Embed Size (px)
Citation preview

SOME, PHYSIOLOGICAL CHARACTERISTICS OF ASCOSPOREACTIVATION IN NEUROSPORA CRASSAI
MARY R. EMERSONKERCKHOFF LABORATORIES OF BIOLoGY, CALIFORNIA INSTITUTE OF TECHNOLOGY,
PASADENA, CALIFORNIA
The application of heat is used to break thedormant state in spores of several species of bacteriaand fungi and ensure germination at a known timeand in predictable numbers. Using the ascospores ofNeurospora tetrasperma, Goddard (3) and Goddardand Smith (5) investigated this type of activation,paying particular attention to the critical temperatureof the activation reaction and to the respiratory in-terrelations existing among dormant, activated, andgerminating spores. A very large increase in therespiratory rate-from ten- to forty-fold-takesplace when dormant spores are subjected to an acti-vating temperature, and this rate doubles whengermination occurs, approximately three hours afteractivation. The respiration of each stage differsqualitatively as well as quantitatively from the otherstages as shown by the dissimilarities in inhibition inresponse to several respiratory poisons. Further,carboxylase activity could not be detected in dor-mant spores, but was found after heat treatment,suggesting that the activation of this enzyme maybe the primary effect of heat which leads to the in-terruption of the dormant state. Acetaldehyde andethanol, alone of many substrates tested, were ableto induce a large increase in the Qo2 of dormantspores, but this increase was far short of thephenomenally large one accompanying activation;and in fact these substrates did not activate thespore, or at least, did not result in germination.Goddard (3, 4) also demonstrated that activatedspores could be returned to the dormant condition bythe inhibition of respiration after activation; thatthey could then be reactivated de novo, and thatthis sequence could be repeated at least several times.
Later Emerson (1) found that furfural is aseffective as heat in inducing activation of ascosporesof N. crassa, and Mefford and Campbell (7) foundthat it is also successful in bringing about the germ-ination of spores of a number of thermophilic andheat tolerant bacteria. The way the activation iseffected is quite unknown, although Mefford andCampbell suggest that in bacterial spores it may bedue to a detoxification of the medium. This wouldnot explain activation in Neurospora where nomedium other than distilled water need be involved.Recently Sussman (8) has tested a large number ofanalogues of furfural, and also many substances ofknown biological activity, for ability to activate dor-mant spores or to inhibit activation and growth.The results suggest that only unsaturated, 5-mem-bered heterocyclic compounds are active (diethylether showed slight activity), and that manyanalogues of active substances (particularly those
1 Received October 23, 1953.
with substituted nitro-groups, and carboxylic acidderivatives) are strongly inhibitory to germinationand to mycelial growth.
The work to be reported here was planned tolearn more about the nature of chemical activators,and of the relationship of chemical to thermal acti-vation. For this general purpose two lines of investi-gation, briefly referred to in a previous publication(1), are more thoroughly explored: (a) the activityof substances related to furfural; and (b) a com-parison of chemically and thermally induced acti-vation with respect to spore age, to reactivation, andto the effects of some respiratory poisons and otherinhibitors.
The term activation is not used synonymouslywith germination; the latter is regarded as the entiresequence of reactions which lead to the appearanceof the germ tubes, i.e., to mycelial growth. Activa-tion is here used to refer only to the initial stages ofthis sequence, which break dormancy.
MATERIALS AND METHODSTwo pairs of wild type strains: IA and 25a and
E5256A and E5297a, primarily the latter, were usedin these experiments, unless otherwise noted. Asco-spores were collected from crosses made in Petridishes on a special medium (10) inducing a heavyproduction of perithecia. When ripe a sample ofspores all of one age was collected by placing a freshcover on the Petri dish when spore shedding was wellstarted. By inverting the Petri plate and placing itin the sun, a particularly large sample could becollected in a few hours. The cover was then re-moved to a fresh base plate containing water andstored in the dark at a temperature of 23 to 250 C.Spore samples collected in this fashion containedrelatively few conidia and produced very littlemycelial growth. For use in tests, small batches ofspores were removed with a microspatula fromseveral regions on the cover, and mixed. The nuumberof spores per experiment varied from under 1000where only solid media were used to between 500,000and a million with liquid media. In the earlier ex-periments spores were washed with a 35 % solutionof Purex (5.25 % sodium hypochlorite) to kill anyconidia present, but it was found that a more carefulcollection of spores, and particularly the use of non-nutrient plating medium (agar and water) made thisstep unnecessary. In tests relating to spore age, thetime since shedding was noted to the half day or less;in all other experiments the spores used were withinthe age range known to be reactive, i.e., from 2 daysto 6 weeks. Any deviation from this rule is noted.In all cases spores were subject to three routinecontrols: plating on blank medium with no activation,
418
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.
EMERSON-ASCOSPORE ACTIVATION IN NEUROSPORA
activation by heat for 30 minutes, and activation by astandard furfural solution, 1.2 x 10A4M (0.01 ml/i).The work reported here was done over a period of 5years; the use of many different collections of sporeswas consequently unavoidable, but the controlsdemonstrated a very high degree of stability in thespores with regard to the reactions which are thesubject of the experiments under consideration.
Two alternative procedures were used exclusivelyin all tests. In the first, spores were plated on a non-autoclaved agar medium containing the test substanceor substances, in Petri dishes of a convenient size.Germination counts were made after an exposure of16 to 20 hours (at 23 to 250 C), which allowed timefor slowly penetrating substances to take effect. Forheat activation these dishes were placed partiallysubmerged on a wire shelf in a constant temperaturebath (600 C) for 30 minutes. The plating media werefreshly made for each test, with glass distilled waterand 1.5 to 3 % agar. For the series testing activecompounds and responses to age, 2 % sucrose andFries No. 3 salt solution were also added to themedia, but these were left out in later experiments.Activation, germination, and some mycelial growthof a normal form, take place in the absence of sugarand salts while growth from contaminating conidiais so reduced that it does not interfere with germi-nation counts, making the use of this type of mediummore convenient for experiments lasting over 24hours. In some cases the pH was noted because thedegree of dissociation of the test substance was ofinterest. In itself, the pH of the medium over therange 3 to 7 was without apparent effect on germi-nation; beyond this range the effect has not beendetermined.
In the second type of procedure the spores weresuspended in a solution of the test substance, andwere usually washed afterwards, but the circum-stances varied and were noted in the data each time.After the activation treatment, the spores were platedon agar and germination was noted as describedabove. The number of spores counted also variedand consequently was noted in each case.
The criterion used to denote activation was theappearance of the germinal buds or tubes, since thisis the only easily available indication that activationhas taken place. A negative result, particularly inthe use of poisons, needs careful interpretation sinceit may be due to an inhibition taking effect afteractivation but before germination is evident.
