Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Species-Level Phylogenetic Analysis of Pterocephaliids (Trilobita, Cambrian) fromthe Great Basin, Western USAAuthor(s): Melanie J. HopkinsSource: Journal of Paleontology, 85(6):1128-1153. 2011.Published By: The Paleontological SocietyDOI: http://dx.doi.org/10.1666/11-002.1URL: http://www.bioone.org/doi/full/10.1666/11-002.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

SPECIES-LEVEL PHYLOGENETIC ANALYSIS OF PTEROCEPHALIIDS(TRILOBITA, CAMBRIAN) FROM THE GREAT BASIN, WESTERN USA
MELANIE J. HOPKINSDepartment of the Geophysical Sciences, University of Chicago, 5734 S. Ellis Ave, Chicago, IL 60637, USA; current address: Department of Geology,
Field Museum of Natural History, 1400 S. Lake Shore Drive, Chicago, Illinois 60605, USA, ,[email protected].
ABSTRACT—Considerable systematic work devoted to late Cambrian trilobites includes very little species-levelphylogenetic analysis. This paper presents the phylogenetic analysis of 36 species representing eight genera assignedto the Family Pterocephaliidae that occur in the Great Basin of the western United States during the Steptoean stage(Furongian). Continuous characters are treated in four different ways: discretization using finite mixture coding,discretization using gap-weighting, ‘‘as such’’ using ranges of values as implemented in the phylogeny program TNT,and exclusion altogether. Results indicate that even the inclusion of only a few continuous characters dramaticallyincreases the resolution of nodes. Despite the different treatments of continuous characters, major features of thetrees are shared across all results. The subfamily Pterocephaliinae is restricted to genera which possess a concaveanterior border. Relative stratigraphic placement was estimated using a composite section built in CONOP and usedto scale the tree topologies and to assess stratigraphic consistency. Although previously hypothesized multispeciesevolutionary series are not supported by the results, tree topology, stratigraphic distribution, and optimizedcharacter state transformation support the interpretation of several sister taxa as direct ancestor-descendent pairs.
INTRODUCTION
LATE CAMBRIAN Laurentian trilobites have arguably one ofthe best-known fossil records of any fossil group at any
time (Adrain and Westrop, 2001). A considerable body ofwork has been devoted to late Cambrian trilobite systematics(e.g., Lochman, 1938, 1940; Lochman and Duncan, 1944;Lochman, 1950; Palmer, 1954, 1955; Lochman and Hu, 1960;Palmer, 1960; Lochman and Hu, 1961; Grant, 1962; Lochmanand Hu, 1962a, 1962b; Palmer, 1962; Lochman, 1964; Palmer,1965b, 1968; Stitt, 1971a; Kurtz, 1975; Stitt, 1977; Westrop,1986; Pratt, 1992; Stitt et al., 1994; Stitt, 1998; Stitt andPerfetta, 2000; Adrain and Westrop, 2005, 2006; Westrop andAdrain, 2007; Westrop et al., 2008; Adrain et al., 2009) and theexploration of patterns of diversification, ecological distribu-tion, and extinction of these clades (e.g., Palmer, 1965a;Lochman-Balk, 1970; Stitt, 1971b, 1975; Palmer, 1979, 1982;Hardy, 1985; Westrop and Ludvigsen, 1987; Westrop, 1988,1989, 1996) but relatively little phylogenetic analysis has beendone, particularly at the species-level, until recently (seeHughes and Rushton, 1990; Westrop and Ludvigsen, 2000;Adrain and Westrop, 2001; Westrop and Adrain, 2009;Westrop et al., 2010). Nonetheless, a phylogenetic frameworkmay be crucial for understanding the evolutionary patternspreserved in the fossil record, particularly in the lateCambrian, when trilobite evolution is characterized byrepeated intervals of diversification and extinction (formerlyreferred to as ‘‘biomeres’’, Palmer, 1965a).
The Pterocephaliidae characterize the Steptoean Stage(Furongian) of Laurentia, which is preserved in its entiretyonly in the Great Basin of the western U.S.A. (e.g., Palmer,1965b) and in some offshore sections in northern Canada(Pratt, 1992) due to a mid-Stage continent-wide regression(Sauk II-Sauk III, Palmer, 1981; Osleger and Read, 1993). Thepurpose of this study is to generate a species-level phylogenyfor genera of the family Pterocephaliidae Kobayashi, 1935,specifically genera from the subfamilies PterocephaliinaeKobayashi, 1935 and Housiinae Hupe, 1953, that may beused to assess previously described evolutionary relationships(this study) and provide a framework for ongoing macroevo-lutionary studies (e.g., Hopkins, in press) while exploring the
effects of coding methods for continuous characters. This isthe first species-level phylogenetic analysis of this clade, andfor the sake of feasibility, the number of included taxa wasnecessarily limited and generally accepts current speciesdesignations. Nonetheless, results provide support for severalpreviously established clades and the examination of existingmaterial as well as new material from field collections formsthe basis for expanded descriptions of several taxa, primarilyspecies of Parahousia Palmer, 1960, Prehousia Palmer, 1960,Sigmocheilus Palmer, 1960, and Strigambitus Palmer, 1965b.
MATERIAL AND METHODS
Taxon selection.—The Pterocephaliidae is currently com-prised of 47 genera (Jell and Adrain, 2003). Two of these 47genera have been assigned to the Chinese subfamily Cilinae(Peng, 1992), four to the subfamily Housiinae (Palmer, 1965b),and seven to the subfamily Pterocephaliinae (Palmer, 1965b;Kurtz, 1975; Hohensee and Stitt, 1989). The remaining 34genera in Jell and Adrain’s classification are not currentlyassigned to any of the named subfamilies. Of these taxa,Anechocephalus Palmer, 1960, is questionably assigned to thefamily, Plesiocilia Yuan and Yin, 1998, has since beensynonymized with Adelogenus (Family Proasaphiscidae, Penget al., 2004), Pterocephalopsinus Jell in Jell and Adrain, 2003,has since been synonymized with Baikadamaspis (FamilyHarpetidae? Peng et al., 2004), and Pterocephalina Resser,1938, was previously synonymized with Litocephalus Resser,1937, (Aphelaspididae) by Palmer (1956). Ventral morphologyhas not yet been described for most of these taxa.
All described species assigned to the subfamily Housiinae(Prehousia, Housia Walcott, 1916, Tumicephalus Palmer, 1960,and Parahousia) were included in the analysis (Table 1, 12species). Housia canadensis (Walcott, 1912) is herein consid-ered a junior synonym of Housia varro (Walcott, 1916),following Hohensee and Stitt (1989). Housia halli (Resser,1935), known only from one pygidium indistinguishable fromH. varro, was questionably assigned to the latter species byPalmer (1965b), and this assignment is followed here. Palmer(1965b) considered Aphelaspis tumifrons Resser, 1938, fromTennessee to be congeneric with Tumicephalus depressus Palmer,
Journal of Paleontology, 85(6), 2011, p. 1128–1153
Copyright ’ 2011, The Paleontological Society
0022-3360/11/0085-1128$03.00
1128

1965b, but this taxonomy does not appear to have been adoptedby subsequent workers (e.g., Lochman-Balk, 1974) and is notfollowed here.
Almost all species of four pterocephaliine genera withoccurrences in the Great Basin (Cernuolimbus Palmer, 1960,Strigambitus, Sigmocheilus, and Pterocephalia Roemer, 1849)were included in the analysis (Table 1, 24 species). Wherepossible, species coding was based on inspection of specimensheld at the Smithsonian, U.S. Geological Survey, the Institutefor Cambrian Studies (University of Chicago) and from fieldcollections. Coding for four species was based solely onpublished descriptions and figures (Cernuolimbus monilis Stittet al., 1994, Pterocephalia norfordi Chatterton and Ludvigsen,1998, Cernuolimbus pegakanthodes Pratt, 1992, Cernuolimbuslongifrons Palmer, 1968). Two species were excluded becausefossil material and published descriptions were either unavail-able or inadequate (Pterocephalia brevifrons Kobayashi, 1938,Sigmocheilus aldanensis Gogin, 1990). Three species question-ably assigned to genera were also excluded (Strigambitus?blepharina Palmer, 1965b, Pterocephalia? punctata Palmer,1965b, Sigmocheilus? compressus Palmer, 1968).
Three additional genera assigned (or questionably assigned)to the Pterocephaliinae include Camaraspis Ulrich and Resser,in Ulrich, 1924, Pulchricapitus Kurtz, 1975, and PelicephalusKurtz, 1975. All occur in Laurentia but outside the GreatBasin and first appear in the uppermost Steptoean (Elvinia
biozone). All differ from other pterocephaliine genera bylacking a concave border but show affinities to some housiinegenera as well as other families (e.g., Elvinidae, Westrop,1986). Because the focus of this study is trilobites of the GreatBasin, all of which belong to older genera, these taxa were notincluded in this analysis. Remaining genera currently assignedto this family are largely from China and to a lesser extentfrom Siberia and Kazakhstan. Neither fossil material norpublished descriptions were available for many of these taxa,and were thus not included. Some descriptive comparison ofthese taxa to those included in the analysis is given in theDiscussion.
Outgroup selection.—Because of their morphological simi-larity to some of the older housiine taxa, their lowerstratigraphic position, and the availability of large samplesizes for study, three species of Dicanthopyge Palmer, 1965b,and three Great Basin species of Aphelaspis Resser, 1935, wereselected as outgroup taxa to the pterocephaliids (Table 1).These species belong to the family Aphelaspididae Palmer,1960, previously assigned subfamilial status among thepterocephaliids (Palmer, 1965b). Each species occurs in thelowermost biozones of the stage (Aphelaspis and Dicantho-pyge) and occur below all other taxa in the analysis with thepossible exceptions of ‘‘Prehousia’’ semicircularis (Palmer,1960), and Cernuolimbus semigranulosus Palmer, 1960, both ofwhich have very poorly constrained first appearances (seeresults). Species of pterocephaliid genera that may have beencandidate outgroup taxa (e.g., Camarapsis, Pulcricapitus,Pelicephalus, Morosa Palmer, 1960, spp.) are all younger thanthe subset of taxa analyzed in this study, occurring in theDunderbergia to Elvinia biozones as well as the overlyingSunwaptan Stage (Palmer, 1965b; Kurtz, 1975; Westrop,1986).
Aphelaspis haguei (Hall and Whitfield, 1877) and Aphelaspissubditus Palmer, 1962, were chosen as outgroup taxa over twoslightly older Aphelaspis species (Aphelaspis buttsi Palmer,1962, and Aphelaspis brachyphasis [Kobayashi, 1936]) becausethey are more widespread in their geographic occurrence andbetter sampled.
Increased sampling in Aphelaspis subditus and A. haguei hasled to systematic uncertainty, however. Palmer (1965b)reported that a narrower cranidial border, less convexpreglabellar field, and lack of pitting could be used todistinguish Aphelaspis subditus from Aphelaspis haguei. Pratt(1992) added that A. haguei also has a prominent occipitalnode, longer palpebral lobes, and a narrower pygidial border.Some of these characteristics are not consistently present (e.g.,some specimens with a narrow anterior margin also havepitting) and that others are highly variable (e.g., the extent ofconvexity of preglabellar field, the prominence of occipitalnode, and the ratio between the sagittal length of the anteriorborder and preglabellar field). As a result, cranidia of thesetwo species are difficult to tell apart, particularly where theyare thought to co-occur. For this study, specimens wereseparated into two groups, one representing ‘‘Aphelaspissubditus’’ and one representing ‘‘Aphelaspis haguei,’’ basedprimarily on the shape and size of the anterior border. Thesetwo species code almost identically with the exception ofdifferences in sample means that result in close but notidentical coding using gap-weighting (described below).Synonymizing these two species did not change the topologyof the consensus trees for any of the character datasets(described below). Similarly, A. brachyphasis and A. buttsi areso similar to these two species, particularly relative to ingrouptaxa, that their substitution as outgroup taxa is not expected
TABLE 1—Species included in the phylogenetic analysis. # indicatesoutgroup species; * indicates species coded entirely from literature.Numbers refer to character matrix in Appendix 2.
No. Species name
1 .Aphelaspis haguei (Hall and Whitfield, 1877)#2 .Aphelaspis longispina Palmer, 1965b#3 .Aphelaspis subditus Palmer, 1962#4 .Cernuolimbus arcticus Palmer, 19685 .Cernuolimbus depressus Palmer, 19606 .Cernuolimbus granulosus Palmer, 1965b7 .Cernuolimbus laevifrons Palmer, 1965b8 .Cernuolimbus longifrons Palmer, 1968*9 .Cernuolimbus monilis Stitt et al., 1994*
10 .Cernuolimbus orygmatos Palmer, 196011 .Cernuolimbus pegakanthodes Pratt, 1992*12 .Cernuolimbus semigranulosus Palmer, 196013 .Dicanthopyge convergens Palmer, 1965b#14 .Dicanthopyge quadrata Palmer, 1965b#15 .Dicanthopyge reductus Palmer, 1965b#16 .Housia ovata Palmer, 196017 .Housia vacuna (Walcott, 1890)18 .Housia varro (Walcott, 1916)19 .Parahousia constricta Palmer, 196020 .Parahousia subequalis Palmer, 1965b21 .Prehousia alata Palmer, 196022 .Prehousia diverta Palmer, 1965b23 .Prehousia impolita Palmer, 1965b24 .Prehousia indenta Palmer, 1965b25 .Prehousia prima Palmer, 1965b26 .‘‘Prehousia’’ semicircularis (Palmer, 1960)27 .Pterocephalia concava Palmer, 195428 .Pterocephalia constricta Palmer, 196829 .Pterocephalia elongata Palmer, 196030 .Pterocephalia norfordi Chatterton and Ludvigsen, 1998*31 .Pterocephalia occidens (Walcott, 1884)32 .Pterocephalia sanctisabae Roemer, 184933 .Pterocephalia tenura Rozova, 196334 .Sigmocheilus delicatus Lazarenko, 196835 .Sigmocheilus flabellifer (Hall and Whitfield, 1877)36 .Sigmocheilus grata (Resser, 1942)37 .Sigmocheilus notha (Resser, 1942)38 .Sigmocheilus pogonipensis (Resser, 1942)39 .Strigambitus bilobus Palmer, 1965b40 .Strigambitus transversus Palmer, 1965b41 .Strigambitus utahensis (Resser, 1942)42 .Tumicephalus depressus Palmer, 1965b
HOPKINS—PTEROCEPHALIID PHYLOGENY 1129

