Embed Size (px)
Citation preview

77
GATA 10(3-4): 77-83, 1993
Specific Cloning of DNA Fragments Unique to the Dog Y Chromosome
S U S A N F L E T C H E R ,
D A V I D D A R R A G H ,
YING FAN,
M I R A N D A D. G R O U N D S ,
C H R I S T O P H E R J. F I S H E R , and
M A N F R E D W. B E I L H A R Z
A novel technique that enabled the specific cloning of a DNA fragment unique to the dog Y chromosome is de- scribed. The method involves competitive hybridization of DNA prepared from male dog lymphocytes with biotin- labeled DNA prepared from female dog lymphocytes. The biotinylated female-female and male-female hybrid DNA fragments were removed by capture with streptavidin-coated paramagnetic particles. Full-length double-stranded DNA was generated from the remaining fragments by using the Klenow fragment of DNA polymerase I, followed by direct cloning using a low-background ligation technique. Anal- ysis of putative recombinant clones derived by this method has led to the identification of a fragment that hybridizes specifically to male dog DNA. The clones were selected initially on the basis of a differential signal obtained when hybridized to dilutions of male and female dog DNA im- mobilized on neutral nylon membrane. To evaluate its suitability as a probe for trans-sexually grafted cells in transplantation studies, the fragment was labeled with di- goxigenin and hybridized in situ to male and female dog tissue sections. The clone designated number 6.2 hybrid- ized strongly to male dog nuclei. The cloning strategy employed could be extended to other studies in which com- petitive reassociation can be used to identify unique DNA sequences.
Introduction Many transplantation studies require the location, movement, and survival of donor cells to be moni- tored, making it desirable to have a cell marker that
From the Australian Neuromuscular Research Institute (S.F., Y.F.), and the Departments of Microbiology (D.D., M.W.B.) and Pathology (M.D.G.), University of Western Australia, Ned- lands; and Biotech International (C.J.F.), Bentley, Western Aus- tralia.
Address correspondence to Dr. S. Fletcher, Australian Neu- romuscular Institute, Queen Elizabeth II Medical Centre, Ned- lands, Western Australia 6009.
Received 12 March 1993; revised and accepted 6 July 1993.
can distinguish donor cells from those of the host. The ideal marker is specific, permanent, nontrans- ferable, and can be identified in situ [1]. Donor cells that have inherent genetic differences from the host ceils fulfill these criteria, but have the major disad- vantage that they may be subject to immunologic rejection. Another approach is the use of nonendo- genous genetic markers, such as those introduced by retroviral vectors; however, these may be affected by recombination events and/or alter the behavior of the cell.
Unlike many applied markers and other endoge- nous genetic markers, Y-chromosome-specific se- quences are not subject to reutilization or dilution, and can be localized to individual cell nuclei. Much of the Y chromosome consists of repeated sequences that are rarely expressed [2]. Immunologic rejection is minimal, since tissues can be trans-sexually grafted between animals of the same strain rather than be- tween strains. Of the endogenous genetic markers, Y-chromosome sequences are ideally suited to the study of multinucleated syncitia, as expression of the DNA sequence is not required and male nuclei can be readily identified by in situ hybridization [1].
Y-chromosome-specific sequences have been ex- ploited to determine gender [3-7], to analyze chi- meric animal models [8], and to assess graft survival in humans [9-13]. More recently, Y-chromosome- specific probes were used in situ to localize trans- sexually isografted cells in mice [1, 14-16] with particular reference to myoblast transplantation stud- ies.
The transfer of myoblasts has recently become of great interest as a potential therapy for myopathies, such as Duchenne muscular dystrophy (DMD). This therapy proposes to alter the genetic and phenotypic constitution of dystrophic muscle by implanting nor- mal myoblasts. There are two main animal models of DMD, the mdx mouse and the canine X-linked muscular dystrophy in golden retrievers [17-20], of which the latter more closely resembles the condition in human patients [21].
