-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
1/19
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
2/19
1111
SPECIES LIMITS IN ANTBIRDS (THAMNOPHILIDAE):
THE WARBLING ANTBIRD (HYPOCNEMIS CANTATOR) COMPLEXM L. I,1,3P R.
I,1 B M. W2
1Department of Vertebrate ZoologyBirds, National Museum of
Natural History, Smithsonian Institution,P.O. Box 37012,
Washington, D.C. 20013, USA; and
2Museum of Natural Science, Louisiana State University, 119
Foster Hall, Baton Rouge, Louisiana 70803, USA
A.Six populations of Warbling Antbird(Hypocnemis cantator)
currentlyconsidered subspecies are more appropriately recognized as
species, given themultiplicity of vocal differences that
distinguishes them. These vocal differencesare as great or greater
than vocal differences documented between syntopic spe-cies-pairs
in the family Thamnophilidae (Isler et al. 1998). Evidence is also
providedthat two forms currently considered subspecies are syntopic
and that two othersare parapatric without apparent physical
barriers. Vocally, the pair of syntopic
taxa were distinguished dramatically, but solely, by common
calls. The finding thatcommon calls were as diversified as songs in
this species-group suggests that callsmay be as important as songs
in reproductive isolation. Future field studies shouldtherefore
focus on clarifying the function of different types of thamnophilid
vocal-izations and elucidating their role in speciation. This is
the first contribution to amultifaceted analysis of the
vocalizations and genetic structure of typical
antbirds(Thamnophilidae) in Amazonia. In future work, results of
the vocal analysis will becompared with a parallel genetic study,
aer which the two analyses will be inte-grated to suggest a
phylogeny. The present study and preliminary studies of
geneticdifferentiation (e.g., Bates et al. 1999) make it clear that
the H. cantatorcomplex hashad a long evolutionary history that
produced greater species-level diversity thanwas recognized
previously. Received 15 January 2005, accepted 12 February
2006.
Key words: biodiversity, Hypocnemis, Neotropics, speciation,
systematics,
Thamnophilidae, vocalizations.
Lmites de Especies en Thamnophilidae: El Complejo de Hypocnemis
cantator
R.Teniendo en cuenta las mltiples diferencias en vocalizaciones
quedistinguen a seis poblaciones deHypocnemis cantatorque
actualmente son consideradascomo subespecies, es ms adecuado
reconocerlas como especies diferentes. Estasdiferencias en
vocalizaciones son iguales o mayores que las documentadas
entrepares de especies sintpicas en la familia Thamnophilidae
(Isler et al. 1998). Tambinse provee evidencia de que dos formas
actualmente consideradas subespecies sonsintpicas, y que otras dos
son paraptricas, sin barreras fsicas evidentes. Vocalmente,el par
de taxones sintpicos se diferenciaron dramticamente, pero nicamente
en susllamadas comunes. El hallazgo de que las llamadas comunes
estn tan diferenciadascomo los cantos en este grupo de especies
sugiere que las llamadas pueden ser tanimportantes como los cantos
en el aislamiento reproductivo. Por lo tanto, estudiosde campo
futuros deberan enfocarse en aclarar la funcin de los distintos
tipos devocalizaciones de los Thamnophilidae, y en elucidar el
papel que stos juegan en laespeciacin. Esta es la primera
contribucin de un anlisis con mltiples facetas de
The Auk124(1):1128, 2007 The American Ornithologists Union,
2007.Printed in USA.
3E-mail: [email protected]
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
3/19
I, I, W12 [Auk, Vol. 124
V a relevant yard-stick (sensuMayr and Ashlock 1991) for
esti-mating reproductive isolation and species statusof sympatric
and allopatric populations of sub-oscine passerines (Isler et al.
1998, Johnson etal. 1999, Baptista and Kroodsma 2001, Helbig etal.
2002, Remsen 2005), though cautions regard-ing individual variation
and clinality apply
(Isler et al. 2005). Recent studies of vocaliza-tions of the
suboscine family Thamnophilidaehave documented extensive,
geographically
based differentiation within some species (e.g.,Isler et al.
1997, 1999, 2001; Whitney et al. 2000;Isler and Isler 2003).
Parallel to these analyses,molecular studies of antbirds have
revealedhigher-than-expected levels of intraspecificgenetic
structure (Capparella 1988; Hacke andRosenberg 1990; Hacke 1993;
Brumfield andCapparella 1996; Bates et al. 1999; Bates 2000,2002).
Questions remain, however, about theextent to which species
diversity as expressed
by vocal characters is consistent with
geneticdifferentiation.
Over a period of nearly two decades, wecollected recordings of
vocalizations of theWarbling Antbird (Hypocnemis cantator)
fromdiverse parts of its extensive range and notedsubstantial
geographically related differencesamong vocalizations. More
recently, explor-atory molecular studies (Bates et al. 1999;Bates
2000, 2002) have revealed high levels ofregional genetic
differentiation among somepopulations of H. cantator. General
concordancein these results led to the inclusion of H. canta-tor in
an ongoing study of thamnophilid dif-ferentiation in Amazonia in
which vocal andmolecular character sets are being developedfor
populations of five widespread species-groups. Vocal and molecular
characters will beemployed independently to derive estimates ofthe
most appropriate taxonomy of each group.These results will be
compared, an effort will
be made to understand the basis for any dis-crepancies between
them, and an estimate of
phylogeny will be constructed from both datasets. Ultimately,
the results of the five studieswill be integrated to provide tests
of hypothesesof historical processes leading to avian
diversi-fication in the Amazon basin.
Although an initial step within this long-termprogram, the
results of the vocal analysis of theH. cantator complex are suffi
ciently robust to
recommend modification of species limits atthis time. The
analysis builds on the methodol-ogy for considering taxonomic
limits in allopat-ric antbird populations established in
studiescited above. The criteria are conservative, andour
conclusions may be revised aer incorpo-ration of the molecular
results. Additionally,
because the geographic distribution of avail-able tissue samples
for H. cantator is much morerestricted than the inventory of tape
recordings,we expect that determinations of distributionallimits of
most populations and, by extension,taxonomic recommendations, will
depend onthe results of analysis of the more robust sampleof
recordings. We expect the molecular analysisto occupy a more
dominant role in construct-ing a phylogeny, the balance depending
on theconcordance of the vocal and molecular resultsand
consideration of morphological distinctionsamong populations. Given
the acceleratinganthropogenic alteration of the landscape in
theNeotropics, empirically based re-evaluations ofspecies
definitions are not only vital for studiesof speciation and
phylogenetics, but are anurgent necessity for ecological and
conservationresearch.