All substances were tested for contamination withfurfural by the aniline acetate test (11). This test isin general very satisfactory since it will show a pinkcolor for furfural, 0.2-0.1 ppm, though with a fewsubstances another color interferes with the detectionof physiologically active amounts of furfural. Moreserious is the lack of any equally sensitive test forother very active substances; however, the more
active of these, 5-methyl furfural and 5-hydroxyl-methyl furfural, are unlikely to be contaminants inthe substances tested for activity. In the series of
substituted benzaldehydes it is quite possible that theslight activity exhibited by some was due to thepresence of unconverted benzaldehyde.
RESULTSCHEMICAL ACTIVATING AGENTS: As a preliminary
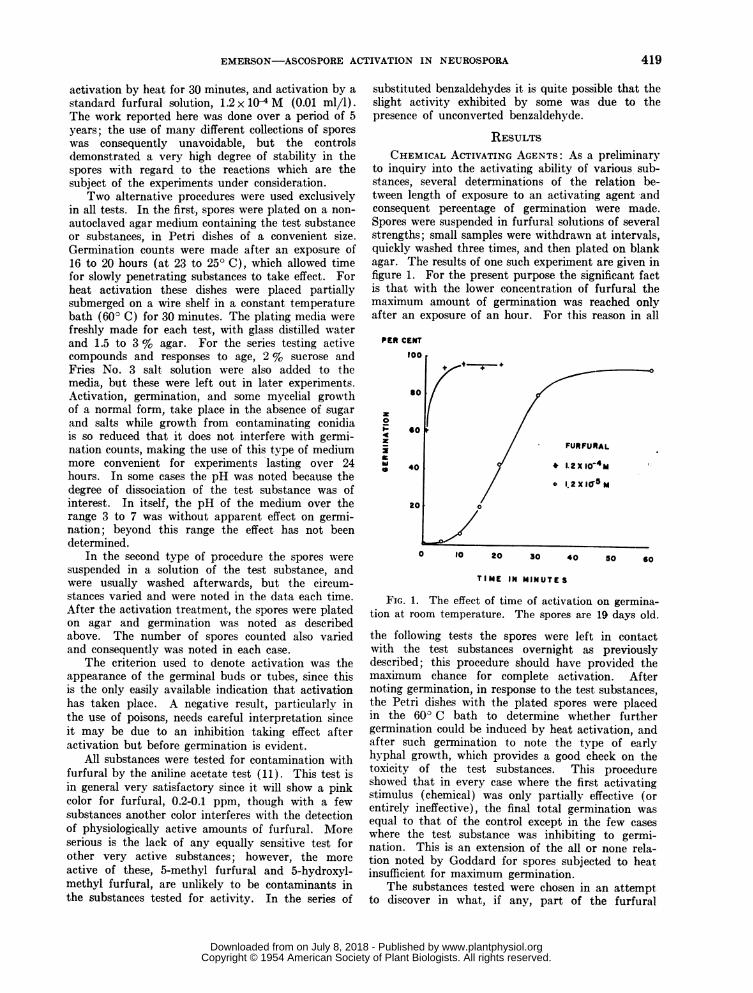
to inquiry into the activating ability of various sub-stances, several determinations of the relation be-tween length of exposure to an activating agent andconsequent percentage of germination were made.Spores were suspended in furfural solutions of severalstrengths; small samples were withdrawn at intervals,quickly washed three times, and then plated on blankagar. The results of one such experiment are given infigure 1. For the present purpose the significant factis that with the lower concentration of furfural themaximum amount of germination was reached onlyafter an exposure of an hour. For this reason in all
PER CENT
100
so
z
0
I9Zhi
FURFURAL
+ I.2X 10-4M
o 1.2XICF5 m
0 10 20 30 40 50
TIME IN MINUTES
FIG. 1. The effect of time of activation on germina-tion at room temperature. The spores are 19 days old.
the following tests the spores were left in contactwith the test substances overnight as previouslydescribed; this procedure should have provided themaximum chance for complete activation. Afternoting germination, in response to the test substances,the Petri dishes with the plated spores were placedin the 600 C bath to determine whether furthergermination could be induced by heat activation, andafter such germination to note the type of earlyhyphal growth, which provides a good check on thetoxicity of the test substances. This procedureshowed that in every case where the first activatingstimulus (chemical) was only partially effective (orentirely ineffective), the final total germination was
equal to that of the control except in the few caseswhere the test substance was inhibiting to germi-nation. This is an extension of the all or none rela-tion noted by Goddard for spores subjected to heatinsufficient for maximum germination.
The substances tested were chosen in an attemptto discover in what, if any, part of the furfural
419
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
TABLE IACTiVATION OF DORMANT SPORES BY VARIOUS COMPOUNDS
NAME: FuJRFuRAL 5-METHYr 5-HYLROXY- FURFURYL BENZ- SALICYL- PYRROLE-2- ACET-FURFURAL FURFURAL ALCOHOL ALDEHYDE ALDEHYDE ALDEHYDE ALDEHYDE
FORMULA: HRCHO * CH3RCHO CH20HRCHO HRCH2OH HR'CHO HOR'CHO R"CHO CHsCHO **
Conc. (M) % germination1.2x 10-2 98 87 .. 132.4 X 10-2 241.2 x 10-' 98 75 94 36 3 71.2 x 10-4 94 98 94 56 t 10 61.2x 10-a 99 96 201.2 x 10' 94 t 64 t 8 ..
* HH0H
R = IV10
R= II-H
** The same set of spores gave no activation with CHsCHO a few days later.t These responses are limited to a short period of maturity.
H--HRif= 11
H
NH
molecule physiological activity resides. Each com-pound was used on several occasions. The resultsshowed little variation except in the case of weakresponses which often could not be repeated with thesame spore sample later. All compounds found tohave any appreciable activity, as well as severalillustrating threshold activity, are listed in table I.The following were inactive: furan, furoic acid,furoin, furfuryldoxime, furoamide, methyl-furyl acro-lein, 5-nitrofurfural, o-amino-benzaldehyde, anisalde-hyde, m-hydroxybenzaldehyde, o-methoxybenzalde-hyde, 3-acetyl-6-methoxybenzaldehyde, cuminalde-hyde, cinnemaldehyde, peach lactone (C11H2002),glyoxal, glyceraldehyde, butyraldehyde and pyridoxal.At a concentration of 1.2 x 10-3 M furoic acid, severalsubstituted benzaldehydes, and pyridoxal each, on oneoccasion, induced from 7 to 15 % germination. At1.2x 1O-3M also, germination after heat activationwas suppressed by 5-nitrofurfural and o-methoxy-benzaldehyde (an emulsion at this concentration),and reduced by 3-acetyl-6-methoxybenzaldehyde.Mycelial growth was suppressed (i.e., germinal budsonly appeared), or reduced by methyl-furyl acrolein,salicylaldehyde, cinnemaldehyde and peach lactone.A concentration of 1.2 x 10-2 M was investigated onlyfor active substances-it was inhibitory to mycelialgrowth, but not to germination, in every case. Noneof the substances listed above was inhibitory tomycelial growth at a concentration of 1.2 x 104 M.
