Embed Size (px)
Citation preview

Spatiotemporal Expression of SSeCKSin Injured Rat Sciatic Nerve
LI CHEN,1,2 JING QIN,2 CHUN CHENG,3 SHUQIONG NIU,1 YONGHUA LIU,3
SHUXIAN SHI,3 HAI’OU LIU,3 AND AIGUO SHEN1*1The Jiangsu Province Key Laboratory of Neuroregeneration, Nantong University,
Nantong, People’s Republic of China2Department of Pathology, Medical School of Nantong University (Former Nantong
Medical College), Nantong, People’s Republic of China3Department of Microbiology and Immunology, Medical School of Nantong University
(Former Nantong Medical College), Nantong, People’s Republic of China
ABSTRACTSSeCKS (src suppressed C kinase substrate) functions in the control
of cell signaling and cytoskeletal arrangement. It is expressed in brainand spinal cord, but little is known about its expression in peripheralnerves. In this study, in rats, real-time polymerase chain reaction andWestern blot analysis showed that expression of SSeCKS in crushed sci-atic nerve reached its highest level 6 hr after crushing, whereas in atransection model, SSeCKS peaked at 2 days in the proximal stump and12 hr in the distal stump. Immunohistochemical analysis demonstratedup-regulation of SSeCKS protein surrounding the crush site and in thetwo stumps of the transected nerve. In addition, SSeCKS colocalized withgrowth-associated protein 43 and with S100, which also changed withtime after injury. These findings support the idea that SSeCKS partici-pates in the adaptive response to peripheral nerve injury and may beassociated with regeneration. Anat Rec, 291:527–537, 2008. � 2008Wiley-Liss, Inc.
Key words: SSeCKS; sciatic nerve; crush; transect; rat
Src suppressed C kinase substrate (SSeCKS) is a highmolecular mass (290/280 kDa) heat-stable protein thatwas identified as a protein kinase C (PKC) substrate/PKC-binding protein. In vitro, SSeCKS mRNA and pro-tein levels are down-regulated in transformed cells, sug-gesting a role in growth regulation (Chapline et al.,1996). In vivo, SSeCKS is expressed by fibroblasts, endo-thelial cells, and mesangial cells and has mitogenic reg-ulatory activity, indicating a role in tumor suppression.During embryogenesis, SSeCKS is involved in control-ling cytoskeletal structure and tissue architecture, form-ing the migratory process and cell migration (Gelmanet al., 2000). The SSeCKS expression pattern is develop-mentally regulated in numerous cell types and tissuesduring perinatal growth, indicating a role in develop-ment and differentiation processes. In the nervous sys-tem of adult rats, SSeCKS is localized to central axonalcollaterals of primary sensory neurons in the cerebel-lum, medulla, and sensory ganglia (including trigeminalganglia) and in the dorsal horn at all spinal levels. Inaddition, SSeCKS is localized to C-fibers and is involvedin nociception (Siegel et al., 2002). No prior studies have
examined the involvement of SSeCKS in peripheralnerve injury; therefore, we examined the expression ofSSeCKS in this condition, a complex process thatremains poorly understood.
Drs. Chen and Qin contributed equally to this work.
*Correspondence to: Aiguo Shen, The Jiangsu Province KeyLaboratory of Neuroregeneration, Nantong University, Nan-tong, People’s Republic of China, 226001. Fax: 86-513-85051999.E-mail: [email protected]
Abbreviations used: BSA = bovine serum albumin; PBS =phosphate-buffered saline; SSeCKS = src suppressed C kinasesubstrate; TBST = Tris-buffered saline with Tween; b2-M = b2-microglobulin; GAP43 = growth-associated protein 43.
Grant sponsor: The National Natural Scientific Foundation ofChina; Grant numbers: 30300099, 30770488; Grant sponsor:Natural Scientific Foundation of Jiangsu Province; Grant num-bers: BK2003035, BK2006547.
Received 1 September 2007; Accepted 17 January 2008
DOI 10.1002/ar.20692Published online 31 March 2008 in Wiley InterScience (www.interscience.wiley.com).
� 2008 WILEY-LISS, INC.
THE ANATOMICAL RECORD 291:527–537 (2008)
The cellular events that follow crushing or transectionof a peripheral nerve have been extensively documented.To achieve successful nerve repair, axons have to re-grow and find their correct target cells. The first step isthe removal of myelin debris, a process known as Wal-lerian degeneration, and is carried out by residentSchwann cells and infiltrating macrophages. The
Schwann cells de-differentiate, proliferate, and alignwithin basal lamina tubes (bands of Bungner), providinga guidance substrate for growing axons. The re-grownaxons are then myelinated by re-differentiated Schwanncells (Kury et al., 2001).In this study, we examined temporal changes of
SSeCKS expression after crush and transection injuryby real-time polymerase chain reaction (PCR) and West-ern blot analysis. Immunohistochemistry was used todetect the spatial expression of SSeCKS in proximal anddistal portions of injured sciatic nerves. Double stainingof SSeCKS with S100 and with growth-associated protein43 (GAP43) was performed to determine its localizationand possible significance. SSeCKS was shown to colocalizewith the two markers, demonstrating that SSeCKS mightcorrelate with nerve injury and regeneration.
