Embed Size (px)
Citation preview

Naunyn-Schmiedeberg's Arch. exp. Path. u. Pharmak. 237, 1--10 (1959)
Aus dem Pharmakologischen Institut der Universit~t G5ttingen (Direktor: Prof. Dr. LEI~DLE)
Spontane Reaktivierung der durch Alkylphosphate (Paraoxon, Systox, Isosystox)
blockierten Cholinesterase in vitro ~ Von
K. D . FRIEDBERG ~ und W. D . ERDMANN
Mit 2 Textabbildungen
(Eingegangen am 15. Juni 1959)
Viele biologische Wirkungen der organischen Phosphors/~ureester {Alkylphosphate) erkl~ren sich daraus, dab diese Verbindungen die Cholinesterase im Organismus blockieren. Im Gegensatz zu der seit langem bekannten reversiblen Esterasehemmung durch Eserin (Physo- stigmin) oder Prostigmin (Neostigmin) gilt diese Fermentblockade als schwer oder gar nicht reversibel. Tats~chlich hemmen einige Alkylphos- phate, z. B. DFP, die Esterase so nachhaltig, dal~ man aus der Erholungs- zeit der Fermentaktivit~t in vivo Riickschliisse auf die Neubfldungsrate des Fermentes ziehen kann (vgl. z. B. VO~HAVS u. KARK 1953). Bei Ver- gfftungen re_it Parathion (Di~thyl-p-nitrophenylthiophosphat, ,,E 605"; vgl. DUBoIS u. Mitarb. 1949) oder mit Paraoxon (Dii~thyl-p-nitrophenyl- phosphat, ,,E 600"; vgl. KEWITZ 1957b) erholt sich die Ferment- aktivit/~t dagegen sehr viel schneller: hier kommt es offensichtlich zu einer Reaktivierung der blockierten Fermentmolekiile in vivo. In vitro hat man eine derartige Reaktivierung in erster Linie nach Zusatz geeig- neter ,,Reaktivatoren" beschrieben (vgl. Ubersichten bei NACttMANSOHN 1955, KEWITZ 1957b).
FRIEDBERG U. SAKAr (1958) haben aber auch nach mehrtagiger Lagerung yon Paraoxon- oder Parathion-vergifteten Tierkadavern in den Kleinhirn- und Blut- homogenaten eine sehr viel hShere Esteraseaktiviti~t gemessen als unmittelbar nach dem Tode. Es kam also zu einer spontanen Reaktivierung. Diese Erscheinung fehlte nach einer TStung der Tiere mit Systox (Di~thylthiophosphors~ureester des ~thylthioglykols, ,,E 1059") und erm5glichte damit eine li~ngere Nachweisbarkeit dieser Vergiftung.
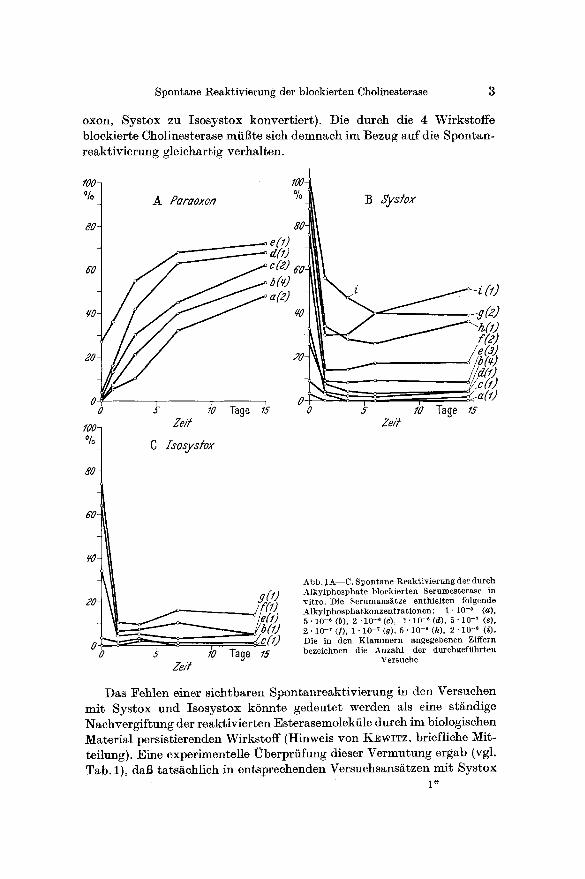
Auch im in vitro-Versuch mit Pferdeserum li~13t sich dieso spontane Reaktivierung bei der Paraoxon-Vergiftung nachweisen. Abb. 1 gibt
* Herrn Prof. Dr. W. S. LOEW% Salt Lake City, zum 75. Geburtstag ge- widme~.