These results suggest that all compounds withappreciable activity possess an unsaturated ring; thefuran ring is not necessary nor is a heterocyclic ring,but the inclusion of a furan ring appears to give thegreatest activity; with one exception (furfurylalcohol) all active substances are aldehydes. Severalfacts suggest that the CHO group is important forthe activation reaction. In the series of substitutedbenzaldehydes tested, every substitution resulted inloss or very great reduction in activity; hydroxy and
methoxy groups in the ortho, but not in the para ormeta, position also increased the toxicity of the com-pound. With the noted exception of furfuryl alcohol,loss of the CHO group or change in its relation tothe ring as in methyl-furyl acrolein and cinnemalde-hyde, also brought loss of activity, with increasedtoxicity in the last two cases. However, furfuraldiacetate was found to be as active as furfural bySussman (8), with a good number of other 5-mem-bered heterocyclic compounds showing less but veryconsiderable activity even though they lacked theCHO group. While it might be possible to explainthe activity of the alcohol and acetate as the resultof substantial oxidation (by air) or hydrolysis, suchan easy conversion is not available for compoundslike 2-thiophene chloride or 2-methyl furan. Also itwas not possible at the end of the tests with furfurylalcohol to find any indication of oxidation to furfuralby means of aniline acetate, though this does not ruleout the possibility that it occurred.
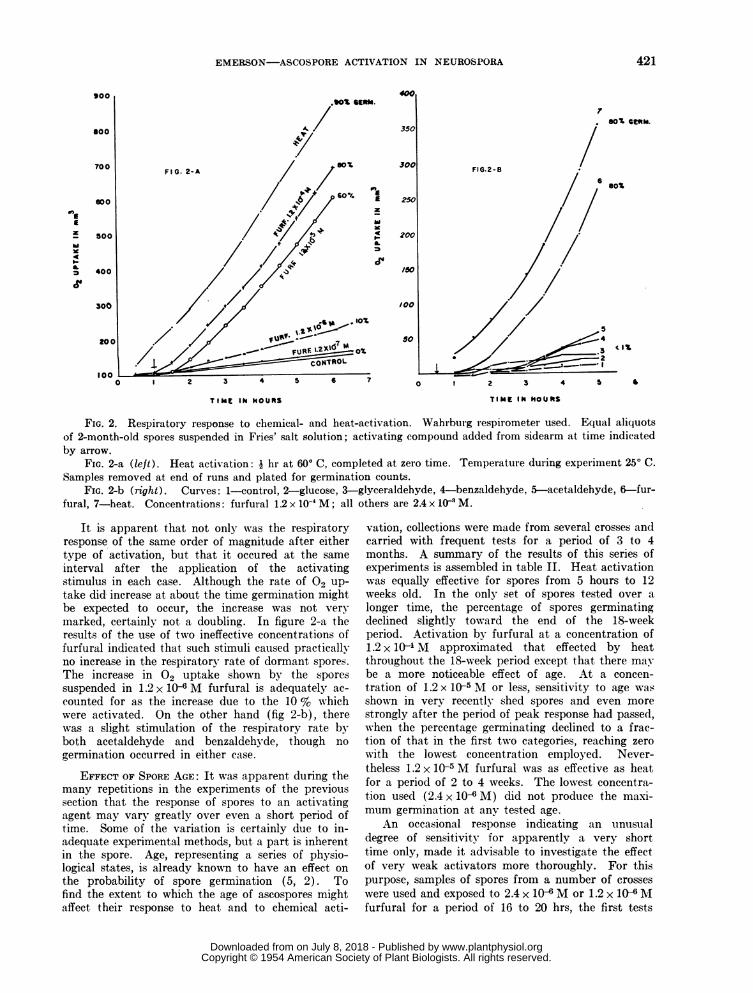
The increase in 02 uptake, after heat activation,by the spores of N. tetrasperma is very marked (4);there is every reason to believe that the samesequence of reactions takes place in N. crassa. Tomake sure of this, but primarily to ascertain whetherthe respiratory response is the same after chemicalas after heat activation, a number of trials weremade using the Warburg respirometer. Spores werefreed as much as possible from conidia by Goddard'smethod of repeated differential settling through waterin deep cylinders. Equal aliqudts (approximately2 mg dry wt) of a heavy suspension of spores wereused in each vessel; the error involved in this methodof apportioning spores is rather large-it is probablyabout 25 % judging by dried samples. Nevertheless,the results are sufficiently orderly to indicate thatthe individual results in any one experiment may beroughly compared. The data from two such experi-ments are represented graphically in figure 2.
420
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.
EMERSON-ASCOSPORE ACTIVATION IN NEUROSPORA
.o0 sea".
/
A.
FIG. 2-A
E 250z
< 2003-
/00
5
TIME IN HOURS TIME IN HOURS
FIG. 2. Respiratory response to chemical- and heat-activation. Wahrburg respirometer used. Equal aliquotsof 2-month-old spores suspended in Fries' salt solution; activating compound added from sidearm at time indicatedby arrow.
FIG. 2-a (left). Heat activation: i hr at 600 C, completed at zero time. Temperature during experiment 250 C.Samples removed at end of runs and plated for germination counts.
FIa. 2-b (right). Curves: 1-control, 2-glucose, 3-glyceraldehyde, 4-benzaldehyde, 5-acetaldehyde, 6-fur-fural, 7-heat. Concentrations: furfural 1.2 x 10' M; all others are 2.4 x 10' M.
It is apparent that not only was the respiratoryresponse of the same order of magnitude after eithertype of activation, but that it occured at the same
interval after the application of the activatingstimulus in each case. Although the rate of 02 Up-take did increase at about the time germination mightbe expected to occur, the increase was not very
marked, certainly not a doubling. In figure 2-a theresults of the use of two ineffective concentrations offurfural indicated that such stimuli caused practicallyno increase in the respiratory rate of dormant spores.The increase in 02 uptake shown by the sporessuspended in 1.2 x 10-6 M furfural is adequately ac-
counted for as the increase due to the 10 % whichwere activated. On the other hand (fig 2-b), therewas a slight stimulation of the respiratory rate byboth acetaldehyde and benzaldehyde, though no
germination occurred in either case.
