Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 227~597-606 (1984)
Somatosensory Thalamocortical Connections in the Racoon: An HRP Study
SUSAN WARREN AND BENJAMIN H. PUBOLS, JR. Department of Anatomy, College of Medicine, Pennsylvania State University, Hershey,
Pennsylvania 17033
ABSTRACT In the North American raccoon (Procyon Zotor), representations of the
glabrous surfaces of the hand digits are found within separate subnuclei of the thalamic ventrobasal complex (VB) and on separate subgyri of the somatosensory cortex (SmI). In the present study, the retrograde transport of horseradish peroxidase from SmI to VB was utilized to study relationships between physiologically identified cortical subgyri and somatotopically cor- responding thalamic subnuclei.
Single large or multiple small injections confined to a single gyral crown led to retrograde labeling of large groupings of cells filling the entire VB subnucleus for the appropriate digit. In the aggregate, the regions of label appeared as thin, wedge-shaped sheets extended in the dorsoventral and anteroposterior dimensions, but flattened mediolaterally, and curving to form a laterally directed convexity; these appear to correspond to the lamel- lae of monkey VB described by others. These large injections led to labeling of approximately 80% of all large (18-30-pm diameter) cells within the lamella. Single, small, focal injections of a gyral crown led to variable amounts of labeling, ranging from an entire digital lamella to only a small focal cluster of cells. No evidence was obtained for the existence of antero- posteriorly extending “rods” of cells, as reported in primates. Finally, there was a sparse, but consistent labeling of cells of the posterior nuclear group (Po) following gyral crown injections.
These results are in agreement with expectations based on prior electro- physiological studies of raccoon VB and SmI, as well as prior anatomical studies of thalamocortical relationships.
Key words: somatosensory cortex, somatosensory thalamus, thalamocortical connections, ventrobasal complex posterior nuclear group
Representation of the glabrous surfaces of the raccoon’s hand is precisely somatotopically organized in both the thalamic ventrobasal complex (VB) and primary somato- sensory cortex (SmI). In VB, digital representations are found within separate subnuclei, demarcated by myeli- nated fiber laminae (Welker and Johnson, ’65). In SmI, each digit is represented on a separate subgyrus-glabrous sur- faces on gyral crowns, hairy surfaces within demarcating sulci (Welker and Seidenstein, ’59). Additionally, it has been reported that thalamocortical afferents terminate preferentially within gyral crowns (Welker and Seiden- stein, ’59).
Recent studies by Jones and co-workers (Jones et al., ’79, ’82; Jones and Friedman, ’82), combining both retrograde and anterograde tract-tracing techniques with microelec- trode-recording procedures, have shown, in both Old World (Macaca) and New World (Saimiri) monkeys, that “rods” of
relay cells running throughout most or all of the anteropos- terior dimension of VB project to discrete cortical cell col- umns. Each rod or column has as its physiological counterpart the representation of a fairly restricted region of the animal’s body surface. Further, the thalamic rods aggregate into gently curving lamellae, or sheets, each corresponding to the representation of a larger body part,
Accepted April 12, 1984.
Susan Warren’s present address is Department of Physiology, New York University School of Medicine, 550 First Avenue, New York, NY 10016. Address reprint requests there.
Benjamin H. Pubols, Jr.’s present address is Neurological Sci- ences Institute, Good Samaritan Hospital and Medical Center, 1120 N.W. Twentieth Avenue, Portland, OR 97209.
0 1984 ALAN R. LISS, INC.

598
such as a hand or foot digit, or the tail. Such lamellae are somatotopically organized in a manner originally described by Mountcastle and colleagues in a number of mammalian species (Rose and Mountcastle, '59). The projection of these thalamic lamellae then defines the plan of somatotopic organization in primary somatosensory cortex (Kaas, '83).
Because of the unique relationship between physiological subdivisions (digital representations) and anatomical sub- dividers (thalamic fiber laminae, cortical sulci), the raccoon seemed an ideal nonprimate animal in which to test the concepts outlined in the preceding paragraph. The retro- grade transport of horseradish peroxidase (HRP) from SmI to VB was utilized with the broad expectation that injec- tions into physiologically identified digital subgyri would result in labeling of neurons in the somatotopically corre- sponding thalamic subnuclei (lamellae). It was further pre- dicted that large injections, encompassing an entire gyral crown, would label most or all of the appropriate subnu- cleus, while more discrete, focal injections would lead to more circumscribed labeling of clusters of thalamic cells. Finally, it was predicted that injections into sulcal fundi (digital hairy skin representations) would lead to more dif- fuse, but sparser labeling within VB than would equivalent injections into gyral crowns (digital glabrous skin representations).
