Embed Size (px)
Citation preview

Plant Physiol. (1991) 96, 467-4720032-0889/91/96/0467/06/$01 .00/0
Received for publication August 29, 1990Accepted January 22, 1991
Short-Term Effects of Carbon Dioxide on Carnation CallusCell Respiration1
Artur Palet, Miquel Ribas-Carb6, Josep M. Argiles, and Joaquim Azc6n-Bieto*Departament de Biologia Vegetal (A.P., M.R., J.A.) and Departament de Bioquimica i Fisiologia (J.M.A.),
Facultat de Biologia, Universitat de Barcelona, Diagonal 645, 08028 Barcelona, Spain
ABSTRACT
The addition of potassium bicarbonate to the electrode cuvetteimmediately stimulated the rate of dark 02 uptake of photomixo-trophic and heterotrophic carnation (Dianthus caryophyllus L.)callus, of Elodea canadensis (Michx) leaves, and of other planttissues. This phenomenon occurred at pH values lower than 7.2to 7.8, and the stimulation depended on the concentration ofgaseous CO2 in the solution. These stimulatory responses lastedseveral minutes and then decreased, but additional bicarbonateor gaseous CO2 again stimulated respiration, suggesting a re-versible effect. Carbonic anhydrase in the solution increased thestimulatory effect of potassium bicarbonate. The C02/bicarbonatedependent stimulation of respiration did not occur in animaltissues such as rat diaphragm and isolated hepatocytes, and wasinhibited by salicylhydroxamic acid in carnation callus cells andE. canadensis leaves. This suggested that the alternative oxidasewas engaged during the stimulation in plant tissues. The cyto-chrome pathway was severely inhibited by C02/bicarbonateeither in the absence or in the presence of the uncoupler carbonyl-cyanide m-chlorophenyl hydrazone. The activity of cytochrome coxidase of callus tissue homogenates was also inhibited by C02/bicarbonate. The results suggested that high carbon dioxidelevels (mainly free C02) partially inhibited the cytochrome path-way (apparently at the oxidase level), and this block in electrontransport elicited a large transient engagement of the alternativeoxidase when present uninhibited.
Effects of carbon dioxide (including under this term freeCO2 and bicarbonate) on plant tissue respiration have beenreported (1 1), and are diverse and sometimes rather contra-dictory. On the one hand, many studies have reported inhib-itory effects of high CO, on dark respiratory CO, evolutionand 02 uptake rates in several organs and tissues: fruits (9,29), roots (21), leaf epidermal strips (26), alfalfa leaves (25),and tissue cultured cells (10). On the other hand, severalauthors have reported that high carbon dioxide levels stimu-late respiratory rates in potato tubers (24), lemon fruits (31),and peanut leaves (30). High CO2 levels also induce cyanideresistant respiration in potato tuber slices ( 13, 24) and wheatseedlings (15), and can also have a synergistic effect on eth-ylene- and wound-induced cyanide resistant respiration (6).
' Supported by grants from the Comisi6n Interministerial de Cien-cia y Tecnologia, Programa Nacional de Investigaci6n Agricola(Grant AGR89-580) and the Comissi6 Interministerial de Ciencia iTecnologia, Generalitat de Catalunya.
The mechanisms by which CO2 differentially affects tissuerespiration are not well understood at present and need to beevaluated, provided that many plant tissues are exposed tovery high CO2 levels under natural conditions (e.g. roots inthe soil and in flooded systems, refs. 4 and 21; stems, ref. 22;bulky organs such as some fruits, refs. 4 and 9) and underartificial conditions (e.g. fruit and vegetable storage, ref. 29;in vitro culture conditions, ref. 12). These mechanisms mightalso be relevant to understand long-term adaptations of plantrespiration to high atmospheric CO2 levels (25) (BG Drake,personal communication).We have studied short-term (minutes) effects of carbon
dioxide (supplied as free CO2 or bicarbonate) on respirationof carnation callus cells and other plant and animal tissues.Inhibitors of electron transport pathways and uncouplers ofoxidative phosphorylation have been used to examine thepossible influence of CO, on the partitioning of electrons tothe cytochrome and alternative pathways. The results suggestthat carbon dioxide affected respiration mainly through aninhibition of the Cyt pathway. A preliminary short report ofthis work was presented elsewhere (23).