EFFECT OF SPORE AGE: It was apparent during themany repetitions in the experiments of the previoussection that the response of spores to an activatingagent may vary greatly over even a short period oftime. Some of the variation is certainly due to in-adequate experimental methods, but a part is inherentin the spore. Age, representing a series of physio-logical states, is already known to have an effect on
the probability of spore germination (5, 2). Tofind the extent to which the age of ascospores mightaffect their response to heat and to chemical acti-
vation, collections were made from several crosses andcarried with frequent tests for a period of 3 to 4months. A summary of the results of this series ofexperiments is assembled in table II. Heat activationwas equally effective for spores from 5 hours to 12weeks old. In the only set of spores tested over a
longer time, the percentage of spores genninatingdeclined slightly toward the end of the 18-weekperiod. Activation by furfural at a concentration of1.2 x 10AM approximated that effected by heatthroughout the 18-week period except that there ma-be a more noticeable effect of age. At a concen-
tration of 1.2 x 10-5 M or less, sensitivity to age was
shown in very recently shed spores and even morestrongly after the period of peak response had passed,when the percentage germinating declined to a frac-tion of that in the first two categories, reaching zero
wvith the lowest concentration employed. Never-theless 1.2 x 10- M furfural was as effective as heatfor a period of 2 to 4 weeks. The lowest concentra-tion used (2.4 x 10 M) did not produce the maxi-
mum germination at any tested age.
An occasional response indicating an uinusualdegree of sensitivity for apparently a very shorttime only, made it advisable to investigate the effectof very weak activators more thoroughly. For thispurpose, samples of spores from a number of crosses
were used and exposed to 2.4 x 10" M or 1.2 x 10-6 Mfurfural for a period of 16 to 20 hrs, the first tests
S00
600
700
421
it49I-a.
500
400
so% GeR.
FIG.2-B6 So%
300
1002
,5
< lS
2 3 4 5 '
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.

422 PLANT PHYSIOLOGY
TVABLE IIPERCENTAGE oF, SPoREs * GERmINATING AT DiFFERENT AGES
5297a x 5256A (9/23/47) 5297a x 5256A (10/19/47) 25a x lA (11/4/47)
FURFURAL FuRs'uRAL FuRFURALAGE
CoN- HEAT 0 CoN- HEAT 0 0 0 CoN- HEATTROL x x x TROL x x x TROL x x x
5 hrs 0 95 ..22 .iday 0 95 . 56 0 98 . 77 . 80 84 82744 days ... . 0 99 ..91 41 .. .iweek .. 8 97 79 9 94 98 91 66 0 88 9 06lwk 6d 0 100 97 97 . .2 wks ... 6 97 85 132 wks 2d 0 95 ..96 44 ... 77 9370'3 wks 1 95 93 874--5wks 5 82 99 73 0 999590::, 15-6wks 3 84 83 602 0 97 9080o1,6 0 92 91 58157 wks 0 94 90 50 .. 96 92 52 2 . .8 wks 0 85 65 44. ......9-10 wks 0 78 69 36 1 . 1 83 8 64.11-12 wks .. 79 67 25 0 98 8 50, 0 75 76 33.15-16 wks 0 84 71 ..... .. ......17-18 wks 0 75 57 10 .. .. ..
* Spores activated on agar medium with or without furfural. Activation treatment by heat--30 min. in 600 Cbath. Each percentage based on spore count of 100 to 500, usually 150 to 250.
taking place immediately after the collection of thenewly shed spores. The results (fig. 3) show thatthere is a short period during the first 2 weeks (inthese strains) after the spores are shed when verylow concentrations of furfural are as effective as anyconcentration or as heat in bringing about maximumgermnination. Before and after this period 1.2 x 1O"M furfural does not provide an effective activatingstimulus. The responses to two concentrations ofbenzaldehyde over a period of three weeks make itseem probable that during the most sensitive periodall activating stimuli'induce unusually strong re-actions. This may explain the occasional activation
PIER CENT FURPURAL iZRsO%a
~~~~60~~ ~ ~ ~ ~~%
00
£ 40,
0 /0 20 /0 20 10 /0
elicited by otherwise ineffective agents such asacetaldehyde.
To test the effect of temperature on this agingprocess, spores were separated after collection intotwo sets one of which was kept at 250 C, the otherat 350 C. The latter temperature presumably leadsto a considerably heightened respiratory or meta-bolic rate and so perhaps to premature aging and anearlier decline in the germination rate. The resultsof these tests (table III) show that the responses ofspores kept at 350 C are in fact similar to those ofolder spores kept at a lower temperature particularlyin the accelerated decline in reactivity to furfural.
ZAX.41W104
20
lo~r
/0 /0 20
*
A
A
a SENZALSIINYDE
LZX1I63N
10 20
SPORE AGE (IN DAYS)
FIG. 3. The germination response to chemical activators applied during the "ripe"~period. All spores except3/15 are from crosses between E5256A and E5297a. 3/15 spores from the cross: 1555a. x Z-35-1-8A (pab-less x pab-less). Each point is based on a count of 150 to 350 spores. Control germination at time represented by last pointson curves, measured by response to 1.2 x 10' M furfural: 1/8, 97 %7; 1/12, 90%O/; 1/26, 96 9o.
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.
EMERSON-ASCOSPORE ACTIVATION IN NEUROSPORA
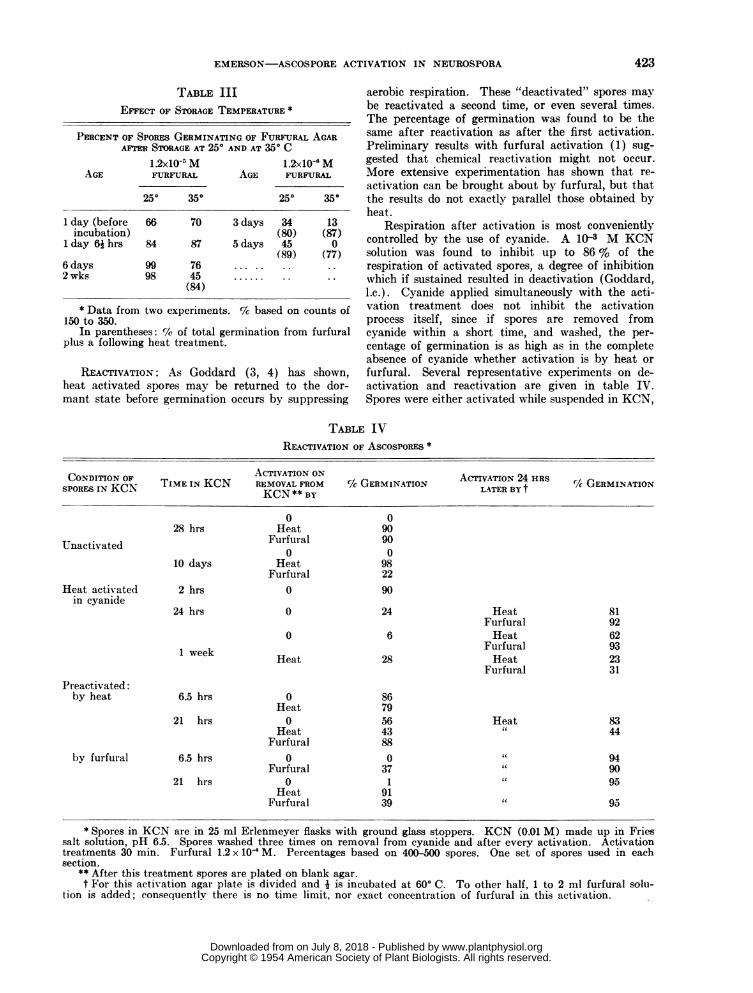
TABLE IIIEFFECT OF STORAGE TEMPERATURE *
PERCENT OF SPORES GERMINATING OF FURFURAL AGARAFTER STORAGE AT 250 AND AT 350 C
AGE1.2x10-5 MFURFURAL
250 350
1.2xl0-" MAGE FURFURAL
250 350
1 day (before 66 70 3 days 34 13incubation) (80) (87)
1 day 6ihrs 84 87 5days 45 0(89) (77)
6 days 99 762 wks 98 45 ......
