Embed Size (px)
Citation preview

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Journal of Insect Behavior, Vol. 17, No. 4, July 2004 ( C© 2004)
Species-Specificity of Chemical Signals: Silk SourceAffects Discrimination in a Wolf Spider(Araneae: Lycosidae)
J. Andrew Roberts1,2 and George W. Uetz1
Accepted March 17, 2004; revised May 10, 2004
Female spiders deposit chemical cues that elicit male courtship behavior withsilk. These cues are often assumed to be species-specific although male spidersmay court in response to chemical cues of closely-related species. We usedbehavioral assays to test the extent of species discrimination of female chemi-cal cues by male Schizocosa ocreata, a wolf spider (Lycosidae). Discrimina-tion, expressed as relative courtship intensity of males, varied significantly withphylogenetic distance. Males did not discriminate between female cues of con-specifics and a sibling species, S. rovneri. Courtship response was intermediatefor another species within the ocreata clade and not different from control forspiders outside the clade. These findings support the sibling species status ofS. ocreata and S. rovneri, and also suggest the composition of female chemicalsignals is conserved across closely related wolf spider species.
KEY WORDS: chemical communication; Schizocosa ocreata; species recognition; silk; wolfspider.
INTRODUCTION
Chemical signals are used in communication across most animal taxa. In par-ticular, signaling in the chemical modality has been widely studied in arthro-pods and is suggested to be the most primitive channel for intraspecies com-munication (Weygoldt, 1977; Bradbury and Vehrencamp, 1998; Wyatt, 2003).
1Department of Biological Sciences, University of Cincinnati, Cincinnati, Ohio 45221-0006.2To whom correspondence should be addressed. E-mail: [email protected].
477
0892-7553/04/0700-0477/0 C© 2004 Springer Science+Business Media, Inc.

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
478 Roberts and Uetz
Most research on chemical communication has focused on arthropod taxa ofeconomic or medical importance such as insects or crustaceans (Weygoldt,1977; Barth, 2002; Wyatt, 2003). Chemical communication is also well es-tablished for spiders, although information concerning the use of chemicalcommunication in spiders is far less extensive than for other arthropod taxa(Kaston, 1936; Crane, 1949; Tietjen and Rovner, 1982; Pollard et al., 1987;Papke et al., 2001; Barth, 2002). Chemical signals are used by spiders in sev-eral contexts, including prey detection (Persons and Uetz, 1996; Punzo andKukoyi, 1997; Jackson et al., 2002), predator avoidance (Suter et al., 1989;Persons et al., 2001, 2002), male/male competition (Ayyagari and Tietjen,1987), and even aggressive mimicry (Stowe et al., 1987; Haynes and Yeargan,1999), but have been studied most extensively in the context of intersexualcommunication.
Intersexual chemical communication in spiders primarily occurs whenfemales signal to males through airborne and/or contact (cuticular or silkbound) chemical compounds, detected by males with specialized sensoryhairs and pit organs concentrated on the legs and palps (Barth, 2002). Thesechemical signals are involved in mediating many social interactions acrossdiverse taxa (reviews in Robinson, 1982; Tietjen and Rovner, 1982; Foelix,1996). Volatile compounds (airborne chemical cues) play a role in mate at-traction, courtship, and mating behaviors in several spider families (Blanke,1973; Olive, 1982; Riechert and Singer, 1995; Miyashita and Hayashi, 1996;Searcy et al., 1999; Papke et al., 2001; Barth, 2002), but contact chemical com-pounds are more widely studied. In funnel-web spiders (Agelenidae) of thegenus Tegenaria, changes in female cuticular chemistry may be associatedwith sexual receptivity (Prouvost et al., 1999), and cuticular compounds playa role in male courtship behavior and reproductive isolation (Trabalon et al.,1997). Male bowl-and-doily spiders, Frontinella pyramitela (Linyphiidae),detect sex differences in pheromones on the cuticle of conspecifics (Suteret al., 1987), and the silk of female webs contains multiple compounds thatattract males and release male courtship (Suter and Renkes, 1982; Suter andHirscheimer, 1986). In the wolf spider genus Rabidosa (Lycosidae), malesuse chemical cues in the draglines of females to track and locate potentialmates (Rovner, 1968; Tietjen, 1977; Tietjen and Rovner, 1980). The silk offemale wolf spiders can elicit courtship behavior in males even in the absenceof other (visual and vibratory) cues (Kaston, 1936; Rovner, 1968; Hegdekarand Dondale, 1969; Richter et al., 1971; Tietjen and Rovner, 1982).
Species-specificity of chemical signals is commonly assumed in many ofthe behavioral studies described above, particularly those involving behav-ioral assays of signal efficacy. This assumption may not be well-founded, asspiders living in separate habitats would not necessarily be expected to usedifferent chemicals for communication (Dobzhansky, 1970). Only a few stud-ies have tested species-specificity of pheromone communication in spiders.

