Embed Size (px)
Citation preview

Vo1.184, No. 3, 1992
May 15,1992
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 1386-I 392
S P E C I F I C A C T I V A T I O N OF A TYROSINE -~ G L Y C I N E MUTANT
OF A5-3-KETOSTEROID I S O M E R A S E BY P H E N O L S
Bob Brooks and William F. Benisek
Department of Biological Chemistry School of Medicine
The University of California Davis, California 95616
Received March 30, 1992
A key unknown still to be explored concerning the mechanism of A5-3 - ketosteroid isomerase from P s e u d o m o n a s t e s t o s t e r o n i is the extent of the proton transfer between tyrosine-14 of the enzyme and the C-3 carbonyl oxygen of the steroid substrate. This report is a preliminary study of a system we are develop- ing to allow us eventually to use a BrCnsted analysis to measure this transfer. We describe the construction of an expression vector and tyrosine-14 ~ glycine-14 mutant of the enzyme and its specific activation, in the manner of chemical res- cue, by a variety of phenolic compounds. We suggest that the binding region of phenol is very tight and that the level of activation may be a result of steric con- straints as well as of differences in the pKa' of the phenol, o ~9~2 Academic ~ ..... ~no.
The A5-3-ketosteroid isomerase (EC 5.3.3.1) from P. t e s t o s t e r o n i catalyzes a 413- 6[3 proton shift in steroid 5-ene-3-ones by a mechanism involving tyrosine-14 and
aspartate-38 (1). The hydroxyl group of Y-14 is thought to facilitate abstraction
of the 413-proton by the base, D-38, by acting either as an acid to protonate the substrate carbonyl oxygen or as a hydrogen bond donor to this oxygen (2-9). Thus, the former role of Y-14 would involve a transfer of the hydroxyl proton between it and the substrate whereas the latter role would not. The goal of our investigation is to distinguish between these two roles. Our approach is to mea- sure this transfer via a Br0nsted analysis (10, 11) of the relationship between the pKa' of various tyrosine side chain analogues and the kcat of the reaction.
Abbreviations: IPTG (isopropy113-D-thiogalactopyranoside), KSI (A5-3-ketosteroid isomerase), MOPS (3-[N-Morpholino]propanesulfonic acid), PCR (polymerase chain reaction), RF (replicative factor), TB ("Terrific broth"), WT (wild-type).
0006-291X/92 $4.00 Copyright © 1992 by Academic Press, Inc. All rights of.reproduction in any form reserved. 1386

Vol. 184, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
In this report we describe a strategy to achieve this which is based on the method of "chemical rescue" (10-19), and we report some of our initial results. In our approach, we used site-directed mutagenesis and an efficient inducible ex- pression system to generate the mutant isomerase, Y14G. In this mutant we then restored the missing tyrosine side chain functionality by providing small phenols of various pKa's to the isomerase during the catalytic reaction. We found that several phenols significantly activated the Y14G mutant isomerase whereas others had little effect. Kinetic studies showed that activation was due to an increase in kcat.
MATERIALS AND METHODS
Construction of plasmid pKSItac: We started with the cloned wild-type 375 bp gene in plasmid pKC2 (20). To this we annealed two DNA primers (Genetic Designs, Inc.)--a 36mer complementary to the 5' end of the ketosteroid isomerase coding region and a 27mer complementary to the 3' end of the gene. The large primer contained an EcoRI sequence adjacent to the ATG start codon of the steroid isomerase coding region, and the smaller primer carried a HindlII se- quence slightly overlapping the TGA stop codon of the steroid isomerase gene. We used the polymerase chain reaction (GeneAmp--Perkin Elmer Cetus) for 25 cycles of 96°C (2 min), 44°C (1 min), and 74°C (1.5 min increasing to 3 min) in a Perkin Elmer Cetus thermocycler to amplify the gene with simultaneous intro- duction of the restriction sites. We then cut the 391 bp PCR product with EcoRI and HindlII and ligated it into pKK223-3 (Pharmacia), which had been cut simi- larly. The resulting 5.0 kb construct, pKSItac, carries the vector tac promoter, inducible by IPTG, and a ribosome binding site 10 bp upstream of the ATG start codon of the isomerase gene. We confirmed the identity and fidelity of the insert by sequencing from pKSItac directly using Sequenase (United States Biochemical). We then transformed the plasmid into E. coli TG1 (Amersham).
