prolonged.
It is not possible in this relatively briefdiscussion to consider in greater detail thearguments for and against the view that sig-
nals arising in the periphery and reflectingnutritional status are an integral part of themechanism by which energy storage is reg-ulated. However, it is important to empha-size that some of the basic assumptions ofthe traditional view that a ventromedial hy-pothalamic “satiety center” exercises inhibi-tory control over a lateral “feeding center”have been repeatedly challenged on ana-tomical as well as on physiological grounds
(6, 69-73). For example, Booth (6) hasshown that rats with apparently adequatelesions placed in the ventromedial nucleishow normal suppression of subsequentfood intake after absorption of a meal. Ratswith similar lesions dorsal and lateral to thefornix did, however, show satiety deficits. Ithas also been reported that ventromediallylesioned rats will eat more in response toinsulin-induced hypoglycemia (74) and will
decrease food intake after intraperitonealadministration of glucose (75). In contrast,rats that have recovered feeding after lateral
hypothalamic injury do not eat more afterinsulin; however, the same animals remainresponsive to other major controls of feed-
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

FOOD INTAKE REGULATION 749
ing, adjusting intake precisely to changes in
the nutrient density of the diet and eatingless in the heat and more in the cold (74).
The foregoing studies seem to be at oddswith the simplistic view that insulin-sensitiveglucoreceptors in the ventromedial areatransduce glucose utilization rates into sa-
tiety messages capable of holding in checklaterally mediated feeding behavior. Nev-ertheless, it is possible to reconcile at leastsome of the apparent discrepancies in theliterature about the hypothalamus by postu-lating the presence of glucoreceptors in thelateral as well as in the ventromedial orperifornical regions. The lateral receptorsmight be specifically responsive to hypogly-cemia, inducing food ingestion as a defenseagainst this metabolic emergency. Indeed,as Panksepp (36) has suggested, it is con-ceivable that the lateral hypothalamic areaembodies the system which responds toshort-term satiety signals, whereas the yen-tromedial area mirrors or responds to theanimal’s nutritional status.
The mechanism we have proposed forintegrating short-term and long-term regula-tion of food intake is obviously oversimpli-fied and highly speculative . But it is custom-ary to rationalize hypotheses of this kind byaffirming that they have, at least, some heu-ristic value , and we do not shrink from mak-ing such a claim. To this end, we should liketo summarize a series of experiments con-ducted by our group that appear to substan-tiate the notion that the status of the carbo-hydrate reserves, at least, can exert a spe-cific and important influence upon the food-motivated behavior of food-deprived ani-mals.
The odd carbon fatty acid-enriched model
In these studies, we have used as a modelthe odd carbon-enriched (OCE) animal de-veloped in our laboratory. In 1969, Camp-bell and Hashim (76) reported that when adiet containing tnundecanoin as the majorfat source is fed to weanling rats for 4 to 6weeks, the animals exhibit substantial en-richment of depot fat with undecanoate andhigher fatty acids with odd-numbered car-bon skeletons. Of particular interest is theobservation , repeatedly confirmed, that ani-
mals so enriched maintain significantly
higher concentrations of liver glycogen and
serum glucose during food deprivation thando controls previously fed a diet containinga conventional fat source (77-83). This ne-markable response appears to occur becausethe terminal three-carbon residues arisingfrom /3-oxidation of the mobilized odd car-bon acids are potentially glucogenic,thereby preventing in part the marked de-creases in liver glycogen and plasma glucose
usually induced by starvation.As might be predicted, OCE rats also
exhibit significantly higher serum levels ofimmunoreactive insulin and lower plasma
concentrations of free fatty acids and ke-tones than do comparably deprived controls(78, 79). Metabolic differences between
OCE and control animals are summarized inFigure 3. Taken together, these findingssuggest that during starvation, the fuel used
by OCE animals contains a higher than
usual proportion of carbohydrate.
In the nonfasted state, liver glycogen andserum glucose , insulin , free fatty acid , ke-tone, and alanine concentrations of OCE
animals do not differ significantly fromthose of controls (77-84). Accordingly, theprincipal metabolic effect of odd carbon
fatty acid enrichment is to expand substan-tially the carbohydrate reserves available toOCE animals during fasting (Fig. 4). Be-cause of the special ability of the OCE ani-mal to withstand carbohydrate depletion un-der nutritional conditions that induce nearexhaustion of the preformed carbohydratesupply in control animals, this preparationseems to provide a useful model in which totest the effect of the carbohydrate status onfood related behavior independently of theeffect of general caloric deprivation.
Behavioral studies
Initially, we found that when the feeding
behavior of OCE and control animals wastested by means of a progressive ratio sched-
ule of reinforcement, the OCE animalsstopped responding at lower ratios after a
48-hr fast than did their nonenriched con-trols (81). On this schedule, the animal inthe operant chamber is required to maketwo presses for the first reward , four for the
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

ODD CARBONFATTY-ACID DERIVED
PROPIONATE UNITS4.2g
MUSCLELACTATE
BLOOD GLUCOSE
GLUCOGENIC
AMINO ACIDS
FIG. 4. Potential sources of liver glycogen and
blood glucose during starvation . Resrves of nonpro-
tein precursors of carbohydrate in the rat can be
markedly expanded when adipose tissue triglycerides
are enriched (in this case to 40%) with undecanoate
and higher fatty acids with odd carbon skeletons.
