Embed Size (px)
Citation preview

ORIGINAL PAPER
Somatic embryogenesis in Alnus glutinosa (L.) Gaertn
Elena Corredoira • Silvia Valladares •
Ma Teresa Martınez • Ana Ma Vieitez •
Ma Carmen San Jose
Received: 11 March 2013 / Revised: 4 July 2013 / Accepted: 9 July 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract Induction of somatic embryos and plant
regeneration was demonstrated for the first time in Alnus
glutinosa. Somatic embryos were initiated from zygotic
embryos collected 1–3 weeks post-anthesis (WPA), i.e.,
when they were at globular or early cotyledonary stage and
were 0.5–1 mm in length. Induction frequency (16.6 %)
and the mean number of somatic embryos (4.5 embryos/
explant) were highest after culture of zygotic embryos,
collected at 3 WPA, on Murashige and Skoog medium
(MS) supplemented with 0.9-lM 2,4-dichlorophenoxy-
acetic acid and 2.22-lM benzyladenine (BA). No
embryogenic induction was observed on medium with BA
alone. Initial somatic embryos differentiated indirectly
from callus tissue formed at the surface of the zygotic
embryos. Embryogenic competence was maintained by
secondary embryogenesis, which was affected by explant
type, plant growth regulators and genotype. Secondary
embryogenesis was induced by culture of small groups of
whole somatic embryos or isolated cotyledon explants on
medium consisting of MS medium (half-strength ma-
cronutrients) supplemented with 0.44-lM BA. Histological
study of isolated cotyledon explants revealed that
secondary embryos developed directly from differentiated
embryogenic tissue on the surface of cotyledons. Somatic
embryos at successive stages of development, including
cotyledonary-stage embryos with shoot and root meris-
tems, were evident. For plantlet conversion, somatic
embryos were transferred to maturation medium supple-
mented with 3 % maltose, followed by 6 weeks of culture
in Woody Plant Medium supplemented with 0.44-lM BA
and 0.46-lM Zeatin (Z). This novel protocol appears
promising for mass propagation, conservation and genetic
transformation of black alder.
Keywords Black alder � Cones � Histology � Secondary
embryogenesis � Somatic embryos � Zygotic embryos
Introduction
Alders grow throughout the northern hemisphere in
woodland and riparian habitats. These species prefer cool
climates, although they can also withstand warmer tem-
peratures when water is adequate (Prada and Arizpe 2008).
The genus is thought to comprise approximately 30 spe-
cies, which are characterized by wind pollination, seed
dispersal by wind and water, rapid colonization of bare
ground and a relatively short life span. The male and
female flowers occur on separate catkins. The female
flowers are reddish-purple in color and develop into hard
cones containing seeds. Although alders are usually not the
focus of forest protection concerns in Europe, because they
are of minor importance to the forest economy, the eco-
logical value of the genus and its high value for land rec-
lamation and reforestation are well recognized (Cech
1998). Alders are capable of fixing atmospheric nitrogen
through symbiotic association with the actinomycete
Communicated by J. Carlson.
S. Valladares and E. Corredoira contributed equally to this work.
E. Corredoira (&) � S. Valladares � M. T. Martınez �A. M. Vieitez � M. C. San Jose
Department of Plant Physiology, Instituto de Investigaciones
Agrobiologicas de Galicia (CSIC), Apartado 122, 15780
Santiago de Compostela, Spain
e-mail: [email protected]
123
Trees
DOI 10.1007/s00468-013-0907-8

Frankia alni (Oliveira et al. 2005), which forms nodules on
the tree roots. Alnus glutinosa (L.) Gaertn, also called
common alder, black alder or European black alder, is the
most common tree species in riparian forests. In addition to
the above-mentioned characteristics, black alder has a use
in flood control, stabilization of riverbanks and in func-
tioning of the river ecosystems (Claessens et al. 2010).
Black alder is also a host to a wide variety of moss and
lichen.
Unfortunately, alder populations are threatened by lethal
alder blight disease, which is caused by the pathogenic
fungus Phytophthora alni (Brasier et al. 2004).The disease
has been particularly destructive in the UK, but is also
found in Austria, Belgium, France, Germany, and Spain,
where it threatens natural riparian populations of alder
(Brasier et al. 1995; Cech 1998; Cavelier et al. 1999;
Streito et al. 2002; Jung and Blaschke 2004; Solla et al.
2010). Various attempts have been made to control the
disease through international research programmes, none
of which has yet succeeded in significantly halting its
spread or reducing its impact (Gibbs et al. 2003; Webber
et al. 2004). Genetic engineering may prove useful for
controlling alder blight disease. Regeneration of plants
from genetically transformed cells is a key step in devel-
oping a protocol for the genetic transformation of alder.
Somatic embryogenesis is an ideal regeneration system
for genetic transformation. This propagation technique is
defined as a process in which a bipolar structure, resem-
bling a zygotic embryo, develops from a somatic cell
without vascular connection to the original tissue (von
Arnold et al. 2002). Somatic embryogenesis, in which the
frequency of chimeras is low and the regeneration rate is
high, is more attractive than organogenesis as a plant
regeneration system (Gaj 2001). In addition, somatic
embryogenesis is considered the best in vitro propagation
method for most tree species (Merkle and Nairn 2005).
This technique could make a substantial contribution to the
conservation of alder species, not only as a means of pro-
ducing transgenic trees with resistance genes, but also for
mass propagation of the resistant genotypes produced by
traditional breeding programmes. However, as far as we
know, no studies have addressed the capacity of black alder
tissue to form somatic embryos. Alder species have only
been micropropagated by multiplication and rooting of
axillary shoots (Garton et al. 1981; Perinet and Lalonde
1983; Tremblay et al. 1986; Perinet and Tremblay 1987;
Barghchi 1988; Welander et al. 1989; Lall et al. 2005; San-
Jose et al. 2012, 2013).
The main objective of this study was, therefore, to
induce somatic embryogenesis from zygotic embryos (ZEs)
in Alnus glutinosa and to define a protocol for maintenance
of embryogenic lines and plantlet regeneration.
Materials and methods
Collection and sterilization of plant material
In the first experiment, cones were collected from three A.
glutinosa trees, of age 20–25 years, during August and
September 2011 (Table 1). The trees, which were desig-
nated Sarela 1, Sarela 2 and Sar 1, were growing in Garelas
park (Sarela 1 and Sarela 2) and Branas del Sar park (Sar 1)
in Santiago de Compostela (Spain). For the second exper-
iment, carried out in 2012, cones were collected from Sa-
rela 2 at weekly intervals during August and September,
i.e., approximately 1–6 weeks post-anthesis (WPA).
Anthesis is defined as the time at which approximately half
of the tree is in bloom.
The cones were washed in running tap water for 30 min
and surface sterilized by immersion for 1 min in 70 %
ethanol followed by immersion for 10 min in a 0.5 %
solution of free chlorine (Millipore� chlorine tablets) plus
1 % Tween�80. The cones were then rinsed twice with
sterile distilled water. After the sterilization process, the
seeds were removed from the cones, and intact zygotic
embryos were excised from seeds under a stereo micro-
scope in a flow laminar chamber. Isolated zygotic embryos
were then used as initial explants and were placed in
individual 20 9 150 mm culture tubes (one zygotic
embryo 9 tube) containing 16 ml of embryo induction
medium. A similar sterilization protocol has been used to
sterilize shoot tips and nodal segments of Paulownia
tomentosa (Corredoira et al. 2008).
