Embed Size (px)
Citation preview

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat)
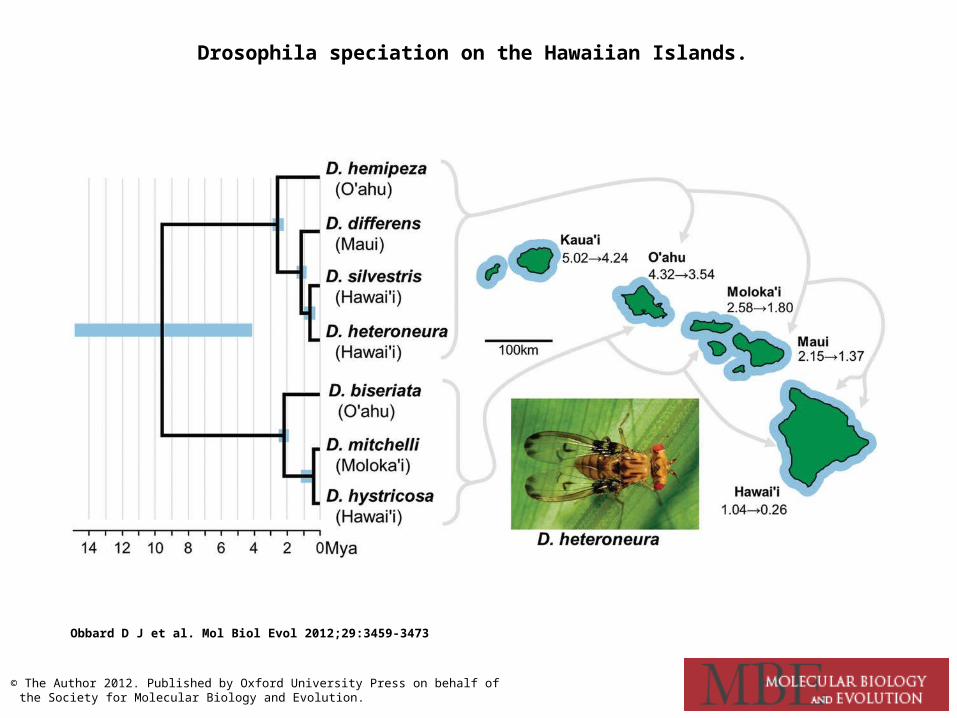
Drosophila speciation on the Hawaiian Islands.
Obbard D J et al. Mol Biol Evol 2012;29:3459-3473
© The Author 2012. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.

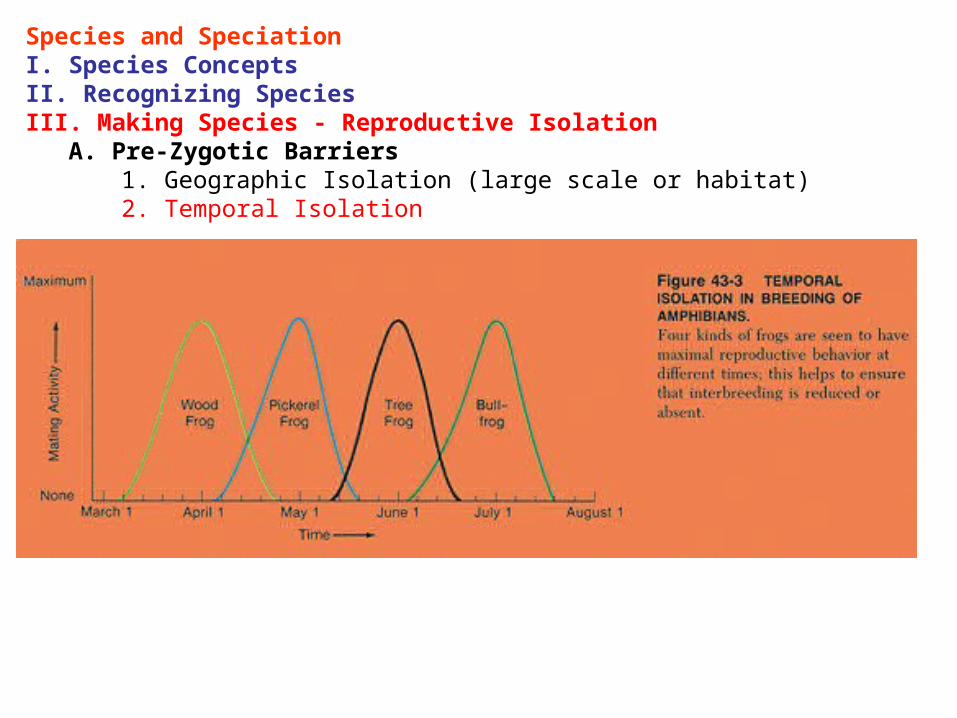
Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates
Western Meadowlark Eastern Meadowlark

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates 4. Mechanical isolation - genitalia don't fit; limit pollinators

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates 4. Mechanical isolation - genitalia don't fit 5. Gametic Isolation - gametes transfered but sperm can't fertilize egg; this
is a common isolation mechanism in species that spawn at the same time

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates 4. Mechanical isolation - genitalia don't fit 5. Gametic Isolation - gametes transfered but sperm can't fertilize egg
B. Post-Zygotic Isolation

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates 4. Mechanical isolation - genitalia don't fit 5. Gametic Isolation - gametes transfered but sperm can't fertilize egg
B. Post-Zygotic Isolation1. Genomic Incompatibility - zygote dies

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates 4. Mechanical isolation - genitalia don't fit 5. Gametic Isolation - gametes transfered but sperm can't fertilize egg
B. Post-Zygotic Isolation1. Genomic Incompatibility - zygote dies 2. Hybrid Inviability - F1 has lower survival
Crazy hybrids

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates 4. Mechanical isolation - genitalia don't fit 5. Gametic Isolation - gametes transfered but sperm can't fertilize egg
B. Post-Zygotic Isolation1. Genomic Incompatibility - zygote dies 2. Hybrid Inviability - F1 has lower survival 3. Hybrid Sterility - F1 has reduced reproductive success
Horse: 64 chromosomesDonkey: 62 chromosomesMule: 63 chromosomes

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive Isolation A. Pre-Zygotic Barriers
1. Geographic Isolation (large scale or habitat) 2. Temporal Isolation 3. Behavior Isolation - don't recognize one another as mates 4. Mechanical isolation - genitalia don't fit 5. Gametic Isolation - gametes transfered but sperm can't fertilize egg
B. Post-Zygotic Isolation1. Genomic Incompatibility - zygote dies 2. Hybrid Inviability - F1 has lower survival 3. Hybrid Sterility - F1 has reduced reproductive success 4. F2 breakdown - F1's survive but F2's have incompatible combo's of genes

AABB x aabb
F1: AaBb = ok
F2:A-B- = okA-bb = noaaB- = noaabb = ok

Species and SpeciationI. Species ConceptsII. Recognizing SpeciesIII. Making Species - Reproductive IsolationIV. Speciation

Speciation
Speciation is not a goal, or a selective product of adaptation. It is simply a consequence of genetic changes that occurred for other reasons (selection, drift, mutation, etc.).

Speciation
I. Modes:
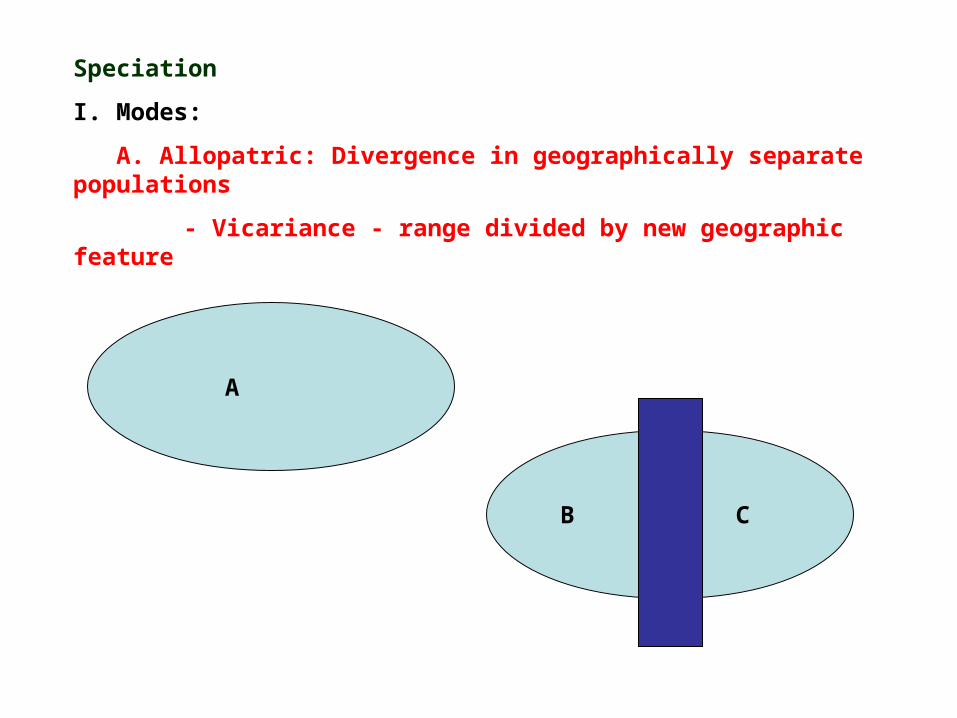
Speciation
I. Modes:
A. Allopatric: Divergence in geographically separate populations
- Vicariance - range divided by new geographic feature
A
B C

