Embed Size (px)
Citation preview

TSINGHUA SCIENCE AND TECHNOLOGY ISSN 100 7-0 214 10 / 18 pp44 0- 446 Volume 11, Number 4, August 2006
Spontaneous Neurotransmitter Release Depends on Intracellular Rather than ER Calcium Stores in Cultured Xenopus NMJ*
GE Song (葛 松), LI Ruxin (李如心), QI Lei (亓 磊), HE Xiangping (何湘平), XIE Zuoping (谢佐平)**
Department of Biological Sciences and Technology, Tsinghua University, Beijing 100084, China
Abstract: Calcium ions are important in many vital neuron processes, including spontaneous neurotransmit-
ter release. Extracellular calcium has long been known to be related to spontaneous neurotransmitter re-
lease, but the detailed mechanism for the effect of intracellular Ca2+ on synaptic release has not yet been
understood. In this research, 1,2-bis-(o-aminophenoxy)-ethane-N, N, N’, N’-tetraacetic acid tetraacetoxy-
methyl ester (BAPTA-AM) was used to combine with cytosolic free Ca2+ in a calcium free medium of cul-
tured Xenopus neuromuscular junctions (NMJ). The spontaneous synaptic current (SSC) frequency was ob-
viously reduced. Then, drugs were applied to interrupt and activate the Ca2+ release channels in the endo-
plasmic reticulum (ER) membrane, but the SSC frequency was not affected. The results show that sponta-
neous neurotransmitter release depends on intracellular rather than ER calcium in cultured Xenopus NMJ
without extracellular calcium.
Key words: spontaneous neurotransmitter; extracellular Ca2+; intracellular Ca2+; endoplasmic reticulum
Introduction
During development, many cell types exhibit sponta-neous neurotransmitter release, with synaptic transmis-sions crucial for normal nervous system activity. Syn-aptic transmissions are initiated when an action poten-tial triggers the neurotransmitter release from a pre-synaptic nerve terminal[1]. An action potential induces the opening of Ca2+ channels, with the subsequent Ca2+ transient stimulating synaptic vesicle exocytosis. At rest, synapses have a finite but low probability of release, causing spontaneous exocytosis events[2]. Im-portant neuronal processes, such as neurotransmitter
release, neuronal excitability and plasticity, neurite outgrowth, synaptogenesis, neuronal survival, and dif-ferentiation, are all Ca2+-dependent processes[3]. There-fore, calcium plays an important role in regulating a great variety of neuronal processes. For a cell, Ca2+ can be extracellular and intracellular. Extracellular Ca2+ has long been known to be related with synaptic spon-taneous release, but the connection between intracellu-lar Ca2+ concentration and synaptic transmission is not well understood. For example, the intracellular Ca2+ concentration is related to the frequency of miniature end plate potentials (MEPPs)[4]. However, the detailed mechanism for the intracellular Ca2+ effect on synaptic release has not yet been understood.
The cytosolic Ca2+ concentration is elevated by channels located on the plasma membrane which transport Ca2+ from the extracellular space, and chan-nels on the endoplasmic reticulum (ER) which regulate the supply of Ca2+ from the organelles themselves[5]. The ER Is a dynamic calcium pool that plays an
﹡
﹡﹡
Received: 2005-09-21; revised: 2005-12-27 Supported by the Natural Science Foundation of Beijing (No. 5052015) To whom correspondence should be addressed. E-mail: [email protected] Tel: 86-10-62788677; Fax: 86-10-62772271

GE Song (葛 松) et al:Spontaneous Neurotransmitter Release Depends … 441
important part in cellular responses to both electrical and chemical signals. The calcium ion channels on the ER are mainly ryanodine receptors (RyR) and inositol 1,4,5-trisphosphate (IP3) receptors (IP3R)[6,7].
CNS synapses are very complex, so the transmission mechanism is difficult to study directly. The neuro-muscular junction (NMJ) provides a simple and easily accessible model to analyze the synaptic transmission mechanisms in great detail, so the Xenopus neuromus-cular junction was used as a model to analyze the in-tracellular Ca2+ induced synaptic spontaneous release mechanism.
