Embed Size (px)
Citation preview

Biotechnol. Appl. Biochem. (2002) 36, 235–239 (Printed in Great Britain) 235
Stabilization of α-chymotrypsin by modification withβ-cyclodextrin derivatives
Michael Ferna! ndez*, Marı!a de Lourdes Villalonga*, Alex Fragoso†,Roberto Cao† and Reynaldo Villalonga*1
*Enzyme Technology Group, Center for Biotechnological Studies, University of Matanzas, Matanzas, C.P. 44740, Cuba, and†Laboratory of Bioinorganic Chemistry, Faculty of Chemistry, University of Havana, Havana 10400, Cuba
Bovine pancreatic α-chymotrypsin was chemicallymodified with two different β-cyclodextrin derivatives,named mono-6-formyl-β-cyclodextrin and mono-6-succinyl-6-deoxy-β-cyclodextrin.The modifiedenzymescontained approx. 3–5 mol of oligosaccharide/mol ofprotein, and retained full proteolytic and esterolyticactivity. The optimum temperature for α-chymotrypsinwas increased by 8 °C and its thermostability wasenhanced by about 4–6 °C after modification. The con-jugated enzymes were also more resistant to thermalinactivation at temperatures ranging from 45 to 55 °C.Additionally, the modified enzymes were 7-fold morestable against incubation at pH 9.0. The possibleinfluence of supramolecular interactions on thethermal stabilization of modified α-chymotrypsins wasalso studied.
Introduction
There is considerable interest in developing new methodsfor increasing the thermal stability of an enzyme bymanipulation of its protein structure. In this regard,strategies for stabilizing enzymes have been mainly addressedto produce genetically or chemically modified proteinvariants [1–4]. Of these, the chemical method appears to bethe most promising, taking into account both the largenumber of recent reports and the improvements in enzymethermostability achieved with this approach [5–7].
In Nature, stabilization of thermophilic proteins isattributable to the combined action of several mechanisms,two of the most relevant being a high number of in-tramolecular cross-links (disulphide bonds, salt bridges andhydrogen bonds) and a low degree of surface hydrophobicity[8]. Considering this fact, the cross-linking of enzyme proteinstructures with hydrophilic compounds, especially carbo-hydrates, has been extensively and successfully used as a toolfor improving enzyme thermostability [2,6,9,10].
Cyclodextrins (CDs) are cyclic oligosaccharides con-taining six (α-cyclodextrin, αCD), seven (β-cyclodextrin,βCD) or eight (γ-cyclodextrin, γCD) α-1,4-linked D-gluco-pyranose units, with hydrophilic hydroxyl groups on theirouter surfaces and a hydrophobic cavity in the centre [11].
CDs can form stable inclusion complexes with hydrophobiccompounds [12], such as several amino acid residues locatedon as enzyme’s surface. In view of this property, a novelstrategy for improving enzyme thermostability could be theformation of intramolecular non-covalent cross-links,mediated by supramolecular interactions at the surface ofCD-modified enzymes.
This paper describes a new method for improving thestability to bovine pancreatic α-chymotrypsin (EC 3.4.21.1)by conjugation with two different βCD derivatives, namedmono-6-formyl-β-cyclodextrin (CD1) and mono-6-succinyl-6-deoxy-β-cyclodextrin (CD2). In addition, the possibleinfluence of supramolecular interactions on the increasedenzymic thermostabilization was also studied.
Materials and methods
MaterialsBovine pancreatic α-chymotrypsin (45 units}mg), N-α-acetyl-L-tyrosine ethyl ester hydrochloride (ATEE), 1-ethyl-3-(3-dimethylaminopropyl)carbodi-imide (EDAC), NaBH4
and Fractogel EMD BioSEC (S) were obtained from Merck(Darmstadt, Germany). βCD was purchased from Amaizoand used as received. All other chemicals were analyticalgrade.
Preparations of the conjugatesα-Chymotrypsin–CD1 CD1 was prepared by DMSO oxi-dation of the corresponding βCD monotosylate [13] in the
Key words : enzyme thermostabilization, neoglycoenzyme, serine protease,supramolecular interaction.
Abbreviations used : CD, cyclodextrin ; αCD, α-cyclodextrin ; βCD, β-cyclodextrin ; γCD, γ-cyclodextrin ; CD1, mono-6-formyl-β-cyclodextrin ;CD2, mono-6-succinyl-6-deoxy-β-cyclodextrin ; ATEE, N-α-acetyl-L-tyrosine ethyl ester hydrochloride ; EDAC, 1-ethyl-3-(3-dimethylaminopropyl)carbodi-imide.