Figures for liver glycogen and depot fat content are
based on studies by J. H. Salter (Am. J. Clin.
Nutr. 8: 535, 1960) in normal Wistar rats, average
weight 261 g.
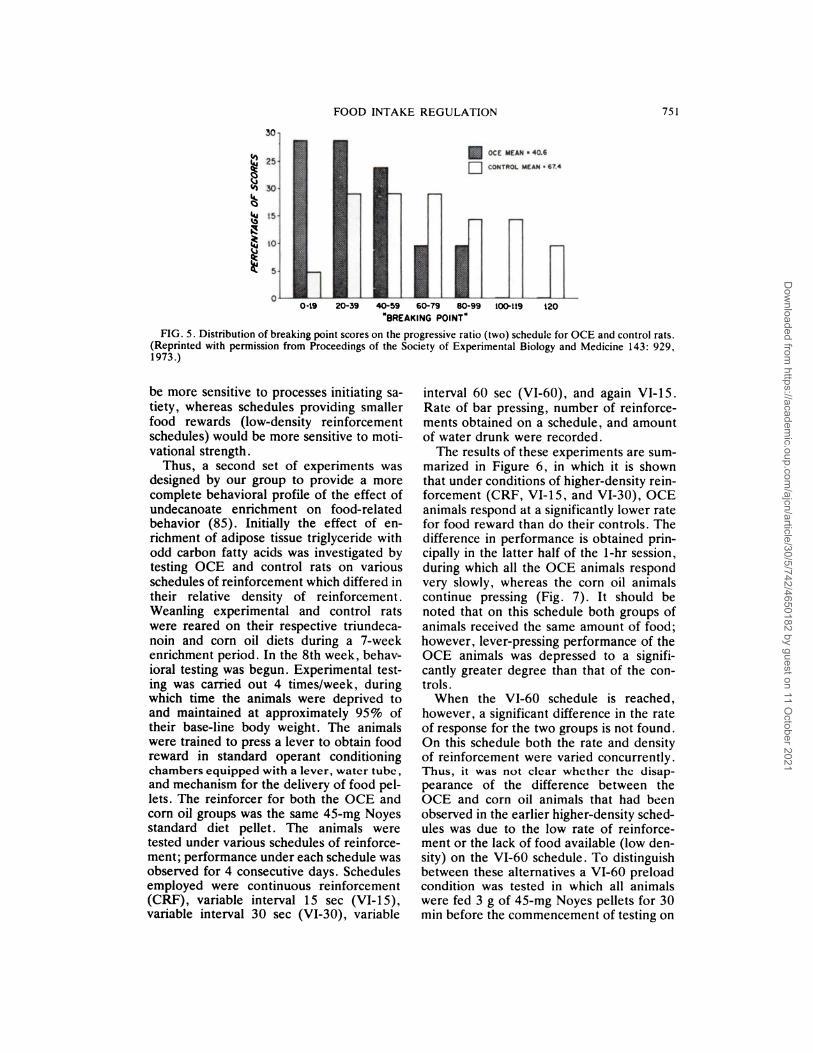
second, six for the third, and so on. The“breaking point” is reached when the ratfails to obtain a reinforcement in 20 mm.The clear difference in distribution of break-
750 VAN ITALLIE ET AL.
FIG . 3 . Comparison of biochemical parameters (mean ± SE) in OCE and corn oil-fed control rats during food
deprivation. All measurements were made at 48 hr starvation unless otherwise indicated (see References 77-79,
84). (Asterisk indicates 24-hr starvation period.)
ing point scores between the two groups ofanimals is shown in Figure 5.
We took this observation to indicate thatthe undecanoate-enriched rats were show-ing a significantly lower drive for food after48 hr of starvation than their controls. Theprogressive ratio test, however, fails ade-quately to distinguish level of drive from thesatiety generated by the food obtained onthe schedule. The more the animal presses,
the more food is obtained, making it unclearwhether it was reduced motivation or morerapid satiation that led to the lowered
breaking point. We recognized that in orderto separate the two processes designated
“hunger” and “satiety” it would be neces-sary to measure performance during food-motivated tasks in which varying amounts offood could be obtained. The effect of availa-bility of food can be systematically studiedby the use of variable interval schedules
which permit variation in the total amount
of food obtained in a fixed time period bymanipulation of the length of the variableinterval: the shorter the interval, the higherthe reinforcement density . Because thesame number of reinforcements can be oh-tamed over a wide range of response rates,variations in response rate observed at agiven reinforcement density should reflectthe specific effect of food rather than thestrength of the drive to obtain food . Accord-ingly , test schedules providing larger quanti-ties of food (high-density schedules) would
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021
0BREAKING POINT”
FOOD INTAKE REGULATION 751
FIG. 5. Distribution of breaking point scores on the progressive ratio (two) schedule for OCE and control rats.
(Reprinted with permission from Proceedings of the Society of Experimental Biology and Medicine 143: 929,
1973.)
be more sensitive to processes initiating sa-tiety , whereas schedules providing smaller
food rewards (low-density reinforcementschedules) would be more sensitive to moti-vational strength.
