Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 89, pp. 9686-9690, October 1992Cell Biology
Sphingosine 1-phosphate, a specific endogenous signaling moleculecontrolling cell motility and tumor cell invasivenessYOSHITO SADAHIRA, FUQIANG RUAN, SEN-ITIROH HAKOMORI, AND YASUYUKI IGARASHIThe Biomembrane Institute, 201 Elliott Avenue West, Seattle, WA 98119; and Department of Pathobiology, University of Washington, Seattle, WA 98195
Communicated by John A. Glomset, June 23, 1992
ABSTRACT Sphingosine 1-phosphate (Sph-1-P), the ini-tial product of Sph degradation by Sph kinase, was shown tobe a strong inhibitor of cell motility and phagokinesis of B16melanoma and other types ofcells at 10-100nM concentration.It also inhibited "chemoinvasion' of tumor cells through a thicklayer of Matrigel on a filter membrane. Such inhibitory effectswere produced minimally or not at all by Sph, N-methylderivatives of Sph, or other related sphingolipids and phos-pholipids. Sph-1-P did not inhibit cell proliferation or proteinkinase C (PKC) activity, in contrast to Sph and N-methyl-Sph,which inhibit PKC activity and cell growth in general. Radio-labeled [3H]Sph and [14CJN-methyl-Sph were rapidly incorpo-rated into B16 melanoma cells. However, [14CJN-methyl-Sphwas not metabolically converted into other compounds,whereas [3H]Sph was efficiently converted within 10 min toSph-1-P, followed by conversion to other sphingoliplds andphospholipids. The inhibitory effect of Sph-1-P on cell motilityand tumor cell invasiveness could be a specific phenomenonindependent of PKC and other known transmembrane signal-ing mechanisms, based on an unknown mechanism. It maydirectly affect organizational assembly of actin filaments. Sinceexogenous Sph is rapidly converted into Sph-1-P, some re-ported effects of Sph may be ascribable to such conversion.
Sphingosine 1-phosphate (Sph-1-P) has been known for manyyears as an intermediate product during degradation of Sphby Sph kinase to ethanolamine 1-phosphate and a long-chainaldehyde (e.g., palmitaldehyde) by a pyridoxal phosphate-dependent lyase reaction (1-4). Zhang et al. (5) demonstratedthat Sph at low concentrations (s10 gM) stimulated growthof mouse 3T3 cells through a protein kinase C (PKC)-independent pathway (5); they subsequently attributed thisgrowth-stimulatory effect to formation of Sph-1-P (6). Sph orits catabolites may enhance cytoplasmic Ca2+ release (7); thiswas also attributed to Sph-1-P (6), in analogy to the effect ofinositol 1,4,5-trisphosphate on Ca2+ movement (8). AlthoughSph-1-P was assumed to induce proliferation in 3T3 cells,particularly in synergy with epidermal growth factor andinsulin (6), the physiological role of this endogenous productof Sph metabolism in cells remains unknown.We now report that Sph-1-P, either in chemically synthe-
sized form or rapidly converted from exogenous Sph,strongly and specifically inhibits chemotactic motility andinvasiveness of tumor cells at very low (nanomolar) concen-tration but does not affect cell proliferation or PKC activityeven at much higher concentrations. These findings suggestthat Sph-1-P controls motility of normal cells and invasive-ness of tumor cells, without affecting cell proliferation, viachanges in transmembrane signaling independent of the PKCpathway. In contrast, N-methyl derivatives ofSph are knownto inhibit cell proliferation, via blocking of the PKC pathwayor some unknown mechanism (9, 10).*
EXPERIMENTAL PROCEDURESCell Lines and Culture. Mouse melanoma B16 variants
(B16/F1 and B16/F10) were obtained from Isaiah J. Fidler(University of Texas M.D. Anderson Cancer Center, Hous-ton). Human HT1080 fibrosarcoma cells, cow pulmonaryartery endothelial cells (CPAEs), human melanomaMalme-3M cells, and human osteosarcoma MG-63 cells werefrom the American Type Culture Collection. Human umbil-ical vein endothelial cells (HUVECs) and their culture me-dium were purchased from Cell Systems (Kirkland, WA).Mouse BALB/c 3T3 fibroblasts were from T. Megidish ofthis institute. All these cell lines (except HUVECs) werecultured in Dulbecco's modified Eagle's medium (DMEM)supplemented with 10o fetal calf serum (FCS) (HyClone), 2mM L-glutamine, pyruvic acid (110 mg/liter), D-glucose (4.5g/liter), penicillin G (100 units/ml), and streptomycin (100pg/ml). Mouse stromal endothelial cell line SPE-1 was es-tablished from C3H/He mouse spleen culture (Y.S., unpub-lished data) and maintained in RPMI 1640 medium supple-mented with 10% FCS.