M
Hypocnemis cantator is distributed widely inAmazonia and the
Guianan region (Zimmerand Isler 2003). Eleven subspecies
wereaccepted by Peters (1951), and a 12th wasdescribed subsequently
by Pinto (1966). Allwere diagnosed by plumage characters basedon
specimen comparisons. We accepted existing
las vocalizaciones y la estructura gentica de los Thamnophilidae
en la Amazona. Enestudios futuros, los resultados de los anlisis de
vocalizaciones sern comparadoscon los de un estudio gentico
paralelo, para luego integrar los dos anlisis y sugerir
una filogenia. El presente estudio, junto con anlisis
preliminares de variacin gentica(e.g., Bates et al. 1999) indican
claramente que el complejo de H. cantatorha tenido unahistoria
evolutiva prolongada que produjo una mayor diversidad de especies
que laque ha sido reconocida hasta ahora.
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
4/19
Warbling Antbird Species LimitsJanuary 2007] 13
subspecies and morphological distinctions todefine a baseline
taxonomy and, to simplifyexposition, we employed subspecies
names
without initials as to genus or species and abbre-viated
subspecies names to the first three leersin maps and tables.
Because of the possibility,however, that existing, plumage-based
subspe-cies definitions may not fully reflect distinctionsamong
independently evolving populations asreflected in their
vocalizations, we further dis-aggregated the inventory of vocal
recordings tosearch for possible diagnostic vocal differenceswithin
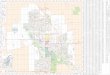
the geographic range of named subspe-cies. Twenty-six geographic
clusters (Fig. 1),identified in the text as subpopulations,
weredefined on the basis of existing knowledge ofgeographic limits
of thamnophilid antbirds.
For example, if a major river bisecting therange of an H.
cantator subspecies was knownto demarcate the ranges of sister
populations ofother thamnophilid taxa, recordings obtainedon
opposite sides of the river were analyzedas subpopulations.
Initially, pairwise vocalcomparisons were made between
subpopula-tions of currently described subspecies (e.g.,
fla1 was compared with fla2). Then, data forsubpopulations whose
vocal data did not differdiagnostically were aggregated, and
pairwisecomparisons were made of the vocal character-istics of all
subspecies and vocally diagnosticsubpopulations. The final results
provided the
basis for taxonomic recommendations, includ-ing a test of the
extent of geographic congruencewith plumage-based subspecies.
Tape recordings were compiled from ourown inventories, from
currently unarchivedcontributions by other individuals, and fromthe
Macaulay Library (ML, at the CornellLaboratory of Ornithology), the
NationalSound Archive (NSA, at The British Library),and the Florida
State Museum (FSM, at theUniversity of Florida). Appendix 1
(online; seeAcknowledgments) provides a list of recordingsemployed
in the study (774 recordings from 196localities) with locality
names, recordists, andarchival locations. No recordings were
availablefrom within the range of H. c. perflava.
We reviewed every recording to identify thenumber and sex of
individuals vocalizing andto label every vocalization as to type.
We identi-fied five vocalization types that we termed maleloudsong,
female loudsong, common call, chirr,and chitcall. Loudsongs
(following Willis 1967)
refer to the ringing series of notes delivered ina consistent
paern oen described simply assong. Common calls are also multinote
vocal-
izations delivered in a consistent paern but aremuch briefer
than and structurally distinct fromloudsongs. Common calls are
sometimes sub-
ject to individual variation (e.g., in duration orfrequency
structure), but atypical common callsare almost always interspersed
within lengthyseries of typical ones. Chirrs are single,
vibrant,harsh notes, and chitcalls are abrupt notes thatare
typically repeated rapidly in clusters of twoto four. Stereotypy of
placement of spectral ele-ments within these vocal types (relative
posi-tion, following Remane [1952]) allowed us toplace
vocalizations into these categories witha relatively high degree of
certainty that they
were homologous.CANARY, version 1.2 (Bioacoustics Research
Program, Cornell Laboratory of Ornithology),was employed to make
a spectrogram of everyvocalization type delivered by each
individualof either sex on every recording. Visual char-acters were
obtained by examining printedcopies of all clearly delineated
spectrograms(i.e., sampling was not employed). We con-sidered a
character to be diagnostic visuallywhen examination of the
character completelydistinguished every spectrogram of one
popula-tion from another. If there was any uncertaintyin this
regard, we conducted blind tests inwhich spectrograms were stripped
of any iden-tification except a randomly selected code num-
ber and sorted visually into groups accordingto perceived
differences. The groups were thenconsidered to differ diagnosably
if they provedto be separated geographically.
We employed the following terms in visualdescriptions of note
shape. Downslurredmeans that the note drops in frequency (i.e.,
thespectrogram trace goes from a higher to a lowerfrequency). An
upslurred note goes in theopposite direction. A flat note shows
lile orno change in frequency (a horizontal line on
thespectrogram). A rectangular note is a flat notewith enough band
width that the note appearsas a rectangle on a spectrogram at the
scaleemployed here. Frequency rises and falls in aninverted U note,
and the spectrogram trace isdistinctly rounded near the point of
peak fre-quency. A chevron is like an inverted U butshows a sharp
peak at the highest frequency,resembling an inverted V. Sometimes,
inverted
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
5/19
I, I, W14 [Auk, Vol. 124
U notes or chevrons are not symmetrical;one side of the note may
be more intense (asexpressed by wider band width on the
spectro-gram), in which case we refer to the le-handor right-hand
side of the note. Lengthydownward extensions to lower frequencies
are
called le- or right-hand tails. Note shapepaern describes how
the shapes of notes,including note duration, change throughouta
multinote vocalization. Clear notes arerepresented on a spectrogram
by distinct edges,whereas a note is said to become raspy when a
F. 1. Geographic distribution of recordings and definitions of
hypothetical populations ana-lyzed in Stage 1. Recording localities
are assigned to geographic sectors using methods described
by Isler (1997). Solid circles = recording locations. Heavy
lines connect recording locations includedin each population and
encircle other recording locations in the population. Areas outside
theheavy lines contain populations for which we had no recordings.
Currently defined subspecies:aff= H. c. affinis, can= H. c.
cantator, col= H. c. collinsi,fla= H. c. flavescens, imp= H. c.
implicata, not=H. c. notaea, och= H. c. ochrogyna,per= H. c.
peruviana, sat= H. c. saturata, str= H. c. striata, and sub=H. c.
subflava. No recordings were available for H. c. perflava. Numbers
after abbreviations identifyhypothesized populations within
subspecies ranges. Vocalizations ofper3and color subhave
beenrecorded at sites in the hatched area, and labels for these
populations are repeated on each sideof the hatched area to locate
where the population occurs. Bothper3and subhave also been
tape-recorded at the location marked A on the map. Loudsong
recordings (n= 2) from Ilha de Maraj,Par, Brazil (identified in the
figure as aff2), were omitted from the aggregated analysis because
of
uncertainty regarding the population to which they should be
referred.