MATERIALS AND METHODS
Animals and Tissue Preparation
Adult (200–300 g) male/female Sprague-Dawley ratswere obtained from the experimental animal center ofNantong University. Experiments were performed in ac-cordance with the National Institutes of Health Guide-lines for the Care and Use of Laboratory Animals. Ani-mals were maintained at constant temperature and hu-midity and kept on a 12-hr day/12-hr night schedule.Rats were anesthetized with pentobarbital (50 mg/kg,i.p.). All procedures were performed on animals in anunconscious state. For animals receiving axotomies, thesciatic nerve was exposed in the mid-gluteal region,crushed unilaterally at the mid-point with a small hemo-stat for 10 sec then unclamped for 10 sec. This proce-
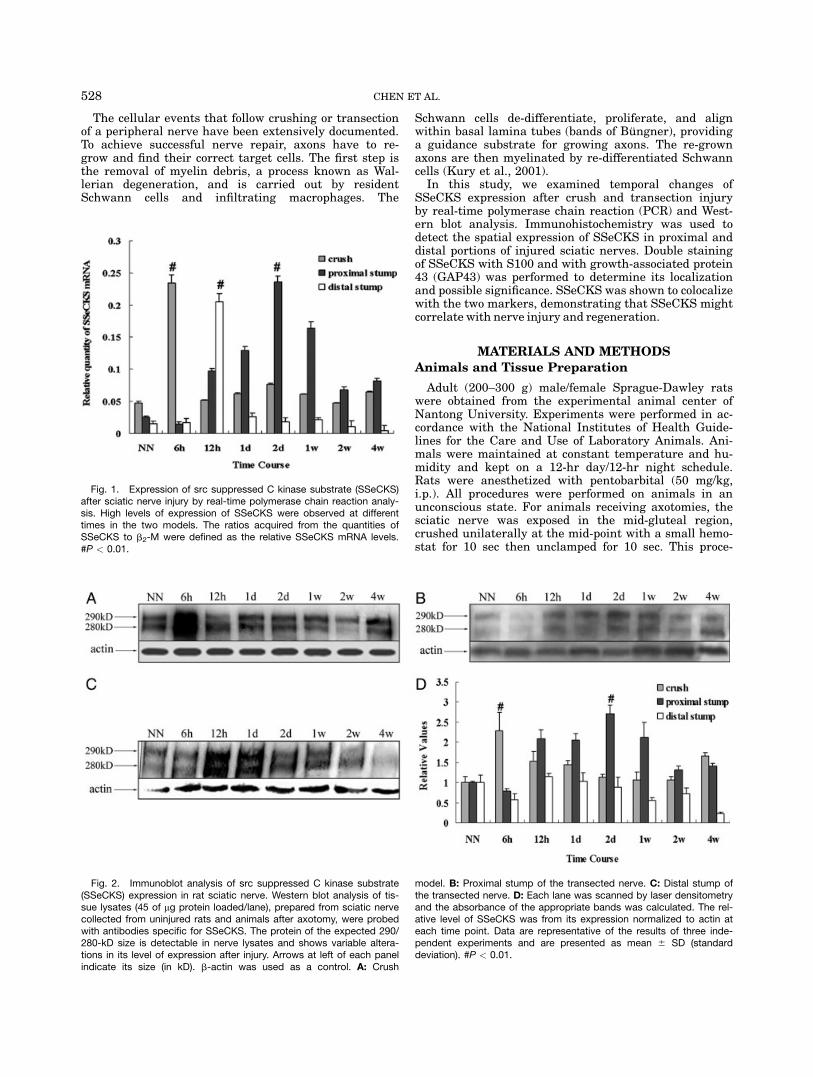
Fig. 2. Immunoblot analysis of src suppressed C kinase substrate(SSeCKS) expression in rat sciatic nerve. Western blot analysis of tis-sue lysates (45 of mg protein loaded/lane), prepared from sciatic nervecollected from uninjured rats and animals after axotomy, were probedwith antibodies specific for SSeCKS. The protein of the expected 290/280-kD size is detectable in nerve lysates and shows variable altera-tions in its level of expression after injury. Arrows at left of each panelindicate its size (in kD). b-actin was used as a control. A: Crush
model. B: Proximal stump of the transected nerve. C: Distal stump ofthe transected nerve. D: Each lane was scanned by laser densitometryand the absorbance of the appropriate bands was calculated. The rel-ative level of SSeCKS was from its expression normalized to actin ateach time point. Data are representative of the results of three inde-pendent experiments and are presented as mean 6 SD (standarddeviation). #P < 0.01.
Fig. 1. Expression of src suppressed C kinase substrate (SSeCKS)after sciatic nerve injury by real-time polymerase chain reaction analy-sis. High levels of expression of SSeCKS were observed at differenttimes in the two models. The ratios acquired from the quantities ofSSeCKS to b2-M were defined as the relative SSeCKS mRNA levels.#P < 0.01.
528 CHEN ET AL.
dure was repeated 3 times. In the transection model, thesciatic nerve was transected at the sciatic notch, andreflected caudally to prevent regeneration. Wounds weresutured and animals were allowed to recover. Water andfood were provided ad libitum until animals were killed.The tissues of corresponding regions from three to fouranimals were pooled. All samples were kept on ice beforeprotein/RNA isolation. For immunohistochemistry, ratswere anesthetized and perfused intracardially with 0.9%saline, followed by fixative (4% paraformaldehyde). Thesciatic nerves were immediately removed, immersion-fixed overnight at 48C, in the same fixative, and subse-quently processed for sectioning.
RNA Isolation and Reverse Transcription
Total RNA was isolated from 100 mg of sciatic nerveusing Trizol. RNA concentration was determined byabsorption at 260 nm, and the 260/280 nm absorption
ratio of the samples was verified to be >1.9. Reversetranscription (RT) was performed on 5 mg RNA per sam-ple using the ThermoScript RT System (Fermentas).cDNA was diluted 1:1 and 2 ml was used in each 20-mlPCR reaction.
Real-time PCR
Total RNA was extracted by Trizol. PCR primers forSSeCKS, and for b2-microglobulin (b2-M) were designedcorresponding to coding regions, as follows: SSeCKS pri-mers, sense 50-AAGTGCTGGCTTCGGAGAAAG-30 andantisense 50-TGACTTCAGGAACTTCAAGGCTC-30; b2-Mprimers, sense 50-GTCTTTCTACATCCTGGCTCACA-30and antisense 50-GACGGTTTTGGGCTCCTTCA-30. Taq-Man probes for SSeCKS, and b2-M were designed cor-responding to coding regions, as follows: SSeCKSprobe, 50(FAM)-AGCCTGTCCAGT- CTCAGAGCCCTGTG-(TAMRA)30 and b2-M probe, 50(FAM)-CACCCACCGA
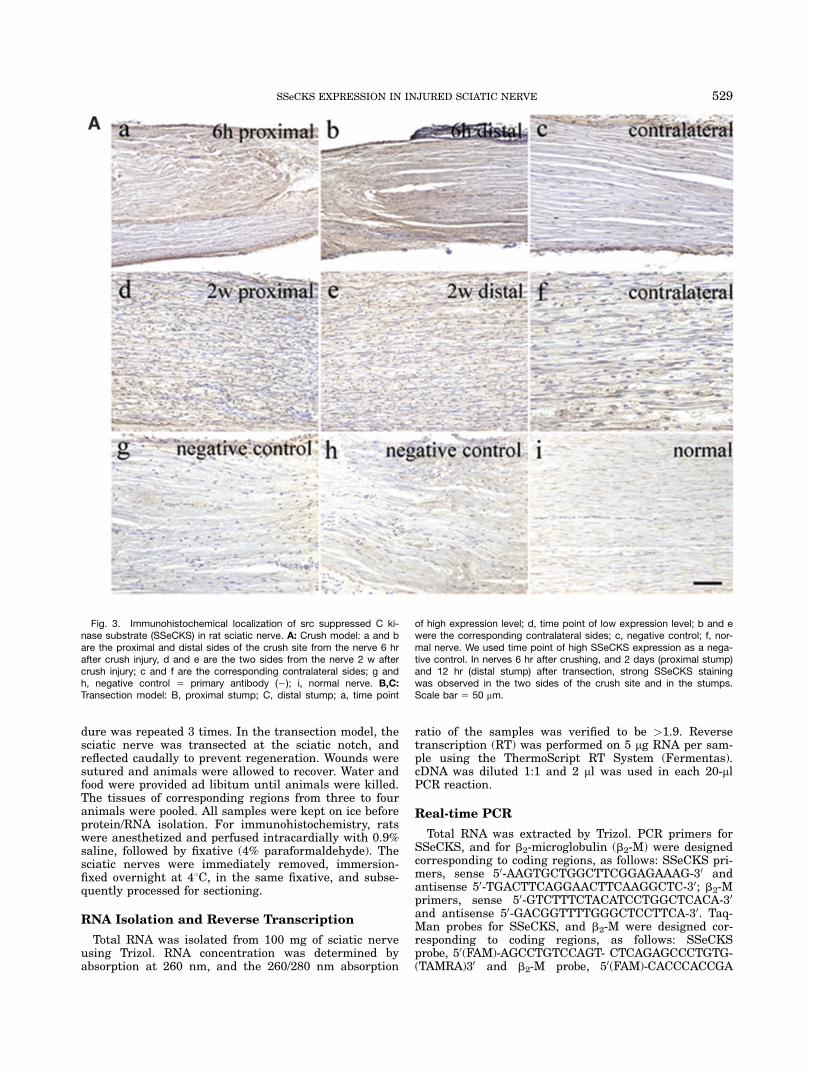
Fig. 3. Immunohistochemical localization of src suppressed C ki-nase substrate (SSeCKS) in rat sciatic nerve. A: Crush model: a and bare the proximal and distal sides of the crush site from the nerve 6 hrafter crush injury, d and e are the two sides from the nerve 2 w aftercrush injury; c and f are the corresponding contralateral sides; g andh, negative control 5 primary antibody (2); i, normal nerve. B,C:Transection model: B, proximal stump; C, distal stump; a, time point
of high expression level; d, time point of low expression level; b and ewere the corresponding contralateral sides; c, negative control; f, nor-mal nerve. We used time point of high SSeCKS expression as a nega-tive control. In nerves 6 hr after crushing, and 2 days (proximal stump)and 12 hr (distal stump) after transection, strong SSeCKS stainingwas observed in the two sides of the crush site and in the stumps.Scale bar 5 50 mm.