1 Materials and Methods 1.1 Culture preparation
Xenopus nerve-muscle cultures were prepared accord-ing to the procedure described previously[8]. In brief, the neural tube and the associated myotomal tissue of Xenopus embryos at stages 20 to 22 were dissociated in Ca2+ and Mg2+ free saline which contains 67 mmol/L NaCl, 1.6 mmol/L KCl, 8 mmol/L Hepes, and 1 mmol/L EDTA, pH 7.8 for 15-20 min. The cells were then grown on glass coverslips for 24 h at room temperature of 20-22. The culture medium consisted of (v/v) 30% Leibovitz L-15 medium (Sigma), 1% FBS (GIBCO), and 69% Ringer’s solution (115 mmol/L NaCl, 2 mmol/L CaCl2, 2.5 mmol/L KCl, and 10 mmol/L Hepes, pH 7.4). Various drugs were applied directly to the culture media at the time of recording.
1.2 Electrophysiological recordings
Synaptic currents were recorded at room temperature in the culture medium from myocytes innervated by spinal motoneurons using whole cell, voltage-clamp recording techniques[8]. The solution inside the re-cording pipette contained 150 mmol/L KCl, 1 mmol/L NaCl, 1 mmol/L MgCl2, and 10 mmol/L Hepes buffer, pH 7.2. For experiments performed in the absence of external Ca2+, the culture medium was replaced with a Ca2+-free extracellular solution containing 115 mmol/L NaCl, 2 mmol/L MgCl2, 10 mmol/L Hepes, 3 mmol/L EGTA, and 0.1% BSA. Membrane currents in all re-cordings were monitored by a patch clamp amplifier (Axon-200B), with a 3-kHz current signal filter. The membrane potentials of the muscle cells were gener-ally in the range of −55-−75 mV and were voltage
clamped at −70 mV after measuring the membrane po-tentials. The resistances of whole-cell electrodes were 2-4 MΩ. All recordings were made at room tempera-ture. Data were collected by a patch clamp amplifier (Axopatch 200B, Axon, USA), filtered at 5 kHz, then stored in a personal computer using a Digidata 1320A interface, and analyzed using pCLAMP 9.2 software (Axon, USA). To quantitatively measure the changes in neurotransmitter release, a time course of the spon-taneous synaptic current (SSC) frequency was first constructed on a minute-to-minute basis. The SSC fre-quencies in a 10-min period right before drug applica-tion were averaged as control. The changes in the SSC frequency were measured by averaging a 5-min or 10-min period recording starting from the first minute after drug application.
2 Results 2.1 SSC depends on intracellular calcium store
A cultured Xenopus myocyte was innervated by a spi-nal motoneuron held in −70 mV (Fig. 1b). The SSCs were recorded in both normal (data not shown) and Ca2+-free extracellular solution (Fig. 1a). The mean frequency of the SSCs in the Ca2+-free extracellular solution was 11 events per min. The SSCs were in-duced by spontaneous secretion of individual ACh-containing synaptic vesicles from motor nerve termi-nals independent of action potentials[9]. 1,2-bis-(o-aminophenoxy)-ethane-N, N, N’, N’-tetraacetic acid tetraacetoxy-methyl ester (BAPTA-AM) application to a final concentration of 50 µmol/L significantly re-duced the SSC frequency (Figs. 1a and 1c). This evi-dence indicates that the intracellular calcium store is necessary and sufficient for spontaneous neurotrans-mitter release in cultured Xenopus NMJ.