Preparation of Sph Derivatives and Other Lipids. Sph-1-Pwas prepared both enzymatically and chemically. Enzymaticpreparation was by hydrolysis of sphingosylphosphocholineby Streptomyces chromofuscus phospholipase D as de-scribed (11) (both reagents were obtained from Sigma).Sph-1-P was chemically synthesized [Weiss (12) succeeded insynthesizing sphinganine 1-phosphate, but failed to synthe-size sphingenin 1-phosphate] from 1-0- and N-protected Sphby acylating 3-OH with pivaloyl chloride. Then 2-N-t-Boc-3-O-pivaloyl-D-erythro-Sph was prepared by selective depro-tection of the primary OH group with p-toluenesulfonic acid(tosyl) chloride, followed by phosphorylation of the C1primary OH group. Deprotection of the C3 allytic OH groupand primary NH2 gave the desired Sph-1-P. The enzymati-cally synthesized product was a mixture of L-threo andD-erythro isomers, whereas the chemically synthesized prod-uct consisted only of D-erythro isomer, according to NMRdata (27). Enzymatically and chemically synthesized Sph-1-Pshowed identical mass spectra and indistinguishable biolog-ical properties. In this study, therefore, we used enzymati-cally synthesized Sph-l-P. Sph, N, N-dimethylsphingosine(DMS), and N,N,N-trimethylsphingosine (TMS) were pre-pared as described (13, 14). N-acetyl-Sph and C8 ceramide(Cer) (N-octanoyl-Sph) were synthesized as described (15).These preparations were free of endotoxin as determined bya Limulus amebocyte lysate assay kit (sensitivity limit, 0.125ng/ml; Associates of Cape Cod, Woods Hole, MA). Capnine
Abbreviations: Cer, ceramide; CM, conditioned medium; CMH,ceramide monohexoside; CPAE, cow pulmonary artery endothelialcell; DMS, N,N-dimethylsphingosine; FCS, fetal calf serum; GSL,glycosphingolipid; HUVEC, human umbilical vein endothelial cell;PDMP, 1-phenyl-2-decanoylamino-3-morpholino-1-propanol; PE,phosphatidylethanolamine; PKC, protein kinase C; SM, sphingomy-elin; Sph, sphingosine; Sph-1-P, sphingosine 1-phosphate; TMS,NNN-trimethylsphingosine.*This is paper no. IV in the series Molecular Mechanisms of CellMotility. Paper no. III is ref. 24.
9686
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 89 (1992) 9687
(Sph-related sulfonolipid isolated from Capnocytophaga)(11, 16) was a gift from Walter Godchaux III (University ofConnecticutt, Storrs, CT). Other chemicals were reagentgrade and were purchased from Sigma. Sph-1-P, Sph, TMS,N-acetyl-Sph, and GM3 were dissolved in ethanol/water(1:1, vol/vol). Cer, C8-Cer, Cer-monohexoside (CMH),sphingomyelin (SM), phosphatidylethanolamine (PE), and1-phenyl-2-decanoylamino-3 -morpholino- 1-propanol(PDMP) were dissolved in ethanol. All these compoundswere maintained as 2 mM stock solutions and were appro-priately diluted in culture medium containing 2% FCS. Con-trol experiments were performed with ethanol (<0.5%) asvehicle, which did not affect cell motility. [1-3H]Sph wassynthesized as described (14). [14C]TMS (N,N,N-trimethyl-[N-14C]Sph) was prepared from DMS with 14CH31 and K2CO3reacted in MeOH solution.
Chemotactic Cell Motility Assay and Chemoinvasion AssayUsing Transwell Plates. The ability of cells to penetrateMatrigel (Collaborative Research) coated on polycarbonatefilter membranes is a parameter frequently used in chemoin-vasion studies (e.g., see refs. 17-19). However, it depends onthe quantity of Matrigel applied and on the incubation time.In this context, two general phenotypic parameters can bedistinguished. When the Matrigel coating is thin (i.e., in thisstudy, 1 jug per filter) and incubation time is short (20 hr),migration of cells across the filter reflects random anddirected cell motility (classically known as chemotactic mo-tility), as confirmed by preliminary experiments using"checkerboard" analysis. When the Matrigel coating is thick(10 ,ug per filter) and incubation time is long (-3 days),migration of cells across the filter reflects a combination ofdirected and random cell motility plus the ability of cells tocause matrix degradation (classically known as invasive-ness).The upper chamber of transwell plates with polycarbonate
filters (diameter, 6.5 mm; pore size, 8 Am) (Costar) wasprecoated with 1 ug or 10 ,ug of Matrigel per filter. Cells(100-pl suspension), harvested after brief trypsin treatmentand resuspended inDMEM with 2% FCS, were seeded on theprecoated upper chamber at a concentration of 5 x 105 (formotility assay) or 5 x 104 (for chemoinvasion assay) cells perml. The upper chamber was then fitted onto a lower chambercontaining 0.6 ml of conditioned medium (CM) and varioussphingolipids or other test compounds. (CM consisted ofDMEM with 2% FCS in which murine splenic stromalendothelial SPE-1 cells had been cultured for 48 hr untilconfluence.) The assembly was placed in a CO2 incubator at37°C for 20 hr (motility assay) or 72 hr (chemoinvasion assay).After incubation, cells remaining in the upper chamber werewiped off with a cotton swab, and cells that had migrated tothe lower chamber side of the filter were fixed in MeOH for30 sec and stained with 0.05% toluidine blue. The filter wasremoved, the stain was solubilized in 10%o acetic acid (0.1 mlfor invasion assay, 0.5 ml for motility assay), and colorintensity (optical density) was quantitated by an ELISAreader at 630 nm. A linear relationship was found between thenumber of cells migrating to the lower chamber and theoptical density (data not shown).