529SSeCKS EXPRESSION IN INJURED SCIATIC NERVE
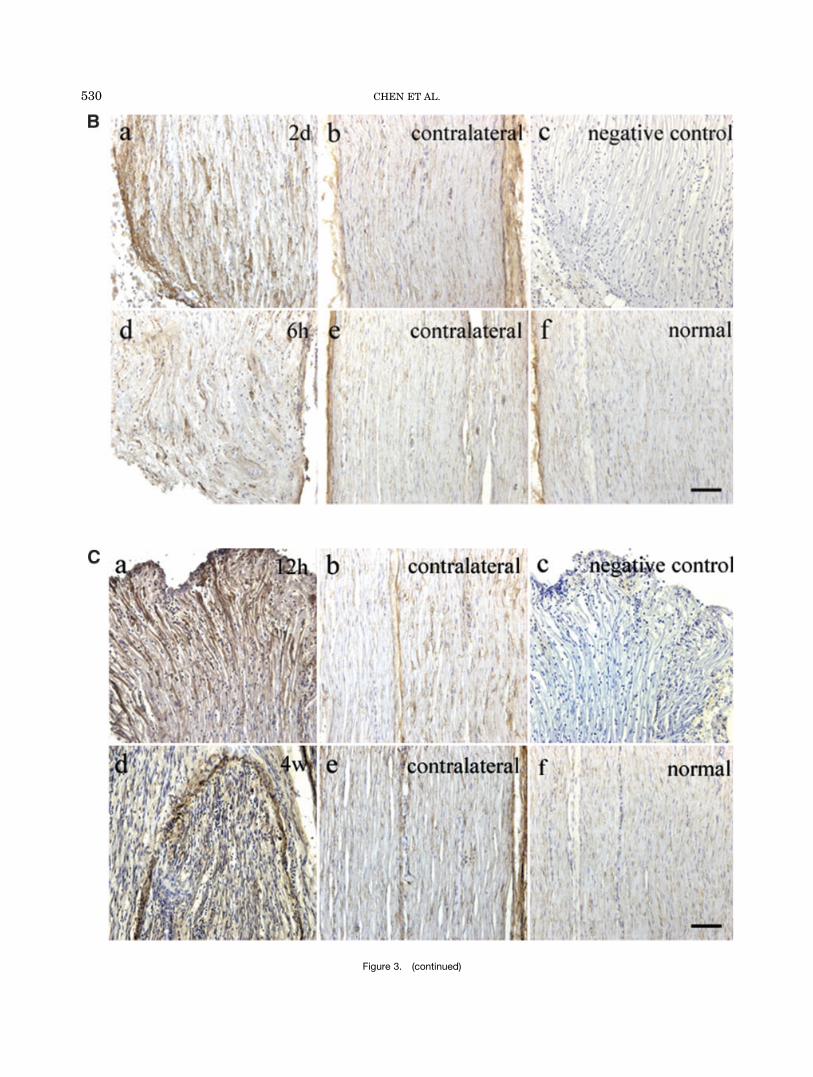
Figure 3. (continued)
530 CHEN ET AL.
GACCGATGTATATGCTTGC-(TAMRA)30. The reactionmixes included 13 PCR buffer, 20 mM magnesium chlo-ride, 0.2 mM deoxyNTP, 10 nmol each TaqMan probewith a pair of 10 nmol SSeCKS primers or 10 nmol b2-Mprimers. Real-time PCR was performed in a Rotor Gene3000 Detector (Perkin-Elmer/Applied Biosystems, FosterCity, CA). The thermal cycling program consisted of3 min at 948C, followed by 40 cycles of 20 sec at 948Cand 1 min at 608C. To account for variability in totalRNA input, the expression of SSeCKS was normalized tob2-M.
Western Blot Analysis
Total protein was isolated from approximately 100 mgof sciatic nerve tissue from normal and injured rats atindicated time points. The length of the sciatic nervewas as follows: (1) in the crush model, the whole nervewas 1 cm and the crush site was in the middle; (2) inthe transection model, the nerve was 1 cm to the stump;(3) the length of the normal sciatic nerve was 2 cmbefore the crotch. Samples were mixed with 2 3 loadingbuffer and dithiothreitol (4:5:1), boiled in water for 5 to
10 min, then cooled on ice. Proteins were resolved on6%/10% sodium dodecyl sulfate-polyacrylamide gel elec-tropheresis gels and transferred to polyvinylidine difluo-ride (Millipore) membranes. The membranes wereblocked with 5% dried skimmed milk in Tris-buffered sa-line with Tween (TBST). After 2 hr at room tempera-ture, filters were washed three times in TBST and thenincubated overnight at 48C with appropriate antibody:sheep polyclonal anti-SSeCKS (Sigma, 1:2,000), mousemonoclonal anti-GAP43 (Sigma, 1:200), mouse monoclo-nal anti-S100 (Sigma, 1:200), or mouse monoclonal anti–b-actin (1:2,000). Finally, rabbit anti-sheep IgG, conju-gated to horseradish peroxidase (Southern Biotech,1:2,000), or goat anti-mouse IgG, conjugated to horserad-ish peroxidase (Pierce, 1:1,000), was added and incu-bated for an additional 2 hours. Blots were developedusing enhanced chemiluminescence (Pierce ChemicalCo., Rockford, IL).
