Embed Size (px)
Citation preview

8/3/2019 S.R. El-Defrawy et al- Functional and Neurochemical Cortical Cholinergic Impairment Following Neurotoxic Lesions of the Nucleus Basalis Magnocellularis in the …
http://slidepdf.com/reader/full/sr-el-defrawy-et-al-functional-and-neurochemical-cortical-cholinergic-impairment 1/6
Neurobiology of Aging, Vol. 6, pp. 325-330, 1985. © Ankho International nc. Printed in the U.S.A. 0197-4580/85 $3.00 + .00
Fu n c t io n a l an d Neu r o ch em ica l Co r t i c a lCho l ine rg ic Impa i rmen t Fo l lowing
N e u r o t o x i c L e s io n s o f th e N u c l e u sBasa l i s Magnoce l lu la r i s in the Rat
S. R . EL-D EFRA WY,* F. COLOM A,* K. JHAMA NDAS, .1 R . J . BOEGM AN,*R. J . BENI NGERt AND B. A. WlRSC HINGt
Department of Pharmacology and Toxicology* and Department of PsychologytQueen's University, Kingston, Ontario, Canada, K7L 3N6
Recei ved 10 Jan uar y 1984
EL-DEFRAWY, S. R., F. COLOMA, K. JHAMANDAS, R. J. BOEGMAN, R. J. BENINGER AND B. A. WIRSCHING.Functiomd and neurochemical cortical cholinergic impairment idhm'ing neurotoxic lesums of the nucleus basalis magnocel-luhu'is in the rat. N EUROBIOL AGING 6(4)32~330, 1985.--The effect of kainic and quinolinic acid on cortical cholinergicfunction was examined following injections of these agents into the nucleus basalis magnocellularis (nbm) or into the fronto-parietal cortex. The release of cortical 3H-acetylcholine aH-ACh), high affinity choline uptake (HACU) and acetylcholinesterasewas measured 7 days following injections of saline (control), kainic acid (4.7 nmoles) and quinolinic acid (60 , 150 and 300nmoles) into the nbm. These cortical cholinergic parameters were also examined after injections of saline (control), kainicacid (9.4 nmoles) and quinolinic acid t300 nmoles) into the fronto-parietal cortex. The release of3H-ACh, HACU and AChEwas significantly reduced in animals injected with kainic or quinolinic acid into the nbm. Histological examinat ion ofstained sections showed a loss of cell bodies in the region of the nbm and the giobus pallidus. The size of the lesionproduced b y quinolinic acid was proportional to the dose injected into the nbm. In animals injected with kainic acid or
Iquino linic acid into the cerebral cortex, the release o f :H-ACh, HACU and AChE was not significant!y reduced whencompared with cont rol animals, although histological examination of stained cortical sections showed a marked loss ofcortical neurons . The results show that quinolinic acid, an endogenous neuroexcitant, produces a deficit of cholinergicfunction similar to that described in the cortical tissue of patients with senile dementia of Alzheimer' s type. The toxiceffects of quinolinic acid on cortical cholinergic function are due to its action on cholinergic cell bodies in the nbm. Thecortical slice preparation from quinolinic acid-treated animals showing impairment of "~H-ACh release, may be useful inassessing the action of drugs designed to improve cholinergic function.
Quino linic acid Kainic acid Nucleus basalis magnocellularisAcetylcholine release Alzheimer's disease
Cholinergic markers
NEUROCHEMICAL studies on autopsy brain of patientssuffering from senile dementia of the Alzheim er type (SDAT)have demonstrated a selective loss of cholinergic markerssuch as choline acetyltransferase (CAT) and acetylcholines-terase, enzymes responsible for the synthesis and degrada-tion of the transmitter acetylcholine (ACh), in hippocampaland cortical areas [2]. The decline in these cholinergic mark-ers is believed to be a major contribu ting factor in the cogni-tive dysfunction seen in patients with SDAT. Recent studieshave demonstrated that the cortical cholinergic loss may bedue to the degeneration of a specific cholinergic pathwaythat originates from the nucl eus basalis of Meynert and pro-jects to the cerebral cortex [2]. The biochemical pathologycharacteristic of SDAT can be mimicked in animals such asthe rat by injection of the excitotoxin, kainic acid, into cer-tain subcortical nuclei. Stereotaxic injections of this.
neurotoxin into the nucleus basalis magnocellularis (nbm),located in the ventral region of the globus pallidus, results ina decrease in CAT, AChE and high affinity choline uptake(HACU) in the frontal and parietal cortex [6,8]. Thissuggests that the nb m cholinergic neurons project to corticalareas. The failure of kainic acid to completel y abolish corti-cal cholinergic markers indicates that some corticalcholinergic neurons may be intrinsic [3, 6, 7] or that corticalcholinergic projections may also arise in different subcorticalregio ns [21].
Because micro injections of the exogenous excitotoxinsuch as kainic acid into the nbm can produce some of theneurochemi cal changes seen in SDAT [1], it is possible thatthe cholinergic damage occurring in this disorder may arisefrom an endogenous neurotoxin generated in the brain. Re-cently, it was reported that quinolini c acid, a mino r metabo-
tRequests for repr ints should be addressed to Dr. K. Jhamandas, Department of Pharmacology and Toxicology, Queen' s Universi ty,Kingston, Ontario, Canada. K7L 3N6.