Myoblast transplantation studies were initially carried out in mice [22, 23], but are now being ex- tended to the canine X-linked model of muscular dystrophy [24]. The Y-chromosome probes currently available are extremely species specific, however, and did not show sex specificity on dog tissue [14]. It was therefore necessary to isolate dog Y-chro- mosome-specific sequences for in situ hybridization studies on trans-sexually transplanted dog myoblasts. In this report, we describe the isolation of a suitable clone by a novel procedure involving competitive
© 1993 Elsevier Science Publishing Co., Inc., 655 Avenue of the Americas, New York, NY 113010
1050-3862/93/$6.00

78
GATA 10(3-4): 77-83, 1993 S. Fletcher et al.
hybridization in which female DNA was biotiny- lated, followed by capture of unwanted autosomal and female DNA with streptavidin-coated paramag- netic particles (SAV-PMPs). The fragments enriched for male-specific DNA were then cloned directly into phagemid by a low-background ligation technique.
Subtraction hybridization has been applied to a wide range of biologic problems such as the selection of genes, the expression of which is altered in (a) the transition from quiescence to proliferation [25], (b) DNA damage [26], and (c) stages in malignant progression [27] and cell transformation [28], tumor suppressor genes [29], and the cloning of fragments absent from the DNA of a male patient with an X- chromosome deletion [30]. We describe here the use of SAV-PMPs to select for DNA sequences unique to the Y chromosome of the dog. Our method pro- vides a simple and reliable alternative to other sub- traction hybridization techniques for the positive se- lection of genes [26, 29, 31 ].
E x p e r i m e n t a l P r o c e d u r e s
Preparation of Genomic DNA Blood samples (20 ml) were collected from individ- ual male and female dogs into tubes containing 3.5 ml of acid-citrate--dextrose solution [32]. The buffy coat was washed in phosphate-buffered saline and resuspended in 10 ml of 10 mM Tris, pH 8.0, 100 mM EDTA, 0.5% SDS, and 20 txg/ml pancreatic RNase A and the mixture incubated for 1 h at 37°C. Proteinase K was added to a final concentration of 100 Ixg/ml and incubated at 50°C for 3 h. After cool- ing to room temperature, the DNA was extracted three times with phenol saturated with 0.5 M Tris, pH 8.0, twice with chloroform, and precipitated by the addition of 0.2 vol of 10 M ammonium acetate and 2 vol ethanol [32]. The precipitated DNA was washed once with 70% ethanol and resuspended in TE (10 mM Tris, pH 8.0, 1 mM EDTA).
Equal amounts of DNA prepared from eight fe- male dogs (one golden retriever and seven mixed- breed dogs) were combined and randomly sheared by sonication to a mean size of 500-2000 base pairs (bp), as determined by comparison to HindlII-cleaved h DNA on a 1% agarose gel. Sheared DNA purified from female dogs (50 p~g) was labeled with photo- biotin acetate (Bresatec) as per the manufacturer's instructions. Similarly, equal amounts of DNA, pre- pared from six male golden retrievers, were com- bined and partially digested with SA U3A 1 (Promega) to a mean size of 509-1000 bp (determined as above).
The DNA was extracted twice with Tris-equilibrated phenol (pH 8.0) and twice with chloroform as de- scribed earlier.
Subtraction Hybridization and Paramagnetic Particle Capture SAU3Al-digested DNA prepared from male dogs (2.0 ixg) and photobiotinylated DNA prepared from female dogs (30 lxg) were mixed, ethanol precipi- tated, washed with 70% ethanol, dried, and resus- pended in 10 Ixl of 0.12 M sodium phosphate buffer, pH 6.8, containing 1 mM EDTA, 0.1% SDS, and 50% deionized formamide in a siliconized microfuge tube. The mixture was overlayed with nuclease-free paraffin oil, denatured at 100°C for 5 min, and in- cubated at 42°C for 5 days.