Phagokinetic Assay Using Gold Sol-Coated Plates. Randomcell motility and phagocytotic activity were jointly estimatedas the area of phagokinetic track on gold sol particle-coatedglass plates as described (20). Phagokinetic tracks wereobserved on a television monitor connected to a light micro-scope (Nikon). Tracks were traced from the screen ontotransparent sheets, which were then photocopied.Phagokinetic activity was quantitated by cutting out andweighing the cleared area.
Effect of Sph Derivatives on PKC Activity and Cell Growth.PKC from F1 cells was partially purified by the method ofKreutter et al. (21) and assayed by the liposome method with
or without Sph and its derivatives as effectors as described(13). The value without phosphatidylserine, 1,2-diolein, andCa2+ was used as a reference blank. For studies on cellgrowth inhibition, 105 F1 cells were seeded on 35-mm plasticplates and incubated in DMEM plus 2% FCS and Sphderivatives at various concentrations. After 3 days, cellnumber was counted and cell viability was checked (bytrypan blue exclusion assay) with a hemocytometer.Metabolism of [3H]Sph in Melanoma Cells. F1 cells (5 X 105)
were cultured in 2 ml ofDMEM supplemented with 10% FCSin 35-mm plastic plates for 20 hr, and the medium wasdiscarded. The cells were then incubated in 1 ml of DMEMcontaining 2% FCS and 1 gM [3H]Sph (8.8 x 104 cpm) or 1,UM [14C]TMS (2.6 x 104 cpm). After various periods, me-dium and cells (collected by rubber scraper) were combinedand extracted as described (22) with minor modification. Theupper fraction, passed through a C18 column (Bond Elute),and the lower fraction were dried and dissolved in MeOH orCHC13/MeOH (2:1) and then applied to TLC plates anddeveloped in CHC13/MeOH/water (65:35:8), CHC13/MeOH/NH4OH (80:20:2), or butanol/acetic acid/water(3:1:1). The bands were identified by staining the controllipids with primulin and were visualized under UV light.After enhancer (Resolution TLC; L.M., Chestnut Hill, MA)treatment of TLC plates, autoradiography was performedwith Kodak X-Omat film at -80'C for 1-14 days.
RESULTSInhibition of Chemotactic Cell Motility and Chemoinvasion
by Sph-1-P. Under chemotactic motility assay conditions,significant inhibition of mouse melanoma F1 cells was pro-duced by Sph-1-P in a dose-dependent manner (Fig. 1A;Table 1). Since F1 and F10 cells are more susceptible toSph-1-P than other types of cells (see below), kinetic studieson inhibition of chemotactic motility and chemoinvasionwere performed with F1 cells. Motility of F1 cells wasinhibited 90% and 100% by 10 and 100 nM concentrations,respectively. Interestingly, at higher concentrations (2-5,LM) Sph-1-P showed relatively weak inhibition. Chemicallysynthesized Sph-1-P (D-erythro) showed a similar inhibitorydose curve.Sph produced significant motility inhibition of B16 cells
only at concentrations of 1 ,uM or more; 10 ,uM was requiredfor complete blocking. This was much higher than the cor-responding concentration of Sph-1-P. TMS produced onlyslight inhibition of motility (Fig. 1A), in contrast to its strongin vitro and in vivo growth-inhibitory effect, reflecting astrong inhibitory effect on PKC activity (ref. 10; see below).Under chemoinvasion assay conditions, the inhibitory
effect of Sph-1-P was again much greater than that of Sph.Sph-1-P at 100 nM produced 80% inhibition of chemoinva-sion. TMS (which had a negligible effect on chemotacticmotility) significantly inhibited chemoinvasion at 10 ,uM (Fig.1B), suggesting that TMS has no effect on cell motility butdoes inhibit the cellular enzymatic activity responsible fordegradation of Matrigel. Susceptibility of motility and inva-sion to Sph-1-P was very high for F1 and F10 cells, but othertumor cell lines and nontumorigenic 3T3 cells also showedmoderate or relatively low susceptibility. In contrast, motil-ity of some cells (e.g., endothelial cell line CPAE) wascompletely unaffected by Sph-1-P (Table 1).Uptake and Metabolic Conversion of Sph vs. TMS. While
Sph-1-P has been identified as a first-step degradation prod-uct of Sph by Sph kinase (1-4), the metabolic fate of Sph intumor cells is of great interest in view of the pronouncedantimotility activity of certain tumor cells (e.g., B16 mela-noma). Both [3H]Sph and [14C]TMS were rapidly and equallyincorporated into F1 cells (-70% during 2- to 3-hr incuba-tion). However, only Sph was rapidly converted into Sph-1-P
Cell Biology: Sadahira et al.