Thus, a second set of experiments wasdesigned by our group to provide a more
complete behavioral profile of the effect ofundecanoate enrichment on food-relatedbehavior (85). Initially the effect of en-nichment of adipose tissue triglyceride withodd carbon fatty acids was investigated bytesting OCE and control rats on variousschedules of reinforcement which differed intheir relative density of reinforcement.Weanling experimental and control rats
were reared on their respective tniundeca-noin and corn oil diets during a 7-weekenrichment period. In the 8th week, behav-ioral testing was begun . Experimental test-ing was carried out 4 times/week, duringwhich time the animals were deprived to
and maintained at approximately 95% oftheir base-line body weight. The animalswere trained to press a lever to obtain foodreward in standard operant conditioningchambers equipped with a lever, water tube,
and mechanism for the delivery of food pel-lets. The reinforcer for both the OCE andcorn oil groups was the same 45-mg Noyesstandard diet pellet. The animals weretested under various schedules of reinforce-ment; performance under each schedule wasobserved for 4 consecutive days. Schedulesemployed were continuous reinforcement
(CRF) , variable interval 1 5 sec (VI-1 5),variable interval 30 sec (VI-30), vaniable
interval 60 sec (VI-60), and again VI-15.Rate of bar pressing, number of reinforce-
ments obtained on a schedule, and amount
of water drunk were recorded.
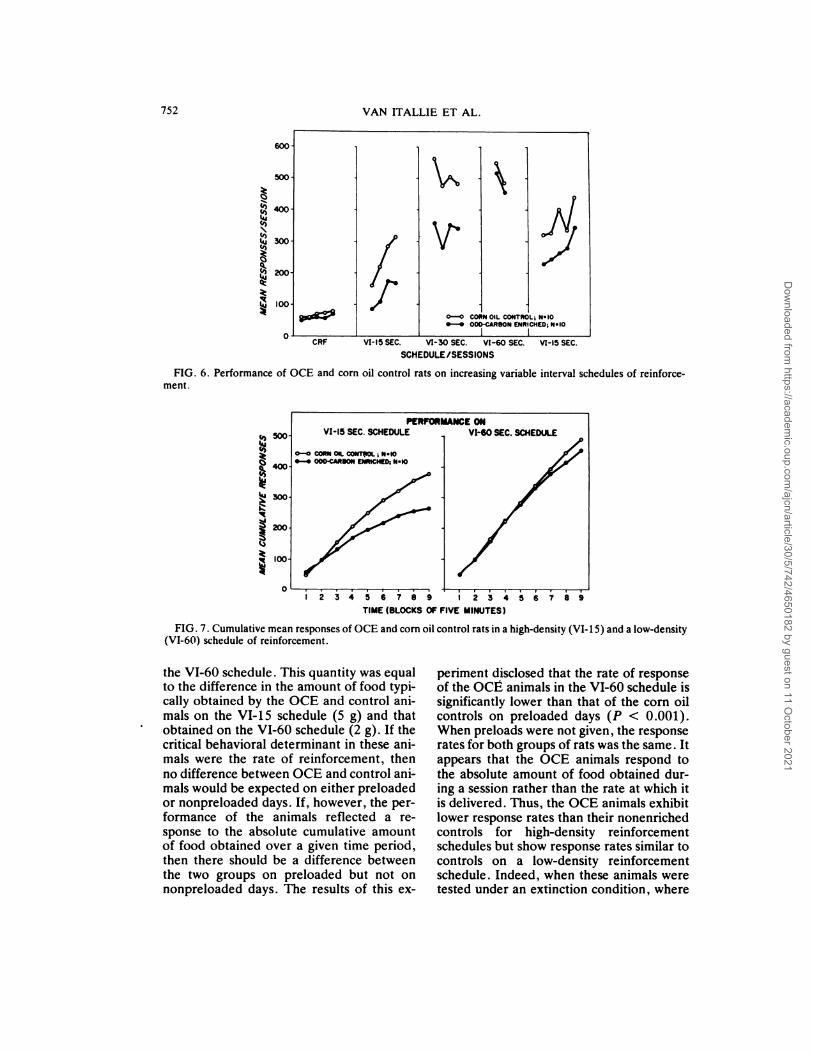
The results of these experiments are sum-manized in Figure 6, in which it is shownthat under conditions of higher-density rein-forcement (CRF, VI-15, and VI-30), OCE
animals respond at a significantly lower ratefor food reward than do their controls. Thedifference in performance is obtained pnin-cipally in the latter half of the 1-hr session,during which all the OCE animals respondvery slowly, whereas the corn oil animals
continue pressing (Fig. 7). It should benoted that on this schedule both groups of
animals received the same amount of food;however, lever-pressing performance of theOCE animals was depressed to a signifi-cantly greater degree than that of the con-trols.