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Discrimination of Species-Specific Chemical Signals in a Wolf Spider 479
Male Latrodectus hesperus (Theridiidae) and Linyphia triangularis (Linyphi-idae) will initiate courtship behaviors after contacting the web of females ofclosely related species (Ross and Smith, 1979; Schulz and Toft, 1993), butin linyphiids the courtship frequency is greatly reduced. Kaston (1936) ex-posed males of several wolf spider (Lycosidae) and jumping spider (Saltici-dae) species to closely related heterospecific females and noted that malesinitially courted on contact with the silk-laden substrate but stopped aftercontacting the female. Males of 30 different jumping spider species weretested for courtship when exposed to conspecific or heterospecific femalesilk and evidence of species-specific discrimination was found in all but twoof the species tested (Jackson, 1987). Chemicals used for communication inwolf spiders may be highly species-specific (Hegdekar and Dondale, 1969;Tietjen, 1977), even in species that are capable of producing hybrids (Costaand Capocasale, 1984; Costa et al., 2000), but this may not be true for somespecies in the genus Schizocosa (Hegdekar and Dondale, 1969; Uetz andDenterlein, 1979; Stratton and Uetz, 1981).
Schizocosa wolf spiders are well-studied behaviorally (reviews in Milleret al., 1998; Uetz, 2000; Uetz and Roberts, 2002), and the genus containsat least two monophyletic species groups (the saltatrix clade and the ocre-ata clade) that contain cryptic species identifiable largely by differences incourtship behavior (Stratton et al., 1996; Miller et al., 1998). Within the ocreataclade, male S. ocreata court conspecific female silk and heterospecific femalesilk of its sibling species S. rovneri with equal frequency (Uetz and Denterlein,1979; Stratton and Uetz, 1981), raising questions about the ability of malesto discriminate. However, this was measured using an all-or-none score, theintensity (i.e. vigor) of courtship was not measured, and males have not beentested on the silk of other members of the ocreata clade. In the current study,we tested the hypothesis of discrimination based on species-specific chemicalcues by examining the response of male S. ocreata to the silk of females fromtaxa that differ in degree of presumed phylogenetic distance. We measuredthe relative intensity of male courtship (latency, rate, and average durationof bouts of exploratory and courtship behaviors) to determine the extent ofmale discrimination of possible species-specific chemical signals, either ex-pressed as an all-or-nothing response (i.e. no response to any heterospecificsilk cues), or as graded variation in behavior.
METHODS
Study Animals
Species are presented throughout the manuscript in order of presumedincreasing phylogenetic distance from Schizocosa ocreata based onphylogenetic relationships in Coddington and Levi (1991), McClintock and

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
480 Roberts and Uetz
Uetz (1996), Stratton et al. (1996), and Miller et al. (1998). The spiders usedin this study are common to North America, and most are found (often sym-patrically) in the deciduous forest and edge zone habitats of eastern NorthAmerica (with the exception of outgroup M. spinipes, which is found predom-inantly along the Pacific Coast and throughout central Mexico). In additionto female S. ocreata, females of other species were used to provide silk fortests of species discrimination: two additional Schizocosa species from theocreata clade (S. rovneri, S. crassipes), one species from the saltatrix clade(S. saltatrix) within the genus Schizocosa, one representative outgroup fromthe “Trochosa” group (Stratton et al., 1996) within the family Lycosidae(Hogna helluo), one outgroup from within the “RTA clade” (those spi-der families sharing the retrolateral tibial apophysis—Coddington and Levi,1991) of the cursorial family Salticidae (Phidippus princeps), and one repre-sentative of the unrelated outgroup family Araneidae (Metepeira spinipes)within the Orbiculariae (the orb-weaving spiders).
Male and female Schizocosa ocreata used in behavioral analyses werecollected from the Cincinnati Nature Center, Rowe Woods, Clermont Co-unty, Ohio, and from the University of Cincinnati Benedict Nature Preserve,Hamilton County, Ohio. Schizocosa rovneri were collected from the OhioRiver flood plain at Sandy Run, Boone County, Kentucky. Schizocosa cras-sipes were collected on Jekyll Island, Glynn County, Georgia. Schizocosasaltatrix and H. helluo were collected from the Cincinnati Nature Center,Rowe Woods, Clermont County, Ohio. Jumping spiders, P. princeps, werecollected in Walton, Boone County, Kentucky. The orb-weaving spiders, M.spinipes, were collected from Asilomar State Beach, Pacific Grove, MontereyCounty, California.