MATERIALS AND METHODS Subjects and preparation
Seven adult raccoons (Procyon lotor) were used for this project. Data will be reported on the results of horseradish peroxidase (HRP) injections into 16 sites in ten hemi- spheres. All animals were anesthetized with pentobarbital sodium, administered intraperitoneally (42 mgkg initial dosage), and the head was secured in a stereotaxic appara- tus. The hand representational area of primary somatosen- sory cortex (SmI) was surgically exposed on one or both sides. Figure 1 gives the cortical exposure necessary for identification and injection of the hand digit subgyri; the exposure extends rostrally to the central sulcus homologue (vertical ramus of coronal sulcus), its lateral margin borders the coronal sulcus, and the exposure extends caudally to the ansate sulcus. The exposed digital subgyri and associ- ated sulci were identified with the aid of an acrylic brain cast (Welker, '67). Identity of the subgyri was confirmed by utilizing a varnish-coated gross electrode to record the evoked potential (slow wave) response to mechanical stim- ulation of the various hand digits (Fig. 1).
HRP injection procedures Injections, each consisting of 0.1-0.6 p1 of a 30-40%
aqueous solution of horseradish peroxidase (Sigma, type VI) were made under direct visual guidance. They were hy- draulically delivered with a micropipette (tip diameter = 60-100 pm) attached via polyethylene tubing to a micro- manipulator-held 10-pl Hamilton syringe. The tubing con- tained the dye oil red 0 in paraffin oil as a visual marker. Injections were made at a depth of 1.5-2.5 mm below the pial surface. The HRP solution was delivered with some residual mineral oil in order to prevent a possible reflux of HRP. A postinjection period of 15-20 minutes was routinely observed prior to withdrawing the pipette.
Tissue processing and histological analysis Following survival times of 24-48 hours, the animals
were reanesthetized and perfused transcardially with a sa-
S. WARREN AND B.H. PUBOLS, JR.
Fig. 1. Photograph of right somatosensory cortex of raccoon, showing cortical exposure necessary for injections of HRP into digital subgyri of the SmI hand area. Anterior a t right, medial a t top. Individual subgyri are identified by number, corresponding to hand digits (1, thumb; 5 , little finger). Scale: each division = 1 mm. Below the photograph is an example of an evoked potentials record used to identify each suhgyrus prior to HRP injection. In the example shown, the contralateral (left) digit 4 was mechan- ically stimulated, and the record was obtained from a surface electrode located over the indicated digit 4 suhgyrus shown in the phototograph. T, 50 msec; A, 100 FV.
line-heparin rinse, followed by a double aldehyde fixative solution consisting of 4% glutaraldehyde and 1% parafor- maldehyde in a 0.1 M phosphate buffer at pH 7.4 (room temperature) and a postfixative rinse of 10% sucrose-0.1 M phosphate buffer solution at 4°C (modified procedure of Rosene and Mesulam, '78). The brain was removed and placed in fresh cold 10% sucrose buffer for 24 hours. Tissue was then transferred to a 30% sucrose buffer solution for 48-72 hours.
Frozen sections in the transverse (N = 21, sagittal (N = 4), or horizontal (N = 1) planes were cut at 50 pm and placed in room temperature 0.1 M phosphate buffer con- taining no sucrose. Every third, fourth, and fifth section was reserved for processing with one of two chromagens: tetramethylbenzidine (TMB; Mesulam, '78) or diaminoben- zidine (DAB; Mesulam and Rosene, '77). After termination of the reaction (acetate buffer-TMB; phosphate buffer-DAB), sections were serially mounted on gelatin-coated slides, subbed with chrome-alum and allowed to air dry. The sec- tions were counterstained (neutral red-TMB; cresyl violet- DAB) to facilitate the identification of the cytoarchitecture of the area under investigation.
Sections were viewed and photographed on an Olympus BHA research microscope with both brightfield and dark-

THALAMOCORTICAL PROJECTIONS IN RACCOON
field illumination capabilities. Cell labeling within the thalamus, indicative of intracellular retrograde transport, appeared as dark blue-black granules (TMB), or dark brown granules (DAB) contained within the cytoplasm of the neu- ronal soma and extending into the dendrites. Sections were examined for the appearance and organization of labeled cells and neuropil in relation to thalamic architecture. Se- rial sections through somatosensory cortex were also ex- amined, to determine the extent of effective injection site.
Normal reference material The brains of three raccoons were available for study of
the normal architecture of the somatosensory thalamus. The brains were embedded in celloidin and cut, one each, in transverse (25 pm), horizontal (30 pm), and sagittal (30 pm) planes. Every ninth and tenth section was reserved for staining, and alternate, adjacent sections were stained by the Nissl method (thionin or cresyl violet) for the study of cytoarchitecture or the Weil method (hematoxylin) for the study of myeloarchitecture. The photomicrographs appear- ing in Figure 2 were taken through a Leitz Aristophot projection enlarger equipped with macro-dia apparatus.