Induction of somatic embryogenesis
In the first set of experiments, the explants were initially
cultured on induction medium consisting of Murashige and
Skoog medium (MS; Murashige and Skoog 1962) supple-
mented with 0.5 g/l casein hydrolyzate, 3 % sucrose, 0.7 %
agar (Difco Bacto, BD, USA), and 2.22-lM benzyladenine
(BA; Sigma-Aldrich, St. Louis, MO, USA) in combination
with 0.9, 2.26, 4.52 or 9.05 lM 2,4-dichlorophenoxyacetic
acid (2,4-D; Duchefa, The Netherlands). Before sterilizing
the medium, by autoclaving at 121 �C for 20 min, the pH
was adjusted to 5.7. For each tree, seeds were collected at
2–3 WPA, and 48 zygotic embryos were cultured for each
concentration of 2,4-D, so that there were 192 explants per
tree. In the second set of experiments, zygotic embryos
were collected from the Sarela 2 tree at 1–6 WPA and
cultured on MS plus 2.22-lM BA, with or without 2,4-D
(0.9 lM). Twenty-four explants were used for each com-
bination of induction medium and collection date, thus
providing 288 explants in total. In addition, on each col-
lection date, another set of 12 seeds and 12 zygotic
Trees
123
embryos was used to evaluate the mean length of seeds and
zygotic embryos.
After 4 weeks of culture on induction medium, the
explants were transferred to 300 ml jars (4 or 5 explants
per jar) containing 50 ml of expression medium consisting
of MS salts supplemented with 0.5 g/l casein hydrolyzate,
3 % sucrose, 0.7 % Difco bacto agar, and 0.44-lM BA.
The explants were transferred monthly to fresh medium
and were examined periodically to record the number of
explants exhibiting callus formation and somatic embryos.
Initial explants were maintained in darkness at 25 �C for
4 weeks and were then subjected to a 16-h photoperiod (30
lmol m-2 s-1 from cool-white fluorescent lamps) with
25 �C light and 20 �C dark temperatures. These conditions
were applied at all subsequent stages of the embryo pro-
liferation, maturation, and germination experiments.
Maintenance of embryogenic cultures
Isolated somatic embryos and embryogenic groups were
excised from the original explants and subcultured sepa-
rately on MS medium supplemented with 0.44-lM BA plus
0.54 lM a-naphthaleneacetic acid (NAA, Duchefa, The
Netherlands). On this medium, embryo proliferation by
secondary embryogenesis was low. Therefore, the capacity
for secondary embryogenesis was investigated by subcul-
turing embryo clumps of 1–3 whole somatic primary
embryos at globular to early cotyledonary stage (embryo-
genic line S1-1) on five proliferation media consisting of
MS medium (half-strength macronutrients) supplemented
with 3-mM glutamine, 3 % sucrose, 0.6 % agar (A-1296,
Sigma-Aldrich, St. Louis, MO, USA) and different com-
binations of plant growth regulators (PGRs) (Table 3).
Similar proliferation media have been used to maintain
different embryogenic lines of Quercus robur (Mallon
et al. 2012).
In a further experiment, the embryo proliferation
capacity of cotyledon explants isolated from somatic
embryos was evaluated using the same proliferation media,
except the medium supplemented with 0.54-lM NAA
(Table 4).
To investigate the influence of genotype, groups of 1–3
whole somatic embryos at globular-early cotyledonary
stage, derived from three different embryogenic lines
named S1-1 (derived from one seed of tree Sarela 1) and
S2-1 and S2-2 (derived from two seeds of tree Sarela 2)
were cultured on proliferation medium supplemented with
0.44-lM BA.
In each experiment, three replicate Petri dishes with six
explants per dish (18 explants in total) were used. Each
Petri dish was considered as a single replicate in a com-
pletely randomized block design, and the experiment was
repeated twice. In embryo proliferation experiments, the
number of explants forming secondary embryos, the
number of embryos per embryogenic explant, and the
multiplication coefficient (MC) were recorded after
6 weeks of culture. The multiplication coefficient was
defined as the product of the proportion of explants pro-
ducing secondary embryos and the mean number of
embryos per embryogenic explant. In the genotype
Table 1 Effect of 2,4-D concentration, tree genotype, and size of zygotic embryo on the induction of somatic embryogenesis in Alnus glutinosa
Tree Collection datea (WPA) Length of zygotic embryo (mm)b Surviving explants with callus (%)c Embryogenesis induction (%)
2,4-D (lM)
0.90 2.26 4.52 9.05
Sarela 1 2 0.8 ± 0.2 50.0 4.2 – – –
Sarela 2 2 0.6 ± 0.1 44.3 10.0 6.6 – –
Sar 1 2 0.9 ± 0.2 45.8 – – – –
Sar 1 3 1.3 ± 0.2 45.8 – – – –
Source of variation df v2 P
2,4-D concentration (A) 3 8.177 B0.05
Tree genotype (B) 2 7.873 B0.05
AXB 9 6.980 ns
For each tree and collection date, 192 explants were cultured
Influence of tree genotype and 2,4-D concentration on embryogenic induction was evaluated by the v2 test; ns not significant
–, no responsea Weeks post-anthesisb Values are means ± standard error of 12 zygotic embryosc After 4 weeks of culture
Trees
123

experiment, for each embryogenic line the total number of
somatic embryos per embryogenic explant and the number
of cotyledonary somatic embryos per embryogenic explant
were recorded after 6 weeks of culture.
Histological analysis
A histological study was performed during secondary
embryo proliferation. Samples were fixed in a mixture of
formalin, glacial acetic acid and 50 % ethanol [1:1:18 (v/v/
v)], dehydrated through a graded n-butanol series and
embedded in paraffin. Sections (8 lm) were cut on a Re-
ichert-Jung rotary microtome and were later stained with
periodic acid-Schiff (PAS)–naphthol blue-black, which is
commonly used to reveal total insoluble polysaccharides
and total protein content of the cells (O0Brien and McCully
1981). The stained sections were mounted with Euckit�,
and photomicrographs were taken with an Olympus DP71
digital camera fitted to a Nikon-FXA microscope.
Macroscopic features were observed in a stereo micro-
scope (Olympus SZX9) and photographed with an Olym-
pus DP10 digital camera.
Embryo maturation and germination
Somatic embryos, developed to the cotyledonary stage,
were carefully separated from the proliferation embryo-
genic explants and cultured on MS medium (half-strength
macronutrients) supplemented with either 6 % sucrose or
3 % maltose. Selection of these media was based on the
findings of previous studies on somatic embryo maturation
in other woody species (Iraqui and Tremblay 2001; Cor-
redoira et al. 2006a). After 5 weeks, somatic embryos were
transferred to glass jars (300 ml) with 25 ml of germination
medium consisting of Woody Plant Medium (WPM; Lloyd
and McCown 1981) supplemented with 2 % glucose, 0.6 %
agar, 0.44-lM BA and 0.46-lM Zeatin trans isomer (Z;
Duchefa, The Netherlands). This germination medium was
selected as the most efficient for the proliferation of axillary
shoot cultures in Alnus glutinosa (San Jose et al. 2013).