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
6/19
Warbling Antbird Species LimitsJanuary 2007] 15
noticeable part of its edges dissolve into noise.Most raspy
notes cover a wide band width andmay or may not contain a distinct
solid area
of high intensity in their core. Terminal raspynotes in
loudsongs were not included in descrip-tions of note shape paern
because individualsoen vary the number of such notes in a seriesof
songs. Visual determinations of change inpeak frequency were made
in reference to ahorizontal line drawn through the peak
frequen-cies of a spectrogram of a multinote vocalization.Peak
frequency refers to the highest frequencyaained by the most intense
trace of an indi-vidual note. Rise, flat, and fall refer tochanges
of peak frequencies among the notesand signify an increase, no
change, and decrease,respectively.
Quantitative measures of continuous vocalcharacters were
restricted to a sample ofrecordings. Sample sizes (reflecting
number ofindividual birds recorded, not number of vocal-izations
measured) are provided in the Tables25 (online; see
Acknowledgments). If morethan six suitable recordings were
available, wesought to distribute the sample throughout theregion
defined for the population. In the finalstage, aer populations were
aggregated, datafor more than six individuals were availablefor
most populations. To obtain quantitativemeasures, spectrograms were
projected onthe screen of a Macintosh G4 computer usingdefault
seings of CANARY (Charif et al. 1995),except that the display was
set to smooth, over-lap was adjusted from 50% to 93.75% dependingon
recording quality, and contrast was adjustedaccording to recording
intensity, with caretaken to retain all elements of the
vocalization.Cursor measurements were made by PRI, typi-cally at
scales of 0.3 s inch1and 4.0 kHz inch1.Spectrograms shown in the
figures were made
by exporting CANARY files into CANVAS,version 9.0.4 (ACD
Systems, Victoria, BritishColumbia).
Whenever possible, we measured three vocal-izations of each
vocalization type for six indi-viduals of every population
identified in Figure1. It was not always possible to achieve this
goal,
because of an inadequate number of recordingsfor some
populations and because a high level ofrecording quality was
required for many of themeasurements. As in the case of visual
descrip-tions, terminal raspy notes were not included
inmeasurements. Vocal characters were defined
on two levels. General characters included(1) number of notes,
(2) duration, (3) pace, (4)change of pace, (5) note shape, (6)
change in
note shape, (7) note duration, (8) change in noteduration, (9)
interval duration, (10) change ininterval duration, (11) peak
frequency, and (12)change in peak frequency. Specific measures(24)
were assigned to general characters andincluded the number of
notes; duration; over-all pace (measured from the beginning of
thesecond note to the beginning of the first raspynote); durations
of first, middle, and last clearnotes and the intervals (spaces)
following them;ratios between note and interval durations; andpeak
frequencies of first, middle, and last clearnote and ratios between
them. Measures aredescribed more fully in earlier papers (Isler et
al.
1998, 1999), and measurement units are providedin table
legends.
In defining vocal characters that relate to spe-cies differences
in antbirds, we are interested inidentifying distinct character
states that havethe potential for unambiguous signal recogni-tion.
Thus, rather than using standard statisti-cal tests, which would
only test for differenceof means, we apply more stringent criteria
thatrequire diagnostic differences in vocal char-acters to
represent discrete, non-overlappingcharacter states, as described
in detail in earlierstudies (Isler et al. 1998, 1999). In the case
ofcontinuous variables, ranges of sample valuescannot overlap, and
the means (x) and stan-dard deviations (SD) of the population with
thesmaller set of measurements (a) and the popula-tion with the
larger set of measurements (b) hadto meet the requirement:
xa+ taSDaxb tbSDb (1)
where ti = t-score at the 97.5 percentile of thet distribution
for n 1 degrees of freedom(except for ratios, for which this
statistical testis inappropriate). Visual and mensural charac-ters
were selected to distinguish populationsof each pair only if they
were considered inde-pendent. This required careful judgments
ofwhether variation in one character could occurwithout resulting
in a different outcome foranother character. If two characters
appearedto be highly dependent, one was selected thatappeared most
appropriate to the vocaliza-tion type. For example, because common
callscomprised only a few notes that afforded a
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
7/19
I, I, W16 [Auk, Vol. 124
limited number of comparable measurements,visually defined
characters were deemed moreinformative.
On the basis of vocal distinctions among con-solidated
populations, we assessed their taxo-nomic status, employing
guidelines developedin previous studies. We recommended
speciesstatus under the Biological Species Concept forpopulations
that differed diagnostically in bothvocalizations and morphology
(Johnson et al.1999). Using a study of vocalizations of syn-topic,
congeneric species-pairs of antbirds andsubsequent applications
(Isler et al. 1998, 1999,2001) as a point of reference, vocal
differences
between allopatric populations were consid-ered diagnostic at
the species level if they werereflected in at least three
independent vocal
characters. As noted above, current subspeciesdefinitions were
accepted as representing diag-nostic morphological differences.
Existing taxathat did not meet vocal requirements for
con-sideration as a distinct species were maintainedas subspecies
pending results of the parallelmolecular study. Future studies will
describethe results of the molecular analysis, review theresults of
the vocal analysis in this context, and,eventually, estimate a
phylogeny for the H. can-tatorcomplex based on both data sets.
R
Diagnostic differences were identified inmale loudsongs, female
loudsongs, commoncalls, and chit calls. No differences in
chirrswere found between populations. Chirrs wererecorded for 17
subpopulations but not forsaturata2, peruviana1, peruviana2,
peruviana4,
peruviana5, subflava, collinsi, and affi nis2. Giventhat chirrs
tended to be the least frequentlyrecorded vocalization type, their
absence fromthe sample of some populations repertoriesmay be a
sampling artifact. However, the lack ofchirrs in the recordings of
subflavaand collinsi,for which there were large samples, may
proveto be taxonomically relevant.
No diagnostic vocal differences were identi-fied in pairwise
comparisons of subpopulations(Fig. 1), with one exception: a
difference incommon calls was found between implicata2andthe
remaining subpopulations of implicata. Someother subpopulation
comparisons, however, wereconstrained by small sample sizes, and
althoughthere were other pairwise comparisons in which
ranges of values did not overlap, these did notmeet our
statistical test. These differences will bere-examined in the
future, when the results of the
ongoing genetic study are available.As a consequence of the
pairwise comparisons
of subpopulations, implicata vocal data weremaintained in two
populations, but data for allother subpopulations were consolidated
into tenexisting subspecies (no recordings were availablefor
perflava) in the following round of analysis.Although vocalizations
of most subspecies dif-fered diagnostically in multiple characters,
pair-wise comparisons revealed no diagnostic vocaldifferences
between notaeaand cantator, saturataand peruviana, collinsi and
subflava, or implicata(excluding implicata2) and affi nis. It is
especiallynoteworthy that neither loudsongs nor common
calls distinguished vocalizations between mem-bers of these four
pairs of taxa.