After incubation, the hybridization mixture was diluted to 500 Ixl with 75 mM sodium chloride and 7.5 mM sodium citrate, pH 7.0 (0.5 x SSC), and added to 250 p,g Magnesphere streptavidin-coated paramagnetic particles (SAV-PMPs) (Promega) that had been washed three times with 0.5 x SSC im- mediately before use and incubated at room temper- ature for 30 min. The SAV-PMPs were captured magnetically and this process repeated with two fresh aliquots of SAV-PMPs. The DNA remaining in so- lution was precipitated by the addition of magnesium chloride and sodium acetate to final concentrations of 1 mM and 300 mM, respectively, and 2.5 vol ethanol.
The precipitated male-enriched DNA was dena- tured and hybridized for 2 days with an additional 8 p,g biotin-labeled DNA purified from female dogs as described earlier. The paramagnetic particle cap- ture of biotin-labeled hybrids, followed by ethanol precipitation, was repeated. The DNA bound to the SAV-PMPs was eluted by incubation in deionized water at 65°C for 5 min. The concentration of eluted DNA and the precipitated "male"-enriched DNA was estimated by agarose gel electrophoresis and com- parison to standards of known concentration.
Low-Background Ligation and Cloning Procedure The strategy used here was adapted from that de- scribed for cloning into h XhoI half-site arms [33]. Full-length double-stranded DNA, enriched for male- specific sequences, was generated by the addition of 50 mM Tris, pH 7.2, 10 mM magnesium sulfate, 0.1 mM DTT, 1 mM dNTPs, and 5 units of Klenow
© 1993 Elsevier Science Publishing Co., Inc., 655 Avenue of the Americas, New York, NY 10010

Dog Y-Chromosome-Specific Sequences
79
GATA 10(3-4): 77-83, 1993
to the DNA in a total volume of 20 ~1. After incu- bation at room temperature for 2 h, the Klenow was inactivated by incubation at 68°C for 10 min. The DNA was again digested with SAU3A1 and a partial end-fill reaction, incorporating dGTP and dATP, was carried out. The reaction included 50 mM Tris, pH 7.2, 10 mM magnesium sulfate, 0.1 mM DTT, 50 Ixg/ml acetylated BSA, 1 mM dGTP and dATP, and 0.5 U of Klenow enzyme (Promega) in a total volume of 10 Ixl. After incubation at room temper- ature for 15 min, the Klenow was inactivated at 68°C for 10 min, the reaction mixture was diluted to 200 Ixl with 0.3 M sodium acetate, pH 5.0, and the DNA extracted once with phenol, twice with chloroform, and precipitated by the addition of 2.5 vol ethanol.
The vector Bluescript II K/S + (Stratagene) was digested with Xhol and partially end-filled with dCTP and dTTP as described for the SAU3A1 digested DNA, extracted, and precipitated as above. The subtracted male-enriched DNA was redissolved and added to a ligation reaction in a siliconized 0.5-ml microfuge tube containing partial end-filled vector in a 1:1 mass ratio (as estimated by agarose gel electrophoresis), 30 mM Tris-HCl, pH 7.8, 10 mM magnesium chlo- ride, 10 mM DTT, 1 mM ATP, and 1 U T4 DNA ligase (Promega) in a final volume of 5 Ixl, which was overlayed with nuclease-free paraffin oil. The ligation was allowed to proceed at room temperature for 1 h and then transferred to 4°C overnight. Each ligation reaction was added to 50 Ixl electrocompetent XL-I Blue cells. Electroporation was performed at 25 p~F and 2.5 kV and recombinants selected on plates supplemented with ampicillin (50 Ixg/ml), te- tracycline (12.5 I~g/ml), Xgal (40 Ixg/ml), and iso- propylthiogalactosidase (0.5 mM). A total of 22 pu- tative recombinant clones were selected and the inserts amplified by the polymerase chain reaction (PCR) [34] using the MI3 forward and reverse sequencing primers (Biotech International), yielding only two clones containing inserts between --100 and 600 bp. The remaining 20 clones appeared to be false recom- binants or to have small inserts that were not apparent by agarose gel electrophoresis. The plasmids from the putative recombinant clones were purified by al- kaline lysis [32] and 100 ng of each was labeled with deoxycytidine 5'-[ot-32p]tdphosphate by nick trans- lation (Promega) and hybridized to slot blots of ge- nomic DNA purified from male and female dogs and sheep (0.1 ng-1.0 Ixg) in 250 mM sodium phosphate, pH 7.2, containing 50% formamide, 250 mM sodium chloride, 7% SDS, and 1 mM EDTA for 18 h at 42°C.