All cursorial spiders were collected as juvenile or penultimate instarsin the field and raised to adulthood under identical laboratory conditions.Schizocosa saltatrix, H. helluo, and P. princeps are all potential predatorsof S. ocreata during some portion of its life cycle, so all of these specieswere maintained in the laboratory for at least 6 weeks prior to silk col-lection to minimize the risk of potential kairomones (chemical cues asso-ciated with previously digested prey) altering the courtship behavior of S.ocreata (Persons et al., 2001). Spiders were maintained at room temperature(22–25◦C) with stable humidity and a 13:11 h light:dark photoperiod. EachSchizocosa wolf spider and Phidippus jumping spider was visually isolatedfrom other spiders in an opaque plastic container (deli-dish, 10 cm diame-ter, round), fed 2–3 one-week-old cricket nymphs (Acheta domesticus) twiceweekly, and supplied with water ad libitum. Individual H. helluo, a larger wolfspider, were maintained in large opaque plastic containers (17 cm× 17 cm×5 cm), fed two sub-adult crickets twice weekly, and provided water ad libitum.As M. spinipes is a colonial orb-weaving spider, individuals collected as penul-timate instars were maintained with other females in a 1 m3 colony cage,

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Discrimination of Species-Specific Chemical Signals in a Wolf Spider 481
supplied with fruit flies (Drosophila melanogaster, D. heidii, and/or D. ro-busta) and houseflies (Musca domestica) as prey ad libitum, and providedwater twice daily by overhead mist.
Silk Collection
To examine whether male S. ocreata are able to detect species-specificcues associated with various female silks, adult males were chosen at randomfrom the laboratory population and exposed to either conspecific female silk,one of six heterospecific silk treatments, or to a blank filter paper control(N = 80, 10 per treatment). All male spiders ranged from 2 to 4 weeksof maturity, female spiders from 1 to 3 weeks of maturity, and all spiderswere unmated at the time of the study. In all cases where female silk cueswere collected for study, they were gathered from each spider individuallyby allowing the female to locomote and deposit dragline silk (and associatedcues) for 24 h on Fisherbrand filter paper (9 cm diameter) in a glass Petri-dish (9 cm diameter, round). Females were not used within 24 h of being fedto reduce contamination from feces (Ayyagari and Tietjen, 1987), and anyfilter paper disk with more than two fecal stains was not used in this study.One half of each circle of filter paper was used per trial, thus silk-associatedcues of each female were used with two different male S. ocreata controllingfor individual variation among females.
Behavioral Assay Trials
Behavioral trials were conducted in clear plastic containers (13 cm ×7 cm × 7 cm) that provided a visual arena suitable for videotaping. Theplastic arenas were cleaned before each trial using a Kimwipe and 70%ethanol to remove all traces of silk, feces, and chemical cues from previoustrials. Filter paper containing female silk cues was placed on the floor ofthe container. Males were placed gently onto the silk-laden filter paper fromabove and videotaped (RCA Autoshot VHS video camera, Model CC4352)for 5 min after introduction into the arena.
The following male courtship elements, adapted from Stratton and Uetz(1986), were scored for each trial using event-recording software designed forthe collection and detailed analysis of behavioral data (The Observer, ver-sion 4.1); (1) Chemoexplore—male exploratory behavior where the male rubsthe substrate with the dorsal palp surface while slowly locomoting, and (2)Jerky Tap—active courtship element where the male locomotes with rapid,“jerky” movements, actively tapping the forelegs on the substrate and of-ten striking the substrate with the ventral body surface, while producing

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
482 Roberts and Uetz
a seismic courtship component (stridulation) with specialized structures inthe pedipalps. An additional, noncourtship display behavior was scored; LegRaise—also called “arch” and/or “wave,” the male raises a foreleg above par-allel to the substrate. Three parameters were determined for each of thesebehaviors; Latency—the time from introduction of the male to the arena tothe initiation of a behavior (a score of 300 s was used if a behavior was notseen during the 5 min trial period), Mean Duration—the average time spentin each behavioral bout during the trial period, and Rate—the number ofbehavioral bouts per minute of the trial. For each parameter, the effects ofsilk treatment on male behavior were examined using one-way ANOVA. Allparameters were square root transformed for analysis, significance tests wereconducted using a Bonferroni-adjusted critical value (α = 0.017) to accountfor multiple comparisons (Shaffer, 1995), and post hoc comparisons weremade with Tukey-Kramer HSD tests. All statistical analyses were carriedout using JMP version 4.02 (SAS Institute).
