Embed Size (px)
Citation preview

1
Short Title: Vascular plant-Mucoromycotina fungal symbiosis 1
2
* Corresponding author: Katie J. Field ([email protected]); Tel: +44 (0)113 343 2849 3
4
Mucoromycotina fine root endophyte fungi form nutritional mutualisms with vascular 5
plants 6
7
Grace A. Hoysted1, Alison S. Jacob2,3, Jill Kowal4, Philipp Giesemann5, Martin I. Bidartondo2,3, 8
Jeffrey G. Duckett4, Gerhard Gebauer5, William R. Rimington2,3,4, Sebastian Schornack6, Silvia 9
Pressel4, and Katie J. Field1* 10
11
1 Centre for Plant Sciences, Faculty of Biological Sciences, University of Leeds, Leeds, LS2 12
9JT, UK 13
2 Comparative Plant & Fungal Biology, Royal Botanic Gardens, Kew, Richmond TW9 3DS, 14
UK 15
3 Department of Life Sciences, Imperial College London, London, SW7 2AZ, UK 16
4 Department of Life Sciences, Natural History Museum, London SW7 5BD, UK 17
5 Laboratory of Isotope Biogeochemistry, Bayreuth Center of Ecology and Environmental 18
Research (BayCEER), University of Bayreuth, Bayreuth, Germany 19
6 Sainsbury Laboratory, University of Cambridge, Cambridge, CB2 1LR 20
21
One sentence summary: Mucoromycotina fine root endophyte fungi transfer soil nutrients, 22
notably N, to lycophyte host plants in exchange for photosynthetically-fixed C and colonise 23
diverse neighbouring vascular plants. 24
25
Author contributions 26
K.J.F., S.P., S.S., M.I.B., and J.G.D. conceived and designed the investigation. S.P., J.K., 27
J.G.D., M.I.B., A.S.J., G.G., and G.A.H collected plant material. G.A.H. and K.J.F. undertook 28
Plant Physiology Preview. Published on July 29, 2019, as DOI:10.1104/pp.19.00729
Copyright 2019 by the American Society of Plant Biologists
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

2
the physiological analyses. A.S.J., W.R.R., and M.I.B. undertook the molecular anlayses. S.P. 29
undertook the cytological analyses with assistance from J.K. P.G. and G.G. analysed and 30
interpreted the stable isotope data. G.A.H. led the writing, all authors discussed results and 31
commented on the manuscript. K.J.F. agrees to serve as the author responsible for contact 32
and ensure communication. 33
34
Abstract 35
Fungi and plants have engaged in intimate symbioses that are globally widespread and have 36
driven terrestrial biogeochemical processes since plant terrestrialisation >500 Mya. Recently, 37
hitherto unknown nutritional mutualisms involving ancient lineages of fungi and non-vascular 38
plants have been discovered, although their extent and functional significance in vascular 39
plants remain uncertain. Here, we provide evidence of carbon-for-nitrogen exchange between 40
an early-diverging vascular plant (Lycopodiella inundata L.) and Mucoromycotina 41
(Endogonales) fine root endophyte fungi. Furthermore, we demonstrate that the same fungal 42
symbionts colonize neighbouring non-vascular and flowering plants. These findings 43
fundamentally change our understanding of the physiology, interrelationships, and ecology of 44
underground plant–fungal symbioses in modern terrestrial ecosystems by revealing the 45
nutritional role of Mucoromycotina fungal symbionts in vascular plants. 46
47
Key Words 48
Arbuscular mycorrhizas, Endogonales, lycophytes, nitrogen, carbon. 49
50
Introduction 51
Plant terrestrialisation >500 Mya (Morris et al., 2018) was facilitated by the formation of 52
mutualistic symbioses with fungi, through which the earliest plants gained access to mineral 53
nutrients in exchange for photosynthetically-fixed carbon (C). It was long hypothesised that 54
this ancient mycorrhiza-like symbiosis was closely related to, and subsequently evolved into, 55
widespread modern-day arbuscular mycorrhizas (AM) formed with plant roots by 56
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

3
Glomeromycotina fungi (Pirozynski and Malloch, 1975; Redecker et al., 2000). However, 57
recent molecular, cytological, physiological, and paleobotanical evidence has strongly 58
indicated that early fungal associates were likely to be more diverse than has previously been 59
assumed (Bidartondo et al., 2011; Field et al., 2015a; Field et al., 2015b). Members of the 60
earliest diverging clade of an ancient land plant lineage, Haplomitriopsida liverworts, are now 61
known to form mycorrhiza-like associations with Mucoromycotina fungi (Bidartondo et al., 62
2011; Field et al., 2012; Field et al., 2015b) which also colonise other early diverging plant 63
lineages, namely hornworts, lycophytes, and ferns, sometimes co-occurring with 64
Glomeromycotina fungi in the same plant host (Desirò et al., 2013; Rimington et al., 2015). 65
Mucoromycotina represents an ancient fungal lineage considered to branch earlier than or 66
sister to the Glomeromycotina AM fungi (James et al., 2006; Lin et al., 2014). The recent 67
identification of Mucoromycotina in a range of modern non-vascular plants (Bidartondo et al., 68
2011) and plant fossils (Krings et al., 2007; Strullu‐Derrien et al., 2014) supports the idea that 69
the colonisation of Earth’s land masses by plants was facilitated not only by Glomeromycotina 70
AM, but also by Mucoromycotina fungal symbionts (Field et al., 2015a). The latest discoveries 71
of putative Mucoromycotina fungi in vascular land plants (Rimington et al., 2015; Rimington et 72
al., 2016; Orchard et al., 2017a) indicate that root symbiotic versatility and diversity (Hoysted 73
et al., 2018) has been grossly underestimated across extant plants. 74
Although Mucoromycotina fungal symbioses in non-vascular plants have received the 75
most attention to date, there are now several reports of their occurrence in vascular plants 76
(Rimington et al., 2015; Rimington et al., 2016; Orchard et al., 2017a; Orchard et al., 2017b; 77
Hoysted et al., 2018). It has been suggested that the globally widespread, arbuscule-forming 78
fine root endophytes (FREs) classified as Glomus tenue (or more recently Planticonsortium 79
tenue (Walker et al., 2018)), which occur across a wide range of vascular plant groups 80
(Rimington et al., 2015; Orchard et al., 2017b), are closely related to the Mucoromycotina 81
fungal symbionts of non-vascular plants (Hoysted et al, 2018; Field and Pressel, 2018). If true, 82
there could be major ramifications for our understanding of the past and present diversity and 83
function of plant-fungal nutritional symbioses (Field and Pressel, 2018), suggesting 84
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

4
Mucoromycotina fungal symbiosis is not limited to ancient plant lineages but is in fact 85
widespread throughout extant land plants. However, it remains unclear whether the putative 86
Mucoromycotina FREs detected in vascular plants to date are comparable in terms of function 87
and identity to the mutualistic Mucoromycotina fungal symbionts detected in non-vascular 88
plants. 89
As lycophytes are considered to be the earliest divergent extant vascular plant lineage 90
(Kenrick and Crane, 1997), the discovery of non-Glomeromycotina fungal associates in 91
lycophyte roots and gametophytes is particularly important. For over 100 years, fungal 92
associations in lycophytes have been thought of as being AM-like but with unique “lycopodioid” 93
features (Duckett and Ligrone, 1992; Schmid and Oberwinkler, 1993). However, global 94
analysis of fungal associates in 20 lycophytes (Rimington et al., 2016) has now shown their 95
colonisation is broadly similar to that of hornworts (Desirò et al., 2013), with many species 96
forming single and/or dual associations with both Glomeromycotina arbuscular mycorrhizal 97
(AM) fungi and Mucoromycotina FRE fungi (Rimington et al., 2016). Remarkably, every 98
sample of Lycopodiella inundata - a species found in wet habitats across the Northern 99
Hemisphere - examined so far appears colonised exclusively by Mucoromycotina FRE fungi 100
(Rimington et al., 2016). The fundamental obstacle to studying function of Mucoromycotina 101
FREs has been finding any plants that are not co-colonized by coarse root endophytes (i.e., 102
Glomeromycotina AM fungi). In fact, so far there is no evidence of nutritional mutualism 103
between any vascular plant and Mucoromycotina FREs (Hoysted et al., 2018). Therefore, L. 104
inundata provides a unique and important opportunity to dissect the symbiotic function of 105
FREs in a vascular plant. 106
Here, we investigate the function, cytology, and occurrence of the fungal associates of 107
L. inundata (Figure 1a, b). We use a combination of radio- and stable isotope tracers 108
(Supplemental Figure S1, S2) to test physiology and functioning of vascular plant-109
Mucoromycotina fungal symbioses, detailed cytological analyses to characterise morphology 110
and colonisation patterns of Mucoromycotina fungi, and molecular techniques to identify the 111
fungal associates of a range of vascular plants (including those used in our experiments) 112
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

5
across several field sites. Furthermore, we used natural abundance 13C and 15N signatures 113
to test under field conditions whether Mucoromycotina fungal association affects the direction 114
of carbon and nitrogen fluxes between plants and associated fungi (Gebauer and Meyer, 115
2003). Specifically, we address the following questions: 116
(1) What is the function of Mucoromycotina fungal associations in lycophytes in terms 117
of carbon-for-nutrient exchange? 118
(2) Are there characteristic cytological signatures or features of Mucoromycotina 119
fungal associations in L. inundata and other vascular plants? 120
(3) Do Mucoromycotina fungal symbionts of L. inundata co-occur in neighbouring 121
angiosperm roots and non-vascular plants? 122
123
Results 124
Mucoromycotina-L. inundata symbioses are nutritional mutualisms 125
Isotope tracing experiments conducted in controlled environment chambers confirmed 126
carbon was transferred from field-collected L. inundata to extraradical hyphae of symbiotic 127
Mucoromycotina FRE fungi (Figure 2). Mucoromycotina fungal symbiont identity was 128
confirmed using sequencing of the fungal 18S ribosomal rRNA gene with the broad specificity 129
fungal primer set NS1/EF3 and a semi-nested approach with Mucoromycotina- and 130
Glomeromycotina-specific primers described in Desirò et al., (2013) (Supplemental Figure 131
S3). Plants transferred an average of 79 g ( 49.3 SE) of recent photosynthate to the external 132
fungal hyphal mycelium within the microcosm during the labelling period (Fig. 2a). This 133
represents 0.28% ( 0.14 SE) of the total amount of carbon that was fixed during the labelling 134
period by L. inundata (Fig. 2b). 135
Mucoromycotina fungi within the experimental microcosms transferred between 3-9% 136
of the supplied 33P tracer and 0.6-1% of the supplied 15N tracer to their plant hosts during the 137
isotope labelling period (Figure 2c, d). Mucoromycotina FREs transferred significantly more 138
15N than 33P to L. inundata both in terms of absolute quantities (Figure 2c, P = 0.05; Student’s 139
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