In situ Hybridization The putative Y-chromosome-specific clone (desig- nated clone 6.2) was labeled with digoxigenin and hybridized overnight to sections of male and female dog muscle. Paraffin-embedded tissue, which had been fixed with methanol-acetic acid (3:1) was used; otherwise, the procedure was as described by Grounds et al. [1].
Nucleotide Sequence Analysis The insert of clone 6.2 was sequenced using a Gen- esis cycle sequencing kit (Biotech International) and the products resolved on 6% acrylamide-urea gels.
Results
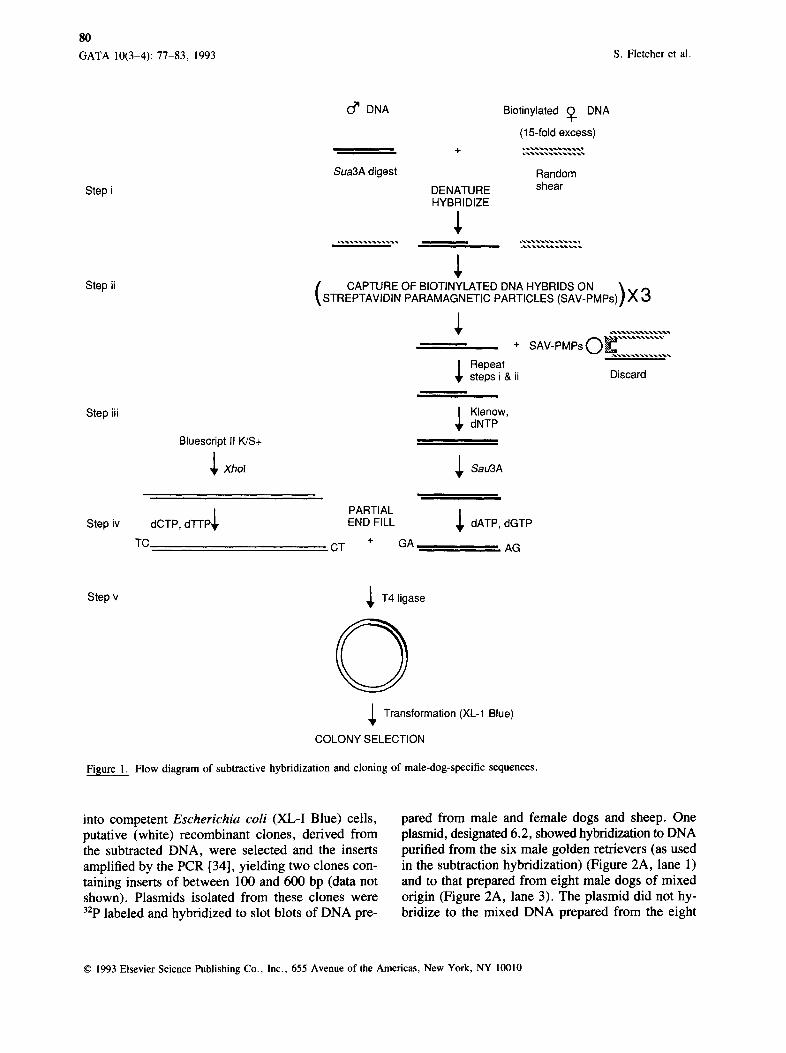
DNA Enriched for Male-Specific Sequences An overview of the competitive hybridization and cloning approaches used is summarized in Figure 1. DNA isolated from male and female dogs was den- atured and allowed to anneal as described in Exper- imental Procedures (Figure 1, step i). DNA purified from female dogs was biotin labeled to enable the binding and subsequent removal of the hybrid frag- ments, containing regions homologous to female se- quences, by streptavidin-coated paramagnetic parti- cles (Figure 1, step ii). Full-length double-stranded DNA generated from the remaining fragments was digested with SAU3A1 and partial end-filled (Figure 1, step iii) and ligated to a partial end-filled, Xhol- digested vector [33] (Figure 1, step iv). In a previous experiment in which a trace of radiolabeled DNA purified from male dogs was included, three se- quential cycles of paramagnetic particle capture led to removal of 96.5% of the labeled DNA originally present. After phenol--chloroform extraction and eth- anol precipitation, < 1% of the labeled DNA (as de- termined by scintillation counting) was recovered (data not shown).