Proc. Natl. Acad. Sci. USA 89 (1992)
A0
V
0
.2'
0
B
02
o 1
0
(-
0V
H
.Da)C
ACER-.
C
Sp .- l 4 -
CMH-PE_
SM7ORG
Sph Derivative (M)
Sph Derivative (M)
FIG. 1. Inhibition of mouse melanoma F1 cell motility andchemoinvasion by Sph derivatives. (A) Chemotactic motility assaythrough polycarbonate filter coated with 1 ,.g of Matrigel (see text fordetails). Sph-1-P produced significant inhibition at much lowerconcentrations (10-100 nM) than Sph or TMS. (B) Chemoinvasionassay through filter coated with 10 ug of Matrigel (see text fordetails). Again, Sph-1-P had a strong inhibitory effect at a much lowerconcentration (100 nM) than Sph and TMS. TMS was effective at 10AM.
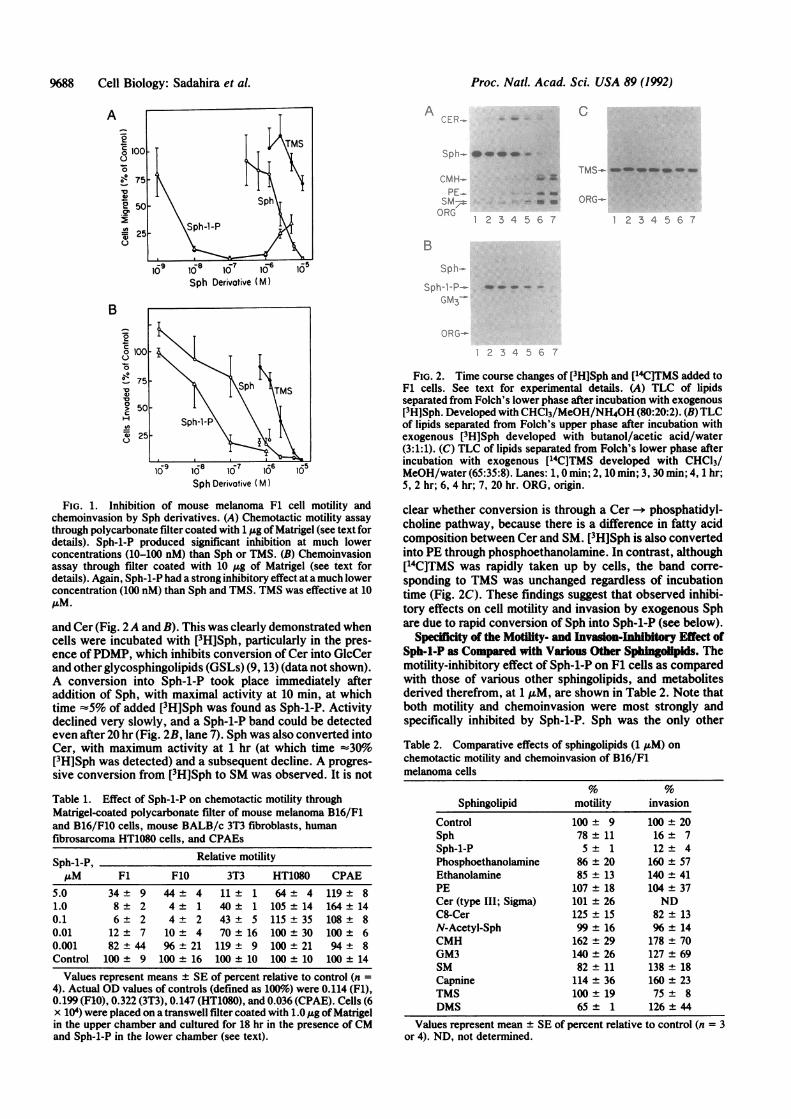
and Cer (Fig. 2A and B). This was clearly demonstrated whencells were incubated with [3H]Sph, particularly in the pres-ence ofPDMP, which inhibits conversion of Cer into GlcCerand other glycosphingolipids (GSLs) (9, 13) (data not shown).A conversion into Sph-1-P took place immediately afteraddition of Sph, with maximal activity at 10 min, at whichtime =5% of added [3H]Sph was found as Sph-1-P. Activitydeclined very slowly, and a Sph-1-P band could be detectedeven after 20 hr (Fig. 2B, lane 7). Sph was also converted intoCer, with maximum activity at 1 hr (at which time ':30%o[3H]Sph was detected) and a subsequent decline. A progres-sive conversion from [3H]Sph to SM was observed. It is not
Table 1. Effect of Sph-1-P on chemotactic motility throughMatrigel-coated polycarbonate filter of mouse melanoma B16/F1and B16/F10 cells, mouse BALB/c 3T3 fibroblasts, humanfibrosarcoma HT1080 cells, and CPAEs
Sph-1-P, Relative motility,uM F1 F10 3T3 HT1080 CPAE
5.0 34 ± 9 44 ± 4 11 1 64 ± 4 119 ± 81.0 8 ± 2 4 ± 1 40 1 105 ± 14 164 ± 140.1 6 ± 2 4 ± 2 43 5 115 ± 35 108 ± 80.01 12 ± 7 10 ± 4 70 16 100 ± 30 100 ± 60.001 82 ± 44 96 21 119 9 100 ± 21 94 ± 8Control 100 ± 9 100 ± 16 100 10 100 ± 10 100 ± 14
Values represent means + SE of percent relative to control (n =4). Actual OD values of controls (defined as 100%6) were 0.114 (Fl),0.199 (F10), 0.322 (3T3), 0.147 (HT1080), and 0.036 (CPAE). Cells (6x 104) were placed on a transwell filter coated with 1.0 ,ug of Matrigelin the upper chamber and cultured for 18 hr in the presence of CMand Sph-1-P in the lower chamber (see text).