RESULTS
Latency of Behavior
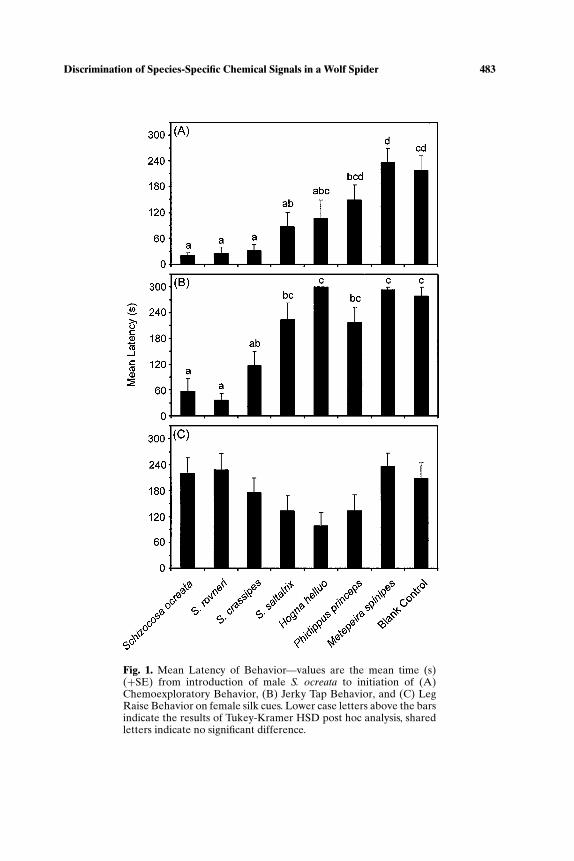
Latency of chemoexploratory behavior varied significantly among treat-ment groups (ANOVA, F7,72 = 9.913, p < 0.001), as did latency of jerky tap(ANOVA, F7,72 = 17.876, p < 0.001). Latency to begin chemoexploring wasshortest on the silk of females within the ocreata clade, intermediate on thesilk of S. saltatrix, and significantly longer (not different from blank controls)for H. helluo, P. princeps, and M. spinipes silks (Fig. 1A). Males began ac-tively courting conspecific and S. rovneri silk quickly, with a short delay onS. crassipes silk, and a response time not different from blank controls forthe remaining treatments (Fig. 1B). Latency of leg raise was not significantlydifferent among treatments (ANOVA, F7,72 = 2.053, p < 0.06), but therewas a trend of decreased latency on the silk of potential sympatric predators(S. saltatrix, H. helluo, P. princeps) (Fig. 1C).
Mean Duration of Behavior
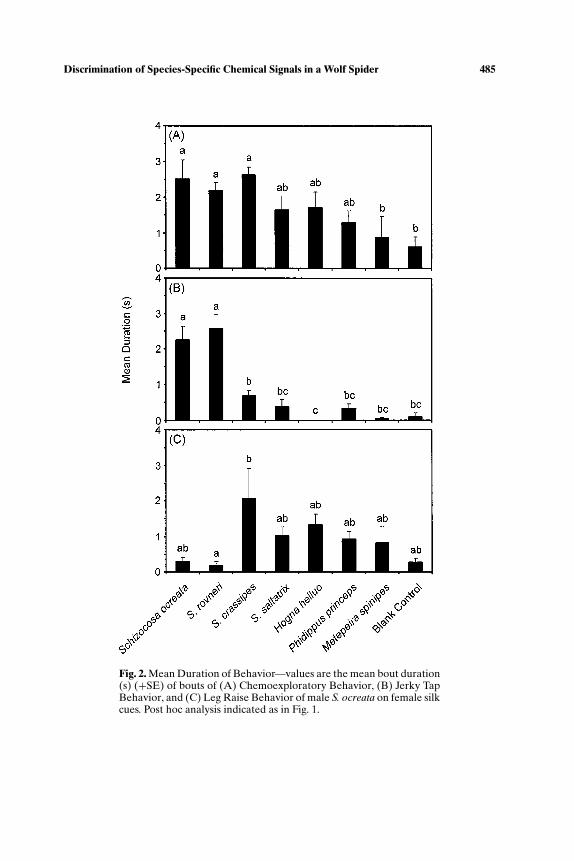
The average time spent in each bout of behavior varied significantly forchemoexplore (ANOVA, F7,72 = 4.850, p < 0.001), jerky tap (ANOVA,F7,72 = 25.036, p < 0.001), and for leg raise (ANOVA, F7,72 = 3.086, p <0.007). The mean duration of bouts of chemoexploring by male S. ocreatawas greatest on silk of females within the ocreata clade, intermediate on
P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Discrimination of Species-Specific Chemical Signals in a Wolf Spider 483
Fig. 1. Mean Latency of Behavior—values are the mean time (s)(+SE) from introduction of male S. ocreata to initiation of (A)Chemoexploratory Behavior, (B) Jerky Tap Behavior, and (C) LegRaise Behavior on female silk cues. Lower case letters above the barsindicate the results of Tukey-Kramer HSD post hoc analysis, sharedletters indicate no significant difference.

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
484 Roberts and Uetz
S. saltatrix, H. helluo, and P. princeps silks, and shortest on the M. spinipesoutgroup and blank control (Fig. 2A). The average time spent in each bout ofactive courtship was highest in the conspecific and S. rovneri treatments, andnot different from blank control for any other treatment (Fig. 2B). While themean duration of bouts of leg raise was significantly longer on S. crassipes silkthan on S. rovneri silk, it was not different than the blank control treatmentfor males exposed to any of the other silk treatments (Fig. 2C).