to affect either character polarity or the resulting treetopologies.
Qualitative character selection and coding.—Forty-fourqualitative characters describing the cranidia, librigena, andpygidia were selected (Appendix 1). Some characters requirespecific notes about their coding and are listed in the order inwhich they appear in Appendix 1:
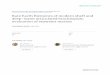
1. Concavity of the border: The subfamily Pterocephalii-nae is defined by the concave shape of the cranidialborder in lateral view (e.g., Fig. 1.2). Species of thesubfamily Housiinae and the family Aphelaspididaeshow either a flat or slightly convex cranidial border(Fig. 1.4).
2. Expression of the anterior border furrow: This trait isoften shared at the genus level. For example, Pteroce-phalia species have a shallow, poorly defined furrow.Sigmocheilus species have a shallow but well definedcranidial border, and Cernuolimbus species have a welldefined and deep cranidial border.
6. Ornamentation along border: Among the species in thisstudy, this character is only present in Cernuolimbus(Fig. 1.5), where the ornamentation is either a row ofcoarse granules (Cernuolimbus depressus Palmer, 1960,and Cernuolimbus granulosus Palmer, 1965b) or a row ofcoarse pits (Cernuolimbus semigranulosus, Cernuolimbusmonilis Stitt et al., 1994, Cernuolimbus longifronsPalmer, 1968, Cernuolimbus arcticus Palmer, 1968). Thischaracter is not unique to this genus across Steptoeantrilobites and is also present in species of LitocephalusResser, 1937 (family Aphelaspididae).
8, 9. Cranidial ornamentation: Whereas caecal markings areconsidered a type of ornamentation, some species showit in addition to another type (e.g., Dicanthopyge speciesshow both pitting and caecal marking on the internalmold). Because these two types of ornamentation maybe expressed on the same specimen, they are notpolymorphisms. In order to make this distinction interms of coding, an additional character describing theabsence or presence of caecal marking was created.
Although Palmer (1965b) describes ‘‘Prehousia’’ semicircu-laris as lacking ornamentation on all sclerites, at least onespecimen shows faint caecal markings on the frontal area(Fig. 1.10).
13. Expression of glabellar furrows: Pterocephalia concavaPalmer, 1954, is distinguished from Pterocephaliasanctisabae Roemer, 1849, by having less conspicuousglabellar furrows (Palmer, 1965b). However, becausethey are still more strongly expressed in this species thanin other genera (such as the housiines), both species arecoded as ‘‘strongly expressed.’’
19. Presence of ocular ridge: The presence of an ocular ridgeis dependent on whether the palpebral lobe reaches theaxial furrow (character 14). For species where thepalpebral lobe abuts the glabella in this fashion (e.g.,Housia), there cannot be an ocular ridge, and characteris coded as ‘‘inapplicable.’’
20. Direction of ocular ridges: For species where the ocularridge is not expressed on the dorsal surface, thedirection of the ocular ridges can usually be determinedby inspecting the internal mold. This character must becoded as ‘‘inapplicable’’ for species without an ocularridge (i.e., the anterior end of the palpebral lobe is incontact with the axial furrow).
21. Course of facial suture in front of glabella: InSigmocheilus and Pterocephalia, the anterior facialsuture is intramarginal on the dorsal surface until closeto the axial line, where it crosses the anterior marginand runs posteriorly on the ventral surface until it meetsthe ventral median suture. The submarginal facialsuture can be observed directly on the ventral side ofthe cranidium where the anterior margin curvesventrally (Fig. 1.6–1.9) or indirectly through compari-son of the shape of the facial suture on the librigena anddorsal surface of the cranidium. If some portion of thefacial suture is submarginal, this species was coded as‘‘1’’; if no part of the facial suture is known to besubmarginal (it meets the rostral suture or ventralmedian suture at the anterior margin), it was consideredto be ‘‘intramarginal only.’’
27. Ornamentation on genal spine: This character wascoded as ‘‘inapplicable’’ for species that lost the genalspine during development (some Housia species).
28. Ventral median suture: The ventral median suture ishistorically considered to be a symplesiomorphy of thefamily Pterocephaliidae (Fortey and Chatterton, 1988)and a key synapomorphy of the Asaphida (Fortey andChatterton, 1988; Fortey, 1997). Herein the ventralmedian suture is coded as present if any portion of thelibrigenal doublures meet medially. Among the speciesin this analysis, either a ventral median suture has beenobserved or the morphology of the librigenal doublureis consistent with a ventral median suture for thefollowing: Sigmocheilus grata (Resser, 1942) (Fig. 2.3),Sigmocheilus notha (Resser, 1942) (Fig. 2.1), Cernuo-limbus longifrons (Palmer, 1968, pl. 8, fig. 13), Cernuo-limbus semigranulosus (not figured), Housia vacuna(Walcott, 1890) (e.g., Westrop, 1986, pl. 26, fig. 8),Housia varro (e.g., Lochman, 1964, pl. 12, fig. 27),Pterocephalia norfordi (Chatterton and Ludvigsen,1998, fig. 28.1), Pterocephalia concava (not figured),Pterocephalia elongata Palmer, 1960 (Palmer, 1960, pl.9, fig. 20), Pterocephalia sanctisabae (Fig. 2.4), Strigam-bitus bilobus Palmer, 1965b, (Fig. 2.6), and Strigambitusutahensis (Resser, 1942) (not figured). Palmer (1960,p. 74) stated that a rostral plate was present in oldergenera of Housiinae and illustrated it diagrammaticallyfor Prehousia (Palmer, 1960, p. 64), but the morphologyof the librigena in comparison with the cranidium inPrehousia alata Palmer, 1960, is more consistent with aventral median suture (Fig. 2.2, see also SystematicPaleontology). Thus, aside from Parahousia, at least onemember of each pterocephaliid genus included in thisanalysis is known to have a ventral median suture.Because the ventral median suture may change duringdevelopment (e.g., Park and Choi, 2009) and may haveevolved independently in different trilobite clades (e.g.,Ludvigsen, 1991; Whittington, 2003; Zhu et al., 2007;Adrain et al., 2009), no assumption is made that allpterocephaliid species in the analysis had ventralmedian sutures and for many taxa, this character iscoded as ‘‘?’’. Where known, the expression of theventral median suture is straight, except in Pterocephalianorfordi, where it is zigzagged (Chatterton and Ludvig-sen, 1998).
Species that do not have a ventral median suture have areduced rostral plate (Aphelaspis, Dicanthopyge). At least onespecies with a ventral median suture (Pterocephalia norfordi)
1130 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

FIGURE 1—Representative specimens showing some cranidial character states mentioned in text: 1, 2, Sigmocheilus notha (Resser, 1942), cranidium,dorsal and right lateral views, USNM 543767, 310; 3, 4, Dicanthopyge quadrata Palmer, 1965b, cranidium, dorsal and right lateral views, USNM543765, 38. Arrows in 2 and 4 indicate the location of the anterior border furrow; 5, Cernuolimbus arcticus Palmer, 1968, internal mold of cranidiumshowing pitting in anterior border furrow, USNM 146800, 35; 6–9 Sigmocheilus notha: 6, 7, anterior and ventral views of anterior margin of cranidiumshowing submarginal suture, USNM 543768, 315; 8, view of anterior margin of cranidium showing submarginal suture, USNM 543767, 325; 9, ventralview of anterior margin of cranidium showing submarginal suture, USNM 543770, 325. Arrows in 6–9 indicate submarginal suture. Specimens in 7–9are oriented with posterior end of palpebral lobes closer to viewer than anterior end in order to better expose the margin (i.e., inconsistent with standardorientation recommended by Shaw [1957]); 10, ‘‘Prehousia’’ semicircularis Palmer, 1960, close-up of right anterior portion of cranidium showing faintcaecal marking, USNM 136868, 325; 1, 2, 6–9, all from the McGill section, Duck Creek Range, NV; 3, 4 from the Schell Creek Range, NV; 5, from theHillard Peak area, AK; 10, from Windfall Canyon, Eureka district, NV.
HOPKINS—PTEROCEPHALIID PHYLOGENY 1131

also has a rostellum, though it is unclear from the figuredspecimens of this species in Chatterton et al. (1994) andChatterton and Ludvigsen (1998) whether the triangular pieceof exoskeleton on the ventral side is separated from the anteriormargin of the cranidium by a true suture or if it is actually partof the cranidium as seen in Sigmocheilus notha (Fig. 1.6–1.9).This possibility is worth noting because the rostellum inPterocephalia norfordi has been argued to be a putativemorphological link between earlier trilobite species withreduced rostral plates (e.g., the ptychopariids) and later trilobitespecies with only a ventral suture (e.g., trinucleoids) (Chattertonet al., 1994). A possible rostellum has only been documented inPt. norfordi; thus, for the purposes of this study, the absence orpresence of this character was not included in the analysis (i.e.,it was effectively treated as an apomorphy).
29. Presence of ankylosed thoracic segment: Housia pygidiaoften but not always possess an ankylosed thoracic
segment. In these specimens, the thoracic segment isseparated from the pygidium by a suture line butremains articulated. In fact, because the thoracicsegments are spinose, Housia specimens were originallyplaced in the genus Ceratopyge, which has true pygidialspines (Walcott, 1924). Because not all specimens of thesame species possess (or preserve) an ankylosed thoracicsegment, they are coded as ‘‘commonly present’’.
41. Shape of posterior border: In Dicanthopyge species, theabsence or presence of a posterior median inbend on thepygidium is obscured by the posterior spines. However,the border appears to be smoothly curved between thespines (Fig. 2.5) and is coded as lacking no inbend onthe pygidium. Whereas Palmer (1965b) describesStrigambitus transversus Palmer, 1965b, as having asmooth pygidial border, there is a slight inbend onseveral specimens and has thus been coded accordingly.Specimens of St. transversus with an inbend on the
FIGURE 2—Representative specimens showing some librigenal and pygidial character states mentioned in text: 1, Sigmocheilus notha (Resser, 1942),librigena, USNM 543771, McGill section, Duck Creek Range, NV, 325; 2, Prehousia alata Palmer, 1960, partial librigena, USNM 543776, SnakeRange, NV, 35; 3, Sigmocheilus grata (Resser, 1942), librigena, USNM 543772, Windfall Canyon, Eureka district, NV, 33; 4, Pterocephalia sanctisabaeRoemer, 1859, partial librigenal doublure, USNM 543777, Shingle Pass, NV, 35; 5, Dicanthopyge quadrata Palmer, 1965b, pygidium, USNM 543766,Schell Creek Range, NV, 310; 6, Strigambitus bilobus Palmer, 1965b, librigena, USNM 543778, Ruby Range, NV, 38; 7, Housia varro (Walcott, 1916),ventral side of pygidium showing terrace lines on doublure, FMNH PE60959, Goshute Mountains, NV, 35; 8, Sigmocheilus grata, partially exfoliatedpygidium showing terrace lines on ventral side of doublure, USNM 543773, Windfall Canyon, Eureka district, NV, 37.
1132 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

pygidial border are still easily distinguished fromcongeneric species, which have lobes on the pygidialborder defined by deeper inbends (see SystematicPaleontology).
43. Terrace lines on border of pygidium: Where knownfrom exposed ventral surfaces or external molds, allspecies show terrace lines on the ventral side of thepygidium (e.g., Fig. 2.7, 2.8, see also Fig. 11.9). There isvariation, however, in the extent of terrace lines acrossthe dorsal surface. Where no terrace lines are visible onthe dorsal surface, even along the margin, this characterwas coded as ‘‘absent.’’
Qualitative characters not used.—Sclerites other than thecranidium, pygidium, and librigena have not been identifiedfor most species in the analysis. Thus, characters describingthe hypostome, thoracic segments, and eyes were not included.In addition, because articulated material is available for only afew species, the relative size of sclerites could not be coded.
Several characters were not included because they are sharedby all taxa. In particular, all species have a tapering forward andmoderately convex glabella, band-like eye socles where known,a preglabellar field, and strongly defined axial furrows.
Many autapomorphic characters are incorporated into thecharacter matrix via multistate characters, for example thealae on the pygidium of Prehousia alata (character 44, state 2)or the long median inbend on Strigambitus bilobus (character41, state 3). Autapomorphic characters not used in the analysisinclude a median swelling on the preglabellar field (onlyTumicephalus depressus), strongly expressed fossulae on thecranidium (only Parahousia subequalis Palmer, 1965b), pairedgranules on the pygidial axis (only Cernuolimbus orygmatosPalmer, 1960), and a zigzag shaped median suture (onlyPterocephalia norfordi).
Finally, several characters were not used because they co-vary with other characters included in the analysis. Forexample, species that do not have a ventral median suture haverostral plates; therefore a character describing the presence orabsence of a rostral plate is not included. In addition, speciesthat have wide cranidial borders also have wide librigenalborders, so no attempt was made to quantify the latter.Finally, species with a concave cranidial border also have aconcave border on the librigena and the pygidium, so this wasdescribed only for the cranidium.
Continuous character selection and coding.—In addition tothe 44 qualitative characters (Appendix 1), eight additionalcharacters expressing continuous variation were identified.These characters are:
45. The length (sag.) of the anterior border: the length (sag.)of the entire frontal area (G1/F1);
46. The width (tr.) of the glabella at the occipital furrow:the length (sag.) of the glabella (K2/B1);
47. The length of the palpebral lobe (the chord): the length(sag.) of the glabella (C1/B1);
48. The width (tr.) of the glabella at the occipital furrow:the width (tr.) of the cranidium at the anterior end of thepalpebral lobe (K2/J4);
49. The length (sag.) of the cranidium: the width (tr.) of thecranidium at the anterior end of the palpebral lobe (A1/J4);
50. The length (sag.) of the frontal area: the length (sag.) ofthe cranidium (F1/A1);
51. The width (tr.) of the pygidium at its widest point: thelength (sag.) of the pygidium (W/X);
52. The length (sag.) of the pygidial axis: the width (tr.) ofthe pygidial axis at its widest point (Y/X);
where notation follows Shaw (1957) (Fig. 3).Despite debate over how continuous characters should be
coded, they are demonstrably phylogenetically informative(Chappill, 1989; Stevens, 1991; Thiele, 1993; Rae, 1998;Swiderski et al., 1998). There are numerous methods forcoding quantitative data into ‘discrete’ character states forinclusion in phylogenetic analysis, including various types ofgap-coding (Michevich and Johnson, 1976; Colless, 1980;Almeida and Bisby, 1984; Thorpe, 1984; Archie, 1985; Baum,1988; Goldman, 1988; Chappill, 1989; Guerrero et al., 2003),statistical methods (e.g., analysis of variance and multiplerange test [ANOVA-MRT], Guerrero et al., 2003), and modelfitting (e.g., finite mixture coding, Strait et al., 1996).
In this dataset, there is considerable overlap in variationamong the taxa. In addition, the number of specimensavailable for inspection varies widely across species. Thusmeans are obtainable for each taxon, but the range ofvariation within each taxon is much greater for those samplesrepresented by more specimens. This is problematic for anymethods that rely on measures of character ranges, standarddeviations, or confidence intervals to identify character states.However, it is possible to apply character state assignments toindividual specimens using finite mixture coding (FMC);species that contain specimens assigned to more than one
FIGURE 3—Length measurements on cranidia (1) and pygidia (2) usedfor continuous characters. Notation after Shaw (1957). Scale bar51 mm.
HOPKINS—PTEROCEPHALIID PHYLOGENY 1133

character state are then treated as polymorphic. To apply thismethod, different numbers of normal distributions are fit tothe dataset of all specimens from all species in the analysis andthe quality of the fit is judged using the bias-correctedAkaike’s Information Criterion (AICc, Akaike, 1974; Hurvichand Tsai, 1989; Burnham and Anderson 2000). Once thenumber of best-fit distributions has been determined, eachspecimen that falls into the 95% confidence interval for anyone distribution is assigned to the same group. Wherespecimens fall into the confidence interval for more than onenormal distribution or fall outside of the distributions, thespecimen is assigned to a group based on the probabilities thatit belongs to each distribution (Strait et al., 1996; Sheets,2003).
For this analysis, all continuous characters are comprised oflength:length ratios. The ratios were transformed by log(x + 1)as recommended when variances are not equal and in order toavoid negative characteristics in the logarithms (Thiele, 1993;Sokal and Rohlf, 1995; Kitching et al., 1998). Many of theratios measured may change allometrically through ontogenyin trilobite species. All specimens selected were holaspids, i.e.,considered to have reached the terminal stage of maturitybased on the cessation of new thoracic segment release.However, species with a relatively large degree of variation oroutliers were tested for allometry by comparing the ratios toanother size measure, commonly the overall length of thecranidium (data not shown), and the smallest specimens wereremoved until allometry was no longer evident in the dataset.
For some characters, the spread of ratio values is broken byone or more obvious gaps (e.g., character 48, Fig. 4.1). Forthese characters, specimen assignments using FMC result infew or no polymorphic species. For other characters, there areno gaps in the dataset but values are clustered in such a waythat the data can be described by more than one normaldistribution (e.g., character 45, Fig. 4.2). Finally, in somecases, the variation within species is very large compared tothe variation among taxa (e.g., character 47, Fig. 4.3). In thesesituations, so many species are coded as polymorphic that thecharacter is essentially phylogenetically uninformative. Thespread of data might also be distributed in such a way that themost strongly supported number of normal distributions isone. Four of the eight continuous characters fell into the firsttwo categories and searches were performed just using thesefour characters (Appendices 2, 3). FMC was accomplishedusing FMCBox (Sheets, 2003).
Another way to reduce the influence of unequal sampling isto apply character coding to species means using gap-weighting (Thiele, 1993), which can be applied to anycharacter, including those that cannot be coded using FMC(Appendices 2, 4). One problem with using gap-weighting isthat it can create characters with an order of magnitude morestates than the qualitative characters. In order to maintainparity, gap-weighted characters must be down-weightedrelative to other characters (Kitching et al., 1998, p. 35),requiring the worker to make a priori assumptions about therelative importance of some morphological transitions com-pared to others. Nonetheless, there are theoretical consider-
FIGURE 4—Examples of the variation in continuous characters amongtaxa. Each species represented by a boxplot; boxplots are ordered bywithin-species mean: 1, character 48 (K2/J4) has an obvious gap in thedistribution of values among species; as a result, very few species are codedas polymorphic using FMC; 2, character 45 (G1/F1) has no obvious gaps
r
but the values are clustered in such a way that the data can be described bymore than one normal distribution; as a result, character states can beidentified using FMC but many species are considered polymorphic; 3,character 47 (C1/B1) has so much variation that even if more than onecharacter state is identified using FMC, so many species are coded aspolymorphic, that the character is phylogenetically uninformative.
1134 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