325

8/3/2019 S.R. El-Defrawy et al- Functional and Neurochemical Cortical Cholinergic Impairment Following Neurotoxic Lesions of the Nucleus Basalis Magnocellularis in the …
http://slidepdf.com/reader/full/sr-el-defrawy-et-al-functional-and-neurochemical-cortical-cholinergic-impairment 2/6
326 E L - D E F R AW ~ £ ~ A i
a - - U l I . E g l O I4 E D l i D [
i " + = , : . + = .+
] + "
• .+
I " I '
• , , - - . . . . . , - - p " . , . ,
.0 . , , . % . , % . o . 0 =
"I'ii~olloFIG . 1 . The outf low of radioac t iv i ty from superfused frontopa rie ta lcort ica l s l ices of animals tha t had rec e ived a uni la tera l 1 / .d infus ionof: (A) 0.9% saline; (B) 4.7 nmoles kainic acid; (C) 60 nmolesquinolinic acid; (D) 150 nmoles quinolinic acid; (E) 300 nmoles
quinol inic ac id in to the nbm. After pre incubat ion w ith 3H-chol ine ,s l ices were superfused with K rebs-He nsele i t and s t im ula ted bya
5-ra in exposure to medium conta ining 35 mM K -. Points representthe mean outf low-+SEM (n=4). Ordina te : outf low of radioac t iv i typer t ime period as pe rcent o f to ta l s l ice content . Sol id and dashedl ines represent re lease from the unles ioned and les ioned hemi-spheres respect ive ly.
l i t e o f t he e s s e n t i a l a m i n o a c i d t r y p t o p h a n , p r o d u c e s a x o n -s p a r i n g l e s i o n s o f h i p p o c a m p a l a n d s t r i a ta l n e u r o n s s i m i l a r t ot h o s e p r o d u c e d b y k a l n i c a c i d [ 16 ]. T h u s , i t i s o f in t e r e s t t od e t e r m i n e w h e t h e r q u i n o l i n i c a c i d p r o d u c e s a f u n c t i o n a ld a m a g e o f c h o l i n e r g i c n e u r o n s p r o j e c t i n g to t h e c o r t e x .
T h e r e l e a s e o f A C h f r o m t h e c e r e b r a l c o r t e x c a n b e u s e da s a n i n d i c a t o r o f t h e f u n c t i o n a l s t a t e o f c h o l i n e rg i c n e u r o n s .I n d e e d , t h e s y n t h e s i s a n d r e l e a s e o f A C h i n r e s p o n s e t o K *d e p o l a r i z a t i o n i s r e d u c e d f r o m c o r t i c a l t i s s u e o b t a i n e d f r o mp a t i e n t s w i t h S D AT [ 1 8 , 1 9 ] . I n t h e p r e s e n t s t u d y w e t e s t e dw h e t h e r f o c a l a d m i n i s t r a t i o n o f q u i n o l in i c a c i d i n t o t h e b r a i no f e x p e r i m e n t a l a n i m a l s p r o d u c e s a d e f i c i t i n th e s p o n t a n e -o u s a n d t h e K ÷ - e v o k e d r e l e a s e o f A C h . A C h E a c t i v i t y a n dH A C U w a s c o m p a r e d t o t h a t o f k a i n i c a c i d . A p r e li m i n a r ya c c o u n t o f t h i s w o r k h a s a l r e a d y b e e n r e p o r t e d ( E I - D e f r a w ye t a l . , So (" Ne uros c i Abs t r10 :1187 , 1984) .
METHOD
S t e r e o t a x i c I n j e c t i o n s
M a l e S p r a g u e - D a w l e y r a t s ( 2 7 5 -3 5 0 g ) w e r e a n e s t h e t i z e dw i t h h a l o t h a n e ( H a l o c a r b o n , M a l t o n , O n t a r i o ; 2% h a l o t h a n e ,9 8 % o x y g e n v i a in h a l a t i o n ) a n d p o s i t i o n e d i n a N a r a s h i g h e
s m a l l a n i m a l s t e r e o t a x i c a p p a r a t u s . F o r i n j e c t i o n s i n t o t h en b m , t h e f o l lo w i n g c o o r d i n a t e s w e r e u s e d : 0 . 8 m m p o s t e r i o rt o b r e g m a ; 2 . 6 m m l a t e r a l a n d 8 . 0 m m v e n t r a l t o t h e s u r f a c eo f t h e s k u l l w i t h t h e i n c i s o r b a r s e t a t - 3 . 3 m m [ 13 ]. C o r t i c a ll e s i o n s w e r e m a d e b y i n j e c t i n g n e u r o t o x i n a t t w o s i t e s a l o n gt h e f r o n t o - p a r i e t a l c o r t e x ( ! . 7 m m a n t e r i o r t o b r e g m a , 2 . 0m m l a t e r a l , 2 . 6 m m v e n t r a l ; a n d 1 .8 m m p o s t e r i o r to b r e g m a ,3 . 0 m m l a t e r a l , 2 .3 m m v e n t r a l ).
K a i n i c a c i d ( L o t 3 2 F - 0 8 6 7, S i g m a ) o r q u i n o l i n ic a c i d ( L o t7 2 F - 0 2 8 3 , S i g m a ) w a s d i s s o l v e d i n 0 . 9 % s a l i n e a n d t i t r a t e d t op H 7 . 4 w i t h 1 N N a O H . K a i n i c a c i d ( l p .l o f 4 .7 n m o l e s / ~ . l ) o rq u i n o l i n i c a c i d ( l ~ 1 o f 6 0 , 1 5 0 o r 3 0 0 n m o l e s / ~ l ) w a s i n f u s e d
N B M L E S I O N C O ~ i C ~ L L E S I O h
[ ] U N I N J f C T I O ( l e | ! l t l e )
[ ] , . , , c , = o ,. . . . . . . . . , T T
..