We confirmed expression of ketosteroid isomerase from pKSItac by growing pKSItac/TG1 at 37°C, 250 rpm, in MOPS media (21) supplemented with 0.4% glucose, 1 ~g/ml thiamine, and 50 ktg/ml ampicillin. At the onset of sta- tionary phase we introduced either 0, 0.01, 0.1, 1.0, or 10 mM IPTG into the medium and allowed the cultures to incubate overnight. Growth was monitored at 420 nm (21). The next day we sonified the cells in the culture medium for 1 min and assayed the extracts. We also obtained similar induction when the cells were grown in LB or TB (22) supplemented with 25-100 mg/ml ampicillin, with IPTG added at the time the medium was inoculated with the cells. Construction of plasmid pKSItac.Y14G: We used an 18mer oligonucleotide (Protein Structure Lab, UC Davis) complementary to the region around the TAT codon of tyrosine-14 except a glycine codon, GGT, was substituted for TAT. We annealed this to M13 single-strand DNA carrying the 391 bp isomerase gene fragment and used the Bio-Rad mutagenesis kit, which is based on the method of Kunkel (23, 24), to obtain the Y14G mutation. The modified gene was excised from M13 RF DNA and ligated into pKK223-3 to create pKSItac.Y14G, the identity of which was confirmed by sequencing it directly using the TaqTrack kit from Promega.
1387

Vol. 184, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Assays of wild-type and Y14G forms of ketosteroid isomerase: TG1 cells carry- ing either pKSItac or pKSItac.Y14G were grown overnight in TB supplemented with 0.5 mM IPTG and 100 ~g/ml ampicillin. They were harvested, washed once in 10 mM tris, pH 7.0, 4°C, and then in 15.4 mM tris, pH 7.5, 1.5 mM EDTA. We sonified them for 1 min, keeping the extracts on ice. After a brief spin to pellet the debris, glycerol was added to bring the final solution to 10 mM tris (pH 7.5), 1 mM EDTA, 35% glycerol. The reactions (3 ml) were done in cuvettes containing (in order of addition to the cuvette) buffer, phenols (if any), A5-an - drostene-3,17-dione (substrate) in methanol, and 10 [tl cell extract at 30°C. The final methanol concentration was 1.7%. Extract was added to start the reaction after a background isomerization rate had been determined for 1 rain. The buffer contained 34 mM potassium phosphate, either pH 7.0 or 6.0, and 2.5 mM EDTA. The pH 6.0 assays and some of the pH 7.0 assays also contained KC1 to provide an ionic strength of 0.1. The activity of the isomerase was not affected by the presence or absence of KC1. All assays were done spectrophotometrically
by following the appearance of the product, A4-androstene-3,17-dione, at 248 nm as described previously (25). Incidentals: UV spectra were taken using a Cary 210 spectrophotometer at 30°C. Sample and reference cuvettes contained buffer, 2 mM phenol and enzyme ex- tract. Only the sample cuvette contained substrate.
Proteins were determined by a commercial Coomassie Blue dye binding method (Bio-Rad). Phenol was from Amresco; the other phenols were from Aldrich. All were of the highest purity available. All reported uncertainties in the data are + one standard deviation.
R E S U L T S AND D I S C U S S I O N
The vector, pKSItac, in E. coli TG1 cells, expressed wild-type steroid iso- merase when induced with 0.1-1 mM IPTG (26). The induced levels ranged from 24- fold to 64-fold over uninduced cells, depending on the point in the growth curve at which IPTG was added. Similarly, cells transformed with pKSItac.Y14G expressed isomerase activity in an IPTG-inducible manner. The level of isomerase activity in the latter case was 10 -5 that of wild-type. Presumably, the lower activity is due to the absence of the tyrosine-14 side chain, the hydroxyl group of which has been shown to be critical for the activity of the wild-type enzyme since kcat for Y14F isomerase is only 10 -4.7 that of wild-type
enzyme (6). The Y14G form of steroid isomerase was unstable in crude cell extract. In
earlier experiments the activity was nearly gone 24 hours after preparation of the extract. However, we found that extracts that included 25-50% glycerol or 30 mM phenol were stable for at least one week. For the results reported herein all extracts were stabilized by 35% glycerol.