When the VI-60 schedule is reached,however, a significant difference in the rateof response for the two groups is not found.On this schedule both the rate and densityof reinforcement were varied concurrently.Thus, it was not clear whether the disap-
pearance of the difference between theOCE and corn oil animals that had beenobserved in the earlier higher-density sched-ules was due to the low rate of reinforce-ment or the lack of food available (low den-
sity) on the VI-60 schedule. To distinguishbetween these alternatives a VI-60 preloadcondition was tested in which all animalswere fed 3 g of 45-mg Noyes pellets for 30mm before the commencement of testing on
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021
600
500
0-0 CORN OIL CONTROL; N.tO
.-. ODO�C*R8ON ENRICHED; N.IO
PENFONMANCE ONVI-I5 SEC. SCHEDULE VI-60 SEC. ScHEDUII
0-0 co�w ost. coNT�oL, NelO.-. OOO�CARSON LNNtCHtD; N.�O
‘I) 500
�4O0
�300
�200
�I00
0
752 VAN ITALLIE ET AL.
�4OO
�3OO
�2OO
�lOO
CRF VI-15 SEC. V1-30 SEC. Vt-SO SEC. VI-15 SEC.
SCHEDULE/SESSIONS
FIG. 6. Performance of OCE and corn oil control rats on increasing variable interval schedules of reinforce-
ment.
2345�7i� �
TIME (BLOCKS OF FIVE MINUTES)
FIG. 7. Cumulative mean responses of OCE and corn oil control rats in a high-density (VI-iS) and a low-density
(VI-60) schedule of reinforcement.
the VI-60 schedule . This quantity was equalto the difference in the amount of food typi-cally obtained by the OCE and control ani-mals on the VI-15 schedule (5 g) and thatobtained on the VI-60 schedule (2 g). If thecritical behavioral determinant in these ani-mals were the rate of reinforcement, then
no difference between OCE and control ani-mals would be expected on either preloaded
or nonpreloaded days. If, however, the per-formance of the animals reflected a re-sponse to the absolute cumulative amountof food obtained over a given time period,then there should be a difference betweenthe two groups on preloaded but not onnonpneloaded days. The results of this ex-
periment disclosed that the rate of responseof the OCE animals in the VI-60 schedule issignificantly lower than that of the corn oil
controls on preloaded days (P < 0.001).When preloads were not given , the responserates for both groups of rats was the same . Itappears that the OCE animals respond tothe absolute amount of food obtained dur-ing a session rather than the rate at which itis delivered. Thus, the OCE animals exhibitlower response rates than their nonenrichedcontrols for high-density reinforcementschedules but show response rates similar to
controls on a low-density reinforcementschedule. Indeed, when these animals weretested under an extinction condition , where
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

HOURS OF DEPRIVATION
FOOD INTAKE REGULATION 753
no food is available , the performance of thetwo groups was virtually indistinguishable,supporting the impression that the amountof food available is the critical behavioraldeterminant.
In the experimental conditions describedabove , animals were always tested at 95%of base-line body weight, a level typicallyachieved after a 24-hr food deprivation pe-nod. Van Itallie and Khachadurian (77) andPi-Sunyer (79) have shown, however, thatthe plasma glucose concentrations of OCEanimals remained higher than those of con-trols during fasts lasting up to 144 hr. Wefelt that before ascribing the behavioral dif-ferences between OCE and control rats to
processes or events related to their differingcarbohydrate status, it would be necessary,
as a first step, to demonstrate that thesebehavioral differences could be observedover the same range of deprivation intervalsfound in the metabolic studies to producesignificant liver glycogen and plasma glu-cose differences. Accordingly, the OCE andcontrol animals were deprived for variousperiods of time and then tested on the VI-15sechedule of reinforcement already fQund
( see above) to disclose differences in food-motivated behavior between the two
groups.
Animals were first refed ad libitum ontheir appropriate triundecanoin or corn oildiet for at least 1 week before deprivation
and behavioral testing were begun . Fourdeprivation periods were employed: 6, 24,48, and 96 hr. Four groups each of OCEand control animals were tested after theseintervals in a different counterbalanced se-
quence.After testing on each interval, animals
were refed ad libitum on their appropriatediets until base-line body weight had been
regained. They were then deprived and re-tested. Testing was carried out on a VI-15schedule of reinforcement once after eachdeprivation period in a 1-hr session. Rate ofresponse, number of reinforcements, andwater intake were recorded for each animal.
The results are presented in Figure 8 . Ttests for independent groups comparing themean responses per session were significantfor the 6-hr (P < 0.05), 24-hr (P < 0.05),
and 48-hr (P < 0.025) period but not for
the 96-hr period. An additional repeat test
after a 96-hr deprivation interval did pro-duce significant differences (P < 0.05) be-tween OCE and control animals. Appan-ently the effect of the odd carbon enrich-
ment is weak after 96 hr and not readilyobserved. In general, the results of the dep-nivation study are consistent with theearlier metabolic findings: OCE animals re-
sponded at lower rates for food rewardafter a fast than did controls. The largestbehavioral difference between OCE andcontrol animals was observed after a 48-hrfast . This result is consistent withpreviously reported metabolic findings that
demonstrated the largest blood sugar and
liver glycogen differences between the twogroups after a 48-hr fast.
Additional experiments carried out withthese animals suggest that the effects oh-
served in food-related tasks do not appearto be artificially mediated by different activ-ity levels of the OCE animals, because nodifference could be observed between thetwo groups either in activity wheel runningor exploration in an open field.