• I o I 1 0 I 0 O $ o t +n m + + l =
I:
+ c e
e r,, ,i,
i l A L I N [ K & I N I C O U I N O t J t N Q U I N S A L I N E II & I N t C O U I N4 ? II 0 11 0 : lO 0 ~ * 4 ~ I o o n m o l l l
tFIG. 2 . High aff in i ty chol ine uptake (HACU) and ace tylchol ines-te rase a c t iv i ty (ACHE) in the c erebra l corte x fol lowing in ject ions ofsa l ine , ka inic ac id and quinol inic ac id in to the nb m an d the fronto-parie ta l cortex. In the la t te r case an in jec t ion (1 /~l) of ka ina te (4 .7
nmoles) or quinol inic ac id 050 nmoles) was made in to the r ightfronta l cortex and in to the parie ta l cortex, n=3-5 animals . *Indi-ca tes the va lue is s ignif icant ly different f rom the uninjec ted controls ide (p<0.05) .
a t t h e s p e c i f i c s i t e o v e r a p e r i o d o f 2 r a i n , 25 s e c v i a a H a m i l -t o n c a n n u l a ( 0 . 3 5 m m o . d . ) . A f t e r t h e i n f u s i o n , a p e r i o d o f 3m i n w a s a l l o w e d f o r d i f fu s i o n . T h e c a n n u l a w a s w i t h d r a w na n d t h e s c a l p w a s a p p o s e d w i t h s u t u r e s . S h a m - l e s i o n e d r a tsr e c e i v e d a n i n f u s i o n o f 1 /z l o f 0 .9 % s a l i n e . To m i n i m i z e o rp r e v e n t d a m a g e t o r e m o t e a r e a s 2 0 m g / k g p e n t o b a r b i t a l w a sa d m i n i s t e r e d t o a n i m a l s i m m e d i a t H y a f t e r r e c o v e r y f r o ma n e s t h e s i a . T h e a n i m a l s w e r e k i l le d 7 d a y s p o s t - l e s i o n i n g f o rA C h r e l e as e e x p e r i m e n t s , a s s e s s m e n t o f A C h E a c t iv i t y a n dH A C U , a n d f o r h i s to l o g y.
R e l e a s e E x p e r i m e n t s
T h e r e l e a s e o f ~ H - a c e t y l c h o l in e ( 3 H -A C h ) f r o m s l i c es o ff r o n t o p a r i e t a l c o r t e x w a s i n v e s t i g a t e d u si n g a m o d i f i c a t i o no f th e m e t h o d o l o g y d e s c r i b e d b y S o m o g y i a n d S z e r b [ 20 ]a n d L e h m a n n a n d S c a t t o n [ 9] . T h e r a t s w e r e d e c a p i t a t e d , th eb r a i n r a p i d l y r e m o v e d a n d h e m i s e c t e d . A s l a b o f f r o n t o -p a r i e t a l c o r t e x w a s d i s s e c t e d f r o m e a c h h e m i s p h e r e a n ds l i c e d (0 .3 m m t h i c k ) c o r o n a l l y w i t h a M c I l i w a i n t i s s u ec h o p p e r . D i s p e r s e d b r a i n s l i c e s w e r e p r e i n c u b a t e d f o r 5 m i ni n K r e b s - H e n s e l e i t b u f f e r ( K R B ) a t 3 7 °C i n a D u b o n o f fm e t a b o l i c s h a k i n g i n c u b a t o r u n d e r a n a t m o s p h e r e o f 9 5 %
0 2 - 5 % C O : . U n l e s s o t h e r w i s e s p e c i f i e d , t h e K R B w a s o f t h ef o l l o w i n g c o m p o s i t i o n ( m M ) : N a C l , 118 ; K C I , 4 . 8 ; K H .~ PO 4 ,1 .2 ; C aC I ~ , 1 .3 ; N a H C O 3 , 2 5 ; g l u c o s e , 1 0 a n d p h y s o s t i g m i n e ,0 .0 1 . T h e p r e i n c u b a t i o n m e d i u m w a s e x c h a n g e d f o r m e d i u mc o n t a i n i n g 5 x l 0 - ~ M m e t h y l : a l l - c h o l i n e c h l o r i d e ( s p e c if i ca c t i v i t y 80 C i / m m o l e ) a n d t h e i n c u b a t i o n c o n t i n u e d f o r af u r t h e r 1 5 m i n . T h e s l i c e s w e r e r i n s e d t w i c e a n d f o u r s l i c e s( t o t a l w e i g h t 8 .5 - * 0 .5 m g ) w e r e t r a n s f e r r e d i n t o a s u p e r f u s i o nc h a m b e r .
B r a i n s l i ce s w e r e s u p e r f u s e d f r o m t h e t o p o f th e c h a m b e ra t a r a t e o f 0 .7 5 m l / m i n . A f t e r a 3 0 - r a i n p e r i o d ( t = 3 0 m i n ) ,s a m p l e s o f s u p e r f u s a t e w e r e c o l l e c t e d a t 2 . 0 - m i n i n t e rv a l s

8/3/2019 S.R. El-Defrawy et al- Functional and Neurochemical Cortical Cholinergic Impairment Following Neurotoxic Lesions of the Nucleus Basalis Magnocellularis in the …
http://slidepdf.com/reader/full/sr-el-defrawy-et-al-functional-and-neurochemical-cortical-cholinergic-impairment 3/6
C H O L I N E R G I C F U N C T I O N A N D N B M L E S I O N S 327
A
Il I~(~IA -.8 m m
ilI~GMA -1. 0 m m
[II:EGMA 1.2mm
FIG. 3 . Schematic representat ion of the ex ten t o f damage producedby infusions of: (A) kainic acid, 4.7 nmoles and (B) quinolinic acid 150nm oles (right side) into the nbm , as assessed by cresyl violet histol-ogy. Five coronal sections are depicted with numbers below refer-ring to distance (ram) posterior to bregma.
f o r th e d u r a t i o n o f t h e e x p e r i m e n t . A t 4 0 m i n ( t = 4 0 r a in ) , t h es li ce s w e r e e x p o s e d t o K R B m e d i u m c o n t a in i n g 3 5 m M K ÷f o r 5 r a in ( w h e n e l e v a t e d K ÷ m e d i u m w a s u s e d , i s o s m o l a r i t yw a s m a i n t a i n e d b y a n e q u i m o l a r r e d u c t i o n i n N a ~ ) . A t t h ee n d o f t h i s p e r i o d t h e s l i c e s w e r e s u p e r f u s e d w i t h n o r m a lm e d i u m f o r a n o t h e r 2 0 m in . T h e t i s s u es f r o m e a c h c h a m b e rw e r e s o l u b i l i z e d a n d t h e r a d i o a c t i v i t y c o u n t e d . Tw o s a m p l e sf r o m e m p t y s u p e r f u s e d c h a m b e r s w e r e c o l le c t ed f o r r es i d ua lr a d i o a c t i v i t y. R a d i o a c t i v i t y i n a l l s a m p l e s w a s c o u n t e d i n al iq u i d s c i n ti l la t i o n c o u n t e r u s i n g t h e e x t e r n a l s t a n d a r d sc h a n n e l r a ti o m e t h o d a n d q u e n c h c o r r e c ti o n . E f f ic i e n c y o fco u n t in g w as 3 0 - -3 5 %.