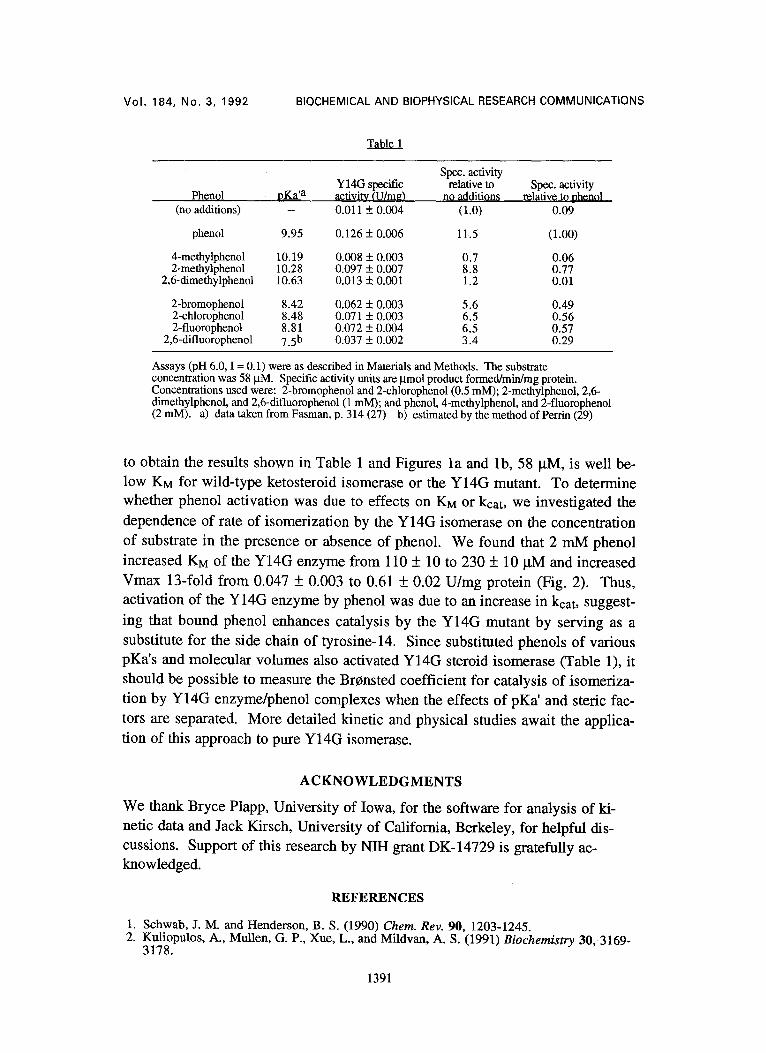
We investigated the effects of phenol and various substituted phenols on the specific activity of the Y14G mutant isomerase in a single crude cell extract. We found that 2 mM phenol increased the specific activity by about ten-fold (Table
1388
Vol. 184, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
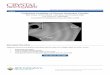
1). Other phenols that are monosubstituted in the 2-position also activated the en- zyme, but to a lesser extent than did phenol itself. Interestingly, 4-methylphenol and 2,6-dimethylphenol had no significant effects on the specific activity. All of the compounds listed in Table 1 were used at concentrations at which their acti- vating effects were maximal. In separate sets of similar experiments, using a dif- ferent cell extract, 1-4 mM 4-fluorophenol activated the enzyme 4.2-fold, 0.5 mM aniline provided a two-fold activation, 0.5 mM 3,4-difluorophenol gave a two-fold activation, and 0.5 mM cyclohexanol had no effect.
By process of elimination we concluded that the increase in activity of the Y14G form of ketosteroid isomerase was consistent with a chemical rescue model in which certain phenols bind to a region in the Y14G active site and thereby re- store the hydroxyl functionality during catalysis. Various alternative explana- tions of the results were ruled out. For example, we found that activation of Y14G steroid isomerase required both substrate and crude extract containing Y14G enzyme: omitting either one el iminated the phenol activation. Furthermore, there was no phenol activation (or activity) associated with an ex- tract from TG1 containing pKK223-3 alone. These observations proved that phenol activation was not due to non-enzymatic catalysis by phenol of substrate isomerization, nor was it due to stimulation of some other enzyme in crude cell extract whose action would lead to an increase in absorbance at 248 nm. We found that activities of neither wild-type isomerase (Fig. la) nor a low activity mutant, D38L, were affected by phenol, proving that the phenol effect is specific for the Y14G mutant isomerase. We also ruled out the unlikely possibility that phenol may have been antagonizing some previously unknown inhibitor of the enzyme since there was no phenol activation of wild-type isomerase even though it was assayed in the presence of all soluble cellular components in the extract.