Carbohydrate status: effect on “satiability”
The difference in behavior during condi-tions of deprivation observed to date in
these two groups of animals appears to be
restricted to situations in which the availa-
FIG. 8. Effect of deprivation period on perform-
ance (mean responses per session ± SE) on a variableinterval 1 5-sec schedule of reinforcement in OCE and
corn oil control rats.
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

754 VAN ITALLIE ET AL.
bility of food is varied. The reduced per-formance of OCE animals on high-densityreinforcement schedules in which theamount of food ingested is large and the
opportunity to satiate is increased suggeststhat the OCE animal satiates more rapidlythan does the control animal. Conversely,
the fact that the performance of the OCEanimal is comparable to the control’s onlow-density reinforcement schedules andunder extinction conditons in which little onno food is ingested and the opportunity tosatiate is decreased or absent suggests thatthe OCE animal is as motivated for food asthe control. However, we recognize that dif-
ferences in hunger motivation may also existbetween OCE and controls that have notbeen disclosed by the behavioral measures
used thus far. It is also conceivable that thetwo types of behavior we commonly desig-nate as “hunger” and “satiety” may actuallybe parts of one continuum.
In the past, it has been difficult to designchronic experiments that could dissociatephysiological responses to carbohydrate de-
pletion from those due to nonspecific energydeprivation . The OCE animal preparationappears to permit an appreciable separationof these two critical variables. Our findingsto date suggest that under identical condi-
tions of energy deprivation, OCE rats, withtheir greater ability to resist carbohydratedepletion, satiate at a faster rate thannonenriched controls.
Before attempting to consider these ob-servations in relation to the general problem
of food intake regulation it may be useful toreview briefly some of the major metabolicadaptations that occur when the body beginsto shift from a “fed” to a “fasted” economy.
The hallmarks of the fasting state seemindisputably to be an increasing reliance bythe body upon its reserves of triglyceridesand protein , together with a reduced rate ofglucose utilization . Insulin plays an impor-tant role in this change of metabolic empha-
sis. As Cahill (86) has pointed out: “Insulinserves as the body’s signal for the fed orfasted state. High insulin levels, the ‘fed’signal, initiate tissue uptake and storage offuels. Low insulin levels, the ‘fasted’ signal,initiate mobilization of stored fuels fromtissue stores, the rate being proportional tothe lowness of insulin.”
During food deprivation, the OCE animalmaintains a metabolic economy that is ap-preciably closer to the “fed” state than itsmore conventionally nourished control.When this fact is considered in relation tothe observation that after deprivation, OCErats satiate more rapidly than their controls,it is tempting to attribute the behavioraldifferences between the two groups of ani-mals to their metabolic differences. Fromthe model described earlier, it could havebeen predicted that OCE animals would ex-hibit greater satiability after deprivationthan their nonenriched controls.
Because studies with the OCE prepara-
tion are still at an early stage , it would behazardous to overinterpret the preliminaryresults just described. There is every indica-tion that the mechanism by which food in-take is regulated is very complicated, withnumerous redundancies built into the sys-tern. However, food-related behavior of thedeprived OCE rat is consistent with thenotion that the response of the “satietysystem” to short-term signals generated bythe eating process and by ingested food issubject to modulation by corrective signalsreflecting the animal’s nutritional state . Itwill require a continuing multidisciplinaryeffort to identify these signals with greatercertainty and to unravel their complex in-terrelationships.
References
1 . HETHERINGTON, A. W. , AND S. W. RANSON. Ex-
perimental hypothalamico-hypophysial obesity inthe rat. Proc. Soc. Exptl. Biol. Med. 41: 465,
1939.
2. BROBECK, J. R., J. TEPPERMAN AND C. N. H.LONG. Experimental hypothalamic hyperphagia in
thealbinorat. YaleJ. Biol. Med. 15: 831, 1943.
3. ANAND, B. K., AND J. R. BROBECK. Hypotha-
lamic control of food intake in rats and cats. Yale
J. Biol. Med. 24: 123, 1951.
4. DELGADO, J. D. R., AND B. K. ANAND. Increase
of food intake induced by electrical stimulation of
the lateral hypothalamus. Am. J. Physiol. 172:
162, 1953.5. BROBECK, J. R. Mechanism of the development of
obesity in animals with hypothalamic obesity.
Physiol. Rev. 26: 541, 1964.6. BooTH, D. A., F. M. TOATES AND S. V. Pi�i-r.
Control system for hunger and its implications inanimals and man. In: Hunger: Basic Mechanisms
and Clinical Implications, edited by D. Novin,W. Wyrwicka and G. A. Bray. New York:
Raven Press, 1976, pp. 127-43.
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

FOOD INTAKE REGULATION 75:s
7. STEFFENS, A.B. Blood glucose and FFA levels in
relation to the meal pattern in the normal rat and
the ventromedial hypothalamic lesioned rat. Phys-
iol. Behav. 4: 215, 1969.
8. PAINTAL, A. S. A study of gastric stretch recep-tors. Their role in the peripheral mechanism of
satiation of hunger and thirst. J. Physiol. 126: 255,
1954.