MATERIALS AND METHODS
Plant Material
Heterotrophic callus of carnation (Dianthus carvophyllusL. cv "Corrida") were obtained by culturing petals from youngfloral buds (about 12 mm diameter) in Murashige and Skoogmedium (18) supplemented with 3% sucrose as a source oforganic carbon, and 4.5 ltM 2,4-D and 0.44 Mm benzyladenineas growth regulators. Agar (0.6%) was used. Callus developedfrom the base of the petals. Once cultures were established,cell greening was induced by supplying a quantum flux densityof 90 ,E m-2 s-' (with OSRAM FLUORA fluorescent tubes),and by substituting the 2,4-D with 27 AM naphtaleneaceticacid, and by increasing the concentration of benzyladenine to2.2 AM. Callus cultures were maintained by subculturing every21 d. Some calli were also maintained under complete heter-otrophic conditions (see above) for 2 years. The age of thecallus used in the experiments ranged between 21 and 28 dafter subculture. Leaves of the aquatic plant Elodea canaden-sis (Michx) were also used (1).
Measurement of Oxygen Uptake
Oxygen uptake rates of callus cells and plant tissues weremeasured in the dark at 25°C using a Clark type oxygen
467
www.plantphysiol.orgon June 30, 2020 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 96, 1991
electrode (Rank Brothers, Cambridge, England) in 4 mL ofan air saturated buffer solution (the initial concentration ofoxygen was considered to be 253 AM). About 0.3 g of calluscells was placed in the electrode cuvette. Buffer solutions usedwere 30 mm Hepes (pH 7.2 and 7.8) and 30 mM Mes (pH 5.7and 6.5). A nylon net separated the callus from the stirrer barand the electrode. The depletion of the oxygen concentrationin the rapidly stirred solution of the closed cuvette was linearwith time, except at low oxygen concentrations (less than 50,iM). Measurements were made in the dark between 253 and75 AM 02. Potassium bicarbonate and SHAM2 were addedfrom 1 M stock aqueous solutions, potassium cyanide from a0.4 M stock solution, and CCCP from a 2 mm stock ethanolicsolution. These compounds were added with a Hamiltonmicrosyringe to the electrode cuvette. In the experiments inwhich carbonic anhydrase was used, the 30 mM Mes buffersolution (pH 6.5) included 0.2 mg mL-' of this enzyme(supplied by Sigma). In the experiments in which the effectsof different concentrations of bicarbonate were tested, thebuffer used was 50 mm Mes (pH 5.7 and 6.5). CO2 and HCO-present in the solution were calculated according to Buch (5),assuming that they were in equilibrium.Oxygen uptake rates of intact, freshly extracted, rat dia-
phragm muscle tissue were measured similarly to those ofplant tissues, except that the assay medium temperature was30°C and it contained 137 mm NaCl, 5 mm glucose, 0.2 mMCaCl,, and 30 mm Hepes buffer (pH 6.6 and 7.0). Thediaphragm was rapidly extracted after sacrifice of a youngWistar rat, and then was rinsed briefly with assay mediumand placed into the electrode cuvette. The initial oxygenconcentration was considered to be 230 ,uM, and the measure-
2 Abbreviations: SHAM, salicylhydroxamic acid; CCCP, carbonyl-cyanide m-chlorophenyl hydrazone.
Table I. Effect of 20 mM Potassium Bicarbonate on the Rate of DarkOxygen Uptake of Photomixotrophic and Heterotrophic CarnationCallus
Measurements were made at several pH values as shown. The pHvalues refer to the initial ones measured in the buffer solution. Theaddition of bicarbonate may slightly increase the pH (a maximum of0.3 unit), especially when the solution is rather acid. The valuesshown are means ± SE of four to seven replicates.