Rate of Behavior
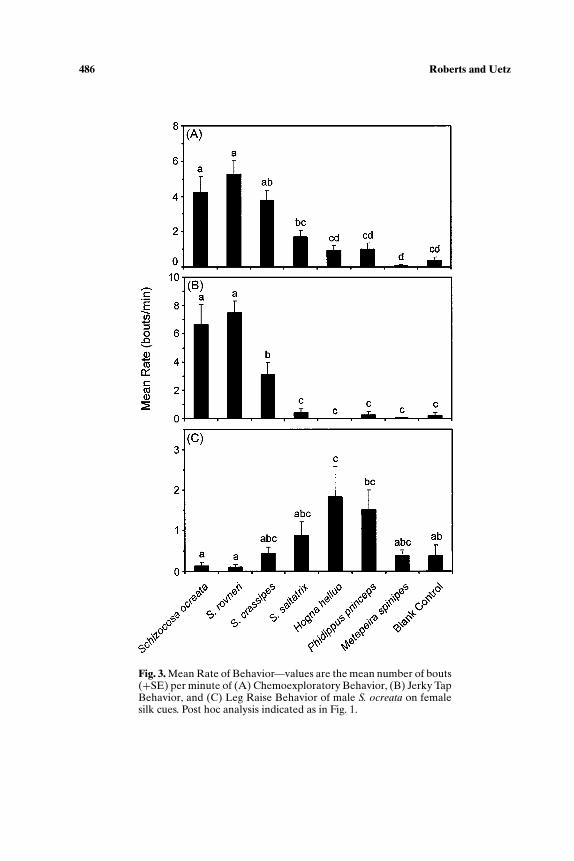
As in the analysis of mean duration of behaviors, rates of chemoexplore(ANOVA, F7,72 = 17.975, p < 0.001), jerky tap (ANOVA, F7,72 = 28.213,p < 0.001), and leg raise (ANOVA, F7,72 = 4.497, p < 0.001) all variedsignificantly among treatments. Males exhibited more bouts of chemoex-ploratory behavior per minute on ocreata clade silks than on any of the othersilk treatments, which were not different from blank controls (Fig. 3A). Rateof jerky tapping was highest and not significantly different on conspecific andS. rovneri silk, intermediate on S. crassipes silk (which was significantly differ-ent from all other treatments), and not different from blank controls for theremaining four silk treatments (Fig. 3B). The rate of leg raises was higheston H. helluo silk and significantly different from conspecific, S. rovneri, andblank control, but it was not significantly different from the blank control forany silk treatment other than H. helluo (Fig. 3C).
DISCUSSION
This study indicates that male S. ocreata court differentially in responseto variation in silk cues among species, both within and above the genus level.However, the presence or absence of male behaviors is not strictly an “all-or-nothing” occurrence by species as would be predicted if males responded onlyto species-specific chemical signals of female conspecifics. Male courtship re-sponse was not different from blank controls for species outside of the ocreataclade, but was indistinguishable for a sibling species (S. rovneri) and inter-mediate for one other species within the clade (S. crassipes). In most animaltaxa, specificity of chemical signals used in communication is determined byunique blends of compounds (Wyatt, 2003). This provides a plausible mecha-nism to explain the courtship responses seen in this study, where more closelyrelated species would likely have similar blends of chemicals. If a key chemicalcompound releasing courtship in S. ocreata is present but at a different con-centration in species within the clade, then male courtship response would beexpected to reflect that difference. The levels of chemoexploratory behavior
P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Discrimination of Species-Specific Chemical Signals in a Wolf Spider 485
Fig. 2. Mean Duration of Behavior—values are the mean bout duration(s) (+SE) of bouts of (A) Chemoexploratory Behavior, (B) Jerky TapBehavior, and (C) Leg Raise Behavior of male S. ocreata on female silkcues. Post hoc analysis indicated as in Fig. 1.
P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
486 Roberts and Uetz
Fig. 3. Mean Rate of Behavior—values are the mean number of bouts(+SE) per minute of (A) Chemoexploratory Behavior, (B) Jerky TapBehavior, and (C) Leg Raise Behavior of male S. ocreata on femalesilk cues. Post hoc analysis indicated as in Fig. 1.

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Discrimination of Species-Specific Chemical Signals in a Wolf Spider 487
varied with phylogenetic distance across the ocreata clade, the genus Schizo-cosa, the family Lycosidae, and the order Araneae, but the behavior waspresent in all treatments. This may indicate that the composition of chemicalsignals responsible for the release of chemoexploratory behavior are of verylow specificity, highly conserved across spider taxa, or that mechanical cuesalone are sufficient to elicit a significant chemoexploratory response.
Schizocosa ocreata Clade
Response of male S. ocreata to conspecific female silk or silk cues ofthe closely related heterospecific, S. rovneri, was not different for any of thebehavioral parameters measured, supporting the status of these spiders asmore closely related to each other than to other species within the ocreataclade (i.e. sibling species) and likely indicating that females of these speciesdeposit nearly identical chemical cues to elicit male courtship. Silk of femaleS. crassipes educed normal chemoexploratory behavior in male S. ocreata, butactive courtship parameters differed significantly from activity on the silk ofconspecifics or S. rovneri, allowing rejection of the null hypothesis of nondis-crimination based on silk cues. Male S. ocreata required additional time tobegin courtship on S. crassipes silk, and the rate and duration of behavioralbouts were significantly less on S. crassipes silk than on silk of conspecifics orS. rovneri. The rate of courtship bouts was significantly higher than the re-sponse on any silk treatment outside of the ocreata clade and the latency to be-gin courting was significantly lower suggesting that male S. ocreata do discrim-inate S. crassipes females as more closely related than other species tested. Asdiscussed above, this may indicate that some chemical component depositedby females that is necessary to release courtship in male S. ocreata is presenton S. crassipes silk, but is only present at concentrations sufficient to elicit areduced courtship response. Schizocosa crassipes are similar in size to S. ocre-ata and S. rovneri (Dondale and Redner, 1990), but we cannot rule out thepossibility that males are responding to mechanical differences in silk such asstrand size or composition, or to cues associated with occasional fecal deposits(based on the methods used in this study). However, a qualitative or quantita-tive divergence of chemical compounds associated with silk remains the mostplausible explanation of decreased courtship response on S. crassipes silk.