In the final stage, subspecies for which therewere no
diagnosable vocal differences wereconsolidated, resulting in nine
taxa, which,using the principle of priority, were desig-nated as:
cantator (including notaea), flavescens (including perflava, on the
basis of range andplumage characteristics), peruviana
(includingsaturata), subflava (including collinsi),
ochrogyna,implicata, taxon novum(= implicata2), striata, andaffi
nis. Each taxon was distinguished from everyother by at least two
vocal characters, except forimplicataand affi nis, which were
maintained astaxa because their geographic ranges appearedto be
disjunct, separated by striata. Geographicranges are depicted in
Figure 2. Range bound-aries were based on specimen as well as
onvocalization information and reflect resolu-tion of some
ambiguities or inaccuracies (e.g.,contrary to Peters [1951], the
Rio Madeira andRio Tapajs delimited the western and easternlimits,
respectively, of implicata).
Table 1 provides the aggregated numbers ofdiagnostic vocal
characters distinguishing pairsof taxa. Vocal characters underlying
these num-
bers are provided in Appendix 2 (online; seeAcknowledgments),
along with supporting data.The following descriptive summary points
outsalient, primarily visual, differences in diagnosticcharacters.
It uses groupings of taxa to facilitatecomparisons. These groupings
do not necessar-ily reflect estimates of phylogeny, which will
bedeveloped subsequently.
Male loudsongs.All populations delivereda series of clear notes
ending with one or more
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
8/19
Warbling Antbird Species LimitsJanuary 2007] 17
raspy notes (which were omied occasionally).Measurements were
confined to the series ofclear notes because of intra-individual
inconsis-tency in number and structure of raspy notes.Male
loudsongs (Fig. 3) fell into three groupsaccording to change in
pace. Male loudsongs offive taxa (cantator, taxon novum, implicata,
striata,and affi nis) accelerated (except for one individualeach of
cantatorand implicata, which had aberrantsongs); those of three
taxa (flavescens, peruviana,
and subflava) decelerated; and ochrogynahad rela-tively evenly
paced loudsongs.
Male loudsongs of taxon novum, implicata,striata, and affi nisof
southeast Amazonia beganwith a long downslurred note followed byan
accelerating series of abrupt notes. Theserapidly delivered
vocalizations almost alwaysconsisted of at least 7 clear notes, and
usually9 or 10. Notes following the initial note weredownslurred or
boxy. Within the group, only
F. 2. Geographic distributions of taxa found to differ in at
least one vocal character from allother populations; vocalizations
of implicata and affinis were not distinguishable. Open circles
=specimen locations; open stars = recording locations, most of
which are also specimen locations;1 = cantator, 2 = flavescens, 3 =
peruviana, 4 = subflava, 5 = ochrogyna, 6a = taxon novum, 6b =
implicata,6c = striata, and 6d = affinis. Ranges of
subflavaandperuvianaoverlap in the hatched area and nearthe
location labeled A where these taxa have been observed in sympatry;
double lines to thenorth and south of A represent apparent
altitudinal parapatry. Parapatry without any majorphysical barrier
also appears to occur between flavescensand cantator in the
southeastern cornerof Venezuela, located by letter B on map.
Question marks identify regions lacking data where aform in the
complex may occur.
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
9/19
I, I, W18 [Auk, Vol. 124
male loudsongs of striatadiffered diagnosticallyin having more
abrupt notes whose frequencypeaks fell toward the end of the
series.
Differences between male loudsongs of can-tator (the fih
population whose notes acceler-ated) and the above southeastern
Amazoniantaxa were evident visually. Notes in the cantatorloudsong
could be placed in three groups: a shortinitial note, two to six
notes of similar shape andpeak frequency but at a lower peak
frequencythan the initial note, and an abrupt shi to athird group
of one to four similar notes at aneven higher peak frequency than
the initial note.
Male loudsongs ofperuvianaand subflavawerenot distinguishable
from one another. The decel-erating loudsongs of these two taxa
differed fromthe accelerating loudsongs of taxa in southeast-
ern Amazonia in multiple characters, but wereclosest to those of
cantator. In contrast to cantatornotes, which abruptly changed
shape and peakfrequency, note changes inperuvianaand
subflavaloudsongs were gradual, broadening in approxi-mately even
steps. In uncommon examples of
peruvianaand subflavathat had sudden shis innote shape, only a
single shi occurred, ratherthan two as in cantator, and subsequent
noteswidened gradually as in typical songs.
Like those of peruviana and subflava, maleloudsongs of
flavescens decelerated, but shapesof initialflavescens notes were
boxy, with a nar-row frequency range. Through the middle por-tion
of the vocalization, notes became broader,frequency-modulated, and
downslurred, endingwith distinctive long, burry, and
downslurrednotes. Note peaks remained flat in frequency orgradually
rose slightly while the boom of thenotes extended downward in
frequency.
The final population, ochrogyna, showed theleast change of pace
in male loudsong. It wasreadily distinguished from all other taxa
byfrequency paern, with the peaks of its initialfour to six notes
increasing gradually. Exceptfor southeastern Amazonian
populations,loudsongs of ochrogyna also differed by hav-ing an
initial note that was substantially longerthan the clear notes that
followed. It was dis-tinguished from the southeastern
Amazonianpopulations by its U-shaped notes.
Female loudsongs.Female loudsongs (Fig.4) of all populations
were similar, a descend-ing series of 6 to 9 (extremes 412) notes,
easilydistinguished from male loudsongs. They wereoen delivered in
response to loudsongs of
T.1.Num
berofvocalcharactersdistinguishingpopulationsidentifiedasdistinctinStage1.Initialnumberisthetotalnumber
ofindepen
dentdiagnosticcharactersandisfollowedbynumberofdiagnosticcharacter
sforeachvocalizationtype.M=male
loudsongs
;F=femaleloudsongs;C=calls,includingbothcommoncallsandchitcalls.Ifaleerisomied,nodiagnostic
differencesinthatvocalizationtypewerefound.
CharactersareidentifiedinAppendix
2(online;seeAcknowledgments).
flavescens
peruviana
subflava
ochropyga
taxonnov.
implicata
striata
affinis
cantator
5/M3C2
3/M1C2
4/M1F1C2
6/M2F2C2
7/M4F1C2
8/M4
F2C2
9/M5F2C2
6/M4C2
flavescens
3/M1C2
5/M2C3
8/M2F4C2
9/M4F3C2
6/M1
F3C2
13M6F5C2
6/M2F2C2
peruviana
3/C3
4/M3F1
11/M7F2C2
7/M3
F2C2
12/M8F2C2
10/M6F2C2
subflava
9/M4F2C3
11/M8C3
6/M3
C3
13/M7F3C3
9/M5F1C3
ochrogyna
6/M3F1C2
4/M2
C2
9/M5F2C2
4/M2C2
taxonnovum
2/C2
4/M2C2
2/C2
implicata
3/M2C1
0
striata
2/M1C1
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
10/19
Warbling Antbird Species LimitsJanuary 2007] 19
F.3.Maleloudson
gsoftheHypocnemiscantatorcomplex
.Arrowsidentifythefirstnotesthatw
ereconsideredraspy(seetext).(A)cantator
90kmnorthofManaus,Amazonas,Brazil(WhitneyISL-BM
W.189:24).(B)flavescensFrentedelaI
slaCigarrn,Amazonas,Venezuela(S
chwartz
ML62045).(C)peruvia
naSantaRita,Loreto,Peru(WhitneyISL-BMW.187:02).(D)subflavaSana
marca,Ayacucho,Peru(WiddowsonSterling
ISL-BW&JS.1:06).(E)ochrogynaVilaBeladaSantssimaTrin
dade,MT,Brazil(WhitneyISL-BMW.190:10).(F)implicataBorba,Amazona
s,Brazil
(WhitneyISL-BMW.18
8:12).(G)taxonnovumViladoCarmo
,Amazonas,Brazil(WhitneyISL-BMW.197:17).(H)striata15kmsouthofA
ltamira,
Par,Brazil(WhitneyISL-BMW.178:06).(I)affinisSerradosC
arajs,Par,Brazil(WhitneyISL.BMW.6:12).