RESULTS Normal architecture of raccoon
ventrobasal complex The normal cyto- and myeloarchitecture of the raccoon
ventrobasal complex and surrounding regions has been characterized by Welker and Johnson (‘651, and more re- cently summarized by Sakai (‘82). In order to facilitate description of the present experimental results, salient ar- chitectural features of VB will be briefly summarized here and illustrated in Figure 2, which depicts VB and surround- ing nuclei in fiber- and cell-stained material in the horizon- tal plane.
The most striking feature of the thalamic ventrobasal complex of the raccoon is the aggregation of cell clusters into distinct subnuclei, or lamellae, separated one from another by intervening myelinated fiber laminae. Such subnuclei are recognizable in Nissl material in transverse, horizontal, and sagittal planes, and the fiber laminae are prominent in all three planes in Weil material. Welker and Johnson (’65) have shown that these fiber laminae serve to demarcate the representations of adjacent hand digits, as well as to separate the representation of digit 5 from that of the leg laterally, and the representation of digit 1 from the trigeminal representation medially (the classical ar- cuate lamina).
Examination of Nissl material (Fig. 2B) reveals large (18- 30-pm diameter), darkly staining perikarya arranged in clusters. Intermingled with the clusters are smaller (10-12 pm in diameter), pale-staining cells (approximately 25% of the total number of cells). Examination of Weil material (Fig. 2A) indicates two types of myelinated fiber groupings: (1) the curviplanar laminae, or sheets, referred to above, which represent the final resorting of medial lemniscal fibers within VB before their termination in the somatotop- ically appropriate subnucleus; and (2) more thickly myeli- nated thalamocortical and corticothalamic fiber bundles, arcing laterally and rostrally in route to and from the internal capsule.
The nucleus ventralis lateralis (VL) is located dorsal and anterior to VB; cells are less densely packed, more uni- formly distributed, and paler staining than those of VB. Laterally, VB is segregated from the thalamic reticular
599
nucleus by the external medullary lamina, and medially it lies adjacent to the nucleus centrum medianum (CM). Ven- trally, VB is bounded by the nucleus ventralis posterior inferior (VPI) and the ventral continuation of the external medullary lamina. It is only at its posterior border that the boundaries of VB are somewhat blurred. Among nuclei near and posterior to VB is the posterior nuclear complex (Po) of Poggio and Mountcastle (‘60).
Characteristics of injection sites Regardless of the number of injections per gyral crown or
the volume of HRP per injection site, the extent of effective injection (region of tissue damage, plus central core of dense reaction product, surrounded by a zone of less dense second- ary diffusion) never exceeded 2-3 mm in diameter, and the reaction product was always confined to the intended subgyrus. Figure 3A illustrates the effective injection site of the digit 3 subgyrus following injection of a large amount of HRP (0.6 pl). This response is typical of the reaction found following large (0.4-0.6 pl) injection quantities, whether delivered at one or multiple sites within the gyrus. Injection of smaller quantities of HRP (0.1-0.3 p1 single injections) led to variable spread within the cortex. Occa- sionally the extent of cortical diffusion was sufficient to yield an effective injection site approaching in size that seen after larger injections, but more typically (Fig. 3B,C), the effective site remained small and focal. Nevertheless, in five of ten such cases (see below), there was widespread, but relatively sparse, labeling within a VB lamella.
Labeling of ventrobasal lamellae (subnuclei) Single large or multiple small injections (total volume =
0.4-0.6 pl) confined to a single gyral crown lead to the retrograde labeling of a large grouping of thalamic cells traversing much or all of both the anteroposterior and dor- soventral extents of VB, but relatively restricted in the mediolateral dimension. Viewed in transverse section, the labeled cells form a crescent-shaped lamella; its concave border is directed medially and its convexity directed lat- erally. The lamella is expanded dorsally and tapers ven- trally to form a wedge.
Figure 4 shows the VB labeling in low-power, transverse sections, following large injections into the crowns of two digital subgyri, those containing the representations of dig- its 3 and 5 . The more lateral crescent (to the right) corre- sponds to the digit 5 subnucleus and the more medial crescent (to the left) corresponds to the digit 3 subnucleus. The unlabeled region between them is the region of repre- sentation of digit 4.