~ 2 0 0 0
~ 1600
1200 tO
"3
~: 400 I -
• . 1 " , • i ' ' , i , ' ' i , . , i ' . " i .
phenol . . . . . 4-methylphenol
1 . 0 • • i • • • i - • • i • | . i | • • i - i • i
0.9
0.8
._~ 0.7 • ~ 0.6 0
0.5 tO
0.4
~ 0.3
(5 0.2 4-methylphenol ; °.'F . . . . . =
0 . . I . . . l . . . l . . . l , . . l . . . i 0 , 0 1 . . . I • . • I • . . I , . * I . . , I . . . I .
0.0 1.0 2.0 3.0 4.0 5.0 6.0 0.0 1.0 2.0 3.0 4.0 5.0 6.0
Phenolic compound (raM) Phenolic compound (raM)
Figure la and lb. Assays (pH 7.0, no adjustment for ionic strength) were as described in Materials and Methods and contained either phenol, 4-methylphenol, or no additions. The substrate concentration was 58 ~tM. Error bars are + one standard deviation for replicate assays.
1389
VOI. 184, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
We observed that 1 mM pyridine and 1 mM benzene had either slight or 50% inhibitory effects, respectively, thus suggesting that the hydroxyl group was re- quired for enzyme activation. Finally, we observed that the UV spectrum of the product of Y14G + 2 mM phenol-catalyzed isomerization of AS-androstene-3,17- dione was virtually identical to that of authentic A4-androstene-3,17-dione, proving that the increase in A248 was not due to enzymatic conversion of phenol itself to some other, more strongly UV-absorbing compound.
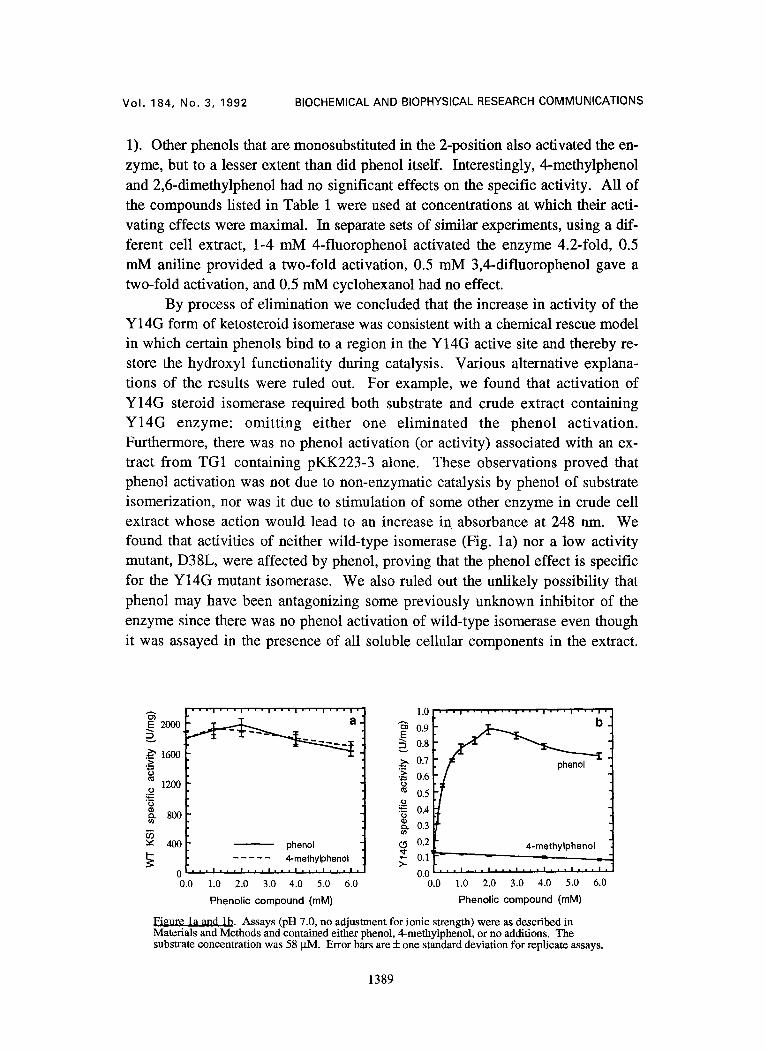
The activating effect of phenol appeared to be a saturable process specific to the Y14G mutant isomerase, a property not exhibited by 4-methylphenol on either wild-type or mutant isomerase (Fig. la and lb). Apparently, activation by phenol is non-linear, following a hyperbolic-like curve that reaches half-maximal activation at about 250 lxM phenol and maximal activation at 1-2 mM phenol (Fig. lb). These results suggest that the activating effect of phenol on the Y14G enzyme is due to a specific complex that is formed between phenol and the Y14G isomerase, and that this complex cannot be formed with the bulkier 4-methylphe- nol. We noted that the smaller 4-fluoro substituent provided significant activa- tion (see above) though less than that of phenol. The pKa' of 4-fluorophenol is nearly identical to that of phenol (27). Thus the binding site for phenols is highly sensitive to bulk in the 4-position.