T h e o u t f l o w o f r a d i o a c t i v i t y f r o m t h e t is s u e w a s c a l c u -l a t e d f r o m t h e r a d i o a c t i v i t y r e l e a s e d d u r i n g e a c h s a m p l ep e r i o d a n d e x p r e s s e d a s a p e r c e n t a g e o f to t a l s li c e c o n t e n t a tt h e t i m e o f co l l e c t i o n o f t h a t s a m p l e .
B i o c h e m i c a l A n a l y s e s
M e a s u r e m e n t s o f H A C U a n d A C h E in e ac h a n im a l w e rem a d e o n t h e u n i n j e c t e d s i d e a n d i n j e c t e d s i d e . S i n c e e a c ha n i m a l s e r v e d a s i t s o w n c o n t r o l t h e i n f l u e n c e o f v a r i a b il i tyb e t w e e n d i f f e re n t g r o u p s o f a n i m a l s w a s m i n i m i z e d .
T h e r a t e o f H A C U w a s m e a s u r e d a c c o r d i n g t o m o d i fi c a-t i o n s o f t h e m e t h o d s o f S i m o net a l . [ 1 7] a n d H a r t g r a v e se ta l. [ 5 ] . S y n a p t o s o m e s w e r e p r e p a r e d b y c e n t r i f u g i n g c o r t i c a lh o m o g e n a t e s a t 1 0 0 0 × g a n d th e s u p e r n a t a n t a t 1 7 , 0 0 0 × g .3 H - C h o l i n e ( 1 0 0 n M ) w a s a d d e d t o t h e r e s u s p e n d e d p e l l e t o fs y n a p t o s o m e s a n d u p t a k e w a s a l l o w e d t o p r o c e e d f o r 4 ra ina t 3 70 C . B l a n k s w e r e r u n i n t h e s a m e f a s h i o n e x c e p t t h ei n c u b a t io n m e d i u m c o n t a i n e d n o N a ~ . T h e r e a c ti o n w a ss t o p p e d b y f i lt r at i o n t h r o u g h n i t r o c e l l u lo s e - c e l l u l o s e - a c et a t ef ' d ter s ( p o re s ize 0 .4 5 ~ .m) an d th e r ad io ac t iv i ty r e ta in ed o n
. . . . . U E U ~ I ~ I V 4 ~
A
@0 '
Q@,
It O
It O
I O [ "
N u l l K *
' l / l iE (m ~ )
C
FIG . 4. Th e outflow o f radioactivity from superfused frontoparietalcortical slices of animals th at had rec eived a I t~l infusion o f saline;kainic acid, 4.7 nm oles or quinolinic acid , 150 nm oles at each of twopoin ts along the frontoparietal corte x. (A) 0.9% saline; (B) kainicacid; (C) quirlolinic acid. A fter preincubation with 3H -choline, sliceswe re superfused w ith Krebs-Henseleit and stimulated b y a 5-rainexpo sure to medium containing 35 mM K +. Points represent meanou tflow ±S EM (n--4). Ordinate: outflow of radioactivity p er timeperiod as percent o f total slice content. Solid and dashed lines repre-
sent release from unlesioned a nd lesioned hem ispheres respectively.
t h e f 'd t e r c o u n t e d . A C h E a c t i v i t y w a s d e t e r m i n e d b y t h em e t h o d o f E ll m a ne t a l . [ 4 ] w h i l e p r o t e i n w a s m e a s u r e d a c -c o r d i n g t o L o w r yet a l . [12] .
H i s t o l o g y
A f te r 1 0 - -2 0 d ay s o f f ix a t io n in 1 0 % fo rmal in , th e b r a in sw e r e f r o z e n o n a c r y o s t a t c h u c k a n d s e c t i o n e d ( 4 0 t zm t h i ck -n ess ) to r ev ea l th e le s io n s i t e . Tis su e s ec t io n s w ere s ta in edf o r N i s s l s u b s t a n c e w i t h c r e s y l v i o l e t a n d s e r i al s e c t i o n sw e r e u s e d t o m a p t h e s i z e o f th e l e s io n .
RESULTS
n b m I n j e ct io n s
3 H - A c e t y l c h o l i n e r e l e a s e .T h e r e l e a s e o f 3 H - A C h f r o mc o r t i ca l s l ic e s p r e p a r e d f r o m a n i m a l s w i t h s h a m i n j e c t i o n si n t o t h e n b m i s s h o w n i n F i g . I A . N o s i g n i f i ca n t d i f fe r e n c e i ns p o n t a n e o u s o r K + - e v o k e d r e l e as e o f r a d i o a ct i v it y w a s o b -s e r v e d b e t w e e n i n j e c t e d a n d u n i n j e c t e d s i d e s .