Outgroups
Male S. ocreata chemoexploratory response was generally intermediatefor the saltatrix clade representative, S. saltatrix, the other lycosid outgroup,H. helluo, and the salticid outgroup, P. princeps, falling between response

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
488 Roberts and Uetz
to members of the ocreata clade and response to the Orbiculariae outgrouprepresentative (and blank control). Courtship behavior was not significantlydifferent from blank control for any of the outgroups. Interestingly, whilethe level of jerky tap activity was very low (but present) in most treatments,even on M. spinipes silk and blank control, male S. ocreata never courted inresponse to H. helluo silk. Hogna helluo are large wolf spiders and general-ist predators that will include S. ocreata in their diet (Wise and Chen, 1999;personal observation). While we attempted to reduce the potential impactof kairomones on S. ocreata behavior (see methods), these spiders may beunder high selection pressure to recognize cues (mechanical or chemical) leftbehind by significant potential predators. Pardosa milvina wolf spiders recog-nize cues associated with silk of H. helluo and respond with reduced activity,particularly to H. helluo that have previously fed on P. milvina (Persons et al.,2001). Schizocosa ocreata may respond similarly with suppressed courtshipactivity, or increased latency to court (beyond the 5 min trial period), as ameans of avoiding predation. Schizocosa ocreata also exhibit a higher rateof leg raises on the silk of H. helluo than in response to any of the othertreatments. Leg raise behavior has been previously reported as a precursorto retreat behaviors in a study of male/male agonistic interactions in thisspecies (Aspey, 1976), and it seems logical that a readiness to retreat wouldbe adaptive in the presence of cues associated with a potential predator.
Overall, these results suggest that the assumption of species-specificityof chemical signaling should be applied with caution for spiders in the fam-ily Lycosidae. Female chemical signals deposited with silk that elicit malecourtship may be conserved across closely related genera. The placement ofS. ocreata and S. rovneri as sibling species, more closely related to each otherthan to other members of the ocreata clade, still holds (Uetz and Denterlein,1979; Stratton and Uetz, 1981; Stratton et al., 1996). However, behavioralresponse of males to heterospecific female chemical signals and/or the spe-cific composition of female chemical signals may provide a useful characterto further elucidate relationships within the currently monophyletic ocreataspecies group and other monophyletic groups within the family Lycosidae.
ACKNOWLEDGMENTS
This work was supported by the American Arachnological Society(JAR), the University of Cincinnati Research Council and Department of Bi-ological Sciences (JAR), and the National Science Foundation (IBN 9906446and IBN 9414239 to GWU). The research presented here was submitted inpartial fulfillment of the requirements for completion of the PhD degree inBiological Sciences at the University of Cincinnati. We thank the Cincinnati

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Discrimination of Species-Specific Chemical Signals in a Wolf Spider 489
Nature Center, Rowe Woods, the State of California, Department of Parksand Recreation, and the Pebble Beach Corporation for permission to collectspiders on their properties. Bruce Cutler and Sean Walker identified manyof the species used in this study. Voucher specimens are on deposit at theNational Museum of Natural History in Washington, DC and in the personalcollection of the corresponding author (JAR). Many thanks to all of thegraduate and undergraduate students whom have helped in the collectionand rearing of spiders for this work. We are especially grateful to J. Rovner(for access to his reference collection), and K. Roberts, A. Cady, C. Har-ris, B. Jayne, S. Norton, K. Petren, M. Polak, J. Rovner, J. Shann, P. Taylor,S. Walker, and two anonymous reviewers for their advice and/or editorialcomments.
REFERENCES
Aspey, W. P. (1976). Wolf spider sociobiology: I. Agonistic display and dominance-subordinancerelationships in adult male Schizocosa crassipes. Behaviour 62: 102–141.
Ayyagari, L. R., and Tietjen, W. J. (1987). Preliminary isolation of male-inhibitory pheromoneof the spider Schizocosa ocreata (Araneae, Lycosidae). J. Chem. Ecol. 13: 237–244.
Barth, F. G. (2002). A Spider’s World: Senses and Behavior, Springer-Verlag, Heidelberg.Blanke, R. (1973). Nachweis von pheromonen bei netzspinnen. Die Naturwissenschaften 60: 481.Bradbury, J. W., and Vehrencamp, S. L. (1998). Principles of Animal Communication, Sinauer
Associates, Sunderland.Coddington, J. A., and Levi, H. W. (1991). Systematics and evolution of spiders (Araneae).