** Stipendiat der Deutschen Forschungsgemeinschaft. Naunyn-Schmiedeberg's Arch. exp. Path. Pharmak., Bd. 237 1

K. D. FRIEDBERG und W. D. ERD.MAN~ :
die manomet r i sch gewonnenen Wer te der Esteraseakt iv i t / i t von Pferde-
seren wieder, denen Pa raoxon (Abb. 1 a), Systox (Abb. 1 b) oder Isosys tox (isomere Thiolform yon Systox, Abb. 1 c) zum Zei tpunkt Null zugesetzt
worden war (Aufbewahrung bei Zimmer tempera tur ) . Die Wer te sind in Prozen t der Akt iv i t i i t in den Kontro l lans~tzen des jeweiligen Pferde-
serums ausgedriickt. Die Messung der Es teraseakt iv i t / i t erfolgte nach der manomet r i schen Methode yon AMMON (1934), etwas modifiziert (aus- fiihrlich beschrieben bei FRIEDBERG U. SAKAI 1958)1.
Versuchsansatz. 19,8 ml Pferdeserum + 0,2 ml Alkylphosphat, gel6st in reinem J~thylalkohol bzw. 0,2 ml reiner ~thylalkohol fiir die Kontrollen. Bei der Ver- giftung wurde das Serum also praktiseh nicht verdiinnt. Die gepriiften End- konzentrationen der Alkylphosphate betrugen im Ansatz 1 - 10 5 bis 5. 10 -7 fiir Paraoxon, 1 • 10 -5 bis 2 - 10 s fiilr Systox und 5 - 10 s bis 1 • l0 v fiir Isosystox. Fiir die manometrische Bestimmung der Esteraseaktiviti~t wurden diese Ans~tze vor Versuchsbeginn 1 : 10 verdiinnt. Bei der Vereinigung mit dem Substrat (im Warburg- Trog) erfolgte eine weitere Verdiinnung 1 : 4.
Aus Abb. 1 a ist abzulesen, dai~ sich die durch Paraoxon blockierte Es terase innerhalb einer Beobachtungszei t bis zu 15 Tagen bis auf 50 bis
70°/0 der Normalakt ivi t i~t erholt. Die Reakt iv ie rung verli iuft besonders bei den niedrigen Konzen t ra t ionen von Paraoxon ers taunhch schnell 2.
Demgegeni iber kann in den Versuchen mi t Systox (Abb. 1 b) oder Isosy- s tox (Abb. 1 c) diese spontane Reak t iv ie rung nicht beobachte t werden.
Bei der Vergiftung des Serums mit Systox oder Isosystox nimmt die Esterase- hemmung in den ersten 24 Std sogar noch zu und verhMt sich somit gegenl~ufig zu der Paraoxon-Vergiftung. Da sieh lsosystox (vgl. Abb. 1 e) ganz so wie Systox verhiilt, kann die in den ersten Stunden zunehmende Esterasehemmung dureh Systox nicht als Ausdruck einer langsam verlaufenden Umwandlung dieses Alkyl- phosphates in vitro in seine Thiolform (,,Konversion") angesehen werden. Naeh WIRTH (1958) unterliegen beide Systox-Isorneren in vivo einer Oxydation zu den entsprechenden Sulfoxyden und Sulfonen. Derartige Umwandlungen vermSgen die zunehmende Esterasehemmung bei unseren Serumversuehen (Abb. 1 b u. e) jedoeh aueh nieht zu erkl~ren, da diese Verbindungen die Cholinesterasen in wesentlich geringerem AusmaBe hemmen als die urspriinglichen Wirkstoffe (WmT~ 1958).
Das untcrsehiedliche Verhal ten von Paraoxon und Para th ion einer- seits und Systox und Isosystox andererseits in bezug auf die spontane Reak t iv ie rung des blockier ten Fermentes in vi t ro ist bemerkenswert , da die 4 gcnannten Alkylphosphate nach der Theorie von WILSO~ und NAeHMA-XSOH~ ~ (vgl. NACHMA~SOH~ 1955, KEWITZ 1957b) den gleiehen
Di /~thylphosphoryl -Fermentkomplex bilden (wenn Para th ion zu Para-
1 Ffir die I)berlassung der Warburg-Apparatur und einer Zentrifuge danken wir der Deutschen Forschungsgemeinsehuft.
2 Auf entsprechende Versuehe mit dem Alkylphosphat Parathion wurde ver- ziehtet, weil Parathion in vitro keine und in vivo nur naeh Konversion (zu seinem biologischen Oxydationsprodukt Paraoxon) deuthche Hemmwirkungen auf die Esterase besitzt. Bei der Aufbewahrung von Blut Parathion-vergifteter Meer- sehweinchen haben wir erwartungsgem~B ebenfaUs eine spontane Reaktivierung beobachtet.
Spontane Reaktivierung der blockierten Cholinesterase
oxon, Systox zu Isosystox konvertiert). Die durch die 4 Wirkstoffo blockierte Cholinesterase miiBte sich demnach im Bezug auf die Spontan- reaktivierung gleichartig verhalten.
1007 I 1o0~ °1° t A faraoxon % B Syslox
/~------~ <us.~ ql l t 60 l , m '<,illt
! i i -b(#) . .