F i g u r e I B s h o w s t h e r e le a s e o f 3 H - A C h f ro m t h e c e r e b r a lc o r t e x o f a n i m a l s i n j e c t e d w i t h k a i n i c a ci d i n t o t h e n b m . T h es p o n t a n e o u s r e l e a s e f r o m t h e c o r t e x i p s i l at e ra l t o t h e i n j e c -t io n ( ip s i la te r a l co r tex ) d id n o t d i f f e r s ig n i f ican t ly f ro m th er e l e a se o c c u r r i n g f r o m t h e c o r t e x c o n t r a l a te r a l t o t h e i n -j e c t e d s i d e ( c o n t r a l a t e r a l c o r t e x ) . T h e K ÷ - e v o k e d r e l e a s e o f3 H - A C h f r o m t h e i p si l at e ra l c o r t e x w a s r e d u c e d b y a b o u t7 0 % w h e n c o m p a r e d t o t h e c o n t r a l a t e r a l c o r t e x . N o d i ff e r-e n c e w a s a p p a r e n t i n t h e s p o n t a n e o u s r e l e a se f r o m t h e t w oc o r t i c e s d u r i n g th e p o s t - s t i m u l u s r e c o v e r y p h a s e . T h e r e s u l t so f r e l e a s e e x p e r i m e n t s p e r f o r m e d o n q u i n o l i n ic a c i d - l e s io n e da n i m a ls a re s h o w n i n F ig . I C E . T h e s p o n t a n e o u s a n d K ~ -e v o k e d r e l e a s e o c c u r r i n g f r o m t h e i p s i l a te r a l a n d c o n t r a l a t -e r a l c o r t e x i n a n i m a l s i n j e c t e d w i t h 6 0 n m o l e s o f q u in o l i n i ca c i d d i d n o t d i f f er s i g n i fi c a n tl y. H o w e v e r , f o l l o w i n g i n j e c t io no f a h i g h e r d o s e o f q u in o l i n i c a c i d ( 1 5 0 o r 3 0 0 n m o l e s ) , t h eK + - e v o k e d r e l e a s e f r o m t h e i p s il a te r a l c o r t e x w a s s ig n if i-c a n t l y r ed u c e d w h e n c o m p a r e d t o th e c o n t r a la t e ra l c o r t e x .T h e s p o n t a n e o u s r e l e a s e o c c u r r i n g b e f o r e o r a f t e r K ÷ a p p l i-c a t i o n w a s n o t s i g n i f i c a n t l y d i f f e r e n t . A l t h o u g h t h e t w o
8/3/2019 S.R. El-Defrawy et al- Functional and Neurochemical Cortical Cholinergic Impairment Following Neurotoxic Lesions of the Nucleus Basalis Magnocellularis in the …
http://slidepdf.com/reader/full/sr-el-defrawy-et-al-functional-and-neurochemical-cortical-cholinergic-impairment 4/6
328 EL-DEFRAWY L7 A~..
A B
FIG. 5. Schematic representation of the extent of damage producedby infusions at two sites along the frontoparietal cortex (fight side)of: (A) kainic acid, 4.7 nmoles/site; (B) quinolinic acid. 150 nmoles/site as assessed by cresyl violet histology. Five coronal sectionsare depicted with numbers referring to the distance (ram) anterior(+) or posterior (-) to bregma.
doses of quinolinic acid that reduced K÷-evoked release dif-fered by 100%, their effect on the evoked release was com-parable.
A Ch E a c t i v i t y a n d H A CU .The effects of kainate andquinolinic acid injections were also investigated on AChEactivity and HACU, two markers of cholinergic function.The tissue samples used in this pan of the study were ob-tained from the tissue block prepared for the 3H-ACh releaseexperiments described above. The results are shown in Fig.2A and C. In saline-injected animals there was no differencein these cholinergic markers whe n the ipsilateral and con-tralateral cortices were compared. Following injection ofkainic acid, the HACU and AChE activity in the ipsilateralcortex decreased by 47% and 70% respectively. Quinolinicacid injections (150 and 300 mmoles) produced a similar ef-fect on these markers. Injection of 60 nmoles, the lowestdose empl oyed, did not produce a significant decre ase inHACU or ACHE. However, following 150 nmoles, theHACU and AChE activity decreased by 64% and 58% re-spectively. A higher dose of quinolinic acid, 300 nmoles,produced comparable effects on these two biochemicalmarkers. Thus, as was observed in the preceding releaseexperiments, these two doses of quinolinic acid produ~:edeffects of comparable magnitude on HACU and AChE ac-tivity.
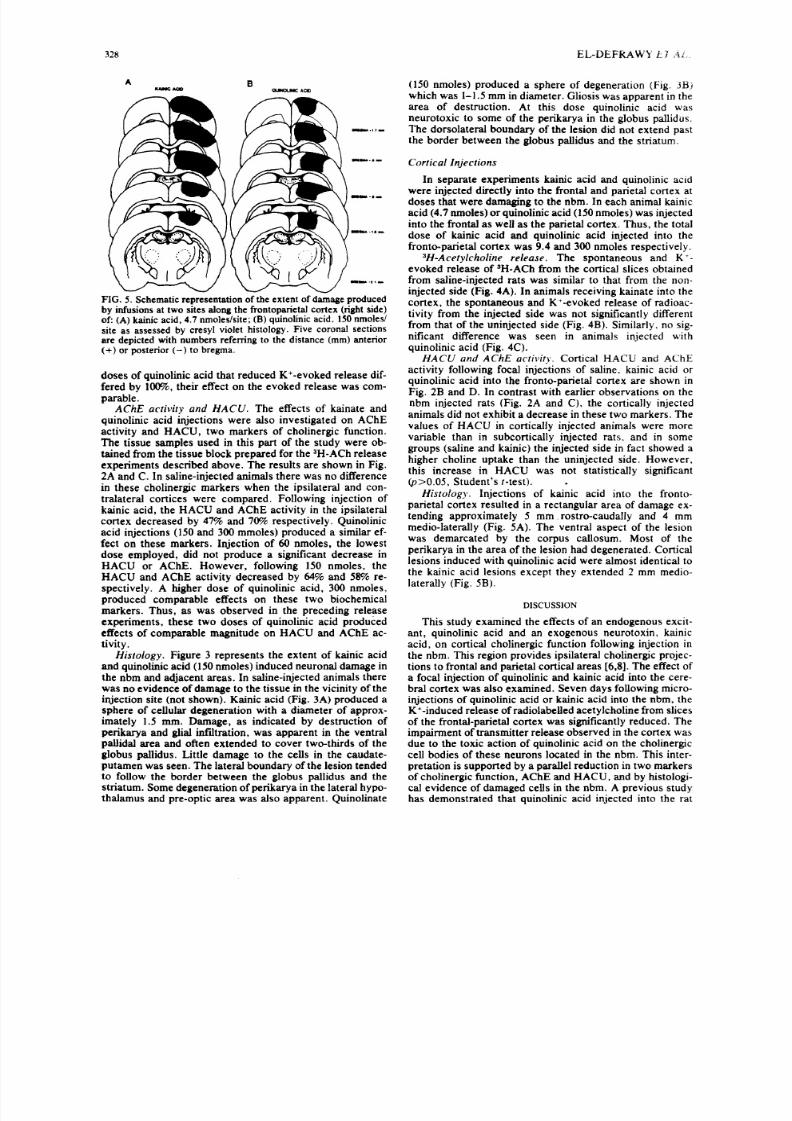
His to logy. Figure 3 represents the extent of kainic acidand quinolinic acid (150 nmoles) induced neuronal damage in