1 To whom correspondence should be addressed (e-mailreynaldo.villalonga!umcc.cu).
# 2002 Portland Press Ltd

236 M. Ferna! ndez and others
presence of 2,4,6-trimethylpyridine at 130 °C under nitro-gen, followed by precipitation with ethanol [14]. The productwas characterized by conventional NMR techniques andalso by conversion into its oxime derivative [14]. Forattaching CD1 to α-chymotrypsin, 10 mg of the enzyme,dissolved in 5 ml of 50 mM sodium phosphate buffer, pH 6.0,was mixed with 50 mg of CD1 and stirred at 4 °C in the darkfor 1 h. NaBH4 (30 mg) was added and the reaction mixturewas kept under continuous stirring for 4 h. The solution wasfurther dialysed against 20 mM sodium acetate buffer,pH 5.0, and then applied to a Fractogel EMD BioSEC (S) gel-filtration column (2.6 cm¬60 cm), equilibrated in the samebuffer containing 100 mM NaCl. The fractions containing thepolysaccharide–enzyme complex were pooled and kept at4 °C.
α-Chymotrypsin–CD2 To a cold solution of succinic acid(2.4 g, 20 mmol) in dry dimethylformamide (25 ml),dicyclohexylcarbodi-imide (6.2 g, 30 mmol) was added andthe solution was stirred at 5 °C for 30 min. To this solution,mono-6-amino-6-deoxy-βCD (2.3 g, 2 mmol) [15] wasadded in one portion and the solution was stirred at 25 °C.After 24 h, the insoluble precipitate of dicyclohexylurea wasfiltered off and the remaining solution was precipitated byaddition over acetone (300 ml), washed several times withacetone and dried under high vacuum. The CD2 derivativesynthesized by this procedure was characterized by con-ventional NMR techniques. The conjugate α-chymotrypsin–CD2 was further prepared by adding 50 mg of EDAC to areaction mixture containing 50 mg of the activated oligo-saccharide and 10 mg of protease dissolved in 5 ml of 50 mMsodium phosphate buffer, pH 6.0. The solution was stirredfor 1 h at room temperature and for 16 h at 4 °C, and finallydialysed and purified as described previously for the α-chymotrypsin–CD1 complex.
AssaysEsterolytic activity of native and modified α-chymotrypsinswas determined at 25 °C in 50 mM sodium phosphate buffer,pH 7.0, using ATEE as a substrate [16]. One unit ofesterolytic activity is defined as the amount of enzyme thathydrolyses 1.0 µmol of ATEE per min at 25 °C. Proteolyticactivity was determined as described by Laskowski [17] usingmilk casein as a substrate. Total carbohydrates weredetermined by the phenol}H2SO4 method [18] using glucoseas a standard. Protein concentration was estimated asdescribed by Lowry et al. [19] using BSA as a standard. Thedegree of modification of amino groups was determined bymeasuring the amount of free amino groups with o-phtalaldehyde using glycine as a standard [20].
Results and discussion
In the present work, two different strategies were used tomodify the free amino groups of α-chymotrypsin withsynthesized βCD derivatives (Figure 1) : (i) reductive alkyl-ation of the protein with CD1 in the presence of NaBH4
and (ii) attachment of CD2 via amide bonds using EDAC asa coupling agent. Table 1 reports the structural and catalyticproperties of the enzyme conjugates prepared.
According to the measured carbohydrate content, theCD-modified enzymes have an average of 3 and 5 mol ofCD}mol of protein, respectively, for the CD1 and CD2conjugates. These results correlate well with the degree ofmodification estimated for these adducts by determining theamount of free amino groups in the enzyme before and afterthe attachment of the oligosaccharide moieties [20]. Thedifferences in carbohydrate content in both conjugates mightbe due to the different conditions employed to attach thecarbohydrate. Whereas amide formation with CD2proceeds smoothly, reductive ammination requires theprevious formation of an unstable aliphatic Schiff-baseintermediate that might be decomposed as the reactiontakes place. This could be the reason why CD1-conjugatesshowed a lower carbohydrate content.
The specific activity of α-chymotrypsin towards caseinwas not significantly affected after modification. Additionally,the α-chymotrypsin–CD2 complex retained a high estero-lytic activity towards ATEE. Interestingly, the esterolyticactivity of the enzyme was increased to about 122% aftermodification with the CD1 derivative. These results suggestthat the active conformation of the enzyme was not affectedby the attachment of the oligosaccharide residues. Similarcatalytic behaviour has been previously reported for α-chymotrypsin in organic media after co-lyophilization withcyclodextrins [21].