Twenty-five somatic embryos (five embryos per jar) were
used for each maturation treatment, and the experiment was
repeated twice. The numbers of somatic embryos that
developed roots only and those that successfully converted
into plantlets were recorded after 6 weeks of culture.
Statistical analysis
The influence of the 2,4-D concentration and tree origin on
the percentage of somatic embryogenesis (Table 1) was
analyzed using a v2 test (P \ 0.05). The interaction
between both factors was analyzed using a log-linear
model (P \ 0.05). The influence of the collection date on
the percentage of somatic embryogenesis was evaluated
using a v2 test (P \ 0.05) (Table 2). Factors influencing
secondary embryogenesis (PGRs and the embryogenic line
genotype) were analyzed by one-way analysis of variance
(ANOVA I). Test for normality and homogeneity of vari-
ance were performed prior to ANOVA, and the least sig-
nificant difference (LSD) or Dunnett’s T3 test (P B 0.05)
was used to compare means in the case of homogeneous or
non-homogeneous variances, respectively. All data were
analyzed using IBM SPSS statistical version 19.0 for
Windows (Chicago, USA).
Results
Induction of somatic embryogenesis
In the first set of experiments, zygotic embryos collected
during 2 WPA (Fig. 1a, b) from two (Sarela 1 and Sarela 2)
of the three trees tested, proved amenable to induction of
somatic embryogenesis via a straightforward procedure of
Table 2 Embryogenic response (percentage and mean number of somatic embryos) of Alnus glutinosa zygotic embryos (Sarela 2 tree) as a
function of collection date (weeks post-anthesis, WPA), length of seeds and of zygotic embryos, and developmental stage of the explants
Collection datea
(WPA)
Length of seed
(mm)bLength of zygotic
embryo (mm)bDevelopmental stage of the
zygotic embryo
Embryogenesis
induction (%)
Number somatic embryos
per explant
1 2.0 ± 0.1 0.5 ± 0.1 Globular 10.0 1.5 ± 0.3
2 2.3 ± 0.1 0.7 ± 0.1 Globular-early heart 11.1 2.0 ± 0.5
3 2.5 ± 0.2 1.0 ± 0.1 Early cotyledonary 16.6 4.5 ± 0.3
4 2.8 ± 0.1 1.6 ± 0.1 Cotyledonary – –
5 2.9 ± 0.1 2.2 ± 0.2 Cotyledonary – –
6 2.9 ± 0.1 2.5 ± 0.1 Cotyledonary – –
Influence of collection date on the embryogenic induction was evaluated by the v2 test (P B 0.01)
–, no responsea Weeks post-anthesisb Values are means ± standard error of 12 seeds or zygotic embryos
Trees
123

culturing these explants on medium containing the auxin
2,4-D and the cytokinin BA (Table 1).
The contamination frequency ranged between 5 and
50 % (Table 1) and was related to the tree of origin (e.g., in
Sar 1 contamination rate reached 50 %). After 4 weeks of
culture on induction medium, the proportion of surviving
explants with callus ranged from 44 to 50 % (Table 1).
After transfer of explants to the expression medium sup-
plemented with 0.44-lM BA and incubation under light
conditions, callus culture progressed, and somatic embryos
and embryogenic structures were generated at the surface
of white nodules that developed from the initial callus
(Fig. 1c, d). Somatic embryos appeared after 2–4 months
of culture on expression medium. Somatic embryo
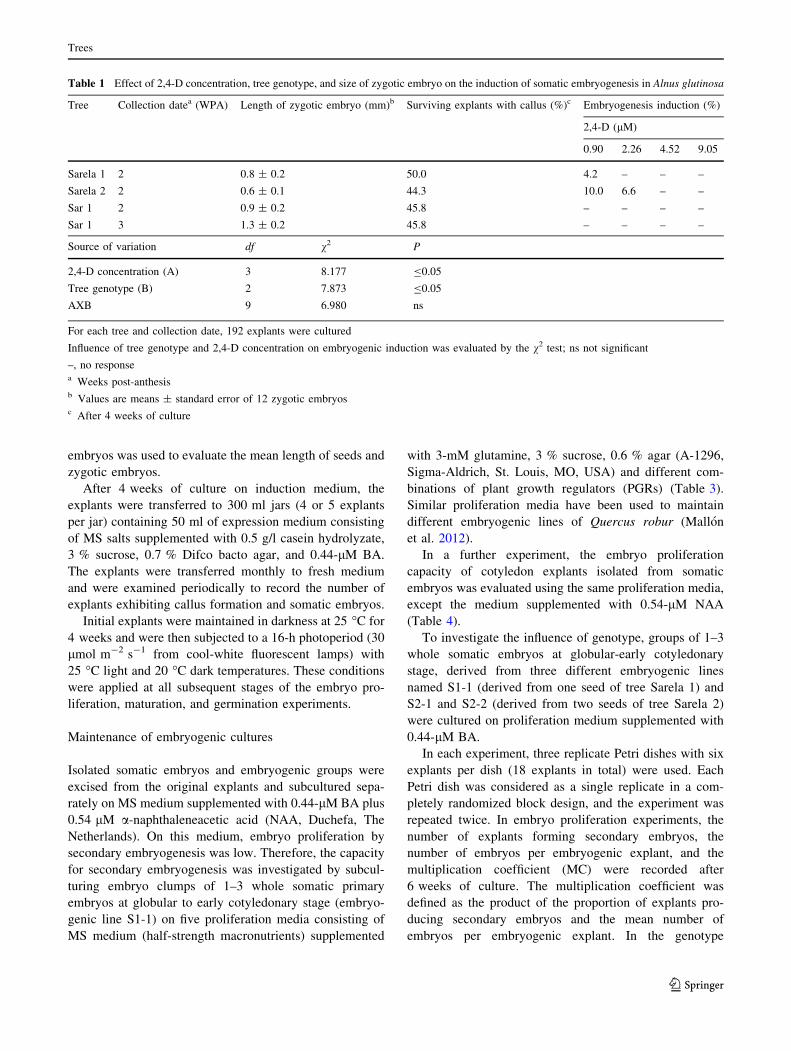
Fig. 1 Somatic embryogenesis in zygotic embryos of Alnus
glutinosa. a, b Appearance of a seed (a) and of an immature zygotic
embryo (b) collected during 2 WPA; c somatic embryos initiated
from immature zygotic embryo cultured on induction medium
supplemented with 0.90 lM 2,4-D; d somatic embryo developed
from the upper surface of a cotyledon of the zygotic embryo;
e secondary somatic embryos after 6 weeks of the culture on the
proliferation medium; f somatic embryos developed from a isolated
cotyledon explant after 6 weeks on proliferation medium; g plantlet
from a somatic embryo maturated on maltose 3 % and germinated on
WPM medium with 0.44-lM BA and 0.46-lM Z; h acclimatized
somatic plantlets following 1 month in the growth chamber. Scale
bars a–e 1 mm and h 1 cm
Trees
123

development was asynchronous and somatic embryos at
different stages of development, from globular-through
cotyledonary-shaped embryos, appeared by this time.
Although somatic embryos were formed indirectly from
embryogenic calli, direct somatic embryogenesis also
occasionally occurred in parts of the zygotic embryos that
were not in contact with the medium. Somatic embryos
were morphologically similar to zygotic embryos. How-
ever, anomalous morphologies, including embryos with
fused cotyledons and embryos with more cotyledons than
the usual number, were also observed.