9. SMITH, G. P., J. GmBs AND R. C. YOUNG. Chole-cystokinin and intestinal satiety in the rat. Federa-
tion Proc. 33: 1146, 1974.
10. ANTIN, J. GIBBS, J. HOLT, R. C. YOUNG AND
G. P. SMITH. Cholecystokinin elicits the com-
plete behavioral sequence of satiety in rats . J.Comp. Physiol. Psychol. 89: 784, 1975.
11. SCHALLY, A. V., T. W. REDDING, H. W. LUCIEN
AND J. MEYER. Enterogastrone inhibits eating by
fasted mice. Science 157: 210, 1967.
12. HUNT, J. N. The duodenal regulation of gastric
emptying. Gastroenterology 45: 149, 1963.
13. SCHWARTZBAUM, J. S. , AND H. P. WARD. An
osmotic factor in the regulation of food intake in
the rat. J. Comp. Physiol. Psychol. 51: 555, 1958.
14. SHUFORD, E. H., J�. Palatability and osmotic pres-
sure of glucose and sucrose as determinants of
intake. J. Comp. Physiol. Psychol. 52: 150, 1959.
15. JACOBS, H. L. Evaluation of the osmotic effects of
glucose loads in food satiation. J. Comp. Physiol.
Psychol. 57: 309, 1964.
16. MELLINKOFF, S. M., M. FRANKLAND AND M.GREIPEL. Effect of amino acid and glucose inges-tion on arteriovenous blood sugar and appetite . J.Appl. Physiol. 9: 85, 1956.
17. PANKSEPP, J., AND D. A. BOoTH. Decreased feed-
ing after injections of amino acids into the hypo-
thalamus. Nature 233: 341, 1971.18. WURTMAN, R. J., AND J. D. Fernstrom. Effect of
the diet on brain neurotransmitters. Nutr. Rev.
32: 193, 1974.
19. VAN ITALLIE, T. B., R. BEAUDOIN AND J. MAYER.
Arteriovenous glucose differences, metabolic hy-poglycemia and food intake in man. J. Clin. Nutr.
1: 208, 1953.
20. STUNKARD, A. J., AND H. G. WOLFF. Studies on
the physiology of hunger. 1 . The effect of intrave-
nous administration of glucose on gastric hunger
contractions in man. J. Clin. Invest. 35: 954,
1956.
21. STUNKARD, A. J., T. B. VAN ITALLIE AND B. B.REIss. Effect of glucagon on gastric hunger con-
tractions in man. Proc. Soc. Exptl. Biol. Med. 89:
258, 1955.
22. BOOTH, D. A. Feeding inhibition by glucose loads,
compared between normal and diabetic rats. Phys-
iol. Behav. 8: 801, 1972.
23. NOVIN, D., J. D. SANDERSON AND D. A. WAN-
DERWEELE. The effect of isotonic glucose on eating
as a function of feeding conditions and infusion
site. Physiol. Behav. 13: 3, 1974.
24. NIUIMA, A. Afferent impulse discharges from glu-
coreceptors in the liver of the guinea pig. Ann.
N.Y. Acad. Sci. 157: 690, 1969.25. RUSSEK, M., A. M. RODRIGUEZ-ZIDEJAS AND S.
PINA. Hypothetical liver receptor and the anorexia
caused by adrenaline and glucose . Physiol. Behav.
3: 249, 1968.
26. BERNSTEIN, L. M., AND M. I. GROSSMAN. An
experimental test of the glucostatic theory of regu-
lation of food intake. J. Clin. Invest. 35: 627,
1956.
27. MELLINKOFF, S. Digestive system. Ann. Rev.
Physiol. 19: 195, 1957.
28. KOOPMANS, H. S. Jejunal signals in hunger satiety.
Behav. Biol. 14: 309, 1975.
29. KOOPMANS, H. S. In press.
30. EDHOLM, 0. G. Energy expenditure and caloric
intake in young men. Proc. Nutr. Soc. 20: 71,
1961.
31. KHOSLA, T., AND W. Z. BILLEwICz. Measurement
of change in body weight. Brit. J. Nutr. 18: 227,
1964.
32. RoBINSoN, M. F., AND P. E. WATSON. Day-to-day
variations in body weight of young women . Brit . J.Nutr. 19: 225, 1965.
33. ADOLPH, E. F. Urges to eat and drink in rats. Am.
J. Physiol. 151: 110, 1947.
34. CAMPBELL, R. G., S. A. HASHIM AND T. B. VAN
ITALLIE. Studies of food-intake regulation in man.
Responses to variations in nutritive density in lean
and obese subjects. New Engl. J. Med. 285: 1402,
1971.
35. COHN, C., AND D. JOSEPH. Influence of body
weight and body fat on appetite of “normal” and
obese rats. Yale J. Biol. Med. 34: 598, 1962.36. PANKSEPP, J. Hypothalamic regulation of energy
balance and feeding behavior. Federation Proc.
33: 1150, 1974.
37. SIMs, E. A. H., E. DANFORTH, E. S. HORTON,G. A. BRAY, J. A. GLENNON AND L. B. SALANS.
Endocrine and metabolic effects of experimental
obesity in man. Recent Progr. Horm. Res. 29:
457, 1973.