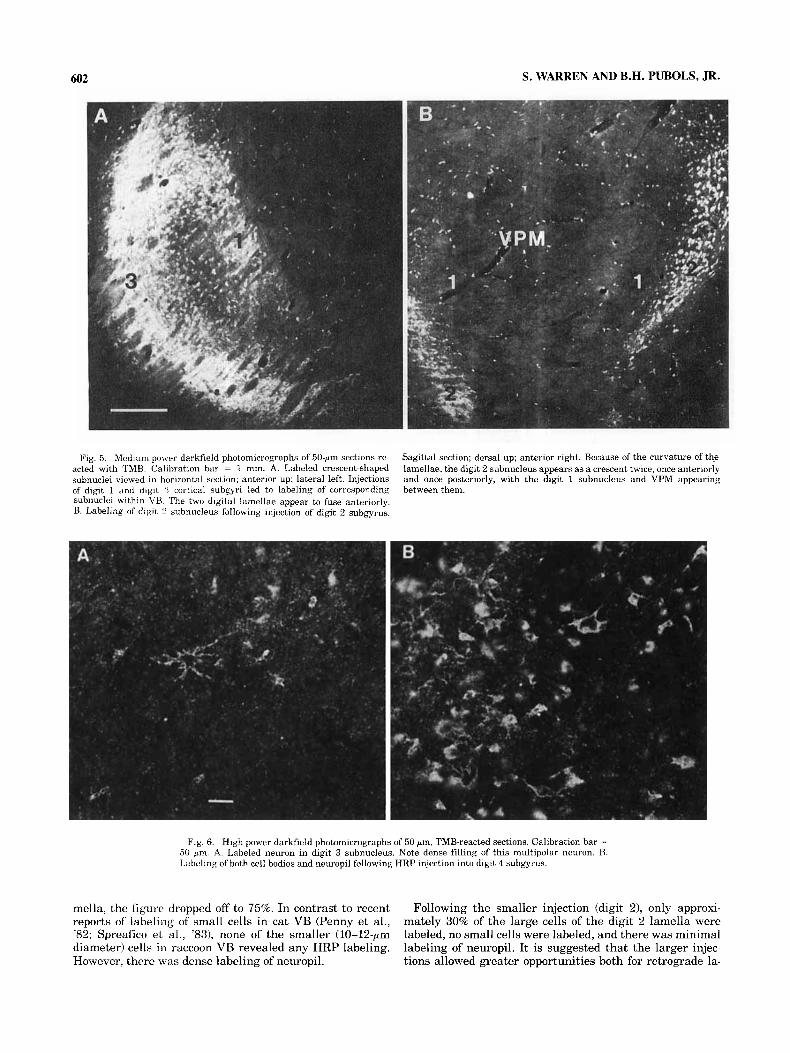
Occasionally, smaller, single injections (total volume = 0.1-0.3 pl), exhibiting minimal diffusion within the gyral crown, would also lead to labeling of an entire lamella. Figure 5A is a medium-power, darkfield photomicrograph of VB labeling following single small injections into the digit 1 and digit 3 cortical subgyri. This is a horizontal section, with lateral to the left and anterior to the top. Two distinct labeled lamellae can be seen-that of digit 3 lat- erally, that of digit 1 medially. Anteriorly, the two lamellae appear to fuse.
Figure 5B shows labeling of the digit 2 subnucleus, in sagittal section, following injection of the digit 2 subgyrus. Because of the “saucer shape” appearance of the lamellae, the digit 1 representation, as well as that of the trigeminal system (VPM), appear as if contained within the digit 2 crescent or subnucleus.

Fig. 2. Photoinicro@~~phs of' adjacent 30-pm horizontal sections through VB. Lateral toward left, anterior a t top. Calibration bar = 1 mm. A. Hematoxylin stain. W. Cresyl violet stain. The laterally convex anteropos- teriorly extending laminae are prominent in A. They separate the subnuclei
identified in B. L, leg representation within VB; 5 , 4 , 3 , 2 , 1, representations of hand digits 5-1, respectively; VPM, nucleus ventralis posteromedialis: VL, ventralis lateralis; MD, medialis dorsalis; CM, nucleus centrum me- dianum; Po, posterior nuclear group; MGN, medial geniculate nucleus.
A C
Fig. 3. Effective HRP injectlon sites in three digital subgyri. The core of the injection (tissue injury plus dense reaction product) is indicated by solid black, regions of secondary diffusion by shading. All drawings traced from DAB-reacted tissue sections, counterstained with cresyl violet. Drawings are of sections showing the greatest amount of spread of HRP. In each case, the micropipette track passed through nearby sections. A. Transverse sec- tion, showing large 10.6 pli injection into digit 3 subgyrus. There is major spread of HlZP so that the effective injection encompasses nearly the entire
subgyrus. VB labeling is shown in Figure 4. B. Sagittal section, showing small (0.1 pl) injection into digit 4 subgyrus. Although there is some spread, it is not as great as in A, and includes only portions of the suhgyral crown. VB labeling is shown in Figure 6B. C. Horizontal section, showing small (0.1 pl) injection into digit 3 subgyrus. There is no secondary diffusion, so the effective injection site remains small and focal. VB labeling is shown in Figures 5A and 6A. A, anterior; D, dorsal; L, lateral.