Induction of somatic embryos was significantly affected
by the 2,4-D concentration (P B 0.05) and tree genotype
(P B 0.05), but there was no interaction between the two
factors. Induction of somatic embryos was achieved in two
of the three trees evaluated, and the frequency of induction
was highest in the material collected from the Sarela 2 tree
(16.6 %). Somatic embryos were only initiated after cul-
ture on media supplemented with 0.90 and 2.26 lM 2,4-D
(Table 1), and the best results (10.0 %) were obtained with
0.90 lM 2,4-D.
A second set of experiments was carried out to identify
the ‘developmental window’ during which the induction of
somatic embryogenesis is possible in black alder. Based on
the results obtained in the first experiment, the best tree
(Sarela 2) and induction medium supplemented with
0.90 lM 2,4-D with or without 2.22-lM BA were chosen
for this induction experiment. Cones were collected from
Sarela 2 at weekly intervals between 1 and 6 WPA. On
each collection date, zygotic embryos were identified by
morphological measurements including size and develop-
mental stage (Table 2). For embryo induction in medium
including 0.9 lM 2,4-D, the collection date significantly
affected (P B 0.01) the induction response, although there
were no significant differences for collection dates of
1–3 WPA. Immature zygotic embryos collected 1–3 weeks
following anthesis proved to be the best explants for
induction of somatic embryogenesis (Table 2). During this
period, the zygotic embryos are at globular (0.5 mm) to
early cotyledonary stages (1 mm). The highest frequency
of embryogenic cultures was observed with zygotic
embryos at early cotyledonary stage and of mean length
1.0 mm. This type of explant also produced the highest
number of somatic embryos (4.5 per explants) (Table 2).
In cones collected in September (4–6 WPA), zygotic
embryos were at the cotyledonary stage and were charac-
terized by a marked and concomitant increase in the length
of the zygotic embryos (1.6–2.5 mm) and seeds
(2.8–2.9 mm). These embryos had lost their ability to
induce somatic embryos and most of them failed to pro-
duce callus and rapidly became necrotic (Table 2).
When initial explants (1–6 WPA) were cultured on
medium including BA alone, no SEs were produced.
However, the zygotic embryos collected during 6 WPA
(i.e., mature seeds) geminated successfully on this medium.
Maintenance of embryogenic cultures
Initial somatic embryos were easily separated from original
explants and were used to establish different embryogenic
lines of A. glutinosa, which were multiplied by secondary
embryogenesis (Fig. 1e). Each embryogenic line was
derived from a single zygotic embryo and, therefore, the
genotype of each line is different.
To optimize the proliferation rates, groups of 1–3 whole
somatic embryos (globular to early cotyledonary stage)
from embryogenic line S1-1 were cultured in media sup-
plemented with different combinations of PGRs and also
on PGR-free medium (Table 3). The PGRs treatment had a
significant effect on the frequency of explants that gener-
ated secondary embryos (P B 0.0001) and on the mean
number of secondary embryos (P B 0.0001) produced.
Although secondary embryos were formed on all prolifer-
ation media tested (44.4–100 %), the highest frequency
was obtained with medium supplemented with 0.44-lM
BA (100 %) (Table 3). Proliferation media supplemented
with NAA produced the smallest number of explants that
formed secondary embryos. The largest numbers of SEs
were obtained with 0.44-lM BA plus 0.46-lM Z (9.0 per
explant) and with 0.44-lM BA (8.7 per explant), although
the best medium in terms of somatic embryo productivity
(MC) was that containing only BA. When alder somatic
Table 3 Effect of different secondary proliferation media on the
percentage of explants producing secondary embryos, mean number
of embryos per explant and coefficient (MC) in whole somatic
embryos (embryogenic line S1-1) of Alnus glutinosa after 6 weeks of
culture on different proliferation media
PGRs (lM) Explants producing
secondary embryos
(%)a
Number of
somatic
embryos per
explantb
MCc
No PGR 91.7 ± 5.1b 5.3 ± 0.7b 4.8
BA 0.44 100.0 ± 0.0b 8.7 ± 0.6c 8.9
BA 0.44 ? Z 0.46 97.2 ± 2.5b 9.0 ± 0.5c 8.7
BA 0.44 ? NAA 0.54 83.3 ± 5.4b 5.7 ± 0.4b 4.7
NAA 0.54 44.4 ± 3.2a 3.1 ± 0.4a 1.4
F test P B 0.0001 P B 0.0001
Values are means ± standard error of three replicates from two
experiments, with six explants in each replicate
Within each column, values followed by the same letter are not sig-
nificantly different at P = 0.05, according to T3 Dunnett’s T3 testa or
LSD testb
c MC multiplication coefficient, defined as the proportion of explants
producing secondary embryos and the mean number of embryos per
embryogenic explant
Trees
123
embryos were subcultured on a PGR-free medium or on
medium supplemented with 0.44-lM BA plus 0.54-lM
NAA, the multiplication values were intermediate, while
proliferation medium with NAA alone yielded the lowest
values in terms of MC.
Morphological observations showed that secondary
somatic embryos mainly arose from the abaxial surface of
the cotyledonary region (Fig. 1f), and therefore in the
subsequent experiment, isolated cotyledons were tested for
embryo proliferation. Cotyledons were subcultured on the
same proliferation medium evaluated for whole somatic
embryos, with exception of the medium containing 0.54-
lM NAA (Table 4), which yielded the lowest proliferation
rates (Table 3). The frequency of isolated cotyledons pro-
ducing somatic embryos ranged from 31 to 88 %, with a
mean number of 2.9–6.0 embryos, and both parameters
were significantly affected by the treatment (Table 4).
Taking into account the three parameters evaluated, the
best results were achieved with the 0.44-lM BA treatment.
Once the 0.44-lM BA treatment was defined as the most
suitable medium for proliferation of somatic embryos of
embryogenic line S1-1, we evaluated its effectiveness for
the other embryogenic lines. The genotype and total
number of somatic embryos produced per explants were
not significantly different, although the genotype had a
significant effect on the number of cotyledonary somatic
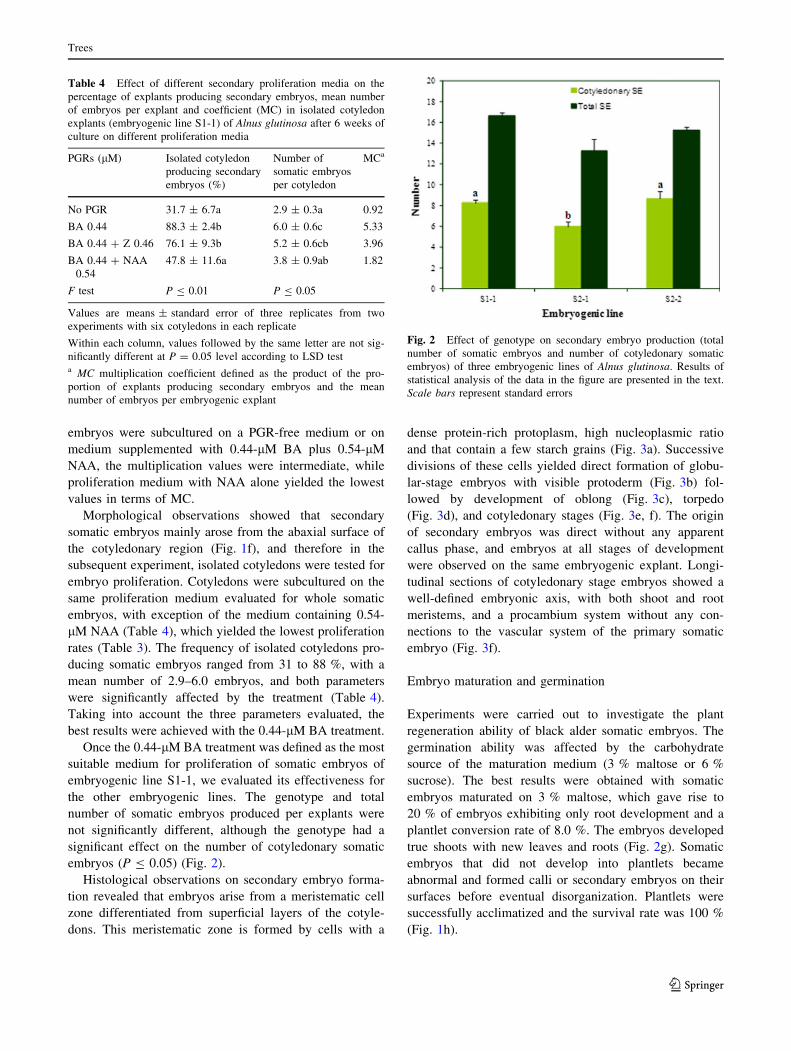
embryos (P B 0.05) (Fig. 2).