~,o ' / 2 / ~4 III " ~ --g(~; / / / . / L L _ - - - p "
0 .J" sO Tage 15 0 5 IO Tage 15 Ze# Ze#
/001° 2 Io C fsosys/ox
8o
60
20 . - _ , f ' ( s )
o 5 10 Tage ;'5 Ze#
A b b . 1 A - - C . S p o n t a n e 1%eak$iv ierung d e r d u r c h A l k y l p h o s p h a t e b l o c k i e r t e n S e r u m e s t e r a s e i n v i t r o . D i e S e r u m a n s ~ t z e e n t h i e l t e n f o l g e n d e A l k y l p h o s p h a t k o n z e n t r a t i o n e n : 1 "10 -a (a ) , 5 . 1 0 - * ( b ) , 2 - 1 0 - e ( c ) , 1 . 1 0 - ~ ( d ) , 5 . 1 0 -~ (e) , 2 . 1 0 -7 (]) , 1 • 10 -~ (g), 5 • 10 -8 (h), 2 - 10 - s ( i ) . D i e in d e n K l a m m e r n a n g e g e b e n e n Z i f f e r n b e z e i c h n e n d ie A n z a h l d e r d u r c h g e f i t h r t e n
V e r s u c h e
Das ~ehlen einer sichtbaren Spont~nroaktivierung in den Versuchon mit Systox und Isosystox k6nnte gedeutet werden als eine st/indigo Nachvergiftung der reaktiviorten Esterasemolokfilo dutch im biologischon Material persistierenden Wirkstoff (Hinweis yon KEWITZ, briefliche Mit- teilung). Eine experimentello Uborprfifung dieser Vermutung ergab (vgl. Tab. 1), dab tats~chlich in ontsprechenden Versuchsans/~tzen mit Systox

4 K. D. FRIEDBERG und W. D. ERI)MA~:
sogar noch n a c h 30 T a g e n f iberschfissige Mengen eines e s t e r a s e h e m m e n d e n Stoffes e n t h a l t e n sind, w/ ih rend in d e n e n m i t P a r a o x o n bis zu r e l a t i v
h o h e n K o n z e n t r a t i o n e n schon nach 24 S t d ein de r a r t i ge r N a c h w e i s n i ch t m e h r ge l ing t . D e r N a c h w e i s w u r d e m i t e iner y o n SA~:AI a u s g e a r b e i t e t e n
Tabelle 1. Nachweis von ,,paraesteratischem" Alkylphosphat im Serum nach einer Vergi/tung mit Paraoxon (2d Std) bzw. Systox (30 Tage) mit Hil/e von nativem Serum a - Aktiviti~t im Kontrollserum = 100°/0; b --Aktivit i~t im vergifteten Serum; c := Aktiviti~t in einer Mischung yon a und b. N~here Erli~uterungen : Siehe im Text !
a -i= b K o n z e n - b c - - ~ =
t r a t i o n
% °/o %
24 Std 10 5 Paraoxon-vergiftetes 10 6
Serum 10 7
i 30 Tage 10 5
Systox-vergiftetes 10 6 i
Serum 10 7
8 18 64
55 58 81
0 3 0 7 9 22
a + b - - C
54 2 59 (d. h. kein ,,freier" 82 Esterasebloeker
nachweisbar !)
50 a ÷ b . . . . . . . . . . > C
50 2 55 (d. h. ,,freier" Esterase-
blocker naehweisbar !)
M e t h o d e (in A n l e h n u n g an NACHMANSOHN U. FELD 1947, n o c h unve r - 6f fen t l ich t ) gof i ih r t :
Versuchsausatz. Verglichen wurde jeweils die Esteraseaktivitgt des unvergifteten Kontrollserums (a -- 100°/0) und die des vergifteten Serums (b) mit einer Mischung beider Seren zu gleichen Teilen (c). Wenn die Esteraseaktiviti~t im Ansatz c geringer
a @ b ist als der Wert in - ~ , so liegt im Ansatz b mit Sieherheit ein LTbersehug an
esterasehemmendem Wirkstoff vor. Mit Riicksieht auf den Vergiftungsverlauf im Serum bei Paraoxon und Systox (vgl. Abb. l a u. l b) wurden die Mischungen (Ansatz c) verschieden lang inkubiert: Paraoxon 15--60 min, Systox 16--24 Std.
D ie v o r g o l e g t e n Versuchse rgobn i s se wer fen zwei F r a g e n a u f :
1. Wie ist der erhebliche Unterschied in der Bestiindigkeit von Paraoxon und Isosystox (bzw. Systox) im Serum zu erkliiren ?