the nbm and adjacent areas. In saline-injected animals therewas no evide nce of damage to the tissue in the vicinity of theinjection site (not shown). Kainic acid (Fig. 3A) produced asphere of cellular degeneration with a diameter of approx-imately 1.5 ram. Damage, as indicated by destruction ofperikarya and glial infiltration, was apparent in the ventralpallidal a rea and often e xtended t o cove r two-thirds of theglobus pallidus. Little damage to the cells in the cauda te-putamen was seen. The lateral boundary of the lesion tendedto follow the border between the globus pallidus and thestriatum. Some degeneration of perikarya in the lateral hypo-thalamus and pre-optic area was also apparent. Quinolinate
(150 nmoles) produced a sphere of degeneration (Fig. 3B)which was I-1. 5 mm in diameter. Gliosis was apparent in thearea of destruction. At this dose quinolinic acid wasneurotoxic to some of the perikarya in the globus pallidus.The dorsolateral boundary of the lesion did not extend pastthe borde r between the globus pallidus and the striatum.
Cortica l Injec tions
In separate experiments kainic acid and quinolinic acidwere injected directly into the frontal and parietal cortex atdoses that were damaging to the nbm. In each animal kainicacid (4.7 mol e s ) or quinolinic acid (150 nmoles) was injectedinto the frontal as well as the parietal cortex. Thus, the totaldose of kainic acid and quinolinic acid injected into thefronto-parietal cortex was 9.4 and 300 nmoles respectively.
3H-Acetylcholine re lease .The spontane ous and K *-evoked release of 3H-ACh from the cortical slices obtainedfrom saline-injected rats was similar to that from the non-injected side (Fig. 4A). In animals receiving kainate into thecortex, the spontaneous and K~-evoked release of radioac-tivity from the injected side was not significantly differentfrom that of the uninjected side (Fig. 4B). Similarly, no sig-
nifican t difference was seen in animals injected withquinolinic acid (Fig. 4C).H A CU a n d A Ch E a c t i v i t y.Cortical HACU and AChE
activity following focal injections of saline, kainic acid orquinolinic acid into the fronto-parietal cortex are shown inFig. 2B and D. In contrast with earlier observations on thenbm injected rats (Fig. 2A and C), the cortically injectedanimals did not exhibit a decrease in these two markers. Thevalues of HACU in cortically injected animals were morevariable than in subcortically injected rats, and in somegrou ps (saline and kainic) the injected side in fact showe d ahigher choline uptake than the uninjected side. However,this increase in HACU was not statistically significant(p>0.05, Student's t-test).
His to logy. Injections of kainic acid into the fronto-
parietal cortex resulted in a rectangular area of damage ex-tending approximately 5 mm rostr o-caud ally and 4 mmmedio-laterally (Fig. 5A). The ventral aspect of the lesionwas demarcated by the corpus callosum. Most of theperikarya in the area of the lesion had degenerated. Corticallesions induced with quinolinic acid were almost identical tothe kainic acid lesions except they extended 2 mm medio-laterally (Fig. 5B).
DISCUSSION
This study examined the effects of an endogenous excit-ant, quinolinic acid and an exogenous neurotoxin, kainicacid, on cortical cholinergic function following injection inthe nbm. This region provides ipsilateral cholinergic projec-tions to frontal and parietal coni cal areas [6,8]. The effect of
a focal injection of quinolinic and kainic acid into the cere-bral cortex was also examined. Seven days following micro-injections of quinolinic acid or kainic acid into the nbm, theK*-induced release o f radiolabelled acetylcholine from slicesof the frontal-parietal cortex was significantly reduced. Theimpairment of transmitter release observed in the cortex wasdue to the toxic action of quinolinic acid on the cholinergiccell bodies of these neurons located in the nbm. This inter-pretation is suppor ted by a parallel reduction in two marker sof cholinergic function, AChE and HACU, and by histologi-cal evidence of damaged cells in the nbm. A previous studyhas demonstrated that quinolinic acid injected into the rat

8/3/2019 S.R. El-Defrawy et al- Functional and Neurochemical Cortical Cholinergic Impairment Following Neurotoxic Lesions of the Nucleus Basalis Magnocellularis in the …
http://slidepdf.com/reader/full/sr-el-defrawy-et-al-functional-and-neurochemical-cortical-cholinergic-impairment 5/6
CHOLINERGIC FUNCTION AND NBM LESIONS 329
striatum reduces A ChE and HACU, as well as CAT activity[16]. This cholinergic enzyme marker was not measured inthe present study, however , it has been measured in a follow-ing study and was reduced by quinolinate injections (seefollowing report).