Oxygen UptakepH Increase
-HCO3 +HCO3
nmol 02 g9-' fresh %wt-min1
Photomixotrophic callus5.7 51 ± 2 104 ± 8 109 ± 66.5 36 ± 5 55 ± 7 52 ± 137.2 45±2 59±2 32±47.8 52±8 54±2 4±3
Heterotrophic callus5.7 24 ± 6 47 ± 12 97 ± 86.5 22 ± 3 29 ± 5 35 ± 147.2 34±6 35±6 2±27.8 29±6 29±6 0
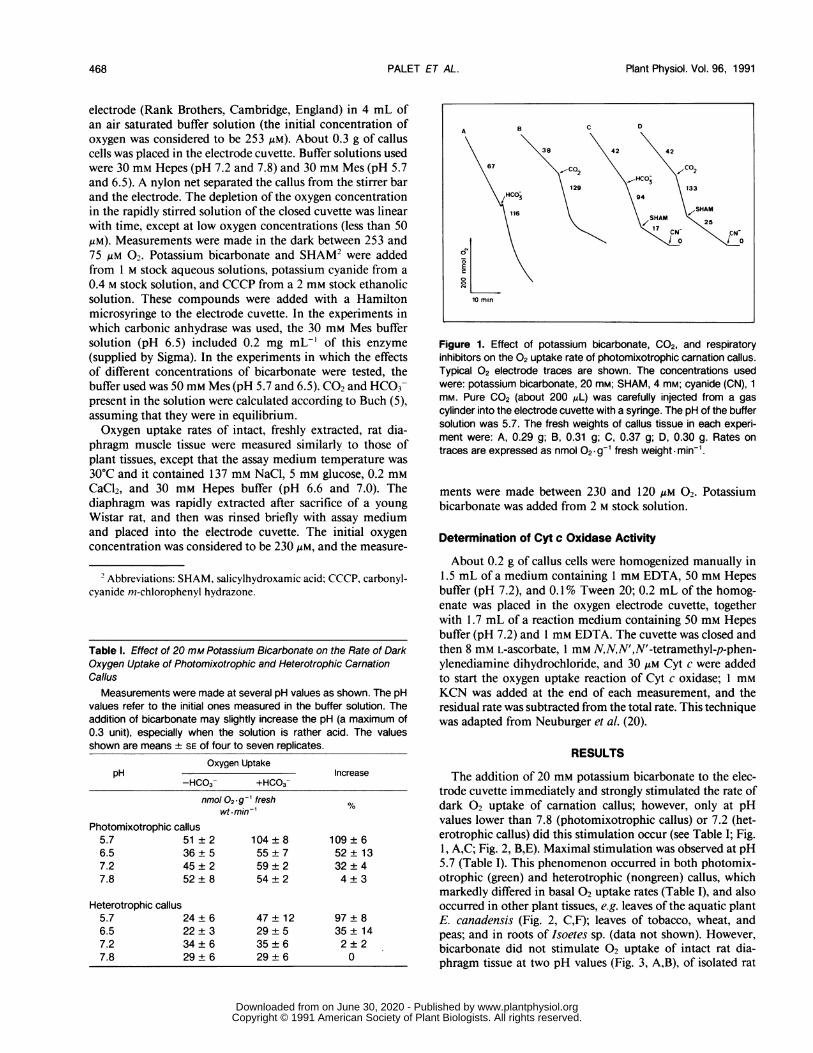
Figure 1. Effect of potassium bicarbonate, C02, and respiratoryinhibitors on the 02 uptake rate of photomixotrophic carnation callus.Typical 02 electrode traces are shown. The concentrations usedwere: potassium bicarbonate, 20 mM; SHAM, 4 mm; cyanide (CN), 1mm. Pure C02 (about 200 AL) was carefully injected from a gas
cylinder into the electrode cuvette with a syringe. The pH of the buffersolution was 5.7. The fresh weights of callus tissue in each experi-ment were: A, 0.29 g; B, 0.31 g; C, 0.37 g; D, 0.30 g. Rates ontraces are expressed as nmol 02- g-1 fresh weight- min-'.
ments were made between 230 and 120 ,sM 02. Potassiumbicarbonate was added from 2 M stock solution.
Determination of Cyt c Oxidase Activity
About 0.2 g of callus cells were homogenized manually in1.5 mL of a medium containing 1 mm EDTA, 50 mM Hepesbuffer (pH 7.2), and 0.1S% Tween 20; 0.2 mL of the homog-enate was placed in the oxygen electrode cuvette, togetherwith 1.7 mL of a reaction medium containing 50 mM Hepesbuffer (pH 7.2) and 1 mm EDTA. The cuvette was closed andthen 8 mM L-ascorbate, 1 mm N,N,N',N'-tetramethyl-p-phen-ylenediamine dihydrochloride, and 30 ,gM Cyt c were addedto start the oxygen uptake reaction of Cyt c oxidase; 1 mM
KCN was added at the end of each measurement, and theresidual rate was subtracted from the total rate. This techniquewas adapted from Neuburger et al. (20).