THALAMOCORTICAL PROJECTIONS IN RACCOON 601
Fig. 4. Labeling of the digit 3 (left) and digit 5 (right) subnuclei of VB resulting from large HRP injections into the cortical digit 3 and digit 5 subgyral crowns. Four transverse sections (A-D) in anteroposterior sequence; brightfield photomicrographs of 50-pm sections reacted with TMB but not counterstained. The unlabeled region between the digit 3 and digit 5 subnuclei is the digit 4 subnu- cleus. Calibration bar = 1 mm.
Viewed at higher power (Fig. 6A,B), labeled VB cells are seen to be relatively large, but of variable diameter (18-30 pm), multipolar in shape, and densely filled with reaction product. Figure 6B also illustrates labeling of neuropil.
Labeling of ventrobasal cell clusters Focal single injections of digital cortex lead, in half of the
cases, to circumscribed labeling of discrete cell clusters within the appropriate digital subnucleus of VB. In Figure 7 are shown a series of four sagittal planes through VB, progressing lateral to medial, indicating the labeling of the digit 5 crescent following multiple small injections (total volume = 0.5 p l ) of the digit 5 subgyrus, and the labeling of a small portion of the digit 2 subnucleus following a single small (0.1 pl) injection of the digit 2 subgyrus. In Figure 7A, only the lateralmost convexity of the digit 5 crescent is visible. Progressing medially (Fig. 7B-D), the crescent divides into anterior and posterior segments (cf. Fig. 5B), and, appearing in Figure 7C and D, a small spot is seen, localized within the digit 2 subnucleus. In Figure 7B and C, located anterodorsally, fibers traversing the tha-
lamic reticular nucleus, to and from the internal capsule, can be seen. Our material did not reveal any rods coursing throughout the entire anteroposterior extent of VB, as re- ported by Jones and colleagues (Jones et al., '79, '82).
Density of cell labeling within the ventrobasal complex
Not only were there differences in the extent of a lamella labeled as a function of the size of the injection, but there were differences in proportions of labeled cells. The case depicted in Figure 7 showed HRP labeling following both large (digit 5 ) and small (digit 2) cortical injections and the tissue was counterstained with neutral red, making possi- ble a count of both labeled and unlabeled cells. All cells were counted in sections spaced 0.25 mm apart throughout the entire approximately 3 mm mediolateral extent of la- beling. Following the large injection (digit 5 ) 80% of the large-diameter cells (18-30-pm diameter) showed HRP granules. The density of labeling varied with position within the lamella. In the center, approximately 85% of the large cells were labeled, while toward the edge of the la-
602 S. WARREN AND B.H. PUBOLS, JR.
Fig. 5. Mediun-power darkfield photomicrographs of 50-pm sections re- acted with TMB. Calibration bar = 1 mm. A. Labeled crescent-shaped subnuclei viewed in horizontal section; anterior up; lateral left. Injections of digit 1 and digit 3 cortical suhgyri led to labeling of corresponding subnuclei within VH. The two digital lamellae appear to fuse anteriorly. B. Labeling of digit 2 subnucleus following injection of digit 2 subgyrus.
Sagittal section; dorsal up; anterior right. Because of the curvature of the lamellae, the digit 2 subnucleus appears as a crescent twice, once anteriorly and once posteriorly, with the digit 1 subnucleus and VPM appearing between them.
Fig. 6. High-power darkfield photomicrographs of 50-pm, TMB-reacted sections. Calibration bar = 50 p m . A. Labeled neuron in digit 3 subnucleus. Note dense filling of this multipolar neuron. B. Labeling of both cell bodies and neuropil following HRP injection into digit 4 subgyrus.
mella, the figure dropped off to 75%. In contrast to recent reports of labeling of small cells in cat VB (Penny et al., '82; Spreafico et al., '831, none of the smaller (10-12-pm diameter) cells in raccoon VB revealed any HRP labeling. However, there was dense labeling of neuropil.
Following the smaller injection (digit 21, only approxi- mately 30% of the large cells of the digit 2 lamella were labeled, no small cells were labeled, and there was minimal labeling of neuropil. It is suggested that the larger injec- tions allowed greater opportunities both for retrograde la-

THALAMOCORTICAL PROJECTIONS IN RACCOON 603
Fig. 7. Brightfield, low-power photomicrographs of four sagittal sections (A-D) in lateral to medial sequence, showing labeling of digit 5 subnucleus after multiple small injections into digit 5 subgyrus (lateral), and labeling of digit 2 subnucleus after a single small focal injection into digit 2 subgyrus (medial); 50-pm sections, reacted with TMB and counterstained with Neutral red. Calibration bar = 1 mm.
beling of the thalamic cell processes and for anterograde labeling of terminals of corticothalamic afferents, an oppor- tunity not present following the smaller volume injections.