(84)
* Data from two experiments. % based on counts of150 to 350.
In parentheses: %lk of total germination from furfuralplus a following heat treatment.
REACTIVATION: As Goddard (3, 4) has shown,heat activated spores may be returned to the dor-mant state before germination occurs by suppressing
aerobic respiration. These "deactivated" spores maybe reactivated a second time, or even several times.The percentage of germination was found to be thesame after reactivation as after the first activation.Preliminary results with furfural activation (1) sug-gested that chemical reactivation might not occur.More extensive experimentation has shown that re-activation can be brought about by furfural, but thatthe results do not exactly parallel those obtained byheat.
Respiration after activation is most convenientlycontrolled by the use of cyanide. A 10Q3 M KCNsolution was found to inhibit up to 86 % of therespiration of activated spores, a degree of inhibitionwhich if sustained resulted in deactivation (Goddard,l.c.). Cyanide applied simultaneously with the acti-vation treatment does not inhibit the activationprocess itself, since if spores are removed fromcyanide within a short time, and washed, the per-
centage of germination is as high as in the completeabsence of cyanide whether activation is by heat orfurfural. Several representative experiments on de-activation and reactivation are given in table IV.Spores were either activated while suspended in KCN,
3LE IVREACTIVATION OF AscosPoREs *
CONDITION OF TMINKN ACTIVATION ON ACTIVATION 24 HRS GEMNTOSPORES IN KCN TIME IN KCN REMOVAL FROM % GERMINATION LATER BY t C/( GERMINATIONKCN ~~~~KCN** BY
0 028 hrs Heat 90
Unactivated Furfural 900 0
10 days Heat 98Furfural 22
Heat activated 2 hrs 0 90in cyanide
24 hrs 0 24 Heat 81Furfural 92
0 6 Heat 62Furfural 93
Heat 28 Heat 23Furfural 31
Pr-eactivated:by heat 6.5 hrs 0 86
Heat 7921 hrs 0 56 Heat 83
Heat 43 44Furfural 88
by furfural 6.5 hrs 0 0 " 94Furfural 37 " 90
21 hrs 0 1 " 95Heat 91
Furfural 39 " 95
* Spores in KCN are in 25 ml Erlenmeyer flasks with ground glass stoppers. KCN (0.01 M) made up in Friessalt solution, pH 6.5. Spores washed three times on removal from cyanide and after every activation. Activationtreatments 30 min. Furfural 1.2 x 10i M. Percentages based on 400-500 spores. One set of spores used in eachsection.
** After this treatment spores are plated on blank agar.t For this activation agar plate is divided and i is incubated at 600 C. To other half, 1 to 2 ml furfural solu-
tion is added; consequently there is no time limit, nor exact concentration of furfural in this activation.
423
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
or were placed in the cyanide solution after acti-vation. This latter sequence avoids the irrelevantcomplication of the combined effects of high tempera-ture and cyanide, and is the preferred method; butthere appears to be no marked difference in the re-sults whichever course was followed. The results ofthese reactivation tests were quite reproducible,though varying in actual magnitude between experi-ments and with different sets of spores.
In considering the results there are several com-plications which should be kept in mind. A tem-perature of 600 C, for even 15 minutes will killactively growing cells; the concentrations of furfuralused in activation exert no detectable inhibition. Asecond point arises from the possibility of interactionbetween furfural, as an aldehyde, and KCN withformation of furfural cyanohydrin. The apparentlycomplete deactivation after even a short time incyanide, of spores previously activated by furfural,could be due as much or more to the withdrawal offurfural from the activation site by chemical com-bination with cyanide, as to the effect of the cyanidein repressing aerobic oxidation. In either case, thespores became dormant again, but the different re-sponse they showed to reactivation by heat and byfurfural might be explained in the latter case by amasking of the activation site by the cyanohydrin-which though not removed by washing, is destroyedor otherwise eliminated by the 600 C temperature ofheat activation. While such an explanation fits theone experimental sequence adequately, it does notapply equally well, if at all, to the others; the re-sults, however, are parallel in all cases. A final con-sideration concerns the degree and speed of pene-tration of the compounds in question, particularlythe different effects of the two temperatures.
The data show that reactivation by either heat orfurfural does occur successfully after original acti-vation by either agent, but with several partial ex-ceptions. Heat applied before any residual actioninitiated by the first activation has been completed,appears to be lethal; spores so treated do not re-spond to either type of activation the next day,although other classes with a small amount of germi-nation do respond with much increased germination.This lethal effect was absent, or slight, in the spores6.5 hrs in cyanide; nor is it shown in the reactivationexperiments reported by Goddard.
Spores originally activated by furfural, then de-activated, are only indifferently well reactivated bythis agent. This relation is particularly evident in theexperiment in which activation preceded immersionin KCN. Here, under entirely similar conditions(except for the original activation), reactivation byfurfural was good after heat, poor after furfuralactivation. This illustrates a difference between thetwo activators which must be considered in any ex-planation of activation. A further point shown ineach experiment is the stimulating effect of heat onany subsequent response to furfural. The time in thesequence at which heat is applied appears to be of
little importance-it may precede or follow cyanidepoisoning. The depressing effect of cyanide on sub-sequent activation by furfural (with no interventionof heat) is shown quite markedly by dormant as wellas by deactivated spores. Here again it does notseem likely that there could be a large enough carry-over of cyanide throughout washing, to inactivatethe furfural chemically.
The reactivation by heat of heat activated spores,while much better than the furfural-furfural se-quence, is usually considerably short of the maximumpossible even when immediate, and possibly lethal,heat treatment is avoided. This result again doesnot agree with the complete reactivation found byGoddard.