The covalent modification of α-chymotrypsin by theCD derivatives yields to several positive changes on thefunctional stability of the protease under various denaturing
Figure 1 Modifying cyclodextrin derivatives
# 2002 Portland Press Ltd
β-Cyclodextrin derivatives stabilize α-chymotrypsin 237
Table 1 Structural and catalytic properties of native and modified α-chymotrypsin preparations
Data are means from three measurements ; relative errors were ! 5%.
Enzyme form Carbohydrate content (%, w/w)Degree of modification ofNH2 groups (%, mol/mol) Proteolytic activity (katal/kg) Esterolytic activity (units/mg)
α-Chymotrypsin – – 5.6¬10−2 44.6α-Chymotrypsin–CD1 14 18 5.9¬10−2 54.6α-Chymotrypsin–CD2 23 30 5.2¬10−2 42.7
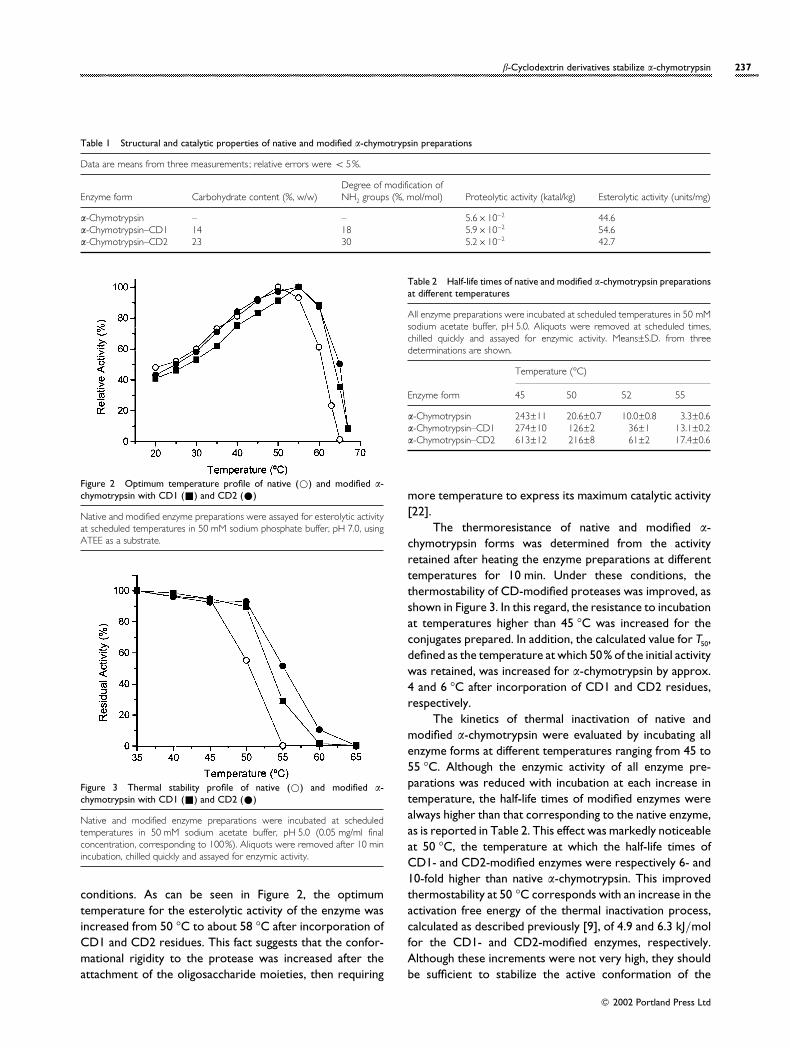
Figure 2 Optimum temperature profile of native (D) and modified α-chymotrypsin with CD1 (+) and CD2 (E)
Native and modified enzyme preparations were assayed for esterolytic activityat scheduled temperatures in 50 mM sodium phosphate buffer, pH 7.0, usingATEE as a substrate.