Histological observations on secondary embryo forma-
tion revealed that embryos arise from a meristematic cell
zone differentiated from superficial layers of the cotyle-
dons. This meristematic zone is formed by cells with a
dense protein-rich protoplasm, high nucleoplasmic ratio
and that contain a few starch grains (Fig. 3a). Successive
divisions of these cells yielded direct formation of globu-
lar-stage embryos with visible protoderm (Fig. 3b) fol-
lowed by development of oblong (Fig. 3c), torpedo
(Fig. 3d), and cotyledonary stages (Fig. 3e, f). The origin
of secondary embryos was direct without any apparent
callus phase, and embryos at all stages of development
were observed on the same embryogenic explant. Longi-
tudinal sections of cotyledonary stage embryos showed a
well-defined embryonic axis, with both shoot and root
meristems, and a procambium system without any con-
nections to the vascular system of the primary somatic
embryo (Fig. 3f).
Embryo maturation and germination
Experiments were carried out to investigate the plant
regeneration ability of black alder somatic embryos. The
germination ability was affected by the carbohydrate
source of the maturation medium (3 % maltose or 6 %
sucrose). The best results were obtained with somatic
embryos maturated on 3 % maltose, which gave rise to
20 % of embryos exhibiting only root development and a
plantlet conversion rate of 8.0 %. The embryos developed
true shoots with new leaves and roots (Fig. 2g). Somatic
embryos that did not develop into plantlets became
abnormal and formed calli or secondary embryos on their
surfaces before eventual disorganization. Plantlets were
successfully acclimatized and the survival rate was 100 %
(Fig. 1h).
Table 4 Effect of different secondary proliferation media on the
percentage of explants producing secondary embryos, mean number
of embryos per explant and coefficient (MC) in isolated cotyledon
explants (embryogenic line S1-1) of Alnus glutinosa after 6 weeks of
culture on different proliferation media
PGRs (lM) Isolated cotyledon
producing secondary
embryos (%)
Number of
somatic embryos
per cotyledon
MCa
No PGR 31.7 ± 6.7a 2.9 ± 0.3a 0.92
BA 0.44 88.3 ± 2.4b 6.0 ± 0.6c 5.33
BA 0.44 ? Z 0.46 76.1 ± 9.3b 5.2 ± 0.6cb 3.96
BA 0.44 ? NAA
0.54
47.8 ± 11.6a 3.8 ± 0.9ab 1.82
F test P B 0.01 P B 0.05
Values are means ± standard error of three replicates from two
experiments with six cotyledons in each replicate
Within each column, values followed by the same letter are not sig-
nificantly different at P = 0.05 level according to LSD testa MC multiplication coefficient defined as the product of the pro-
portion of explants producing secondary embryos and the mean
number of embryos per embryogenic explant
Fig. 2 Effect of genotype on secondary embryo production (total
number of somatic embryos and number of cotyledonary somatic
embryos) of three embryogenic lines of Alnus glutinosa. Results of
statistical analysis of the data in the figure are presented in the text.
Scale bars represent standard errors
Trees
123
Discussion
Induction of somatic embryos has been obtained for the
first time in a species of the genus Alnus using immature
zygotic embryos as the starting material. The main factors
determining the induction of somatic embryogenesis in
black alder are collection date (closely related to the
developmental stage of zygotic embryos), concentration of
PGRs and genotype. Alder somatic embryos were obtained
by culture of zygotic embryos on medium supplemented
with 2,4-D. The important role of 2,4-D in the induction of
somatic embryogenesis when zygotic embryo was used as
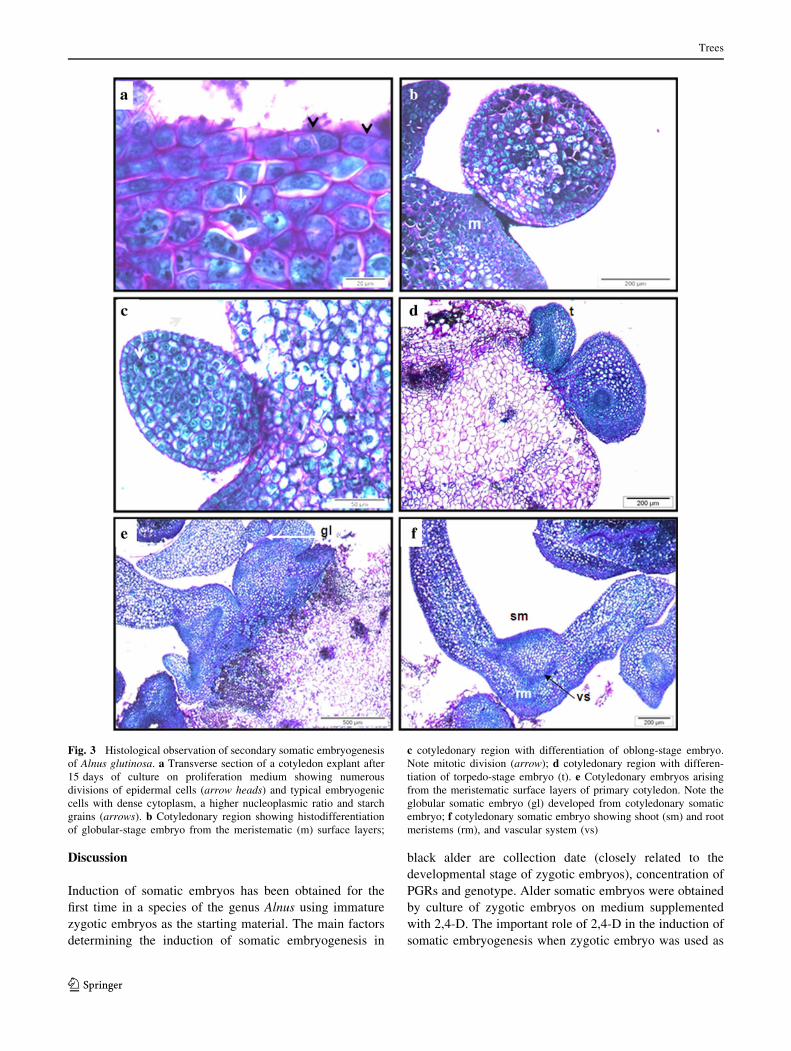
Fig. 3 Histological observation of secondary somatic embryogenesis
of Alnus glutinosa. a Transverse section of a cotyledon explant after
15 days of culture on proliferation medium showing numerous
divisions of epidermal cells (arrow heads) and typical embryogenic
cells with dense cytoplasm, a higher nucleoplasmic ratio and starch
grains (arrows). b Cotyledonary region showing histodifferentiation
of globular-stage embryo from the meristematic (m) surface layers;
c cotyledonary region with differentiation of oblong-stage embryo.