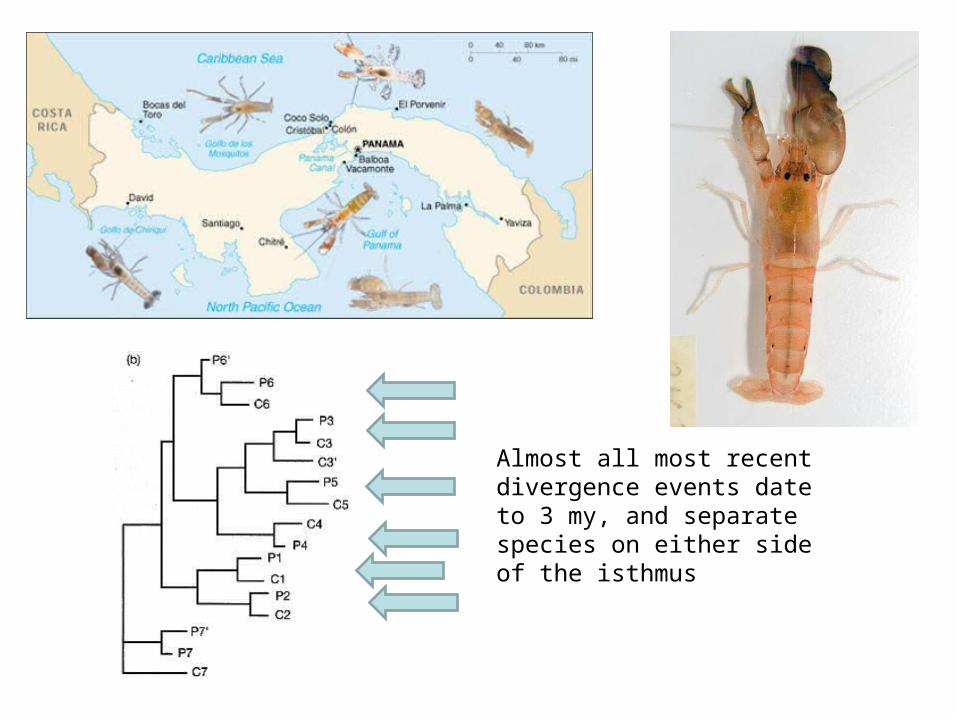
Almost all most recent divergence events date to 3 my, and separate species on either side of the isthmus
Speciation
I. Modes:
A. Allopatric: Divergence in geographically separate populations
- Vicariance - range divided by new geographic feature
- Peripatric - divergence of a small migrant population
A
B

Abert’s Squirrel Kaibab Squirrel
Crossed grand canyon to the north during Ice Age and isolated.

Speciation
I. Modes:
A. Allopatric: Divergence in geographically separate populations
- Vicariance - range divided by new geographic feature
- Peripatric - divergence of a small migrant population
B. Parapatric - neighboring populations diverge, even with gene flow
Speciation
I. Modes:
A. Allopatric: Divergence in geographically separate populations
- Vicariance - range divided by new geographic feature
- Peripatric - divergence of a small migrant population
B. Parapatric - neighboring populations diverge, even with gene flow

B. Parapatric - neighboring populations diverge, even with gene flow
Hybrid
Hybrid Backcross??
Speciation
I. Modes:
A. Allopatric: Divergence in geographically separate populations
- Vicariance - range divided by new geographic feature
- Peripatric - divergence of a small migrant population
B. Parapatric - neighboring populations diverge, even with gene flow
C. Sympatric: Divergence within a single population

C. Sympatric: Divergence within a single population
Maynard Smith (1966) - hypothesized this was possible if there was disruptive selection within a population - perhaps as a specialist herbivore/parasite colonized and adapted to a new host.

C. Sympatric: Divergence within a single population
Maynard Smith (1966) - hypothesized this was possible if there was disruptive selection within a population - perhaps as a specialist herbivore/parasite colonized and adapted to a new host.
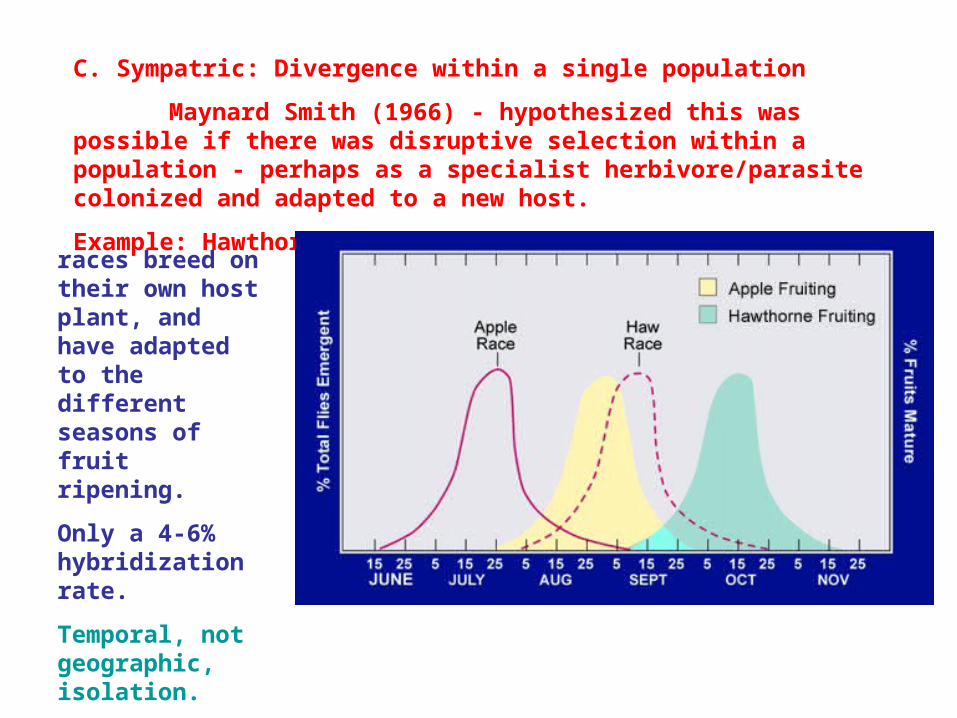
Example: Hawthorn/Apple Maggot Fly (Rhagoletis pomonella)
Hawthorn maggot fly is a native species that breeds on Hawthorn (Crataegus sp.)

C. Sympatric: Divergence within a single population
Maynard Smith (1966) - hypothesized this was possible if there was disruptive selection within a population - perhaps as a specialist herbivore/parasite colonized and adapted to a new host.
Example: Hawthorn/Apple Maggot Fly (Rhagoletis pomonella)
Europeans brought apples to North America. They are in the same plant family (Rosaceae) as Hawthorn.

C. Sympatric: Divergence within a single population
Maynard Smith (1966) - hypothesized this was possible if there was disruptive selection within a population - perhaps as a specialist herbivore/parasite colonized and adapted to a new host.
Example: Hawthorn/Apple Maggot Fly (Rhagoletis pomonella)
Europeans brought apples to North America. They are in the same plant family (Rosaceae) as Hawthorn.
In 1864, apple growers noticed infestation by Apple Maggot flies...which were actually just "hawthorn flies"...
C. Sympatric: Divergence within a single population
Maynard Smith (1966) - hypothesized this was possible if there was disruptive selection within a population - perhaps as a specialist herbivore/parasite colonized and adapted to a new host.
Example: Hawthorn/Apple Maggot Fly (Rhagoletis pomonella)
races breed on their own host plant, and have adapted to the different seasons of fruit ripening.
Only a 4-6% hybridization rate.
Temporal, not geographic, isolation.