Ff i r das schnel le V e r s c h w i n d e n a u c h eines groBen P a r a o x o n - U b e r -
schusses i m S e r u m (ira Gegensa t z zu Sys tox ) lassen s ich v e r s c h i e d e n e Erk l /~ rungsmSgl i chke i t en in B e t r a c h t z iehen :
a) Es k S n n t e eine h6here H y d r o l y s e g e s c h w i n d i g k e i t oder eine st/~rkere F l i i c h t i g k e i t y o n P a r a o x o n gegenf iber S y s t o x ode r I s o s y s t o x vor l iegen .
b) Das P a r a o x o n k S n n t e ( lurch die Cho l ines te rase schnel le r v e r b r a u c h t w e r d e n als S y s t o x oder I sosys tox .
c) E i n im S e r u m vo r l i egendes F e r m e n t k 5 n n t e P a r a o x o n , n i ch t abe r S y s t o x ode r I s o s y s t o x sehr schnel l ze rs t6ren .

Spontane Reaktivierung der bloekierten Cholinesterase
Z u a. Ein Vergleich der Hydro lysegeschwind igke i t und der F l i ich t ig- ke i t yon P a r a o x o n (bzw. Pa ra th ion ) m i t den en t sprechenden W e r t e n yon Sys tox (bzw. I sosys tox) (vgl. ~([UHLMANIq U. SOHRADER 1958, HWOHT U. WIDTH 1950, WIRTK 1953 U. a.) g ib t keinen A n h a l t s p u n k t dafiir , dab P a r a o x o n au f diese Weise im Serum schneller verschwinden kSnnte als
Tabelle 2. Cholinesterase-Restaktivit6t yon P/erdeserum nach Mischen mit Rinder- serum, das verschieden lange Zeiten mit den Alkylphosphaten Paraoxon oder Systox
in steigenden Konzentrationen inkubiert wurde
1 Std 1 Tag 2 Tage 14 Tage Rinderserum lni t
' Io °Io °Io °Io
Paraoxon 1. i0 4 0 82 88 - -
1 - 1 0 - 5 0 81 96 - -
1 • 1 0 6 4 1 1 0 4 1 0 2 - -
6 Systox 1 • 10 -6 1 • 10 -7
3 18 21
7 29
Sys tox oder I sosys tox , da F l t i eh t igke i t und Hydro lysegeschwind igke i t gerade bei P a r a o x o n (und Pa ra th ion) im in teress ierenden Tempera tu r - bere ieh von 20- -40° C a m ger ings ten sind.
Z u b. Zur Pri i fung, ob die Cholinesterase fiir die schnelle Bese i t igung yon P a r a o x o n im Serum ve ran twor t l i eh ist, s ind Versuche m i t Rinder - serum geeignet , da dieses Serum keine Cholinesterase enth/t l t .
Versuchsansatz. Es wurden jeweils 19,8 ml Rinderserum mit je 0,2 ml Paraoxon oder Systox versetzt (10 2, 10 3, 10 4 und 10 -5 in reinem _~thylalkohol); die End- konzentration der Alkylphosphate betrug demnaeh jeweils 10 4, 10-~, 10-6 und 10 -7. Nach 1 Std, 24 Std, 48 Std und 14 Tagen wurden Proben des vergifteten Rinder- serums mit gleiehen Teilen unvergifteten Pferdeserums versetzt, und die Cholin- esterase-Aktivit/it dieser Ans~tze bestimmt.
Aus Tab. 2 geh t hervor , dab P a r a o x o n auch in hoher K o n z e n t r a t i o n sehr sehnell und fas t vo l l s t and ig aus dem Rinde r se rum verschwinde t . Dies is t bei Sys tox n ich t der Fal l . Ff i r die schnelle Zers t6rung y o n P a r a o x o n im Serumansa tz is t demnach die Cholinesterase n ich t ve ran t - wort l ich.
Z u c. Als F e r m e n t fiir die Zers tSrung yon P a r a o x o n im Serum k o m m t eine besonders yon AL~GUSTI~CSSON U. H~IM]3~)I~GER (1954/55) ausff ihr l ich un te r such te , ,Phosphory lphospha t a se" in Bc t raeh t . AUGUSTI~qSSON zeigte, dab es in Gegenwar t dieses F e r m e n t e s zu einer schnellen S p a l t u n g yon P a r a o x o n (und einigen anderen A lky lphospha t en ) k o m m t . Sys tox und I sosys tox (sowie einige wei tere A lky lphospha te ) werden durch das F e r m e n t n icht angegriffen. I n eigenen Versuehen k o n n t e n wir uns d a v o n fiberzeugen, dab P a r a o x o n im e rh i t z ten Pfe rdese rum (58 ° C ffir 16 S td ; F e r m e n t i n a k t i v i e r u n g ; Chol inesteraseakt ivi t /~t = Null) ebenso lange nachweisbar b le ib t wie Sys tox im na t i ven Serum (vgl. Tab. 3). Die Ver: suche wurden wie im A b s c h n i t t b (siehe oben) durchgeff ihr t .