Figure 3 Thermal stability profile of native (D) and modified α-chymotrypsin with CD1 (+) and CD2 (E)
Native and modified enzyme preparations were incubated at scheduledtemperatures in 50 mM sodium acetate buffer, pH 5.0 (0.05 mg/ml finalconcentration, corresponding to 100%). Aliquots were removed after 10 minincubation, chilled quickly and assayed for enzymic activity.
conditions. As can be seen in Figure 2, the optimumtemperature for the esterolytic activity of the enzyme wasincreased from 50 °C to about 58 °C after incorporation ofCD1 and CD2 residues. This fact suggests that the confor-mational rigidity to the protease was increased after theattachment of the oligosaccharide moieties, then requiring
Table 2 Half-life times of native and modified α-chymotrypsin preparationsat different temperatures
All enzyme preparations were incubated at scheduled temperatures in 50 mMsodium acetate buffer, pH 5.0. Aliquots were removed at scheduled times,chilled quickly and assayed for enzymic activity. Means±S.D. from threedeterminations are shown.
Enzyme form
Temperature (°C)
45 50 52 55
α-Chymotrypsin 243±11 20.6±0.7 10.0±0.8 3.3±0.6α-Chymotrypsin–CD1 274±10 126±2 36±1 13.1±0.2α-Chymotrypsin–CD2 613±12 216±8 61±2 17.4±0.6
more temperature to express its maximum catalytic activity[22].
The thermoresistance of native and modified α-chymotrypsin forms was determined from the activityretained after heating the enzyme preparations at differenttemperatures for 10 min. Under these conditions, thethermostability of CD-modified proteases was improved, asshown in Figure 3. In this regard, the resistance to incubationat temperatures higher than 45 °C was increased for theconjugates prepared. In addition, the calculated value for T50,defined as the temperature at which 50% of the initial activitywas retained, was increased for α-chymotrypsin by approx.4 and 6 °C after incorporation of CD1 and CD2 residues,respectively.
The kinetics of thermal inactivation of native andmodified α-chymotrypsin were evaluated by incubating allenzyme forms at different temperatures ranging from 45 to55 °C. Although the enzymic activity of all enzyme pre-parations was reduced with incubation at each increase intemperature, the half-life times of modified enzymes werealways higher than that corresponding to the native enzyme,as is reported in Table 2. This effect was markedly noticeableat 50 °C, the temperature at which the half-life times ofCD1- and CD2-modified enzymes were respectively 6- and10-fold higher than native α-chymotrypsin. This improvedthermostability at 50 °C corresponds with an increase in theactivation free energy of the thermal inactivation process,calculated as described previously [9], of 4.9 and 6.3 kJ}molfor the CD1- and CD2-modified enzymes, respectively.Although these increments were not very high, they shouldbe sufficient to stabilize the active conformation of the
# 2002 Portland Press Ltd
238 M. Ferna! ndez and others
protease, taking into account that the net free energy forstabilization of proteins is, in general, small [23].
It should be noted that the conjugate α-chymotrypsin–CD1 was slightly less thermoresistant than the α-chymotrypsin–CD2, probably associated with the higheramount of CD residues attached to the latter complex.
The stability against thermal denaturation induced bythe attachment of the highly hydrophilic oligosaccharidemoieties to the protease could be mainly due to theprevention of interactions between hydrophobic clusterslocated on the protein surface and surrounding watermolecules. In fact, this kind of energetically unfavourableinteraction has been reported as one of the most importantfactors contributing to protein denaturation at hightemperatures [1]. In this regard, a similar mechanism ofstabilization has been reported previously for the sameprotease modified with several hydrophilic compounds [24].
Additionally, the ability of CD to form inclusioncomplexes with hydrophobic compounds [12] could beanother explanation for the improved thermal resistanceshown by the modified α-chymotrypsin. In this sense, theformation of inclusion complexes between the attached CDgroups and the hydrophobic amino acid residues located inthe vicinity of the covalent modification points could induceadditional conformational stabilization to α-chymotrypsin. Inorder to test this last hypothesis, the thermal inactivationprocesses of native and CD-modified α-chymotrypsins wereevaluated at 50 °C in the presence and absence of a saturatedsolution of 1-adamantanol (approx. 10−4 M). This moleculeforms a highly stable inclusion complex with βCD, with anassociation constant in the order of 104 M−1 [25]. For thisreason, it is expected that the presence of 1-adamantanol inthe conjugated enzyme solutions could disrupt any stabilizingsupramolecular interaction occurring in the conjugates.