Labeling following sulcal injections In three hemispheres, small (0.1-0.2 pl) injections were
made into the fundus of one of the branches of the triradiate sulcus (which separates the various digital subgyri). When compared with the labeling resulting from equivalent injec- tions into gyral crowns, the thalamic cell labeling was relatively sparse and diffuse. In each case, cells were la- beled in adjacent subnuclei corresponding to the subgyri on either side of the injections site (e.g., those for digits 3 and 4).
Labeling of cells of the posterior nuclear group (Po) In addition to VB labeling, injections of gyral crowns also
consistently led to sparse labeling of both perikarya and neuropil within the posterior nuclear group (Po). In every case, five to 15 neurons were labeled within a small region extending approximately 1 mm rostrocaudally, and 0.5-0.7 mm mediolaterally. Po labeling is depicted in both low-
power, darkfield, and high-power, brightfield photomicro- graphs in Figure 8.
Fiber labeling Labeled thalamocortical and corticothalamic fibers were
consistently observed, in addition to the thalamic cell bod- ies and neuropil labeling described in the preceding sec- tions. These fibers were seen to traverse VB in bundles which also passed through the external medullary lamina and thalamic reticular nucleus, in route to and from the internal capsule.
DISCUSSION The results of the present investigation are in agreement
with expectations based on prior electrophysiological stud- ies of raccoon VB and SmI, as well as with prior anatomical studies of VB-SmI connectivity. There exists in the raccoon a system of discrete, well-organized projections from VB hand subnuclei, or lamellae, to SmI hand subgyri, with a preferential projection to gyral crowns-the regions of rep- resentation of the glabrous surfaces of the hand. Retrograde transport of HRP reveals that cortical subgyri electrophys-

604 S. WAIUWN AND B.H. PUBOLS, JR.
Fig. 8. Photomicrographs of labeling in Po; 50-pm, TMB-reacted sections. A. Darklield photomicro- p a p h showing the tight clustering of labeled Po cells. Calibration bar = 10 pm. B. Higher-power hrightfield photomicrograph. Po cells, although smaller than labeled cells of VB, tend to have a hroader range of' sizes; they appear not to have prominently labeled processes. Calibration bar = 10 urn.
iologically identified as being associated with a specific hand digit receive axonal projections from thalamocortical relay cells located within the ventrobasal subnucleus asso- ciated with that same digit.
These results are in complete agreement with the earlier comprehensive retrog-ade degeneration data of Welker and Johnson ('651, who demonstrated the appropriate correla- tions between raccoon ventrobasal and cortical tail, hind- limb, palm, and head regions, as well as corresponding regions for each ofthe hand digits.
Welker and Seidenstein ('59), on the basis of observation of hematoxylin-stained raccoon somatosensory cortex, noted a dense penetration of myelinated fibers into gyral crowns but that few fibers seemed to enter cortex in sulcal fundi. A complementary arrangement was revealed in the present study by the finding that HRP injections into sulcal fundi (branches of the triradiate sulcus) led to more widespread but sparser labeling than did comparable injections of gyral crowns.
The present results extend to the raccoon recent reports, based on studies of retrograde transport of HRP, of a projec- tion of neurons of the posterior nuclear group (Po) upon SmI in cats (Spreafico et al., '81, '83). Results of earlier studies, utilizing a variety of' anatomical techniques, including the Nauta degeneration method and anterograde transport of
tritiated amino acids, had led to the conclusion that Po does not project to SmI, in either cats or monkeys (e.g., Heath and Jones, '71; Burton and Jones, '76). Apparently the first report of a small projection of Po to SmI was based on an electrophysiological study utilizing the technique of anti- dromic activation of thalamocortical relay cells in cats (Curry, '72). Out of 88 Po neurons activated orthodromically by electrical stimulation of the dorsal columns, 15 could be antidromically activated from the second somatosensory area (SmII), five from SmI, and one from both SmI and SmII. This small number of Po-to-SmI relay cells (7% of total sample) led Curry to suggest that "the projection to SI is too small to be demonstrated anatomically" (p. 475). A recent HRP investigation of relationships between cortical SmI and SmII, and various thalamic nuclei in the raccoon (Herron, '83), failed to reveal a labeling of Po neurons following SmI injections. Thus, the present results reveal for the first time an anatomically demonstrable, small but reliable projection of Po upon SmI in the raccoon.