Annu. Rev. Ecol. Syst. 22: 565–592.Costa, F. G., and Capocasale, R. M. (1984). Lycocsa carbonelli, sp. nov.: una etoespecie sim-
patrida, sibilina de Lycosa thorelli (Keyserling) (Araneae, Lycosidae). J. Arachnol. 11:423–431.
Costa, F. G., Viera, C., and Francescoli, G. (2000). A comparative study of sexual behavior in twosynmorphic species of the genus Lycosa (Araneae, Lycosidae) and their hybrid progeny. J.Arachnol. 28: 237–240.
Crane, J. (1949). Comparative biology of salticid spiders at Rancho Grande, Venezuela, part IV.an analysis of display. Zoologica 34: 159–215.
Dobzhansky, T. (1970). Genetics of the Evolutionary Process, Columbia University Press, NewYork.
Dondale, C. D., and Redner, J. H. (1990). The Insects and Arachnids of Canada Part 17: The WolfSpiders, Nurseryweb Spiders, and Lynx Spiders of Canada and Alaska Araneae: Lycosidae,Pisauridae, and Oxyopidae, Minister of Supply and Services Canada, Ottawa, Canada.
Foelix, R. F. (1996.) Biology of Spiders, Oxford University Press, New York.Haynes, K. F., and Yeargan, K. V. (1999). Exploitation of intraspecific communication systems:
Illicit signalers and receivers. Ann. Entomol. Soc. Am. 92: 960–970.Hegdekar, B. M., and Dondale, C. D. (1969). A contact sex pheromone and some response
parameters in lycosid spiders. Can. J. Zool. 47: 1–4.Jackson, R. R. (1987). Comparative study of releaser pheromones associated with silk of jumping
spiders (Araneae, Salticidae). N. Z. J. Zool. 14: 1–10.Jackson, R. R., Clark, R. J., and Harland, D. P. (2002). Behavioural and cognitive influences of
kairomones on an araneophagic jumping spider. Behaviour 139: 749–775.Kaston, B. J. (1936). The senses involved in the courtship of some vagabond spiders. Entomol.
Am. 16: 97–167.

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
490 Roberts and Uetz
McClintock, W. J., and Uetz, G. W. (1996). Female choice and pre-existing bias: Visual cuesduring courtship in two Schizocosa wolf spiders (Araneae: Lycosidae). Anim. Behav. 52:167–181.
Miller, G. L., Stratton, G. E., Miller, P. R., and Hebets, E. (1998). Geographic variation in malecourtship behaviour and sexual isolation in wolf spiders of the genus Schizocosa. Anim.Behav. 56: 937–951.
Miyashita, T., and Hayashi, H. (1996). Volatile chemical cue elicits mating behavior of cohabitingmales of Nephila clavata (Araneae, Tetragnathidae). J. Arachnol. 24: 9–15.
Olive, C. D. (1982). Sex pheromones in two orbweaving spiders, (Araneae, Araneidae): Anexperimental field study. J. Arachnol. 10: 241–245.
Papke, M. D., Riechert, S. E., and Schulz, S. (2001). An airborne female pheromone associatedwith male attraction and courtship in a desert spider. Anim. Behav. 61: 877–886.
Persons, M. H., and Uetz, G. W. (1996). Wolf spiders vary patch residence time in the presenceof chemical cues from prey (Araneae, Lycosidae). J. Arachnol. 24: 76–79.
Persons, M. H., Walker, S. E., and Rypstra, A. L. (2002). Fitness costs and benefits of antipredatorbehavior mediated by chemotactile cues in the wolf spider Pardosa milvina (Araneae:Lycosidae). Behav. Ecol. 13: 386–392.
Persons, M. H., Walker, S. E., Rypstra, A. L., and Marshall, S. D. (2001). Wolf spider predatoravoidance tactics and survival in the presence of diet-associated predator cues (Araneae:Lycosidae). Anim. Behav. 61: 43–51.
Pollard, S. D., Macnab, A. M., and Jackson, R. R. (1987). Communication with chemicals:Pheromones and spiders. In Nentwig, W. (ed.), Ecophysilogy of Spiders, Springer-Verlag,Berlin, pp. 133–141.
Prouvost, O., Trabalon, M., Papke, M., and Schulz, S. (1999). Contact sex signals on web andcuticle of Tegenaria atrica (Araneae, Agelenidae). Arc. Insect Biochem. Physiol. 40: 194–202.
Punzo, F., and Kukoyi, O. (1997). The effects of prey chemical cues on patch residence time inthe wolf spider Trochosa parthenus (Chamberlin) (Lycosidae) and the lynx spider Oxyopessalticus Hentz (Oxyopidae). Bull. Br. Arachnol. Soc. 10: 323–326.
Richter, C. J. J., Stolting, H. C. J., and Vlijm, L. (1971). Silk production in adult females of thewolf spider Pardosa amentata (Lycosidae, Araneae). J. Zool., Lond. 165: 285–290.