ations that indicate that gap-weighting may be preferable toother methods, particularly to other gap-coding methods. Ingeneral, other gap-coding methods implicitly weight characterstate transitions by creating intervals in which change isconsidered phylogenetically uninformative and ‘gaps’ in whichchange is considered phylogenetically informative (Thiele,1993). In contrast, gap-weighting does not ignore variation inthe sizes of gaps, variation within character state ranges, orstate ranges that are longer than the gaps themselves (Wiens,2001). By ordering the character (Wagner optimization), gap-weighting incorporates information about the distance be-tween states, particularly for characters where gaps are notevenly distributed (Wiens, 2001). In addition, Garcia-Cruzand Sosa (2006) compared gap weighing to four other gap-coding methods (but not FMC) and found that gap-weightingproduced the greatest number of informative characters, theleast number of most parsimonious trees with better cladesupport, and better results in terms of phylogenetic signalestimators.
Because continuous characters are typically additive (i.e.,ordered), they may also be analyzed using Farris’s (1970)algorithms for optimization (Goloboff et al., 2006). Asimplemented in the phylogeny program TNT (Goloboff etal., 2008), continuous characters may take values between 0and 65 with up to three decimal values. In effect, this methodis comparable to discretizing continuous characters into a verylarge number (65,001) of ordered character states, but does soin a computationally tractable way for datasets with largernumbers of taxa (cf. step-matrix method of Wiens, 2001). Thusit appears that one benefit of this might be that the amount ofvariation ‘contained’ within each character state is dependenton measurement precision, whereas in gap-weighting, theamount of variation ‘contained’ within each character state islargely dependent on (and therefore subject to) the range ofvariation captured in the overall dataset. An additionaladvantage that this method has over gap-weighting is theoption to include a range of values for each taxon (ranges areeffectively treated as polymorphisms). Here, ranges were set atone standard deviation above and below the mean value; taxafor which only one specimen was available for measurementare simply assigned the measurements taken from thatspecimen (Appendix 2).
In this study, searches were run separately for matricesincluding characters coded by FMC, for matrices includingcharacters coded by gap-weighting and for matrices includingrange values for continuous characters (Appendices 2–4). Theresults of these searches were compared to trees producedwhen continuous characters are excluded entirely.
Search method.—Heuristic searching in PAUP*4.0b10(Swofford, 1998) and TNT 1.1 (Goloboff et al., 2008) wasused to find the optimal tree according to the maximumparsimony criterion. Inapplicable characters were treated asmissing data. Taxa were added by random sequence additionwith 20 replicates and branch swapping was performed usingthe tree bisection reconnection option (TBR). When gap-weighted characters were included, all characters were scaledfor equal character weighting (characters were weightedrelative to their number of character states). In TNT, theoutgroup is limited to one taxon, and in this case, Aphelaspishaguei was selected. Bremer support values (Bremer, 1988)were calculated for resulting trees.
Stratigraphic consistency.—Stratigraphic ranges for eachtaxon were estimated using a composite section of con-tinent-wide (Laurentian) occurrences of Steptoean taxa builtusing constrained optimization (CONOP, Sadler et al., 2003).
The composite section was constructed using taxa other thanthe Great Basin pterocephaliid and aphelaspid trilobites of thisstudy. The reason for doing this was to increase theindependency between the construction of the timescale(represented by the composite section) and the estimates ofstratigraphic range for the taxa under study. This reduces thenumber of taxa used to estimate the composite section butcomparison with the composite section built using all of thedata indicates that the results are robust to the exclusion ofthese taxa (Spearman rank correlation of first appearances50.984, P%0.001; Spearman rank correlation of stratigraphicdurations50.981, P%0.001). Confidence intervals were esti-mated by compiling numerous equally good solutions to thecorrelation problem (for a detailed discussion, see Hopkins,2010). Stratigraphic consistency was estimated using the GapExcess Ratio (GER), calculated using the program Ghost 3.2(Wills, 1999). Consensus trees were pruned to include only taxafor which first and last appearance data could be estimatedfrom the Laurentian composite section. Because the results arebased on different sets of characters, the GERs are not directlycomparable. They can be ranked, however, by the significanceof the GERs through randomization tests (here 1000 iterations).Significance values indicate approximate uncertainty thatcongruence is significantly better than random. Analyses wererun under the worst-case scenario for polytomies.
RESULTS
All trees have low consistency indices, moderately highretention indices, similar degrees of stratigraphic consistency,and identical GER significance values (Table 2). The additionof even a few continuous characters (e.g., the characters codedusing finite mixture coding) dramatically decreased thenumber of most parsimonious trees, and thus increased theresolution of the analysis (Figs. 5–9). In scaling compositetrees to stratigraphic placement, nodes that could not bepinned to stratigraphic occurrences were given an arbitrarilyshort length that did not obscure the graphical depiction ofthe cladogram. A complete list of unambiguous charactertransformations is included in Appendix 5, available from theJournal of Paleontology’s Supplemental Data Archive, www.journalofpaleontology.org.
Coding characters using gap-weighting and treating contin-uous characters ‘‘as such’’ in TNT produce almost identicaltrees with the exception of the placement of a few adjacentbranches within genera. Thus, treating the continuouscharacters ‘‘as such’’ did not perform differently than gap-weighting (it neither produced a very different topology norimproved the result as measured by the above indices). Ingeneral, however, this method was found to be more flexiblebecause it can accommodate within-species variation andwould not require re-coding if additional taxa or specimensare added in the future.
TABLE 2—Search results using only qualitative characters (NONE), usingFMC-coded continuous charactres (FMC) and gap-weighted continuouscharacters (GW), and treating continuous characters as such (TNT).Char5number of characters; MPT5number of most parsimonious trees;TL5tree length; CI5consistency index; RI5retention index; GER5gapexcess ratio of Wills, 1999; GER sig.5significance of gap excess ratio,statistically significant values bolded.
Char MPT TL CI RI GER GER sig.
NONE 44 796 264 0.337 0.638 0.540 0.1FMC 48 9 290 0.390 0.640 0.750 0.1GW 52 1 197.7338 0.321 0.650 0.690 0.1TNT 52 1 248.073 0.287 0.637 0.704 0.1
HOPKINS—PTEROCEPHALIID PHYLOGENY 1135

DISCUSSION
Consistencies across trees.—Results vary across datasets butthere are some features that are shared by all or many of thetrees regardless of how continuous characters are treated.Notably, the genus Housia is strongly supported by all analyses,even as less parsimonious trees are accepted (see Bremersupport values, Figs. 6–9). In addition, all trees support thefollowing sister pairs: 1) Cernuolimbus laevifrons, Palmer 1965b,and C. depressus; 2) Cernuolimbus semigranulosus and C.granulosus; 3) Parahousia constricta Palmer, 1960, and P.subequalis; 4) Dicanthopyge reductus Palmer, 1965b, andDicanthopyge convergens Palmer, 1965b; and 5) Housia varroand H. vacuna. Prehousia indenta Palmer, 1965b, and Prehousiaalata, and Sigmocheilus flabellifer (Hall and Whitfield, 1877)and Sigmocheilus pogonipensis (Resser, 1942) are supported assister pairs when continuous characters are included and in themajority of most parsimonious trees when they are excluded.All trees support the following monophyletic groups: Para-housia, Housia, Dicanthopyge, and Strigambitus. For all butStrigambitus, the internal topology of these clades is consistentacross all trees. Housia is united by having strongly ventrally-sloping axial furrows anterior to the palpebral lobes (character12), a semicircular pygidium commonly ankylosed with theposterior-most thoracic segment (character 29), and palpebrallobes that contact the axial furrow at the anterior end (character14), resulting in a lack of ocular ridges and an exceptionallynarrow interocular area. All of these characters have beendescribed previously for this genus. All trees support a cladecomprised of Pterocephalia, Strigambitus, and Sigmocheilus; theinclusion of Cernuolimbus in this clade is supported whencontinuous characters are included and unresolved otherwise.Where resolved, the subfamily Pterocephaliinae is united byhaving a concave anterior border (character 1), Pterocephalia
FIGURE 6—Adams consensus tree found when only qualitative characters are included: 1, cladogram; 2, consensus tree scaled to relative stratigraphic placementbased on the composite section built in CONOP. Error bars indicate 95% confidence intervals around first appearance datum for each species.
FIGURE 5—Strict consensus tree found when only qualitative charactersare included, showing Bremer support values greater than 1 (circledvalues) and the distribution of characters where states are unambiguous(i.e., occur at the same node under both ACCTRAN and DELTRANoptimization), confined to a single node, and shared by all terminal tipsconnected by that node (black bars).
1136 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

and Sigmocheilus are united by having a submarginal anteriorfacial suture (character 21), and Cernuolimbus is united byhaving a row of coarse ornamentation on the anterior borderfurrow of the internal mold (character 6). The basal position ofCernuolimbus relative to other pterocephaliine genera isconsistent with stratigraphy.
Species of the subfamily Housiinae have short, straightpalpebral lobes located anterior to the glabellar midlength andpoorly defined by the palpebral furrows. However, there is nounique character that distinguishes this subfamily from other taxain the analysis. This may be in part due to the method of codingcontinuous characters (one of which describes relative palpebrallobe length). In addition, although the pterocephaliine Strigambi-tus has longer and better defined palpebral lobes than housiinespecies, they are also located anterior to the glabellar midlength.
Nonetheless, the results support a subfamilial distinctionbetween Pterocephaliinae and Housiinae as well as thePterocephaliidae as a monophyletic group. The inclusion ofAphelaspis longispina Palmer, 1965b, in the Housiinae shouldbe interpreted with caution because the character that unitesthe Pterocephaliidae (the ventral marginal suture) is unknownin this taxon. In addition, this study includes only three of 36described species of Aphelaspis, and the inclusion of additionalspecies of Aphelaspis (or other aphelaspids) may help tostabilize the position of this species.
Ornamentation and pygidial characteristics are the mostimportant for designation of congenerics.
Inconsistencies across trees.—The primary difference be-tween the trees, particularly those including data from
continuous characters, is the status of Prehousia, Pterocepha-lia, and Cernuolimbus as monophyletic clades.
Prehousia is monophyletic in trees constructed using FMCand paraphyletic in the tree constructed using gap-weighting.The opposite is true for Pterocephalia and Cernuolimbus.However, if characters 45 or 47 are excluded from the analysisusing gap-weighting, the resulting most parsimonious tree isalmost identical to those produced using FMC. Similarly,monophyly of Pterocephalia is lost (either Strigambitus orSigmocheilus is nested within the genus) if either characters 47or 49 are excluded from the analysis in TNT using continuouscharacters. This suggests that the length of the palpebral loberelative to the glabella (character 47), the length of the anteriorborder relative to the frontal area (character 45), and the lengthof the frontal area relative to the entire cranidium (character 49)are important in determining major structural differencesbetween the results. Coding characters 45 and 50 using FMCactually results in a loss of information concerning thedistribution of this character across taxa. For example, despitethe high range in variation that pterocephaliine species express(see coding for gap-weighting) in character 45, all of them arecoded identically using FMC. This is particularly important forPterocephalia, which is easily distinguished from other pter-ocephaliine species by the exceptionally long anterior border(sag.) relative to the length of the preglabellar field (sag.) andoverall length (sag.) of the cranidium. The inclusion of thesecharacters reverses the polarity of the branch containingPterocephalia and Sigmocheilus (compare Figs. 7, 8) andsupports Pterocephalia as a monophyletic clade.
FIGURE 7—Strict consensus tree found when continuous characters are coded using FMC: 1, cladogram showing Bremer support values greater than 1(circled values) and the distribution of characters where states are unambiguous (i.e., occur at the same node under both ACCTRAN and DELTRANoptimization), confined to a single node, and shared by all terminal tips connected by that node (black bars); 2, consensus tree scaled to relativestratigraphic placement based on the composite section built in CONOP. Error bars indicate 95% confidence intervals around first appearance datum foreach species.
HOPKINS—PTEROCEPHALIID PHYLOGENY 1137

The question of whether Cernuolimbus is monophyletic restsin the placement of C. pegakanthodes, which varies acrossresults. However, the row of ornamentation on the internalmold that defines this genus is unknown from this species, thusC. pegakanthodes is retained in the genus until additionalmaterial can be described.
‘‘Prehousia’’ semicircularis was originally assigned to Pre-housia but the results indicate that ‘‘Pr.’’ semicircularis is moreclosely aligned with other housiine species (its retention inPrehousia makes the genus polypheletic in trees resulting fromFMC or gap-weighted coding of continuous characters). Withonly a handful of specimens designated to this taxon,‘‘Prehousia’’ semicircularis remains a poorly known species.However, some comparisons can be made. In particular,‘‘Prehousia’’ semicircularis is similar to Parahousia in havingan upsloping interocular area (character 22) and prominentpalpebral lobes (character 16) but lacks the concave axialfurrows (character 11), the strongly tapering anterior borderthat gives this part of the cranidium a triangular shape, theunconnected border furrows on the librigena (character 26),and the strongly arched pygidium characteristic of Parahousia.‘‘Prehousia’’ semicircularis forms a natural group withParahousia only in trees built using gap-weighting. Thus, theresults of the analysis do not justify broadening the definitionof Parahousia in order to accommodate this species. In fact,this species may represent a new genus of housiine, but thepaucity of material currently available precludes erection of anew genus.
Bremer support values are high (2) for the clade includingPrehousia alata, Prehousia indenta, Prehousia diverta, and
Prehousia impolita when continuous characters are included.The question of whether Prehousia sensu stricto is monophy-letic, rather than paraphyletic, rests in the placement ofPrehousia prima Palmer, 1965b, which varies across results.The palpebral lobes are longer relative to the glabella in thisspecies compared to cogenerics. Because exclusion of thischaracter from the analysis using gap-weighted charactersresults in trees almost identical to those based on FMC, mostof the Prehousia species could be united by further restrictingthe size of the palpebral lobes relative the glabella. However,because Prehousia prima is similar in cranidial morphology toAphelaspis and because A. longispina is placed immediatelybasal to this species, generic reassignment is best deferred tofuture analyses also addressing the systematics of Aphelaspis.Thus no action to reassign this species is taken here.
Previously hypothesized evolutionary transitions.—Palmer(1965b) hypothesized evolutionary transitions between someof the Great Basin species based on morphological similarityand relative stratigraphic placement. Without stating soexplicitly, his discussion suggests that these might be examplesof anagenetic trends or ancestor-descendent pairs. Thecombination of phylogenetic analysis and composite stratig-raphy presented above provides an opportunity to examinesome of these putative transitions. The discussion below isbased on character reconstructions on one randomly selectedmost parsimonious tree.
Palmer suggested that Prehousia may have arisen fromAphelaspis, specifically that there was a transition fromAphelaspis subditus to Prehousia prima Palmer, 1965b, to Pr.indenta to Pr. alata. This series is characterized by a
FIGURE 8—Most parsimonious tree found when continuous characters are coded using gap-weighting: 1, cladogram showing Bremer support valuesgreater than 1 (circled values) and the distribution of characters where states are unambiguous (i.e., occur at the same node under both ACCTRAN andDELTRAN optimization), confined to a single node, and shared by all terminal tips connected by that node (black bars); 2, consensus tree scaled torelative stratigraphic placement based on the composite section built in CONOP. Error bars indicate 95% confidence intervals around first appearancedatum for each species.
1138 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

morphological shift to a narrower interocular area, smallerpalpebral lobes, and an increasingly unevenly curved posteriormargin on the pygidium, culminating in the expression of alaeon Pr. alata (Fig. 10, see also Palmer, 1965b, fig. 12). Almostall trees support a sister relationship between Pr. alata and Pr.indenta and a basal position for Pr. prima relative to these twospecies. In many trees, however, both Pr. alata and Pr. indentaare more closely related to Prehousia impolita Palmer, 1965b,and Prehousia diverta Palmer, 1965b, than to Pr. prima. As anoutgroup, Aphelaspis subditus is, to some extent, constrainedto a position basal to Prehousia. However, results for allcharacter datasets were the same when only Aphelaspis hagueiwas designated as the outgroup. Thus, the tree structuresuggests it is unlikely that any Prehousia species descendeddirectly from Aphelaspis subditus. In contrast, Aphelaspislongispina appears more closely related to Prehousia.
Nonetheless, both regional stratigraphy and tree topologyare consistent with a phyletic transformation (i.e., anagenesis)from Prehousia indenta to Pr. alata. These two taxa differ inthe coding of four qualitative characters. A transition from Pr.indenta to Pr. alata would require acquisition of a polymor-phism from a fixed character state (character 8), an increase inthe number of pygidial axial rings and pleural furrowsexpressed in the pygidium (character 32 and 37, respectively),and the appearance of alae on the antero-lateral margin of thepygidium (character 44). Because a phyletic transition betweenPr. indenta and Pr. alata does not require an increase in thebranch length relative to evolution from a common ancestorregardless of whether one assumes accelerated or delayed
transformation of character states (Fig. 10), it is no lessparsimonious than the hypothesis that both evolved cladoge-netically from a common ancestor. In fact, it could beconsidered more parsimonious because it does not requirean ad hoc extension of the stratigraphic range of Pr. alata.
Though not discussed by Palmer (1965b), both treetopography and stratigraphy support the hypothesis thatParahousia constricta evolved anagenetically from P. subequa-lis, that Cernuolimbus depressus evolved from C. laevifrons,and (on some trees) that Strigambitus bilobus evolved fromStrigambitus utahensis. Based on character reconstructionalong the tree and including the apomorphy for P. subequalisexcluded from the character matrix (see Methods), all are lessparsimonious than cladogenesis but only require one to threeadditional transitions depending on whether one assumesaccelerated or delayed transformation.
Tree topology and stratigraphy is also consistent with directdescent through budding cladogenesis of Cernuolimbus granulo-sus from C. semigranulosus, and (on some trees) of Sigmocheiluspogonipensis from S. flabellifer. In the former case, thishypothesis is equally parsimonious to that of cladogenesis, butthe latter requires the reversal of at least half a dozen characterstates unique to S. pogonipensis among other cogenerics.
Finally, Palmer (1965b) suggested that Dicanthopyge mayhave descended from Aphelaspis longispina and that specieswithin Dicanthopyge formed an ‘‘evolutionary series’’ whereDicanthopyge quadrata Palmer, 1965b, preceded D. convergens,which in turn preceded D. reductus. Though not originallyincluded in the phylogenetic analysis in order to evaluate this
FIGURE 9—Most parsimonious tree found when continuous characters are treated as such as implemented in TNT: 1, cladogram showing Bremersupport values greater than 1 (circled values) and the distribution of characters where states are unambiguous (i.e., occur at the same node under bothACCTRAN and DELTRAN optimization), confined to a single node, and shared by all terminal tips connected by that node (black bars);2, consensus tree scaled to relative stratigraphic placement based on the composite section built in CONOP. Error bars indicate 95% confidence intervalsaround first appearance datum for each species.
HOPKINS—PTEROCEPHALIID PHYLOGENY 1139