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
11/19
I, I, W20 [Auk, Vol. 124
F.4.FemaleloudsongsoftheHypocnemiscantatorcompl
ex.Arrowsidentifythefirstnotesthatwereconsideredraspy(seetext).(A)canta-
torBrownsbergNatureReserve,Suriname(WhitneyISL-BM
W.119:26).(B)flavescensFrentedelaIslaCigarrn,Amazonas,Venezuela(Schwartz
ML62045).(C)peruvianaYarapaReserve,Loreto,Peru(WhitneyISL-BMW.163:05).(D)subflavaSan
amarca,Ayacucho,Peru(WiddowsonSterling
ISL-BW&JS.1:06).(E)oc
hrogynaSerradosPacasNovos,Rondnia,Brazil(WhittakerISL-AW.7:23).
(F)implicataCaima,Par,Brazil(WhitneyISL-
BMW.158:17).(G)taxon
novumViladoCarmo,Amazonas,Brazil(WhitneyISL-BMW.197:14).(H)stria
taRurpolis,Par,Brazil(P.IslerML4
7910).(I)
affinisCaxiuan,Par,Brazil(ZimmerISL-KJZ.85:22).
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
12/19
Warbling Antbird Species LimitsJanuary 2007] 21
their mates and typically started immediatelybefore or as the
male loudsong ended. Noteshapes were similar in all populations;
initial
notes were usually downslurred and shadedgradually into terminal
notes, which wereshaped like an inverted U. Note shapes
offlave-scenswere most divergent. These were typicallyrectangular
in shape, as shown in the figure, buta few individuals delivered
notes resemblingthose of other populations, and,
consequently,differences were not considered diagnostic.
Diagnostic differences in female loudsongswere fewer than those
found in males: 44 pair-wise comparisons as compared to 132.
Themost prevalent character distinguishing femaleloudsongs
(Appendix 2) was a shi in note dura-tion, expressed by a ratio
between durations of
initial and terminal clear notes. As is evident inthe
spectrograms, females of the southeasternAmazonian taxa (taxon
novum, implicata, striata,and affi nis) and, to a lesser extent,
neighbor-ing ochrogyna, initiated their loudsongs with along note
and ended with an abrupt note (set-ting aside terminal raspy
notes). Differencesin ratios between first and last notes in
canta-tor, peruviana, and subflava were diagnosticallysmaller and,
at the extreme, notes of flavescensloudsongs increased in duration.
Comparisonsof note duration and overall pace also
provideddiagnostic differences, but they were rarer givenlarge
within-taxa variances. Note that the inter-vals between notes
diminished at a similar rate inall taxa, and a difference in
interval duration wasdiagnostic in only one pairwise
comparison.
With regard to frequency paern, peak fre-quencies gradually
declined in female loud-songs of all taxa, though peaks of the
first twonotes were constant or nearly so for most taxa.However,
peaks of the first three notes of ochrog-
ynaascended and those of cantatordescended infrequency.
Loudsongs of affi nis were lowest infrequency, diagnostically lower
in initial notesthan cantatorand subflava.
We documented no diagnostic differencesin pairwise comparisons
of female loudsongsamong cantator, flavescens, peruviana, and
sub-
flava. Similarly, no diagnostic differences werefound among
female loudsongs of taxon novum,implicata,striata, and affi
nis.
Common call.Observing Warbling Antbirdsin the wild over many
years has revealed thatthe common call (Fig. 5) is an important
andfrequently delivered vocalization that may be
employed by both sexes and all age classes in var-ious
intraspecific contexts, such as maintenanceof auditory contact
between members of pairs or
families and encounters between neighbors. Itwas typically a
short (usually 0.51.0 s) series oftwo to five notes. Although
female calls of sometaxa tended to contain fewer notes, common
callsshowed no other sex-linked differentiation, andsamples from
both sexes were combined in theanalyses. Despite the variety of
contexts in whichan individual may deliver this call and varia-tion
in the number of notes, the great majorityof common calls were
highly consistent withinpopulations. Vocal characters were defined
visu-ally rather than measured because common callscontained few
notes. Except in pairwise compari-sons ofperuvianaand ochrogynaand
implicataand
affi nis, common calls of all populations differeddiagnostically
in either note shape or frequencypaern, and both were diagnostic in
31 of 36pairwise comparisons. Thus, of all vocalizationtypes,
common calls revealed the most consistentvocal differences among
taxa.
Common calls of cantatorconsisted of a briefrepetitive series of
two to four (most oen three)similar, clear notes having essentially
level peakfrequencies (Fig. 5A). Final notes oen becameraspy, but
maintained the width and spacing ofclear notes (Fig. 5B). Notes
sounded upslurred
because their intensity was greatest at thebeginning of the
note. Calls of peruviana, likethose of cantator, typically
consisted of two tothree notes (rarely four to six) of
approximatelythe same note-duration and spacing. Unlike can-tator,
however, the initial two notes differed inshape, the second
starting at a higher frequencythan the first and becoming more
downslurred(Fig. 5E). When present, the third note was usu-ally
identical to the second (Fig. 5F), but thethird note sometimes
began at an even higherfrequency and became more downslurred
thanthe second note (Fig. 5G). Occasionally, secondand third notes
were rounded like cantator, butthe emphasis remained on the
downward sideof the note (Fig. 5H). Rarely, the initial note
wasreplicated in the second note. In addition, endnotes of
peruviana calls rarely became raspy (2of 70 recordings), whereas
cantator calls typi-cally ended in raspy notes (22 of 29
recordings).Calls of ochrogyna (Fig. 5KL) fit the paern of
peruviana in having notes following the initialnote becoming
more downslurred. However,notes of peruviana calls maintained the
same
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
13/19
I, I, W22 [Auk, Vol. 124
F. 5. Calls of the Hypocnemis cantatorcomplex. A through S are
common calls and variants. Thelatter illustrate the range of
variation and are almost always delivered within series of common
calls.T and U exemplify chirrs, which were not found to distinguish
taxa. V is an example of a chitcall,which was not found to differ
among taxa except for subflava(W). Caption continued on next
page.