The activating effect of phenol may be due either to a lowering of KM or to an augmentation of Vmax of the Y14G enzyme. The substrate concentration used
0.35 . . . . , . . . . , . . . . , . . . . , . . . . , . . . . , . . . . , . . . .
= 0.30 2
o
0.25
/ ::t .~ 0.20 a >
o.1, / / 0.10 - 1 ~ - ' - ' e
0.0 1.0 2.0 3.0
~/[s] ~- 0.05 no phenol
~ , , i . . . . i , i , , i . . . . 0.00 0 50 100 150 200 250 300 350 400
Substrate (~tM)
Figure 2. Assays (pH 7.0, I = 0.1) were as described in Materials and Methods except the
methanol concentration was 3.3%. Substrate was A5-androstene-3,17-dione. We observed no precipitation of steroid subslrate in the concentration range investigated (28-224 JAM). Kinetic data were analyzed using MS-DOS-based computer programs according to Cleland (28). Error bars are + one standard deviation for replicate assays. Inset: (o) 2 m M phenol in assay, (a) no phenol.
Horizontal axis values are 1/[S] x 102.
1390
Vol. 184, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Table 1
Spec. activity Y14G specific relative to Spec. activity
Phenol pKa 'a activitv (U/m~) no additions r~l~tive to phenol (no additions) -- 0.011 + 0.004 (1.0) 0.09
phenol 9.95 0.126 ± 0.006 11.5 (1.00)
4-methylphenol 10.19 0.008 + 0.003 0.7 0.06 2-methylphenol 10.28 0.097 + 0.007 8.8 0.77
2,6-dimethylphenol 10.63 0.013 + 0.001 1.2 0.01
2-bromophenol 8.42 0.062 + 0.003 5.6 0.49 2-chlorophenol 8.48 0.071 + 0.003 6.5 0.56 2-fluorophenol 8.81 0.072 + 0.004 6.5 0.57
2,6-difluorophenol 7.5 b 0.037 + 0.002 3.4 0.29
Assays (pH 6.0, I = 0.1) were as described in Materials and Methods. The substrate concentration was 58 ktM. Specific activity units are ~tmol product formed/mirdmg protein. Concentrations used were: 2-bromophenol and 2-chlorophenol (0.5 mM); 2-methylphenol, 2,6- dimethylphenol, and 2,6-difluorophenol (1 mM); and phenol, 4-methylphenol, and 2-fluorophenol (2 mM). a) data taken from Fasman, p. 314 (27) b) estimated by the method of Perrin (29)
to obtain the results shown in Table 1 and Figures la and lb, 58 lxM, is well be- low KM for wild-type ketosteroid isomerase or the Y14G mutant. To determine whether phenol activation was due to effects on KM or kcat, we investigated the
dependence of rate of isomerization by the Y14G isomerase on the concentration of substrate in the presence or absence of phenol. We found that 2 mM phenol increased KM of the Y14G enzyme from 110 + 10 to 230 + 10 JaM and increased Vmax 13-fold from 0.047 + 0.003 to 0.61 + 0.02 U/mg protein (Fig. 2). Thus, activation of the Y14G enzyme by phenol was due to an increase in kcat, suggest-
ing that bound phenol enhances catalysis by the Y14G mutant by serving as a substitute for the side chain of tyrosine-14. Since substituted phenols of various pKa's and molecular volumes also activated Y14G steroid isomerase (Table 1), it should be possible to measure the Br~nsted coefficient for catalysis of isomeriza- tion by Y14G enzyme/phenol complexes when the effects of pKa' and steric fac- tors are separated. More detailed kinetic and physical studies await the applica- tion of this approach to pure Y14G isomerase.