trend, the presence of these species as part of the outgroupallows some assessment. All trees support a sister relationshipbetween D. convergens and D. reductus and the placement ofD. quadrata directly basal to this pair. Although thishypothesis is consistent with the order of first appearances,duration estimates extracted from the composite section
(Figs. 6–8) imply that these three species coexisted eventhough they have not yet been observed to co-occur locally(this result is also supported by the composite section builtusing all taxa with the exception that the last appearance of D.quadrata precedes the last appearance of the other two taxa).The composite stratigraphy is inconsistent with a strictphyletic transformation between Dicanthopyge species but itdoes not preclude the possibility that the younger speciesbudded off from the older species before extinction (i.e.,cladogenesis of Wagner and Erwin, 1995; and buddingcladogenesis of Foote, 1996). For example, Dicanthopygeconvergens and D. reductus differ in the coding of threequalitative characters. Because of the character distributionwithin this genus, the branch leading to D. reductus contains aloss of pitted ornamentation (character 8), a shift from asubquadrate-shaped pygidium to a more semicircular pygid-ium (character 30) and a shift in the type of ornamentation onthe pygidium (character 42) regardless of whether D reductus ishypothesized to have evolved from an ancestor common withD. convergens or from a population of D. convergens itself.Thus both hypotheses are equally parsimonious. In contrast,budding of D. convergens from D. quadrata would requireadditional character state transitions and is not supported byparsimony.
In summary, almost half of the putative ancestor-descendentpairs described above are supported by both tree topology andrelative stratigraphic placement and are equally likely tocladogenesis under an assumption of parsimony. In addition,most of the other pairs require only a few additional characterstate transitions. This is possible in this study because species-level differences exist primarily in character states unique withinthe genus rather than across all species (e.g., ornamentation). Inaddition, many of these characters are multistate and speciesdifferences are dominated by character state transitions ratherthan reversals. Thus, depending on the polarity of characteracquisition as well as character reconstruction on internalnodes, some species pairs may be interpreted as phyletictransitions without suffering a decrease in parsimony. None-theless, the multispecies evolutionary series posited by Palmer(1965b) are not supported by the results.
Affinities between included and excluded pterocephaliid taxa.—Because Camaraspis, Pulchricephalus, and Pelicephalus do notpossess the concave border that defines the Pterocephaliinae(Palmer, 1965b, p. 57) and is the uniting characteristic of thissubfamily in this analysis, they are hereafter excluded from thePterocephaliinae (sensu Palmer, 1965b). Of the genera currentlyassigned to the Pterocephaliidae, Cernuolimbus, Sigmocheilus,Pterocephalia, and Strigambitus possess a concave border and areretained within Pterocephaliinae. Inspection of published figuresindicates that eight additional genera, primarily from China, alsoappear to have concave borders and are thus questionablyassigned to this subfamily. These include Beigongia Qiu, in Qiuet al., 1983, Dikelocephalioides Qian, 1994, Dikelocephalites Sun,1935, Dingxiangaspis Zhang, in Qiu et al., 1983, JubileiaKobayashi, 1938, Prodikelocephalites Yuan and Yin, 1998,Yokusenia Kobayashi, 1935, and Zhenania Luo, 1983.
Camaraspis, Pulchricapitus, and Pelicephalus do share somesimilarities with some housiine genera as do other pteroce-phaliids not yet assigned to a subfamily. For example, likeother housiines, Pulchricapitus and Pelicephalus have relativelyshort palpebral lobes compared to glabellar length (sag.).Pelicephalus and Morosa Palmer, 1960, are similar to Para-housia in having strongly tapering anterior borders (character3) and upsloping interocular areas (character 22), but lack theconcave axial furrows that are diagnostic of Parahousia
FIGURE 10—Reconstruction of character state transitions for Prehousiaindenta Palmer, 1965b, and Prehousia alata Palmer, 1960, based onphylogenetic analysis and relative stratigraphic placement: 1, hypothesis ofevolution from common ancestor (cladogenesis); 2, hypothesis of phyletictransition from Pr. indenta to Pr. alata (anagenesis).
1140 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

(character 11). Morosa and Luotuolingia Lu and Qian, 1983,have wide and well defined pygidial borders similar to Housiaand Tumicephalus. Xiaoshiella Lu and Qian, 1983, is similar toHousia in having a strongly tapering glabella, palpebral lobesabutting the axial furrow (character 14), narrow interocularareas and a poorly defined anterior border, but the anteriorpart of the glabella and frontal area do not appear to bestrongly ventrally sloping (character 12). Petalocephalus Luand Qian, 1983, has narrow interocular areas and poorlydefined palpebral lobes similar to Housia, but the glabella doesnot taper as strongly, and the pygidium is longer (sag.) thanwide (tr.) with a median inbend along the posterior margin(character 41). In addition, Pulchricapitus and StenambonPalmer, 1965b, differ from other housiine genera by acombination of characters, including a strongly definedanterior border furrow (character 2), a consistently wideanterior border (character 3), wide (tr.) frontal and interocularareas relative to cranidial length (sag.) (character 49), andstrongly expressed granular ornamentation. Thus, thoughsome of the genera not currently assigned to a subfamily mayultimately fall within the Housiinae, their inclusion is unlikelyto affect the status of the included genera as monophyleticclades. This could be tested in future analyses by adding thesegenera to the analysis, but because this would increase thenumber of taxa by more than 50% (more than 100% if theChinese taxa with concave borders are added as well), isbeyond the scope of the current study.
SYSTEMATIC PALEONTOLOGY
The following includes revisions or additions to previoussystematic descriptions for taxa included in this analysis basedon examination of previously collected material as well as newmaterial from field collections. See Appendix 6 (available fromthe Journal of Paleontology’s Supplemental Data Archive atwww.journalofpaleontology.org) for literature sources andmaterial examined for species included in the phylogeneticanalysis but not amended below.
Terminology.—Morphological terminology follows Whit-tington and Kelly (Palmer, 1997). Abbreviations used: USNM5National Museum of Natural History (Smithsonian);FMNH5Field Museum of Chicago; USGS5U.S. GeologicalSurvey collection, detailed locality information in Palmer(1960, 1965b); ICS5Institute for Cambrian Studies collection,University of Chicago. All remaining collection numbers referto field collections made from 2005 to 2008. Detailed localityinformation for ICS and field collections may be found inHopkins (2010).
Family PTEROCEPHALIIDAE Kobayashi, 1935Subfamily HOUSIINAE Hupe, 1953Genus PARAHOUSIA Palmer, 1960
Not figured
Parahousia PALMER, 1960, p. 76; PALMER, 1965b, p. 66.
Type species.—Parahousia constricta Palmer, 1960.Other species.—Parahousia subequalis Palmer, 1965b.Diagnosis.—Amended from Palmer (1965b). Anterior bor-
der longer (tr.) abaxially, tapers strongly laterally. Glabellatapers strongly anteriorly; axial furrows gently concavebetween palpebral lobes in dorsal view. Node on occipital ring.
Discussion.—The strongly tapering anterior border gives theanterior border a triangular shape. Neither this nor the gentlyconcave axial furrows have been explicitly described asdiagnostic for this genus but both features have previouslybeen recognized as genus-level characteristics (e.g., see KeyNo. 26 in Palmer, 1965b, p. 24) and are visible in previously
published figures of the holotypes (see Palmer, 1960, 1965b).In addition, both serve to distinguish this genus fromPrehousia and Housia. Parahousia is further distinguishedfrom these genera by the strongly arched pygidium.
Parahousia constricta and P. subequalis are currently theonly two species belonging to this genus. A wider (sag.)preglabellar field relative to border width (sag.), lack of coarsepitting on cephalon, more prominent fossulae, and moreextensive coverage of terrace lines across the pygidial borderdistinguish P. subequalis from P. constricta. The two specieshave been found in the same locality only at Shingle Pass,where P. constricta occurs above P. subequalis (Palmer,1965b). Parahousia subequalis was uncertainly identified fromthe Mackenzie Mountains of Canada (Pratt, 1992); inMissouri, P. subequalis has been found in subsurface depositsand P. constricta has been found in outcrop (Kurtz, 1975).Description of both species in Palmer (1960, 1965b) aresufficient.
Genus PREHOUSIA Palmer, 1960
Prehousia PALMER, 1960, p. 77; PALMER, 1965b, p. 67.
Type species.—Prehousia alata Palmer, 1960.Other species.—Prehousia diverta Palmer, 1965b; Prehousia
impolita Palmer, 1965b; Prehousia indenta Palmer, 1965b;Prehousia prima Palmer 1965b.
PREHOUSIA ALATA Palmer, 1960Figures 2.2, 11.4, 11.7, 11.8, 11.10
Prehousia alata PALMER, 1960, p. 78, pl. 7, figs. 10, 12, 13;PALMER, 1965b, p. 67, pl. 13, figs. 2, 5, 8, 9, 12, 13.
Diagnosis.—Palmer (1965b) is sufficient.Description.—Amended from Palmer (1965b). Slight angu-
lation in anterior margin of cranidium at axial line. Anteriorfacial suture divergent; curves medially at border, cuttingmargin at axial line. Librigenal doublure terminated bystraight margin perpendicular to anterior margin. Pygidialposterior margin unevenly curved; alae present at anterolat-eral margins. Fixigena and pleural regions of the pygidiacovered with fine, shallow pits; internal mold of cranidiashows coarse pitting on frontal area and fixigena.
Material examined.—NEVADA: Snake Range, White PineCounty: USGS 1197-CO, USGS 1435-CO, USGS 1441-CO,ICS-1838, ICS 1839. Shingle Pass, Lincoln County: USGS2313-CO. Desert Range, Clark County: DR07. PahranagatRange, Lincoln County: PR49, PR51, PR53. Spring Moun-tains, Clark County: ICS-1390. UTAH: House Range, MillardCounty: OR38.
Discussion.—No complete specimen of Prehousia alata is yetknown but a comparison of librigena and cranidia indicatethat the anterior facial suture more likely cut the anteriormargin at the axial line rather than near the axial line, aspreviously described (Palmer, 1960, 1965b). In particular, theaxial suture of the doublure of the librigena is perpendicular tothe anterior margin (Fig. 2.2), and the shape of the anteriormargin on the cranidium is slightly angular at the axial marginrather than abaxial to the axial line as visible in other generawith rostral plates, such as Aphelaspis or Dicanthopyge (cf.Fig. 11.10 with Figs. 1.3, 11.3). This distinction is importantbecause it implies that Prehousia alata had a ventral mediansuture rather than a small subquadrate rostrum as illustrateddiagrammatically in Palmer (1960, p. 64). One librigenalspecimen of Prehousia prima suggests a similar ventral suturebut the adaxial margin is not well preserved (Fig. 11.5).Otherwise, the nature of the librigenal doublure is unknown
HOPKINS—PTEROCEPHALIID PHYLOGENY 1141

for other Prehousia species. Prehousia species are so similar incranidial shape, however, that it is likely that the ventralmedian suture is a genus level characteristic.
Identification of Prehousia to species level can be verydifficult. Prehousia diverta is most easily distinguished by thewide cranidial border and undefined border furrow and bylacking pitting or granulation on any sclerites. Prehousiaimpolita has the narrowest border of the Prehousia spp.(relative to both frontal area and cranidial length) and finegranulation on the cranidium. Prehousia impolita is thesmallest Prehousia species: the largest cranidia examined were6.2 mm in sagittal length. In contrast, the largest cranidia
examined for Pr. alata, Pr. diverta, Pr. indenta, and Pr. primawere 13.3, 16.1, 15.2, and 13.0 mm, respectively.
Prehousia indenta, Pr. alata, and Pr. prima are very similar toone another in cranidial shape. There is a gradient in overallmorphology from older Pr. prima to younger Pr. prima to Pr.indenta to Pr. alata: the fixed cheeks become narrower, thepalpebral lobes shorten relative to the glabella and shiftanteriorly relative to the glabellar midline, and the divergencebetween the anterior branch of the facial suture and the palpebrallobe decreases (cf. Fig. 11.10–11.12). Palmer (1965b) recognizedthis gradient in the comparative lengths of the palpebral lobe,width of fixed cheeks, and length of glabella (his fig. 12).
FIGURE 11—Prehousia Palmer, 1960, species: 1, ‘‘Prehousia’’ semicircularis Palmer, 1960, cranidium, USNM 543781,Windfall Canyon, Eurekadistrict, NV, 312; 2, ‘‘Prehousia’’ semicircularis, cranidium, USNM 136868, Windfall Canyon, Eureka district, NV, 312; 3, Aphelaspis haguei (Hall andWhitfield, 1877), cranidium with arrow indicating point where anterior facial suture crosses anterior margin, FMNH PE60960, McGill section, DuckCreek Range, NV, 312; 4, Prehousia alata Palmer, 1960, librigena, USNM 543783, Snake Range, NV, 33; 5, Prehousia prima Palmer, 1965b, librigena,USNM 543785, Shingle Pass, NV, 35; 6, Prehousia indenta Palmer, 1965b, pygidium, FMNH PE60961, Mormon Mountains, NV, 37; 7, Prehousiaalata, pygidium, USNM 543782, Snake Range, 35; 8, Prehousia alata, pygidium, USNM 136867b, Snake Range, NV, 34; 9, Prehousia prima, pygidium,USNM 543784, Shingle Pass, NV, 37; 10, Prehousia alata, cranidium with arrow indicating point where anterior facial suture crosses anterior margin,FMNH PE60962, Orr Ridge, House Range, UT, 36; 11, Prehousia prima, cranidium, USNM 543786, McGill section, Duck Creek Range, NV, 38;12, Prehousia indenta, cranidium, FMNH PE 60972, Mormon Mountains, NV, 35.
1142 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