As was observed by Schwarcz et a l . [16], who reported adose related reduction in striatal CAT activity after injec-
tions o f 60--600 nmoles o f quinolinic acid, the lowes t dose (60nmoles) injected into the nbm study failed to modify the twocholinergic markers or the release of aH-ACh from the cere-brad cortex in this study. However, two higher doses (150and 300 mo le s) clearly produced significant reductions in allpresynaptic markers of cholinergic function. In the presentstudy the maximal neurotoxic effect of quinolinic acid onaH-ACh release and cholinergic markers occurred at 150nmoles. In contrast, the peak effect of quinolinic acid onstriatal cholinergic neurons reported by Sc hwarcz et al. [16]occur red at 600 nmoles. This difference m ay be due to differ-ences in the sensitivity of cholinergic neurons to the action o fquinolinic acid in the striatum and the nbm. Variations inregional sensitivity of brain neurons to the excitant action ofquinolinic acid after microinotophoretic application has beenreported by Perkins and Stone [14,15]. It may also be due tothe use of halothane anesthesia in this study and barbiturateanesthesia, which is known to restrict the degree ofneurotoxicity, in the Schwar cz study [16].
Quinolinic acid in the present study was considerably lesspotent than kainic acid. Injections of 150 nmoles quinolinicacid produced effects on cholinergic function comparable insize to the effects produced by 4.7 nmoles kainic acid. Thisdifference in potency could be related to different receptorsactivated by these two agents. Evidence from elec-trophysiological [14, 15, 22] and transmitter release [9] ex-periments indicates that quinolinate preferentially activatesMg-sensitive NMDA (N-methyI-D-aspartate)-type aminoacid receptors, while kainate preferentially activates Mg-insensitive kainate-type receptors to produce neuronal exci-tation.
The failure of quinolinate or kainate to abolish corticalcholinergic function following nbm injection suggests thatthe residual function might be due to intrinsic corticalcholinergic neurons [3]. This provided the basis forintracortical injections. In previous work injections of kain-ate into the cerebral cortex yielded conflicting results.Johnston et a l . [7] reported a 30% reduction in CAT activityafter injecting 9.4 nmoles into rat cortex. In contrast, follow-ing an intracortical kainate injection, Lehmann et al. [8]failed to observe a significant reduction in CAT activity al-though the level of glutamic acid decarboxylase was re-duced. In the present study injections of quinolinic acid orkainic acid into the fronto-parietal cortex, at twice the dosethat pr oduced damage in the nbm, failed to decreas e :~H-AChrelease, AChE activity or HACU. An apparent increase incholine uptake observed after intracortical injections mightbe due to factors such as mechanical tissue irritation result-ing from such injections.
A lack of effect of cortically administered toxins oncholinergic neurons further confirms the axon-sparring
neurotoxic action of these agents. Despite a relatively highdose of kainate (9.4 nmoles) injected into the fronto-parietalcortex, the presynaptic cholinergic function in the injectedside was not reduced. Histological assessment, however,provided clear evidence of damage to the cerebral cortex.Thus, failure of intracortical injections to induce cholinergicdamage was not due to the lack of delivery of an adequate
dose of the neurotoxin. The results of the present studywould suggest that intrinsic cholinergic neurons either areresistant to the action of these toxins or that such elementsmake a modest contribution to the total release of 3H-AChevoked by K*-depolarization. The contribution for suchneurons to HACU and AChE activity also may be quitesmall and difficult to detect considering the intergroup vari-ance in these two markers. Eckenstein and Thoenen [3] haverecently demonstrated cholinergic cell bodies in the cerebralcortex. The sensitivity of such intrinsic cholinergic neuronsto neurotoxins remains to be examined in future experi-ments.
Injections of quinolinic acid into the nbm in this studyproduced the functional cholinergic deficit which previouslyhas been observed in the biopsied cerebral cortex o f humanpatients with Alzheimer's disease. Sims et a l . [17,18]demonstrated that K+-stimulated (31 raM) release ofradiolabeiled acetylcholine from slices ofbiopsied neocortexof Alzheimer's patients was reduced when compared withnon-disease controls. In the present study, 35 mM K*-evoked release of acetylcholine from cortical slices ofquinolinic or kainic acid treated animals was also reducedwhen compared to contralateral controls. The spontaneousreleas e of '~H-acetylcholine f rom slices was not signifi cantlyaffected in lesioned rats. LoConte et a l . [I0] observed an18% reduction in the in vivo release of cortical acetylcholine,over a 20 minute period, in rats following electrolyti c lesionsof the forebrain. A lack of effect on the spontaneous releasein the present study could be due to file fact that the releasefrom slices is already low, owing to a lack of impulse flowfrom the subcortical cell bodies, and small changes in thisrelease may not be readily apparent. Measurements of thisrelease over extended periods may reveal a deficit as is ob-served in the K" release. The observation that quinolinatecauses cholinergic dysfunction raises the possibility thatdamage to cholinergic neurons in AIzheimer's disease mightresult from an abnormal production or disposition of an en-dogenous neurotoxin such as quinolinic acid, or from an ab-normal sensitivity to its synaptic actions. Quinolinic acid isfound in human and animal brain [ 23] and its concent rati on isapparently increased by administration of tryptophan or inaging Ill].
ACKNOWLEDGEMENTS
This research was supported by a grant f rom the Ontario MentalHealth Foundation. R. J. Beninger and S. R. EI-Defrawy were sup-ported by the Ontario Ministry of Health and the Canadian Geriat-rics Research Society. The authors wish to thank Paul Payson andBrenda Scarth for their technical assistance, and Ms. D. Browne fortyping the manuscript.
REFERENCES
1. Coyle, J. T., D. L. Price and M. R. Delong. Alzheimer's dis-ease: A disorder of cortical cholinergic innervation. Science
219: 1184-1190, 1983.
2. Davies. P. Neurotransmitter-related enzymes in senile dementiaof the Alzheimer's type. Brain Res 171: 319-327, 1979.

8/3/2019 S.R. El-Defrawy et al- Functional and Neurochemical Cortical Cholinergic Impairment Following Neurotoxic Lesions of the Nucleus Basalis Magnocellularis in the …
http://slidepdf.com/reader/full/sr-el-defrawy-et-al-functional-and-neurochemical-cortical-cholinergic-impairment 6/6
330 E L - D E F R A W YE] AL
3 . E ck en s t e in , F. an d H . T h o en en . C h o l in e rg i c n eu ro n s i n th e r a tce reb ra l co r t ex d em o n s t r a t ed b y im m u n o h i s to ch em ica l l o -cal izat ion o f cho l ine acety l t ransferase .Neurosci Lett 3 6 : 2 11 -215, 1983.