There is one major aspect of the present results which is at variance with expectations based on previous observa- tions by others. On the basis of recent retrograde and anter- ograde transport studies of Jones and associates (Jones et al., '78, '82; Jones and Friedman, '82), it was anticipated that large injections (or multiple small injections) encom-

THALAMOCORTICAL PROJECTIONS IN RACCOON 605
passing all or most of a gyral crown would label all or most of the corresponding ventrobasal subnucleus, but that more focal injections would result in labeling of a “rod” of cells coursing through the entire anteroposterior extent of VB, or at least the entire anterposterior extent of the appropri- ate digital subnucleus (see Fig. 2; as well as Welker and Johnson, ’65: Fig. 14). The first of the predictions was veri- fied (e.g., Fig. 4). However, small, focal injections sometimes led to labeling of an entire VB lamella (Fig. 5A), and some- times to the labeling of only a small cluster of cells, rather than an entire “rod” (Fig. 7C,D). One possible explanation for this variability in extent of VB cell labeling may relate to local variations in the density of thalamocortical axon terminal arborization fields responsible for the uptake of the injected HRP (Jones, ’75). However, Saporta and Kruger (‘77), in an HRP study of rat ventrobasal thalamocortical relay neurons, also noted that small injections (0.005-0.5 pl) into rat somatosensory cortex often led to labeling of entire lamellae within VB, extending over a considerable anteroposterior range. Donoghue and Ebner (‘81) reported that small, focal injections of HRP into opossum somatosen- sory cortex also led to widespread anteroposterior labeling of VB. The primary somatosensory cortex of raccoon (Welker and Seidenstein, ’59), rat (Welker, ’711, and opossum (Pubols et al., ’76) consists, in each case, of a single body represen- tation, in contrast to the multiple representation found in simian primates (Kaas, ’83). It might be hypothesized that the organization of thalamocortical projection systems dif- fers fundamentally in animals having one versus multiple cortical representations receiving input from a single tha- lamic representation, and that the differences noted above are one manifestation of such organizational differences.
The present results, as well as those of the laboratories of Mountcastle (Mountcastle, ’57; Rose and Mountcastle, ’59; Poggio and Mountcastle, ’63), Welker (Welker and Seiden- stein, ’59; Welker and Johnson, ’651, Jones (e.g., Jones et al., 82), and others lead to a general picture of the organi- zation of mammalian somatosensory thalamocortical rela- tionships, including the following salient features: (1) Both the thalamic ventrobasal complex and primary somatosen- sory cortex are somatotopically organized; (2) VB projects to SmI in a topologically organized fashion; (3) concentri- cally arranged lamellae exist within VB: corresponding to representations of major body parts; (4) whatever the over- all somatotopic arrangement, cortical functional subdivi- sions representing major body subdivisions are composed of columns of cells receiving input from circumscribed body regions.
However, certain species differences must also be noted: (5) Simian primates possess multiple body representations in primary somatosensory cortex, while other species (both prosimian and nonprimate) appear to possess but a single body representation; (6) in simian primates, ventrobasal lamellae are composed of anteroposteriorly running “rods” of cells corresponding to subdivisions of major body parts, while such rods have not been demonstrated in animals with only a single cortical representation; (7) in simian primates, ventrobasal rods project to cortical cell columns, while in other mammals, an entire ventrobasal lamella may project to a circumscribed region of cortex. Thus, there appear to be fundamental differences in the thalamocorti- cal organization of mammals possessing one versus multi- ple cortical representations of the body surface within primary somatosensory cortex.
ACKNOWLEDGMENTS We acknowledge the technical assistance of B. J. Pettit
and thank Dianna Grimaldi for her care in typing the manuscript. This study was supported in part by research grants NS-13418 and NS-19486, United States Public Health Service. The paper is based in part on a dissertation submitted by S.W. in partial fulfillment of the require- ments for the Ph. D. degree at Pennsylvania State University.
LITERATURE CITED Burton, H., and E.G. Jones (1976) The posterior thalamic region and its
cortical projection in New World and OId World monkeys. J. Comp. Neurol. 168249-301
Curry, M.J. (1972) The effects of stimulating the somatic sensory cortex on single neurones in the posterior group (PO) of the cat. Brain Res. 44:463- 481.
Donoghue, J.P., and F.F. Ebner (1981) The organization of thalamic projec- tions to the parietal cortex of the Virginia opossum. J. Comp. Neurol. 198t365-388.
Heath, C.J., and E.G. Jones (1971) An experimental study of ascending connections from the posterior group of thalamic nuclei in the cat. J. Comp. Neurol. 141r397-425.