RESULTS
The addition of 20 mm potassium bicarbonate to the elec-trode cuvette immediately and strongly stimulated the rate ofdark 02 uptake of carnation callus; however, only at pHvalues lower than 7.8 (photomixotrophic callus) or 7.2 (het-erotrophic callus) did this stimulation occur (see Table I; Fig.1, A,C; Fig. 2, B,E). Maximal stimulation was observed at pH5.7 (Table I). This phenomenon occurred in both photomix-otrophic (green) and heterotrophic (nongreen) callus, whichmarkedly differed in basal 02 uptake rates (Table I), and alsooccurred in other plant tissues, e.g. leaves of the aquatic plantE. canadensis (Fig. 2, C,F); leaves of tobacco, wheat, andpeas; and in roots of Isoetes sp. (data not shown). However,bicarbonate did not stimulate 02 uptake of intact rat dia-phragm tissue at two pH values (Fig. 3, A,B), of isolated rat
10 min
PALET ET AL.468
www.plantphysiol.orgon June 30, 2020 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

C02 EFFECTS ON RESPIRATION
Figure 2. Effect of potassium bicarbonate and respiratory inhibitorson dark 02 uptake by photomixotrophic carnation callus (A, B, D, andE) and E. canadensis leaves (C and F). Oxygen uptake was measuredin solution at pH 5.7 (A, B, and C) and at pH 7.8 (D, E, and F). Typical02 electrode traces are shown. The concentrations used were: po-tassium bicarbonate, 20 mM; SHAM, 4 mM; potassium cyanide (CN),1 mm. The fresh weights of tissue in each experiment were: A, 0.53g; B, 0.71 g; C, 0.17 g; D, 0.56 g; E, 0.59 g; F, 0.17 g. Rates ontraces are expressed as nmol 02 -g-1 fresh weight- min-'.
hepatocytes, and ofa blank solution containing only the bufferused (30 mm Mes, pH 5.7 and 6.5) (results not shown).The above mentioned effect was apparently due to the
anion species, because the substitution ofpotassium bicarbon-ate by sodium bicarbonate did not alter the stimulatoryresponse. The substitution of bicarbonate by another anion(e.g. formate, acetate) did not reproduce the effects of bicar-bonate. The only similar compound found that reproducedthe stimulatory effect was gaseous CO2 injected with a syringeinto the cuvette (Fig. 1, B,D).The strong stimulation of 02 uptake by bicarbonate or CO,
Figure 3. Effect of potassium bicarbonate and cyanide (CN) additionson 02 uptake by intact rat diaphragm tissue. Oxygen uptake wasmeasured in solution at pH 7.0 (A) and 6.6 (B). Typical 02 electrodetraces are shown. The fresh weights of tissue were: A, 0.11 g; B,0.07 g. Rates on traces are expressed as nmol 02-g-1 fresh weightmin-'.
lasted for several min, and then decreased gradually (Fig. 1,A,B). The eventual addition of more bicarbonate or CO,again stimulated 02 uptake, suggesting the existence of areversible effect. The stimulatory effect of bicarbonate on 02uptake of callus was higher and lasted longer when carbonicanhydrase was present in the solution (Table II).The stimulatory effect of added bicarbonate on 02 uptake
depended on its concentration in the cuvette, and also on thepH of the medium (Fig. 4A). Theoretical calculations ofestimated concentrations in equilibrium of bicarbonate andCO, in the medium at different pH values suggest that thestimulation of 0, uptake mainly depended on the concentra-tion of free carbon dioxide in the medium (Fig. 4B).The bicarbonate or CO, dependent stimulation of 02 up-
take was fully inhibited by SHAM in carnation callus cells(Fig. 1, C,D; Fig. 2B; Table III, experiment 2), in E. canadensisleaves (Fig. 2C), and in other plant tissues examined (e.g. pealeaves; not shown). This suggests that the mitochondrial al-ternative pathway was mainly involved during the stimulationby bicarbonate or CO2 (see also "va1," in Table IV), being fullyengaged (p = 1; see Table IV). The SHAM-resistant 02 uptakerate, which is an estimate of the activity of the Cyt pathway(v,,,), was normally lower in the presence of bicarbonate thanin its absence (Table III, experiments 1 and 2; Table IV). Apossible negative effect of SHAM on the Cyt path is not likelybecause SHAM, at the concentration used, had usually littleor no effect on 02 uptake in the absence of bicarbonate (Fig.2, A,D; Table III, experiments 1 and 3), and did not affect 02uptake of callus in the presence of bicarbonate when the pHof the solution was 7.8 (Fig. 2, E,F; Table III, experiment 2).We believe that SHAM inhibition may be used under ourconditions as a good indicator of alternative oxidase activity.The uncoupler CCCP greatly stimulated 0, uptake, either
in the absence (Table III, experiment 4) or in the presence ofSHAM (Table III, experiment 3), suggesting adenylate controlof respiration and Cyt pathway. However, CCCP did notsignificantly affect cyanide-resistant respiration rate (TableIII, experiment 5). Estimates of the Cyt and alternative path-ways suggest that CCCP stimulated the activity of both path-ways, but did not significantly affect the capacity of thealternative pathway (Valt; see Tables III and IV). However,bicarbonate strongly inhibited the uncoupled 0, uptake ratein the presence ofSHAM (which is an estimate ofthe capacity
Table II. Effect of Carbonic Anhydrase on the BicarbonateDependent Stimulation of Dark Oxygen Uptake of PhotomixotrophicCarnation Callus
Carbonic anhydrase was added to the electrode cuvette from thestart of the measurements, and used at a final concentration of 0.2mg/mL. The concentration of potassium bicarbonate was 10 mm.The pH of the buffer (20 mm Mes) was adjusted to 6.5. The valuesare means ± SE of five replicates.