INHIBITORS: A number of analogues of furfuraland a number of known enzyme poisons were used inattempts to inhibit the activation reaction, particu-larly in the hope of finding evidence concerning therelation of the two types of activation. The usualdifficulties in interpreting the effects of poisons ap-plied in vivo are increased here by the necessity ofusing a high temperature during some part of anexperiment. A further complication is that the in-dicator of activation-the germination of the spore-may be inhibited during the period after activation,but preceding the appearance of germ tubes. Thedata presented are thought to contain some usefuland pertinent information as well as many obviousuncertainties.
The following analogues of furfural were usedwith furfural in proportions of 100 and 1000 to onewithout resulting in any lowering of the rate ofgermination: furan, furoamide, and acetaldehyde;and the following poisons were also without appreci-able affect: CUSO4 - 5 H2(10 3 M); AgNO3 (10 3M); iodoacetic acid (10 3M); iodoacetamide (2 x1O4 M); and KCN (10-3 M) though all but the firstinhibit the further growth of germ tubes to a greateror lesser extent if not removed after activation.More extensive tests were made with the furfuralanalogue, 5-nitrofurfural, with 2,4-dinitrophenol,NaHSO3, and NaN3. The results of these testswere quite reproducible when care was taken toequalize the washing, particularly after presoakingthe spores in the poison. Because, at the concentra-tions used, all of the inhibiting substance could bewashed out again, the method used (1 to 3 centrif-ugations, 15 ml H20) was designed to clean the ex-teriors as quickly as possible with a minimum ofoutward diffusion of the poison.
Several experiments illustrating the affect of 5-nitro-furfural, 2,4-dinitrophenol, and NaHSO3 onactivation, are presented in table V. In the presenceof these compounds heat activation resulted in someamount of irreversable inhibition, while furfuralactivation was unaffected except by the stronger con-centrations of NaHSO3. When spores were presoakedin 5-nitrofurfural or 2,4-dinitrophenol, and washedbefore activation, it is clear that a partial ex-planation of this difference is complete lack of pene-
424
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.

EMERSON-ASCOSPORE ACTIVATION IN NEUROSPORA
TABLE V
INHIBITION OF ACTIVATION BY 5-NITROFURFURAL, 2,4-DINITROPHENOL, AND NAHSO3
INHIBITOR: 5-NITROA 2,4-DINITROPHENOL t NAHSOSt
CONCENTRATION 12x10-8M 10-3M 10-4M 5x10' M 10-1M 10-2M 103MCONCNTRAION .PEDp S* P S P S P S P S P S P S
% germinationActivated by:
Furfural 30 min1.2 x 10-aM 97 96 .. 84 .. .. .. .. .. ..1.2x 10 {M(15 min NaHSO0) .. 71 + 98 631: .. 96 4 + 96 33 .. 96
Heat30 min 75tt 99 55 tt 98 9215mi .. .. .. . .. .. Ott 28 tt 0 52 0 93 87
Heat + furfural30 min (15 minNaHSO0) 69tt .. 5Stt 100 .. .. Ott 41tt .. .. ..
* P = inhibitor present duiring activation; S = spores presoaked in 5-nitrofurfural and 2,4-dinitrophenol 3 hrs, inNaHSOs i hr. Spores washed 3 times after activation, and 3 times after soaking in inhibitor, plated on agar.
** In aqueous solution, pH 4.5.t In Fries' salt solution, pH 4.5 for 2,4-dinitrophenol and 5.5 for NaHSO3.
ft No reactivation or ungerminated spores by either furfural or heat 24 hrs later.t Good, or moderately good, reactivation by either heat or furfural.
tration of either of these substances in the shorttime of the activation treatment, except at the highertemperature. The results also suggested that furfuralactivation is more sensitive than heat activationwhich is unaffected in each case by the amount ofinhibitor present in the spores after 3 hours presoak-ing, followed by washing. This inhibition can be re-versed by increasing the concentration of furfuralused to activate; it cannot be reversed when theconcentration of the inhibitor is appreciably greaterthan 1O- MI, no doubt partly because above thisconcentration furfural also entered the inhibitoryrange.
5-Nitrofurfural is much more toxic to conidial andmycelial growth than to the activation of dormantascospores; only a very slight reversal of such in-hibition by furfural was achieved in the hyphalgrowth from germinating ascospores. There was noreversal of inhibition of conidial growth induced by1.2 x 1jO M 5-nitro-furfural by 1.2 x 1F3 M furfural.In the presence of sugar these inhibitions are lessextreme.
The selection of NaHSO3 as a poison was due tothe hope that an aldehyde reagent might help clarifya possible function of the CHO group in activation.The results do not rule out this possibility, but canbe interpreted in too many ways to lend much sup-port to it. Loss of activity by furfural in thepresence of high concentrations of bisulfite is to beexpected; it could be due entirely to an externalreaction between the two substances, with the re-sulting combined form inactive, or failing to pene-trate, or both. Reactivation of spores in this se-quence was only fairly good, and better by heat thanby furfural (86 vs. 72 %; after M/2 NaHSO3). This
suggests that though the furfural-bisulphite compoundis not an activator, it does enter the spore to someextent, causing an inhibition which appears to affectfurfural activation particularly. The concentration ofbisulphide present in spores after a short presoakingdoes not affect furfural activation but exerts a strongand irreversible inhibition of heat activation. Thedegree of inhibition is not affected, or only slightlyso, by the presence of furfural during heat treatment.Unfortunately this possible protective effect was nottested against the lower concentrations of NaHSO3.
The superior penetration undoubtedly accompany-ing the 60° C temperature does not seem a sufficientreason to account for the irreversible nature of theinhibitions produced with heat if activation alone isin question. The percentage of spores affected inthe presence of heat may be no greater than thatshowing reversible furfural inhibition at 250 C. Itwould seem likely that increased penetration of poisonshould increase the percent inhibited, as well as thedegree of inhibition. If activation alone is affected,there should be some reversible inhibition in each casealso. Consequently it is considered possible that hightemperature may make other sites open to attack,which otherwise are innert in the dormant spore.
In table VI are data from two experiments illus-trating the effect of sodium azide on the two typesof activation. Since the inhibitory action of azidehas been shown to vary directly with the amount ofundissociated hydrozoic acid present (6), the solutionsused were pH 6.0 or less. No chemical reaction be-tween NaN3 and furfural could be demonstrated; inany case the molecular concentrations used wouldmake a chemical inactivation of azide by furfuralunlikely.
425
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
TABLE VIEFFcT OF SODIUM AZIDE ON ACTIVATION
% SPORESTIME IN NAN. ACTIVATED BY GERMINATING
EXPER. CONTROL
1 hr
2 hrs
Activation period
15 min30 min15 min30 min15 min
30 min
Series A *HeatFurfur alHeat + furfuralHeatFurfuiralHeat + furfural
Series B **HeatHeatFurfur alFurfur alHeat + furfural
Heat + fuirfural
486793.5
7371
8993
.. .
.. .
.. .
. .