TMS-..- inqw_ ea -m
-23-4 -5
QRG-
1 2 3 4 5 C. 7
B
Sph-
Sph-l,-P_ _ w ,,
GM3-
ORG-
2 3 4 6 7
FIG. 2. Time course changes of [3H]Sph and [14C]TMS added toF1 cells. See text for experimental details. (A) TLC of lipidsseparated from Folch's lower phase after incubation with exogenous[3H]Sph. Developed with CHC13/MeOH/NH40H (80:20:2). (B) TLCof lipids separated from Folch's upper phase after incubation withexogenous [3H]Sph developed with butanol/acetic acid/water(3:1:1). (C) TLC of lipids separated from Folch's lower phase afterincubation with exogenous [14C]TMS developed with CHCl3/MeOH/water (65:35:8). Lanes: 1, 0 min; 2, 10 min; 3, 30 min; 4, 1 hr;5, 2 hr; 6, 4 hr; 7, 20 hr. ORG, origin.
clear whether conversion is through a Cer -- phosphatidyl-choline pathway, because there is a difference in fatty acidcomposition between Cer and SM. [3H]Sph is also convertedinto PE through phosphoethanolamine. In contrast, although[14C]TMS was rapidly taken up by cells, the band corre-sponding to TMS was unchanged regardless of incubationtime (Fig. 2C). These findings suggest that observed inhibi-tory effects on cell motility and invasion by exogenous Sphare due to rapid conversion of Sph into Sph-1-P (see below).
Specificity of the Motility- and Invasion-Inhibitory Effect ofSph-1-P as Compared with Various Other Sphingdipids. Themotility-inhibitory effect of Sph-1-P on F1 cells as comparedwith those of various other sphingolipids, and metabolitesderived therefrom, at 1 ,uM, are shown in Table 2. Note thatboth motility and chemoinvasion were most strongly andspecifically inhibited by Sph-1-P. Sph was the only other
Table 2. Comparative effects of sphingolipids (1 AM) onchemotactic motility and chemoinvasion of B16/F1melanoma cells
Sphingolipid
ControlSphSph-1-PPhosphoethanolamineEthanolaminePECer (type III; Sigma)C8-CerN-Acetyl-SphCMHGM3SMCapnineTMSDMS
motility invasion
100 ± 978 ± 115± 1
86 ± 2085 ± 13107 ± 18101 ± 26125 ± 1599 ± 16162 ± 29140 ± 2682 ± 11114 ± 36100 ± 1965± 1
100 ± 2016± 712± 4
160 ± 57140 ± 41104 ± 37ND
82 ± 1396 ± 14178 ± 70127 ± 69138 ± 18160 ± 2375 ± 8126 ± 44
Values represent mean ± SE of percent relative to control (n = 3or 4). ND, not determined.
%88 Cell Biology: Sadahira et al.

Proc. Natl. Acad. Sci. USA 89 (1992) 9689
metabolically or chemically related compound showing sig-nificant inhibition at 1 AuM in the chemoinvasion assay.Interestingly, capnine (Sph-related sulfonolipid; 2-amino-3-hydroxy-15-methylhexadecane-1-sulfonic acid), which hassimilar chemical properties (e.g., solubility, charge) to Sph-1-P (11, 16), showed no inhibitory effect. The inhibitory effectof Sph in the chemoinvasion assay may involve conversion ofSph to Sph-1-P as described above.
Inhibition of Phagokinetic Activity of Tumor Cells by Sph-1-P. To confirm the antimotility effect of Sph-1-P, we usedanother experimental model: the phagokinetic track assay.