6
T-Test) and when normalised to plant biomass (Figure 2d, P = 0.03, Student’s T-Test). 140
To test the potential nutritional role of Mucoromycotina fungi in L. inundata in the field, 141
we analysed the natural abundance 13C and 15N stable isotope signatures of leaves and roots 142
of L. inundata and Juncus bulbosus, both of which were shown to host Mucoromycotina FREs 143
in their roots (Supplemental Figure S4-S9). In addition, five plant species representing three 144
different types of mycorrhizal associations were sampled to serve as reference plants: two 145
ericoid mycorrhizal species (Erica tetralix, collected on six plots; Calluna vulgaris, collected on 146
three plots), two ectomycorrhizal species (Pinus sylvestris and Betula pendula seedlings, both 147
from one plot) and one arbuscular mycorrhizal species (Molinia caerulea from six plots). 148
All leaf δ13C values from plants collected from the same site as those collected for our 149
isotope tracing experiments ranged between -26.2 and -30.1 ‰ and root δ13C values between 150
-24.5 and -28.9 ‰, while leaf δ15N values ranged from 3.3 to -10.0 ‰ and root δ15N values 151
from 3.1 to -5.9 ‰ (Figure 3). Leaves of the three groups, L. inundata (n = 6), Juncus bulbosus 152
(n = 6) and reference plants, five species representing three types of mycorrhizal associations, 153
i.e. arbuscular, ericoid and ectomycorrhizal species (n = 17), also collected from the same site 154
as plants used in our isotope tracing experiments, were significantly different in δ13C (H(2) = 155
8.758; P = 0.013) and δ15N (H(2) = 21.434; P < 0.001, Figure 3a). L. inundata leaves were 156
significantly depleted in 13C compared to J. bulbosus leaves (Q = 2.644, P < 0.05) and a 157
significant depletion of L. inundata leaves compared to reference plant leaves (Q = 2.662, P 158
< 0.05, Figure 3a) was found. The J. bulbosus leaves were not significantly different from 159
reference plants in δ13C. No significant difference was discovered for δ15N in L. inundata and 160
J. bulbosus leaves (Q = 1.017, P > 0.05), while leaves of both species were significantly 161
enriched in 15N compared to the reference plants (Q = 2.968, P < 0.05; Q = 4.205, P < 0.05, 162
Figure 3a). For the roots, only δ15N showed significant differences between the three groups 163
under comparison (F(2) = 34.815; P < 0.001, Figure 3b). The L. inundata and J. bulbosus 164
roots were not significantly distinguished in δ15N, however, roots of both species were 165
significantly enriched in 15N compared to reference plant roots (q = 10.109, P < 0.001; Q = 166
8.515, P < 0.001, Figure 3b). 167
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

7
168
Mucoromycotina fine root endophytes of L. inundata show distinctive cytology 169
Trypan blue staining and SEM of wild-collected plants (a liverwort, two grasses, and a 170
rush) from the same site as the L. inundata plants used in our isotope tracer and stable isotope 171
studies (except for the grass Holcus lanatus, see Supplemental Table S1), revealed two 172
distinct fungal symbiont morphologies. These consisted of either coarse hyphae (>3 µm 173
diameter) and large vesicles (>20 µm diameter) or fine branching hyphae (<2 µm diameter) 174
with small swellings/vesicles (usually 5-10 but up to 15 µm diameter) (Figures 4-5). Both 175
morphologies were regularly observed, often co-occurring in the same sample, in the 176
gametophyte of the liverwort Fossombronia foveolata (Figures 4a, b, 5a; Supplemental Figure 177
S10a), in the roots of the grasses Holcus lanatus (Figure 4f) and Molinia caerulea (Figure 4g, 178
h), and the rush Juncus bulbosus (Figure 5h, i). In the colonised roots of both freshly collected 179
L. inundata and those incubated in growth chambers, only fine hyphae with small 180
swelling/vesicles were invariably detected (Figures 4c-e, 5f, g). As in the other plants 181
analysed, these fine hyphae were aseptate and formed both intercalary and terminal 182
swellings/vesicles but, in contrast to the grasses (Supplemental Figure S10b), never 183
arbuscules. Similar fungal morphology was also observed in protocorm cells of newly 184
developing sporophytes (Figure 5b, c) and in gametophytes of L. inundata (Supplemental 185
Figure S11). However, in these early developmental stages, fungal colonization consistently 186
exhibits a distinct zonation: an outer intracellular zone and a more central, strictly intercellular 187
zone (Figure 5d, e; Supplemental Figure S11b-g). In the intracellular zone, fungal colonization 188
is the same as in the sporophyte roots and consists of fine hyphae with intercalary and terminal 189
swellings/vesicles (Figure 5b, c; see also Supplemental Figure S11i). Unique to the 190
gametophyte generation, in the outermost cortical layers, the fungus also forms tightly wound 191
coils (hyphae up to 2.5 µm in diameter) with larger vesicles (15-20 µm) (Supplemental Figure 192
S11d), as described before in Lycopodium clavatum (Schmid and Oberwinkler, 1993). Both 193
gametophyte and early developmental stages of the sporophyte generation develop a 194
conspicuous central system of large, mucilage-filled intercellular spaces. In this region, the 195
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

8
fungus becomes strictly intercellular (Figure 5d, e; Supplemental Figures S11e-g). The 196
intercellular hyphae are initially fine and with small swellings/vesicles (Figure 5d, 197
Supplemental Figure S11e), as their intracellular counterparts, but soon enlarge and 198
eventually reach diameters in excess of 3 µm (Supplemental Figure S11f), with no 199
swellings/vesicles present at this stage. 200
201
Mucoromycotina fungal symbionts are shared by neighbouring angiosperms 202
Analysis of L. inundata plants collected from the same site, at the same time as plants 203
used in the above investigations, confirmed that they were colonised by Mucoromycotina 204
fungi. Glomeromycotina sequences were not detected. Mucoromycotina OTUs were detected 205
before and after the experiments (Supplemental Table S2); these same OTUs had previously 206
been identified in wild-collected lycophytes from diverse locations (Rimington et al., 2015). 207
Diverse and shared Mucoromycotina fungi OTUs were detected in wild L. inundata, 208
liverworts, and angiosperms growing adjacently in the same UK locations (Supplemental 209
Table S2, Supplemental Figures S2-8) in the following combinations: L. inundata, F. foveolata, 210
M. caerulea, and J. bulbosus (Thursley Common, Surrey); L. inundata, F. foveolata, and J. 211
bulbosus (Norfolk); F. foveolata and H. lanatus (Lynn Crafnant, Wales). Mucoromycotina 212
OTUs were also detected in L. inundata from Studland Heath, Dorset. 213
214
Discussion 215
Our results show that the symbiosis between L. inundata and Mucoromycotina FREs are 216
nutritionally mutualistic, with the fungus gaining plant-fixed C and the plant gaining fungal-217
acquired P and N (Figure 2a-d). Cytological analyses of the fungus colonising the roots of L. 218
inundata revealed a characteristic morphology consisting invariably of fine, aseptate 219
branching hyphae with terminal and intercalary swellings/vesicles. This morphology matches 220
that described previously in a range of angiosperms colonized by FREs (Orchard et al., 2017a; 221
Orchard et al., 2017b) and here in grasses, a rush, and a liverwort, all harbouring fungi 222
identified molecularly as Mucoromycotina (Supplemental Figure S3). Our results provide 223
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

9
compelling evidence for Mucoromycotina FREs being shared by plants occupying key nodes 224
in the land plant phylogeny - from early liverworts and vascular lycophytes to the later diverging 225
angiosperms - and demonstrate that this association represents a nutritional mutualism as 226
much in vascular plants as it does in non-vascular plants (Field et al., 2015b; Field et al., 227
2016). 228
Our findings raise important questions regarding the ecology and evolution of 229
mycorrhizal associations and the nature of widespread Mucoromycotina FRE fungal 230
symbioses, chief among which is how have these associations persisted and why are they so 231
widespread today? We can now begin to address this with the demonstration that a vascular 232
plant assimilates relatively large amounts of 15N tracer via its Mucoromycotina fungal symbiont 233
when compared to the Mucoromycotina fungal-acquired 33P tracer (Fig. 2c), suggesting a 234
potential role for Mucoromycotina FREs in vascular plant nitrogen uptake, complementary to 235
the role of Glomeromycotina AM fungi (Field et al., 2019). This nutritional role could help to 236
explain the persistence of Mucoromycotina FREs across nearly all modern land plant lineages. 237
238
Costs and benefits of hosting Mucoromycotina fungi 239
Our data demonstrate that L. inundata transfers carbon to symbiotic Mucoromycotina 240
FRE (Figure 2a, b). However, when compared to other vascular plants with Glomeromycotina 241
AM fungal associates in similar experimental systems (Field et al., 2012), it is clear that the 242
relative C “cost” of maintaining Mucoromycotina fungal symbionts in L. inundata is at least on 243
a par with, if not greater than, that of maintaining Glomeromycotina fungi. It is not entirely clear 244
from our experiments why this might be and it is important to note that the C “cost” of hosting 245
Mucoromycotina FREs has only been tested in one vascular plant species to date and thus 246
represents an important area for future research. It is possible that the carbon-for-nutrient 247
exchange dynamics between plant and Mucoromycotina FREs varies according to plant and 248
fungal identity, in addition to abiotic factors, as it does for Glomeromycotina AM (Field and 249
Pressel, 2018). 250
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