Oxygen UptakeTreatment Increase
-HCO3- +HCO3-nmol 02 9-1 fresh
wt-min-
None 42 ± 2 57 ± 3 36 ± 3+Carbonic anhydrase 37 ± 6 66 ± 7 81 ± 11
B.-Tissue
720
\-_35 mM HCO0
602
\ .._-25 mM HCO3
490mM CN0
469
www.plantphysiol.orgon June 30, 2020 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 96, 1991
100o
Q)0
a)C>
Xa)
0
CD)
0
a1)(9)
C)
C:
0)C)
0-m)
50p
0L0 5 1 0 1 5 20 25 30 35
Added HC03- (mM)
5 10 15 20 25
Calculated CO2 (mM)
Figure 4. Effect of added potassium bicarbonate (A) and estimatedCO2 (B) concentrations on dark 02 uptake of photomixotrophic car-nation callus. The pH values of the buffer solutions were 5.7 (0) and6.5 (0). The effect is shown as the percentage of increase in the rateOf 02 uptake. The calculations of the estimated equilibrium concen-trations of CO2 and bicarbonate present in solution at both pH valuesis described in "Materials and Methods." The values shown are means± SE of three to seven replicates. Analysis of variance of the calculateddata indicated that the effect of estimated bicarbonate concentrationon 02 uptake was statistically different (P < 0.001) at pH 5.7 and 6.5(data not shown). However, the effect of estimated CO2 concentrationon 02 uptake was not statistically different (P < 0.05) at both pHvalues (see B).
of the Cyt pathway, Va-,; Table III, experiment 3; Table IV).Thus, bicarbonate addition inhibited both the activity (v",1)and the capacity ( Vat) ofthe Cyt pathway in callus cells (TableIV). Similarly, the addition of potassium bicarbonate alsoinhibited the Cyt path in intact rat diaphragm (Fig. 3, A,B),but in this case no SHAM was previously added becausehigher animal cells do not have the alternative oxidase.
Consistently, the enzymatic activity of Cyt c oxidase ofcallus tissue homogenates was also inhibited by about 40%by short-term exposure to bicarbonate (Table V) or CO2 (datanot shown). Similarly, oxidation of ascorbate/N,N,N',N'-te-tramethyl-p-phenylene diamine dihydrochloride through theCyt pathway was also inhibited by bicarbonate addition inpurified pea leaf mitochondria (our unpublished results).The capacity of the alternative pathway (Valt), which was
estimated as the cyanide-resistant 02 uptake rate, was notsignificantly affected by a short-term exposure to bicarbonate(Table III, experiment 5; Table IV). Longer exposures tobicarbonate also reduced Vait, but less and later than the Cytpathway.
DISCUSSION
The short-term effects of "carbon dioxide" (mainly bicar-bonate and free CO2 in our conditions) on plant tissue respi-
ration at different pH values suggest that external free CO2 isthe species mainly responsible for these effects (Table I). Sincethe stimulatory response of dark oxygen uptake to calculatedfree CO2 (in contrast to bicarbonate) was very similar at thetwo pH values studied (see Fig. 4), this confirms that externalfree CO2 initiated the observed effects on respiration. A similarexperimental approach was used by Jones and Osmond (8) toelucidate that free CO2 was the species utilized by leaf slicesin photosynthesis during 02 electrode measurements. Addi-tional support to this idea derived from the experiments usingcarbonic anhydrase (Table II; see also ref. 8) and from thoseusing injected gaseous C02 (Fig. 1). However, it is difficult toknow which is the active species (free CO2 or bicarbonate)responsible for these effects within the cell, but it is clear thatincubation of carnation callus tissue with radioactive bicar-bonate results in active decarboxylation and fixation rates(results not shown).High carbon dioxide levels in solution induced an imme-
diate and strong reversible stimulation of dark 02 uptake rate,which was due to increased alternative oxidase engagement
Table Ill. Respiration of Photomixotrophic Carnation Callus inResponse to Various Combinations of Inhibitors, Bicarbonate, andUncoupler
All measurements were made at pH 5.7, except experiments 1and 2, which were also made at pH 7.8. The concentrations usedwere: SHAM, 4 mm; cyanide, 1 mM; potassium bicarbonate, 20 mM;CCCP, 1 ,lM. The values are means ± SE of four to eight replicates.