DNA Cloning The DNA enriched for male-specific fragments was cloned into the phagemid vector Bluescript II K/S + . The partial end-fill reaction used in the cloning pro- cedure [33] ensures a low nonrecombinant back- ground. To maximize the recovery of DNA for sub- sequent transformation, all reactions with male- enriched DNA were carded out in siliconized, small- volume microfuge tubes. Following transformation
© 1993 Elsevier Science Publishing Co., Inc., 655 Avenue of the Americas, New York, NY 10010
80
GATA 10(3-4): 77-83, 1993 S. Fletcher et al.
Step i
Step ii
Step iii
0 ~ DNA
Sua3A digest
DENATURE HYBRIDIZE
Biotinylated (~. DNA
(15-fold excess)
Random shear
CAPTURE OF BIOTINYLATED DNA HYBRIDS ON \ . STREPTAVIDIN PARAMAGNETIC PARTICLES (SAV-PMPs)] X 3
÷ SAV-PMPsO . . . . . . . . . . .
~ Repeat steps i & ii Discard
Klenow, dNTP
Bluescript II K/S+
~ Xhol ~ Sau3A
Step iv ~, PARTIAL
dCTP, d'l-l'P END FILL dATP, dGTP
TC CT + GA AG
Step v
Fisure 1.
~ T 4 ligase
0 ~ Transformation (XL-1 Blue)
COLONY SELECTION
Flow diagram of subtractive hybridization and cloning of male-dog-specific sequences.
into competent Escherichia coli (XL-I Blue) cells, putative (white) recombinant clones, derived from the subtracted DNA, were selected and the inserts amplified by the PCR [34], yielding two clones con- taining inserts of between 100 and 600 bp (data not shown). Plasmids isolated from these clones were 32p labeled and hybridized to slot blots of DNA pre-
pared from male and female dogs and sheep. One plasmid, designated 6.2, showed hybridization to DNA purified from the six male golden retrievers (as used in the subtraction hybridization) (Figure 2A, lane 1) and to that prepared from eight male dogs of mixed origin (Figure 2A, lane 3). The plasmid did not hy- bridize to the mixed DNA prepared from the eight
© 1993 Elsevier Science Publishing Co., Inc., 655 Avenue of the Americas, New York, NY 10010

Dog Y-Chromosome-Specific Sequences
81
GATA 10(3-4): 77-83, 1993
Fisure 2. Hybridization of 32p-labeled clone 6.2 to male and female dog (A) and sheep (B) genomic DNA. Linearized probe was labeled with 32p dCTP by random priming and annealed to DNA prepared from male golden retrievers (blot A, lane 1 ), DNA pre- pared from female dogs of mixed breeds (lane 2 ), and DNA prepared from male dogs of mixed breeds (lane 3); and to DNA pre- pared from male and female sheep (blot B, lanes 1 and 2, respectively).
1.0
0.5
0.1
A Dog DNA
1 2 3
5.0
2.5
1.0
B Sheep DNA
4 5
female dogs (Figure 2A, lane 2) (as used in the sub- traction hybridization) or to DNA prepared from male and female sheep (Figure 2B).