10
Lycophytes represent a critical node in land plant phylogeny, widely considered as a 251
diversification point in the mid-Paleozoic (480-360 Ma) characterised by the evolution of roots, 252
leaves, stomata, and associated vasculature (Kenrick and Crane, 1997). Given that all the 253
plants sampled grew in close proximity and all follow the C3 photosynthetic pathway, the trend 254
for lower δ13C values in root tissues versus leaves (Figure 3a, b) is most likely caused by 255
systematic differences in 13C abundance in photosynthetic vs. non-photosynthetic tissues 256
(Gebauer and Schulze, 1991; Cernusak et al., 2009). The depletion of 13C observed in the 257
leaves of L. inundata (Figure 3a) relative to the other, non-Mucoromycotina associated plants 258
sampled is unlikely to be related to C gains from its Mucoromycotina fungal symbiont (Bago 259
et al., 2000) as one would expect (myco)heterotrophic carbon gains to result in 13C enrichment 260
(Press et al., 1987; Schulze et al., 1991; Gebauer and Meyer, 2003); rather, it may indicate 261
that L. inundata leaves regulate their stomata differently from J. bulbosus or the reference 262
plants tested, as δ13C in tissues of terrestrial plants is, among other factors, driven by the 263
water use efficiency of the plant (Farquhar et al., 1982; Farquhar et al., 1989). 264
Alongside increased capacity for regulation of water relations and C capture and 265
fixation, we hypothesise that the increasing size and structural complexity of land plants 266
across the land plant phylogeny and evolutionary time (Field et al., 2012) result in greater 267
plant nutrient demand. Glomeromycotina AM are associated with facilitation of plant P uptake 268
and occur commonly in soils with low P availability (Smith et al., 2015; Albornoz et al., 2016). 269
The amount of 33P transferred to L. inundata plants in our experiments was much less than 270
has previously been recorded for Mucoromycotina-associated liverworts (Field et al., 2016) or 271
for Glomeromycotina-associated ferns and angiosperms (Field et al., 2012) despite the same 272
amount of 33P being made available in comparable experimental systems. This suggests 273
Mucoromycotina fungi may not play a critical role in lycophyte P nutrition. Our results contrast 274
with the view that Mucoromycotina FREs enhance plant P uptake, at least in soils with very 275
low P (Ryan and Kirkegaard, 2012; Orchard et al., 2017b). Previous experiments with 276
Mucoromycotina fungi-associated liverworts suggest that in addition to supplying host plants 277
with P, Mucoromycotina fungal associates also play a role in plant N nutrition (Field et al., 278
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

11
2015b; Field et al., 2016; Field et al., 2019). 279
Nitrogen is an essential element for plants that is available in soils in plant-inaccessible 280
organic forms and as plant-accessible inorganic nitrate and ammonium (Krapp, 2015). Our 281
results show that Mucoromycotina FRE symbionts transfer significantly more 15N tracer 282
compared to 33P (Figure 2c, d). Up to 145 times more 15N was transferred to L. inundata (0.3-283
1% of the supplied tracer) than to Haplomitriopsida liverworts in comparable experiments 284
which assimilated between 0.05 and 0.2 % of the supplied tracer (Field et al., 2016). Using 285
analysis of natural abundance 15N signatures, we show that Mucoromycotina FRE-associated 286
L. inundata and J. bulbosus were 15N enriched compared to co-occurring reference plants with 287
different mycorrhizal fungal partners (Figure 3). This 15N enrichment could be caused by 288
temporal and spatial variations in N availability and changes in plant N demand over time 289
(Gebauer and Meyer, 2003). However, these factors are unlikely in our case, because all 290
plants sampled from this field collection grew in close spatial proximity and were collected at 291
the same time. Presence of multiple N sources with distinct isotopic values and their utilization 292
by different mycorrhizal associations are known as additional drivers of variations in plant 15N 293
isotope abundance (Bidartondo et al., 2004). While this distinction in N isotope abundance 294
between plants with different mycorrhizas is almost or completely lost in conditions of high N 295
availibility (Gebauer and Meyer, 2003), it may become prominent under severe N limitation 296
(Schulze et al., 1994). Given that our plants were collected at a heathland field site which was 297
likely N-limited (von Oheimb et al., 2010) despite substantial atmospheric N deposition, the 298
separation of L. inundata and J. bulbosus in their natural abundance 15N from neighbouring 299
plants with other fungal associations supports our hypothesis that plants hosting 300
Mucoromycotina symbionts benefit from fungal-acquired N. 301
Some Glomeromycotina AM fungi transfer N to their associated hosts (Farquhar et al., 302
1989); however, the ecological relevance of AM-facilitated N uptake is widely debated, in 303
particular the amounts of N transferred to hosts compared to the overall N requirements of the 304
plant (Smith and Smith, 2011). Exclusive plant-Mucoromycotina FRE symbioses seem to be 305
rare, having been reported before only in the earliest-diverging Haplomitriopsida liverworts 306
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

12
(Field et al., 2015a; Field et al., 2015b), while all other plants, including other lycophytes 307
(Rimington et al., 2015), that form associations with these fungi appear able to do so also with 308
Glomeromycotina, often simultaneously (Rimington et al., 2015). It is possible that the large 309
input to Lycopodiella N-nutrition and minor contribution to P-nutrition by Mucoromycotina 310
FREs reflect a specialised relationship, particulary pertinent when considering heathland 311
habitats have very low plant-available N. Nevertheless, our present data combined with 312
previous demonstrations of N transfer in liverwort-Mucoromycotina symbioses (Field et al., 313
2015b; Field et al., 2016) and emerging evidence that Mucoromycotina FREs, but not 314
Glomeromycotina AM fungi, are able to transfer N to host liverworts from organic sources 315
(Field et al. 2019), all point to a critical role of Mucoromycotina FREs in host plant N nutrition. 316
Indeed, our cytological analyses show that, differently from Lycopodiella roots where only fine 317
endophytes were observed (Figures 4, 5; Table 1), all other co-occurring plants (F. foveolata, 318
J. bulbosus, and M. caerulea) were also colonised by coarse endophytes with cytology typical 319
of Glomeromycotina (Figures 4, 5; Table 1). The finer functional details, in terms of N and P 320
transfer, of this partnership in other vascular plants from a broader range of habitats remain 321
to be established; the challenge here will be to separate the nutritional contributions of 322
Mucoromycotina FREs and Glomeromycotina to host plants that are co-colonized by both 323
fungi (in addition to the contributions made by any mutualistic Ascomycetes and 324
Basidiomycetes) as that seems to be the prevailing condition in vascular plants, especially 325
angiosperms. 326
327
Mucoromycotina fine root endophytes 328
Mucoromycotina fungi within Endogonales colonising the gametophytes of liverworts 329
(F. foveolata) and lycophytes (L. inundata), the sporophytic protocorms and roots of 330
lycophytes (L. inundata), and the roots of angiosperms (J. bulbosus, M. caerulea, and H. 331
lanatus), all display the same characteristic morphology attributed previously to FREs 332
(Orchard et al., 2017b; Walker et al., 2018). This contrasts with that typical of 333
Glomeromycotina AM fungal associations, consisting of coarse hyphae (>3 µm diameter) and 334
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

13
larger vesicles, which we observed in Fossombronia, Juncus, Molinia, and Holcus but not in 335
L. inundata (Table 1). These observations, together with the molecular identification of 336
Mucoromycotina clades shared by these phylogenetically distant plant lineages presented 337
here, support previous suggestions that vascular plants’ FREs are closely related to the 338
Mucoromycotina mycorrhizal-like symbionts of non-vascular plants (Rimington et al., 2016). 339
Here, we show that the same Mucoromycotina FREs have the capacity to be nutritionally-340
mutualistic across different land plant phyla. 341
Our demonstration of an extensive intercellular phase of fungal colonisation in the 342
gametophytes and protocorms of L. inundata is in line with other lycophytes (Schmid and 343
Oberwinkler, 1993; Rimington et al., 2015) and strongly recalls the gametophytes of the 344
Haplomitriopsida liverwort Treubia (Duckett et al., 2006) and several hornworts (Desirò et al., 345
2013), all of which have also been shown to associate with Mucoromycotina fungi (Bidartondo 346
et al., 2011; Desirò et al., 2013). Differently from their fine intracellular counterparts, 347
intercellular hyphae become swollen, eventually reaching more than 3 µm in diameter. Tightly 348
wound hyphal coils up to 2.5 µm in diameter with somewhat larger terminal vesicles (up to 20 349
µm in diameter) are also prominent in the outer cortical layers of L. inundata gametophytes 350
but were not observed in either protocorms or roots. Thus, Mucoromycotina FREs display 351
considerable phenotypic plasticity in their interactions with diverse lineages of land plants 352
which appears to relate to the developmental stage of the host and whether it produces an 353
extensive network of mucilage-filled intercellular spaces. The putative occurrence of 354
Mucoromycotina FREs in early land plants and their presence in both extant early and later 355
diverging plant lineages now point to a prominent role of these fungi, not only in plant 356
terrestrialization (Field et al., 2015a), but also in current ecosystem functioning. Indeed, 357
Mucoromycotina FREs have been shown to occur worldwide across many ecosystems, 358
particularly in the roots of crop and pasture species, where colonization levels may be high, 359
even as dense as the biomass of coarse Glomeromycotina arbuscular mycorrhizal fungi 360
(Orchard et al., 2017b). 361
362
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