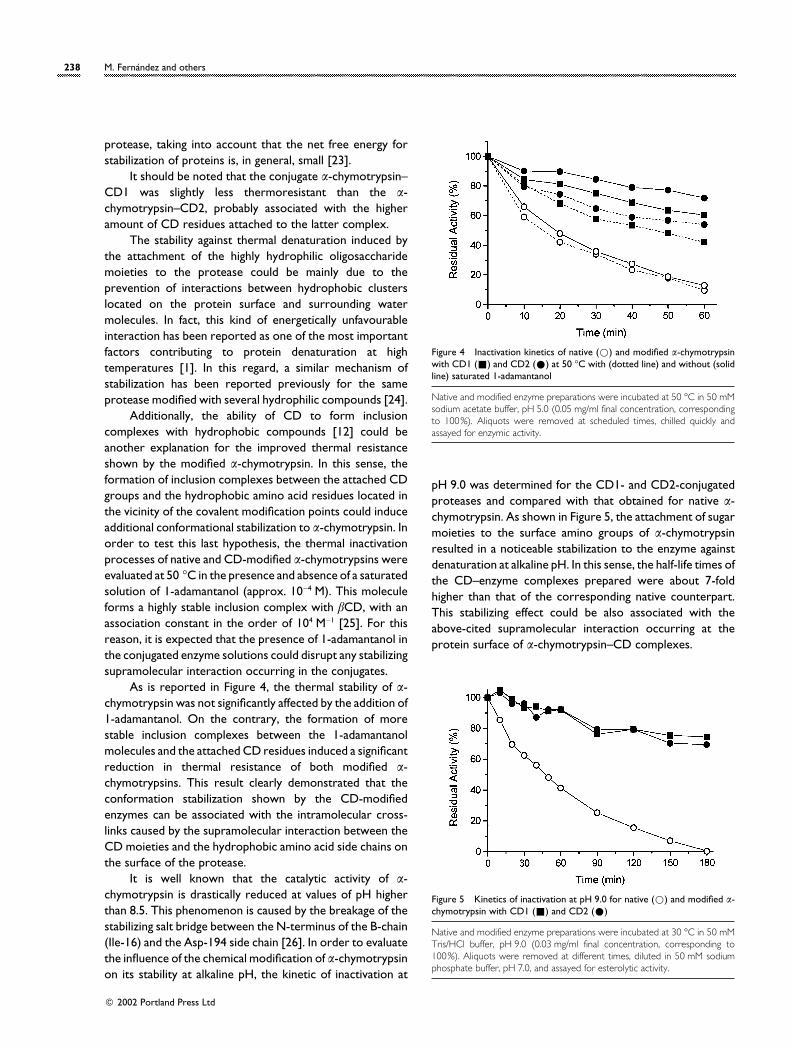
As is reported in Figure 4, the thermal stability of α-chymotrypsin was not significantly affected by the addition of1-adamantanol. On the contrary, the formation of morestable inclusion complexes between the 1-adamantanolmolecules and the attached CD residues induced a significantreduction in thermal resistance of both modified α-chymotrypsins. This result clearly demonstrated that theconformation stabilization shown by the CD-modifiedenzymes can be associated with the intramolecular cross-links caused by the supramolecular interaction between theCD moieties and the hydrophobic amino acid side chains onthe surface of the protease.
It is well known that the catalytic activity of α-chymotrypsin is drastically reduced at values of pH higherthan 8.5. This phenomenon is caused by the breakage of thestabilizing salt bridge between the N-terminus of the B-chain(Ile-16) and the Asp-194 side chain [26]. In order to evaluatethe influence of the chemical modification of α-chymotrypsinon its stability at alkaline pH, the kinetic of inactivation at
Figure 4 Inactivation kinetics of native (D) and modified α-chymotrypsinwith CD1 (+) and CD2 (E) at 50 °C with (dotted line) and without (solidline) saturated 1-adamantanol
Native and modified enzyme preparations were incubated at 50 °C in 50 mMsodium acetate buffer, pH 5.0 (0.05 mg/ml final concentration, correspondingto 100%). Aliquots were removed at scheduled times, chilled quickly andassayed for enzymic activity.
pH 9.0 was determined for the CD1- and CD2-conjugatedproteases and compared with that obtained for native α-chymotrypsin. As shown in Figure 5, the attachment of sugarmoieties to the surface amino groups of α-chymotrypsinresulted in a noticeable stabilization to the enzyme againstdenaturation at alkaline pH. In this sense, the half-life times ofthe CD–enzyme complexes prepared were about 7-foldhigher than that of the corresponding native counterpart.This stabilizing effect could be also associated with theabove-cited supramolecular interaction occurring at theprotein surface of α-chymotrypsin–CD complexes.