Herron, P. (1983) The connections of cortical somatosensory areas I and I1 with separate nuclei in the ventroposterior thalamus in the raccoon. Neuroscience 8243-257.
Jones, E.G. (1975) Possible determinants of the degree of retrograde neu- ronal labeling with horseradish peroxidase. Brain Res. 85.249-253.
Jones, E.G., and D.P. Friedman (1982) Projection pattern of functional components of thalamic ventrobasal complex on monkey somatosensory cortex. J. Neurophysiol. 48521-544.
Jones, E.G., D.P. Friedman, and S.H.C. Hendry (1982) Thalamic basis of place- and modality-specific columns in monkey somatosensory cortex: A correlative anatomical and physiological study. J. Neurophysiol. 48:545-568.
Jones, E.G., S.P. Wise, and J.D. Coulter (1979) Differential thalamic rela- tionships of sensory-motor and parietal cortical fields in monkeys. J . Comp. Neurol. 183:833-881.
Kaas, J.H. (1983) What, if anything, is SI? Organization of first somatosen- sory area of cortex. Physiol. Rev. 63:206-231.
Mesulam, M.-M. (1978) Tetramethyl henzidine for horseradish peroxidase neurohistochemistry: A noncarcinogenic blue reaction product with superior sensitivity for visualizing neural afferents and efferents. J. Histochem. Cytochem. 26: 106-117.
Mesulam, M.-M., and D.L. Rosene (1977) Differential sensitivity between blue and brown reaction procedures for HRP neurohistochemistry. Neu- rosci. Lett. 5:7-14.
Mountcastle, V.B. (1957) Modality and topographic properties of single neu- rons of cat’s somatic sensory cortex. J. Neurophysiol. 2Ot408-434.
Penny, G.R., K. Itoh, and I.T. Diamond (1982) Cells of different sizes in the ventral nuclei project to different layers of the somatic cortex in the cat. Brain Res. 24255-65.
Poggio, G.F., and V.B. Mountcastle (1960) A study of the functional contri- butions of the lemniscal and spinothalamic systems to somatic sensibil- ity: Central nervous mechanisms in pain. Bull. Johns Hopk. Hosp. 106:266-316.
Poggio, G.F., and V.B. Mountcastle (1963) The functional properties of ven- trobasal thalamic neurons studied in unanesthetized monkeys. J. Neu- rophysiol. 26775-806.
Pubols, B.H., L.M. Pubols, D.J. DiPette, and J.C. Sheely (1976) Opossum somatic sensory cortex: A microelectrode mapping study. J. Comp. Neu- rol. 165.229-245.
Rose, J.E., and V.B. Mountcastle (1959) Touch and kinesthesis. In J. Field, H.W. Magoun, and V.E. Hall (eds): Handbook of Physiology. Section I, Neurophysiology, Vol. I. Washington, DC: American Physiological Soci- ety, pp. 387-429.
Rosene, D.L., and M.-M. Mesulam (1978) Fixation variables in horseradish peroxidase neurohistochemistry: I. The effects of fixation time and per- fusion procedures upon enzyme activity. J. Histochem. Cytochem. 2628- 39.
Sakai, S.T. (1982) The thalamic connectivity of the primary motor cortex (MI) in the raccoon. J. Comp. Neurol. 204.238-252.

606 S. WARREN AND B.H. PUBOLS, JR.
Saporta, S., and L. Kruger (1977) The organization of thalamocortical relay neurons in the rat ventrobasal complex studied by the retrograde trans- port of horseradish peroxidase. J. Camp. Neural. 174r187-208.
Spreafico, R., N.L. Hayes, and A. Rustioni (1981) Thalamic projections to the primary and secondary somatosensory cortices in cat: Single and double retrograde tracer studies. J. Camp. Neural. 203r67-90.
Spreafico, R., D.E. Schmechel. L.C. Ellis, and A. Rustioni (1983) Cortical relay neurons and interneurons in the n. ventralis posterolateralis of cats: A horseradish peroxidase, electron-microscopic, Golgi and immu- nocytochemical study. Neuroscience 9:491-509.
Welker, C. (1971) Microelectrode delineation of fine grain somatotopic orga- nization of SmI cerebral neocortex in albino rat. Brain Res. 26t259-275.
Welker, W.I. (1967) A method for preparing brain casts. Anat. Rec. 158r239- 244.
Welker, W.I., and J.I. Johnson (1965) Correlation between nuclear morphol- ogy and somatotopic organization in ventrobasal complex of the rac- coon's thalamus. J. Anat. (Lond.) 99:761-790.
Welker, W.I., and S. Seidenstein (1959) Somatic sensory representation in the cerebral cortex of the raccoon (Procyon lotor). J. Camp. Neurol. 111r469-501.