14
Conclusions 363
More ammunition for the mycorrhizal revolution 364
Our findings provide conclusive evidence that Mucoromycotina FREs form nutritional 365
mutualisms not only with non-vascular liverworts (Field et al., 2015b; Field et al., 2016), but 366
also with a vascular plant. We have found that the Mucoromycotina FRE associates of L. 367
inundata receive up to 189 times more photosynthesis-derived C from the plant than the 368
Mucoromycotina fungal associates of non-vascular plants (Field et al., 2016). In return, L. 369
inundata hosts receive a relatively large amount of N from their Mucoromycotina FRE partners 370
- ca. 145 times more than non-vascular plants receive from their Mucoromycotina fungal 371
symbionts (Field et al., 2016). Together with our discovery that the same Mucoromycotina 372
fungal symbionts are shared with neighbouring grasses, rushes and liverworts, and recent 373
findings of functional complementarity between Mucoromycotina FREs and Glomeromycotina 374
AM (Field et al., 2019), our findings point towards a unique physiological niche for the 375
persistence of Mucoromycotina fungi, both in single and dual colonisations with 376
Glomeromycotina AM. 377
378
Materials and Methods 379
Plant material and growth conditions 380
Lycopodiella inundata (L.), neighbouring angiosperms (the grasses Holcus lanatus, 381
Molinia caerulea, and the rush Juncus bulbosus), and a liverwort (Fossombronia foveolata) 382
were collected from Thursley National Nature Reserve, Surrey, UK (SU 90081 39754), a 383
heathland site, in June 2017. The L. inundata plants were planted directly into pots (90 mm 384
diameter x 85 mm depth) containing a homogenous mixture of acid-washed silica sand and 5 385
% pot volume compost (Petersfield No.2, Leicester, UK) to aid water-retention properties of 386
the substrate and to provide minimal nutrients. Soil surrounding plant roots was left intact to 387
prevent damage to the roots and to act as a natural inoculum, including symbiotic fungi and 388
associated microorganisms. Pots were weeded regularly to remove other plant species. All 389
plants used throughout this investigation were collected from the wild. Each microcosm was 390
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

15
a homogenised mixture of acid-washed sand and soil collected from around the plant roots 391
from the collection site. All plants used in this investigation (isotope tracing, cytology, and 392
stable isotope studies) were collected from Thursley Nature Reserve, with additional plants 393
from three other UK field sites (Supplemental Table S1), used for additional cytological and 394
molecular analyses. 395
Based on the methods of Field et al. (2012; 2015a), three windowed cylindrical plastic 396
cores covered in 10 µm nylon mesh (Supplemental Figure S1) were inserted into the substrate 397
within each experimental pot. Two of the cores were filled with the same homogenous mixture 398
of acid-washed silica sand and compost (Petersfield No.2, Leicester, UK), together making up 399
95% of the core volume as made up with microcosm bulk soil together with native soil gathered 400
from around the roots of wild plants (4% of the core volume) to ensure cores contained the 401
same microbial communities as bulk soil in pots, and fine-ground tertiary basalt (1% core 402
volume) to act as fungal bait. The third core was filled with glass wool to allow below-ground 403
gas sampling throughout the 14C-labelling period to monitor soil community respiration. Plants 404
were watered every other day with no additional applications of nutrient solutions. Microcosms 405
shared a common drip-tray within each cabinet throughout the acclimation period which 406
ensured a common pool of rhizospheric microorganisms in each microcosm. 407
A total of 20 L. inundata microcosms were maintained in controlled environment 408
chambers (Micro Clima 1200, Snijders Labs, The Netherlands) with a light cycle of 16 h 409
daytime (20 °C and 70 % humidity) and 8 h night-time (at 15 °C and 70 % humidity). Day-time 410
PAR, supplied by LED lighting was 225 µmol photons m-2 s-1. Atmospheric CO2 (a[CO2]) 411
concentrations were set at 440 ppm. a[CO2] was monitored using a Vaisala sensor system 412
(Vaisala, Birmingham, UK), maintained through addition of gaseous CO2. Cabinet settings and 413
contents were alternated every four weeks, and all pots were rotated within cabinets to control 414
for cabinet and block effects. Plants were acclimated to chamber/growth regimes (see 415
Supplemental Data) for four weeks to allow establishment of mycelial networks within pots 416
and confirmed by hyphal extraction from soil and staining with trypan blue (Brundrett et al., 417
1996). Additionally, roots were stained with acidified ink for the presence of fungi, based on 418
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

16
the methods of Brundrett et al., (1996). 419
420
Molecular identification of fungal symbionts and phylogenetic analysis 421
All plants (Supplemental Table S1) were processed for molecular analyses within one 422
week of collection. Genomic DNA extraction and purification from all specimens and 423
subsequent amplification, cloning and Sanger sequencing were performed according to 424
methods from Rimington et al., (2015). The fungal 18S ribosomal rRNA gene was targeted 425
using the broad specificity fungal primer set NS1/EF3 and a semi-nested approach with 426
Mucoromycotina- and Glomeromycotina-specific primers described in Desirò et al., (2013) for 427
the experimental L. inundata plants and all other field collected plant material. Resulting partial 428
18S rDNA sequences ca. 400-700bp were edited and preliminarily identified with BLAST in 429
Geneious v. 8.1.7 (Kearse et al., 2012). Chimeric sequences were detected using the 430
UCHIME2 algorithm (Edgar, 2016) in conjunction with the most recent non-redundant SSU 431
SILVA database (SSU Ref NR 132, December 2017, www.arb-silva.de). Sequences identified 432
as Mucoromycotina sp. were aligned with MAFFT prior to removing unreliable columns using 433
the default settings in GUIDANCE2 (http://guidance.tau.ac.il). The best-fit nucleotide model 434
for phylogenetic analysis was calculated using Smart Model Selection (Kumar et al., 2016). 435
Maximum likelihood (ML) with 1,000 replicates was performed using PhyML 3.0 (Guindon and 436
Gascuel, 2003). Bayesian inference analysis was conducted in Mr Bayes version 3.2.6 437
(Ronquist and Huelsenbeck, 2003) with four Markov chain Monte Carlo (MCMC) strands and 438
106 generations. Consensus trees were produced after excluding an initial burn-in of 25% of 439
the samples (Supplemental Figs. S3-9). Representative DNA sequences were deposited in 440
GenBank (see below). 441
442
Cytological analyses 443
Lycopodiella inundata gametophytes (n = 15), young sporophytes (protocorms) (n = 444
15), and roots of mature plants (both wild (n = 20) and experimental (n = 20), roots of 445
angiosperms (Holcus lanatus, Molinia caerulea and Juncus bulbosus), and liverwort 446
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

17
gametophytes (Fossombronia foveolata) (30 samples for each species) were either stained 447
with trypan blue (Brundrett et al., 1996) and photographed under a Zeiss Axioscope (Zeiss, 448
Oberkochen, Germany) equipped with an MRc digital camera, or processed for scanning 449
electron microscopy (SEM) within 48 hrs of collection. For SEM, we followed the protocol of 450
Duckett et al., (2006) (see Supplemental Data). 451
452
Quantification of C, 33P and 15N fluxes between lycophytes and fungi 453
After the four-week acclimation period, microcosms were moved to individual drip-454
trays immediately prior to isotope labelling to avoid cross-contamination of the isotope tracers. 455
100 µl of an aqueous mixture of 33P-labelled orthophosphate (specific activity 111 TBq mmol-456
1, 0.3 ng 33P added; Hartmann analytics, Braunschweig, Germany) and 15N-ammonium 457
chloride (1 mg ml-1; 0.1 mg 15N added; Sigma, Dorset, UK) was introduced into one of the soil-458
filled mesh cores in each pot through the installed capillary tube (Supplemental Fig. S2a). In 459
half (n = 10) of the pots, cores containing isotope tracers were left static to preserve direct 460
hyphal connections with the lycophytes. Fungal access to isotope tracers was limited in the 461
remaining half (n = 10) of the pots by rotating isotope tracer-containing cores through 90°, 462
thereby severing the hyphal connections between the plants and core soil. These were rotated 463
every second day thereafter, thus providing a control treatment that allows us to distinguish 464
between fungal and microbial contributions to tracer uptake by plants. Assimilation of 33P 465
tracer into above-ground plant material was monitored using a hand-held Geiger counter held 466
over the plant material daily. 467
At detection of peak activity in above-ground plant tissues (21 days after the addition 468
of the 33P and 15N tracers), the tops of 33P and 15N-labelled cores were sealed with plastic caps 469
and anhydrous lanolin and the glass wool cores were sealed with rubber septa (SubaSeal, 470
Sigma, Dorset, UK). Before cabinet lights were turned on at 0800, each pot was sealed into a 471
3.5 l, gas-tight labelling chamber and 2 ml 10% (w/v) lactic acid added to 30 µl NaH14CO3 472
(specific activity 1.621 GBq/mmol-1; Hartmann Analytics, Braunschweig, Germany), releasing 473
a 1.1-MBq pulse of 14CO2 gas into the headspace of the labelling chamber (Supplemental Fig. 474
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

18
S2b). Pots were maintained under growth cabinet conditions, and 1 ml of headspace gas was 475
sampled after 1 h and every 1.5 h thereafter. Below-ground respiration was monitored via gas 476
sampling from within the glass-wool filled core after 1 h and every 1.5 h thereafter for ~16 h. 477
478
Plant harvest and sample analyses 479
Upon detection of maximum below-ground flux of 14C, approximately 16 h after the 480
release of the 14CO2 pulse, each microcosm compartment (i.e., plant material and soil) were 481
separated, freeze-dried, weighed, and homogenised. The 33P activity in plant and soil samples 482
was quantified by digesting in concentrated H2SO4 (see Supplemental Data) and liquid 483
scintillation (Tricarb 3100TR liquid scintillation analyser, Isotech, Chesterfield, UK). The 484
quantity of 33P tracer that was transferred to the plant by it’s fungal partner was then calculated 485
using previously published equations (Cameron et al., 2007) (see Supplemental Data). To 486
determine total symbiotic fungal-acquired 33P transferred to L. inundata, the mean 33P content 487
of plants which did not have access to the tracer because cores into which the 33P was 488
introduced were rotated, thus severing the hyphal connections between plant and core 489
contents, was subtracted from the total 33P in each plant that did have access to the isotopes 490
within the core via intact fungal hyphal connections where isotope-containing cores remained 491
static throughout the labelling period. This calculation controls for diffusion of isotopes and 492
microbial nutrient cycling in pots, ensuring only 33P gained by the plant via intact fungal hyphal 493
connections is accounted for and therefore serves as a conservative measure of the minimum 494
fungal transfer of tracer to the plant. 495
Between two and four mg of freeze-dried, homogenised plant tissue was weighed into 496
6 x 4 mm2 tin capsules (Sercon Ltd. Irvine, UK) and 15N abundance was determined using a 497
continuous flow IRMS (PDZ 2020 IRMS, Sercon Ltd. Irvine, UK). Air was used as the reference 498
standard, and the IRMS detector was regularly calibrated to commercially available reference 499
gases. The 15N transferred from fungus to plant was then calculated using equations published 500
previously (Field et al., 2016) (see Supplemental Data). In a similar manner as for the 33P 501
tracer, the mean of the total 15N in plants without access to the isotope because of broken 502
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