Immunohistochemistry
Immunohistochemistry was performed on 14-mm sec-tions of sciatic nerve, 6 hr after crushing, or on trans-
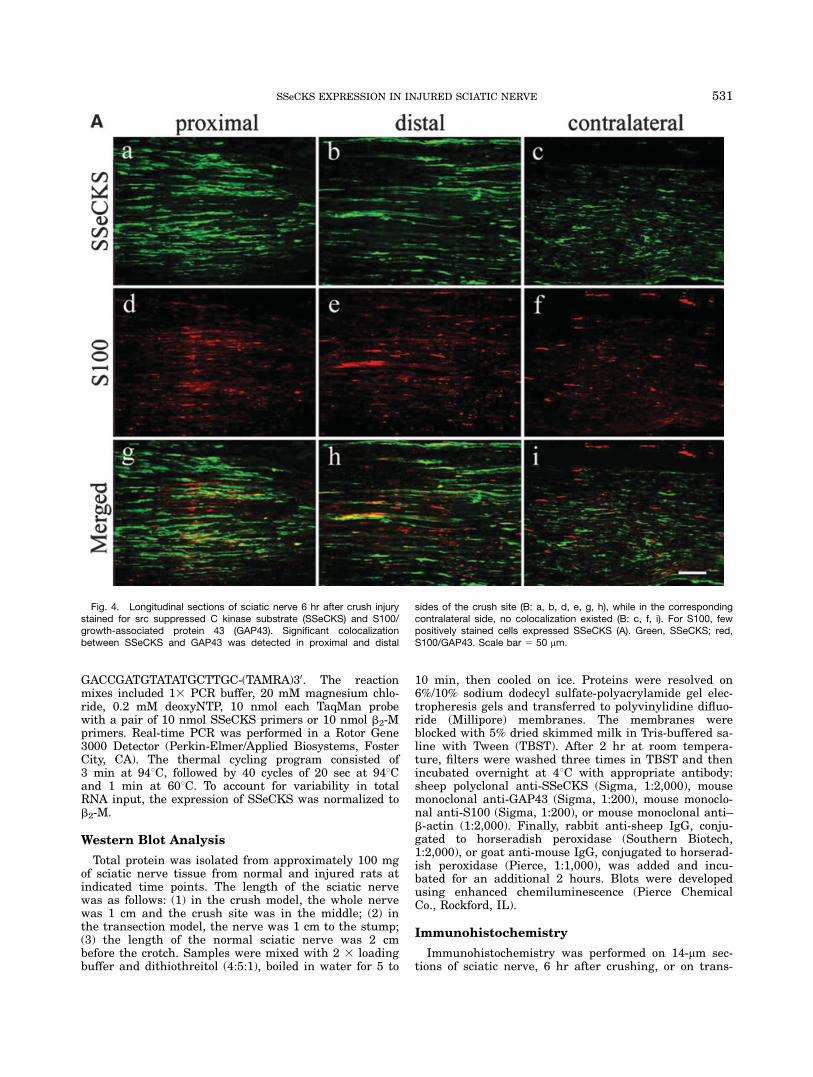
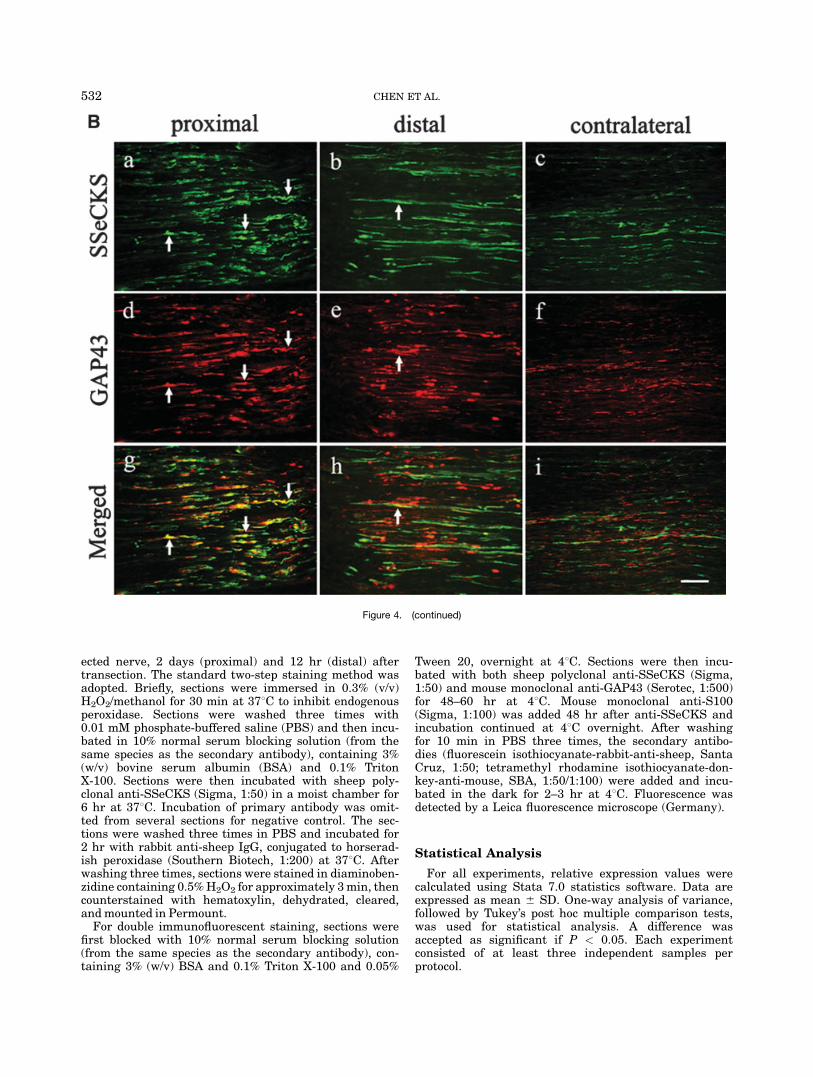
Fig. 4. Longitudinal sections of sciatic nerve 6 hr after crush injurystained for src suppressed C kinase substrate (SSeCKS) and S100/growth-associated protein 43 (GAP43). Significant colocalizationbetween SSeCKS and GAP43 was detected in proximal and distal
sides of the crush site (B: a, b, d, e, g, h), while in the correspondingcontralateral side, no colocalization existed (B: c, f, i). For S100, fewpositively stained cells expressed SSeCKS (A). Green, SSeCKS; red,S100/GAP43. Scale bar 5 50 mm.