2.2 SSC does not depend on the ER calcium store
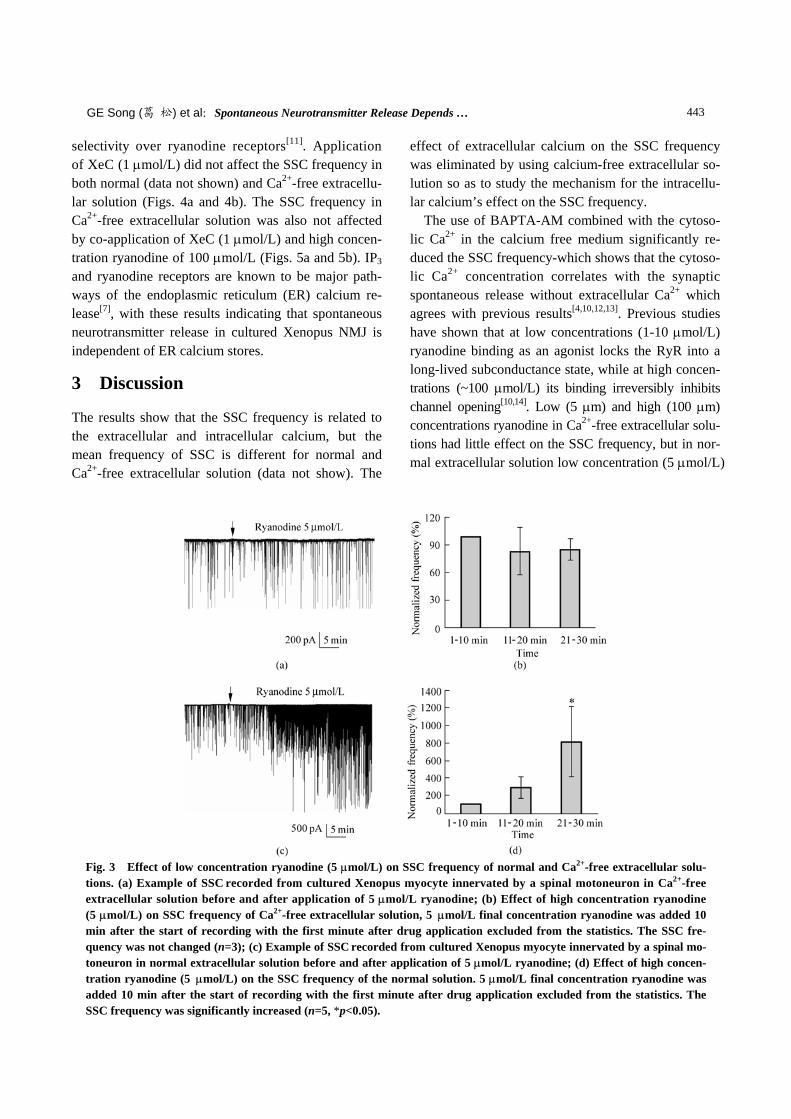
A low concentration of ryanodine (2.5-5 µmol/L) may be used as a ryanodine receptor agonist, while a high concentration (100 µmol/L) serves as an antagonist[10]. In the Ca2+-free extracellular solution, the application of high concentration ryanodine of 100 µmol/L (Figs. 2a and 2b) and low concentration ryanodine of 5 µmol/L (Figs. 3a and 3b) did not affect the SSC fre-quency, but in the normal extracellular solution, the low concentration ryanodine (5 µmol/L) significantly

Tsinghua Science and Technology, August 2006, 11(4): 440-446
442
increased the SSC frequency (Figs. 3c and 3d). The in-crease may be attributed to Ca2+-induced Ca2+ release which is usually associated with the activity of
ryanodine receptors in the presence of extracellular Ca2+. Xestospongin C (XeC) is a potent, membrane permeable IP3 receptor blocker exhibiting a high
Fig. 1 Spontaneous neurotransmitter release depending on intracellular calcium. (a) An example of SSC recorded from cultured Xenopus myocyte innervated by a spinal motoneuron in Ca2+-free extracellular solution before and after BAPTA-AM application of 50 µmol/L; (b) Phase contrast photo of a Xeonpus myocyte innervated by a spinal motoneu-ron clamped by a patch-clamp pipette; (c) Effect of BAPTA-AM on SSC frequency of Ca2+-free extracellular solution. A 50-µmol/L final concentration BAPTA-AM was added 5 min after the start of recording with the first minute after drug application excluded from the statistics. The SSC frequency significantly decreased after the BAPTA-AM application (n=4, *p<0.005).
Fig. 2 Effect of high concentration ryanodine (100 µmol/L) on the SSC frequency of the Ca2+-free extracellular solution. (a) Example of SSC recorded from cultured Xenopus myocyte innervated by a spinal motoneuron in Ca2+-free extracellu-lar solution before and after ryanodine application of 100 µmol/L; (b) Effect of high concentration ryanodine (100 µmol/L) on SSC frequency of Ca2+-free extracellular solution. 100 µmol/L final concentration ryanodine was added 10 min after the start of recording with the first minute after drug application excluded from the statistics. The SSC frequency was not changed (n=6).
GE Song (葛 松) et al:Spontaneous Neurotransmitter Release Depends … 443
selectivity over ryanodine receptors[11]. Application of XeC (1 µmol/L) did not affect the SSC frequency in both normal (data not shown) and Ca2+-free extracellu-lar solution (Figs. 4a and 4b). The SSC frequency in Ca2+-free extracellular solution was also not affected by co-application of XeC (1 µmol/L) and high concen-tration ryanodine of 100 µmol/L (Figs. 5a and 5b). IP3 and ryanodine receptors are known to be major path-ways of the endoplasmic reticulum (ER) calcium re-lease[7], with these results indicating that spontaneous neurotransmitter release in cultured Xenopus NMJ is independent of ER calcium stores.