Note mitotic division (arrow); d cotyledonary region with differen-
tiation of torpedo-stage embryo (t). e Cotyledonary embryos arising
from the meristematic surface layers of primary cotyledon. Note the
globular somatic embryo (gl) developed from cotyledonary somatic
embryo; f cotyledonary somatic embryo showing shoot (sm) and root
meristems (rm), and vascular system (vs)
Trees
123

initial explants has been reported for many woody species
(Carraway and Merkle 1997; Canhoto et al. 1999; Capuana
et al. 2007; Vengadesan and Pijut 2009; Rocha et al. 2012).
This auxinic herbicide appears to create a double effect by
acting directly as an auxin while also triggering the pro-
duction of endogenous indole-3-acetic acid and even by
acting as a stress factor (Gaj 2004). Parallel activation of
auxin and stress signaling may be key events in cellular
adaptation via reprogramming the gene expression pattern,
cellular metabolism, and physiology (Pasternak et al.
2002). Thomas and Jimenez (2006) suggested that early
stages of somatic embryogenesis are characterized by the
induction of many stress-related genes, leading to the
hypothesis that SE is an extreme stress response in cultured
plant cells (Dudits et al. 1995).
In the present study, the highest frequency of somatic
embryogenesis was obtained in induction medium supple-
mented with 0.90 lM 2,4-D. This concentration of auxin
has also been found to be the most effective for inducing
somatic embryogenesis in zygotic embryos of elm (Cor-
redoira et al. 2002). Feher (2006) pointed out that auxins
and cytokinins are often used to induce somatic embryos
because they regulate the cell cycle and trigger cell divi-
sion. In alder, somatic embryogenesis took place in med-
ium containing 2,4-D and BA, but not in medium
supplemented with BA alone, suggesting that the presence
of 2,4-D was critical for embryogenesis. Similarly, Cor-
redoira et al. (2002) found that culture of zygotic embryos
of Ulmus minor and U. glabra on medium supplemented
with BA was not effective for induction of somatic
embryos. Although in some species, cytokinins rather than
auxins were found to be effective for induction of somatic
embryos (Kim et al. 2003; Chung et al. 2005; Pacheco et al.
2007), somatic embryogenesis is usually induced in the
presence of auxins alone or in combination with cytokinins.
The type of explant and the development stage appear to
be important factors determining the embryogenic capacity
(Gaj 2004). In black alder, there is a short period during
development of the zygotic embryo when it is possible to
establish embryogenic cultures. Zygotic embryos collected
in August (1–3 WPA), i.e., when they were at the globular
to early cotyledonary stages and of length 0.5–1 mm, were
responsive. The induction frequencies and the mean num-
ber of somatic embryos were highest when the zygotic
embryos were collected at early cotyledonary stage, before
deposition of storage reserves. We have previously found
that for elm, the best embryogenic induction rates were
achieved with zygotic embryos at cotyledonary stage,
before storage proteins began to accumulate (Corredoira
et al. 2002). In other species, such as Cercis canadensis
(Trigiano et al. 1999) and Tilia cordata (Karkonen 2000),
the zygotic embryos are embryogenically competent only
prior to maturation, and the appearance of significant
protein reserves signals a decline in embryogenic compe-
tence. The length of alder zygotic embryos collected in
September (4–6 WPA) increased considerably due to
elongation of cotyledons and the accumulation of reserve
substances (associated with seed maturation), and these
embryos were not able to initiate somatic embryos. Once
histodifferentiation of alder zygotic embryos up to the
cotyledonary stage has occurred (approximately 3 WPA),
the subsequent maturation during the remaining weeks
involves rapid elongation of the zygotic embryo, and
therefore length may be a good parameter for selecting
explants for initiating somatic embryogenesis.
Somatic embryos of black alder showed the potential to
initiate new cycles of somatic embryogenesis by repetitive
or secondary embryogenesis. Although proliferation med-
ium lacking plant growth regulators has been used to
maintain the embryogenic ability of various woody species
(Hernandez et al. 2003; Mauri and Manzanera 2005; Pintos
et al. 2008), in alder the application of embryo proliferation
medium supplemented with cytokinin (BA with or without
Z) significantly increased embryo production. Cytokinins
have been shown to promote secondary embryogenesis in
several woody species (Jimenez 2005). BA is commonly
used alone or in combination with other growth regulators,
mainly auxins, for the proliferation of somatic embryos
(Corredoira et al. 2003a, 2012; Valladares et al. 2006;
Cerezo et al. 2011); however, in black alder, the results
obtained with NAA alone or in combination with BA
indicated that exogenously supplied auxin was not neces-
sary for secondary embryo proliferation. Similarly, Mallon
et al. (2012) reported that the addition of NAA to the
proliferation medium has a negative effect on the multi-
plication of pedunculate oak somatic embryos. There are
evidences that once embryogenesis is induced, the role of
auxin changes and embryos begin to synthesize their own
auxins and, thus, require less additional auxin (Zimmerman
1993).
Secondary embryos were also obtained from isolated
cotyledons cultured on different proliferation media.
However, the multiplication coefficient values were lower
than those obtained when whole somatic embryos were
used as explants. The differences in proliferation ability
suggest that isolated cotyledons cannot synthesize the
appropriate PGRs or they do not produce sufficient levels
of PGRs. In addition, the number of embryogenic cells
involved in generating new secondary embryos is possibly
higher in whole somatic embryos than in isolated cotyledon
explants.
As expected, genotype had a clear influence on the
induction of black alder somatic embryos and also on the
embryo proliferation stage as the three different embryo-
genic lines tested yielded significantly different numbers of
cotyledonary somatic embryos. Hernandez et al. (2003)
Trees
123

also observed a genotype effect on the secondary embryo
multiplication of Quercus suber somatic embryos, based on
significant differences in the relative increase in fresh
weight and number of ‘detachable’ cotyledonary embryos.
We have also found significant differences in the prolif-
eration rates of four embryogenic lines derived from elm
zygotic embryos (Corredoira et al. 2003b).
Determination of the origin of the cell or groups of cells
involved in plant regeneration events is essential for the use
of somatic embryogenesis in clonal propagation and in
genetic transformation protocols (Rocha et al. 2012). In
black alder, histological observations showed that sec-
ondary embryos differentiated from the meristematic zones
originated from surface layers of cotyledon explants. These
meristematic zones were composed by cells with similar
embryogenic attributes as those reported for other woody
species, such as coffee (Quiroz-Figueroa et al. 2002),
pedunculate oak (Corredoira et al. 2006b), peach palm
(Steinmacher et al. 2007), and white oak (Corredoira et al.