Oxygen UptakeSequential Additions
pH 5.7 pH 7.8
nmol 02.g fresh wt.min1Exp. 1None 72 ± 5 78 ± 11+SHAM 70 ± 5 70 ± 7+Cyanide 6 ± 2 15 ± 3
Exp. 2None 67 ± 5 67 ± 10+Bicarbonate 116 ± 7 67 ± 10+SHAM 42 ± 5 67 ± 10+Cyanide 2 ± 2 7 ± 2
Exp. 3None 67 ± 6+SHAM 62 ± 1+CCCP 101 ± 5+Bicarbonate 49 ± 4+Cyanide 0
Exp. 4None 68 ± 2+CCCP 120 ± 3+Cyanide 70 ± 6+SHAM 0
Exp. 5None 77 ± 3+Cyanide 81 ± 11+CCCP 73 ± 8+Bicarbonate 67 ± 4+SHAM 0
A
.
PALET ET AL.470
www.plantphysiol.orgon June 30, 2020 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

CO2 EFFECTS ON RESPIRATION
(see Table IV). The alternative oxidase was often fully ex-pressed, probably as a result of the inhibition (30-40%) of theCyt pathway activity (vcy1) by high levels of carbon dioxide(Table IV). This behavior is consistent with the model of Bahrand Bonner (2), which predicts that the alternative oxidasecan be engaged when the redox state of the ubiquinone poolis reduced above a certain critical level, and this can easilyoccur when the Cyt pathway is either fully saturated or fully/partially restricted. Recent evidence obtained with a polaro-graphic technique, able to continuously monitor the redoxstate of the ubiquinone pool in isolated mitochondria underseveral conditions (7), confirmed the Bahr and Bonner (2)model. Thus, it seems likely that high levels of carbon dioxidecould increase the reduction level of the ubiquinone pool inorder to fully engage the alternative oxidase. In this sense, theeffects of high carbon dioxide levels on alternative oxidaseexpression can be similar (but not as drastic because theinhibition of the Cyt pathway was always partial) to those ofclassic inhibitors of the Cyt pathway, such as cyanide andhydrogen sulfide (1, 2). Additional support of the inhibitoryeffects of carbon dioxide on the Cyt pathway is obtained bythe strong inhibition of the "capacity" of the Cyt pathway(V,vt; estimated with the use of uncoupler; see Tables III andIV), and by a quantitatively similar inhibition of the enzymicactivity of Cyt c oxidase in callus homogenates (Table V) andpurified pea leaf mitochondria (see above). Interestingly,Miller and Evans (16) also reported inhibition of plant Cyt coxidase activity by sodium bicarbonate, but at higher concen-trations and pH values than in this study.The partial inhibition of the Cyt pathway by high carbon
dioxide probably decreased cellular ATP levels and synthesisrate (unless the higher engagement of the alternative oxidasecan somewhat compensate ATP synthesis during sometime),
Table IV. Estimation of Respiratory Pathways in PhotomixotrophicCarnation Callus
The values have been calculated from the experiments describedin Table ll. vc,4 is the activity of the Cyt pathway, estimated bymeasuring 02 uptake in the presence of SHAM. vm, is the activity ofthe alternative pathway, estimated by measuring the rate of 02 uptakesensitive to SHAM. Vc,y is the capacity of the Cyt path, estimated bymeasuring 02 uptake in the presence of CCCP and SHAM. Valt is thecapacity of the alternative path, estimated by measuring 02 uptakein the presence of cyanide. p is the fraction of the maximum capacityof the alternative path' that is expressed: p = va,t/Vait. The residualrate of 02 uptake resistant to both SHAM and cyanide was takeninto account for calculating vcyt, Vcyt, and Valt. The asterisk means thatthe difference due to the addition of bicarbonate is statistically signif-icant (P < 0.05).
Oxygen Uptake
pH 5.7 pH 7.8
-HCO3- +HCO3- -HCO3 +HCO3-nmol 02-g' fresh wt-min-1
vcyt 62 ± 1 40 ± 5* 54 ± 4 60 ± 11Vait 4 ± 3 74 ± 1* 8 ± 8 0vcyt 101 ± 5 49 ± 4*Va,t 74 ± 6 67 ± 4p <0.1 1
Table V. Effect of Potassium Bicarbonate (10 mM) on Cyt c Oxidase(Cytox) Activity of Photomixotrophic Carnation Callus HomogenatesSee "Materials and Methods" for experimental details. Values are
means ± SE of six replicates. The asterisk means that the differencedue to the addition of bicarbonate is statistically significant (P < 0.05).