Some pygidia of Prehousia indenta have unevenly curvedposterior margins (Fig. 11.6) but alae are underdevelopedcompared to Pr. alata, where the alae are defined by a kink inthe anteriormost pleural furrow in addition to the shape of themargin (Fig. 11.7, 11.8). Prehousia alata also differs from Pr.indenta in having narrower fixed cheeks, more poorly definedpalpebral lobes and an anterior facial suture which divergesfrom the palpebral lobe at a smaller angle. In addition, Pr.alata has a smooth or very faintly pitted cranidium, whereasboth Pr. indenta and Pr. prima have a coarsely pittedcranidium. Coarse pitting appears on the internal mold ofPr. alata. Within Housiinae, species with a gentle inbend in thepygidial border are confined to Prehousia but this is not aunique character across the entire set of taxa.
Thus, though identification can be difficult, there appear to beno characters or suite of characters that justify synonymizing orsplitting any of these five species. This system might benefit fromfurther study, particularly if more silicified material is discovered.
PREHOUSIA? PRIMA (Palmer, 1965b)Figure 11.5, 11.9, 11.11
Prehousia prima PALMER, 1965b, p. 68, pl. 12, figs. 16–18.
Diagnosis.—Palmer (1965b) is sufficient.Material examined.—NEVADA: Duck Creek Range, White
Pine County: USGS 2501-CO, USGS 2502-CO, USGS 2503-CO,USGS 2504-CO. Shingle Pass, Lincoln County: USGS 2560-CO,USGS 2561-CO. Pahranagat Range, Lincoln County; PR13.
Discussion.—Questionably retained in Prehousia. See dis-cussion of phylogenetic results and Prehousia alata above.
‘‘PREHOUSIA’’ SEMICIRCULARIS (Palmer, 1960)Figures 1.10, 11.1, 11.2
Prehousia semicircularis PALMER, 1960, p. 78, pl. 7, figs. 11,14, 15, 19; 1965b, p. 69, pl. 12, figs. 24–26.
Diagnosis.—Amended from Palmer (1965b). Anterior bor-der narrower (sag.) than preglabellar area, defined by narrow,
FIGURE 12—Sigmocheilus Palmer, 1960, species: 1–3, Sigmocheilus pogonipensis (Resser, 1942): 1, cranidium, FMNH PE60963, Cherry Creek Range,NV, 38; 2, cranidium, from FMNH PE60964, Desert Range, NV, 315; 3, pygidium, from FMNH PE60965, Pahranagat Range, NV, 35; 4, 5,Sigmocheilus notha (Resser, 1942): 4, pygidium, USNM 543787, Cherry Creek Range, NV, 38; 5, cranidium, FMNH PE60966, Pahranagat Range, NV,35; 6, 7, Sigmocheilus grata: 6, cranidium, USNM 543774, Windfall Canyon, Eureka district, NV, 38; 7, latex cast of pygidium, USNM 543775.Windfall Canyon, Eureka district, NV, 34; 8, 9, Sigmocheilus flabellifer (Resser, 1942): 8, cranidium, FMNH PE60967, Cherry Creek Range, NV, 35;9, pygidium, FMNH PE60968, Desert Range, NV, 39.
HOPKINS—PTEROCEPHALIID PHYLOGENY 1143

deep furrow. Glabella tapering anteriorly, glabellar furrowspoorly defined. Axial furrows strongly defined, straight.Anterior border furrow divergent. Interocular areas upslop-ing. Palpebral lobe one third length of glabella (sag.),prominent, well defined by palpebral furrow. Node onoccipital ring. Transversely subovate pygidium, three or fouraxial furrows behind articulating furrow, no border furrow.Smooth to roughened exoskeletal surface. Faint caecalmarkings on frontal area.
Material examined.—NEVADA: Windfall Canyon, EurekaCounty: USGS 2294-CO, USNM 136868, USNM 136869a,b, c.
Discussion.—There is very little material assigned to‘‘Prehousia’’ semicircularis and this species has only beenfound in one collection made from the Dunderberg Formationat Windfall Canyon in the Eureka district of Nevada (USGS2294-CO). Cranidia are very small with no known specimenlarger than 3.0 mm. No specimens have any pitting, but somespecimens have a ‘roughened’ appearance. This is in contrastto previous species descriptions which described this crani-dium as smooth (Palmer, 1960, 1965b), but it is unclear fromthe specimens available whether this is the actual exoskeletalsurface texture or an artifact of preservation. Ornamentationis considered a species-level characteristic herein because it isimportant for species designation in closely related genera.
Phylogenetic analysis suggests that this species is moreclosely aligned with other housiine genera than with Pre-housia. ‘‘Prehousia’’ semicircularis differs from Prehousia andHousia species by having upsloping interocular areas (charac-ter 22), prominent palpebral lobes (character 16), stronglydeveloped occipital node (character 24), more pleural fur-rows (character 37), a wider glabella (tr.) relative to cranidialwidth (tr.) (character 48), and caecal marking on the frontalarea (character 9). ‘‘Prehousia’’ semicircularis also lacksthe strongly ventrally-sloping axial furrows anterior to thepalpebral lobes (character 12), semicircular pygidium com-monly ankylosed with the posterior-most thoracic segment(character 29), and palpebral lobes that contact the axialfurrow at the anterior end (character 14) diagnostic of Housia.Characters 22, 16, and 24 are shared with Parahousia but‘‘Pr.’’ semicircularis lacks the concave axial furrows andstrongly tapering frontal area diagnostic of Parahousia.Tumicephalus depressus more closely resembles ‘‘Pr.’’ semi-circularis but has a median swelling on the preglabellar field, athin and well defined pygidial border furrow, and only twoaxial rings. Thus this species is clearly different from otherclosely related genera, including Prehousia, and may representa new genus of housiine. However, the paucity of materialavailable precludes erecting a new genus at this time.
Subfamily PTEROCEPHALIINAE Kobayashi, 1935
Diagnosis.—Palmer (1965b) is sufficient.Assigned genera.—Cernuolimbus Palmer, 1960; Pterocepha-
lia Roemer, 1849; Sigmocheilus Palmer, 1960; StrigambitusPalmer 1965b; ?Beigongia Qiu, 1983; ?Dikelocephalioides Qian,1994; ?Dikelocephalites Sun, 1935; ?Dingxiangaspis Zhang,1983, ?Jubileia Kobayashi, 1938; ?Prodikelocephalites Yuanand Yin, 1998; ?Yokusenia Kobayashi, 1935; and ?ZhenaniaLuo, 1983.
Discussion.—This subfamily is distinguished from otherpterocephaliids primarily by the presence of a concave anteriorborder. All previously assigned genera that did not have thischaracter are now excluded. Several taxa which appear to haveconcave borders based on inspection of published figures arequestionably included.
Genus CERNUOLIMBUS Palmer, 1960Figure 1.5
Cernuolimbus PALMER, 1960, p. 84; 1965, p. 69; 1968, p. B71;PRATT, 1992, p. 58, 59; PTITT ET AL., 1994, p. 519.
Type species.—Cernuolimbus orygmatos Palmer, 1960.Other species.—Cernuolimbus arcticus Palmer, 1968; Cer-
nuolimbus depressus Palmer, 1960; Cernuolimbus granulosusPalmer, 1965b; Cernuolimbus laevifrons Palmer, 1965b; Cer-nuolimbus longifrons Palmer, 1968; Cernuolimbus monilis Stittet al., 1994; Cernuolimbus pegakanthodes Pratt, 1992; Cernuo-limbus semigranulosus Palmer, 1960.
Diagnosis.—Amended from Palmer (1965b) and Stitt et al.(1994). Anterior border concave. Preglabellar field down-sloping to noticeably convex. Coarse pits or granules in rowalong anterior border furrow of internal mold (character 6).
Discussion.—The most notable addition to the diagnosis ofthis genus is the presence of a row of coarse pits or granulesalong the anterior border furrow in all species. This characterunites all species in this genus relative to other pterocepha-liine taxa; the combination of this character and the con-cave anterior border unites Cernuolimbus species relative toother pterocephaliid trilobite taxa. In addition, the anterior borderof Cernuolimbus is narrow (sag.) with a distinct border furrow.
Cernuolimbus also differs from Sigmocheilus, Strigambitus,and Pterocephalia in having a more transversely subovatepygidium with a narrower posterior border relative to the pleuralregion (not figured). Almost all Cernuolimbus species showgranulation on the genal spine (C. longifrons is smooth); otherpterocephaliine species show strong terrace lines across the spine.
Genus SIGMOCHEILUS Palmer, 1960Figures 1.1, 1.2, 1.6–1.9, 12
Sigmocheilus PALMER, 1960, p. 89, text fig. 20.
Type species.—Sigmocheilus flabellifer (Hall and Whitfield,1877) (5Sigmocheilus serratus Palmer 1960).
Other species.—Sigmocheilus aldanensis Gogin, 1990; Sig-mocheilus delicatus Lazarenko and Nikiforov, 1968; Sigmo-cheilus grata (Resser, 1942); Sigmocheilus notha (Resser, 1942);Sigmocheilus pogonopensis (Resser, 1942).
Diagnosis.—Amended from Palmer (1960). Pleural ridgesextend postero-laterally, curve posteriorly at border furrow,extend to the posterior margin in some species.
Discussion.—Palmer (1965b) described the pleural ridges asextending onto but not across the border. This is true inSigmocheilus notha and S. grata which have smoothly curvedposterior margins but not for S. flabellifer and S. pogonipensiswhich have pleural ridges extending across the border to thespinose margin. In both species, the ridges meet the posteriorborder in between adjacent spines (Fig. 12.3, 12.9).
As described in Palmer (1960), the facial suture isintramarginal until close to the axial line, where it crossesthe anterior margin and continues posteriorly until it meets theventral marginal suture. At least in Sigmocheilus notha, there isno rostrum and the anterior margin of the cranidium curvesventrally following the submarginal facial suture (Fig. 1.6–1.9). Pterocephalia is also known to have a submarginal suture(Palmer, 1960, 1965), but this characteristic has not beenobserved in Cernuolimbus or Strigambitus.
Sigmocheilus may be distinguished from Pterocephaliaby having a better-defined anterior border furrow on thecranidium and a considerably narrower (sag.) anterior borderrelative to the preglabellar field (in Pterocephalia, the bordercan be up to 10 times longer (sag.) than the preglabellar field).The strongly convex preglabellar field on Sigmocheilus pogoni-
1144 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

pensis is similar to Cernuolimbus species but S. pogonipensislacks ornamentation on the anterior border furrow of theinternal mold.
Strigambitus may be distinguished from Sigmocheilus mostreadily by having smaller palpebral lobes and more stronglyexpressed, densely packed ornamentation. With the exceptionof Sigmocheilus pogonipensis, Sigmocheilus species may also bedistinguished from Strigambitus species by having a longer(sag.) anterior border relative to the preglabellar field.Sigmocheilus pogonipensis differs from Strigambitus in pygidialcharacters, with which it is more closely allied with otherSigmocheilus species. In particular, Sigmocheilus has moreaxial rings and pleural furrows on the pygidium and thepleural furrows extend across the border, almost reaching theposterior margin. Sigmocheilus pogonipensis and S. flabelliferboth have spines along the pygidial margin associated with thefused segments that form the pygidium.
Genus STRIGAMBITUS Palmer, 1965bFigure 13
Strigambitus PALMER, 1965b, p. 75, 76.
Type species.—Strigambitus transversus Palmer, 1965b,p. 77, pl. 16, figs. 6–10.
Other species.—Strigambitus bilobus Palmer, 1965b; Stri-gambitus utahensis (Resser, 1942).
Diagnosis.—Palmer (1965b) is sufficient.Description.—Amended from Palmer (1965b). Shallow
anterior border furrow present in smaller specimens. Mid-length of palpebral lobe positioned anterior to glabellamidlength. Post-axial ridge present in some species.
Discussion.—Palmer described the anterior border of thisgenus as ‘‘poorly differentiated’’ from the preglabellar field.This is true for larger specimens but is a feature that appearsto change through ontogeny as smaller specimens show ashallow but defined furrow (cf. Fig. 13.4, 13.5 from the same
FIGURE 13—Strigambitus Palmer, 1965b, species: 1, Strigambitus bilobus Palmer, 1965b, pygidium, USNM 543769, McGill section, Duck CreekRange, NV, 312; 2–5, Strigambitus utahensis (Resser, 1942): 2, pygidium, FNMH PE60969, Pahranagat Range, NV, 38; 3, pygidium, USNM 543788,Cherry Creek Range, NV, 35; 4, cranidium, FMNH PE60970, Pahranagat Range, NV, 310; 5, cranidium, FMNH PE60971, Pahranagat Range, NV,310. 6, Strigambitus bilobus, pygidium, USNM 543780, Ruby Range, NV, 35; 7, Strigambitus transversus Palmer, 1965b, counterpart to USNM 543790,USNM 543791, Snake Range, NV, 35; 8, Strigambitus bilobus, pygidium, USNM 543779, Ruby Range, NV, 35; 9, 10, Strigambitus transversus: 9,pygidium, USNM 543789, Snake Range, NV, 310; 10, pygidium, USNM 543790, Snake Range, NV, 35.
HOPKINS—PTEROCEPHALIID PHYLOGENY 1145

collection). This feature still distinguishes Strigambitus fromSigmocheilus, which has a deeper anterior border furrow, inaddition to the other features previously described (seediscussion above).
In his description of Strigambitus, Palmer (1965b) also listeda post-axial ridge as a characteristic of the genus. However,the holotype pygidium of Strigambitus transversus, the typespecies, does not show a post-axial ridge (Palmer, 1965b, pl.16, fig. 9). Nor do other examined specimens of St. transversusshow a ridge that extends onto the border (Fig. 13.7, 13.9,13.10). In contrast, Strigambitus bilobus and St. utahensisshow low-relief ridges that extend onto the border but not tothe posterior margin (Fig. 13.1–13.3, 13.6, 13.8).
STRIGAMBITUS TRANSVERSUS Palmer, 1965bFigure 13.7, 13.9, 13.10
Strigambitus transversus PALMER, 1965b, p. 77, pl. 16, figs. 6–10.
Diagnosis.—Amended from Palmer (1965b). Posteriormargin smoothly curved to gently inbent.
Material examined.—NEVADA: Bastian Peak, White PineCounty: USGS 3003-CO. Snake Range, White Pine County:USGS 1196-CO. Duck Creek Range, White Pine County:USGS 3022-CO. UTAH: Deep Creek Range, Tooele County:USGS 2612-CO.
Discussion.—Palmer (1965b) described the posterior marginof the pygidium in Strigambitus transversus as ‘‘convexbackward’’ and that this species could be distinguished fromcogenerics by ‘‘lacking a median inbend.’’ More precisely, St.transversus has a smoothly curved to gently inbent posteriormargin such that the posterior border is shortened (sag.) alongthe axial line (Fig. 13.7, 13.9, 13.10). The inbend is not strongenough, however, that the pleural region form ‘‘lobes.’’ Incontrast, St. utahensis and St. bilobus have medium and longinbends, respectively, large enough that the pleural regions ofthe posterior margin do form ‘lobes’. Because in both St.utahensis and St. bilobus the posterior margin curves smoothlyat the axial margin rather than meeting at a point, this featureis better described as an inbend rather than a notch (Fig. 13.1–13.3, 13.6, 13.8). In addition to this feature, both St. utahensisand St. bilobus have a subquadrate pygidium rather than thetransversely ovate pygidium seen in St. transversus. Pygidia forthese species can also be distinguished from each other by therelative proportions of length and width: in St. utahensis, thepygidium is wider (tr.) than it is long (sag.) and has a shortinbend forming shallow lobes (Fig. 13.2, 13.3) whereas in St.bilobus, the pygidium is longer (sag.) than it is wide (tr.)(Fig. 13.1, 13.6, 13.8).
Strigambitus bilobus is further distinguished from either St.transversus or St. utahensis by having granular ornamentationon the axial rings but not the pleural regions of the pygidiumand a short genal spine. Cranidia are very similar among thethree species, although the anterior branch of the facial suturediverges at a greater angle from the sagittal axis in St.utahensis.
ACKNOWLEDGMENTS
Thank you to M. Webster and S. Westrop for discussion oncharacter selection and coding as well as guidance inphylogenetic analysis in general. Thank you also to M. Foote,D. Jablonski, and M. LaBarbara for reviewing earlier drafts ofthis manuscript. K. McKinney of the U.S. Geological Surveyand J. Thompson and D. Levin of the National Museum ofNatural History (Smithsonian) generously provided specimenloans. F. Sundberg and N. Hughes provided helpful reviewsand S. Westrop provided much appreciated editorial support,
all of which greatly improved the final version of this paper.Finally, this work could not have been possible without theextensive field work and specimen description undertaken byA. Palmer in the Great Basin.
REFERENCES
ADRAIN, J. M., S. E. PETERS, AND S. R. WESTROP. 2009. The Marjumantrilobite Cedarina Lochman: Thoracic morphology, systematics, andnew species from western Utah and eastern Nevada, U.S.A. Zootaxa,2218:35–58.
ADRAIN, J. M. AND S. R. WESTROP. 2001. Stratigraphy, phylogeny, andspecies sampling in time and space, p. 291–322. In J. M. Adrain, G. D.Edgecombe, and B. S. Lieberman (eds.), Fossils, Phylogeny, and Form.Topics in Geobiology, 19. Kluwer Academic/Plenum Publishers, NewYork.
ADRAIN, J. M. AND S. R. WESTROP. 2005. Late Cambrian ptychaspididtrilobites from western Utah: Implications for trilobite systematics andbiostratigraphy. Geological Magazine, 142:377–398.
ADRAIN, J. M. AND S. R. WESTROP. 2006. Notchpeakia, a new genus ofUpper Cambrian (Sunwaptan) ‘‘entomaspidid’’ trilobites. Journal ofPaleontology, 80:1152–1171.
AKAIKE, H. 1974. A new look at the statistical model identification. IEEETransactions on Automatic Control, 19:716–723.
ALMEIDA, M. T. AND F. A. BISBY. 1984. A simple method for establishingtaxonomic characters from measurement data. Taxon, 33:405–409.
ARCHIE, J. W. 1985. Methods for coding variable morphological featuresfor numerical taxonomic analysis. Systematic Zoology, 34:326–345.
BAUM, B. R. 1988. A simple procedure for establishing discrete charactersfrom measurement data, applicable to cladistics. Taxon, 37:63–70.
BREMER, K. 1988. The limits of amino acid sequence data in angiospermphylogenetic reconstruction. Evolution, 42:795–803.
BURNHAM, K. P. AND D. R. ANDERSON. 2002. Model Selection andMultimodel Inference (second edition). Springer-Verlag, Inc., NewYork, 488 p.
CHAPPILL, J. A. 1989. Quantitative characters in phylogenetic analysis.Cladistics, 5:219–234.
CHATTERTON, B. D. E., G. D. EDGECOMBE, S. E. SPEYER, A. S. HUNT,AND R. A. FORTEY. 1994. Ontogeny and relationships of Trinucleodea(Trilobita). Journal of Paleontology, 68:523–540.
CHATTERTON, B. D. E. AND R. LUDVIGSEN. 1998. Upper Steptoean(Upper Cambrian) trilobites from the McKay Group of southeasternBritish Columbia, Canada. Paleontological Society Memoir, 49:1–43.
COLLESS, D. H. 1980. Congruence between morphometric and allozymedata for Menidia species: A reappraisal. Systematic Zoology, 29:288–299.
FARRIS, J. 1970. Methods for computing Wagner trees. SystematicZoology, 19:83–92.
FOOTE, M. 1996. On the probability of ancestors in the fossil record.Paleobiology, 22:141–151.
FORTEY, R. A. 1997. Classification, p. 289–302. In R. L. Kaesler (ed.),Treatise of Invertebrate Paleontology. Pt. O. Trilobita (Revised).Geological Society of America and University of Kansas Press,Lawrence.
FORTEY, R. A. AND B. D. E. CHATTERTON. 1988. Classification of thetrilobite suborder Asaphina. Palaeontology, 31:165–222.
GARCIA-CRUZ, J. AND V. SOSA. 2006. Coding quantitative character datafor phylogenetic analysis: A comparison of five methods. SystematicBiology, 31:302–309.
GOGIN, I. YA. 1990. Novye pozdnekembriyskie trilobity Sette-Dabana(New Late Cambrian trilobites of the Sette-Daban). EzhegodnikVsesoyuznogo paleontologicheskogo obshchestva, 33:140–152. (InRussian)
GOLDMAN, N. 1988. Methods for discrete coding of morphologicalcharacters for numerical analysis. Cladistics, 4:59–71.
GOLOBOFF, P. A., J. S. FARRIS, AND K. C. NIXON. 2008. TNT, a freeprogram for phylogenetic analysis. Cladistics, 24:774–786.
GOLOBOFF, P. A., C. I. MATTONI, AND A. S. QUINTEROS. 2006.Continuous characters analyzed as such. Cladistics, 22:589–601.
GRANT, R. E. 1962. Trilobite distribution, upper Franconia Formation(Upper Cambrian), southeastern Minnesota. Journal of Paleontology,36:965–998.
GUERRERO, J. A., E. DE LUNA, AND C. SANCHEZ-HERNANDEZ. 2003.Morphometrics in the quantification of character state identity for theassessment of primary homology: An analysis of character variation ofthe genus Artibeus (Chiroptera: Phyllostomidae). Biological Journal ofthe Linnean Society, 80:45–55.
HALL, J. AND R. P. WHITFIELD. 1877. Paleontology: Fossils of thePotsdam Group. U.S. Geological Exploration 40th Parallel, 4:199–231.
1146 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