Phagokinetic activity of F10 cells on gold sol-coated plateswas inhibited by Sph-1-P (but not related compounds) atconcentrations as low as 0.01 ,uM (Fig. 3; Table 3). Similarinhibition of phagokinesis was observed with F1 cells, humanmelanoma Malme-3M cells, and human osteosarcoma MG-63cells. Interestingly, no inhibition of phagokinesis was ob-served with normal HUVECs or CPAEs (Table 3).
Effects of Sph Derivatives on PKC Activity and Growth ofF1Cells. Sph-1-P had no inhibitory effect on PKC activityisolated from F1 cells, even at 75 ,M, whereas both Sph andTMS showed strong inhibition [66% ± 7% and 84% ± 3%(mean ± SD), respectively; n = 4] at this concentration.Using the mixed-micelle assay method, Bell's group alsoreported no inhibition of PKC activity by Sph-1-P (11). TMSand Sph showed, respectively, strong (89% ± 1%) andmoderate (22% ± 4%) growth-inhibitory effects on F1 cells at10 ,uM, whereas Sph-1-P showed no growth-inhibitory effectat this or lower concentrations at which motility was stronglysuppressed. Toxicity of these compounds to F1 cells was alsoexamined by the trypan blue exclusion assay after 1 hr ofincubation with the compounds. Sph-1-P showed weak tox-icity against the cells at 45-50 ,uM, whereas Sph was verytoxic at this concentration when incubated in the absence ofbovine serum albumin (data not shown).
DISCUSSIONCell motility is an integral phenomenon in the physiologicalprocesses of development, organogenesis, and wound heal-ing, as well as pathological processes such as inflammation,and tumor invasiveness and metastasis. The molecular mech-
-.' ;*fttstS¢3
,A . B .. ^ .. C
:-..
Lo'-:X̂' :: :
A B c
FIG. 3. Inhibitory effect of Sph-1-P on phagokinetic activity ofF10 cells. Cells were plated on gold particle-coated plates in culturemedium supplemented with CM from splenic stromal endothelialcells (A), CM plus 0.1 ,uM Sph (B), or CM plus 1.0 jiM Sph-1-P (C).Photographs were taken after 18 hr. White areas are particle-free dueto particle removal by the cells. (Upper) Photographs of represen-tative cells. (Lower) Tracings of cells copied from television monitor.(Bar = 90 gm.)
Table 3. Effect of Sph, Sph-1-P, and TMS on phagokineticactivity of B16/F10 melanoma and other cells
Cleared area, % ofCell line Treatment x 103 Am2 control*B16/F10 Control 6.9 + 3.4 100
Sph (1 AM) 2.5 ± l.Ot 8Sph-1-P (1 AM) 2.4 ± l.lt 6Sph-1-P (0.1 ,M) 3.4 ± 1.3t 27Sph-1-P (0.01 AM) 4.1 ± 1.4t 42Sph-1-P (0.001 AM) 6.0 ± 2.7 81TMS (1 tiM) 5.6 ± 2.7f 73
B16/F1 Control 12.8 ± 6.4 100Sph-1-P (1 AM) 4.9 ± 2.5t 26
Malme-3M Control 8.1 ± 2.8 100Sph-1-P (1 ,M) 4.4 ± 2.9t 0
MG-63 Control 3.6 ± 1.5 100Sph-1-P (1 ,M) 2.4 ± l.Ot 10
HUVEC Control 8.4 ± 2.9 100Sph-1-P (1 ,uM) 8.3 ± 2.8 >90
CPAE Control 14.0 ± 6.1 100Sph-1-P (1 ,M) 11.8 ± 5.5 >90
*Percent motility of control, calculated by [CA-t (CM+) - CAO(CM-)/CAO (CM+) - CA0 (CM-)] x 100, where CAt is cleared areawith test substance, CA0 is cleared area without test substance, andCM+ and CM- refer to presence and absence of CM.tP < 0.001 compared to control.tP < 0.05 compared to control.
anism of cell motility is complex, being controlled by anumber of synergistic factors-e.g., growth factors and theirreceptors, adhesive proteins and their receptors (integrin andimmunoglobulin families), cytoskeletal components, andjunctional units connecting the cytoskeletal network andmembrane receptors (refs. 23 and 24 and references therein).