0899060 (buds
only)0 +
* Series A: Spores suspended in 0.01 M solution ofNaN8 at pH 4.0, then washed once, activated, washedthree times, and plated on agar. % based on 500 spores.Furfural: 12 x 10' M. Activation time: 15 min.
** Series B: Spores activated in 0.01 M NaN8 in Fries'salt solution at pH 6.0, washed three times, and platedon agar. Furfural: 1.2 x 10' M. %o based on 400 to 700spores.
t Approximately 50 %o reactivated by furfural; noneby heat.
It No reactivation by heat; less than 1 9o by furfural.* No reactivation by heat; approximately 20% by
furfural.
Activation by furfural is comparatively unaffectedby NaN3, while heat activation is greatly or com-
pletely inhibited by this poison. Since the resultsare very much the same whether the spores are pre-treated with NaN3 and washed before the activatingtreatment, or activated in the presence of NaN3, itis evident that this difference is not due to a tem-perature effect on permeability. Spores activated byheat while simultaneously poisoned by azide, cannot,after washing and plating, be reactivated by anotherheat treatment 24 hours later, though they may beto some degree reactivated by furfural. When fur-fural is present during the first heat activation, thespores are protected to a large extent against theinhibiting effect of azide. It will be noted that inone experiment quoted, only germinal buds result,showing that though in at least 60 % of the sporesthe germination process had been completed, themycelial metabolism is inhibited. It may well bethat some of the reduction in the amount of germi-nation is due to a stronger inhibition at this otherphysiological site rather than to an interference withthe activation reaction. This type of inhibition islimited to spores heated while suspended in a solu-tion of azide-those showing reduced germinationafter presoaking in azide grow in an uninhibitedfashion if they are able to germinate. Altogether,
the data suggest that furfural is able to act to cir-cumvent poisoning by NaN3 with respect to theactivation reaction, but less successfully or not at allwith regard to any inhibition induced in the regularmetabolic cycles.
Since in the presence of either NaHSO3 or NaN3the spores may have been responding to the Na+ ion,the effect of 0.5 and 0.01 molar NaCl solutions onactivation by heat was tested. There was no changein the percent of germination at either concentration.
DISCUSSIONGoddard has suggested that the primary effect of
heat on the dormant spore may be the reversibleconversion of some inactive substance to an activestate, possibly to a precursor of, or directly to, acatalytically active substance; this theory continuesto fit the available data. The simplest explanationof chemical activation is that an activator suppliedexternally is identical with, or very like, this activesubstance released within the spore after heat treat-ment, or that it is identical with or very similar to acritical substance of which the heat-released substance1S a precursor.
There are, however, differences between the acti-vation resulting from heat and that due to externallyapplied chemical agents which must be explainedbefore any substantial identity in the two types ofactivation can be assumed; namely, the sensitivityshown by chemical activation to spore age as op-posed to the comparative independence of age shownby heat activation, differences in reactivation, andtoward some inhibitors. Though the age effect dis-cussed here refers mainly to older spores it must beremembered that the same discrimination betweenactivators is shown by young but ripe spores still inthe ascal sac, and by freshly shed spores (1). If oneexplanation applies to both age groups then it can-not be the accumulation of an inhibiting substanceduring the period of spore dormancy. It is possible,however, that the causes in the two cases are differentso that such an explanation is not ruled out for theolder spores. The reactions of young spores are notchanged by washing with Purex, which makes it im-probable that there is a substance present at the timeof shedding which prevents too early germination, ashappens in the case of many seeds.
A change in the permeability of the spore withage would be a satisfactory explanation, but theevidence available does not easily fit such an as-sumption. The spores used in the respiratory runswere two months old and showed an already con-siderably reduced amount of germination. This con-dition, however, did not prevent a reaction nearly asimmediate as that after heat activation when fur-fural was added from the side arm of the respiratoryvessel (fig 2-a).
Other types of substances have been found toenter the dormant spore in effective amounts:acetaldehyde and pyruvate have also been shown bytheir effect to enter easily, as has H202 (4, 5).
426
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.

EMERSON-ASCOSPORE ACTIVATION IN NEUROSPORA
Sussman (8) found that old spores, no longer veryreactive to furfural, gave maximum results frompoisoning with furfural analogues as gauged by areduction in the germination rate after heat activa-tion. Since the spores were suspended in the in-hibiting substances for 6 hours, the latter may haveentered slowly, but the spores were evidently notimpermeable to them. These facts make it appearvery unlikely that the large changes in reaction to a20 hour exposure to furfural shown by spores atdifferent ages can be due solely or primarily tochanges in permeability.
If permeability is not a matter of critical impor-tance, it is possible that variations with age reflectchanges within the spore more immediately relatedto the activation reaction. If permeability were themain factor, the lowest concentration of furfural,i.e., 1.2 x 10-6 M which will produce maximum germi-nation at the most sensitive time, could be consideredas the concentration necessary to produce this resultat any time, considering the activation reaction byitself. Instead it appears more probable that thespore undergoes a ripening process during which itapproaches, reaches, and declines from a state ofoptimum efficiency in its response to an activatingagent; and that it is only during a brief period ofripeness that the metabolism is triggered to reactwith maximum effect to a minimum stimulus. Whileit is probable that the internal production of activatoris always in excess of what is needed to activate thespore unless the heat treatment is very short, thisfact would account only partially for the observeddifferences between the two types of activation sinceincreasing the concentration of externally appliedactivator will counteract the effect of age to a certaindegree only.
It is possible that the difference between inter-nally produced and externally applied activators isless real than it appears; that it is due to a secondaryeffect of heat on the metabolism of the spore. Thiscritical secondary effect is assumed to be a change insome part of the spore metabolism which will resultin a state closely resembling the " ripe " state, thoughless extreme. It is also assumed that this state can bebrought about by heat over a much longer period oftime than that during which the natural reactivityof the spore remains at a high level. There is nodirect evidence supporting this theory, but it fits theavailable data. In the experiments with deactivationand reactivation, the probable effect of cyanide inlowering the general metabolic level may be the mainexplanation for the consequent comparative lack ofresponse to furfural of those spores to which no heatwas applied at any time. This effect is particularlynoticeable in the case of dormant spores (suspendedin cyanide for some time before they are activated)where it results in a large decrease in the amount ofgermination due to furfural while leaving that dueto heat unaffected. Further, after an original acti-vation by heat the sensitive state so induced pre-sumably lasts for some time even in the presence of
cyanide since spores so treated are still responsive tofurfural. From this it follows that it is not primarilythe heightened respiratory rate accompanying heatwhich is responsible for the resulting "ripe" state.This probability is strengthened by a comparison ofthe respiratory rates of dormant spores of differentages (4). That of the youngest and most reactiveage (three weeks) was only 1/2 of the Qo2 of sporesnine weeks old. The critical characteristic of theyounger spores may, however, be the forty-fold in-crease in the respiratory rate they show on activationas against the ten-fold increase in the nine weeks oldspores.