C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977a and 1977b. Science.
Two species of green lacewings - generalist insect predators
Chrysopa downesi has one generation in early spring
C. carnea breeds and has three generations in summer
C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977a and 1977b. Science.
Two species of green lacewings - generalist insect predators
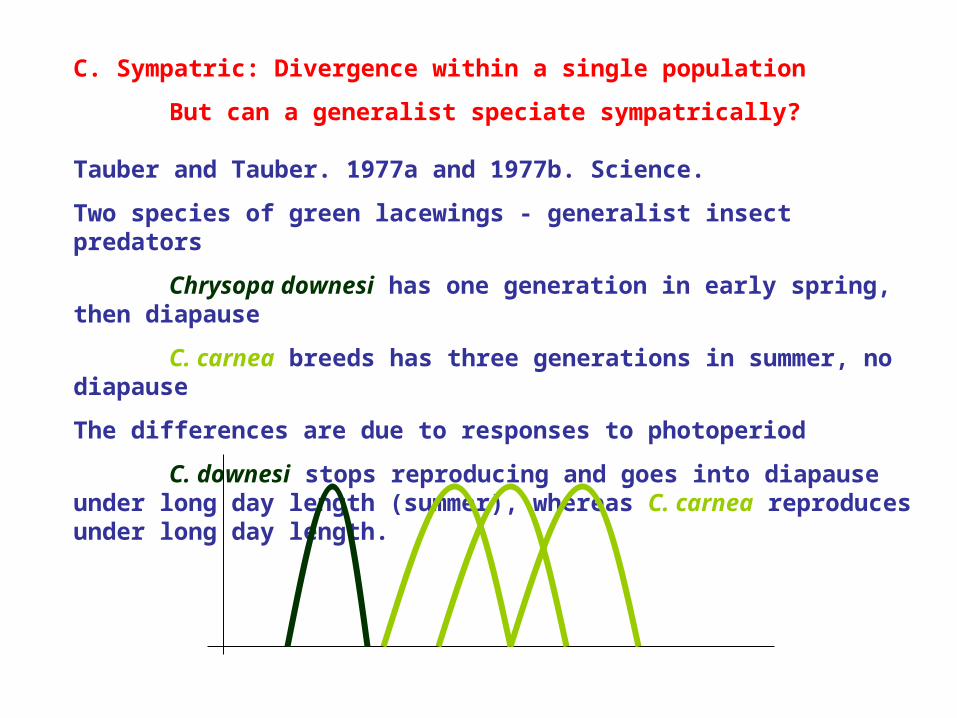
Chrysopa downesi has one generation in early spring, then diapause
C. carnea breeds has three generations in summer, no diapause
The differences are due to responses to photoperiod
C. downesi stops reproducing and goes into diapause under long day length (summer), whereas C. carnea reproduces under long day length.

C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977a. Science 197:592.
The species are completely interfertile in the lab:
Did reciprocal matings: C. downesi x C. carea
Reared F1 offspring under long day length (16L:8D). Found all F1 did not enter diapause (C. carnea photoperiod response is dominant).

C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977a. Science 197:592.
Did F1 x F1 cross: Found 7% (~1/16) of F2 exhibited diapause at 16L:8D. This is consistent with a model of 2 independently assorting autosomal genes with complete dominance at each and an additive effect.
AABB x aabb
F1 all A-B- phenotype
F2 A-B- = 9/16
A-bb = 3/16
aaB- = 3/16
aabb = 1/16.... ~ 7%
C. carnea photoperiod
C. downesi photoperiod

C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977a. Science 197:592.
F1 x C. downesi backcross had 3:1 ratio, as expected of model.
AaBb x aabb
AaBb = .25
Aabb = .25
aaBb = .25
aabb = .25
C. carnea photoperiod
C. downesi photoperiod

C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977b. Science 197:1298.
How did this temporal separation get established?
C. downesi is dark green and prefers hemlock forests
C. carnea is light green and prefers fields and meadows
Difference governed by a single locus where dark is incompletely dominant.

C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977b. Science 197:1298.
How did this temporal separation get established?
C. downesi is dark green and prefers hemlock forests
C. carnea is light green and prefers fields and meadows
Difference governed by a single locus where dark is incompletely dominant.
Hypothesize that selection for different morphs in different habitats created the stable dimorphism, reinforced by inbreeding within the habitats.
intermediate heterozygote

C. Sympatric: Divergence within a single population
But can a generalist speciate sympatrically?
Tauber and Tauber. 1977b. Science 197:1298.
How did this temporal separation get established?
C. downesi is dark green and prefers hemlock forests
C. carnea is light green and prefers fields and meadows
Difference governed by a single locus where dark is incompletely dominant.
Hypothesize that selection for different morphs in different habitats created the stable dimorphism, reinforced by inbreeding within the habitats.
Selection then favored early breeding in C. downesi, as that is when insects feeding on conifers are most abundant.

Speciation
I. Modes
II. Mechanisms

Speciation
I. Modes
II. Mechanisms
A. Progressive Genomic Incompatibility
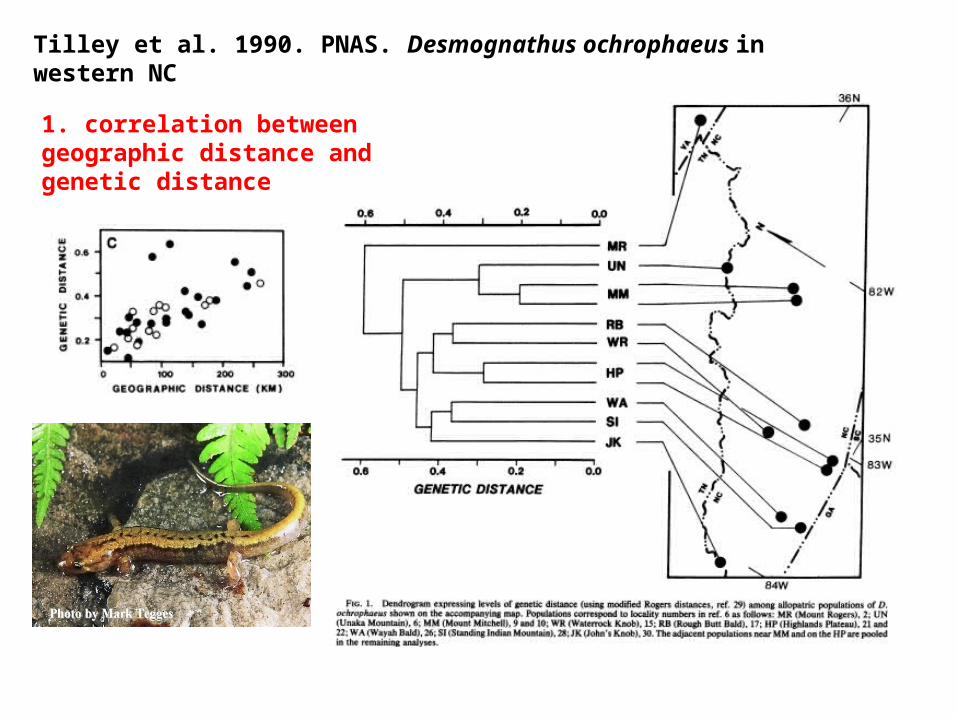
Tilley et al. 1990. PNAS. Desmognathus ochrophaeus in western NC
1. correlation between geographic distance and genetic distance
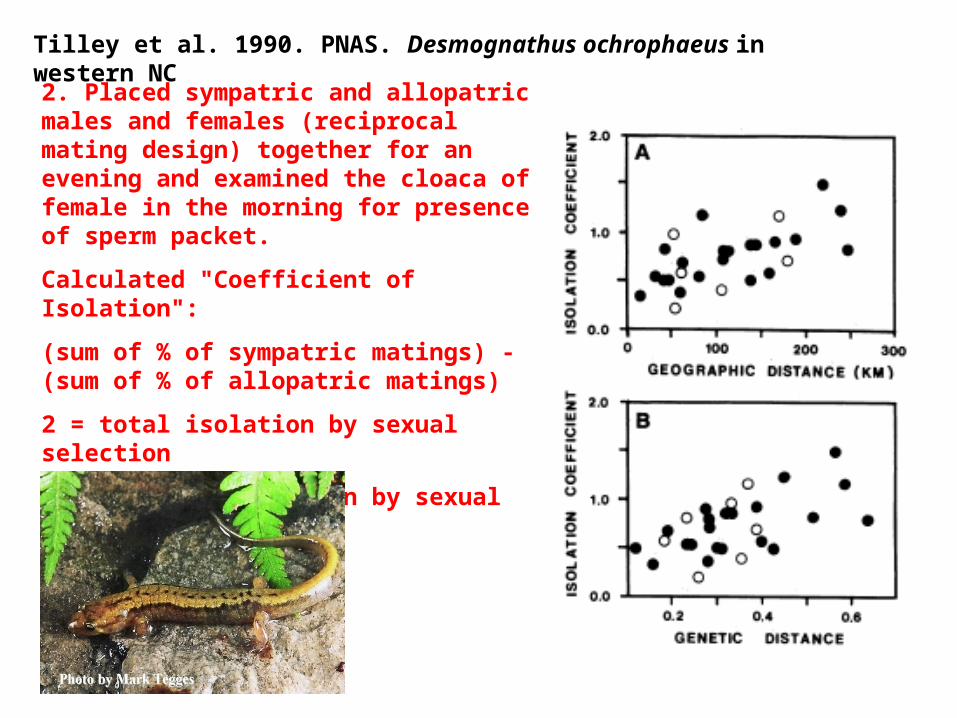
Tilley et al. 1990. PNAS. Desmognathus ochrophaeus in western NC
2. Placed sympatric and allopatric males and females (reciprocal mating design) together for an evening and examined the cloaca of female in the morning for presence of sperm packet.
Calculated "Coefficient of Isolation":
(sum of % of sympatric matings) - (sum of % of allopatric matings)
2 = total isolation by sexual selection
0 = no differentiation by sexual selection