In an earlier study, we found that fibronectin-mediated cellmotility was highly dependent on RGDS recognition ofintegrin receptors in only 2 of 10 cell lines examined, althoughall 10 cell lines showed an equal degree of fibronectin/integrin-dependent cell adhesion. Motility in the majority ofthe cell lines, therefore, was considered to be controlled bysome unknown mechanism (23). Motility is also enhanced byan unknown signaling mechanism based on GSL recognition(e.g., GSL-GSL interaction) (24).The present study reveals an entirely different endogenous
signaling molecule, Sph-1-P, which plays a specific role incell motility. Motility of seven tumor cell lines so far exam-ined was always inhibited (although to varying degrees) bythe presence of Sph-1-P. Interestingly, motility of two endo-thelial cell lines (CPAEs and HUVECs) was totally unaf-fected by Sph-1-P. The number of cell lines we studied formotility- and phagokinesis-inhibitory effects of Sph-1-P islimited, and the inhibitory effect of Sph derivatives wastested by using cells stimulated by a common CM frommurine splenic stromal endothelial cells. Nevertheless, wehave shown that there are tumor cell types highly susceptibleand moderately susceptible, and a normal cell type (endo-thelial cells) completely nonsusceptible to Sph-1-P. Thissuggests the existence of an unknown target mechanismsensitive to Sph-1-P, which is well-developed in most tumorcells but absent in some normal cells. The antimotility andantiinvasive effect ofSph-1-P is unrelated to its effect on PKCactivity or on cell growth. In contrast, DMS and TMSstrongly inhibit PKC activity and thereby block cell prolif-eration but have less effect on cell motility.Exogenous Sph can generally be converted via three
metabolic pathways: (i) resynthesis to Cer (25) (route 1 in Fig.4), which is utilized in synthesis of SM and GSLs; (ii)N-methylation by methyltransferase (9, 25, 26) (route 2 inFig. 4); (iii) rapid conversion to Sph-1-P (refs. 1 and 6, and
Cell Biology: Sadahira et al.

9690 Cell Biology: Sadahira et al.
this work) (route 3 in Fig. 4), which is degraded into phos-phoethanolamine and palmitaldehyde. Among all these me-tabolites, only Sph-1-P (and to a much lesser extent Sph)showed significant antimotility and antiinvasive effects onthe majority of tumor cell lines so far examined. The anti-motility effect of Sph is presumably due to its rapid conver-sion to Sph-1-P. A conversion of Sph into DMS was notdemonstrated in B16 melanoma cells but has been docu-mented in CTLL cells (9) and A431 cells (28). Therefore, celltype-specific sphingolipid metabolism may help define sus-ceptibility of cell proliferation and motility to external stim-uli. Whereas DMS and its chemically synthesized analogueTMS inhibit cell motility at micromolar concentrations, andCer does not inhibit motility at any concentration, Sph-1-P iseffective at 10 nM, which is more meaningful physiologically.The relative importance of the three conversion pathwayslisted above may vary among different cell types, and sus-ceptibility of cell motility and cell growth to specific sphin-golipids also varies, presumably depending on the relativesusceptibility of yet-unidentified mechanisms controlling mo-tility and of a multiple series of protein kinases involved incell function.The target molecule or mechanism sensitive to Sph-1-P is
unknown at present. While Sph-1-P does not directly affectPKC activity, addition of 2.5-40 ng of phorbol 12-myristate13-acetate (a PKC promoter) per ml suppresses the motility-inhibitory effect of Sph-1-P in a dose-dependent manner. Themechanism underlying this phenomenon is also unknown.The target molecule of Sph-1-P could be a crucial transmem-brane receptor controlling cell motility, highly sensitive toCa2+, since Sph-1-P is reported to cause intracellular releaseof Ca2+, in analogy to inositol trisphosphate (8). The targetmolecule could also be a cytoplasmic junction protein havingsrc homology sequence-e.g., tensin (29)-linking the cyto-skeletal network and a receptor protein involved in motility(30, 31). In preliminary experiments, we have observed thatSph-1-P effectively inhibits the process of pseudopod pro-trusion (32) that occurs in melanoma cells in the presence ofCM. Thus, Sph-1-P may directly affect the organization ofactin filament meshwork.
Glycosphingollpid
11GicCeramide
+ Gic for UDP.Gc t GIC(inhibition by D-PDMP)
SphingornyelinPC j
HN sC_ §7A11 __/ Phosphoryichollne° ceramide
(1) IA OH
Fatty acid = NH,
PE
HO-P-O-HH2
Phosphoethanolamine
Sph
OH
(3) _H(2)
HiCH,)2OH
O DMS
HO-P-0l-05-CMHVOH NH,
Sph-1-P
0
PH-aIm--it H27
Palmitaidehyde
FIG. 4. Pathways for metabolic conversion of Sph. See text forexplanation. PC, phosphatidylcholine.
The motility-inhibitory effect of Sph-l-P, particularly incertain tumor cell lines, suggests the possibility that thiscompound or its mimetics could be useful agents for sup-pression of metastasis. Results ofa preliminary study supportthis idea (S. Kawa, K. Wataya, Y.S., F.R., S.H., and Y.I.,unpublished data).
The authors thank Drs. Tatsushi Toyokuni for [3H]Sph synthesis,Naoya Kojima for help with the phagokinetic assay, and StephenAnderson for scientific editing and preparation of the manuscript.This study was supported by funds from The Biomembrane Institute,in part under a research contract with Otsuka Pharmaceutical. S.H.is the recipient of National Cancer Institute Outstanding InvestigatorGrant CA42505.