HARDY, M. C. 1985. Testing for adaptive radiation: The ptychaspid(Trilobita) biomere of the Late Cambrian, p. 379–397. In J. W.Valentine (ed.), Phanerozoic Diversity Patterns. Princeton UniversityPress.
HOHENSEE, S. R. AND J. H. STITT. 1989. Redeposited Elvinia zone (UpperCambrian) trilobites from the Collier Shale, Ouachita Mountains, west-central Arkansas. Journal of Paleontology, 63:857–879.
HOPKINS, M. J. 2010. Intraspecific variation, its geographic structure, andthe relationship between variation and duration, with examples fromCambrian trilobites and Recent fiddler crabs. Unpublished Ph.D.dissertation, University of Chicago, 375 p.
HOPKINS, M. J. In press. How species longevity, intraspecific morpho-logical variation, and geographic range size are related: A comparisonusing late Cambrian trilobites. Evolution.
HUGHES, N. C. AND A. W. A. RUSHTON. 1990. Computer-aidedrestoration of a Late Cambrian ceratopygid trilobite from Wales, andits phylogenetic implications. Palaeontology, 33:429–445.
HUPE, P. 1953. Classe des trilobites. Traite de Paleontologie, 3:44–246.HURVICH, C. M. AND C.-L. TSAI. 1989. Regression and time series model
selection in small samples. Biometrika, 76:297–307.JELL, P. A. AND J. M. ADRAIN. 2003. Available generic names for
trilobites. Memoirs of the Queensland Museum, 48:331–553.KITCHING, I. J., P. L. FOREY, C. J. HUMPHRIES, AND D. M. WILLIAMS.
1998. Cladistics: The Theory and Practice of Parsimony Analysis(second edition). Systematics Association Publication No 11, 228 p.
KOBAYASHI, T. 1935. The Cambro–Ordovician formations and faunas ofSouth Chosen. Paleontology. Pt. 3: Cambrian faunas of South Chosenwith a special study on the Cambrian trilobite genera and families.Journal of the Faculty of Science, Imperial University of Tokyo(Section II), 4:49–344.
KOBAYASHI, T. 1936. On the Parabolinella fauna from Province Jujuy,Argentina, with a note on the Olenidae. Japanese Journal of Geologyand Geography, 13:85–102.
KOBAYASHI, T. 1938. Upper Cambrian fossils from British Columbia witha discussion of the isolated occurrences of the so-called ‘‘Olenus’’ bedsof Mt. Jubilee. Japanese Journal of Geology and Geography, 15:150–192.
KURTZ, V. E. 1975. Franconian (Upper Cambrian) trilobite faunas fromthe Elvins Group of southeast Missouri. Journal of Paleontology,49:1009–1043.
LAZARENKO, N. P. AND N. I. NIKIFOROV. 1968. Complexes of trilobitesfrom the Upper Cambrian deposits of the Kulumbe River (northwesternSiberian Platform). Uchenye Zapiski, 23:20–80. (In Russian)
LOCHMAN, C. 1938. Upper Cambrian faunas of the Cap MountainFormation of Texas. Journal of Paleontology, 12:72–85.
LOCHMAN, C. 1940. Fauna of the basal Bonneterre Dolomite (UpperCambrian) of southeastern Missouri. Journal of Paleontology, 14:1–53.
LOCHMAN, C. 1950. Upper Cambrian faunas of the Little RockyMountains, Montana. Journal of Paleontology, 24:322–349.
LOCHMAN, C. 1964. Upper Cambrian faunas from the subsurfaceDeadwood Formation, Williston Basin, Montana. Journal of Paleon-tology, 38:33–60.
LOCHMAN, C. AND D. DUNCAN. 1944. Early Upper Cambrian faunasof central Montana. Geological Society of America Special Papers, 54,181 p.
LOCHMAN, C. AND C.-H. HU. 1960. Upper Cambrian faunas from thenorthwest Wind River Mountains, Wyoming, Pt. I. Journal ofPaleontology, 34:793–834.
LOCHMAN, C. AND C.-H. HU. 1961. Upper Cambrian faunas from thenorthwest Wind River Mountains, Wyoming, Pt. II. Journal ofPaleontology, 35:125–146.
LOCHMAN, C. AND C.-H. HU. 1962a. An Aphelaspis zone faunule fromLogan, Montana. Journal of Paleontology, 36:431–441.
LOCHMAN, C. AND C.-H. HU. 1962b. Upper Cambrian faunas from thenorthwest Wind River Mountains, Wyoming, Pt. III. Journal ofPaleontology, 36:1–28.
LOCHMAN-BALK, C. 1970. Upper Cambrian faunal patterns on the craton.Geological Society of America Bulletin, 81:3197–3224.
LOCHMAN-BALK, C. 1974. Late Dresbachian (Late Cambrian) biostratig-raphy of North America. Geological Society of America Bulletin,85:135–140.
LU, Y.-H. AND Y.-Y. QIAN. 1983. New zonation and correlation of theUpper Cambrian Changshanian Stage in North China. Acta Palaeon-tologica Sinica, 22:235–257.
LUDVIGSEN, R. 1991. The median suture of Asaphida: A monophyleticcharacter? The Trilobite Papers, 3:11–13.
LUO, H. 1983. New finds of trilobites from Late Cambrian in westernYunnan, p. 1–30. In Contributions to the Geology of the Qinghai-Xizang (Tibet) Plateau.
MICHEVICH, M. F. AND M. S. JOHNSON. 1976. Congruence betweenmorphological and allozyme data in evolutionary inference andcharacter evolution. Systematic Zoology, 25:260–270.
OSLEGER, D. AND J. F. READ. 1993. Comparative analysis of methodsused to define eustatic variations in outcrop: Late Cambrianinterbasinal sequence development. American Journal of Science,293:157–216.
PALMER, A. R. 1954. The faunas of the Riley Formation in central Texas.Journal of Paleontology, 28:709–786.
PALMER, A. R. 1955. Upper Cambrian Agnostidae of the Eureka District,Nevada. Journal of Paleontology, 29:86–101.
PALMER, A. R. 1956. The type species of Litocephalus Resser, 1937(Trilobita). Journal of Paleontology, 30:608–610.
PALMER, A. R. 1960. Trilobites of the Upper Cambrian DunderbergShale, Eureka District, Nevada. U.S. Geological Survey ProfessionalPaper, 334-C:53–109.
PALMER, A. R. 1962. Glyptagnostus and associated trilobites in the UnitedStates. U.S. Geological Survey Professional Paper, 374-F, 45 p.
PALMER, A. R. 1965a. Biomere: A new kind of biostratigraphic unit.Journal of Paleontology, 39:149–153.
PALMER, A. R. 1965b. Trilobites of the Late Cambrian Pterocephaliidbiomere in the Great Basin, United States. U.S. Geological SurveyProfessional Paper, 493, 105 p.
PALMER, A. R. 1968. Cambrian trilobites of east-central Alaska. U.S.Geological Survey Professional Paper, 559-B, 115 p.
PALMER, A. R. 1979. Biomere boundaries re-examined. Alcheringa, 3:33–41.
PALMER, A. R. 1981. Subdivision of the Sauk sequence, p. 160–162. InM. E. Taylor (ed.), Short Papers for the Second InternationalSymposium on the Cambrian System. U.S. Geological Survey Open-File Report 81-743.
PALMER, A. R. 1982. Biomere boundaries: a possible test for extraterres-trial perturbation of the biosphere, p. 469–476. In L. T. Silver and P. H.Schultz (eds.), Geological Implications of Impacts of Large Asteroidsand Comets on the Earth. Geological Society of America Special Paper190.
PARK, T.-Y. AND D. K. CHOI. 2009. Post-embryonic development of theFurongian (Late Cambrian) trilobites Tsinania canens: implications forlife mode and phylogeny. Evolution and Development, 11:441–455.
PENG, S. 1992. Upper Cambrian biostratigraphy and trilobite faunas ofthe Cili-Taoyuan area, northwestern Hunan, China. Memoirs of theAssociation of Australasian Palaeontologists, 13, 119 p.
PENG, S., J. A. BABCOCK, AND H. LIN. 2004. Polymerid Trilobites fromthe Cambrian of Northwestern Hunan, China. Vol. 2: Ptychopariida,Eodiscida, and undetermined forms. Science Press, Beijing, 355 p.
PRATT, B. R. 1992. Trilobites of the Marjuman and Steptoean stages(Upper Cambrian), Rabbitkettle Formation, southern MackenzieMountains, northwest Canada. Paleontographica Canadiana, No. 9,179 p.
QIAN, Y.-Y. 1994. Trilobites from Middle Upper Cambrian (ChangshanStage) of north and northeast China. Palaeontologia Sinica, Vol. 183,190 p.
QIU, H.-A., Y.-H. LU, Z.-L. ZHU, D.-C. BI, T.-R. LIN, Z.-Y. ZHOU, Q.-Z.ZHANG, Y.-Y. QIAN, T.-Y. JU, N.-R. HAN, AND X.-Z. WEI. 1983.Trilobita, p. 28–254, 574–609, pl. 211–288. In Paleontological Atlas ofEast China, 1, Early Paleozoic Vol. Geological Publishing House,Beijing. (In Chinese)
RAE, T. C. 1998. The logical basis for the use of continuous characters inphylogenetic systematics. Cladistics, 14:221–228.
RESSER, C. E. 1935. Nomenclature of some Cambrian trilobites.Smithsonian Miscellaneous Collections, 93, 46 p.
RESSER, C. E. 1937. Third contribution to nomenclature of Cambriantrilobites. Smithsonian Miscellaneous Collections, 95, 29 p.
RESSER, C. E. 1938. Fourth contribution to nomenclature of Cambrianfossils. Smithsonian Miscellaneous Collections, 97, 43 p.
RESSER, C. E. 1942. New Upper Cambrian trilobites. SmithsonianMiscellaneous Collections, 103, 136 p.
ROEMER, F. 1849. Texas, mit besondered Rucksicht auf deutscheAuswanderung und die physischen Verhaltnisse des Landes nacheigener Beobachtung geschildert, Bonn, 464 p.
ROZOVA, A. V. 1963. Biostratigraphic scheme of the upper and upper-middle Cambrian and new upper Cambrian trilobites. Geologiya iGeofizika, 9:3–20.
SADLER, P. M., W. G. KEMPLE, AND M. A. KOOSER. 2003. Contents ofthe compact disc: CONOP9 Programs for solving the stratigraphiccorrelation and seriation problems as constrained optimization, p. 461–465. In P. J. Harries (ed.), Approaches in High-Resolution StratigraphicPaleontology, Topics in Geobiology, 21. Kluwer Academic Publishers,Dordrecht.
HOPKINS—PTEROCEPHALIID PHYLOGENY 1147

SHAW, A. B. 1957. Quantitative trilobite studies II. Measurement of thedorsal shell of non-agnostidean trilobites. Journal of Paleontology,31:193–207.
SHEETS, H. D. 2003. FMCBox. Canisius College, Buffalo, New York.SOKAL, R. R. AND F. J. ROHLF. 1995. Biometry (third edition). W. H.
Freeman and Co., New York, 887 p.STEVENS, P. F. 1991. Character states, morphological variation, and
phylogenetic analysis: A review. Systematic Botany, 16:553–583.STITT, J. H. 1971a. Cambrian–Ordovician trilobites, western Arbuckle
Mountains. Oklahoma Geological Survey Bulletin, 110, 83 p.STITT, J. H. 1971b. Repeating evolutionary patterns in Late Cambrian
trilobite biomeres. Journal of Paleontology, 45:178–181.STITT, J. H. 1975. Adaptive radiation, trilobite paleoecology, and
extinction, Ptychaspid Biomere, Late Cambrian of Oklahoma. Fossilsand Strata, 4:381–390.
STITT, J. H. 1977. Late Cambrian and earliest Ordovician trilobites,Wichita Mountains area, Oklahoma. Oklahoma Geological SurveyBulletin, 124, 79 p.
STITT, J. H. 1998. Trilobites from the Cedarina dakotaensis zone,lowermost part of the Deadwood Formation (Marjuman State, UpperCambrian), Black Hills, South Dakota. Journal of Paleontology,72:1030–1046.
STITT, J. H. AND P. J. PERFETTA. 2000. Trilobites, biostratigraphy, andlithostratigraphy of the Crepicephalus and Aphelaspis zones, lowerDeadwood Formation (Marjuman and Steptoean stages, UpperCambrian), Black Hills, South Dakota. Journal of Paleontology,74:199–223.
STITT, J. H., J. D. RUCKER, N. D. BOYER, AND W. D. HART. 1994. NewElvinia zone (Upper Cambrian) trilobites from new localities in theCollier Shale, Ouachita Mountains, Arkansas. Journal of Paleontology,68:518–523.
STRAIT, D. S., M. A. MONIZ, AND P. T. STRAIT. 1996. Finite mixturecoding: A new approach to coding continuous characters. SystematicBiology, 45:67–78.
SUN, Y.-C. 1935. The Upper Cambrian trilobites faunas of North China.Palaeontologia Sinica series B, Vol. 7, 93 p.
SWIDERSKI, D. L., M. L. ZELDITCH, AND W. L. FINK. 1998. Whymorphometrics is not special: Coding quantitative data for phylogeneticanalysis. Systematic Biology, 47:508–519.
SWOFFORD, D. L. 1998. PAUP*: Phylogenetic analysis using parsimony*4.0. Sinauer, Sunderland, MS.
THIELE, K. 1993. The holy grail of the perfect character: The cladistictreatment of morphometric data. Cladistics, 9:275–304.
THORPE, R. S. 1984. Coding morphometric characters for constructingdistance Wagner networks. Evolution, 38:244–255.
ULRICH, E. O. 1924. Notes on new names in table of formations and onphysical evidence of breaks between Paleozoic systems in Wisconsin.Transactions of the Wisconsin Academy of Science, Arts, and Letters,21:71–107.
WAGNER, P. J. AND D. H. ERWIN. 1995. Phylogenetic patterns as tests ofspeciation models, p. 87–122. In D. H. Erwin and R. L. Anstey (eds.),New Approaches to studying speciation in the fossil record. ColumbiaUniversity Press, New York.
WALCOTT, C. D. 1884. Paleontology of the Eureka District. U.S.Geological Survey Monographs, 8, 298 p.
WALCOTT, C. D. 1890. Description of new forms of Upper Cambrianfossils. U.S. National Museum Proceedings, 13:267–279.
WALCOTT, C. D. 1912. New York Potsdam-Hoyt fauna. SmithsonianMiscellaneous Collections, 57:251–304.
WALCOTT, C. D. 1916. Cambrian Geology and Paleontology III. No. 5,Cambrian Trilobites. Smithsonian Miscellaneous Collections, 64:303–456.
WALCOTT, C. D. 1924. Cambrian Geology and Paleontology V. No. 2,Cambrian and Ozarkian trilobites. Smithsonian Miscellaneous Collec-tions, 75:53–60.
WESTROP, S. R. 1986. Trilobites of the Upper Cambrian SunwaptanStage, southern Canadian Rocky Mountains, Alberta. Paleontogra-phica Canadiana, No. 3, 179 p.
WESTROP, S. R. 1988. Trilobite diversity patterns in an Upper Cambrianstage. Paleobiology, 14:401–409.
WESTROP, S. R. 1989. Macroevolutionary implications of mass extinction-evidence from an Upper Cambrian stage boundary. Paleobiology,15:46–52.
WESTROP, S. R. 1996. Temporal persistence and stability of Cambrianbiofacies: Sunwaptan (Upper Cambrian) trilobite faunas of NorthAmerica. Palaeogeography, Palaeoclimatology, Palaeoecology, 127:33–46.
WESTROP, S. R. AND J. M. ADRAIN. 2007. Bartonaspis new genus, atrilobite species complex from the base of the Upper CambrianSunwaptan Stage in North America. Canadian Journal of EarthSciences, 44:987–1003.
WESTROP, S. R. AND J. M. ADRAIN. 2009. The late Cambrian (Furongian;Steptoean) trilobite genus Xenocheilos Wilson, 1949: Systematics andbiostratigraphic signficance. Memoirs of the Association of Austral-asian Palaeontologists, 37:351–368.
WESTROP, S. R., J. D. EOFF, T.-W. NG, A. A. DENGLER, AND J. M.ADRAIN. 2008. Classification of the Late Cambrian (Steptoean) trilobitegenera Cheilocephalus Berkey, 1898 and Oligometopus Resser, 1936from Laurentia. Canadian Journal of Earth Sciences, 45:725–744.
WESTROP, S. R. AND R. LUDVIGSEN. 1987. Biogeographic control oftrilobite mass extinction at an Upper Cambrian ‘‘biomere’’ boundary.Paleobiology, 13:84–99.
WESTROP, S. R. AND R. LUDVIGSEN. 2000. The Late Cambrian (Marju-man) trilobites genus Hysteropleura Raymond from the Cow HeadGroup, western Newfoundland. Journal of Paleontology, 74:1020–1030.
WESTROP, S. R., R. A. W. POOLE, AND J. M. ADRAIN. 2010. Systematicsof Dokimocephalus and related trilobites from the Late Cambrian(Steptoean; Millardan and Furongian series) of Laurentian NorthAmerica. Journal of Systematic Palaeontology, 8:545–606.
WHITTINGTON, H. B. 2003. The Upper Cambrian trilobites Plethopeltisand Leiocoryphe: Morphology, paedomorphism, classification. Journalof Paleontology, 77:698–705.
WHITTINGTON, H. B. AND S. R. A. KELLY. 1997. Morphological termsapplied to Trilobita, p. 313–329. In R. L. Kaesler (ed.), Treatise onInvertebrate Paleontology. Pt. O. Arthropoda 1, Trilobita, Revised.Geological Society of America and University of Kansas Press,Lawrence.
WIENS, J. J. 2001. Character analysis in morphological phylogenetics:Problems and solutions. Systematic Biology, 50:689–699.
WILLS, M. A. 1999. Congruence between phylogeny and stratigraphy:Randomization tests and the gap excess ratio. Systematic Biology,48:559–580.
YUAN, J.-L. AND G.-Z. YIN. 1998. New polymerid trilobites from theChefu Formation in early Late Cambrian of eastern Guizhou. ActaPalaeontologica Sinica, 37:138–172.
ZHU, X.-J., N. C. HUGHES, AND S.-C. PENG. 2007. On a new species ofShergoldia Zhang & Jell, 1987 (Trilobita), the family Tsinaniidae andthe order Asaphida, p. 243–353. In J. R. Laurie and J. R. Paterson(eds.), Papers in Honour of John H. Shergold 1938–2006. Memoirs ofthe Association of Australasian Palaeontologists, 34.
ACCEPTED 22 MAY 2011
1148 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