A C K N O W L E D G M E N T S
We thank Bryce Plapp, University of Iowa, for the software for analysis of ki- netic data and Jack Kirsch, University of California, Berkeley, for helpful dis- cussions. Support of this research by NIH grant DK-14729 is gratefully ac- knowledged.
REFERENCES
1. Schwab, J. M. and Henderson, B. S. (1990) Chem. Rev. 90, 1203-1245. 2. Kuliopulos, A., MuUen, G. P., Xue, L., and Mildvan, A. S. (1991) Biochemistry 30,3169-
3178.
1391

Vol. 184, No. 3, 1992 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
3. Kuliopulos, A., Talalay, P., and Mildvan, A. S. (1990) Biochemistry 29, 10271-10280. 4. Xue, L., Talalay, P., and Mildvan, A. S. (1990) Biochemistry 29, 7491-7500. 5. Bevins, C. L., Bantia, S., Pollack, R. M., Bounds, P. L., and Kayser, R. H. (1984) J. Am.
Chem. Soc. 106, 4957-4962. 6. Kuliopulos, A., Mildvan, A. S., Shortle, D., and Talalay, P. (1989) Biochemistry 28, 149-
159. 7. Malhotra, S. K. and Ringold, H. J. (1965) J. Am. Chem. Soc. 87, 3228-3236. 8. Zeng, B. and Pollack, R. M. (1991) J. Am. Chem. Soc. 113, 3838-3842. 9. Zeng, B., Bounds, P. L., Steiner, R. F., and Pollack, R. M. (1992) Biochemistry 31, 1521-
1528. 10. Toney, M. D. and Kirsch, J. F. (1989) Science 243, 1485-1488. 11. Toney, M. D. and Kirsch, J. F. (1992) Protein Science 1, 107-119. 12. Smith, H. B. and Hartman, F. C. (1991) Biochemistry 30, 5172-5177. 13. den Blaauwen, T., van de Kamp, M., and Canters, G. W. (1991) J. Am. Chem. Soc. 113,
5050-5052, 14. Ehrig, T., Hurley, T. D., Edenberg, H. J., and Bosron, W. F. (1991) Biochemistry 30,
1062-1068, 15. Eriksson, A. E., Baase, W. A., Zhang, X. J., Heinz, D. W., Blaber, M., Baldwin, E. P.,
and Matthews, B. W. (1992) Science 255, 178-183. 16. Kirsch, J. F., Toney, M. D., and Goldberg, J. M. (1990) in Protein and Pharmaceutical
Engineering, pp 105-118, Wiley-Liss, Inc., New York. 17. Paranawithana, S. R., Tu, C., Laipis, P. J., and Silverman, D. N. (1990) J. Biol. Chem.
265, 22270-22274. 18. Tu, C. and Silverrnan, D. N. (1989) Biochemistry 28, 7913-7918. 19. Tu, C., Paranawithana, S. R., Jewell, D. A., Tanhauser, S. M., LoGrasso, P. V., Wynns,
G. C., Laipis, P. J., and Silverman, D. N. (1990) Biochemistry 29, 6400-6405. 20. Choi, K. Y. and Benisek, W. F. (1988) Gene 69, 121-129. 21. Neidhardt, F. C., Bloch, P. L., and Smith, D. F. (1974) J. Bacteriol. 119, 736-747. 22. Tartof, K. D. and Hobbs, C. A. (1987) Focus 9:2, 12. 23. Kunkel, T. A. (1985) Proc. Natl. Acad. Sci. U.S.A. 82, 488-492. 24. Kunkel, T. A., Roberts, J. D., and Zakour, R. A. (1987) Methods Enzymol. 154, 367-382. 25. Jarabak, R., Colvin, M., Moolgavkar, S. H., and Talalay, P. (1969) Methods Enzymol. 15,
642-651. 26. Brooks, B. and Benisek, W. F. (1991) FASEB J. 5, 1520. 27. Fasman, G. D., Eds. (1976) Handbook of Biochemistry_ and Molecular Biology, Physical and
Chemical Data, 3 rd edition, vol. 1, CRC Press, Cleveland. 28. Cleland, W. W. (1979) Methods Enzymol. 63, 103-138. 29. Perrin, D. D., Dempsey, B., and Serjeant, E. P. (1981) P--g-a Prediction for Organic Acids and
Bases, pp 1-146, Chapman and Hall, London.
1392