Speciation
I. Modes
II. Mechanisms
A. Progressive Genomic Incompatibility
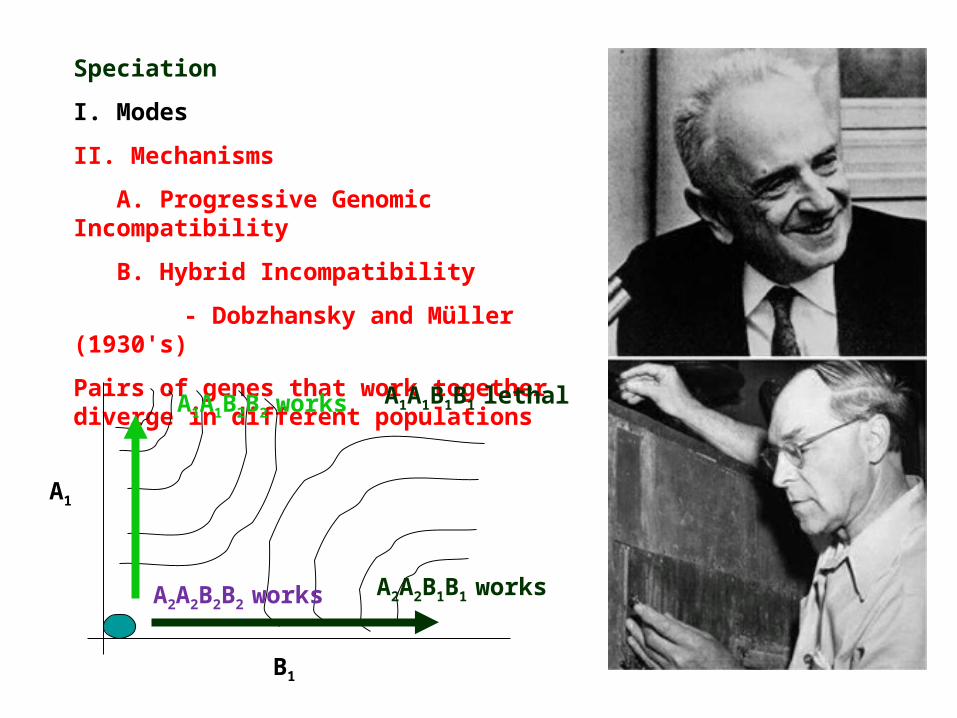
B. Hybrid Incompatibility
- Dobzhansky and Müller (1930's)
Pairs of genes that work together diverge in different populations
Speciation
I. Modes
II. Mechanisms
A. Progressive Genomic Incompatibility
B. Hybrid Incompatibility
- Dobzhansky and Müller (1930's)
Pairs of genes that work together diverge in different populations
A1
B1
A1A1B2B2 works
A2A2B1B1 worksA2A2B2B2 works
A1A1B1B1 lethal

B. Hybrid Incompatibility
D. melanogaster and D. simulans

B. Hybrid Incompatibility
D. melanogaster and D. simulans
Cross female D. mel. x male D. sim - no sons

B. Hybrid Incompatibility
D. melanogaster and D. simulans
Cross female D. mel. x male D. sim - no sons
- Watanabe - 1970 - isolated a mutant strain of D. simulans (w) that could make males with D. melanogaster....

B. Hybrid Incompatibility
D. melanogaster and D. simulans
Cross female D. mel. x male D. sim - no sons
- Watanabe - 1970 - isolated a mutant strain of D. simulans (w) that could make males with D. melanogaster....
- Hypothesized that this strain had a mutant gene partner that reestablished function with the D. melanogaster partner gene... called it "lethal hybrid rescue" (lhr).

B. Hybrid Incompatibility
D. melanogaster and D. simulans
Cross female D. mel. x male D. sim - no sons
- Watanabe - 1970 - isolated a mutant strain of D. simulans (w) that could make males with D. melanogaster....
- Hypothesized that this strain had a mutant gene partner that reestablished function with the D. melanogaster partner gene... called it "lethal hybrid rescue" (lhr).
- Ashburner - 1980 - isolated a mutant strain of D. melanogaster (a) females that could breed with D. simulans males and produce sons ...called it "hybrid male rescue" - hmr - X-linked
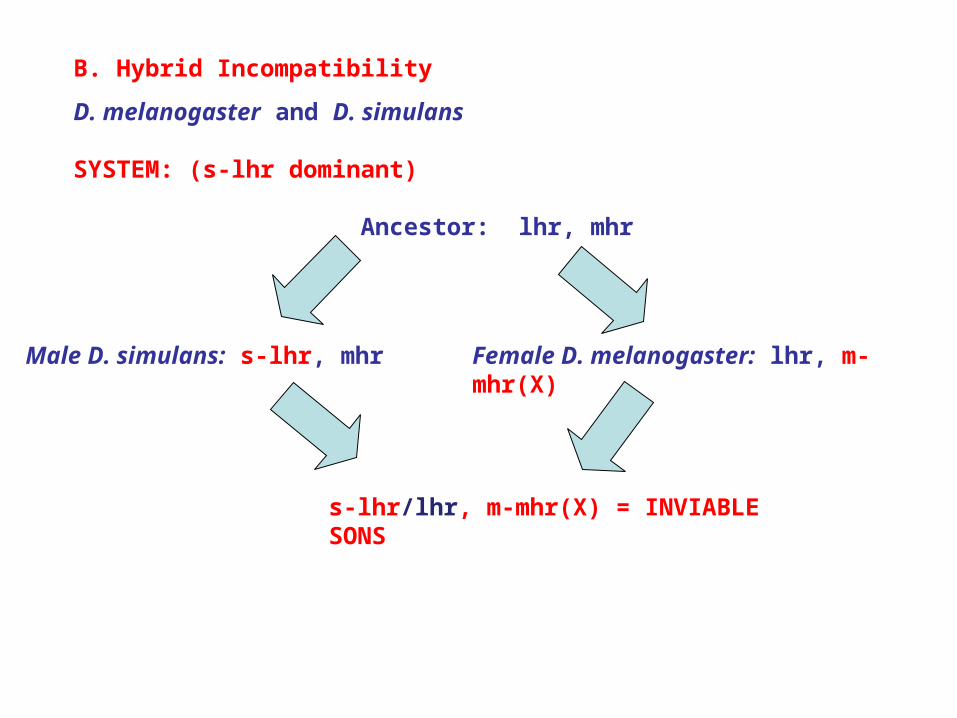
B. Hybrid Incompatibility
D. melanogaster and D. simulans
SYSTEM: (s-lhr dominant)
Ancestor: lhr, mhr
Male D. simulans: s-lhr, mhr Female D. melanogaster: lhr, m-mhr(X)
s-lhr/lhr, m-mhr(X) = INVIABLE SONS

B. Hybrid Incompatibility
D. melanogaster and D. simulans
SYSTEM: (s-lhr dominant)
D. sim = s-lhr, hmr (X) x D. mel = lhr, m-hmr (X) SONS GET : s-lhr/lhr, m-hmr/Y (only X) .... INVIABLE

B. Hybrid Incompatibility
D. melanogaster and D. simulans
SYSTEM:
D. sim = s-lhr, hmr (X) x D. mel = lhr, m-hmr (X) SONS GET : s-lhr/lhr, m-hmr (only X) .... INVIABLE
(w)D. sim = lhr/s-lhr, hmr (X) x D. mel = lhr, m-hmr (X) 1/2 SONS GET lhr/lhr, m-hmr (ONLY X) = VIABLE

B. Hybrid Incompatibility
D. melanogaster and D. simulans
SYSTEM:
D. sim = s-lhr, hmr (X) x D. mel = lhr, m-hmr (X) SONS GET : s-lhr/lhr, m-hmr (only X) .... INVIABLE
(w)D. sim = lhr/s-lhr, hmr (X) x D. mel = lhr, m-hmr (X) 1/2 SONS GET lhr/lhr, m-hmr (ONLY X) = VIABLE
D. sim = s-lhr, hmr (X) x (a) D. mel = lhr, m-hmr(X)/hmr (X) 1/2 SONS GET: s-lhr/lhr, hmr (only X) = VIABLE
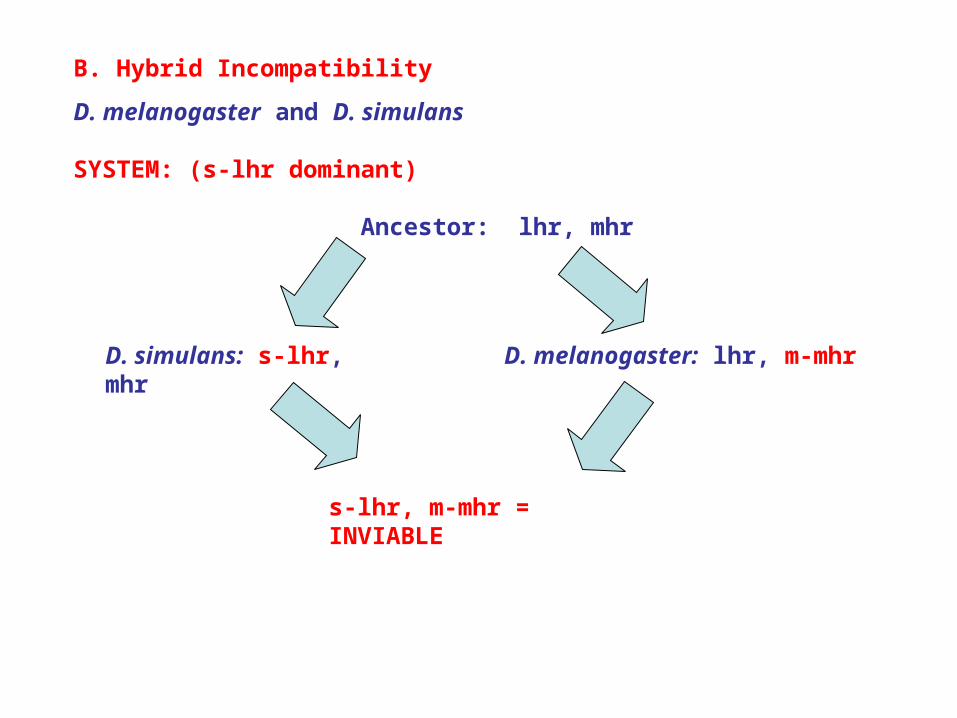
B. Hybrid Incompatibility
D. melanogaster and D. simulans
SYSTEM: (s-lhr dominant)
Ancestor: lhr, mhr
D. simulans: s-lhr, mhr D. melanogaster: lhr, m-mhr
s-lhr, m-mhr = INVIABLE