2012). Black alder secondary embryos originated directly
from cotyledonary tissues, whereas the somatic embryos
originated from initial zygotic embryo explants have an
indirect pattern of differentiation and arise from callus
tissue formed from these zygotic embryos. Direct regen-
eration of secondary embryos is important for both plant
propagation and genetic transformation studies because
direct regeneration is less likely to generate somaclonal
variation than indirect regeneration from callus tissue (Gaj
2001).
In general, maturation of somatic embryos can be
achieved through treatment with abscisic acid (ABA) and/
or osmotic agents (such as high concentrations of sugars or
sugar alcohols), which induce water stress in the culture
medium (von Arnold et al. 2002). In black alder, somatic
plantlets were obtained after culture of somatic embryos on
maturation medium supplemented with 3 % maltose,
although conversion rates were low. Therefore, further
research on the maturation and germination of alder
somatic embryos is still required. In several woody species,
embryo conversion and germination are the limiting steps
in embryogenic systems (Merkle et al. 2005; Cerezo et al.
2011; Vieitez et al. 2012).
The protocol defined in this study may help to enhance
the propagation of Alnus glutinosa and other alder species,
and it will also provide a regeneration system for future use
in genetic manipulation for the introduction of genes con-
ferring resistance to pathogens such as Phytophthora alni.
However, application of the black alder embryogenic sys-
tem as a standard procedure or for use in genetic engi-
neering is still limited by the low plantlet conversion rate.
Efforts towards improving embryo maturation and
increasing the conversion rate should be made in future
research.
Acknowledgments We thank Ma Jose Cernadas and Jose Carlos
Suarez for their excellent technical and laboratory assistance. This
research was partially funded by INLUDES (Diputacion Provincial de
Lugo) and Xunta de Galicia (Spain) through Project
09MRU002400PR.
References
Barghchi M (1988) Micropropagation of Alnus cordata (Loisel.)
Loisel. Plant Cell Tissue Organ Cult 15:233–244
Brasier CM, Rose J, Gibbs JN (1995) An unusual Phytophthora
associated with widespread alder mortality in Britain. Plant
Pathol 44:999
Brasier CM, Kirk SA, Delcan J, Cooke DL, Jung T, Man in‘t Veld M
(2004) Phytophthora alni sp nova and its variants: designation of
a group of emerging heteroploid hybrid pathogens. Mycol Res
108:1172–1184
Canhoto JM, Lopes ML, Cruz GL (1999) Somatic embryogenesis
induction in Bay Laurel (Laurus nobilis L.). In: Jain SM, Gupta
PK, Newton RJ (eds) Somatic embryogenesis in woody plants,
vol 4. Kluwer Academic Publishers, Dordrecht, pp 341–367
Capuana M, Petrini G, Di Marco A, Giannini R (2007) Plant
regeneration of common ash (Fraxinus excelsior L.) by somatic
embryogenesis. In Vitro Cell Dev Biol Plant 43:101–110
Carraway DT, Merkle SA (1997) Plantlet regeneration from somatic
embryos of American chestnut. Can J Forest Res 27:1805–1812
Cavelier M, Claessens H, Etienne M (1999) Premier signalement du
Phytophthora de l’aulne (Alnus glutinosa) en Belgique. Parasi-
tica 55:63–71
Cech TL (1998) Phytophthora decline of alder (Alnus spp.) in Europe.
J Arboriculture 24:339–343
Cerezo S, Mercado JA, Pliego-Alfaro F (2011) An efficient regen-
eration system via somatic embryogenesis in olive. Plant Cell
Tissue Organ Cult 106:337–344
Chung HH, Chen JT, Chang WC (2005) Cytokinins induce direct somatic
embryogenesis of Dendrobium chiengmai pink and subsequent
plant regeneration. In Vitro Cell Dev Biol Plant 41:765–769
Claessens H, Oosterbaan A, Savill P, Rondeux J (2010) A review of
the characteristics of black alder (Alnus glutinosa (L.) Gaertn.)
and their implications for silvicultural practices. Forestry
83:163–175
Corredoira E, Vieitez AM, Ballester A (2002) Somatic embryogenesis
in elm. Ann Bot 89:637–644
Corredoira E, Ballester A, Vieitez AM (2003a) Proliferation,
maturation and germination of Castanea sativa Mill. somatic
embryos originated from leaf explants. Ann Bot 92:129–136
Corredoira E, Vieitez AM, Ballester A (2003b) Proliferation and
maintenance of embryogenic capacity in elm embryogenic
cultures. In Vitro Cell Dev Biol Plant 39:394–401
Corredoira E, Ballester A, Vieitez FJ, Vieitez AM (2006a) Somatic
embryogenesis in chestnut. In: Mujib A, Samaj J (eds) Somatic
embryogenesis. Plant cell monographs. Springer, Berlin,
pp 177–199
Corredoira E, Valladares S, Vieitez AM (2006b) Morphohistological
analysis of the origin and development of somatic embryos from
leaves of mature Quercus robur. In Vitro Cell Dev Biol Plant
42:525–533
Corredoira E, Ballester A, Vieitez AM (2008) Thidiazuron-induced
high-frequency plant regeneration from leaf explants of Pau-
lownia tomentosa mature trees. Plant Cell Tissue Organ Cult
95:197–208
Corredoira E, San-Jose MC, Vieitez AM (2012) Induction of somatic
embryogenesis from different explants of shoot cultures derived
from young Quercus alba trees. Trees 26:881–891
Trees
123

Dudits D, Gyorgyey J, Bogre L, Bako L (1995) Molecular biology of
somatic embryogenesis. In: Thorpe TA (ed) In vitro embryo-
genesis in plants. Kluwer Academic Publishers, Dordrecht,
pp 267–308
Feher A (2006) Why somatic plant cells start to form embryos? In:
Mujib A, Samaj J (eds) Somatic embryogenesis. Plant cell
monographs. Springer, Berlin, pp 85–101
Gaj MD (2001) Direct somatic embryogenesis as a rapid and efficient
system for in vitro regeneration of Arabidopsis thaliana (L.)
Heynh. Plant Cell Tissue Organ Cult 64:39–46
Gaj MD (2004) Factors influencing somatic embryogenesis induction
and plant regeneration with particular reference to Arabidopsis
thaliana (L.) Heynh. Plant Growth Regul 43:27–47
Garton S, Hosier MA, Read PE, Farnham RS (1981) In vitro
propagation of Alnus glutinosa Gaertn. Hortic Sci 16:758–759
Gibbs JN, van Dijk C, Webber JF (2003) Phytophthora disease of
alder in Europe. Forestry Commission Bulletin 126. Forestry
Commission, Edinburgh
Hernandez I, Celestino C, Alegre J, Toribio M (2003) Vegetative
propagation of Quercus suber L. by somatic embryogenesis. II.