38. KEYS, A., J. BROZEK, A. HENSHEL, 0. MICKEL-
SEN AND H. L. TAYLOR. The Biology of Human
Starvation. 1950, vol. 1. Minneapolis; University
of Minnesota Press.
39. HAUSBERGER, F. X. Uber die innervation der fet-
torgans. Z. Mikroskop. Anat. Forsch. 36: 231,
1934.40. WERTHEIMER, E. , AND B. SHAPIRO. The physiol-
ogy ofadipose tissue. Physiol. Rev. 28: 451, 1948.
41 . KENNEDY, G. C. The role of depot fat in the
hypothalamic control of food intake in the rat.
Proc. Roy. Soc. London (B) 140: 578, 1953.
42. Biu�Y, G. A., AND L. A. CAMPFIELD. Metabolic
factors in the control of energy stores. Metabolism
24: 99, 1975.
43. BJORNTORP, P. Disturbances in the Regulation of
Food Intake. Adv. Psychosom. Med. 7: 116,1972.
44. SMITH, V. Effects of glucose and insulin on lipoly-
sis rates in human fat cells of different sizes. FEBSLett. 11: 8, 1970.
45. SALANS, L. B., J. L. KNITTLE AND J. HIRSCH. The
role of adipose cell size and adipose tissue insulin
sensitivity in the carbohydrate intolerance of hu-
man obesity. J. Clin. Invest. 47: 153, 1968.
46. KARAM, J. H. , G. M. GRODSKY AND P. H. FoR-
SHAM. Excessive insulin response to glucose in
obese subjects as measured by immunochemical
assay. Diabetes 19: 656, 1970.47. HERVEY, G. R. Regulation of energy balance.
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

756 VAN ITALLIE ET AL.
Nature 222: 629, 1969.48. BAILE, C. A., C. W. SIMpsoN, S. M. BEAN, C. L.
MCLOUGHLIN AND H. L. JAcoBs. Prostaglandins
and food intake of rats: a component of energy
balance regulation? Physiol. Behav. 10: 1077,
1973.
49 . PFAFF, D . W . Histological differences between
ventromedial neurons of well fed and underfed
rats. Nature 223: 77, 1969.
50. POWLEY, T. L. Hypothalamic cytoarchitectonics
and body weight regulation. In: Proceedings of theInternational Symposium on Hunger and Regula-
tion of Body Energy. Ememonville, France, 1973.
5 1 . NIcOiAIDI5, S. A possible molecular basis of regu-
lation of energy balance. In: Physiology of Food
and Fluid Intake. XXVI International Congress of
Physiological Sciences. Jerusalem Satellite Sympo-
sia, 1974, p. 97 (abstr.).
52. WOLF, A. Z. Thirst: Physiology of the Urge to
Drink and Problems of Water Lack. Springfield,
Illinois: Charles C Thomas, 1958.
53. BLA55, E. Cellular dehydration thirst: physiologi-
cal, neurological and behavioral correlates. In:
Neurophysiology of Thirst. New Findings and Ad-
vances in Concepts, edited by A. N. Epstein, H.
R. Kissileff and E. Stellar. Washington, D.C.: V.
H. Winston and Sons, 1973, pp. 37-72.
54. HOEBEL, B. G. Feeding: Neural control of intake.
Ann. Rev. Physiol. 33: 533, 1971.
55. HOEBEL, B. G. Satiety: hypothalamic stimualtion,
anorectic drugs and neurological substrates . In:
Hunger: Basic Mechanisms and Clinical Implica-tions, edited by D. Novin, W. Wyrwicka and
0. A. Bray. New York: Raven Press, 1976,
pp. 33-50.
56. DEBONS, A. F., I. KRIMSKY, A. FROM et al. Rapideffects of insulin on the hypothalamic satiety cen-
ter.Am. J. Physiol.217: 1114, 1969.57. DEBONS, A. F., I. KRIMSKY AND A. FROM. A
direct action of insulin on the hypothalamic satiety
center. Am. J. Physiol. 219: 938, 1970.58. Sz�.ao, 0., AND A. J. SZABO. Studies on the nature
and mode of action of the insulin-sensitive gluco-
regulator receptor in the central nervous system.
Diabetes 24: 328, 1975.
59. HATFIELD, J. S., W. J. MILLARD AND C. J. V.SMrrH. Short-term influence of intra-ventromedial
hypothalamic administration of insulin on feeding
in normal and diabetic rats. Pharmacol. Biochem.
Behav. 2: 223, 1974.
60. M�i..aussE, W. J., AND F. MALAISSE-LAGAC. Stim-
ulation of insulin secretion by noncarbohydrate
metabolites. J. Lab. Clin. Med. 72: 438, 1968.61 . UNGER, R. H., H. KETrERER, J. DUPRE AND A.
M . EIsENra�ur. The effects of secretin , pan-
creaozymin , and gastrin on insulin and glucagon
secretion in anesthetized dogs . J . Clin . Invest . 46:630, 1967.
62. RABINOwITz, D., J. J. MERIMEE, R. MAFFEZZOLI
AND J. A. BURGESS. Patterns of hormonal release
after glucose , protein , and glucose plus protein.