3 Discussion
The results show that the SSC frequency is related to the extracellular and intracellular calcium, but the mean frequency of SSC is different for normal and Ca2+-free extracellular solution (data not show). The
effect of extracellular calcium on the SSC frequency was eliminated by using calcium-free extracellular so-lution so as to study the mechanism for the intracellu-lar calcium’s effect on the SSC frequency. The use of BAPTA-AM combined with the cytoso-lic Ca2+ in the calcium free medium significantly re-duced the SSC frequency-which shows that the cytoso-lic Ca2+ concentration correlates with the synaptic spontaneous release without extracellular Ca2+ which agrees with previous results[4,10,12,13]. Previous studies have shown that at low concentrations (1-10 µmol/L) ryanodine binding as an agonist locks the RyR into a long-lived subconductance state, while at high concen-trations (~100 µmol/L) its binding irreversibly inhibits channel opening[10,14]. Low (5 µm) and high (100 µm) concentrations ryanodine in Ca2+-free extracellular solu-tions had little effect on the SSC frequency, but in nor-mal extracellular solution low concentration (5 µmol/L)
Fig. 3 Effect of low concentration ryanodine (5 µmol/L) on SSC frequency of normal and Ca2+-free extracellular solu-tions. (a) Example of SSC recorded from cultured Xenopus myocyte innervated by a spinal motoneuron in Ca2+-free extracellular solution before and after application of 5 µmol/L ryanodine; (b) Effect of high concentration ryanodine (5 µmol/L) on SSC frequency of Ca2+-free extracellular solution, 5 µmol/L final concentration ryanodine was added 10 min after the start of recording with the first minute after drug application excluded from the statistics. The SSC fre-quency was not changed (n=3); (c) Example of SSC recorded from cultured Xenopus myocyte innervated by a spinal mo-toneuron in normal extracellular solution before and after application of 5 µmol/L ryanodine; (d) Effect of high concen-tration ryanodine (5 µmol/L) on the SSC frequency of the normal solution. 5 µmol/L final concentration ryanodine was added 10 min after the start of recording with the first minute after drug application excluded from the statistics. The SSC frequency was significantly increased (n=5, *p<0.05).

Tsinghua Science and Technology, August 2006, 11(4): 440-446
444
ryanodine increased the SSC frequency as in Refs. [10,14]. The effect of the IP3 receptor inhibitor XeC on the SSC frequency was indistinguishable from the control. Therefore, these two channels have a
complementary function to some degree. In addition, the use of ryanodine and XeC together resulted in essen-tially the same SSC frequency as the control. Thus, the synaptic spontaneous release rate is independent of the
Fig. 4 Effect of XeC on the SSC frequency of Ca2+-free extracellular solution. (a) Example of SSC recorded from cul-tured Xenopus myocyte innervated by a spinal motoneuron in Ca2+-free extracellular solution before and after applica-tion 1 µmol/L XeC; (b) Effect of XeC on the SSC frequency of Ca2+-free extracellular solution. 1 µmol/L final concentra-tion XeC was added 10 min after the start of recording with the first minute after drug application excluded from the sta-tistics. The SSC frequency was not changed (n=6).
Fig. 5 Effect of co-application of XeC (1 µmol/L) and ryanodine (100 µmol/L) on the SSC frequency of Ca2+-free ex-tracellular solution. (a) Example of SSC recorded from cultured Xenopus myocyte innervated by a spinal motoneuron in Ca2+-free extracellular solution before and after co-application of XeC (1 µmol/L) and ryanodine (100 µmol/L); (b) Effect of co-application of XeC (1 µmol/L) and ryanodine (100 µmol/L) on the SSC frequency of Ca2+-free extracellular solution. 1 µmol/L XeC and 100 µmol/L final concentration ryanodine were both added 10 min after the start of recording with the first minute after drug application excluded from the statistics. The SSC frequency was not changed (n=4).