531SSeCKS EXPRESSION IN INJURED SCIATIC NERVE
ected nerve, 2 days (proximal) and 12 hr (distal) aftertransection. The standard two-step staining method wasadopted. Briefly, sections were immersed in 0.3% (v/v)H2O2/methanol for 30 min at 378C to inhibit endogenousperoxidase. Sections were washed three times with0.01 mM phosphate-buffered saline (PBS) and then incu-bated in 10% normal serum blocking solution (from thesame species as the secondary antibody), containing 3%(w/v) bovine serum albumin (BSA) and 0.1% TritonX-100. Sections were then incubated with sheep poly-clonal anti-SSeCKS (Sigma, 1:50) in a moist chamber for6 hr at 378C. Incubation of primary antibody was omit-ted from several sections for negative control. The sec-tions were washed three times in PBS and incubated for2 hr with rabbit anti-sheep IgG, conjugated to horserad-ish peroxidase (Southern Biotech, 1:200) at 378C. Afterwashing three times, sections were stained in diaminoben-zidine containing 0.5%H2O2 for approximately 3min, thencounterstained with hematoxylin, dehydrated, cleared,andmounted in Permount.For double immunofluorescent staining, sections were
first blocked with 10% normal serum blocking solution(from the same species as the secondary antibody), con-taining 3% (w/v) BSA and 0.1% Triton X-100 and 0.05%
Tween 20, overnight at 48C. Sections were then incu-bated with both sheep polyclonal anti-SSeCKS (Sigma,1:50) and mouse monoclonal anti-GAP43 (Serotec, 1:500)for 48–60 hr at 48C. Mouse monoclonal anti-S100(Sigma, 1:100) was added 48 hr after anti-SSeCKS andincubation continued at 48C overnight. After washingfor 10 min in PBS three times, the secondary antibo-dies (fluorescein isothiocyanate-rabbit-anti-sheep, SantaCruz, 1:50; tetramethyl rhodamine isothiocyanate-don-key-anti-mouse, SBA, 1:50/1:100) were added and incu-bated in the dark for 2–3 hr at 48C. Fluorescence wasdetected by a Leica fluorescence microscope (Germany).
Statistical Analysis
For all experiments, relative expression values werecalculated using Stata 7.0 statistics software. Data areexpressed as mean 6 SD. One-way analysis of variance,followed by Tukey’s post hoc multiple comparison tests,was used for statistical analysis. A difference wasaccepted as significant if P < 0.05. Each experimentconsisted of at least three independent samples perprotocol.
Figure 4. (continued)
532 CHEN ET AL.
RESULTS
Expression of SSeCKS in Injured Rat
Sciatic Nerve
To reveal any change in the expression of SSeCKS inpostinjury rat sciatic nerve, we chose real-time PCR toanalyze mRNA levels. Our time points included 6 hr,12 hr, 1 days, 2 days, 1 week, 2 weeks, and 4 weeksafter injury. Sciatic nerves from normal rats were used asa control. Total RNA samples were freshly isolated fromnerves of the two different models. As shown in Figure 1,the relative quantity of SSeCKS changed with time afterinjury. In the crush model, it was significantly higher at6 hr after injury. In the transection model, the highestlevel was at 2 days in the proximal stump; whereas in thedistal stump, its peak was at 12 hr after injury.To corroborate our findings, Western blot analysis
was used to determine the abundance of the protein.In crushed sciatic nerve, it reached its highest levelat 6 hr post injury, and then decreased gradually(Fig. 2A). In the proximal stump of the transectednerve, SSeCKS was almost undetectable at 6 hr afterthe operation, and gradually increased and reached itspeak at 2 days (Fig. 2B). Thereafter, the level de-creased. In the distal stump (Fig. 2C), SSeCKS peakedat a relative early stage, 12 hr after transection, andthen levels declined. These results were in good agree-ment with those from real-time PCR analysis, andtogether indicated that the expression of SSeCKS
changed with time after sciatic nerve injury, andin different models, the pattern of expression wasdifferent.
Immunohistochemical Localization of SSeCKS
in Injured Rat Sciatic Nerve
To observe the localization of SSeCKS in the sciaticnerve, we used immunohistochemistry. In the crushmodel, no immunoreactivity was found at the crush site,immediately after crushing. Six hours after crush injury,striking staining of SSeCKS was found on both proximaland distal sides of the crush site (Fig. 3A:a,b). While inthe contralateral side, the staining of SSeCKS wasweaker and its distribution was relatively uniform(Fig. 3A:c). After 2 weeks, the sciatic nerve was under-going repair and had become thicker and the crush sitewas not apparent. At this time, expression was uniformon both sides of the crush site and the correspondingcontralateral side (Fig. 3A:d–f). The expression level2 weeks after crush injury was lower than that after 6 hr.In the transection model, we investigated the expres-
sion pattern of SSeCKS in the proximal and distalstumps (Fig. 3B,C). Intensive staining of SSeCKS wasobserved in the proximal stump of the sciatic nerve2 days after transection (Fig. 3B:a), and it was in a clus-ter distribution. In the same position of the nerve 6 hrafter transection (Fig. 3B:d), much weaker signals weredetected. In the contralateral side of both models, and in
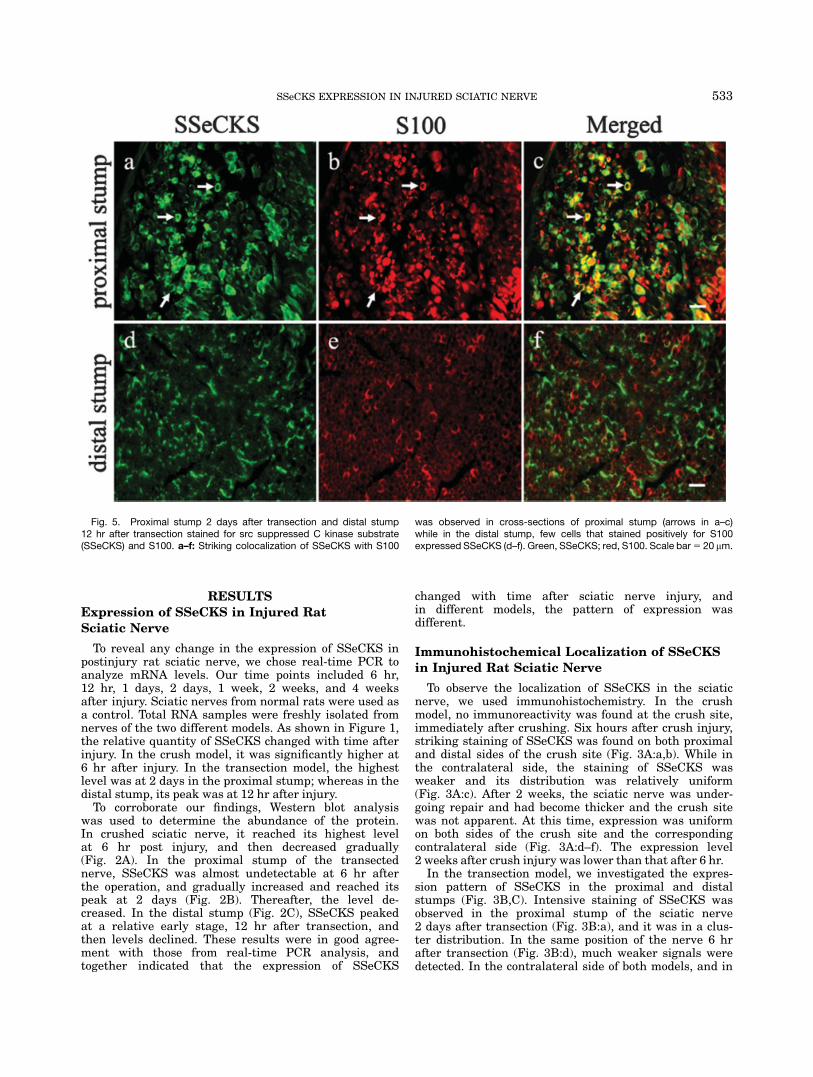
Fig. 5. Proximal stump 2 days after transection and distal stump12 hr after transection stained for src suppressed C kinase substrate(SSeCKS) and S100. a–f: Striking colocalization of SSeCKS with S100
was observed in cross-sections of proximal stump (arrows in a–c)while in the distal stump, few cells that stained positively for S100expressed SSeCKS (d–f). Green, SSeCKS; red, S100. Scale bar5 20 mm.