K. D. FRIEDBERO u n d W. D. ERDMANN:
2. Wird durch die Beseitigung des Sys tox (bzw. Isosystox-)- Oberschusses im Serum eine spontane Realctivierung sichtbar, und ist diese der Erholung des Fermentes in den Paraoxonans~itzen vergleichbar, wie es nach der Phospho-
rylierungstheorie (siehe S. 2) zu /ordern wdre ?
Zur Beseit igung yon fiberschfissigem Isosystox bzw. Systox im Serum, also yon Alkylphosphat-Molekfilen, die n icht mi t der Cholinesterase reagiert haben, bieten sich methodisch mehrere M6glichkeiten an :
a) Gefr ier t rocknung mi t anschliel]ender Chloroform-Extraktion, b) Dialyse gegen unvergif tetes Serum.
Tabelle 3. Cholinesterase-Restalctivitiit von nativem P/erdeserum nach Zusatz yon erhitztem P/erdeserum, das verschieden lange Zeiten mit Paraoxon inlcubiert wurde
1 Std ! Tag 8 Tage Erhi tz tes Pferdeserum mi t % % %
Paraoxon 1 - 10 4 1 • 10 -a 1 .10 ~
2 3
10
1 1
12
0 2
69
Zu a. Eine geeignete Methode zur Beseit igung iiberschtissiger Alkyl- phospha tmengen im Gewebe ist naeh KEWITZ (1957a) sowie KEWITZ u. NACHMANSOtIN (1957) die Chloroform-Extrakt ion des Gewebes nach vor- heriger Trocknung im Vakuum (vgl. KEWITZ 1957b). Bei Uber t ragung dieser Methode auf unsore Versuche haben wir das Serum gefriergetrocknet.
Versuchsansatz. Verglichen wurde jeweils die Nachweisbarkeit yon fiber- schfissigem Wirkstoff im Paraoxon- und Isosystox-vergifteten Serum (durch Zusatz yon Kontroll-Pferdeserum wie oben beschrieben), und zwar 1. vor der Gefrier- trocknung (bei Paraoxon erfolgte der Zusatz zum unverdfinnten Serum 1 Std, bei Systox 16 Std vor der Gefriertrocknung), 2. nach der Gefriertrocknung und Wieder- aufl6sen des Trockenpr~parates und 3. nach Gefriertrocknung und anschlie[3ender 16stfindiger Chloroform-Extraktion des Trockenpr~parates.
Aus Tab. 4 geht hervor, da6 der mi t unserer Methodik nachweisbare iiberschfissige Wirkstoff in den Serumansii tzen mi t Paraoxon (5 • 10 -6) schon nach der Gefr ier t rocknung ent fern t ist, w~hrend in den Ans~tzen mi t Systox (5 • 10 -6) das Alkylphosphat auch naeh Gefrier trocknung und anschliel]ender Chloroform-Extrakt ion nachweisbar bleibt.
Auch an diesen gefriergetrockneten sowie gefriergetrockneten und chloroform- extrahierten Ansatzen wurde die Spontanreaktivierung der Esterase fiber mehrere Wochen verfolgt. Es zeigte sich wie bei den Ans~tzen mit nativem Serum (vgl. Abb. 1) eine deutliche Reaktivierung der mit Paraoxon vergifteten Cholinesterase; die mit Systox vergiftete Cholinesterase reaktivierte nicht. Auch dieser Befund l~6t sich durch einen persistierenden (,,paraesteratischen") 0berschuB von Systox erkl~ren.
Zu b. Nachdem es dureh Gefr ier t roeknung und Chloroform-Extrak- t ion nicht gelungen war, e inen Systox-tJbersehu6 im Serum zu beseitigen, wurden n u n m e h r Dialyse-Versuche durchgef/ihrt.
Spontane Reaktivierung der blockierten Cholinesteraae
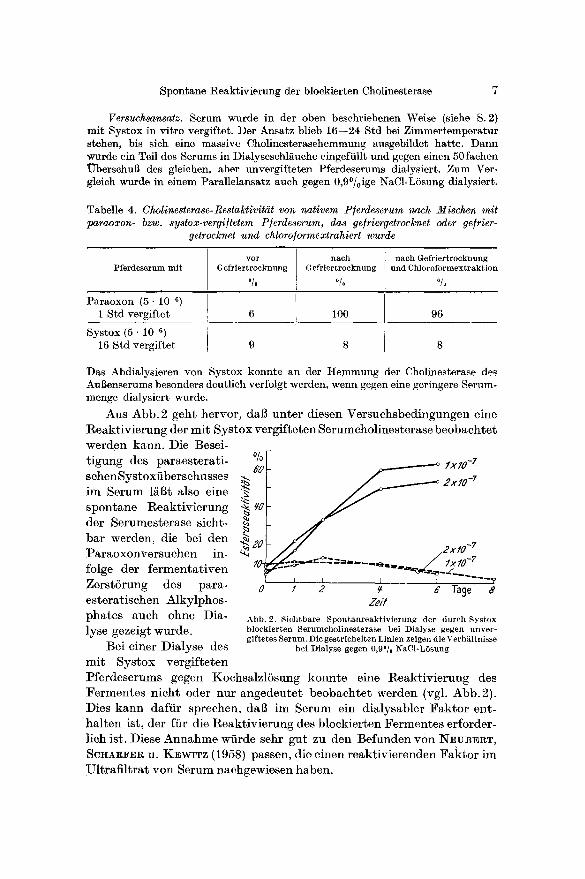
Versuchsansatz. Serum wurde in der oben beschriebenen Weise (siehe S. 2) mit Systox in vitro vergiftet. Der Ansatz blieb 16--24 Std bei Zimmertemperatur stehen, bis sich eine massive Cholinesterasehemmung ausgebfldet hatte. Dann wurde ein Teil des Serums in Dialyseschl~uche eingeftillt und gegen einen 50fachen ~berschuB des gleichen, aber unvergifteten Pferdeserums dialysiert. Zum Ver- gleich wurde in einem Parallelansatz auch gegen 0,9°/oige NaC1-LSsung dialysiert.