In situ Hybridization
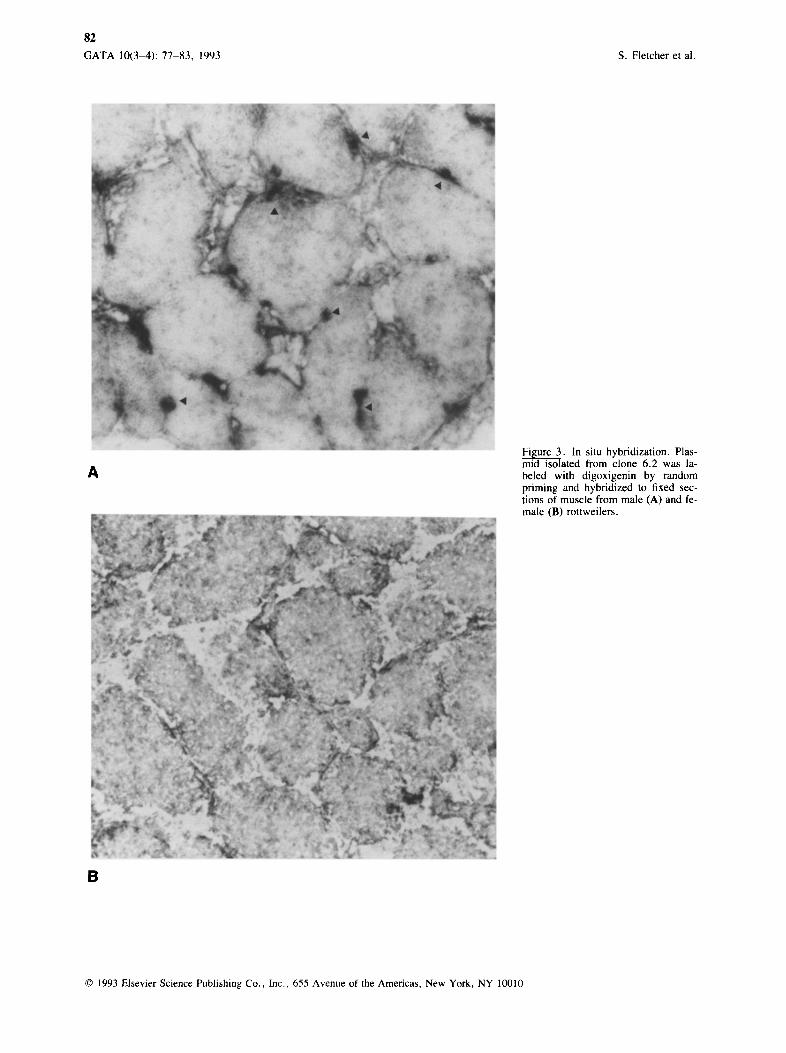
Linearized plasmid DNA isolated from clone 6.2 was digoxigenin labeled by random priming. The labeled plasmid was hybridized to sections of muscle from a male (Figure 3A) and a female (Figure 3B) Rott- weiler. The signal on nuclei in the male dog muscle section was intense, while no reaction on female dog muscle nuclei was observed.
Nucleotide Sequence Analysis
Sequence analysis revealed that clone 6.2 contains a 94-bp insert (Figure 4). A GenBank database search showed no highly probable matches to published se- quences. Although the insert contains three possible open reading frames, these are unlikely to be of sig- nificance because of the small size of the fragment.
Discussion Cloning of reassociated molecules derived from a subtraction-hybridization reaction has led to the iso- lation of DNA sequences specific for the Y chro- mosome of the dog. The methods described here are, in principle, applicable to any situation where DNA sequences in one cell type are amplified when com- pared with another.
The use of streptavidin-coated paramagnetic par- ticles to remove the hybrid DNA containing biotin- labeled fragments combines the efficiency of solution hybridization with the speed and convenience of magnetic separation. In this experiment, the number of SAV-PMPs in the first capture could have been increased, thereby eliminating one of the subsequent
separation steps. The first two captures removed equivalent amounts of DNA as determined by aga- rose gel electrophoresis, and this amount probably exceeded the binding capacity of the particles. Al- though the particles are reported to have a high bind- ing capacity for biotinylated oligonucleotides, this is likely to vary with different DNA preparations and must be determined for each specific application.