533SSeCKS EXPRESSION IN INJURED SCIATIC NERVE
normal sciatic nerve, the level of SSeCKS was low anddistribution was relatively uniform (Fig. 3B:b,e,f). Theexpression pattern of SSeCKS in the distal stump of thesciatic nerve, 12 hr and 4 weeks after transection, wassimilar to that of the proximal stump.
Double Immunofluorescent Staining
for SSeCKS and S100 or GAP43
in Injured Sciatic Nerve
To further investigate the localization of SSeCKS incrushed or transected sciatic nerve and to investigatepossible function, we carried out double immunofluores-cent staining to detect whether SSeCKS colocalizes withS100 or with GAP43. Little colocalization betweenSSeCKS and S100, a marker for Schwann cells, wasfound in longitudinal sections of sciatic nerve 6 hr aftercrush or from cross-sections of sciatic nerve 12 hr (distalstump) after transection (Figs. 4A:g,h; Fig. 5f). But inthe proximal stump of the nerve 2 days after transec-tion, striking colocalization was detected in the cross-sections (Fig. 5c). Next, we labeled GAP43, a growth-associated protein, in the injured sciatic nerve. We foundSSeCKS prominently colocalized with GAP43, both incrushed and transected nerve (Figs. 4B, 6). In the sec-tions of all corresponding contralateral nerves, no coloc-alization was observed in either model. We, therefore,
only show the contralateral side of the crush model(Fig. 4c,f,i) as a control.
GAP43 and S100 Also Changed in Injured RatSciatic Nerve
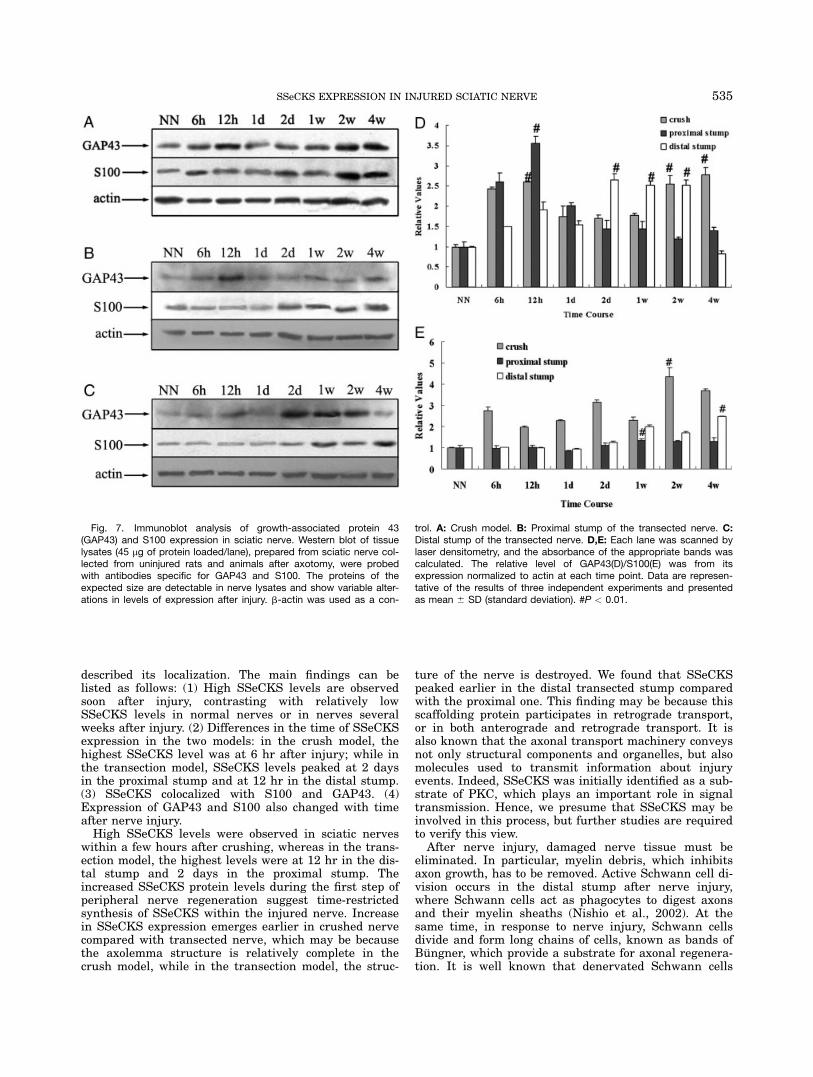
Because the expression of SSeCKS changed afterinjury and we found it colocalized with GAP43/S100 atdifferent time points in different models, we investigatedwhether GAP43 and/or S100 were also up-regulated atthe selected time points. Therefore, Western blot analy-sis was performed to observe the expression pattern ofthe two molecules. The results showed that, in the crushmodel, the level of GAP43 was high 12 hr after injury,and in later stages the protein also had a high level. Inthe transection model, levels peaked at 12 hr in theproximal stump, while in the distal stump, the highlevel was at 2 days and this level was maintained up to2 weeks after injury. The highest level of S100 wasobserved at 2 weeks in the crush model. In the proximalstump, the protein was relatively high 2 days afterinjury and persisted to the end of our experiment. In thedistal stump, the highest level was at 4 weeks (Fig. 7).