Cytox ActivityInhibition
-HCO3 +HC03nmol °2 *9-1 fresh wt *min-' %
817 ± 77 467 ± 23* 38 ± 7
and this should stimulate glycolysis and the use of respiratorysubstrates. Consistently, ATP levels and dark 02 uptake werefound to decrease in epidermal strips of Commelina commu-nis in response to high CO2 levels (26). The strong stimulatoryeffect of the uncoupler CCCP on carnation callus respirationin the absence of inhibitors (Table III) suggested that calluscell respiration was not limited by substrate supply. Thus, itwas not surprising to find a full engagement of alternativeoxidase when Cyt pathway was partially inhibited by eitherfree CO2 or bicarbonate. Under these conditions, the alter-native oxidase functioned as a transient "over-reduction valveor poise" of the electron chain. This can prevent a largereduction level build-up of electron transport componentswhen the Cyt pathway cannot accommodate all the electronsfrom excess substrate oxidation to the demand. Thus, thealternative oxidase seems able to automatically react to rapidchanges in Cyt pathway activity (and not necessarily fullinhibition), as demonstrated by the effects of high carbondioxide levels, to contribute to electron transport "flexibility."When the alternative oxidase is inhibited or not present (suchas in animal cells), then this flexibility is not as evident.
Effects of high CO2 levels on respiratory enzymes otherthan Cyt c oxidase are frequently found in the literature.Inhibitory effects ofC02/bicarbonate have been reported withthe activities of malic enzyme (19), succinate dehydrogenase(3, 27, 31), and both ATP and PPi dependent phosphofruc-tokinases (9, 10). Miller and Hsu (17) suggested a generalsensitivity of mitochondrial dehydrogenases to CO2-bicarbon-ate mixtures. Such widespread effects of carbon dioxide onrespiratory metabolism are not easy to explain from a mech-anistic point of view, but either intracellular pH changes (4,28) or carbamate formation in proteins (14) in response tohigh CO2 might be involved. As Lorimer (14) pointed out,given the ubiquity of carbon dioxide and the facility withwhich carbamates can be formed on proteins, it is reasonableto suppose that this reversible mechanism can be an importantpotential form of regulation of respiration, as it occurs inother well-known systems (e.g. ribulose bisphosphate carbox-ylase-oxygenase and hemoglobin; see ref. 14), which surelydeserves further investigation in plant and animal cells.
ACKNOWLEDGMENTS
We are grateful to Drs. Joe A. Berry, Bert G. Drake, and JosepPefiuelas for useful comments, and to Prof. Jordi Aguila for hissupport and advice. We also thank Dr. Marcello Buiatti for providingus the original callus line, Neus Carb6 for kindly supplying isolated
471
www.plantphysiol.orgon June 30, 2020 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 96, 1991
rat hepatocytes, and Miguel A. Gonzalez-Meler for technical support.This work forms part of the Ph.D. thesis project of Artur Palet.
LITERATURE CITED
1. Azcon-Bieto J, Ribas-Carbo M, Gonzalez-Meler MA, PenuelasJ (1989) Sulfide-resistant respiration in leaves of Elodea can-adensis Michx. Comparison with cyanide-resistant respiration.Plant Physiol 90: 1249-1251