1. Stoffel, W., Heimann, G. & Hellenbroich, B. (1973) Hoppe-Seyler'sZ. Physiol. Chem. 354, 562-566.
2. Stoffel, W., Hellenbroich, B. & Heimann, G. (1973) Hoppe-Seyler'sZ. Physiol. Chem. 354, 1311-1316.
3. Stoffel, W., Bauer, E. & Stahl, J. (1974) Hoppe-Seyler's Z. Physiol.Chem. 355, 61-74.
4. Louie, D. D., Kisic, A. & Schroefer, G. J., Jr. (1976)J. Biol. Chem.251, 4557-4564.
5. Zhang, H., Buckley, N. E., Gibson, K. & Spiegel, S. (1990) J. Biol.Chem. 26S, 76-81.
6. Zhang, H., Desai, N. N., Olivera, A., Seki, T., Brooker, G. &Spiegel, S. (1991) J. Cell Biol. 114, 155-167.
7. Ghosh, T. K., Bian, J. & Gill, D. L. (1990) Science 248, 1653-1656.8. Berridge, M. J. (1983) Biochem. J. 212, 849-858.9. Felding-Habermann, B., Igarashi, Y., Fenderson, B. A., Park,
L. S., Radin, N. S., Inokuchi, J., Strassmann, G., Handa, K. &Hakomori, S. (1990) Biochemistry 29, 6314-6322.
10. Endo, K., Igarashi, Y., Nisar, M., Zhou, Q. & Hakomori, S. (1991)Cancer Res. 51, 1613-1618.
11. van Veldhoven, P. P., Foglesong, R. J. & Bell, R. M. (1989)J. LipidRes. 30, 611-616.
12. Weiss, B. (1957) J. Am. Chem. Soc. 79, 5553-5557.13. Igarashi, Y., Hakomori, S., Toyokuni, T., Dean, B., Fujita, S.,
Sugimoto, M., Ogawa, T., El-Ghendy, K. & Racker, E. (1989)Biochemistry 28, 6796-6800.
14. Toyokuni, T., Nisar, M., Dean, B. & Hakomori, S. (1991) J.Labelled Comp. Radiopharm. 29, 567-574.
15. Vunnam, R. R. & Radin, N. S. (1979) Biochim. Biophys. Acta 573,73-82.
16. Godchaux, W., III, & Leadbetter, E. R. (1980) J. Bacteriol. 144,592-602.
17. Albini, A., Iwamoto, Y., Kleinman, H. K., Martin, G. R., Aaron-son, S. A., Kozlowski, J. M. & McEwan, R. N. (1987) CancerRes.47, 3239-3245.
18. Repesh, L. A. (1989) Invasion Metastasis 9, 192-208.19. Rosen, E. M., Goldberg, I. D., Liu, D., Setter, E., Donovan,
M. A., Bhargava, M., Reiss, M. & Kacinski, B. M. (1991) CancerRes. 51, 5315-5321.
20. Albrecht-Buehler, G. (1977) Cell 11, 395-404.21. Kreutter, D., Kim, J. Y. H., Goldenring, J. R., Rasmussen, H.,
Ukomadu, C., DeLorenzo, R. J. & Yu, R. K. (1987) J. Biol. Chem.262, 1633-1637.
22. Bligh, E. G. & Dyer, W. J. (1959) Can. J. Biochem. Physiol. 37,911-917.
23. Straus, A. H., Carter, W. G., Wayner, E. A. & Hakomori, S. (1989)Exp. Cell Res. 183, 126-139.
24. Kojima, N. & Hakomori, S. (1991)J. Biol. Chem. 266, 17552-17558.25. Goldkorn, T., Dressler, K. A., Muindi, J., Radin, N. S., Mendel-
sohn, J., Menaldino, D., Liotta, D. C. & Kolesnick, R. N. (1991) J.Biol. Chem. 266,16092-16097.
26. Igarashi, Y. & Hakomori, S. (1989) Biochem. Biophys. Res. Com-mun. 164, 1411-1416.
27. Ruan, F., Sadahira, Y., Hakomori, S. & Igarashi, Y. (1992) Bioorg.Med. Chem. Lett., in press.
28. Igarashi, Y., Kitamura, K., Toyokuni, T., Dean, B., Fenderson,B. A., Ogawa, T. & Hakomori, S. (1990) J. Biol. Chem. 265,5385-5389.
29. Davis, S., Lu, M. L., Lo, S. H., Lin, S., Butler, J. A., Druker,B. J., Roberts, T. M., An, Q. & Chen, L. B. (1991) Science 252,712-715.
30. Carraway, K. L. & Carraway, C. A. C. (1989) Biochim. Biophys.Acta 988, 147-171.
31. Sharift, A. & Luna, E. J. (1991) Science 256, 245-247.32. Guirguis, R., Margulies, I., Taraboletti, G., Shiffman, E. & Liotta,
L. A. (1987) Nature (London) 329, 261-263.
Proc. Natl. Acad Sci. USA 89 (1992)





