DISCUSSION
In the present study, we characterized the alterationsof SSeCKS expression at gene and protein levels in twodifferent injury models of the sciatic nerve, and also
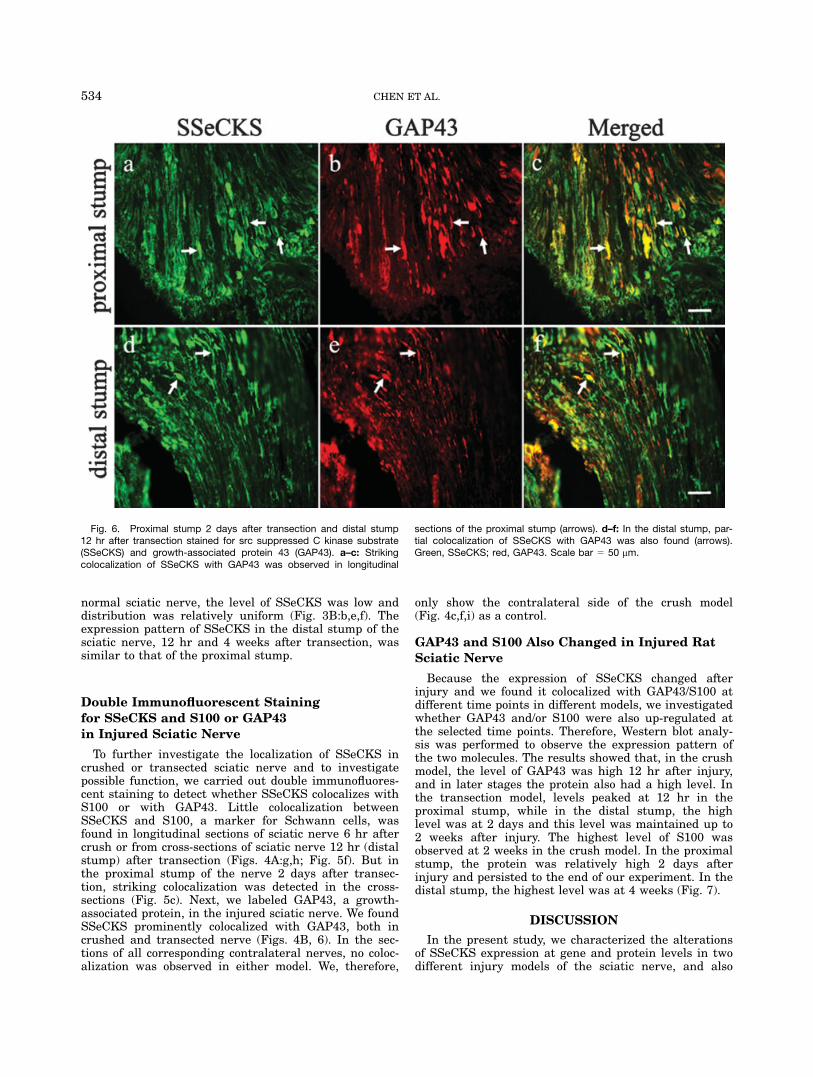
Fig. 6. Proximal stump 2 days after transection and distal stump12 hr after transection stained for src suppressed C kinase substrate(SSeCKS) and growth-associated protein 43 (GAP43). a–c: Strikingcolocalization of SSeCKS with GAP43 was observed in longitudinal
sections of the proximal stump (arrows). d–f: In the distal stump, par-tial colocalization of SSeCKS with GAP43 was also found (arrows).Green, SSeCKS; red, GAP43. Scale bar 5 50 mm.
534 CHEN ET AL.
described its localization. The main findings can belisted as follows: (1) High SSeCKS levels are observedsoon after injury, contrasting with relatively lowSSeCKS levels in normal nerves or in nerves severalweeks after injury. (2) Differences in the time of SSeCKSexpression in the two models: in the crush model, thehighest SSeCKS level was at 6 hr after injury; while inthe transection model, SSeCKS levels peaked at 2 daysin the proximal stump and at 12 hr in the distal stump.(3) SSeCKS colocalized with S100 and GAP43. (4)Expression of GAP43 and S100 also changed with timeafter nerve injury.High SSeCKS levels were observed in sciatic nerves
within a few hours after crushing, whereas in the trans-ection model, the highest levels were at 12 hr in the dis-tal stump and 2 days in the proximal stump. Theincreased SSeCKS protein levels during the first step ofperipheral nerve regeneration suggest time-restrictedsynthesis of SSeCKS within the injured nerve. Increasein SSeCKS expression emerges earlier in crushed nervecompared with transected nerve, which may be becausethe axolemma structure is relatively complete in thecrush model, while in the transection model, the struc-
ture of the nerve is destroyed. We found that SSeCKSpeaked earlier in the distal transected stump comparedwith the proximal one. This finding may be because thisscaffolding protein participates in retrograde transport,or in both anterograde and retrograde transport. It isalso known that the axonal transport machinery conveysnot only structural components and organelles, but alsomolecules used to transmit information about injuryevents. Indeed, SSeCKS was initially identified as a sub-strate of PKC, which plays an important role in signaltransmission. Hence, we presume that SSeCKS may beinvolved in this process, but further studies are requiredto verify this view.After nerve injury, damaged nerve tissue must be
eliminated. In particular, myelin debris, which inhibitsaxon growth, has to be removed. Active Schwann cell di-vision occurs in the distal stump after nerve injury,where Schwann cells act as phagocytes to digest axonsand their myelin sheaths (Nishio et al., 2002). At thesame time, in response to nerve injury, Schwann cellsdivide and form long chains of cells, known as bands ofBungner, which provide a substrate for axonal regenera-tion. It is well known that denervated Schwann cells
Fig. 7. Immunoblot analysis of growth-associated protein 43(GAP43) and S100 expression in sciatic nerve. Western blot of tissuelysates (45 mg of protein loaded/lane), prepared from sciatic nerve col-lected from uninjured rats and animals after axotomy, were probedwith antibodies specific for GAP43 and S100. The proteins of theexpected size are detectable in nerve lysates and show variable alter-ations in levels of expression after injury. b-actin was used as a con-
trol. A: Crush model. B: Proximal stump of the transected nerve. C:Distal stump of the transected nerve. D,E: Each lane was scanned bylaser densitometry, and the absorbance of the appropriate bands wascalculated. The relative level of GAP43(D)/S100(E) was from itsexpression normalized to actin at each time point. Data are represen-tative of the results of three independent experiments and presentedas mean 6 SD (standard deviation). #P < 0.01.
535SSeCKS EXPRESSION IN INJURED SCIATIC NERVE

increase production of several neurotrophic molecules,including nerve growth factor (Heumann et al., 1987)and brain-derived neurotrophic factor (Meyer et al.,1992). Axon–Schwann cell interaction, mediated by neu-rotrophic factors, may play a pivotal role in peripheralnerve regeneration (Liu et al., 1995). Increased produc-tion of trophic factors by Schwann cells may substitutefor the usual target-derived trophic factors. Using anti–S-100 antibody for Schwann cell identification, we foundthat few Schwann cells showed SSeCKS immunostain-ing within the first few hours after injury, but strongSSeCKS staining in Schwann cells was seen in cross-sec-tions of the proximal stump 2 days after transection.This finding is consistent with the early stage of sciaticnerve injury, when most Schwann cells undergo apopto-sis and are not in their activated state. However, sometime after injury, Schwann cells start to divide, prolifer-ate, and play related biologic activity. The strongSSeCKS immunostaining observed in Schwann cells atthis time indicated that SSeCKS might contribute to themechanism of these complicated changes that areinduced by different signals.Normally, successful axonal regeneration is accompa-
nied by the appearance of numerous, functionallydiverse families of molecules that regulate surface-cytos-keletelal interaction (Makwana and Raivich, 2005).Zheng and others suggested that intra-axonal transla-tion of cytoskeletal components was required for sustain-ing growth cones in regenerating axons (Zheng et al.,2001). GAP43 is a membrane- and cytoskeletal-associ-ated phosphoprotein (Benowitz and Routtenberg, 1987;Skene, 1989; Coggins and Zwiers, 1991) that isexpressed at high levels in neurons during developmentand is concentrated in axonal growth cones (Biffo et al.,1990; Fitzgerald et al., 1991). After neural injury in theadult, however, GAP43 is re-expressed and is rapidlytransported along the axons of those neurons wherethere is successful regeneration (Bisby, 1988; Tetzlaffet al., 1989; Van der Zee et al., 1989; Woolf et al., 1990).The correlation between the presence of GAP43 and thegrowth state of neurons has led to the use of GAP43 asa marker for axonal growth, and evidence from in vitrostudies suggest that GAP43 is directly involved in suchgrowth (Zuber et al., 1989; Yanker et al., 1990; Sheaet al., 1991; Jap Tjoen San et al., 1992; Aigner and Car-oni, 1993; Widemer and Caroni, 1993). Thus, the colocal-ization of SSeCKS with GAP43 was performed to deter-mine whether SSeCKS associated with regeneration.Our results showed that the two molecules colocalized incrushed nerve and in the two transected stumps. Thisfinding indicates that SSeCKS may have a function inthe regeneration process of the injured nerve. For exam-ple, it may participate in the transmission of some sig-nals, effecting the reformation of the cystoskeleton andso on. The observation raises the potential role ofSSeCKS in regenerative tissue. However, the demonstra-tion of a direct role of SSeCKS in nerve regenerationrequires further investigation.Taken together, these findings indicate that SSeCKS
participates in the adaptive response to peripheral nerveinjury and may play a role in nerve regeneration. How-ever, its exact implication has yet to be defined. Maybe,it participates in part by means of partial replication ofthe molecular and cellular mechanisms that operate dur-ing development. A better understanding of its contribu-
tion may generate new strategies for enhancing periph-eral nerve regeneration.