4 . E U m an , G . L . , K . C o u r tn ey, S . A n d res and R . F ea th e r s to n e . An ew rap id ca lo r im e t r i c d e t e rm in a t io n o f ace ty l ch o l in es t er a seact iv i ty. Biochem Pharmacol 7: 88-95, 1961.
5 . Har tg raves , S . L . , P. L . Mensah and P. H. Kel ly. Reg ional
decre ases o f co r t ical cho l ine acety l t ransferase af ter les ions o fthe sep tal area and in the area o f nucleus basal is magn ocel lar is .Neuroscience 7: 2369-2376, 1982.
6 . Jo h n s to n , M . V. , M . M cK in n ey an d J . T. C o y le . E v id en ce fo r acho l inerg ic p ro ject ion to the neocor tex f rom neurons in thebasal fo reb raln .Proc Natl Acad Sci USA 76: 5392-5396, 1979.
7 . Jo h n s to n , M . V. , M . M cK in n ey an d J . T. C o y le . N eo co r t i ca lcho l inerg ic innervat ion : A descr ip t ion o f ex t r ins ic and in t r ins icco m p o n en t s i n t h e r a t.Exp Brain Res 43: 159-172, 1981.
8 . L eh m an n , J . , J . I . N ag y, S . A tm ad ja an d H . C . F ib ig e r. T h enucleus basal is m agnocel lu lar is : The o r ig in o f a cho l inerg ic p ro -j ec t i o n to t h e n eo co r t ex o f t h e r a t.Neuroscience 5 :1161-1174 ,1980.
9 . Lehm ann , J . and B . Scat ton . Character iza t ion o f the exci ta to ryamino a cid recep to r m ed iated re leas e o f '~H-acety lcho l ine f romrat striatal slices.Brain Res 252: 77-89, 1982.
10 . LoConte , G . , L . Bar to l in i , F. Casament i , I . Marconcin i -Pepeu
and G. Pepe u . L esions o f cho l inerg ic fo reb rain nuclei : Changesin av o id an ce b eh av io u r an d sco p o lam in e ac t i o n s .PharrnacolBiochem Behav 17: 933-9 37, 1982.
11. Lom bard i , G . , R . Corradet t i , V. Car la , C . Ald in io and F. Mo-ron i . Ma ss-f ragm entograph ic iden t i f icat ion o f the exci to tox inqu ino l in ic acid : On toge net ic changes and modu lat ion o f i ts con-cen tra t ion in the ra t b rain by t ryp tophan and byp-ch lo ropheny lalan ine.Soc Neurosci Abstr 9 :1184 , 1983 .
1 2 . L o w ry, O . H . , N . J . R o sen b ro u g h , A . L . F a r r an d R . S . R an -dal l . P ro tein measurements wi th the fo l in pheno l method .J BiolChem 193: 265-275, 1951.
13. Pax inos, G . and C . W atson .The Rat Brain m 5tereotaxi,.Coordinates. Toron to , Canada: Academic Press , 1982 .
14 . Perk ins , M. N. and T. W. S tone. Pharmaco logy and reg ionalvar ia t ions o f qu ino l in ic acid -evoked exci ta t ions in the ra t cen tra ln e rv o u s sy s t em .J Pharmacol Exp Ther 226: 551-557, 1983.
15. Perk ins , M. N . and T. W . S tone. Qu ino l in ic acid : Reg ional var-ia t ions in neuronal sensi t iv i ty.Brain Res 259: 172-176, 1983.
16. Schw arcz, R . , W . O. Whetsel l and R . M. Mangano . Q u ino l in ic
acid : An endogenou s m etabo l i te that p roduces axo n-spar ing te-s ions in ra t b rain .Science 219: 316-318, 1983.17. S imon , J . R . , S . Atw eh and M. J . Kuhar. So d ium-d ependen t
h igh aff in i ty cho l ine up take: A regu lato ry s tep in the syn thesiso f acety lcho l ine.J Neurochem 26: 909-922, 1976.
18 . S ims, N. R . , O . M. Bowen ,C . C . T. S m i t h , R . H . A . H a c k , A .N . D av i so n , J . S . S n o w d en an d D . N ea ry. G lu co se m e tab o l i smand acety icho l ine systhesis in re la t ion to neuronal act iv i ty inA lzh e im er ' s d i sease .Lancet 1: 333-3 36. 198Q
19 . S ims, N. R . , D . M. Bowen and A. N. Dav ison . '~Acety lcho l inesyn thesis and ~4C carbon d iox ide p roduc t ion f rom (U- '~C) g lu -co se b y t i s su e p r i sm s o f h u m an n eo co r t ex .Biochern J 196:867-876, 1981.
20 . Som ogyi , G . T. and J . C . Szerb . D emo nstra t ion o f acety lcho l inerelease by measu r ing label led cho l ine eff iux f rom cerebral co r t i -cal slices.J Neurochem 19: 2667-2677, 1972.
21 . Vincen t , S . R . , K . Satoh , D. M. Arm strong and H. C . F ib iger.
Substance P in the ascend ing cho l inerg ic re t icu lar system. Na-ture 306: 688-691, 1983.22 . Watk ins , J . C . P harm aco logy o f exci ta to ry amino acid recep-
tors. In: Glutamate: Transmitter in the Central Nervous Sys-tem, ed i ted by P. J . Rober ts , J . S to rm-Math isen and G. A. R .Johnston . Ch iche ster, U .K . : John Wiley, 1981 , pp . 1 -24 .
2 3. W o l fen sb e rg e r, M . , U . A m s le r, M C u en o d , A . C . F o s t e r, W.D. Whetsel l and R . Sch warcz. Iden t i f icat ion o f qu ino l in ic acidin ra t and human b rain t i ssue.Neurosci Lett 41: 247-252, 1983.