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
14/19
Warbling Antbird Species LimitsJanuary 2007] 23
width and were almost never raspy, whereasequivalent notes of
ochrogyna shortenedand usually became raspy (Fig. 5K; 14 of 20
individuals). We suspect that ochrogyna callswithout raspy
endings (Fig. 5L), whose notesshortened like the raspy calls, may
have beengiven by females, although notes lengthenedin one aberrant
call identified as given by afemale. Without more knowledge of the
sourcesof possibly overlapping calls of peruviana andochrogyna, we
do not consider that calls of thesetaxa differ diagnosably at this
time.
There were also no diagnosable differencesbetween calls of
implicata(Fig. 5MO) and affi nis(Fig. 5S). They shared a common
call consistingof a relatively long whistled note that was flat
ornearly flat in frequency followed by a distinct
interval and an clear, abrupt, downslurred notestarting at a
higher frequency than the first. Thesecond note was followed
instantaneously by asharply descending, raspy note which
typically
began at an even higher frequency (Fig. 5M, S).The abrupt, clear
note before the terminal raspynote usually had no overtones.
Occasionally, theraspy note was omied (Fig. 5N), and sometimesa
downslurred note was inserted, producinga more distinctly
three-noted call (Fig. 5O).Common calls of striata(Fig. 5QR) were
similar
to those of implicata1and affi nis, though separablein 100% of
blind trials. The first note was shortand usually in the shape of a
shallow inverted
U (sometimes flaened), and the following notewas raspy and
complex (Fig. 5Q). The second andthird notes (if present) were
almost always raspy;occasionally, the second note was
intermediate
between a clear and raspy note, consisting of ashort clear
segment and overlapping harmonicsin a broad frequency band (Fig.
5R).
The common call of flavescens consisted of amedium-to-long,
clear note followed by three tonine abrupt, sharply downslurred
notes usuallystarting at a higher frequency than the first
note(Fig. 5C). The abrupt notes typically maintainedthe same shape
and frequency range, but short-ened in length as the series
accelerated. In lon-
ger calls of four or more notes, the abrupt notesalso usually
dropped in intensity toward theend. Calls of males appeared to be
more rapidthan those of females, but sample sizes were toosmall to
confirm this difference. Occasionally,female calls were shortened
to three or even twonotes and slowed to the point where they
weresimilar to those of other populations (Fig. 5D),
but these aberrant calls were always given inseries with more
typical calls. No recorded callsofflavescens included raspy
terminal notes.
F. 5. Continued. Examples: (A) cantator common call without
raspy ending; 90 km north ofManaus, Amazonas, Brazil (Bierregaard
ML 42840). (B) cantator common call with raspy ending;Iwokrama
Forest Reserve, Guyana (Whitney ISL-BMW.145:31).
(C)flavescenscommon call; Cao LaUrbana, Bolvar, Venezuela (Schwartz
ML 62038). (D) flavescens variant given by female; P. N.
N.Chiribiquete, Caquet, Colombia (M. Alvarez ISL-MAR.4:36). (E)
peruviana common two-notedcall; Humait, Amazonas, Brazil (Whitney
ISL-BMW.195:05). (F) peruviana common three-notedcall; Limoncocha,
Napo, Ecuador (Coffey ISL-BBC.1:24). (G) peruviana variant
three-noted call;Catuaba Reserva, Acre, Brazil (Whitney
ISL-BMW.141:12). (H)peruvianavariant three-noted call; ElDorado,
Loreto, Peru (Whitney ISL-BMW.185:10). (I) subflavacommon call;
Amazonia Lodge, Madrede Dios, Peru (Zimmer ISL-KJZ.109:14). (J)
subflavacommon call; Catuaba, Acre, Brazil (WhitneyISL-BMW.141:20)
(It is unclear whether this call or I is more typical.) (K)
ochrogynacommon call;26 km east of Vila Bela da Santssima Trindade,
Mato Grosso, Brazil (Whitney ISL-BMW.190:14).(L) ochrogynavariant
without raspy ending; 26 km east of Vila Bela da Santssima
Trindade, MatoGrosso, Brazil (Whitney ISL-BMW.190:14). (M)
implicatacommon call; 25 km west, by road, of Boim,Par, Brazil
(Whitney ISL-BMW.107:30). (N) implicatavariant without raspy
ending; Prainha Nova,Amazonas, Brazil (Whitney ISL-BMW.195:30). (O)
implicatathree-note variant without raspy end-ing; Prainha Nova,
Amazonas, Brazil (Whitney ISL-BMW.196:08). (P) taxon novum common
call;Manicor, AM, Brazil (Whitney ISL-BMW.187:21). (Q)
striatacommon call with raspy chirr; Portodo Meio, Par, Brazil
(Whitney ISL-BMW.175:11). (R) striatacommon call with raspy
overtones; SoBenedito, Par, Brazil (Whitney ISL-BMW.173:24). (S)
affiniscommon call; Caxiuan, Par, Brazil(Zimmer ISL-KJZ.83:04). (T)
ochrogyna example of a chirr; Florida, Santa Cruz, Bolivia
(WhitneyISL-BMW.199:16). (U) cantatorexample of a chirr; Foengoe
Eiland, Suriname (Whitney ISL-BMW.120:06). (V)peruviana example
ofchitcall; Quebrada Sucusari, Loreto, Peru (Parker ML 29101). (W)
sub-
flava example ofchitcall; Humait Reserva, Acre, Brazil (Whitney
ISL-BMW.142:25).
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
15/19
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
16/19
Warbling Antbird Species LimitsJanuary 2007] 25
by D. Lebbin and W. Tori (pers. comm.), who,in studies of the
avifauna of Guadua bamboo,need 27 individuals of subflava inside
patches
of bamboo, whereas net lines (maintainedfor fewer hours) in
nearby forest outside the
bamboo need five peruviana and no subflava.However, further
study by N. Seddon and J.Tobias (pers. comm.) at this site found
the habi-tat distinction to be less strict. For example, theyfound
peruviana territories in the understory ofpalm swamps and in areas
containing bamboo,and subflava territories in tangled riverine
veg-etation and in areas lacking bamboo; and theyfound that the two
forms occasionally inhabitedadjacent territories. Thus,
distinctions in habi-tat preferences between the two forms
remainunclearan important subject for future field
work. Syntopy of peruviana and subflava hasalso been reported in
Peru from near Pozuzo(1004S, 7532W), Pasco (T. Schulenbergpers.
comm.), and at Cuzco Amazonica Lodge(1233S, 6903W), Madre de Dios
(T. J. Davispers. comm.), but habitat distinctions were notstudied
at these sites.
Whether they inhabit adjacent territoriesor not, it appears that
reproductive-isolationmechanisms between peruviana and subflavahave
remained operative following secondarycontact over a fairly
extensive area in the south-western corner of Amazonia. Given this
rangeoverlap, and the close similarities in loudsongsand behavior
of these taxa, we expect that somevery limited hybridization
between peruvianaand subflava eventually may be documented,as
suspected in the mixed pair observed bySchulenberg, cited above.