Tabelle 4. Cholinesterase-Restaktivitiit von nativem P/erdeserum nach Mischen mit paraoxon- bzw. systox-vergi/tetem P/erdeserum, das ge/riergetrocknet oder ge[rier-
getrocknet und chloro :ormextrahiert wurde
vor nach naeh Oefrier trocknung Pferdeserum mit Gefriertrocknung Gefriertrocknung und Ohloroformextraktion
% °h %
Paraoxon (5. lO -~) 1 Std vergiftet 6 100 96
9 8 8 Systox (5 • 10 -6)
16 Std vergiftet
])as Abdialysieren yon Systox konnte an der Hemmung der Cholinesterase des AuBenserums besonders deutlieh verfolgt werden, wenn gegen eine geringere Serum- menge dialysiert wurde.
Aus Abb. 2 geht hervor , dal~ un te r diesen Versuchsbedingungen eine Reak t iv i e rung der mi t Sys tox vergf f te ten Serumchol ines terase b e o b a c h t e t werden kann. Die Besei- t igung des pa raes t e ra t i - schenSys toxi ibersehusses im Serum l~l~t also eine spon tane l~eakt iv ie rung der Serumesterase sieht- b a r werden, die bei den Pa raoxonve r suchen in- folge der f e rmen ta t i ven Zers t6rung des para - es te ra t i schen Alky lphos - pha tes aueh ohne Dia- lyse gezeigt wurde.
B e i einer Dia lyse des mi t Sys tox verg i f te ten
% [ - r
2xlO
L I I 1 l - ' - -? 0 I 2 ~ 6' Tage #
Ze/f
Abb. 2. Sichtbare Spontaareaktivierung der durch Systox bloekierten Serumcholinesterase bei Dialyse gegen unver- giftetes Serum. Die gestrichelten Linien zeigen die Verh~iltnisse
bei Dialyse gegen 0 , 9 ° NaCI-L6sung
Pferdeserums gegen Kochsa lz lSsung konn te eine Reak t iv i e rung des F e r m e n t e s n ich t oder nur angedeu te t beoba c h t e t werden (vgl. Abb .2 ) . Dies k a n n daffir sprechen, dab im Serum ein d i a lysab le r F a k t o r ent- ha l t en ist, der fiir die Reak t iv i e rung des b lock ie r ten F e r m e n t e s erforder- l ich ist. Diese A n n a h m e wiirde sehr gut zu den Befunden yon N~UB]~T, SCHAEFER U. K•WlTZ (1958) passen, die einen r eak t iv i e renden F a k t o r im Ul t r a f i l t r a t yon Serum nachgewiesen h~ben.

K. D. FRIEDBERG und W. D. ERDMANN:
Diskussion
Es ist sicher, dab auch bei einer partiellen Alkylphosphatvergfftung ein gro6er Tell des Giftes nicht an die Cholinesterase gebunden wird : Das AusmaB der Esterasehemmung ist abh/ingig yon der Giftkonzentration. Eine sichtbare Reaktivierung des blockierten Fermentes setzt voraus, dab zun~chst die paraesteratische Giftmenge beseitigt wird. Auch im Pferde- serum in vitro ist diese MSglichkeit fiir bestimmte Alkylphosphate ge- geben : z. B. werden Paraoxon, Tabun, nicht aber Systox, Isosystox und einige andere fermentativ durch eine Phosphorylphosphatase zerst6rt (AuGuSTINSSON u. ttEIMUORC, ER 1954/55). Somit ist -- wenn fiberhaupt -- nur fiir die erstgenannte Alkylphosphatgruppe die M6glichkeit einer sichtbaren Fermentreaktivierung gegeben, es sei denn, dab man -- wie in unseren Versuehen -- auf andere Weise (durch Dialyse gegen Serum) den AlkylphosphatfibersehuB beseitigt. Nach der Theorie yon WILson u. N.¢CHMANSOHN (siehe S.2) muB gefordert werden, da6 durch Paraoxon und Systox bzw. Isosystox blockierte Cholinesterasen sich im Hinblick auf die Reaktivierung gleichartig verhalten. Dies ist nach Beseitigung der paraesteratischen Alkylphosphatmengen tats~chlich der Fall. AUGUSTINSSON zieht in Erw~gung, dab die Phosphorylphosphatase a uch direkt ffir die Reaktivierung der bloekierten Cholinesterase verantwort- lich ist, wahrend N n v ~ T , SCHAEFER U. KEWITZ (1958) hierffir einen besonderen, ultrafiltrierbaren Serumfaktor verantwortlieh machen. Unsere Dialyseversuche zeigen, dab auch die (lurch Systox blockierte Cholinesterase reaktiviert wird, obwohl die Phosphorylphosphatase mit diesem Alkylphosphat nieht zu reagieren vermag. Dies seheint daher mehr for die yon NEUBERT, SCHAEFER U. KEWITZ (1958) angeregte Be- trachtungsweise zu sprechen.