B. Hybrid Incompatibility
D. melanogaster and D. simulans
Brideau et al. 2006. Science 314: 1292-1295
- isolated location of lhr gene. - put NORMAL D. simulans gene into D. melanogaster. - mated these D. melanogaster with Watanabe's mutant strain of D. simulans.
- IF these two genes are partners, then 3/4 hybrids should die.
(w) D. sim = lhr/s-lhr, hmr (X) x (b)D. mel = s-lhr/lhr, m-hmr (X) (doesn't die....)
1/4 SONS GET : lhr/lhr, m-hmr (only X) .... VIABLE
3/4 get some other combination including s-lhr and m-hmr.. INVIABLE

Speciation
I. Modes
II. Mechanisms
A. Progressive Genomic Incompatibility
B. Hybrid Incompatibility
C. Differential Selection

C. Differential Selection - Assumed to be primary, but few studies documenting that reproductive isolation of phenotypes correlates with fitness differential in different environments.
Rundle et al. (2000). Science 287:306.
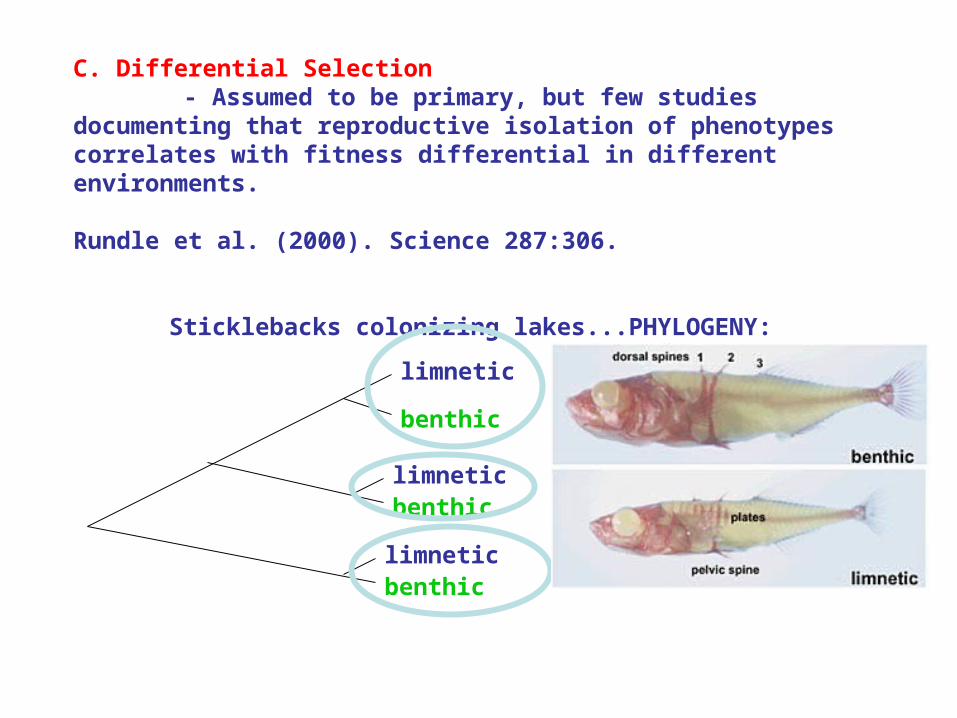
C. Differential Selection - Assumed to be primary, but few studies documenting that reproductive isolation of phenotypes correlates with fitness differential in different environments.
Rundle et al. (2000). Science 287:306.
Sticklebacks colonizing lakes...PHYLOGENY:
benthic
benthic
benthic
limnetic
limnetic
limnetic
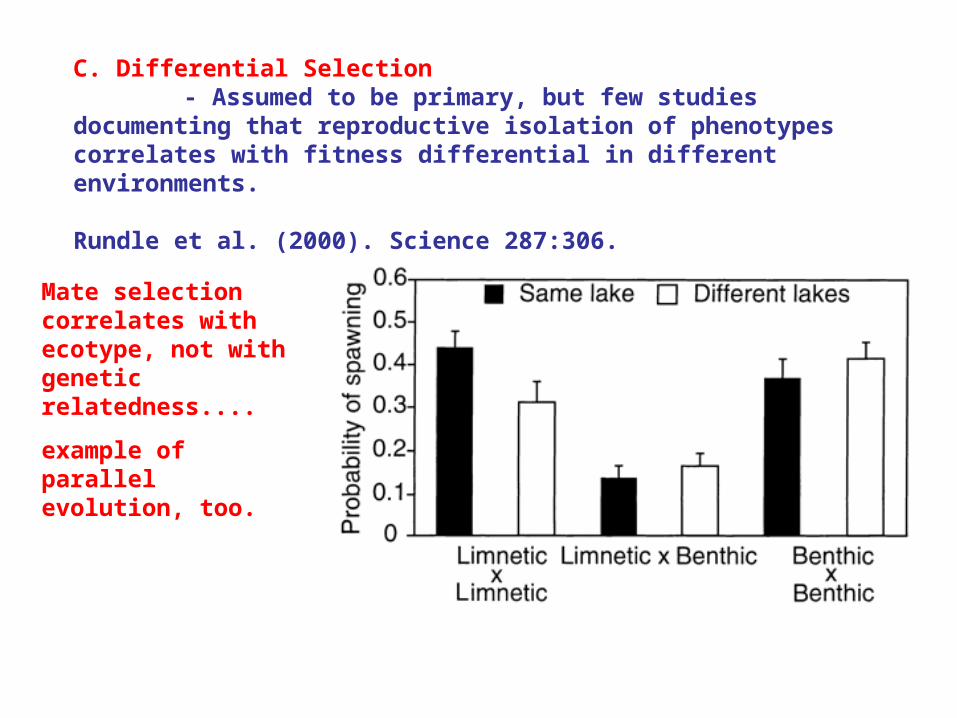
C. Differential Selection - Assumed to be primary, but few studies documenting that reproductive isolation of phenotypes correlates with fitness differential in different environments.
Rundle et al. (2000). Science 287:306.
Mate selection correlates with ecotype, not with genetic relatedness....
example of parallel evolution, too.

Speciation
I. Modes
II. Mechanisms
A. Progressive Genomic Incompatibility
B. Hybrid Incompatibility
C. Differential Selection
D. Hybridization

D. Hybridization - When hybridization occurs, it show increase gene flow between populations. How are hybrids stabilized as a reproductively isolated group?

- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
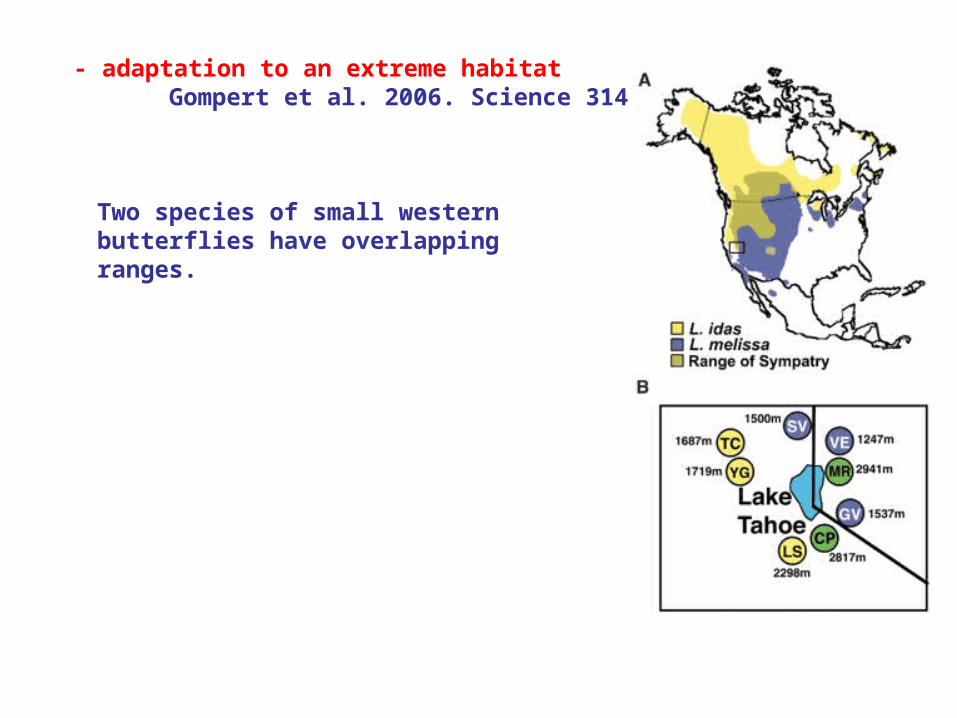
- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
Two species of small western butterflies have overlapping ranges.
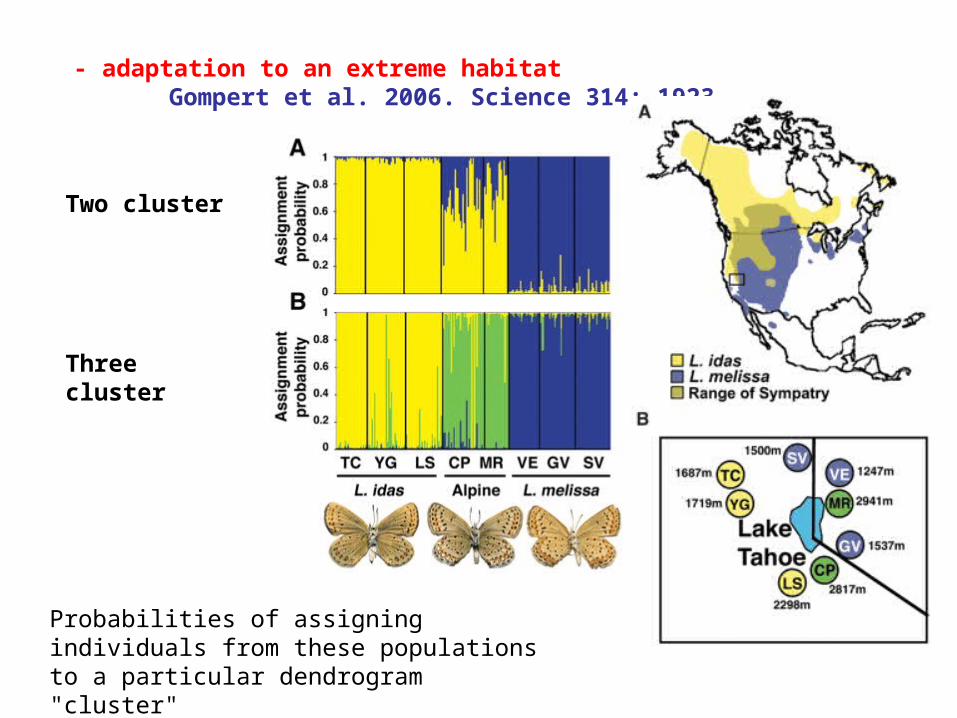
- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
Two cluster
Three cluster
Probabilities of assigning individuals from these populations to a particular dendrogram "cluster"
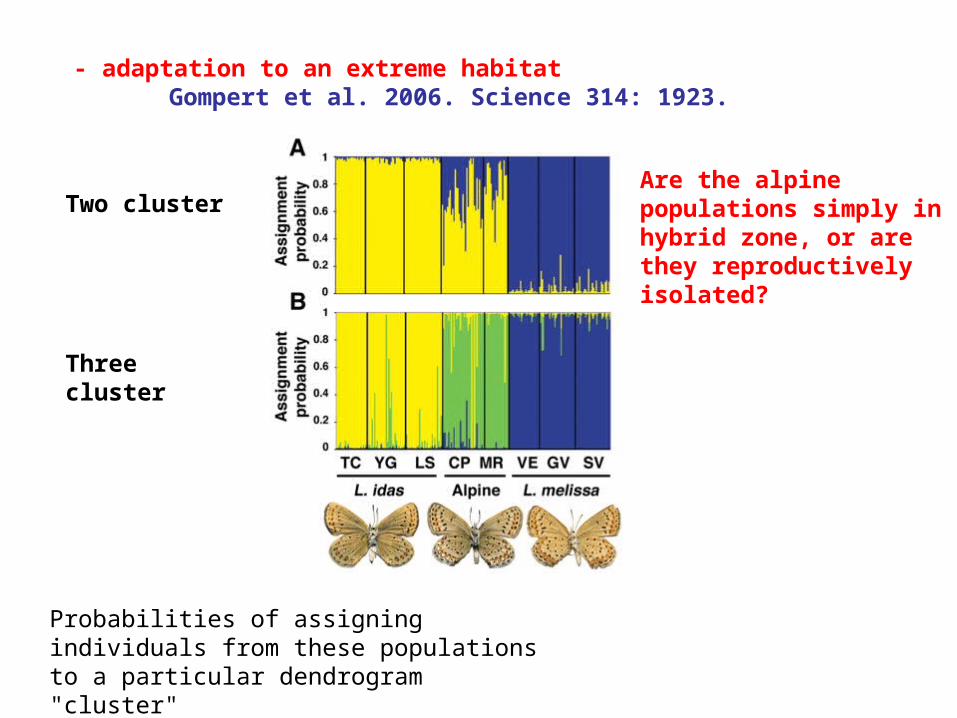
- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
Two cluster
Three cluster
Probabilities of assigning individuals from these populations to a particular dendrogram "cluster"
Are the alpine populations simply in hybrid zone, or are they reproductively isolated?
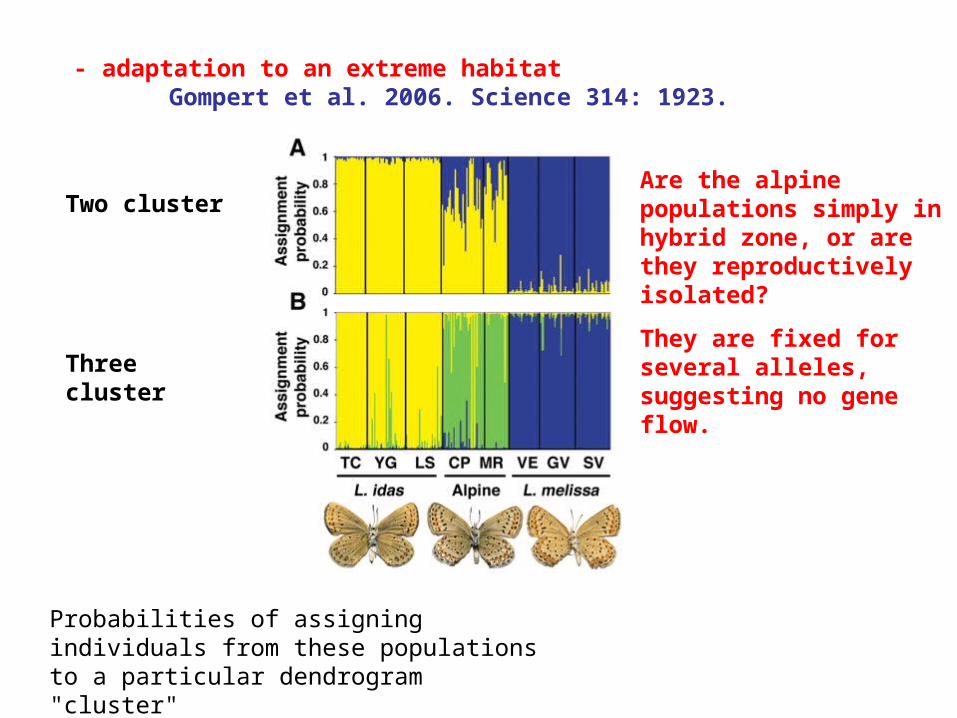
- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
Two cluster
Three cluster
Probabilities of assigning individuals from these populations to a particular dendrogram "cluster"
Are the alpine populations simply in hybrid zone, or are they reproductively isolated?
They are fixed for several alleles, suggesting no gene flow.
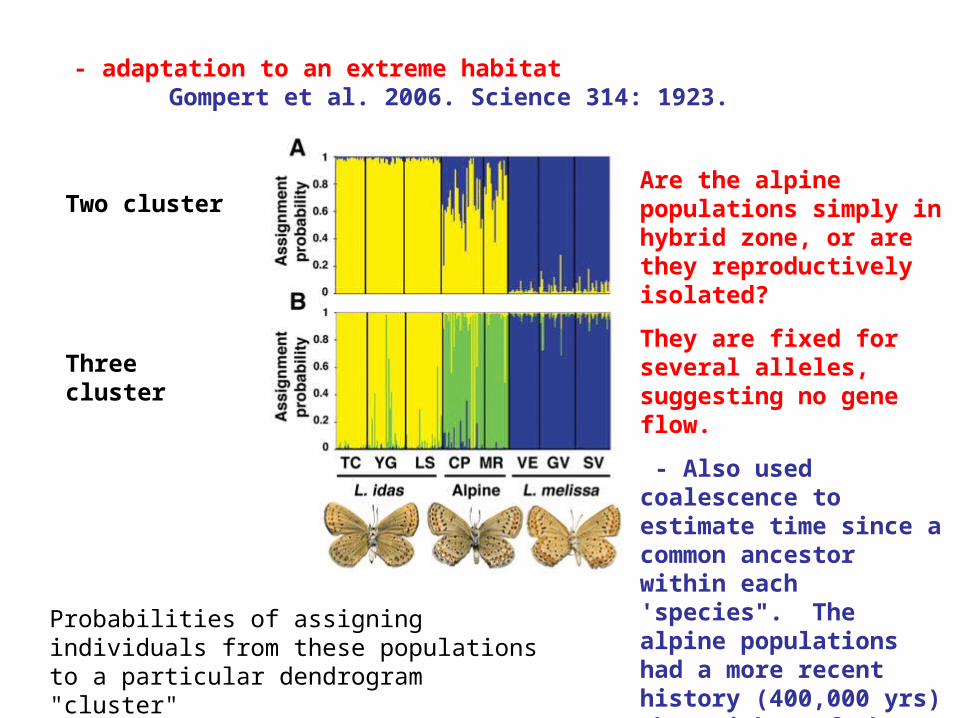
- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
Two cluster
Three cluster
Probabilities of assigning individuals from these populations to a particular dendrogram "cluster"
Are the alpine populations simply in hybrid zone, or are they reproductively isolated?
They are fixed for several alleles, suggesting no gene flow.
- Also used coalescence to estimate time since a common ancestor within each 'species". The alpine populations had a more recent history (400,000 yrs) than either of the others (1.2-1.9 my)

- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
- What maintains this genetic uniqueness?
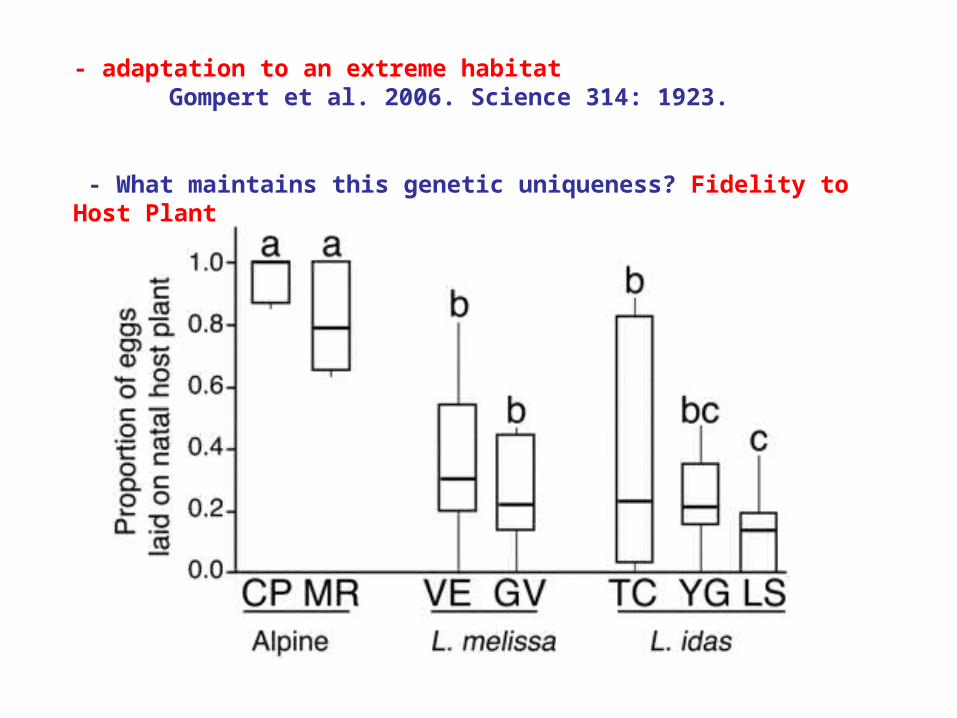
- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
- What maintains this genetic uniqueness? Fidelity to Host Plant

- adaptation to an extreme habitatGompert et al. 2006. Science 314: 1923.
- What maintains this genetic uniqueness?
Fidelity to Host Plant
Also, their eggs don't stick to the leaf; they drop off into litter.
This may be adaptive, as winds blow leaves a long way from original plant at high elevations. The host plant is a perennial, so dropping into the leaf litter keeps it close to host plant.
Other species, even if they used the plant, would have eggs dispersed from the host plant. That's bad for butterflies, 'cuz caterpillars don't disperse too far...

D. Hybridization - When hybridization occurs, it show increase gene flow between populations. How are hybrids stabilized as a reproductively isolated group?
- adaptation to extreme habitat - sexual selection
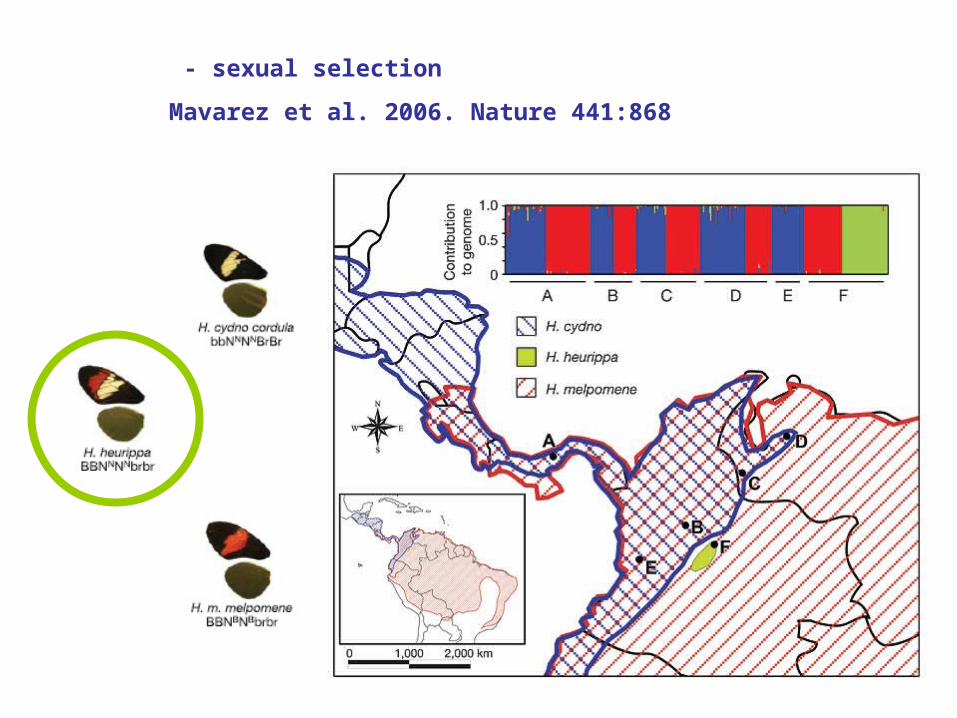
- sexual selection
Mavarez et al. 2006. Nature 441:868
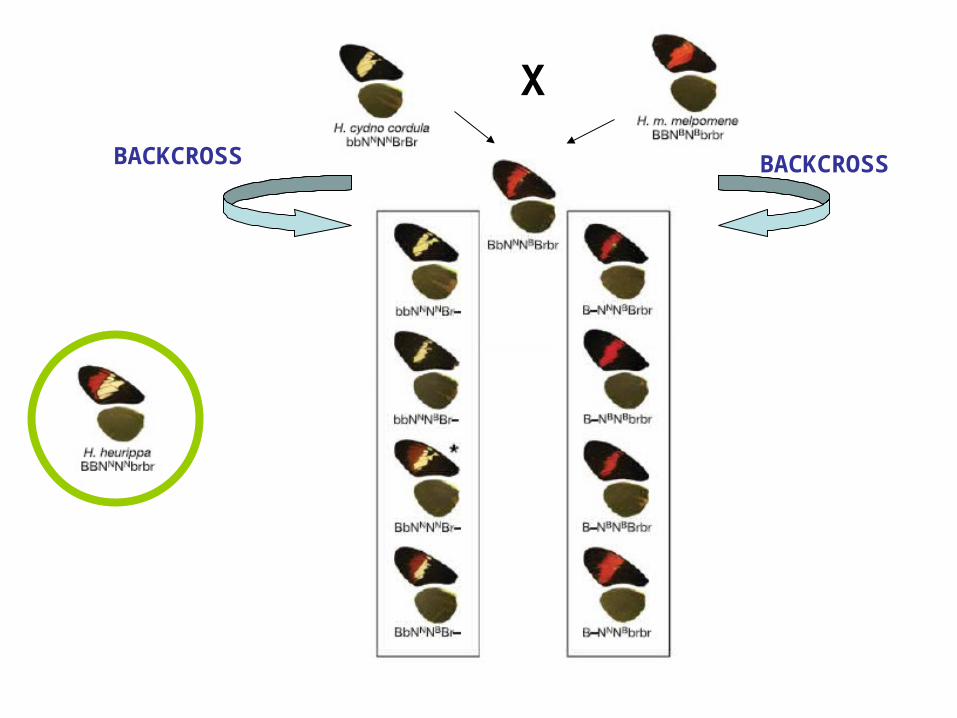
X
BACKCROSS BACKCROSS
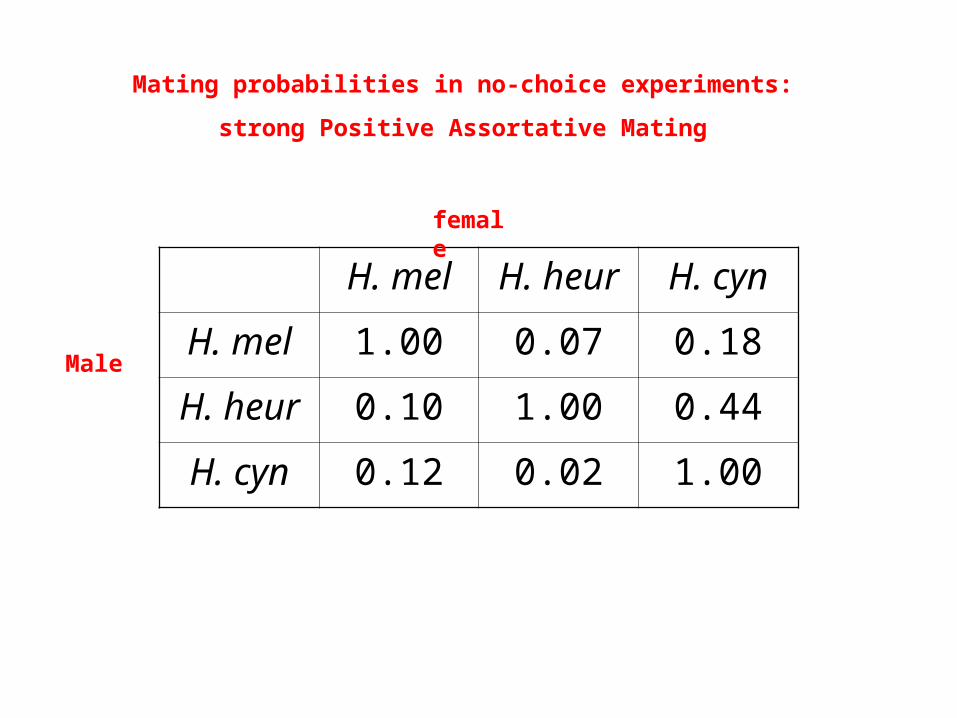
H. mel H. heur H. cyn
H. mel 1.00 0.07 0.18
H. heur 0.10 1.00 0.44
H. cyn 0.12 0.02 1.00
Mating probabilities in no-choice experiments:
strong Positive Assortative Mating
Male
female