Phenotypic distinc-tions we have been able to identify betweenthem
in the field are white versus yellow throatand breast, leg color
(usually bluish-gray in
peruviana, greenish-yellow in subflava), thehighly
differentiated common call (Fig. 5, EHvs. IJ), the difference in
chitcall, and the appar-ent absence of a chirr call in the vocal
repertoireof subflava.
Parapatry of cantator and flavescens.Evidence of parapatry
between these taxain southeastern Venezuela was provided
by specimens deposited in the ColeccinOrnitologia Phelps (COP).
Taxon identifica-tion was substantiated by the authors
usingspecimen photographs. Both taxa have beencollected on or
adjacent to a plateau (knownas the Macizo de Chimant) situated
between
the southwestern corner of La Gran Sabanaand the Sierra
Pacaraima in the headwaters ofthe Ro Caron (region identified by B
in Fig.
2). Specimens of cantatorwere collected on thewestern and
northern edge of this plateau at RoCaruai (= Kariay), Ro Apcara,
and Chimant-tepu. Specimens of flavescens were obtainedat Salto
Arebuchi (= Aripichi?), Acopn-tepu,Ro Icabar, Santa Elena de Uairn,
and Parai-tepu. In particular, the Ro Caruai (location forcantator)
adjoins Acopn-tepu (location for fla-vescens) at its base. Although
precise geographiccoordinates are unavailable for most of
theselocalities, they do not appear to be isolated bymajor
topographic features, though they mayrepresent different
ecosystems. Additional fieldwork is needed to understand the
biogeography
of cantatorandflavescens in this region.
D C
The number of vocal characters distinguish-ing most pairs of H.
cantator subspecies wassubstantial, in most cases greater than
thosedocumented for syntopic pairs of congenericthamnophilid
species in an earlier study (Isleret al. 1998). All three principal
types of vocaliza-tions (male loudsongs, female loudsongs,
andcommon calls) differed diagnosably in 19 of 36pairwise
comparisons (Table 1). Most surprisingwas the finding that calls
differed diagnosablyin all but two pairwise comparisons, and
aspecific result of particular interest was that thevocalizations
of the only pair of subspecies nowknown to exist in syntopy
(peruviana and sub-
flava) differed diagnostically only in the struc-ture of their
calls. Whether these findings reflectthat calls rather than songs
function in matechoice or species recognition is a question
forfuture field studies using experimental designs,such as
mate-removal experiments, necessaryfor understanding vocal behavior
and evolutionin the Thamnophilidae.
On the basis of vocal differences describedherein and
morphological differences described
by earlier authors, we recommend that the H.cantator complex be
considered at this time toconsist of six species:
Hypocnemis cantator(Boddaert)Guianan Warbling-Antbird
H. c. notaeaHellmayrH. c. cantator(Boddaert)
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
17/19
I, I, W26 [Auk, Vol. 124
Hypocnemis flavescensSclaterImeri Warbling-Antbird
H. f. flavescensSclater
H. f. perflavaPinto
Hypocnemis peruvianaTaczanowskiPeruvian Warbling-Antbird
H. p. saturataCarrikerH. p. peruvianaTaczanowski
Hypocnemis subflavaCabanisYellow-breasted Warbling-Antbird
H. s. subflavaCabanisH. s. collinsiCherrie
Hypocnemis ochrogynaZimmerRondonia Warbling-Antbird
Hypocnemis striata(Spix)Spixs Warbling-Antbird
H. s. taxon novumH. s. implicataZimmer
H. s. striata(Spix)H. s. affi nis Zimmer
From the perspective of their vocalizations,we find no reason to
maintain notaea, saturata,and collinsi as taxa. However, given the
pos-sibility that diagnosable differences may befound between them
and cantator,peruviana, andsubflava, respectively, in other suites
of charac-ters, we recommend maintaining these taxa assubspecies,
awaiting the completion of furthermolecular, morphological, and
behavioral stud-ies. We are unable to shed light on the
validityofperflava, given the absence of vocal recordings.We also
recognize the possibility that furthervocal distinctions will be
found within the rangesof proposed species when additional
recordingsprovide improved geographic coverage.
Although recommendations regarding canta-tor, flavescens,
peruviana, subflava, and ochrogynaare straightforward, the
treatment of striataand its four constituent populations is
moreproblematic. A paradox is posed in the rela-tionship between
implicata and affi nis, whosevocalizations could not be
differentiated butwhose ranges appear to be separated by that
ofstriata, whose vocalizations differed from both,though striatawas
found to differ vocally fromimplicata by three characters and from
affi nis
by two, fewer than found in most pairwisecomparisons of taxa
recommended for species
status. The importance of this vocal result isburessed by
parallel morphological findingsof Zimmer (1932), who discovered
minimal
plumage differences between implicata andaffi nis, even though
both differed in plumagefrom the geographically intervening
striata. Themost likely explanation for this paern is
thatstriatahas become genetically isolated relativelyrecently, and
that there remains or recently has
been gene flow between implicataand affi nis inthe essentially
unstudied region south of therange of striata (identified by a
question markin Fig. 2). In the interest of maintaining a
con-servative stance, we recommend that implicataand affi nisbe
maintained as subspecies of striatauntil further information is
available from theregion and the results of molecular analysis
can
be added to the data set.A second diffi culty concerns assessing
the
status of taxon novum. Although differenceswere not identified
in other vocalizations, dif-ferences in the calls of taxon novum
and otherpopulations within striata(including affi nisandthe
remainder of implicata) were substantial andcomparable to the only
known vocal differences
between the syntopicperuvianaand subflava. Wemaintain taxon
novumas a subspecies until it isformally described. It is
noteworthy that taxonnovumand striata(sensu stricto) differed in
fourvocal characters found in male loudsongs aswell as calls.
Further recommendations regarding statusof all taxa within the
Warbling Antbird com-plex and estimates of its phylogeny withinthe
Thamnophilidae await molecular analysesnow underway. It is now
apparent, however,through conservative analysis of
vocalizations,that the H. cantatorcomplex has had a long anddynamic
evolutionary history, and we are con-fident that this initial
revision beer reflects thetaxonomic status of populations in the
complexthan current classifications.
A
Supplementary data for this study are avail-able, as Appendices
1 and 2 and Tables 25,
athp://www.aou.org/suppl/auk_124_1_isler.pdf.We deeply appreciate
the continuing supportof the Macaulay Library, Cornell Laboratoryof
Ornithology (G. Budney); National SoundArchive, The British Library
(R. Rant); andFlorida State Museum. The many recordists
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
18/19
Warbling Antbird Species LimitsJanuary 2007] 27
who contributed to the effort are identified inAppendix 1, and
we are indebted to them all. T.