GE Song (葛 松) et al:Spontaneous Neurotransmitter Release Depends … 445
Ca2+ inside the ER in Ca2+-free extracellular solution. The ER is the largest single intracellular organelle, which is present in all types of nerve cells. The ER is an interconnected, internally continuous system of tu-bules and cisterns, which extends from the nuclear en-velope to the axons and presynaptic terminals, as well as to the dendrites and dendritic spines. The ER fulfills important functions such as the folding and processing of proteins, all of which are strictly calcium dependent. The ER calcium activity is, therefore, relatively high. Calcium stored within the endoplasmic reticulum of the neuron represents an important source of signal Ca2+ that is released upon activation of either the IP3Rs or the RyRs[15,16]. In a recent review, Verkhratsky[17] stated that the Ca2+ release channels and the Ca2+ pumps in the ER membrane provide for synaptic excit-ability. However, this study showed that calcium released from these two receptors, namely, the ER calcium, was not necessary for synaptic spontaneous release in Ca2+-free extracellular solution.
The SSC frequency may also be maintained by other intracellular Ca2+ sources, such as cytosolic dissociated calcium or calcium inside the mitochondria. However, at rest the cytosolic dissociated calcium concentration is very low, only about 100 nmol/L[18], which is not sufficient to maintain synaptic spontaneous release for 20 min or more. Therefore, the mitochondria calcium most likely maintains the synaptic spontaneous release. Mitochondria can be considered as a Ca2+ store under some circumstance[19]. These organelles sequester Ca2+ via a low-affinity, high capacity rapid uniporter[20]. Mi-tochondria release their accumulated Ca2+ largely by a Na+-dependent exchange, which runs at a significantly slower rate than the uniporter[21,22]. Besides the Na+-dependent exchange, a large pore in the mitochondrial inner membrane, which is called the mitochondrial permeability transition pore, is now known to release calcium from mitochondria stores[23]. As a conse-quence, mitochondria can prolong cytosolic Ca2+ sig-nals as they slowly lose their sequestered Ca2+. In some cell types mitochondria appears to have sufficient Ca2+ at rest for them to participate in intracellular Ca2+ sig-nals. Indeed, propagating Ca2+ waves have been ob-served following regenerative activation of the perme-ability transition[24]. Since in the majority of cell types, mitochondrial Ca2+ levels are low at rest, it seems unlikely that the mitochondrial Ca2+-induced Ca2+
release mechanism would be active until substantial Ca2+ accumulation had occurred. However, when the Ca2+ inside the ER is sequestered by drugs, the mito-chondrial Ca2+ may complementarily release. There-fore, the SSC frequency may not notably change when using the ryanodine and XeC in a calcium-magnesium free medium. Further studies will investigate whether Ca2+ from the mitochondria stores would be sufficient and is necessary for the synaptic spontaneous release or whether there are still some other mechanisms in-side the cell.
References
[1] Kato N. The Release of Neural Transmitter Substances. Liverpool: Liverpool University Press, 1969.
[2] Sudhof T C. The synaptic vesicle cycle. Annu. Rev. Neuro-sci., 2004, 27: 509-547.
[3] Pietrobon D. Function and dysfunction of synaptic calcium channels: Insights from mouse models. Curr. Opin. Neuro-biol., 2005, 15(3): 257-265.
[4] Angleson J K, Betz W J. Intraterminal Ca2+ and spon-taneous transmitter release at the frog neuromuscular junc-tion. J. Neurophysiol., 2001, 85(1): 287-294.
[5] Berridge M J, Lipp P, Bootman M D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell. Biol., 2000, 1(1): 11-21.
[6] Collin T, Marty A, Llano I. Presynaptic calcium stores and synaptic transmission. Curr. Opin. Neurobiol., 2005, 15(3): 275-281.
[7] Simpson P B, Challiss R A, Nahorski S R. Neuronal Ca2+ stores: Activation and function. Trends Neurosci., 1995, 18(7): 299-306.
[8] Lu B, Greengard P, Poo M M. Exogenous synapsin I pro-motes functional maturation of developing neuromus-cular synapses. Neuron, 1992, 8(3): 521-529.
[9] Song H, Ming G, Fon E, Bellocchio E, Edwards R H, Poo M. Expression of a putative vesicular acetylcholine trans-porter facilitates quantal transmitter packaging. Neuron, 1997, 18(5): 815-826.
[10] He X, Yang F, Xie Z, Lu B. Intracellular Ca2+ and Ca2+/calmodulin-dependent kinase II mediate acute po-tentiation of neurotransmitter release by neurotrophin-3. J. Cell. Biol., 2000, 149(4): 783-792.