19
hyphal connections between plant and core contents was subtracted from the total 15N in each 503
plant with intact hyphal connections to the mesh-covered core to give fungal-acquired 15N. 504
Again, this provides a conservative measure of 15N transfer from fungus to plant as it ensures 505
only 15N gained by the plant via intact fungal hyphal connections is accounted for. 506
The 14C activity of plant and soil samples was quantified through sample oxidation (307 507
Packard Sample Oxidiser, Isotech, Chesterfield, UK) followed by liquid scintillation. Total C 508
(12C + 14C) fixed by the plant and transferred to the fungal network was calculated as a function 509
of the total volume and CO2 content of the labelling chamber and the proportion of the supplied 510
14CO2 label fixed by plants (see Supplemental Data). The difference in total C between the 511
values obtained for static and rotated core contents in each pot is considered equivalent to 512
the total C transferred from plant to symbiotic fungus within the soil core for that microcosm, 513
noting that a small proportion will be lost through soil microbial respiration. The total C budget 514
for each experimental pot was calculated using equations from Cameron et al., (2006) (see 515
Supplemental Data). Total percent allocation of plant-fixed C to extraradical symbiotic fungal 516
hyphae was calculated by subtracting the activity (Bq) of rotated core samples from that 517
detected in static core samples in each pot, dividing this by the sum of activity detected in all 518
components of each microcosm, then multiplying it by 100. 519
520
Stable isotope signatures of neighbouring plants 521
Lycopodiella inundata and J. bulbosus were collected from Thursley National Nature 522
Reserve, Surrey, together with co-occurring reference plants from six 1 m2 plots in May 2018, 523
following the sampling scheme of (Gebauer and Meyer, 2003). Five plant species representing 524
three different types of mycorrhizal associations served as reference plants: two ericoid 525
mycorrhizal species (Erica tetralix, collected on six plots; Calluna vulgaris, collected on three 526
plots), two ectomycorrhizal species (Pinus sylvestris and Betula pendula seedlings, both from 527
one plot), and one arbuscular mycorrhizal species (Molinia caerulea from six plots). Relative 528
C and N isotope natural abundances of dried and ground leaf and root samples were 529
measured in a dual element analysis mode with an elemental analyser (Carlo Erba 530
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

20
Instruments 1108, Milan, Italy) coupled to a continuous flow isotope ratio mass spectrometer 531
(delta S, Finnigan MAT, Bremen, Germany) via a ConFlo III open-split interface (Thermo 532
Fisher Scientific, Bremen, Germany) as described in Bidartondo et al., (2004). Relative isotope 533
abundances (δ values) were calculated, calibrated and checked for accuracy using methods 534
detailed in Supplemental Data. 535
536
Statistics 537
Isotope tracing data were checked for normality and differences between plant 538
assimilation of 33P and 15N were tested using Student’s t-test in SPSS Version 24 (IBM, New 539
York). Mean values are displayed in figures with standard error. Stable isotope patterns 540
between the groups of Lycopodiella inundata (n = 6), Juncus bulbosus (n = 6) and surrounding 541
angiosperms (n = 17) were tested for normality and equal variance. One-tailed Kruskal Wallis 542
test (Q) was applied for non-parametric data followed by Dunn’s post hoc procedure while 543
One-way ANOVA (F) was applied for parametric data followed by Tukey post hoc procedure 544
(q). Mean values are displayed in figures with standard deviation. 545
546
Accession numbers 547
Representative DNA sequences were deposited in GenBank, accession numbers MK673773-548
MK673803. 549
550
Supplemental Data 551
Supplemental Figure S1. Schematic diagram of mesh-covered core showing dimenions of 552
window (not drawn to scale). 553
Supplemental Figure S2. Schematic diagrams of experimental microcosms showing (a) 15N 554
and 33P and (b) 14C isotope tracing 555
Supplemental Figure S3. Phylogenetic relationships of Mucoromycotina OTUs associated 556
with Lycopodiella inundata grown for isotope tracing and cytology experiments in controlled 557
environment growth chambers. 558
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

21
Supplemental Figure S4. An overview of phylogenetic relationships of Mucoromycotina 559
OTUs associated with bryophytes, lycophytes and angiosperms from various UK locations 560
based on partial 18S gene sequences 561
Supplemental Figure S5. Phylogenetic relationships of partial 18S DNA sequences classified 562
as OTU 2, corresponding to “group A” in Desirò et al., (2013) 563
Supplemental Figure S6. Phylogenetic relationships of partial 18S DNA sequences classified 564
as OTU 1, 3 and 4. 565
Supplemental Figure S7. Phylogenetic relationships of partial 18S DNA sequences 566
clustering within OTU 5. 567
Supplemental Figure S8. Phylogenetic relationships of partial 18S DNA sequences classified 568
as OTU 6, corresponding to “group B” in Desirò et al. ( 2013). 569
Supplemental Figure S9. Phylogenetic relationships of partial 18S DNA sequences 570
clustering with OTU 7, corresponding to “group I” in Desirò et al. (2013). 571
Supplemental Figure S10. Micrographs showing morphology of fungal colonization in 572
Fossombronia foveolata and Holcus lanatus. 573
Supplemental Figure S11. Scanning electron micrographs, except (g) light micrograph of 574
toluidine blue stained semi-thin sections. Gametophyte morphologies in Lycopodiella inundata 575
(from Thursley Common) 576
Supplemental Table S1. Samples of lycophytes, liverworts and angiosperms analysed with 577
their origin. 578
Supplemental Table S2. A summary of Mucoromycotina OTUs associated with liverworts, 579
lycophytes and angiosperms at four UK sites. 580
Supplemental Table S3. A summary of the amounts of carbon, 15N and 33P detected in static 581
and rotated core of microcosms used during carbon-for-nutrient experiments between 582
Lycopodiella inundata and Mucoromycotina FRE fungi. 583
584
Acknowledgements 585
We gratefully acknowledge support from the NERC to KJF, SP, SS (NE/N00941X/1) and MIB 586
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.
22
(NE/N009665/1) and the Leverhume Trust (PLP-2017-079). KJF is funded by BBSRC 587
Translational Fellowship (BB/M026825/1). WRR is supported by a NERC DTP (Science and 588
Solutions for a Changing Planet) studentship. We thank James Giles (Natural England) for 589
field support, Julia Masson and the RSPB for access to the Norfolk site, and The Species 590
Recovery Trust for access to the Dorset site. We are grateful to the anonymous reviewers of 591
this manuscript and to the editor for their constructive comments. 592
593
Table 1. Cytology of colonisation and fungal identity of study species (*) compared to 594
relevant examples from the literature that are referred to in the Discussion. *Results from 595
this study; G = gametophyte generation; S= sporophyte generation; ICSs = intercellular 596
spaces 597
Plant Group G/S Tissue/location Colonization Morphology (diameter) Fungus ID
References
Liverworts
Treubia1 G several ventral cell layers
intracellular coils (0.5-1.5 µm) with ‘lumps’/swellings (up to 15 µm), arbuscule-like short-side branches on coiled hyphae
M2 Duckett et al. 2006 Bidartondo et al. 2011
above intracellular zone
intercellular: large mucilage-filled ICSs
coarse hyphae 2-3 µm, thick-walled fungal structures
Fossombronia* G thallus central strand
intracellular coarse hyphae (2-3 µm); large vesicles (15-30 µm), coils (0.5-1µm), fine hyphae (0.5-1.5 µm) with small swellings/vesicles (5-10 µm), arbuscules
(M&G)*
Lycophytes
Lycopodiella* G outer cortical cell layers several ventral cell layers
intracellular intracellular
coils (up to 2.5µm) with vesicles (15-20µm) fine hyphae (0.5-1.5 µm) with small swellings/vesicles (5-10 µm)
M*
above intracellular zone
intercellular: large mucilage-filled ICSs
coarse hyphae (2->3 µm)
S protocorm: several ventral cell layers central, above intracellular zone
intracellular intercellular: large mucilage-filled ICSs
fine hyphae (0.5-1.5 µm) with small swellings/vesicles (5-10 µm) coarse hyphae (2->3 µm)
M*
S root intracellular and
fine hyphae (0.5-1.5 µm) with small
M1* Rimington et al. 2015
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

23
intercellular, small ICSs
swellings/vesicles (5-15 µm)
Angiosperms
Holcus* S root intracellular and intercellular, small ICSs
coarse hyphae (>3 µm), large vesicles (20-40 µm), fine hyphae (0.5-1.5 µm) with small vesicles (5-10 µm), arbuscules/arbuscule-like structures
(M&G)*
Molinia* S root intracellular and intercellular, small ICSs
coarse hyphae (>3 µm), large vesicles (20-40 µm), fine hyphae (0.5-1.5 µm) with small vesicles/swellings (5-10 µm), arbuscules/arbuscule-like structures
(M&G)*
Juncus* S root intracellular and intercellular, small ICSs
coarse hyphae (>3 µm), large vesicles (20-40 µm), fine hyphae (0.5-1.5 µm) with small vesicles (5-10 µm), arbuscules/arbuscule-like structures
(M&G)*
Trifolium1 S root intracellular and intercellular, small ICSs
coarse hyphae (>3 m),
large vesicles (>30 m)
fine hyphae (>1.5 m), intercalary and terminal vesicles/swellings (5-10
m) and arbuscules/arbuscule-like structures
(M&G)1 Orchard et al. 2017a
Fossils
Horneophyton1 S aerial axes, cortical cells
intracellular coarse hyphae (>3 m), large vesicles (up to 50
m), arbuscule-like structures
G1 Strullu-Derrien et al. 2014
corm intracellular and intercellular
intracellular coils, intercellular coarse
hyphae (11-13 m), thick-walled fungal structures
M1
Nothia1 S aerial and prostrate axes
Intercellular and intracellular
coarse hyphae (up to 15
m) and intercellular
vesicles (>50 m)
? Krings et al. 2007
598
Figure legends 599
Figure 1. Land plant phylogeny and species used in the present study. (a) Land plant 600
phylogeny showing key nodes alongside commonly associated fungal symbionts (Duckett and 601
Ligrone, 1992; Duckett et al., 2006; James et al., 2006; Bidartondo et al., 2011) (b) 602
Lycopodiella inundata at Thursley Common, Surrey, June 2017. 603
604
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