Riechert, S. E., and Singer, F. D. (1995). Investigation of potential male mate choice in a monog-amous spider. Anim. Behav. 49: 715–723.
Robinson, M. H. (1982). Courtship and mating in spiders. Ann. Rev. Entomol. 27: 1–20.Ross, K., and Smith, R. L. (1979). Aspects of the courtship behavior of the black widow spider,
Latrodectus hesperus (Araneae: Theridiidae), with evidence for the existence of a contactsex pheromone. J. Arachnol. 7: 69–77.
Rovner, J. S. (1968). An analysis of display in the lycosid spider Lycosa rabida Walckenaer.Anim. Behav. 16: 358–369.
Schulz, S., and Toft, S. (1993). Identification of a sex pheromone from a spider. Science 260:1635–1637.
Searcy, L. E., Rypstra, A. L., and Persons, M. H. (1999). Airborne chemical communication inthe wolf spider Pardosa milvina. J. Chem. Ecol. 25: 2527–2533.
Shaffer, J. P. (1995). Multiple hypothesis testing. Annu. Rev. Psychol. 46: 561–584.Stratton, G. E., and Uetz, G. W. (1981). Acoustic communication and reproductive isolation in
two species of wolf spiders. Science 214: 575–577.Stratton, G. E., and Uetz, G. W. (1986). The inheritance of courtship behavior and its role as
a reproductive isolating mechanism in two species of Schizocosa wolf spiders (Araneae:Lycosidae). Evolution 40: 129–141.
Stratton, G. E., Hebets, E. A., Miller, P. R., and Miller, G. L. (1996). Pattern and duration ofcopulation in wolf spiders (Araneae, Lycosidae). J. Arachnol. 24: 186–200.
Stowe, M. K., Tumlinson, J. H., and Heath, R. R. (1987). Chemical mimicry: Bolas spiders emitcomponents of moth prey species sex pheromones. Science 236: 964–967.
Suter, R. B., and Hirscheimer, A. J. (1986). Multiple web-borne pheromones in a spider Fron-tinella pyramitela (Araneae: Linyphiidae). Anim. Behav. 34: 748–753.

P1: IKH
Journal of Insect Behavior [joib] pp1309-joir-492408 September 21, 2004 3:52 Style file version Feb 08, 2000
Discrimination of Species-Specific Chemical Signals in a Wolf Spider 491
Suter, R. B., and Renkes, G. (1982). Linyphiid spider courtship: Releaser and attractant functionsof a contact sex pheromone. Anim. Behav. 30: 714–718.
Suter, R. B., Shane, C. M., and Hirscheimer, A. J. (1987). Communication by cuticularpheromones in a linyphiid spider. J. Arachnol. 15: 157–162.
Suter, R. B., Shane, C. M., and Hirscheimer, A. J. (1989). Spider vs, spider: Frontinella pyramiteladetects Argyrodes trigonum via cuticular chemicals. J. Arachnol. 17: 237–240.
Tietjen, W. J. (1977). Dragline-following by male lycosid spiders. Psyche 84: 165–178.Tietjen, W. J., and Rovner, J. S. (1980). Trail-following behavior in two species of wolf spiders:
Sensory and etho-ecological concomitants. Anim. Behav. 28: 735–741.Tietjen, W. J., and Rovner, J. S. (1982). Chemical communication in lycosids and other spiders.
In Witt, P. N., and Rovner, J. S. (eds.), Spider Communication Mechanisms and EcologicalSignificance, Princeton University Press, Princeton, pp. 249–279.
Trabalon, M., Bagneres, A. G., and Roland, C. (1997). Contact sex signals in two sympatricspider species, Tegenaria domestica and Tegenaria pagana. J. Chem. Ecol. 23: 747–758.
Uetz, G. W. (2000). Signals and multi-modal signaling in spider communication. In Espmark,Y., Amundsen, T., and Rosenqvist, G. (eds.), Animal Signals: Signalling and Signal Designin Animal Communication, Tapir Academic Press, Trondheim, pp. 387–405.
Uetz, G. W., and Denterlein, G. (1979). Courtship behavior, habitat, and reproductive isolationin Schizocosa rovneri Uetz and Dondale (Araneae: Lycosidae). J. Arachnol. 7: 121–128.
Uetz, G. W., and Roberts, J. A. (2002). Multisensory cues and multimodal communication inspiders: Insights from video/audio playback studies. Brain Behav. Evol. 59: 222–230.
Weygoldt, P. (1977). Communication in crustaceans and arachnids. In Sebeok, T. A. (ed.), HowAnimals Communicate, Indiana University Press, Bloomington, pp. 303–333.
Wise, D. L., and Chen, B. (1999). Impact of intraguild predators on survival of a forest-floorwolf spider. Oecologia 121: 129–137.
Wyatt, T. D. (2003). Pheromones and Animal Behaviour: Communication by Smell and Taste,Cambridge University Press, Cambridge.





