2. Bahr JT, Bonner WD (1973) Cyanide-insensitive respiration. II.
Control of the alternate pathway. J Biol Chem 248: 3446-34503. Bendall DS, Ranson SL, Walker DA (1960) Effects of carbon
dioxide on the oxidation of succinate and reduced diphospho-pyridine nucleotide by Ricinus mitochondria. Biochem J 76:221-225
4. Bown AW (1985) CO, and intracellular pH. Plant Cell Environ8: 459-465
5. Buch K (1960) Dissoziation der kholensaure, gleichgewichte andpuffersysteme. In W Ruhland, ed, Handbuch der Pflanzenphy-siologie, Vol 1. Springer-Verlag, Berlin, pp 1-11
6. Day DA, Arron GP, Christoffersen RF, Laties GG (1978) Effectof ethylene and carbon dioxide on potato tuber metabolism.Plant Physiol 62: 820-825
7. Dry IB, Moore AL, Day DA, Wiskich JT (1989) Regulation ofalternative pathway activity in plant mitochondria: nonlinearrelationship between electron flux and the redox poise of thequinone pool. Arch Biochem Biophys 273: 148-157
8. Jones HG, Osmond CB (1973) Photosynthesis by thin leaf slicesin solution. I. Properties of leaf slices and comparison withwhole leaves. Aust J Biol Sci 26: 15-24
9. Kerbel EL, Kader AA, Romani RJ (1988) Effects of elevated CO2concentrations on glycolysis in intact "Barlett" pear fruit. PlantPhysiol 86: 1205-1209
10. Kerbel EL, Kader AA, Romani RJ (1990) Respiratory and gly-colytic response of suspension-cultured "Passe Crassane" pearfruit cells to elevated CO2 concentrations. J Am Soc Hortic Sci115: 111-114
1 1. Kidd F ( 1915) The controlling influence ofCO2. III. The retardingeffect of carbon dioxide on respiration. Proc R Soc Lond SerB Biol Sci 89: 136-156
12. Kumar PP, Joy IV RW, Thorpe TA (I1989) Ethylene and carbondioxide accumulation, and growth of cell suspension culturesof Picea glauca (white spruce). J Plant Physiol 135: 592-596
13. Lange H (1970) Respiratory pathways in suberin-synthesizingand proliferating potato tuber tissue after derepression. Planta90: 119-132
14. Lorimer GH (1983) Carbon dioxide and carbamate formation:the makings of a biochemical control system. Trends BiochemSci 8: 65-68
15. McCaig TN, Hill RD (1977) Cyanide-insensitive respiration in
wheat: cultivar differences and effects of temperature. carbondioxide, and oxygen. Can J Bot 55: 549-555
16. Miller GW, Evans HJ (1956) Inhibition of plant cytochromeoxidase by bicarbonate. Nature 178: 974-976
17. Miller GW, Hsu WJ (1965) Effects of carbon dioxide-bicarbon-ate mixtures on oxidative phosphorylation by cauliflower mi-tocondria. Biochem J 97: 615-619
18. Murashige T, Skoog F (1962) A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol Plant 15:473-497
19. Neuburger M, Douce R (1980) Effect of bicarbonate and oxal-oacetate on malate oxidation by spinach leaf mitochondria.Biochem Biophys Acta 589: 176-189
20. Neuburger M, Journet EP, Bligny R, Carde JP, Douce R (1982)Purification of plant mitochondria by isopycnic centrifugationin density gradients of Percoll. Arch Biochem Biophys 217:312-333
21. Nobel PS, Palta JA (1989) Soil 02 and CO2 effects on rootrespiration of cacti. Plant Soil 120: 263-271
22. Osmond CB, Smith SD, Gui-Ying B, Sharkey TD (1987) Stemphotosynthesis in a desert ephemeral, Eriogonum intflatum.Characterization of leaf and stem CO, fixation and H2O vaporexchange under controlled conditions. Oecologia 72: 542-549
23. Palet A, Azc6n-Bieto J, Aguila J (1988) Stimulation of respira-tion and alternative oxidase activity by bicarbonate in carna-tion calli (abstract No. 893b). Plant Physiol 86: S- 150
24. Perez-Trejo MS, Janes HW, Frenkel C (1981) Mobilization ofrespiratory metabolism in potato tubers by carbon dioxide.Plant Physiol 67: 514-517
25. Reuveni J, Gale J (1985) The effect of high levels of carbondioxide on dark respiration and growth of plants. Plant CellEnviron 8: 623-628
26. Shaish A, Roth-Bejerano N, Itai C (1989) The response ofstomata to CO2 relates to its effects on respiration and ATPlevel. Physiol Plant 76: 107-111
27. Shipway MR, Bramlage WJ (1973) Effects of carbon dioxide onactivity of apple mitochondria. Plant Physiol 51: 1095-1098
28. Siriphanich J, Kader AA (1986) Changes in cytoplasmic andvacuolar pH in harvested lettuce tissue as influenced by CO,.J Am Soc Hortic Sci 111: 73-77
29. Smock RM (1979) Controlled atmosphere storage of fruits. Hor-tic Rev 1: 301-336
30. Willmer CM, Hamilton K, Pallas JE Jr (1983) The use of anoxygen electrode for monitoring photosynthesis and respira-tion of leaves. In H Metzner, ed, Photosynthesis and PlantProductivity. Wissenschaftliche Verlagsgesellschaft mbH,Stuttgart, pp 178-181
31. Young RE, Romani RJ, Biale JB (1962) Carbon dioxide effectson fruit respiration. II. Response of avocados, bananas, andlemons. Plant Physiol 37: 416-422
472 PALET ET AL.
www.plantphysiol.orgon June 30, 2020 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.