AP
PE
ND
IX1—
Ch
ara
cter
su
sed
inp
hy
log
enet
ica
naly
sis.
No
.C
hara
cter
des
crip
tio
n
Ch
ara
cter
state
des
crip
tio
n
01
23
4
1.S
ha
pe
of
cra
nid
ial
bo
rder
inla
tera
lv
iew
.No
tco
nca
ve
.Co
nca
ve
..
.
2.E
xp
ress
ion
of
an
teri
or
bo
rder
furr
ow
on
cra
nid
ium
.Dis
tin
cta
nd
of
ap
pro
xim
ate
lyco
nst
an
td
epth
.Wel
ld
efin
edb
ut
shall
ow
.Sh
all
ow
an
dp
oo
rly
def
ined
..
3.W
idth
of
cra
nid
ial
bo
rder
.Gen
era
lly
con
sist
ent
inw
idth
.Dis
tin
ctly
wid
erab
axia
lly
..
.4
.Sh
ap
eo
fcr
an
idia
la
nte
rio
rm
arg
in.E
ven
lycu
rved
.An
gu
lati
on
or
acu
min
ate
..
.
5.S
ha
pe
of
cra
nid
ial
an
teri
or
bo
rder
furr
ow
.Even
lycu
rved
.An
gu
lati
on
.Po
ster
ior
med
ian
inb
end
..
6.D
isti
nct
orn
am
enta
tio
na
lon
gcr
an
idia
lb
ord
erex
pre
ssed
on
inte
rnal
mo
ld
.Ab
sen
t.P
rese
nt
..
.
7.S
ha
pe
of
pre
gla
bel
lar
fiel
din
late
ral
vie
w.N
ot
con
vex
.Co
nv
ex.
..
8.O
rna
men
tati
on
on
cra
nid
ium
.Sm
oo
th.P
itte
d.G
ran
ula
r.
.
9.C
aec
ao
nfr
on
tal
are
a.A
bse
nt
.Pre
sen
t.
..
10
.Ter
race
lin
eso
nb
ord
er.A
bse
nt
.On
ma
rgin
.Acr
oss
bo
rder
..
11
.Sh
ap
eo
fa
xia
lfu
rro
ws
(do
rsa
lv
iew
).S
tra
igh
t(n
ot
curv
ed)
.No
tica
bly
con
cave
bet
wee
np
alp
ebra
llo
bes
(cu
rved
abax
ially)
..
.
12
.Ori
enta
tio
no
fa
xia
lfu
rro
ws
an
teri
or
top
alp
ebra
llo
bes
(la
tera
lv
iew
)
.No
tn
oti
cab
lyd
ow
nsl
op
ing
.Do
wn
slo
pin
g(c
urv
edst
ron
gly
ven
tra
lly)
..
.
13
.Gla
bel
lar
furr
ow
s.S
tro
ng
lyex
pre
ssed
.On
lyp
ost
erio
r-m
ost
stro
ngl
yex
pre
ssed
.All
wea
kly
exp
ress
ed.E
ffa
ced
enti
rely
.1
4.L
oca
tio
no
fa
nte
rio
ren
do
fp
alp
ebra
llo
be
.Do
esn
ot
tou
cha
xia
lfu
rro
wa
nte
rio
rly
.To
uch
esa
xia
lfu
rro
wa
nte
rio
rly
..
.
15
.Po
siti
on
of
mid
len
gth
of
pa
lpeb
ral
lob
ere
lati
ve
tog
lab
ella
.Dis
tin
ctly
an
teri
or
tog
lab
ella
rm
idle
ng
th.O
pp
osi
teo
rp
ost
erio
rto
gla
bel
lar
mid
len
gth
..
.
16
.Palp
ebra
llo
be
sha
pe
.Bu
lbo
us,
cyli
nd
rica
l.F
latt
ened
..
.1
7.D
efin
itio
no
fp
alp
ebra
llo
be
.Po
orl
yd
efin
edb
yp
alp
ebra
lfu
rro
ws
.Wel
ld
efin
edb
yp
alp
ebra
lfu
rro
ws
..
.1
8.S
ha
pe
of
pa
lpeb
ral
furr
ow
.Rel
ati
vel
yst
raig
ht
.Arc
ua
te.
..
19
.Ocu
lar
rid
ges
.Pre
sen
t(s
tro
ng
lyex
pre
ssed
).A
bse
nt
(bu
tm
ayb
evi
sib
leo
nin
tern
alm
old
).
..
20
.Dir
ecti
on
of
ocu
lar
rid
ges
.Ori
ente
dtr
an
sver
sely
.Ori
ente
dp
ost
ero
-la
tera
lly
..
.2
1.C
ou
rse
of
faci
al
sutu
res
infr
on
to
fg
lab
ella
.In
tram
arg
ina
lo
nly
.Su
bm
arg
inal
ad
jace
nt
tov
entr
al
ma
rgin
al
sutu
re.
..
22
.Sh
ap
eo
ffi
xed
chee
ks
(in
tero
cula
rare
a)
inan
teri
or
or
po
ster
ior
vie
w.N
earl
yh
ori
zon
tal
.Up
slo
pin
ga
wa
yfr
om
gla
bel
la.
..
23
.Occ
ipit
al
rin
g.D
efin
edb
yd
eep
furr
ow
.Def
ined
by
shall
ow
furr
ow
(part
icu
larl
yn
ear
axia
lfu
rro
w)
..
.
24
.Occ
ipit
al
tub
ercl
e.A
bse
nt
.Po
orl
yd
evel
op
ed(e
.g.,
on
lyv
isib
leo
nin
tern
al
mo
ld)
.Wel
ld
evel
op
ed.
.
25
.Gen
al
spin
e.P
rese
nt
.Ab
sen
t.
..
26
.Exp
ress
ion
of
furr
ow
so
nli
bri
gen
a.P
ost
erio
rb
ord
erfu
rro
wd
oes
no
tco
nti
nu
est
ron
gly
on
toge
nal
spin
e.F
urr
ow
curv
esb
ack
wa
rds
on
tob
ase
of
gen
al
spin
e,p
art
icu
larl
yp
ost
erio
rb
ord
erfu
rro
w
.Po
ster
ior
bo
rder
furr
ow
an
dla
tera
lb
ord
erfu
rro
wd
on
ot
mee
t
..
27
.Orn
am
enta
tio
no
ng
ena
lsp
ine
.Sm
oo
th(e
xce
pt
for
terr
ace
lin
eso
nm
arg
in)
.Ter
race
lin
esa
cro
sssp
ine
.Gra
nu
lati
on
acr
oss
spin
e.
.
28
.Ven
tra
lm
edia
nsu
ture
.Ab
sen
t.P
rese
nt
..
.2
9.P
yg
idiu
ma
nk
ylo
sed
wit
hp
ost
erio
rmo
stth
ora
cic
seg
men
t.A
bse
nt
.Co
mm
on
lyp
rese
nt
..
.
30
.Sh
ap
eo
fp
yg
idiu
m.S
ub
qu
ad
rate
.Sem
icir
cula
r.T
ran
sver
sely
sub
ov
ate
..
HOPKINS—PTEROCEPHALIID PHYLOGENY 1149

AP
PE
ND
IX1—
Co
nti
nu
ed.
No
.C
hara
cter
des
crip
tio
n
Ch
ara
cter
state
des
crip
tio
n
01
23
4
31
.Gre
ate
stw
idth
of
py
gid
ium
.An
teri
or
pa
rto
fa
xis
.Po
ster
ior
pa
rto
fa
xis
.Po
ster
ior
toa
xis
..
32
.Nu
mb
ero
fd
isti
nct
rin
gfu
rro
ws
beh
ind
art
icu
lati
ng
furr
ow
s.U
pto
two
.Up
toth
ree
.Up
tofo
ur
.Up
tofi
ve
.Up
tose
ven
33
.Py
gid
ial
spin
es.M
arg
ina
lsp
ines
ab
sen
t.S
pin
esp
rese
nt,
each
ass
oci
ate
dw
ith
ap
art
icu
lar
seg
men
t.S
pin
esp
rese
nt
bu
tn
ot
ass
oci
ate
dw
ith
pa
rtic
ula
rse
gm
ents
..
34
.Py
gid
ial
an
teri
or
ma
rgin
at
fulc
rum
.Sm
oo
thly
curv
eda
cro
ssfu
lcru
m.A
ng
ula
ra
tfu
lcru
m.
..
35
.Py
gid
ial
an
teri
or
ma
rgin
at
po
ster
ior
bo
rder
.Sm
oo
thly
curv
eda
tm
eeti
ng
wit
hb
ord
er.A
ng
ula
ra
tb
ord
er.
..
36
.Py
gid
ial
bo
rder
furr
ow
.Pre
sen
t.A
bse
nt
(bo
rder
def
ined
by
orn
am
enta
tio
no
rp
leu
ral
furr
ow
so
rch
an
ge
insl
op
e)
..
.
37
.Nu
mb
ero
fst
ron
gly
def
ined
ple
ura
lfu
rro
ws
.Up
totw
o.U
pto
thre
e.U
pto
fou
r.U
pto
fiv
e.
38
.Ple
ura
lfu
rro
ws
.Exte
nd
on
tob
ord
er.E
ith
erd
on
ot
rea
chb
ord
ero
rd
on
ot
exte
nd
on
toit
..
.
39
.Py
gid
ial
po
sta
xia
lri
dg
e.A
bse
nt
.Pre
sen
t.
..
40
.Sh
ap
eo
fp
yg
idia
lp
ost
erio
rb
ord
er.T
hin
,ev
enb
an
d.T
hic
k,
even
ba
nd
.Ex
pa
nd
ing
late
rall
y.E
xp
an
din
gp
ost
erio
rly
into
lob
eso
rsp
ines
.
41
.Py
gid
ial
bo
rder
no
tch
.Ab
sen
t.G
entl
ein
ben
d.M
ediu
min
ben
d.D
eep
inb
end
.4
2.O
rna
men
tati
on
on
py
gid
ium
.Sm
oo
th.P
itte
d.G
ran
ula
r.
.4
3.T
erra
celi
nes
on
do
rsa
lsi
de
of
py
gid
ium
.Ab
sen
t(m
ay
be
pre
sen
to
nly
on
ven
tra
lsi
de
of
bo
rder
).A
lon
gp
ost
erio
rm
arg
in.P
rese
nt
acr
oss
do
rsa
lsi
de
of
bo
rder
..
44
.‘A
lae’
on
an
tero
-late
ral
ma
rgin
of
py
gid
ium
.Ab
sen
t.E
xp
ress
eda
su
nev
enly
curv
edp
ost
erio
rb
ord
erT
rue
‘ala
e’d
evel
op
ed.
.
1150 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