24
Figure 2. Figure 2. Carbon-for-nutrient exchange between Lycopodiella inundata and 605
Mucoromycotina fine root endophyte fungi. (a) Total plant-fixed carbon (C) transferred to 606
Mucoromycotina fine root endophyte (FRE) fungi by L. inundata. (b) Percent allocation of 607
plant-fixed C to Mucoromycotina FRE fungi (c) Total plant tissue phosphorus (33P) and 608
nitrogen (15N) content (ng) and (d) tissue concentration (ng g-1) of fungal-acquired 33P and 15N 609
in L. inundata tissue. In all panels, error bars denote SEM. In panels (a) and (b) n = 20. In 610
panels (c) and (d) n = 10. n is equivalent to the number of biological replicates used during 611
carbon-for-nutrient exchange experiements. Experiments were carried out three times. * 612
indicates where P <0.05, Student’s T-Test. 613
614
615
Figure 3. Carbon and nitrogen stable isotope natural abundance of Lycopodiella 616
inundata (L.i., n = 6), Juncus bulbosus (J.b., n = 6) and surrounding angiosperms 617
(arbuscular mycorrhizal: Molinia caerulea (M.c., n = 6); ectomycorrhizal: Pinus sylvestris (P.s., 618
n = 1); Betula pendula (B.p., n = 1); ericoid mycorrhizal: Calluna vulgaris (C.v., n = 3), Erica 619
tetralix (E.t., n = 6)) for leaf (A) and root (B) samples, respectively. Mean values are displayed 620
with standard deviation. One-tailed Kruskal Wallis test, followed by Dunn’s post hoc, found 621
significant differences between L. inundata, J. bulbosus and surrounding angiosperms as 622
references in leaf carbon and nitrogen stable isotope natural abundance and in root nitrogen 623
stable isotope natural abundance. 624
625
Figure 4. Light micrographs of trypan blue stained tissues. (a) Branching fine hyphae with 626
small swellings/vesicles in thallus cells and rhizoid (b) of the liverwort Fossombronia foveolata 627
(from Thursely Common) colonized by both Mucoromycotina fine root endophytes (FREs) and 628
Glomeromycotina, in (b) also note the coarse hyphae (arrowhead). (c-e) Fine hyphae with 629
small swellings/vesicles in the root hairs and root cells of the lycophyte Lycopodiella inundata 630
colonized by Mucoromycotina FREs only (field-collected specimens from Thursley Common). 631
(f) Fine hyphae with small swellings/vesicles and large vesicles in a root of the grass Holcus 632
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

25
lanatus (from Lynn Crafnant) colonized by both Mucoromycotina FREs and Glomeromycotina. 633
(g-h) Roots of the grass Molinia caerulea (from Thursley Common) colonized by both 634
Mucoromycotina FREs and Glomeromycotina, showing fine hyphae (g) and coarse hyphae 635
with large vesicles (h). Scale bars: (a, b, d-f) 50 µm, (c, g, h) 100 µm. 636
637
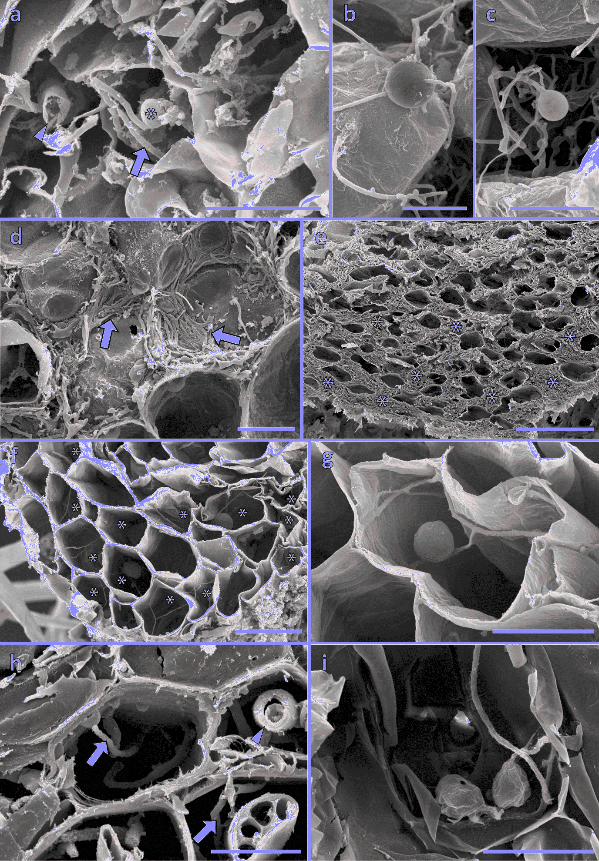
Figure 5. Scanning electron micrographs. (a) Fine hyphae (arrows) with a small 638
swelling/vesicle (*) in the thallus cells of Fossombronia foveolata (from Thursley Common), 639
also note the much coarser hyphae (arrowheads). (b-g) Fungal colonization in Lycopodiella 640
inundata. (b, c) Intercalary (b) and terminal (c) small swellings/vesicles on fine hyphae in the 641
ventral cell layers of a protocorm (from Thursley Common). Centrally and above this 642
intracellular colonization zone, the fungus becomes exclusively intercellular, as evidenced by 643
the presence of abundant, tightly-appressed hyphae surrounding the central protocorm cells 644
(arrows) (d) and eventually completely fills the large, mucilage-filled intercellular spaces 645
present in this zone (e, *). (f, g) Cross sections of roots of experimental plants; (f) several cells 646
colonized exclusively (*) by branching fine hyphae with small swellings/vesicles, enlarged in 647
(g). (h, i) Cross sections of roots of Juncus bulbosus (from Thursley Common) showing fine 648
(arrow) and coarse (arrowheads) hyphae (h) and a fine hypha with small swellings/vesicles 649
(i). Scale bars: (a, d, g, i) 20 µm, (b, c, h) 10 µm, (e) 100 µm, (f) 50 µm. 650
651
Literature Cited 652
Albornoz FE, Lambers H, Turner BL, Teste FP, Laliberté E (2016) Shifts in symbiotic 653
associations in plants capable of forming multiple root symbioses across a long‐654
term soil chronosequence. Ecology and Evolution 6: 2368-2377 655 Bago B, Pfeffer PE, Shachar-Hill Y (2000) Carbon metabolism and transport in arbuscular 656
mycorrhizas. Plant Physiology 124: 949-958 657 Bidartondo MI, Burghardt B, Gebauer G, Bruns TD, Read DJ (2004) Changing partners 658
in the dark: isotopic and molecular evidence of ectomycorrhizal liaisons between 659 forest orchids and trees. Proceedings of the Royal Society of London B: Biological 660 Sciences 271: 1799-1806 661
Bidartondo MI, Read DJ, Trappe JM, Merckx V, Ligrone R, Duckett JG (2011) The dawn 662 of symbiosis between plants and fungi. Biology Letters: rsbl20101203 663
Brundrett M, Bougher N, Dell B, Grove T (1996) Working with Mycorrhizas in Forestry and 664 Agriculture. 665
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

26
Cameron DD, Johnson I, Leake JR, Read DJ (2007) Mycorrhizal acquisition of inorganic 666 phosphorus by the green-leaved terrestrial orchid Goodyera repens. Annals of 667 Botany 99: 831-834 668
Cameron DD, Leake JR, Read DJ (2006) Mutualistic mycorrhiza in orchids: evidence from 669
plant–fungus carbon and nitrogen transfers in the green‐leaved terrestrial orchid 670
Goodyera repens. New Phytologist 171: 405-416 671 Cernusak LA, Tcherkez G, Keitel C, Cornwell WK, Santiago LS, Knohl A, Barbour MM, 672
Williams DG, Reich PB, Ellsworth DS (2009) Why are non-photosynthetic tissues 673 generally 13C enriched compared with leaves in C3 plants? Review and synthesis of 674 current hypotheses. Functional Plant Biology 36: 199-213 675
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in 676 plant ecology. Annual Review of Ecology and Systematics 33: 507-559 677
Desirò A, Duckett JG, Pressel S, Villarreal JC, Bidartondo MI (2013) Fungal symbioses 678 in hornworts: a chequered history. Proceedings of the Royal Society of London B: 679 Biological Sciences 280: 20130207 680
Duckett JG, Carafa A, Ligrone R (2006) A highly differentiated glomeromycotean 681 association with the mucilage-secreting, primitive antipodean liverwort Treubia 682 (Treubiaceae): clues to the origins of mycorrhizas. American Journal of Botany 93: 683 797-813 684
Duckett JG, Ligrone R (1992) A light and electron microscope study of the fungal 685 endophytes in the sporophyte and gametophyte of Lycopodium cernuum with 686 observations on the gametophyte–sporophyte junction. Canadian Journal of Botany 687 70: 58-72 688
Edgar R (2016) UCHIME2: improved chimera prediction for amplicon sequencing. bioRxiv: 689 074252 690
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and 691 photosynthesis. Annual Review of Plant Biology 40: 503-537 692
Farquhar GD, O'Leary MH, Berry JA (1982) On the relationship between carbon isotope 693 discrimination and the intercellular carbon dioxide concentration in leaves. Functional 694 Plant Biology 9: 121-137 695
Field KJ, Bidartondo MI, Rimington WR, Hoysted GA, Beerling DJ, Cameron DD, 696 Duckett JG, Leake JR, Pressel S (2019) Functional complementarity of ancient 697
plant‐fungal mutualisms: contrasting nitrogen, phosphorus and carbon exchanges 698
between Mucoromycotina and Glomeromycotina fungal symbionts of liverworts. New 699 Phytologist https://doi.org/10.1111/nph.15819 700
Field KJ, Cameron DD, Leake JR, Tille S, Bidartondo MI, Beerling DJ (2012) Contrasting 701 arbuscular mycorrhizal responses of vascular and non-vascular plants to a simulated 702 Palaeozoic CO2 decline. Nature Communications 3: 835 703
Field KJ, Pressel S (2018) Unity in diversity: structural and functional insights into the 704 ancient partnerships between plants and fungi. New Phytologist 220: 996–1011 705
Field KJ, Pressel S, Duckett JG, Rimington WR, Bidartondo MI (2015a) Symbiotic 706 options for the conquest of land. Trends in Ecology & Evolution 30: 477-486 707
Field KJ, Rimington WR, Bidartondo MI, Allinson KE, Beerling DJ, Cameron DD, 708 Duckett JG, Leake JR, Pressel S (2015b) First evidence of mutualism between 709 ancient plant lineages (Haplomitriopsida liverworts) and Mucoromycotina fungi and 710 its response to simulated Palaeozoic changes in atmospheric CO2. New Phytologist 711 205: 743-756 712
Field KJ, Rimington WR, Bidartondo MI, Allinson KE, Beerling DJ, Cameron DD, 713 Duckett JG, Leake JR, Pressel S (2016) Functional analysis of liverworts in dual 714 symbiosis with Glomeromycota and Mucoromycotina fungi under a simulated 715 Palaeozoic CO2 decline. The ISME Journal 10: 1514-1526 716
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