Some very recent work appears to bear directlyon this possible secondary effect of heat. Sussman(9) has found that quite old spores of Neurosperatetrasperma, no longer responsive to furfural, can beresensitized by preheating at a less than activatingtemnperature. This freshly induced sensitivity maylast a very short time-a few hours only in 28 monthold spores-though the respiratory increase resultingfrom the preheating continues at the new higher levelfor a considerably longer time. These results are asyet unexplained, and may possibly be unconnectedwith an effect of heat occurring in younger and morereactive spores, though the main result-a sharpenedsensitivity to chemical activators-is the same at eachage.
An effect of heat of a possibly similar kind wasnoted by Evans and Curran (2) who found thatpreheating the spores of mesophilic bacteria greatlyaccelerated their germination when they were subse-quently incubated on an appropriate medium, thoughit did not increase the number of spores germinating.This acceleration was onlv very slightly reduced byan interval of up to a week between the pretreatmentand incubation.
Whereas the main course of activation may wellbe the same following either type of activating stimu-lus, there seems little doubt that the initial steps aredifferent in the two cases. The results of inhibitionwith NaN3 show that this poison can prevent theinitiation of activation by heat, and that this in-hibition can be by-passed in the presence of furfural.If the role of heat is to convert an inactive substancethrough one or more steps to one having catalyticproperties, then apparently the azide intervenes atsome point in this conversion, probably directly,though not necessarily so. Again, it seems mostprobable that the role of furfural in lifting the in-hibition is to take the place of the heat-producedcatalyst rather than to reverse the inhibition moredirectly at the point at which it takes effect.
As a poison azide is known to attack a variety ofenzymatically controlled reactions, very little can besaid about the kind of reaction involved here fromthe data available. Since oxygen is unnecessary foractivation, no direct respiratory inhibition is in ques-tion, and in preliminary trials with a respirometer,the respiratory inhibition, which does in fact occur inthe presence of azide, was not relieved by the added
427
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
presence of furfural. The results from soaking sporesin azide, with washing before activation, suggest thatthe inhibition is either closely limited to the heat-conversion reaction, or, more probable, in view of therespiratory effects, that this particular inhibition isless easily reversed by washing than others whichsurely take place even in the metabolism of dormantspores.
While the simplest explanation of the facts postu-lates a high degree of similarity in form, and conse-quently in action, between internally produced andexternally supplied activator, plus an effect of heat,another explanation has not been ruled out. It isquite possible that the two types of activating stimuliattack the dormant metabolism at different sites, andthat the observed distinctions follow from this cir-cumstance.
SUMMARY1. Compounds able to activate dormant spores,
in very low concentrations, contain an unsaturatedring; the most active have a furan ring.
2. The age of spores is found to affect differentlytheir sensitivity to heat and to chemical activation;sensitivity to the latter declines much faster.
3. At some time within three weeks of sheddingthere is a short period of "ripeness," or great sensi-tivity to furfural and other chemical activators.During this period maximum germination is initiatecdby a solution of 0.1 ppm of furfural.
4. Spores returned to dormancy after activationcan be reactivated by furfural as well as by heat. Itis suggested that a secondary (non-activating) effectof heat is responsible for the differences in reactiva-tion by the two means.
5. 2,4-dinitrophenol and 5-nitrofurfural appear toinhibit activation by furfural to some degree; thiseffect can be partly counteracted by an increase inthe concentration of furfural employed.
6. The results of inhibition by sodium sulfite aredescribed and discussed with reference to the possi-bility that an aldehyde group takes part in the acti-vation reaction.
7. NaN3 is found to inhibit activation by heatbut not by furfural; when heat and furfural treat-ments are combined inhibition is much reduced. Itis suggested that furfural takes the place of an in-ternal catalyst produced by heat, whose production isirreversibly inhibited by NaN3.
8. Except for the initial step-i.e., the productionof a catalyst by heat-, the course of activation isthought to be the same following either type of acti-
vating stimulus. Apparent differences are inter-preted by postulating the induction of a metabolicstate similar to the naturally occurring "ripe" stateas an accompaniment of heat activation.
I wish to express particular thanks to ProfessorA. J. Haagen-Smit for samples of most of the com-pounds tested for activity; to Professor JamesBonner for a sample of 3-acetyl-6-methoxy benzalde-hyde, and to the Eaton Laboratories, Norwich, N. Y.for the 5-nitrofurfural used in these experiments.
I am also indebted to Dr. A. S. Sussman for ac-cess to his paper in press, and to several members ofthe staff of the Kerckhoff Biology Laboratories formuch help and advice.
LITERATURE CITED1. EMERSON, M. R. Chemical activation of ascospore
germination in Neurospora crassa. Jour. Bact. 55:327-330. 1948.
2. EVANS, F. R. and CURRAN, H. R. The acceleratingeffect of sublethal heat on spore germination inmesophilic aerobic bacteria. Jour. Bact. 46: 513-523. 1943.
3. GODDARD, D. R. The reversible heat activation in-ducing germination and increased respiration inthe ascospores of Neurospora tetrasperma. Jour.Gen. Physiol. 19: 45-60. 1935.
4. GODDARD, D. R. The reversible heat activation ofrespiration in Neurospora. Cold Spring HarborSymp. Quant. Biol. 7: 362-376. 1939.
5. GODDARD, D. R. and SMITH, P. E. Respiratory blockin the dormant ascospores of Neurospora tetra-sperma. Plant Physiol. 13: 241-264. 1938.
6. KEILIN, D. The action of sodium azide on cellularrespiration and on some catalytic oxidation reac-tions. Proc. Roy. Soc. (London) B 121: 165-173.1936.
7. MEF0ssW, JR., R. B. and CAMPBELL, JR., L. L. Theactivation of thermophilic spores by furfural.Jour. Bact. 62: 130-132. 1951.
8. SUSSMAN, A. S. The effect of heterocyclic and othercompounds upon the germination of ascospores ofNeurospora tetrasperma. Jour. Gen. Microbiol. 8:211-216. 1953.
9. SUSSMAN, A. S. The sensitization of ascospores tochemical activators by heat treatment. Mycologia.(In press.)
10. WESTERGAARD, M. and MITCHELL, H. K. Neurospora.V. A synthetic medium favoring sexual reproduc-tion. Amer. Jour. Bot. 34: 573-577. 1947.
11. YOUNGBURG, G. E. and PUCKER, G. W. Studies onpentose metabolism. I. A colorimetric method forthe estimation of furfural. Jour. Biol. Chem. 61:741-746. 1924.
428
www.plantphysiol.orgon July 8, 2018 - Published by Downloaded from Copyright © 1954 American Society of Plant Biologists. All rights reserved.





