Figure 5 Kinetics of inactivation at pH 9.0 for native (D) and modified α-chymotrypsin with CD1 (+) and CD2 (E)
Native and modified enzyme preparations were incubated at 30 °C in 50 mMTris/HCl buffer, pH 9.0 (0.03 mg/ml final concentration, corresponding to100%). Aliquots were removed at different times, diluted in 50 mM sodiumphosphate buffer, pH 7.0, and assayed for esterolytic activity.
# 2002 Portland Press Ltd

β-Cyclodextrin derivatives stabilize α-chymotrypsin 239
Acknowledgments
This research was supported by the International Foundationfor Science, Stockholm, Sweden, and the Organization forthe Prohibition of Chemical Weapons, The Hague, TheNetherlands, through a grant to R.V. (grant no. F}3004-1).We thank Ms Maysa Ban4 os for her assistance in thepreparation of CD derivatives.
References
1 Klibanov, A. M. (1983) Adv. Appl. Microbiol. 29, 1–28
2 Villalonga, R., Villalonga, M. L. and Go! mez, L. (2000) J. Mol.
Catal. B Enzymatic 10, 483–490
3 Minagawa, H. and Kaneko, H. (2000) Biotechnol. Lett. 22,
1131–1133
4 Roig, M. G. and Kennedy, J. F. (1992) CRC Crit. Rev. Biotechnol.
12, 391–412
5 Darias, R. and Villalonga, R. (2001) J. Chem. Technol.
Biotechnol. 76, 489–493
6 Sundaram, P. V. and Venkatesh, R. (1998) Protein Eng. 11,
691–698
7 Go! mez, L., Ramı!rez, H. L. and Villalonga, R. (2001) Acta
Biotechnol. 21, 265–273
8 Mozhaev, V. V., Berezin, I. V. and Martinek, K. (1988) CRC Crit.
Rev. Biochem. 23, 235–281
9 Srivastava, R. A. K. (1991) Enzyme Microb. Technol. 13,
164–170
10 Go! mez, L. and Villalonga, R. (2000) Biotechnol. Lett. 22,
1191–1195
11 Szejtli, J. (1998) Chem. Rev. 98, 1743–1754
12 Szejtli, J. (1982) Cyclodextrins and their Inclusion Compounds,
Akade! miai Kiado! , Budapest
13 Zhong, N., Byun, H. S. and Bittman, R. (1998) Tetrahedron Lett.
38, 2919–2920
14 Yoon, J., Hong, S., Martin, K. A. and Czarnik, A. W. (1995) J.
Org. Chem. 60, 2792–2795
15 Fragoso, A., Cao, R. and Villalonga, R. (1995) J. Carbohydr.
Chem. 14, 1389–1396
16 Schwert, G. B. and Takenaka, Y. (1955) Biochim. Biophys. Acta
16, 570–575
17 Laskowski, M. (1955) Methods Enzymol. 2, 26–36
18 Dubois, M. K., Gilles, A., Hamilton, J. K., Rebers, P. A. and Smith,
F. (1956) Anal. Chem. 28, 350–356
19 Lowry, O. H., Rosebrough, N. J., Farr, A. L. and Randall, R.
(1951) J. Biol. Chem. 193, 265–275
20 Bruneel, D. and Schacht, E. (1993) Polymer 34, 2628–2632
21 Ooe, Y., Yamamoto, S., Kobayashi, M. and Kise, H. (1999)
Biotechnol. Lett. 21, 385–389
22 Rajalakshmi, N. and Sundaram, P. V. (1995) Protein Eng. 8,
1039–1047
23 Privalov, P. L. and Gill, S. J. (1988) Adv. Prot. Chem. 39,
191–234
24 Mozhaev, V. V., SC iks) nis, V. A., Melik-Nubarov, N. S., Galkantaite,
N. Z., Denis, G. J., Butkus, E. P., Zaslavsky, B. Y., Mestechkina,
N. M. and Martinek, K. (1988) Eur. J. Biochem. 173, 147–154
25 Cromwell, W. C., Bystro$ m, K. and Eftink, M. R. (1985) J. Phys.
Chem. 89, 326–333
26 Wroblowski, B., Diaz, J. F., Schlitter, J. and Engelborghs, Y.
(1997) Protein Eng. 10, 1163–1174
Received 14 June 2002; accepted 23 July 2002
# 2002 Portland Press Ltd





