In a parallel experiment, the generation of full- length DNA was omitted after the subtraction-hy- bridization step. This method relied on perfectly reas- sociated SAU3A1 fragments being available for lig- ation into the vector, and no recombinant clones were obtained. In the successful procedure, however, complementary DNA fragments of different sizes, generated by partial SAU3A1 digestion, would be expected to prime each other. The subsequent syn- thesis of the full-length strands prior to SAU3A1 digestion and partial end fill enabled highly specific cloning.
The cloned DNA fragments obtained in several attempts at this technique were all relatively small (data not shown). Although the starting DNA was partially SAU3A1 digested to a mean size of 500- 1000 bp, the second digestion would have reduced the sizes further and ligation favors small DNA frag- ments, as would be generated by a frequently cutting enzyme such as SAU3A1.
The low-background cloning strategy makes it vir- tually impossible for the insert to ligate to itself or for the vector to close on itself. This was evidenced by the very small numbers of recombinant colonies observed and by the small number of nonrecombinant clones observed. The limited number of putative re- combinant clones obtained made it feasible to am- plify the inserts and to label the clones for differential hybridization. Dot-blot hybridization to DNA pre- pared from male and female dogs (Figure 2A) and sheep (Figure 2B) DNA and in situ hybridization
© 1993 Elsevier Science Publishing Co., Inc., 655 Avenue of the Americas, New York, NY 10010
82
GATA 10(3-4): 77-83, 1993 S. Fletcher et al.
,6, Figure 3. In situ hybridization. Plas- mid isolated from clone 6.2 was la- beled with digoxigenin by random priming and hybridized to fixed sec- tions of muscle from male (A) and fe- male (B) rottweilers.
B
© 1993 Elsevier Science Publishing Co., Inc., 655 Avenue of the Americas, New York, NY 10010

Dog Y-Chromosome-Specific Sequences
83
GATA 10(3-4): 77-83, 1993
Figure 4. Nucleotide sequence of clone 6.2.
I0 20
GATCT CGTGG CTCTT GTTCC ATATG CTAGA GCACC GAGAA CAAGG TATAC
30 40 50
GCGTC TCTCA TGGAC CTAGT AGTTC CGCAG AGAGT ACCTG GATCA TCAAG
60 70 CTGTG GTCTC CATCT GCCTC GACAC CAGAG GTAGA CGGAG
80 90 ACCTG GAACA CAAAG ACTCT GATC TGGAC CTTGT GTTTC TGAGA CTAG
(Figure 3A) indicated that clone 6.2 is a suitable probe for identifying male nuclei in transplantat ion studies in dogs.
At present, myoblast t ransplantat ion has not been very successful in large animals and the reasons for this are poorly understood. The use of the dog Y- chromosome-specif ic probe will provide precise in-
formation on the survival and movemen t of the trans- planted myoblasts in this species, essential to the development of myoblast transfer therapy in humans , and experiments in the dog model are now under
way.
This work was supported by grants from the Australian Neu- romuscular Research Foundation (M.D.G. and M.W.B.) and the National Health and Medical Research Council of Aus- tralia (M.D.G.). We also thank Dr. Barry Cooper, C.M. Lai, and K.L. Garrett for assistance with aspects of the experi- mental work.