Characters
Taxa 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
1 0 0 1 0 0 0 1 0,1 0 1 0 0 2 0 1 0 1 1 1 1 0 1 0 2 0 1 0 02 0 0 1 1 0 0 0 0,1 0 1 0 0 2 0 1 0 1 1 0 0 0 1 0 1,2 0 1 0 ?3 0 0 1 0 0 0 1 0,1 0 1 0 0 2 0 1 0 1 1 0 1 0 0 0 2 0 1 0 04 1 0 1 1 0 1 ? 2 0 1 0 0 1 0 1 0 1 1 1 1 0 1 0 2 0 0 2 ?5 1 0 1 1 0 1 1 1 0 ? 0 0 1 0 1 0 1 0 0 1 0 1 0 2 ? ? ? ?6 1 0 1 1 2 1 1 2 0 1 0 0 2 0 1 0 1 1 0 1 0 0 0 1 0 0 2 ?7 1 0 1 1 0 ? 1 2 0 1 0 0 1 0 1 0 1 0 0 1 0 1 0 1,2 0 0 2 ?8 1 0 1 1 0 1 ? 0 0 1 0 0 1 0 1 0 1 1 1 1 0 1 0 2 0 0 0 19 1 1 0 1 0 1 1 2 0 ? 0 0 2 0 1 0 1 1 1 1 0 1 0 2 0 0 ? ?10 1 0 1 1 0 ? 0 1 0 ? 0 0 2 0 1 0 1 1 1 1 0 1 0 2 0 0 ? ?11 1 0 1 1 0 ? 0 2 0 ? 0 0 2 0 1 0 1 1 1 1 0 0 ? ? 0 0 ? ?12 1 0 1 1 0 1 1 2 0 1 0 0 2 0 1 0 1 0,1 0 1 0 1 0 1 0 0 2 113 0 0 1 0 2 0 1 1 0 1 0 0 2 0 1 0 0 1 1 1 0 1 0 1 0 0 ? ?14 0 1 1 0 0 0 0 1 0 1 0 0 2 0 1 0 1 1 1 ? 0 0 0 1 0 0 0 015 0 0 1 0 2 0 0 0 0 1 0 0 2 0 1 0 1 1 1 1 0 1 0 1 0 0 1 ?16 0 2 1 1 0 0 0 1 0 1 0 1 3 1 0 1 0 0 NA NA ? 1 1 2 0 2 1 ?17 0 2 1 1 0 0 0 0 0 0 0 1 3 1 0 1 0 0 NA NA ? 0 0 1 1 0 NA 118 0 2 1 1 0 0 0 0 0 0 0 1 3 1 0 1 0 0 NA NA 0 1 0 0 1 0 NA 119 0 1 1 1 0,1 0 0 1 0 0 1 0 2 0 0 0 0 0 1 ? ? 1 0 2 0 2 1 ?20 0 1 1 1 0 0 0 0 0 0 1 0 2 0 0 0 0 0 1 ? ? 1 0 2 0 ? 1 ?21 0 0 1 1 2 0 0 0,1 0 1 0 0 2 0 0 1 0 0 1 ? 0 0 1 1 0 0 1 122 0 2 1 0 0 0 0 0 0 1 0 1 2 0 0 1 0 0 0,1 1 0 0 1 1 0 0 1 ?23 0 0 1 0 0 0 0 2 0 1 0 0 2 0 0 1 0 0 0 1 0 0 1 1 0 0 ? ?24 0 0 1 1 2 0 0 1 0 1 0 0 2 0 0 1 0 0 1 1 0 0 1 1 0 0 1 ?25 0 0 1 0 0 0 0 1 0 1 0 0 2 0 0 0 1 0 0 0 0 0 1 1 0 0 0 ?26 0 0 1 0,1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 ? 0 1 1 2 0 0 1 ?27 1 2 0 0 0 0 0 0,2 1 1 0 0 1 0 1 0 1 1 0 1 1 1 0 2 0 0 1 128 1 2 0 1 0 0 0 ? ? 0 0 0 0 0 1 0 1 1 0 1 1 0 0 2 0 0 ? 129 1 2 0 0 0 0 0 0,2 0 1 0 0 1 0 1 0 1 1 0 1 1 1 0 1 0 1 1 130 1 2 0 0 2 ? 0 0 1 1 0 0 0 0 1 0 1 1 0 1 1 0 0 0 0 0 1 131 1 2 0 0 2 0 0 2 0 ? 0 0 1 0 1 0 1 1 0 0 1 1 0 ? ? ? ? ?32 1 2 0 0 0 0 0 2 0 2 0 0 0 0 1 0 1 1 0 0 1 1 0 2 0 0 1 133 1 2 0 0 0 ? 0 ? ? ? 0 0 2 0 0 ? 1 ? 1 1 1 1 0 1 ? ? ? ?34 1 1 1 0 2 0 0 ? ? 0 0 0 2 0 1 0 1 1 1 1 ? 1 0 ? 0 1 1 ?35 1 1 1 0 0 0 0 2 1 1 0 0 2 0 1 1 1 1 1 1 ? 1 0 2 0 1 1 ?36 1 1 1 0 0 0 0 2 0,1 2 0 0 2 0 1 1 1 1 1 0 ? 1 0 1 0 1 1 137 1 1 1 0 0 0 0 2 1 2 0 0 2 0 1 1 1 1 1 1 1 1 0 1,2 0 1 1 ?38 1 1 1 1 2 0 1 2 1 1 0 0 0 0 1 0 1 0,1 1 1 ? 1 0 2 0 1 1 ?39 1 2 1 1 0 0 0 2 0,1 1 0 0 0 0 0 1 0 0 1 1 0 0 0 1 0 1 1 140 1 2 1 0 0 0 0 2 1 1 0 0 0,2 0 0 0 1 0 0 1 0 0 0 2 0 1 ? ?41 1 2 0 0 0 0 0 2 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 1 142 0 0 1 1 0 ? 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 1 2 2 0 2 0 ?
APPENDIX 2—Continuous characters (log[x + 1] 2 transformed values).
HOPKINS—PTEROCEPHALIID PHYLOGENY 1151

Characters FMC coding Gap-weighted coding
Taxa 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 48 49 50 45 46 47 48 49 50 51 52
1 0 2 0 1 0 1 1 1 1 1 0 0 1 0 ? 0 1,2 0 0 0 8 0 25 0 2 9 26 32 0 0 1 1 0 0 0 1 1 1 1 3 1 0 3 0 1,2 0 0 0 12 8 27 4 5 11 11 83 0 2 0 1 0 1 1 1 1 1 0 0 1 0 1 0 1,2 0 0 0 8 6 23 3 2 6 28 64 0 1 1 2 0 1 1 1 2 0 0 2 0 2 ? 0 2 0 0 0 21 9 7 9 10 10 16 115 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 2 0 0 0 16 8 18 5 0 1 ? ?6 0 2 1 0 0 1 0 1 0 0 0 2 0 2 1 0 1,2 0 0 0 12 8 20 6 6 8 26 37 0 1 1 1 0 1 1 1 1 0 0 2 0 2 1 0 2 0 0 0 15 8 12 5 7 10 11 138 0 2 1 2 0 0 1 1 1 0 0 2 0 0 ? 0 2 0 0 0 27 1 25 3 6 6 28 139 0 1 2 1 0 1 0 1 1 0 1 1 0 2 ? 0 2 0 0 0 17 6 15 8 10 8 16 410 0 1 1 2 0 1 1 1 1 0 0 2 0 1 1 0 2 0 0 0 16 2 21 3 4 5 23 1511 0 2 0 2 0 ? ? 1 1 ? ? 1 ? ? ? 0 2 0 0 0 22 6 29 1 0 8 ? ?12 0 2 1 0 0 0 1 1 0 0 0 2 0 2 1 0 2 0 0 0 17 6 15 2 4 11 24 1213 0 0 0 0 2 0 1 1 0 1 0 4 1 1 3 0 1,2 0 0 0 10 6 19 4 5 7 4 1414 0 0 0 1 2 0 1 1 0 0 0 4 1 1,2 1 0 1,2 0 0 0 11 3 26 5 5 5 9 715 0 1 0 0 2 0 1 1 0 1 0 4 1 2 3 0 1 0 0 0 8 9 18 6 6 9 4 316 1 1 1 3 0 0 0 0 1 1 1 1 0 1 2 0 1,2 1 1 0 10 12 0 28 30 3 10 1117 1 1 1 3 0 0 0 0 0 1 0 1 0 0 0 0 1,2 1 1 0 12 30 7 30 28 9 21 618 1 1 1 2 0 0 0 0 1 1 0 1 0 0 3 0 2 1 1 0 14 21 6 28 26 8 18 419 0 2 1 3 0 1 1 1 1 1 0 1 0 2 1 0 2 0,1 0 0 16 13 10 14 11 2 18 820 0 2 1 3 0 0 1 1 1 1 0 1 0 0 2 0 2 0,1 0 0 18 20 17 13 9 8 13 1221 0 2 0 2 0 1 0 1 0 1 0 1 1 1 1 2 1,2 0 0,1 0 9 9 6 11 11 2 27 522 0 2 1 2 0 1 0 1 1 1 1 1 0 0 ? 0 1,2 0 0 0 13 7 9 8 7 6 19 823 0 2 0 1 0 1 0 1 1 0 0 1 1 2 0 0 1 0 0 0 8 9 9 11 11 2 26 424 0 2 0 1 0 1 0 1 1 1 1 1 1 1 1 1 1,2 0 0 0 9 13 10 9 9 5 30 425 0 2 1 2 0 0 0 1 1 1 0 1 1 1 1 1 1,2 0 0 0 10 6 10 3 5 5 28 626 0 2 1 2 0 0 0 1 2 1 0 1 0 0 1 0 1,2 1 0,1 0 10 11 12 15 12 0 24 827 0 0 1 3 0 0 1 1 1 0 0 2 1 0 2 0 2 0 0 0,1 25 10 23 5 9 17 7 2328 0 2 2 2 0 0 1 1 3 0 1 2 1 0 2 0 2 1 0 0 29 16 21 14 15 12 18 1429 0 0 2 1 0 0 0 1 2 0 1 3 1 0 2 0 2 0 0 0,1 25 9 16 6 12 17 0 1630 0 2 1 4 0 0 0 1 3 0 0 1 0 0 2 0 2 0 0 1 28 21 6 2 9 30 11 3031 0 0 1 2 0 0 1 1 1 0 1 2 1 2 2 0 2 0 0 0 26 10 20 6 9 12 18 1332 0 0 1 4 0 0 1 1 3 0 0 2 1 2 2 0 2 0 0 1 30 16 25 4 10 24 6 2533 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 0 0 0 ? 19 17 10 ? ? ? ?34 0 0 1 1 0 0 1 1 1 0 1 2 1 0 2 0 2 0 0 0 17 10 30 5 8 14 4 1135 0 2 1 3 1 0 0 1 2 0 1 2 0 0 2 0 2 0 0 0 18 7 28 4 6 10 16 1836 0 0 2 2 0 0 1 1 2 0 1 2 1 0 2 0 2 0 0 0 22 9 20 5 6 10 10 2537 0 0 1 3 0 0 0 1 2 1 0 2 0 0 2 0 2 0 0 0 21 4 26 2 5 9 10 2238 0 2 1 2 1 0 0 1 2 0 1 2 0 0 2 0 2 0 0 0 17 11 19 7 4 5 17 1639 0 0 2 1 0 0 1 1 1 0 1 3 3 0,2 2 0 2 0 0 0 19 8 17 6 5 8 4 1140 0 2 1 1 0 0 1 1 1 1 0 2 0,1 2 2 0 2 0 0 0 17 3 9 7 9 5 16 941 0 0 1 0 0 0 1 1 1 1 0 3 2 2 2 0 2 0 0 0 17 8 11 7 6 5 8 1142 0 2 1 0 0 1 0 0 4 1 0 0 0 0 ? 0 0 0 0 0 0 5 10 7 7 4 23 0
APPENDIX 2—Extended.
1152 JOURNAL OF PALEONTOLOGY, V. 85, NO. 6, 2011

Continuous characters
45 46 47 48 49 50 51 52
0.137–0.159 0.208–10.219 0.165–0.182 0.184–0.195 0.342–0.361 0.111–0.131 0.496–0.499 0.303–0.3410.158–0.189 0.228–0.244 0.166–0.193 0.202–0.221 0.359–0.386 0.122–0.136 0.377–0.408 0.334–0.3690.134–0.159 0.220–0.242 0.155–0.180 0.196–0.213 0.344–0.364 0.104–0.125 0.512–0.521 0.302–0.371
0.227 0.236 0.117 0.243 0.405 0.126 0.427 0.370.172–0.219 0.227–0.246 0.137–0.166 0.214–0.224 0.312–0.374 0.082–0.110 ? ?0.162–0.189 0.227–0.245 0.142–0.171 0.219–0.233 0.368–0.388 0.114–0.128 0.473–0.488 0.316–0.3310.188–0.198 0.225–0.245 0.124–0.144 0.214–0.232 0.365–0.390 0.118–0.134 0.463–0.496 0.378–0.3900.265–0.270 0.197–0.233 0.146–0.201 0.194–0.224 0.365–0.398 0.096–0.132 0.479 0.3870.218–0.219 0.219–0.242 0.127–0.162 0.237–0.245 0.383–0.424 0.116–0.122 0.438 0.3250.187–0.204 0.210–0.226 0.157–0.168 0.199–0.214 0.361–0.369 0.108–0.113 0.470–0.479 0.379–0.417
0.235 0.23 0.185 0.192 0.341 0.119 ? ?0.193–0.219 0.223–0.238 0.134–0.153 0.197–0.213 0.363–0.375 0.124–0.136 0.486 0.3750.146–0.179 0.221–0.240 0.130–0.180 0.205–0.227 0.358–0.389 0.106–0.128 0.313–0.371 0.336–0.4210.151–0.177 0.214–0.232 0.161–0.192 0.214–0.227 0.366–0.387 0.102–0.116 0.379 0.3440.145–0.154 0.229–0.246 0.136–0.167 0.223–0.226 0.377–0.378 0.111–0.133 0.318–0.353 0.328–0.3650.146–0.170 0.237–0.253 0.093–0.102 0.331–0.407 0.497–0.571 0.102–0.104 0.363–0.404 0.361–0.3820.171–0.173 0.278–0.311 0.094–0.140 0.357–0.400 0.499–0.571 0.118–0.128 0.456–0.464 0.307–0.367
0.189 0.27 0.117 0.369 0.511 0.119 0.420–0.461 0.316–0.3360.183–0.215 0.241–0.255 0.117–0.135 0.270–0.289 0.400–0.427 0.094–0.108 0.433 0.350.195–0.222 0.244–0.290 0.136–0.165 0.257–0.288 0.382–0.418 0.116–0.125 0.402 0.3750.143–0.171 0.229–0.247 0.105–0.128 0.244–0.260 0.388–0.430 0.091–0.111 0.490–0.523 0.314–0.3480.157–0.200 0.210–0.255 0.088–0.163 0.232–0.253 0.361–0.410 0.112–0.121 0.446–0.458 0.335–0.3670.140–0.158 0.237–0.240 0.114–0.134 0.256–0.262 0.394–0.427 0.092–0.107 0.502–0.503 0.308–0.3500.140–0.165 0.227–0.270 0.112–0.144 0.241–0.283 0.381–0.414 0.100–0.117 0.509–0.549 0.307–0.3440.150–0.167 0.224–0.240 0.120–0.138 0.224 0.355–0.387 0.103–0.118 0.487–0.536 0.321–0.3610.141–0.179 0.226–0.262 0.118–0.152 0.273–0.293 0.385–0.545 0.085–0.103 0.488 0.350.239–0.273 0.220–0.259 0.150–0.186 0.211–0.227 0.372–0.417 0.129–0.166 0.363–0.364 0.428–0.475
0.283 0.256 0.161 0.276 0.434 0.13 0.442 0.3930.243–0.266 0.224–0.255 0.131–0.161 0.225–0.242 0.402–0.437 0.132–0.163 0.294–0.326 0.389–0.4200.271–0.275 0.259–0.282 0.112–0.120 0.195–0.212 0.389–0.407 0.178–0.200 0.357–0.425 0.494–0.4950.253–0.269 0.241 0.16 0.220–0.238 0.401 0.132 0.443 0.383
0.287 0.245–0.269 0.168–0.178 0.208–0.216 0.409 0.169 0.338–0.375 0.419–0.509? 0.266 0.15 0.254 ? ? ? ?
0.206 0.242 0.189 0.221 0.39 0.138 0.343 0.3740.197–0.225 0.221–0.243 0.171–0.194 0.218–0.224 0.367–0.393 0.119–0.135 0.415–0.435 0.397–0.4400.232–0.243 0.221–0.252 0.153–0.162 0.216–0.232 0.369–0.392 0.120–0.133 0.372–0.388 0.417–0.5020.211–0.247 0.204–0.246 0.161–0.192 0.173–0.226 0.356–0.396 0.110–0.139 0.369–0.393 0.404–0.4830.190–0.213 0.233–0.252 0.142–0.168 0.223–0.233 0.352–0.380 0.100–0.120 0.415–0.453 0.391–0.4180.199–0.240 0.222–0.248 0.127–0.169 0.203–0.237 0.359–0.390 0.111–0.128 0.308–0.365 0.353–0.3860.182–0.224 0.222–0.223 0.116–0.136 0.226–0.247 0.392–0.411 0.109–0.113 0.411–0.450 0.349–0.3750.191–0.224 0.227–0.246 0.119–0.145 0.225–0.243 0.368–0.393 0.104–0.119 0.355–0.381 0.347–0.3930.079–0.115 0.209–0.243 0.120–0.134 0.230–0.242 0.374–0.396 0.100–0.115 0.477 0.301
APPENDIX 2—Extended. Continued.
APPENDIX 3—Range of values associated with character states coded using FMC. Corresponding log(x+1)-transformed value in parentheses.
No. Character description
Values at each character state
0 1 2
45 .Length (sag.) of border/length (sag.) of frontalarea of glabella x,0.303 (0.115) 0.303 (0.115).x.0.472 (0.168) x.0.472 (0.168)
48 .Width (tr.) of glabella/width (tr.) of cranidium atanterior end of palpebral lobes x,0.841 (0.265) x.0.841 (0.265)
.
49 .Length (sag.) of cranidium/width (tr.) ofcranidium at anterior end of palpebral lobes x,1.818 (0.450) x.1.818 (0.450)
.
50 .Length (sag.) of frontalarea/length (sag.) of cranidium x,0.413 (0.150) x.0.413 (0.150)
.
APPENDIX 4—Range of values associated with character states coded using gap-weight coding. Corresponding log(x+1)-transformed value in parentheses.
No. Character description
Character state description
Value coded as 0 Value coded as 30
45 .Length (sag.) of border/length (sag.) of frontal area of glabella 0.251 (0.097) 0.935 (0.287)46 .Width (tr.) of glabella/length (sag.) of glabella 0.636 (0.214) 0.971 (0.295)47 .Length of palpebral lobe/length (sag.) of glabella 0.251 (0.097) 0.546 (0.189)48 .Width (tr.) of glabella/width (tr.) of cranidium at anterior end of palpebral lobes 0.547 (0.190) 1.394 (0.379)49 .Length (sag.) of cranidium/width (tr.) of cranidium at anterior end of palpebral lobes 1.194 (0.341) 2.421 (0.534)50 .Length (sag.) of frontal area/length (sag.) of cranidium 0.242 (0.094) 0.545 (0.189)51 .Width (tr.) of pygidium/length (sag.) of pygidium 1.042 (0.310) 2.382 (0.529)52 .Length (sag.) of pygidial axis/width (tr.) of pygidial axis 1.000 (0.301) 1.621 (0.419)
HOPKINS—PTEROCEPHALIID PHYLOGENY 1153