27
Gebauer G, Meyer M (2003) 15N and 13C natural abundance of autotrophic and myco‐717
heterotrophic orchids provides insight into nitrogen and carbon gain from fungal 718 association. New Phytologist 160: 209-223 719
Gebauer G, Schulze E-D (1991) Carbon and nitrogen isotope ratios in different 720 compartments of a healthy and a declining Picea abies forest in the Fichtelgebirge, 721 NE Bavaria. Oecologia 87: 198-207 722
Guindon S, Gascuel O (2003) A simple, fast, and accurate algorithm to estimate large 723 phylogenies by maximum likelihood. Systematic Biology 52: 696-704 724
Hoysted GA, Kowal J, Jacob A, Rimington WR, Duckett JG, Pressel S, Orchard S, 725 Ryan MH, Field KJ, Bidartondo MI (2018) A mycorrhizal revolution. Current 726 Opinion in Plant Biology 44: 1-6 727
James TY, Kauff F, Schoch CL, Matheny PB, Hofstetter V, Cox CJ, Celio G, Gueidan C, 728 Fraker E, Miadlikowska J (2006) Reconstructing the early evolution of Fungi using 729 a six-gene phylogeny. Nature 443: 818-822 730
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, 731 Cooper A, Markowitz S, Duran C (2012) Geneious Basic: an integrated and 732 extendable desktop software platform for the organization and analysis of sequence 733 data. Bioinformatics 28: 1647-1649 734
Kenrick P, Crane PR (1997) The origin and early evolution of plants on land. Nature 389: 735 33 736
Krapp A (2015) Plant nitrogen assimilation and its regulation: a complex puzzle with missing 737 pieces. Current Opinion in Plant Biology 25: 115-122 738
Krings M, Taylor TN, Hass H, Kerp H, Dotzler N, Hermsen EJ (2007) An alternative mode 739 of early land plant colonization by putative endomycorrhizal fungi. Plant Signaling & 740 Behavior 2: 125-126 741
Kumar S, Stecher G, Tamura K (2016) MEGA7: molecular evolutionary genetics analysis 742 version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870-1874 743
Lin K, Limpens E, Zhang Z, Ivanov S, Saunders DG, Mu D, Pang E, Cao H, Cha H, Lin T 744 (2014) Single nucleus genome sequencing reveals high similarity among nuclei of an 745 endomycorrhizal fungus. PLoS Genetics 10: e1004078 746
Morris JL, Puttick MN, Clark JW, Edwards D, Kenrick P, Pressel S, Wellman CH, Yang 747 Z, Schneider H, Donoghue PC (2018) The timescale of early land plant evolution. 748 Proceedings of the National Academy of Sciences: 201719588 749
Orchard S, Hilton S, Bending GD, Dickie IA, Standish RJ, Gleeson DB, Jeffery RP, 750 Powell JR, Walker C, Bass D (2017a) Fine endophytes (Glomus tenue) are related 751 to Mucoromycotina, not Glomeromycota. New Phytologist 213: 481-486 752
Orchard S, Standish RJ, Dickie IA, Renton M, Walker C, Moot D, Ryan MH (2017b) Fine 753 root endophytes under scrutiny: a review of the literature on arbuscule-producing 754 fungi recently suggested to belong to the Mucoromycotina. Mycorrhiza 27: 619-638 755
Pirozynski KA, Malloch DW (1975) The origin of land plants: A matter of mycotrophism. 756 Biosystems 6: 153-164 757
Press MC, Shah N, Tuohy JM, Stewart GR (1987) Carbon isotope ratios demonstrate 758 carbon flux from C4 host to C3 parasite. Plant Physiology 85: 1143-1145 759
Redecker D, Kodner R, Graham LE (2000) Glomalean fungi from the Ordovician. Science 760 289: 1920-1921 761
Rimington WR, Pressel S, Duckett JG, Bidartondo MI (2015) Fungal associations of 762 basal vascular plants: reopening a closed book? New Phytologist 205: 1394-1398 763
Rimington WR, Pressel S, Field KJ, Strullu‐Derrien C, Duckett JG, Bidartondo MI 764
(2016) Reappraising the origin of mycorrhizas. Molecular Mycorrhizal Symbiosis: 31-765 32 766
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under 767 mixed models. Bioinformatics 19: 1572-1574 768
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

28
Ryan MH, Kirkegaard JA (2012) The agronomic relevance of arbuscular mycorrhizas in the 769 fertility of Australian extensive cropping systems. Agriculture, Ecosystems & 770 Environment 163: 37-53 771
Schmid E, Oberwinkler F (1993) Mycorrhiza‐like interaction between the achlorophyllous 772
gametophyte of Lycopodium clavatum L. and its fungal endophyte studied by light 773 and electron microscopy. New Phytologist 124: 69-81 774
Schulze E-D, Chapin FS, Gebauer G (1994) Nitrogen nutrition and isotope differences 775 among life forms at the northern treeline of Alaska. Oecologia 100: 406-412 776
Schulze E-D, Lange O, Ziegler H, Gebauer G (1991) Carbon and nitrogen isotope ratios of 777 mistletoes growing on nitrogen and non-nitrogen fixing hosts and on CAM plants in 778 the Namib desert confirm partial heterotrophy. Oecologia 88: 457-462 779
Smith SE, Anderson IC, Smith FA (2015) Mycorrhizal associations and phosphorus 780 acquisition: from cells to ecosystems. Annual Plant Reviews Online 48: 409-440 781
Smith SE, Smith FA (2011) Roles of arbuscular mycorrhizas in plant nutrition and growth: 782 new paradigms from cellular to ecosystem scales. Annual Review of Plant Biology 783 62: 227-250 784
Strullu‐Derrien C, Kenrick P, Pressel S, Duckett JG, Rioult JP, Strullu DG (2014) 785
Fungal associations in Horneophyton ligneri from the Rhynie Chert (c. 407 million 786 year old) closely resemble those in extant lower land plants: novel insights into 787 ancestral plant–fungus symbioses. New Phytologist 203: 964-979 788
von Oheimb G, Power SA, Falk K, Friedrich U, Mohamed A, Krug A, Boschatzke N, 789 Härdtle W (2010) N: P ratio and the nature of nutrient limitation in Calluna-dominated 790 heathlands. Ecosystems 13: 317-327 791
Walker C, Gollotte A, Redecker D (2018) A new genus, Planticonsortium 792 (Mucoromycotina), and new combination (P. tenue), for the fine root endophyte, 793 Glomus tenue (basionym Rhizophagus tenuis). Mycorrhiza: 1-7 794
795
www.plantphysiol.orgon August 25, 2019 - Published by Downloaded from Copyright © 2019 American Society of Plant Biologists. All rights reserved.

Seedplants
Ferns
Lycophytes
Hornworts
Mosses
Liverworts
Selaginellaceae
Isoetaceae
Lycopodiaceae
Mucoromycotina
Glomeromycotina
Basidiomycota
Ascomycota
Nosymbiosis
b
a

0.0
0.2
0.4
0.6
0.8
1.0
% C
allo
catio
n to
fung
us
33P 15N0.01
0.1
1
10
100
1000
10000
Tota
l pla
nt
33P
and
15N
(ng)
*
0
50
100
150
Tota
l fun
gal
C (µ
g)
[33P] [15N]0.001
0.01
0.1
1
10
100
1000
10000
Plan
t [33
P] a
nd
[15N
] (ng
g-1
)
*
(a)(a) (b)
(c)(c) (d)


Figure 4.