References 1. Grounds MD, Lai MC, Fan Y, Codling JC, Beilharz MW:
Transplantation 52:1101-1105, 1992
2. Cooke H, Schmidtke J, Godsen JR: Chromosoma 87:491- 502, 1982
3. Cooke H: Nature 262:182-186, 1976
4. Nishioka Y, Lamothe E: Genetics 113:417-432, 1986
5. Nishioka Y: Teratology 38:181-185, 1988
6. Jones KW, Singh L, Edwards RG: Hum Reprod 2:439-445, 1987
7. West JD, Angell RJ, Thatcher SS, Godsen JR, Hastie ND, Glasier AF, Baird DT: Lancet 1:1345-1347, 1987
8. Singh L, Matsukuma S, Jones KW: Development 101(Suppl):143-149, 1987
9. Brain A, Purkis P, Coates P, Hackett M, Navsaria H, Leigh I: BMJ 298:917-919, 1989
10. Burt AM, Pallet CD, Sloane JP, O'Hare MJ, Schafler KF, Yardeni P, Eldad A, Clarke JA, Gusterson BA: BMJ 298:915- 917, 1989
11. Durnam DM, Anders KR, Fisher L, O'Quigley J, Bryant E, Thomas ED: Blood 74:2220--2226, 1989
12. Van Dekken H, Haganbeek A, Bauman JG: Leukemia 3:724- 726, 1989
13. Wessman M, Ruutu T, Violin L, Knuutila S: Bone Marrow Transplant 4:283-286, 1989
14. Beilharz MW, Garrett KL, Fan Y, Harvey A, Lai MC, Codling JC, Grounds MD: Proc Aust Physiol Pharmacol Soc 22:168-177, 1991
15. Coulton GR, Skynner M, Smith T, Pagel C, Partridge TA: Histochem J 23:323-327, 1991
16. Harvey AR, Fan Y, Beilharz MW, Grounds MD: Mol Brain Res 12:339-343, 1991
17. Cooper BJ, Valentine BA, Wilson S, Patterson DF, Con- cannon PW: J Hered 79:405-408, 1988
18. Cooper B J, Winand N J, Stedman H, Valentine BA, Hoffman E, Scott MO, Fishbeck KH, Kornegay JN, Avery RJ, Wil- liams JR, Schmickel R, Sylveste JE: Nature 334:154-156, 1988
19. Komegay NJ, Tuler SM, Miller DM, Levesque DC: Muscle Nerve 11:1056-1064, 1988
20. Komegay JN, Sharp NJH, Bartlett RJ, Van Camp SD, Burr CT, Hung WY, Kwock L, Roses AD: In Griggs RC, Karpati G (eds): Myoblast Transfer Therapy. New York, Plenum, 1990, pp 267-272
21. Partridge TA: Neuropathol Appl Neurobiol 17:353-363, 1991 22. Karpati G, Pouliot Y, Zubryoka-Gaarn EE, Carpenter S,
Ray PN, Worton RR, Holland P: Am J Physiol 135:1~, 1989
23. Partridge TA, Morgan JE, Coulton GR, Hoffman EP, Kunkel LM: Nature 337:176-179, 1989
24. Komegay JN, Prattis SM, Bogan DJ, Sharp NJH, Bartlett RJ, Alameddine HS, Dykstra MJ: In Kakulas BA, Howell JM, Roses AD (eds): Duchenne Muscular Dystrophy, Ani- mal Models and Genetic Manipulation. New York, Raven, 1992, pp 203-212
25. Lau LF, Nathans D: EMBO J 4:3145-3151, 1991 26. Fargnoli J, Holbrook NJ, Fomace AJ: Anal Biochem 187:364-
373, 1990 27. Dear TN, Ramshaw IA, Kefford RF: Cancer Res 48:5203-
5209, 1988 28. Scott MRD, Westphal KH, Rigby PWJ: Cell 34:557-567,
1983 29. Lee SW, Tomasetto C, Sager R: Proc Natl Acad Sci USA
88:2825-2829, 1991 30. Kunkel LM, Manoco AP, Middlesworth W, Ochs HD, Latt
SA: Proc Natl Acad Sci USA 82:4778-4782, 1985 31. Yancopeulos GD, Oltz EM, Rathbun G, Berman JE, Smith
RK, Lansford RD, Rothman P, Okada A, Lee G, Morrow M, Kaplan K, Prockop S, Air FW: Proc Natl Acad Sci USA 87:5759-5763, 1990
32. Sambrook J, Fritsch EF, Maniatis T: Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY, Cold Spring Harbor Laboratory, 1989
33. Zabarovsky ER, Allikments RL: Gene 42:119-123, 1986 34. Saiki RK, Gelfand G, Stoffel S: Science 239:487-491, 1988
© 1993 Elsevier Science Publishing Co., Inc., 655 Avenue of the Americas, New York, NY 10010





