LITERATURE CITED
Aigner L, Caroni P. 1993. Depletion of 43-kD growth-associated pro-tein in primary sensory neurons leads to diminished formationand spreading of growth cones. J Cell Biol 123:417–429.
Benowitz LI, Routtenberg A. 1987. A membrane phosphoproteinassociated with neural development, axonal regeneration, phos-pholipid metabolism and synaptic plasticity. Trends Neurosci 10:527–531.
Biffo S, Verhaagen J, Shrama LH, Schotman P, Danho W, MargolisFL. 1990. B-50/GAP-43 expression correlates with process out-growth in the embryonic mouse nervous system. Eur J Neurosci2:487–499.
Bisby MA. 1988. Dependence of GAP-43 (B50,Fl) transport on axo-nal regeneration in rat dorsal root ganglion neurons. Brain Res458:157–161.
Chapline C, Mousseau B, Ramsay K, Duddy S, Li Y, Kiley SC,Jaken S. 1996. Identification of a major protein kinase C-bindingprotein and substrate in rat embryo fibroblasts. Decreasedexpression in transformed cells. J Biol Chem 271:6417–6422.
Coggins PJ, Zwiers H. 1991. Biochemistry and functional neuro-chemistry of a neuron-specific phosphoprotein. J Neurochem 56:1095–1106.
Fitzgerald M, Reynolds ML, Benowitz LI. 1991. GAP-43 expressionin the developing rat lumbar spinal cord. Neuroscience 41:187–199.
Gelman IH, Tombler E, Vargas J Jr. 2000. A role for SSeCKS, amajor protein kinase C substrate with tumor suppressor activity,in cytoskeletal architecture, formation of migratory processes,and cell migration during embryogenesis. Histochem J 32:13–26.
Heumann R, Korsching S, Bandtlow C, Thoenen H. 1987. Changesof nerve growth factor synthesis in nonneuronal cells in responseto sciatic nerve transection. J Cell Biol 104:1623–1631.
Jap Tjoen San ER, Schmidt-Michels M, Oestreicher AB, GispenWH, Schotman P. 1992. Inhibition of nerve growth factor-inducedB-50/GAP-43 expression by antisense oligomers interferes withneurite outgrowth of PC 12 cells. Biochem Biophys Res Commun187:839–846.
Kury P, Stoll G, Muller HW. 2001. Molecular mechanisms of cellularinteractions in peripheral nerve regeneration. Curr Opin Neurol14:635–639.
Liu HM, Yang LH, Yang YJ. 1995. Schwann cell properties: 3. C-fosexpression, bFGF production, phagocytosis and proliferation dur-ing Wallerian degeneration. J Neuropathol Exp Neurol 54:487–496.
Makwana M, Raivich G. 2005. Molecular mechanisms in successfulperipheral regeneration. FEBS J 272:2628–2638.
Meyer M, Matsuoka I, Wetmore C, Olson L, Thoenen H. 1992.Enhanced synthesis of brain-derived neurotrophic factor in thelesioned peripheral nerve: different mechanisms are responsiblefor the regulation of BDNF and NGF mRNA. J Cell Biol 119:45–54.
Nishio Y, Nishihira J, Ishibashi T, Kato H, Minami A. 2002. Role ofmacrophage migration inhibitory factor (MIF) in peripheral nerveregeneration: anti-MIF antibody induces delay of nerve regenera-tion and the apoptosis of Schwann cells. Mol Med 8:509–520.
Shea TB, Perrone-Bizzozero NI, Beermann ML, Benowitz LI. 1991.Phospholipid-mediated delivery of anti-GAP-43 antibodies intoneuroblastoma cells prevents neuritogenesis. J Neurosci 11:1685–l690.
Siegel SM, Grove BD, Carr PA. 2002. SSeCKS immunolabeling inrat primary sensory neurons. Brain Res 926:126–136.
Skene JH. 1989. Axonal growth-associated proteins. Annu Rev Neu-rosci 12:127–156.
Tetzlaff W, Zwiers H, Lederis K, Cassar L, Richardson P, Bisby MA.1989. Axonal transport and localization of B-50/GAP-43-likeimmunoreactivity in regenerating sciatic and facial nerves of therat. J Neurosci 9:1303–l313.
536 CHEN ET AL.

Van der Zee CE, Nielander HB, Vos JP, Lopes da Silva S, Verhaa-gen J, Oestreicher AB, Schrama LH, Schotman P, Gispen WH.1989. Expression of growth-associated protein B-50 (GAP43) indorsal root ganglia and sciatic nerve during regenerative sprout-ing. J Neurosci 9:3505–3512.
Widmer F, Caroni P. 1993. Phosphorylation-site mutagenesis of thegrowth-associated protein GAP-43 modulates its effects on cellspreading and morphology. J Cell Biol 120:503–512.
Woolf CJ, Reynolds M, Molander C, O’Brien C, Lindsay RM, Beno-witz LI. 1990. GAP-43 appears in the rat dorsal horn followingperipheral nerve injury. Neuroscience 34:465–478.
Yanker BA, Benowitz LI, Villa-Komaroff L, Neve RL. 1990.Transfection of PC12 cells with the human GAP-43 gene:effects on neurite outgrowth and regeneration. Mol Brain Res7:39–44.
Zheng JQ, Kelly TK, Chang B, Ryazantsev S, Rajasekaran AK,Martin KC, Twiss JL. 2001. A functional role for intra-axonal pro-tein synthesis during axonal regeneration from adult sensoryneurons. J Neurosci 21:9291–9303.
Zuber MX, Goodman DW, Kams LR, Fishman MC. 1989. The neuro-nal growth-associated protein GAP-43 induces filopodia in non-neuronal cells. Science 244:1193–1195.
537SSeCKS EXPRESSION IN INJURED SCIATIC NERVE