J. Davis, D. Lebbin, N. Seddon, T. Schulenberg, J.
Tobias, A. Whiaker, and K. Zimmer contributedunpublished
information regarding distributionand areas of sympatry of
peruvianaand subflavain Peru, and Whiaker also provided
informa-tion on the distribution of taxon novumin Brazil.We
appreciate the efforts of C. Rodner and C.Milensky to provide
photographs of specimensand related data at Coleccin Ornitologa
Phelps.
J. Bates and K. Zimmer kindly reviewed earlierdras of the
manuscript, and the final manu-script was thoughtfully reviewed by
N. Seddonand an anonymous reviewer. National ScienceFoundation
grant no. DEB 9974104 to J. M. Batesand S. J. Hacke at the Field
Museum of Natural
History helped provide computer equipmentto perform the vocal
analyses. We thank FieldGuides, Inc., for support of B.M.W.s field
work.
L C
B, L. F., D. E. K. 2001. Avianbioacoustics. Pages 1152 in
Handbook ofthe Birds of the World, vol. 6: Mousebirdsto Hornbills
(J. del Hoyo, A. Ellio, and J.Sargatal, Eds.). Lynx Edicions,
Barcelona,Spain.
B, J. M. 2000. Allozymic genetic structureand natural habitat
fragmentation: Datafor five species of Amazonian forest
birds.Condor 102:770783.
B, J. M. 2002. The genetic effects of forestfragmentation on
five species of Amazonian
birds. Journal of Avian Biology 33:276294.B, J. M., S. J. H, J.
M. G.
1999. High levels of mitochondrial DNAdifferentiation in two
lineages of antbirds(Drymophila and Hypocnemis). Auk
116:10931106.
B, R. T., A. P. C. 1996.Historical diversification of birds in
north-western South America: A molecular per-spective on the role
of vicariant events.Evolution 50:16071624.
C, A. P. 1988. Genetic variation inNeotropical birds:
Implications for the spe-ciation process. Pages 16581664 inActa
XIXCongressus Internationalis Ornithologici(H. Ouellet, Ed.).
National Museum ofNatural Sciences, University of OawaPress,
Oawa.
C, R. A., S. M, C. W. C.1995. CANARY 1.2 Users Manual.
CornellLaboratory of Ornithology, Ithaca, New
York.C, C. B., C. E. H. 1924. Catalogue
of birds of the Americas and the adjacentislands.
PteroptochidaeConopophagidaeFormicariidae. Field Museum of
NaturalHistory Zoological Series, no. 13, part III.
H, S. J. 1993. Phylogenetic and biogeo-graphic relationships in
the Neotropicalgenus Gymnopithys (Formicariidae). WilsonBulletin
105:301315.
H, S. J., K. V. R. 1990.Comparison of phenotypic and genetic
dif-ferentiation in South American antwrens(Formicariidae). Auk
107:473489.
H, A. J., A. G. K, D. T. P, G.S, M. C. 2002.Guidelines for
assigning species rank. Ibis144:518525.
I, M. L. 1997. A sector-based ornithologi-cal geographic
information system for theNeotropics. Pages 345354 in Studies
inNeotropical Ornithology Honoring TedParker (J. V. Remsen, Jr.,
Ed.). OrnithologicalMonographs, no. 48.
I, M. L., J. A. A, P. R. I, B. M.W. 2001. A new species of
Percnostolaantbird (Passeriformes: Thamnophilidae)from Amazonian
Peru, and an analysis ofspecies limits within Percnostola
rufifrons.Wilson Bulletin 113:164176.
I, M. L., P. R. I. 2003. Species lim-its in the Pygmy Antwren
(Myrmotherulabrachyura) complex (Aves:
Passeriformes:Thamnophilidae): 1. The taxonomic status of
Myrmotherula brachyura ignota. Proceedingsof the Biological
Society of Washington 116:2328.
I, M. L., P. R. I, R. T. B.2005. Clinal variation in
vocalizations of anantbird (Thamnophilidae) and implicationsfor
defining species limits. Auk 122:433444.
I, M. L., P. R. I, B. M. W.1997. Biogeography and systematics of
theThamnophilus punctatus (Thamnophilidae)complex. Pages 355381 in
Studies inNeotropical Ornithology Honoring TedParker (J. V. Remsen,
Jr., Ed.). OrnithologicalMonographs, no. 48.
I, M. L., P. R. I, B. M. W.1998. Use of vocalizations to
establish
-
8/13/2019 Species Limits in Antbirds (Thamnophilidae)
19/19
I, I, W28 [Auk, Vol. 124
species limits in antbirds (Passeriformes;Thamnophilidae). Auk
115:577590.
I, M. L., P. R. I, B. M. W. 1999.Species limits in antbirds
(Passeriformes;Thamnophilidae): TheMyrmotherula surina-
mensiscomplex. Auk 116:8396.J, N. K., J. V. R, J., C. C.1999.
Resolution of the debate over speciesconcepts in ornithology: A new
comprehen-sive biological species concept. Pages 14701482 inActa
XXII Congressus InternationalisOrnithologici (N. J. Adams and R. H.
Slotow,Eds.). BirdLife South Africa, Johannesburg.
M, E., P. D. A. 1991. Principles ofSystematic Zoology.
McGraw-Hill, New York.
P, J. L. 1951. Check-list of Birds of theWorld, vol. 7. Museum
of ComparativeZoology, Harvard University,
Cambridge,Massachuses.
P, O. M. O. 1966. Estudo crtico e catlogoremissivo das aves de
Territrio Federal deRoraima. Cadernos da Amaznia, InstitutoNacional
de Pesquisas da Amaznia 8:1176.
R, A. 1952. Die Grundlagen des natrli-chen System der
vergleichenden Anatomieund der Phylogenetik. Geest und PortigK.-G.,
Leipzig, Germany.
R, J. V., Jr. 2005. Paern, process, and rigormeet
classification. Auk 122:403413.
R, R. S., G. T. 1994. The Birdsof South America, vol. II: The
SuboscinePasserines. University of Texas Press,Austin.
S, C. G., B. L. M, J. 1990.
Distribution and Taxonomy of the Birdsof the World. Yale
University Press, NewHaven, Connecticut.
W, B. M., J. F. P, D. R. C. B, R. P. 2000. Systematic
revisionand biogeography of the Herpsilochmus
pileatuscomplex, with description of a newspecies from
northeastern Brazil. Auk 117:869891.
W, E. O. 1967. The behavior of bicol-ored antbirds. University
of CaliforniaPublications in Zoology, no. 79.
Z, J. T. 1932. Studies of Peruvian birds.V. The genera
Herpsilochmus, Microrhopias,
Formicivora, Hypocnemis, Hypocnemoides, andMyrmochanes .
American Museum Novitates538:127.
Z, K. J., M. L. I. 2003. FamilyThamnophilidae (Typical
Antbirds). Pages448681 in Handbook of the Birds of theWorld, vol.
8: Broadbills to Tapaculos (J. delHoyo, A. Ellio, and D. Christie,
Eds.). LynxEdicions, Barcelona, Spain.
Associate Editor: K. Yasukawa