Lancet 2: 454, 1966.
63. CARROLL, K. F., AND P. J. NESTEL. Effect of long-
chain triglyceride on human insulin secretion. Dia-
betes 21: 923, 1972.
64. BOSBOOM, R. S., J. ZWEENS AND P. R. BOUMAN.
Effects of feeding and fasting on the insulin secre-tory response to glucose and sulfonylureas in intact
rats and isolated perfused pancreas. Diabetologia
9: 243, 1973.
65. FEUG, P., E. M�iIss AND 0. F. CAHILL JR.
Plasma amino acid levels and insulin secretion in
obesity. New Engl. J. Med. 281: 811, 1969.66. FARQUHAR, J. W., J. OLEFSKY, M. STERN AND G.
M. REAVEN. Obesity, insulin and triglycerides. In:
Obesity in Perspective, edited by G. Bray. Wash-ington, D.C.: Dept. of Health, Education and
Welfare Pub. No.(NIH) 75-708, 1976, vol. 2, Part2, pp. 313-319.
67. OOMURA, Y. , T. NAKAMURA, M. SUGIMo�i AND
Y. YAMADA. Effect of free fatty acid on the ratlateral hypothalamic neurons. Physiol. Behav. 14:
483, 1975.
68. BROBECK, J. R. Nature of satiety signals. Am. J.Clin. Nutr. 28: 806, 1975.
69 . PANKSEPP, J . Is satiety mediated by the ventrome-
dial hyopthalamus? Physiol. Behav. 7: 381 , 1971.
70. RABIN, B. M. Ventromedial hypothalamic control
of food intake and satiety: A reappraisal. Brain
Res. 43: 317, 1972.71. AHLSKOG, J. E., AND B. G. HOEBEL. Overeating
and obesity from damage to a nonadrenergic sys-
tern in the brain. Science 182: 166, 1973.
72. GOLD, R. M. Hypothalamic obesity: the myth ofthe ventromedial nucleus. Science 182: 488, 1973.
73. GROSSMAN, S. P. Role of the hypothalamus in the
regulation of food and water intake . Psychol. Rev.82: 200, 1975.
74. EPSTEIN, A. N., AND P. TEITELBAUM. Specificloss of the hypoglycemic control of feeding in re-
covered lateral rats. Am. J. Physiol. 213: 1159,1967.
75. REYNOLDS, R. W., AND J. KIMM. Effect of glucoseon food intake in hypothalamic hyperphagic rats.
J. Comp. Physiol. Psychol. 60: 438, 1965.76. CAMPBELL, R. G., AND S. A. HASHIM. Deposition
in adipose tissue and transport of odd-numbered
fatty acids. Am. J. Physiol. 217: 1614, 1969.77. VAN ITALLIE, T. B., AND A. K. KHACHADURIAN.
Rats enriched with odd-carbon fatty acids: mainte-
nance of liver glycogen during starvation . Science
165: 811, 1969.
78. PI-SUNYER, F. X. Resistance to ketosis during
prolonged fasting by rats fed on a diet containingundecanoic acid, an odd-carbon-number fatty
acid. Brit. J. Nutr. 35: 41, 1976.79. PI-SUNYER, F. X. Rats enriched with odd-carbon
fatty acids. Effect of prolonged starvation on liver
glycogen and serum lipids, glucose and insulin.
Diabetes 20: 200, 1971.
80. KIM, S. , AND F. X. PI-SUNYER. Beta-cell insulin
secretory response to glucose in odd-carton num-
ber fatty acid-enriched rats. Preservation during
starvation. Diabetes 23: 605, 1974.
81 . QUARTERMAIN, D., M. E. JUDGE AND T. B. VAN
ITALLIE. Hunger drive during starvation in rats
enriched with odd-carbon fatty acids. Proc. Soc.
Exptl. Biol. Med. 143: 929, 1973.
82. Guy, D. G., AND R. C. THEUER. The effect of
feeding various levels of triundecanoin on some
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021

FOOD INTAKE REGULATION 757
blood and tissue rnetabolites. Federation Proc. 31:
67, 1972.83. ANDERSON, R. L., AND R. W. Boaos. Glucogenic
and ketogenic capacities of lard, safflower oil and
triundecanoin in fasting rats. J. Nutr. 105: 185,
1975.
84. PI-SUNYER, F. X., J. M. CONWAY, M. LAvAU, R.G. CAMPBELL AND A. B. EISENSTEIN. Glucagon,
insulin and gluconeogenesis in fasted odd carbon
fatty acid-enriched rats. Am. J. Physiol. 231:
366. 1976.
85. SCHUPF, N., D. QUARTERMAIN AND T. B. VAN
ITALLIE. Food-motivated behavior in the odd-car-
bon enriched rat: Relationship to maintenance of
carbohydrate stores during food deprivation.
Physiol. Behav. 16: 271, 1976.
86. CAHILL, G. F., JR. Physiology of insulin in man.
Diabetes 20: 785, 1971.
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/30/5/742/4650182 by guest on 11 O
ctober 2021





