Parsed CitationsAlbornoz FE, Lambers H, Turner BL, Teste FP, Laliberté E (2016) Shifts in symbiotic associations in plants capable of forming multipleroot symbioses across a long‐term soil chronosequence. Ecology and Evolution 6: 2368-2377
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bago B, Pfeffer PE, Shachar-Hill Y (2000) Carbon metabolism and transport in arbuscular mycorrhizas. Plant Physiology 124: 949-958Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bidartondo MI, Burghardt B, Gebauer G, Bruns TD, Read DJ (2004) Changing partners in the dark: isotopic and molecular evidence ofectomycorrhizal liaisons between forest orchids and trees. Proceedings of the Royal Society of London B: Biological Sciences 271:1799-1806
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bidartondo MI, Read DJ, Trappe JM, Merckx V, Ligrone R, Duckett JG (2011) The dawn of symbiosis between plants and fungi. BiologyLetters: rsbl20101203
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Brundrett M, Bougher N, Dell B, Grove T (1996) Working with Mycorrhizas in Forestry and Agriculture.
Cameron DD, Johnson I, Leake JR, Read DJ (2007) Mycorrhizal acquisition of inorganic phosphorus by the green-leaved terrestrialorchid Goodyera repens. Annals of Botany 99: 831-834
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cameron DD, Leake JR, Read DJ (2006) Mutualistic mycorrhiza in orchids: evidence from plant–fungus carbon and nitrogen transfersin the green‐leaved terrestrial orchid Goodyera repens. New Phytologist 171: 405-416
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cernusak LA, Tcherkez G, Keitel C, Cornwell WK, Santiago LS, Knohl A, Barbour MM, Williams DG, Reich PB, Ellsworth DS (2009) Whyare non-photosynthetic tissues generally 13C enriched compared with leaves in C3 plants? Review and synthesis of currenthypotheses. Functional Plant Biology 36: 199-213
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in plant ecology. Annual Review of Ecology andSystematics 33: 507-559
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Desirò A, Duckett JG, Pressel S, Villarreal JC, Bidartondo MI (2013) Fungal symbioses in hornworts: a chequered history. Proceedingsof the Royal Society of London B: Biological Sciences 280: 20130207
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Duckett JG, Carafa A, Ligrone R (2006) A highly differentiated glomeromycotean association with the mucilage-secreting, primitiveantipodean liverwort Treubia (Treubiaceae): clues to the origins of mycorrhizas. American Journal of Botany 93: 797-813
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Duckett JG, Ligrone R (1992) A light and electron microscope study of the fungal endophytes in the sporophyte and gametophyte ofLycopodium cernuum with observations on the gametophyte–sporophyte junction. Canadian Journal of Botany 70: 58-72
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Edgar R (2016) UCHIME2: improved chimera prediction for amplicon sequencing. bioRxiv: 074252Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Biology 40:503-537
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Farquhar GD, O'Leary MH, Berry JA (1982) On the relationship between carbon isotope discrimination and the intercellular carbondioxide concentration in leaves. Functional Plant Biology 9: 121-137
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Field KJ, Bidartondo MI, Rimington WR, Hoysted GA, Beerling DJ, Cameron DD, Duckett JG, Leake JR, Pressel S (2019) Functionalcomplementarity of ancient plant‐fungal mutualisms: contrasting nitrogen, phosphorus and carbon exchanges betweenMucoromycotina and Glomeromycotina fungal symbionts of liverworts. New Phytologist https://doi.org/10.1111/nph.15819
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Field KJ, Cameron DD, Leake JR, Tille S, Bidartondo MI, Beerling DJ (2012) Contrasting arbuscular mycorrhizal responses of vascularand non-vascular plants to a simulated Palaeozoic CO2 decline. Nature Communications 3: 835
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Field KJ, Pressel S (2018) Unity in diversity: structural and functional insights into the ancient partnerships between plants and fungi.New Phytologist 220: 996–1011
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Field KJ, Pressel S, Duckett JG, Rimington WR, Bidartondo MI (2015a) Symbiotic options for the conquest of land. Trends in Ecology &Evolution 30: 477-486
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Field KJ, Rimington WR, Bidartondo MI, Allinson KE, Beerling DJ, Cameron DD, Duckett JG, Leake JR, Pressel S (2015b) Firstevidence of mutualism between ancient plant lineages (Haplomitriopsida liverworts) and Mucoromycotina fungi and its response tosimulated Palaeozoic changes in atmospheric CO2. New Phytologist 205: 743-756
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Field KJ, Rimington WR, Bidartondo MI, Allinson KE, Beerling DJ, Cameron DD, Duckett JG, Leake JR, Pressel S (2016) Functionalanalysis of liverworts in dual symbiosis with Glomeromycota and Mucoromycotina fungi under a simulated Palaeozoic CO2 decline. TheISME Journal 10: 1514-1526
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gebauer G, Meyer M (2003) 15N and 13C natural abundance of autotrophic and myco‐heterotrophic orchids provides insight intonitrogen and carbon gain from fungal association. New Phytologist 160: 209-223
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gebauer G, Schulze E-D (1991) Carbon and nitrogen isotope ratios in different compartments of a healthy and a declining Picea abiesforest in the Fichtelgebirge, NE Bavaria. Oecologia 87: 198-207
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Guindon S, Gascuel O (2003) A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. SystematicBiology 52: 696-704
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hoysted GA, Kowal J, Jacob A, Rimington WR, Duckett JG, Pressel S, Orchard S, Ryan MH, Field KJ, Bidartondo MI (2018) Amycorrhizal revolution. Current Opinion in Plant Biology 44: 1-6
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
James TY, Kauff F, Schoch CL, Matheny PB, Hofstetter V, Cox CJ, Celio G, Gueidan C, Fraker E, Miadlikowska J (2006) Reconstructingthe early evolution of Fungi using a six-gene phylogeny. Nature 443: 818-822
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C (2012) GeneiousBasic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28:1647-1649
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kenrick P, Crane PR (1997) The origin and early evolution of plants on land. Nature 389: 33Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Krapp A (2015) Plant nitrogen assimilation and its regulation: a complex puzzle with missing pieces. Current Opinion in Plant Biology25: 115-122
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Krings M, Taylor TN, Hass H, Kerp H, Dotzler N, Hermsen EJ (2007) An alternative mode of early land plant colonization by putative

endomycorrhizal fungi. Plant Signaling & Behavior 2: 125-126Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kumar S, Stecher G, Tamura K (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. MolecularBiology and Evolution 33: 1870-1874
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lin K, Limpens E, Zhang Z, Ivanov S, Saunders DG, Mu D, Pang E, Cao H, Cha H, Lin T (2014) Single nucleus genome sequencingreveals high similarity among nuclei of an endomycorrhizal fungus. PLoS Genetics 10: e1004078
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Morris JL, Puttick MN, Clark JW, Edwards D, Kenrick P, Pressel S, Wellman CH, Yang Z, Schneider H, Donoghue PC (2018) Thetimescale of early land plant evolution. Proceedings of the National Academy of Sciences: 201719588
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Orchard S, Hilton S, Bending GD, Dickie IA, Standish RJ, Gleeson DB, Jeffery RP, Powell JR, Walker C, Bass D (2017a) Fineendophytes (Glomus tenue) are related to Mucoromycotina, not Glomeromycota. New Phytologist 213: 481-486
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Orchard S, Standish RJ, Dickie IA, Renton M, Walker C, Moot D, Ryan MH (2017b) Fine root endophytes under scrutiny: a review of theliterature on arbuscule-producing fungi recently suggested to belong to the Mucoromycotina. Mycorrhiza 27: 619-638
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pirozynski KA, Malloch DW (1975) The origin of land plants: A matter of mycotrophism. Biosystems 6: 153-164Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Press MC, Shah N, Tuohy JM, Stewart GR (1987) Carbon isotope ratios demonstrate carbon flux from C4 host to C3 parasite. PlantPhysiology 85: 1143-1145
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Redecker D, Kodner R, Graham LE (2000) Glomalean fungi from the Ordovician. Science 289: 1920-1921Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rimington WR, Pressel S, Duckett JG, Bidartondo MI (2015) Fungal associations of basal vascular plants: reopening a closed book?New Phytologist 205: 1394-1398
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rimington WR, Pressel S, Field KJ, Strullu‐Derrien C, Duckett JG, Bidartondo MI (2016) Reappraising the origin of mycorrhizas.Molecular Mycorrhizal Symbiosis: 31-32
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572-1574Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ryan MH, Kirkegaard JA (2012) The agronomic relevance of arbuscular mycorrhizas in the fertility of Australian extensive croppingsystems. Agriculture, Ecosystems & Environment 163: 37-53
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schmid E, Oberwinkler F (1993) Mycorrhiza‐like interaction between the achlorophyllous gametophyte of Lycopodium clavatum L. andits fungal endophyte studied by light and electron microscopy. New Phytologist 124: 69-81
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schulze E-D, Chapin FS, Gebauer G (1994) Nitrogen nutrition and isotope differences among life forms at the northern treeline ofAlaska. Oecologia 100: 406-412
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schulze E-D, Lange O, Ziegler H, Gebauer G (1991) Carbon and nitrogen isotope ratios of mistletoes growing on nitrogen and non-nitrogen fixing hosts and on CAM plants in the Namib desert confirm partial heterotrophy. Oecologia 88: 457-462
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Smith SE, Anderson IC, Smith FA (2015) Mycorrhizal associations and phosphorus acquisition: from cells to ecosystems. Annual PlantReviews Online 48: 409-440
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Smith SE, Smith FA (2011) Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystemscales. Annual Review of Plant Biology 62: 227-250
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Strullu‐Derrien C, Kenrick P, Pressel S, Duckett JG, Rioult JP, Strullu DG (2014) Fungal associations in Horneophyton ligneri from theRhynie Chert (c. 407 million year old) closely resemble those in extant lower land plants: novel insights into ancestral plant–fungussymbioses. New Phytologist 203: 964-979
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
von Oheimb G, Power SA, Falk K, Friedrich U, Mohamed A, Krug A, Boschatzke N, Härdtle W (2010) N: P ratio and the nature of nutrientlimitation in Calluna-dominated heathlands. Ecosystems 13: 317-327
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Walker C, Gollotte A, Redecker D (2018) A new genus, Planticonsortium (Mucoromycotina), and new combination (P. tenue), for the fineroot endophyte, Glomus tenue (basionym Rhizophagus tenuis). Mycorrhiza: 1-7
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title












![Mucoromycotina fine root endophyte fungi form nutritional ... · early fungal associates were likely to be more diverse than has previously been assumed [5-7]. Members of the earliest](https://img.pdfslide.net/doc/110x75/5f375961b609c5094969a59a/mucoromycotina-fine-root-endophyte-fungi-form-nutritional-early-fungal-associates.jpg)