Plant regeneration from selected cork oak trees. Plant Cell Rep
21:765–770
Iraqui D, Tremblay FM (2001) The role of sucrose during maturation
of black spruce (Picea mariana) and white spruce (Picea glauca)
somatic embryos. Physiol Plant 111:381–388
Jimenez VM (2005) Involvement of plant hormones and plant growth
regulators on in vitro somatic embryogenesis. Plant Growth
Regul 47:91–110
Jung T, Blaschke M (2004) Phytophthora root and collar rot of alders
in Bavaria: distribution, modes of spread, and possible manage-
ment strategies. Plant Pathol 53:197–208
Karkonen A (2000) Anatomical study of zygotic and somatic embryos
of Tilia cordata. Plant Cell Tissue Organ Cult 61:205–214
Kim SW, Oh SC, Liu JR (2003) Control of direct and indirect somatic
embryogenesis by exogenous growth regulators in immature
zygotic embryo cultures of rose. Plant Cell Tissue Organ Cult
74:61–66
Lall S, Mandegaran Z, Roberts AV (2005) Shoot multiplication in
cultures of mature Alnus glutinosa. Plant Cell Tissue Organ Cult
83:347–350
Lloyd GB, McCown BH (1981) Commercially feasible microprop-
agation of mountain laurel (Kalmia latifolia) by use of shoot tip
culture. Proc Int Plant Propagators Soc 30:421–437
Mallon R, Covelo P, Vieitez AM (2012) Improving secondary
embryogenesis in Quercus robur: application of temporary
immersion for mass propagation. Trees 26:731–741
Mauri PV, Manzanera JA (2005) Protocol of somatic embryogenesis:
holm oak (Quercus ilex L.). In: Jain SM, Gupta PK (eds)
Protocol for somatic embryogenesis in woody plants. Springer,
Dordrecht, pp 469–482
Merkle SA, Nairn CJ (2005) Hardwood tree biotechnology. In Vitro
Cell Dev Biol Plant 41:602–619
Merkle SA, Montello PM, Xia X, Upchurch BL, Smith DL (2005)
Light quality treatments enhance somatic seedling production in
three southern pine species. Tree Physiol 26:187–194
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
O0Brien TP, McCully ME (1981) The study of plant structure:
principles and selected methods. Hermacarphi. Pty. Ltd.,
Melbourne
Oliveira RS, Castro PM, Dodd JC, Vosatka M (2005) Synergistic
effect of Glomus intraradices and frankia spp. on the growth and
stress recovery of Alnus glutinosa in an alkaline anthropogenic
sediment. Chemosphere 60:1462–1470
Pacheco G, Gagliardi RF, Carneiro LA, Callado CH, Valls JFM,
Mansur E (2007) The role of BAP in somatic embryogenesis
from seed explants of Arachis species from sections Erectoides
and Procumbentes. Plant Cell Tissue Organ Cult 88:121–126
Pasternak TP, Prinsen E, Ayaydin F, Miskolczi P, Potters G, Asard H,
van Onckelen HA, Duditss D, Feher A (2002) The role of auxin,
pH, and stress in the activation of embryogenic cell division in
leaf protoplast derived cells of alfalfa. Plant Physiol
129:1807–1819
Perinet P, Lalonde M (1983) In vitro propagation and nodulation of
the actinorhizal host plant Alnus glutinosa (L.) Gaertn. Plant Sci
Lett 29:9–17
Perinet P, Tremblay FM (1987) Commercial micropropagation of five
Alnus species. New Forest 3:225–230
Pintos G, Park Y-S, Silva S, Neves L, Araujo C, Santos C (2008)
Factors affecting maintenance, proliferation and germination of
secondary embryos of Eucalyptus globulus Labill. Plant Cell
Tissue Organ Cult 95:69–78
Prada MA, Arizpe D (2008) Alnus glutinosa (L.) Gaertn. In: Prada
MA, Arizpe D (eds) Riparian tree and shrub propagation
handbook. Generalitat Valenciana, Valencia, pp 22–26
Quiroz-Figueroa FR, Fuentes-Cerda CFJ, Rojas-Herrera R, Loyola-
Vargas VM (2002) Histological studies on the developmental
stages and differentiation of two different somatic embryo-
genesis systems of Coffea arabica. Plant Cell Rep 20:
1141–1149
Rocha DI, Vieira LM, Tanaka FAO, Campos da Silva L, Campos
Otoni W (2012) Somatic embryogenesis of a wild passion fruit
species Passiflora cincinnata Masters: histocytological and
histochemical evidences. Protoplasma 249:747–758
San-Jose MC, Romero L, Janeiro LV (2012) Effect of indole-3-
butyric acid on root formation in Alnus glutinosa microcuttings.
Silva Fenn 46:643–654
San-Jose MC, Janeiro LV, Corredoira E (2013) In vitro conservation
of threatened black alder. Silva Fenn (in press)
Solla A, Perez-Sierra A, Corcobado T, Haque MM, Diez JJ, Jung T
(2010) Phytophthora alni on Alnus glutinosa reported for the
first time in Spain. Plant Pathol 59:789
Steinmacher DA, Cangahuala-Inocente GC, Clement CR, Guerra MP
(2007) Somatic embryogenesis from peach palm zygotic
embryos. In Vitro Cell Dev Biol Plant 43:124–132
Streito J-C, Legrand PH, Tabary F, Jarnouen de Villartay G (2002)
Phytophthora disease of alder (Alnus glutinosa) in France:
investigations between 1995 and 1999. Forest Pathol 32:179–191
Thomas C, Jimenez VM (2006) Mode the action of plant hormones
and plant growth regulators during induction of somatic
embryogenesis: molecular aspects. In: Mujib A, Samaj J (eds)
Somatic embryogenesis. Plant cell monographs. Springer, Ber-
lin, pp 157–175
Tremblay FM, Perinet P, Lalonde M (1986) Tissue culture of Alnus
spp. with regard to symbioses. In: Bajaj YPS (ed) Biotechnology
in agriculture and forestry, vol 1. Trees 1. Springer, Berlin,
pp 87–100
Trigiano RN, Buckley LG, Merkle SA (1999) Somatic embryogenesis
in woody legumes. In: Jain SM, Gupta PK, Newton RJ (eds)
Somatic embryogenesis in woody plants. Kluwer Academic
Publishers, Dordrecht, pp 189–208
Valladares S, Sanchez C, Martınez MT, Ballester A, Vieitez AM
(2006) Plant regeneration through somatic embryogenesis from
tissues of mature oak trees: true-to-type conformity of plantlets
by RAPD analysis. Plant Cell Rep 25:879–886
Vengadesan G, Pijut PM (2009) Somatic embryogenesis and plant
regeneration of northern red oak (Quercus rubra L.). Plant Cell
Tissue Organ Cult 97:141–149
Vieitez AM, Corredoira E, Martınez T, San-Jose MC, Sanchez C,
Valladares S, Vidal N, Ballester A (2012) Application of
biotechnological tools to Quercus improvement. Eur J Forest
Res 131:519–539
Trees
123

von Arnold S, Sabala I, Bozhkov P, Dyachok J, Filonova L (2002)
Developmental pathways of somatic embryogenesis. Plant Cell
Tissue Organ Cult 69:233–249
Webber J, Gibbs J, Hendry S (2004) Phytophthora disease of alder.
Forestry Commission Information Note 6. Edinburgh
Welander M, Welander NT, Brackman AS (1989) Regulation of
in vitro shoot multiplication in Syringa, Alnus and Malus by
different carbon sources. J Hortic Sci 64:361–366
Zimmerman JL (1993) Somatic embryogenesis: a model for early
development in higher plants. Plant Cell 5:1411–1423
Trees
123