[11] Gafni J, Munsch J A, Lam T H, Catlin M C, Costa L G, Molinski T F, Pessah I N. Xestospongins: Potent mem-brane permeable blockers of the inositol 1,4,5-trisphosphate receptor. Neuron, 1997, 19(3): 723-733.

Tsinghua Science and Technology, August 2006, 11(4): 440-446
446
[12] Tse F W, Tse A, Hille B, Horstmann H, Almers W. Local Ca2+ release from internal stores controls exocytosis in pi-tuitary gonadotrophs. Neuron, 1997, 18(1): 121-132.
[13] Blochl A, Thoenen H. Localization of cellular storage compartments and sites of constitutive and activity-dependent release of nerve growth factor (NGF) in pri-mary cultures of hippocampal neurons. Mol. Cell. Neuro-sci., 1996, 7(3): 173-190.
[14] Serysheva II H S. Ryanodine binding sites on the sar-coplsmic reticulum Ca2+ release channel, in structure and function of ryanodine receptor. London: Imperial College Press, 1998: 95-104.
[15] Henzi V, MacDermott A B. Characteristics and function of Ca2+- and inositol 1,4,5-trisphosphate-releasable stores of Ca2+ in neurons. Neuroscience, 1992, 46(2): 251-273
[16] Kostyuk P, Verkhratsky A. Calcium stores in neurons and glia. Neuroscience, 1994, 63(2): 381-404.
[17] Verkhratsky A. Physiology and pathophysiology of the calcium store in the endoplasmic reticulum of neurons. Physiol. Rev., 2005, 85(1): 201-279.
[18] Plieth C. Calcium: Just another regulator in the machinery
of life? Ann. Bot. (Lond), 2005, 96(1): 1-8. [19] Bootman M D, Collins T J, Peppiatt C M, Prothero L S,
MacKenzie L, De Smet P, Travers M, Tovey S C, Seo J T, Berridge M J, Ciccolini F, Lipp P. Calcium signalling—An overview. Semin. Cell. Dev. Biol., 2001, 12(1): 3-10.
[20] Nguyen M H, Jafri M S. Mitochondrial calcium signaling and energy metabolism. Ann. N. Y. Acad. Sci., 2005, 1047: 127-137.
[21] Reed P W, Lardy H A. A23187: A divalent cation iono-phore. J. Biol. Chem., 1972, 247(21): 6970-6977.
[22] Carafoli E, Tiozzo R, Lugli G, Crovetti F, Kratzing C. The release of calcium from heart mitochondria by sodium. J. Mol. Cell Cardiol., 1974, 6(4): 361-371.
[23] Bernardi P. Mitochondrial transport of cations: Channels, exchangers, and permeability transition. Physiol. Rev., 1999, 79(4): 1127-1155.
[24] Ichas F, Mazat J P. From calcium signaling to cell death: Two conformations for the mitochondrial permeability transition pore—Switching from low- to high-conductance state. Biochim. Biophys. Acta., 1998, 1366(1-2): 33-50.
Sino-UK Symposium on Developmental Biology and Human Diseases Held in Tsinghua
The Sino-UK Symposium on Developmental Biology and Human Diseases opened in Tsinghua May 6, 2006. The Symposium, which ran through May 8, 2006, was hosted by the Department of Biological Sciences and Biotech-nology, Tsinghua University.
Members of the Royal Society in UK, including Professor Philip W. Ingham from the University of Sheffield, Professor James C. Smith and Professor Michael E.Akam from Cambridge University, Professor David Ish-Horowicz from Cancer Research UK, and Professor Peter W. H. Holalnd from Oxford University attended the symposium.
Also attending the symposium were Professor Meng Anming from Tsinghua University, Professor Zhang Yongqing from Chinese Academy of Sciences, and Professor Zhang Bo from Peking University.
Experts from both China and UK discussed and gave reports on developmental biology and human diseases at a meeting held on May 8, 2006. About 150 researchers and students from universities and research institutes in Bei-jing attended the event that day.
(From http://news.tsinghua.edu.cn)















![Nmj Nov08[2]](https://img.pdfslide.net/doc/110x75/557a8b8fd8b42ac8638b4d86/nmj-nov082.jpg)










![OTOT-NMJ [Compatibility Mode]](https://img.pdfslide.net/doc/110x75/55cf97e4550346d033944380/otot-nmj-compatibility-mode.jpg)