Mate Pairing in Tetrads:
strong Positive Assortative Mating

Speciation
I. Modes
II. Mechanisms
A. Progressive Genomic Incompatibility
B. Hybrid Incompatibility
C. Differential Selection
D. Hybridization
Several ways that new gene combinations can form and become stabilized.
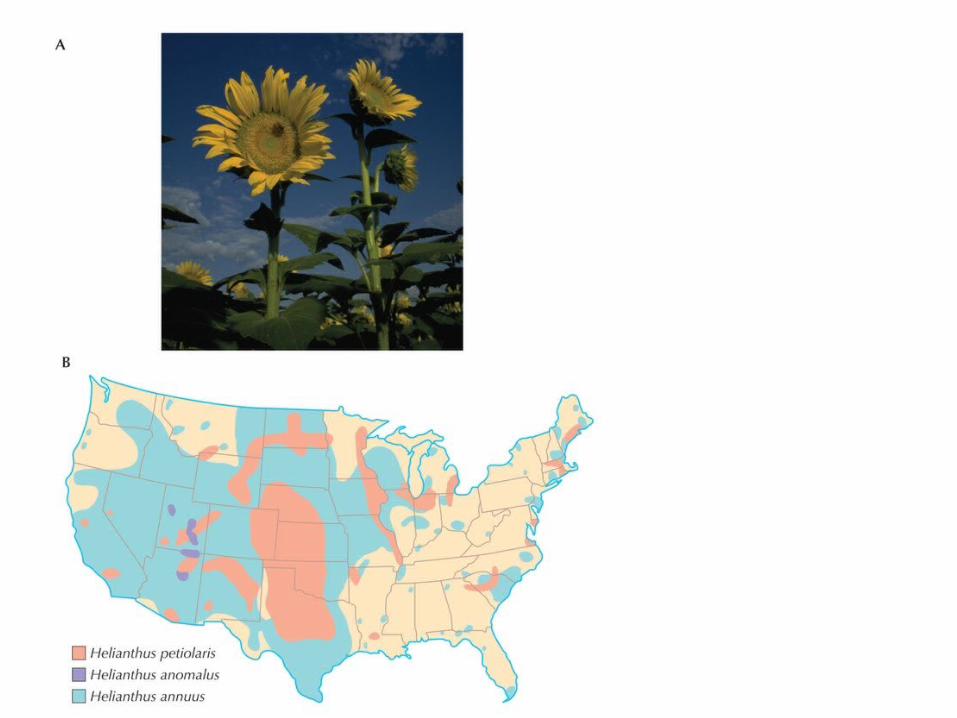
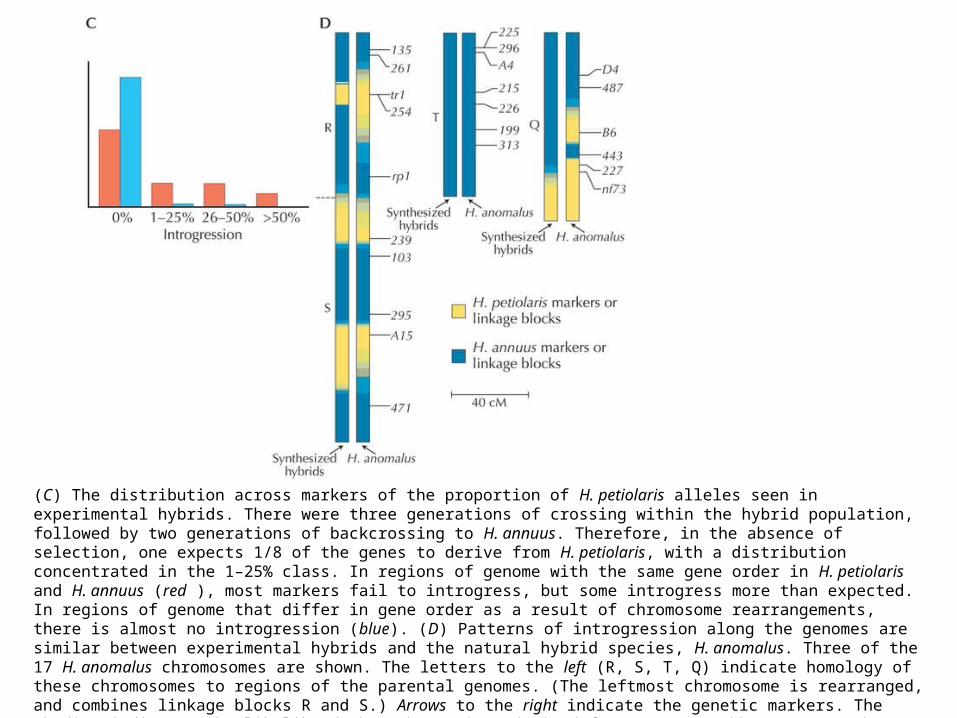
(C) The distribution across markers of the proportion of H. petiolaris alleles seen in experimental hybrids. There were three generations of crossing within the hybrid population, followed by two generations of backcrossing to H. annuus. Therefore, in the absence of selection, one expects 1/8 of the genes to derive from H. petiolaris, with a distribution concentrated in the 1–25% class. In regions of genome with the same gene order in H. petiolaris and H. annuus (red ), most markers fail to introgress, but some introgress more than expected. In regions of genome that differ in gene order as a result of chromosome rearrangements, there is almost no introgression (blue). (D) Patterns of introgression along the genomes are similar between experimental hybrids and the natural hybrid species, H. anomalus. Three of the 17 H. anomalus chromosomes are shown. The letters to the left (R, S, T, Q) indicate homology of these chromosomes to regions of the parental genomes. (The leftmost chromosome is rearranged, and combines linkage blocks R and S.) Arrows to the right indicate the genetic markers. The shading indicates the likelihood that the regions derived from H. annuus (blue) or H. petiolaris (yellow). (A, Courtesy USDA; B, redrawn from Rogers et al. 1982; C, data from Table 1 in Rieseberg et al. 1995a; D, redrawn from Fig. 3 in Rieseberg and Noyes 1998.)





