Aus unseren Untersuehungen ergeben sieh ferner einige praktisch wichtige Gesichtspunkte.
1. Bei der Beurteilung yon Alkylphosphaten pflegen besonders die eholinesterasehemmenden Eigenschaften berficksichtigt zu werden. Aus unseren Versuchen (vgl. Abb. 1) geht hervor, dab sich die Cholinesterase- Aktivit~t des vergifteten Gewebes mit der Vergiftungszeit laufend ver- /indert und je nach Alkylphosphat zunehmen oder abnehmen kann. An- gaben fiber Charakteristika yon Cholinesteraseblockern wie ihre I5o (diejenigen Konzentrationen, die gerade eine 50°/0ige Fermenthemmung bewirken) sind daher bei verschiedenen Autoren nur sehr bedingt ver- gleichbar. Es wird n6tig sein, die Prfifungsbedingungen zu standardi- sieren.
2. Bei der Einwirkung yon Alkylphosphaten auf tierisches Gewebe fiberlagern sich mehrere biochemische Prozesse, z.B. die Hemmung der Cholinesterase, die eventuelle Zerst6rung des paraesteratisehen Alkyl- phosphates durch die Phosphorylphosphatase und andere Fermente

Spontane l%eaktivierung der blockierten Cholinesterase 9
sowie eventuelle physiologische Reaktivierungsvorg/~nge. Durch die bei in vitro-Versuchen vieffach fibliche Verdfinnung yon Serum (oder ande- rem Gewebe) werden diese Reaktionen verschieden beeinfluBt. WKhrend die esterasehemmenden Eigenschaften des Alkylphosphates wohl unver- mindert nachzuweisen sein dfirften, k6nnte die Wirksamkeit der Phos- phorylphosphatase (AuGusT/NSSO~) und des serumeigenen Reaktivators (KEWITZ) durch die Verd/innung eingeschr/£nkt oder aufgehoben sein. Auch in diesem Falle haben also die methodischen Bedingungen einen groBen EinfiuB auf das Versuchsergebnis (siehe unter Punkt 1).
3. Bei der Messung der Cholinesterase-Aktivit/~t yon in vivo vergifte- tem Gewebe k6nnen paraesteratisch gespeicherte Alkylphosphatmengen das Ergebnis verf~lschen, wenn diese bei I{omogenisieren des Gewebes mit der noch aktiven Cholinesteraso reagieren. Es ist daher versucht worden, diese paraesteratischen Alkylphosphatmengen vor dem t tomo- genisieren durch Trocknung und Chloroform-Extraktion aus dem Gewebe zu entfernen (KEwITZ u. NACHMAI~SOH~¢ 1957). Unsere Versuche an systoxvergiftetem Serum zeigen, dal] dieses Verfahren nicht immer ge- eignet ist, einen UbcrschuB an Alkylphosphat zu beseitigen. Eventuell wird Systox an die Serumproteine - - vielleicht fiber Arginin (?) - - ge- bunden, so dab eine Extraktiori durch Chloroform nicht mehr m6glich ist. Andererseits scheint jodoch diese Bindung so locker zu sein, dab es bei Zusatz yon frischem Pferdeserum doch zu einer Esteraseblockierung kommt.
4. Selbst so nahe verwandte Di~thylphosphate wie Paraoxon, Systox und Isosystox k6nnen im isolierten Gewebe ganz verschieden wirksam werden : Paraoxon wird in wenigen Stunden und Tagen entgiftet, Systox und Isosystox dagegen persistieren fiber Wochen. Solche Unterschiede k6nnten unscres Erachtens ffir die Frage der kumulierenden Wirkungen yon Alkylphosphaten, also ffir die chronische Toxicitiit wichtig werden.
Z u s a n l m e n f a s s u n g •
Die durch Paraoxon blockierte Cholinesterase yon Pferdeserum zeigt eine erhebliche spontane Reaktivierung in vitro. Diese Erscheinung fehlt bei der Vergiftung des Serums mit Systox oder Isosystox. Das verschie- dene Verhalten der 3 geprfiften Di/£thylphosphate ist darauf zurfick- zuffihren, dab Paraoxon im Serum sehr schnell fermentat iv zers~Srt wird (und zwar unabh/ingig yon der Cholinesterase). Systox bzw. Isosystox bleiben dagegen im Serum lange erhalten und ffihren so zu einer sti~ndigen Nachvergfftung der reaktivierten Esterasemolekfile. Durch Gefrier- trocknung und Chloroform-Extraktion gelingt es nicht, iiberschfissiges Systox aus dem Serum zu entfernen. Nur nach Dialyse gegen unver- giftetes Serum wird auch hier eine spontane 1%eaktivierung sichtbar.

10 Fl~IEDBERG u. ERDMA~: Reaktivierung der blockierten Cholinesterase
Literatur AMMON, R.: Die fermentative Spaltung des Acetylcholins. Pfliigers Arch. ges.
Physiol. 288, 486 (1934). AVCUSTINSSON, K.-B., and G. HEIMB(~RGER: Enzymatic hydrolysis of organophos-
phorus compounds. I . - -VI . Acta chem. scand. 8, 753, 762, 915, 1533 (1954); 9, 310, 383 (1955).
DVBOm, K. P., Z. DOULL, P. R. SALERNO and J. M. Coo~: Studies on the toxicity and mechanism of action of p-nitrophenyl diethyl thionophosphate (parathion). J. Pharmaeol. exp. Ther. 95, 79 (1949).
FRIEDBERG, K. D., u. F. SAKAI : Spezifischer Nachweis von Vergiftungen mit Alkyl- phosphaten (E 600, E 605, Systox) in Blur und Hirngewebe mit Hilfe eines fermentreaktivierenden Antidots (Pyridin-Aldoxim-Methjodid, PAM). Dtsch. Z. ges. gerichtl. Med. 47,580 (1958).
•ECHT, G., u. W. WIRTH: gut Pharmakologie der Phosphorsaureester. Derivate der Thiophosphors/iure. Naunyn-Schmiedeberg's Arch. exp. Path. Pharmak. 211, 264 (1950).
KEWITZ, H. : A specific antidote against lethal alkylphosphate intoxication. I II . Repair of chemical lesion. Arch. Biochem. 66, 263 (1957a).
JtXEWITZ, H.: Die Wiederherstellung der Cholinesteraseaktivit/~t bei der Alkyl- phosphat-Vergiftung durch ein spezifisches Antidot. Klin. Wschr. 1957 b, 521.
KEWlTZ, H., and D. NACtIMA~SOHZ,~: A specific antidote against lethal alkylphos- phate intoxication. IV. Effects in brain. Arch. Biochem. 66, 271 (1957).
Mt~HLMANN, R., u. G. SCttRADER: Hydrolyse der insekticiden Phosphorsi~ureester. Z. Naturforsch. 12b, 196 (t957).
1N'ACH.~IANSOI4~ ~, D. : Die Rolle des Acetylcholins in den Elementarvorggngen der Nervenleitung. Ergebn. Physiol. 48, 575 (1955).
iXACH~ANSOItN, D., and E. A. FELD: Studies on cholinesterase. IV. On the mecha- nism of diisopropyl fluorophosphate action in vivo. J. biol. Chem. 171, 715 (t947).
NEUBERT, D., J. SCHAEFER U. H. KEWITZ: Reaktivierung der Acetylcholinesterase durch k6rpereigene Stoffe. Naturwissenschaften 45, 290 (1958).
VORHAUS, L. J., and R. M. KAI~K : Serum cholinesterase in health and disease. Amer. J. Med. 14, 707 (1953).
WmTI[, W. : Zur Pharmakologie der Phosphors~ureester. Di/~thylthiophosphor- s~ureester des _~thylthioglykol (,,Systox-Wirkstoff"). Naunyn-Schmiedeberg's Arch. exp. Path. Pharmak. 217, 144 (1953).
~VmTH, W. : Zur Wirkung System-insekticider Phosphors/iure-Ester im Warm- bliiter-Stofl~,echsel. Nsunyn-Schmiedeberg's Arch. exp. Path. Pharmak. 234, 352 (1958).
Dr. K. D. FRIEDBERG, Pharmakologisches Insti tut der Universit/it, GSttingen, GeiststraBe 9
![24546PB SV d v - MSL - Innovative Schlosstechnikmsl-lock.com/.../sFlipLock_PDF/14546PB_SV_D.pdf · · 2016-10-25Einpunktverriegelung mit 1 Riegel und 1 blockierten [Flip-] Kippfalle](https://img.pdfslide.net/doc/110x75/5ae681d07f8b9aee078d0697/24546pb-sv-d-v-msl-innovative-schlosstechnikmsl-lockcomsfliplockpdf14546pbsvdpdf2016-10-25einpunktverriegelung.jpg)




























